
Submitted:
29 September 2024
Posted:
29 September 2024
You are already at the latest version
Abstract
Outbreaks of avian pathogens such as Newcastle disease virus, avian influenza virus and Salmonella have a major impact on economies and food security worldwide. Some pathogens also pose a significant zoonotic potential, especially avian influenza viruses. Vaccination plays a key role in controlling many poultry diseases and there are many vaccines licenced in the United Kingdom for diseases of poultry caused by viruses, bacteria, and parasites. However, these vaccines often do not provide complete protection and can cause unwanted side effects. Several factors affect the potency of poultry vaccines including the type of vaccination used, the mechanism of delivery and the use of adjuvants. Advancements in technology have led to the study and development of novel vaccines and vaccine adjuvants for use in poultry. These induce stronger immune responses compared to current vaccine technology and have the potential to protect against multiple poultry diseases. This review aims to discuss existing poultry vaccine technology, the effect of delivery mechanisms of vaccine efficacy, the use of current and novel adjuvants, the ability to target antigen to antigen presenting cells, the use of probiotics, multivalent vaccines, and nanotechnology to enhance the potency of poultry vaccines.
Keywords:
Poultry
; vaccine potency
; antigen targeted vaccine
; nanotechnology
; probiotics
; adjuvants
1. Introduction
Poultry meat is an important food source for many people, with over 16 million tonnes estimated to be traded in 2022. Poultry meat is also projected to account for over half of all additional meat produced in the next decade. However, the increase in poultry meat trade is slowing down with poultry meat production in 2022 rising by only 0.6% from 2021, the slowest ever pace on record [1,2]. The poultry industry also provides another valuable food source in the form of eggs with egg production in 2020 exceeding 86 million metric tonnes. Global egg production is projected to increase 13% by 2029, with India and China accounting for nearly half of the global increase [2,3].
However, farmed poultry and associated egg production is at risk of severe disease due to infection from viruses such as avian influenza virus (AIV), Newcastle disease virus (NDV) and Marek’s disease virus (MDV) and bacteria such as Salmonella [4,5,6,7]. Outbreaks of infections in poultry flocks is a leading cause of economic loss in the poultry industry worldwide as viral epidemics are often associated with reduced weight gain, decreased egg production and increase the risk potential for zoonosis events [3,8]. The decrease in poultry meat trade seen in 2022 is thought to be in part by the severe widespread outbreaks of highly pathogenic avian influenza virus (HPAIV) that occurred in several of the major poultry farming regions including North and South America and Europe [2].
Vaccination is commonly used to control avian diseases and is primarily focused on the prevention of disease associated morbidity and mortality rather than prevention of the infection itself [9]. Antimicrobial therapy against bacterial infections is also commonly used both a preventative measure and active treatment in poultry farms. However, effective treatment requires diagnosis of the specific infectious agent as well as knowledge regarding dosage and potential interactions with other drugs. There is also a limited number of antimicrobial drugs that are commercially available for poultry therefore, accurate diagnosis is key for effective disease treatment and decrease in associated risk of antimicrobial resistance development [10]. Together, this makes vaccination of poultry a safer and more economical method of preventing disease in poultry.
Preventative measures are being adopted to reduce disease outbreaks. These include diagnostics and surveillance, improvement in biosecurity, reduction of virus spread via culling of infected and at-risk flocks , and better practices in housing, husbandry, vaccination, transport, slaughtering, and waste management [11]. However, poultry vaccines for some diseases, such as avian influenza, remain less effective at blocking virus transmission, allowing to continue circulate in both vaccinated and unvaccinated flocks [8]. Therefore, there is a need to increase the potency of current and future vaccines in poultry.
Most vaccines used in poultry are attenuated or inactivated pathogens (Table 1). However, there are issues with using these technologies as live attenuated vaccines can lead to reversion back to more virulent strains thus negating the vaccination in the first place. Theses vaccines are also associated with adverse reactions, especially when vaccinating young chicks [12]. Another important issue to consider is a need to distinguish between infected and vaccinated animals (DIVA) which is not possible when using attenuated or inactivated vaccines. In addition, attenuated or inactivated vaccines can sometimes become ineffective due to the presence of maternally derived antibodies (MDA) in early age chicks and so vaccination may have to be delayed till they are older [12,13].
2. Current Poultry Vaccine Technology
2.1 Inactivated Virus Vaccines:
One of the oldest methods of vaccine preparation is where native seed virus is cultured in embryonated chicken eggs or in cell culture. Cultured viral particles are then inactivated chemically (e.g. formalin, binary ethylenimine or β-propiolactone) or physically (e.g. heat, gamma or ultraviolet radiation) to destroy infectivity whilst preserving immunogenicity. This method is still used to make up the majority of registered avian influenza vaccines for poultry (Table 1) [3,14]. Inactivated vaccines are safer than live vaccines however they often induce low immunogenicity thus requiring booster doses and formulation with adjuvants (e.g., oil emulsion) for long lasting immunity [15,16]. The persistence of immunity is also often impaired by the age of the bird when the vaccination is given. For example, when given immediately after hatching the response is often lower than in birds immunised at 4 weeks old [17]. The immune response also has a slow onset, is not DIVA compliant and, in young animals, protection can be decreased due to interference from the MDA [3].
2.2 Live Attenuated Vaccines:
Conventional attenuation is typically achieved via the serial passaging of the wildtype pathogen in either irrelevant cell culture or host and has a long history of use against several poultry diseases (Table 1). Live attenuated vaccines are cost effective due to minimal scale up processing costs and induce both humoral and cellular immunity. However, the attenuation process has a long turnaround time and risk of reversion to the initial wildtype form or recombination with circulating pathogenic strains. More recently, reverse genetics has allowed for a more precise and faster method for generating live attenuated vaccines [3]. Coccidiosis is a parasitic infection of poultry caused by Eimeria ssp that is usually controlled by drug treatment however resistance is beginning to appear. To circumvent this, attenuated vaccines have been developed via continuous passage in eggs. However, these vaccines lose effectiveness over time resulting short expiry dates and have potential of reverting back to the more virulent strain [18]. Such live attenuated vaccines have been developed for protection against HPAIV in poultry. Attenuation was achieved via the removal of the haemagglutinin (HA) polybasic cleavage site. Further attenuation was also carried via genetic modification to truncate the NS1 genes. The study showed that vaccines carrying truncated versions of HPAIV NS1 genes completely protected poultry from homologous and heterologous virus challenges [19].
2.3 Subunit Vaccines:
It is not always necessary to include the whole pathogen in the vaccine formulation as some antigens can induce protective immune responses. Recombinant subunit vaccines are DIVA compatible, stable, well established and are considered safe as they don’t contain live viral components which in turn also reduces the likelihood of severe side effects [3,20]. Subunit vaccines can also be engineered to produce responses to several antigens either against serotypes of the same pathogen or against multiple pathogens. The use of several antigens has been shown to increase vaccine efficacy as one antigen is not always sufficient at producing a protective immune response [21,22]. However, subunit protein vaccines also tend to have a low immunogenic response as they can lack the pathogen associate molecular patterns (PAMPs) which are present on the whole virus. To overcome this, subunit vaccines require high dosage, adjuvants and frequent boosters to maintain the protective response [3,23]. The selected antigens can be presented in several types of novel vaccine platforms described below.
2.4 Virus-like Particles (VLPs) Vaccines:
VLPs are non-infectious structures which are made up of viral structural proteins. They have been successfully used in humans against hepatitis B virus and Human papilloma virus [24,25]. VLPs have also been shown to activate dendritic cells and so can stimulate both cellular and humoral immunity [26]. No VLP based vaccines have been licenced for use in poultry. VLPs can be manufactured using eukaryotic and prokaryotic expression systems. VLP vaccines are considered a safer alternative to live inactivated and attenuated vaccines due to the lack of genetic material [27]. Experimental studies have shown high potency of protection in chickens against viral infections such as AIV and NDV [28,29]. A bivalent VLP containing the HA and matrix 1 (M1) proteins of HPAIV H5N1 and the NDV fusion (F) protein was used to vaccinate chickens. The results showed that a single immunisation induced high levels of anti-NDV antibodies and haemagglutinin inhibition (HI) antibody titres against H5N1 AIV [30]. However, VLP production has a high cost associated with expression and purification, cold chain storage and reduced stability in field conditions means their commercial use is so far limited [3,27].
2.5 DNA Vaccines:
DNA vaccines are based on plasmids which encode the vaccine antigen. The antigen is produced by the cells of the vaccinated animal. DNA vaccines can encode genes for the expression of specific or multiple antigens to provide protection against several different pathogens or serotypes of the same pathogen. Chickens vaccinated with DNA plasmids encoding for infectious bronchitis virus (IBV) proteins S1, N or M alone or a combination of the three IBV proteins. The combination vaccine induced significantly higher antibodies compared with the monovalent vaccines [31]. DNA vaccines can also be designed to tigger specific cell mediated immune responses [21]. However, plasmids used to generate DNA vaccines may contain antibiotic resistance genes which confers a risk of transmission of antibiotic resistance to farmed birds [32]. Although the DNA vaccines were first studied in poultry in 1993 [33], only one poultry DNA vaccine has been granted a conditional licence in the USA against H5 HPAIV [3,34]. There are several reasons why DNA vaccines are not reaching the commercial stages. DNA vaccines are often administered intramuscularly and several doses to induce protection, this is not practical in field settings. In addition, studies tend to use SPF birds meaning in the field protection may be reduced due to the presence of MDAs [21,35].
2.6 mRNA Vaccines:
The first proof of principle mRNA vaccine was published over 20 years ago in the mouse model using the mRNA of influenza nucleoprotein (NP) [36]. This technology has been further developed in recent years especially for use in humans in response to the SARS-CoV-2 pandemic [37]. mRNA vaccines are considered to have a high safety profile as they are non-integrating and non-infectious, additionally mRNA is naturally degraded in the cell though the rate of degradation can be regulated via delivery methods. Modifications can also be made to make the mRNA more stable and less likely to be degraded thus more likely to be translated [38,39,40]. mRNA vaccines can also be administered repeatedly and have the potential for rapid and scalable manufacturing for emerging diseases and/or variants [40].
mRNA is the transitional stage between the translation of protein encoding DNA in the cell’s nucleus and protein production by ribosomes in the cell’s cytoplasm. There are two major types of RNA based vaccines currently being studied, these are non-replicating mRNA and self-amplifying RNA (saRNA). Non replicating mRNA vaccines encode the antigen of interest as well as the 5’ and 3’ untranslated regions (UTRs). saRNA vaccines encode the antigen of interest and machinery required for viral replication, this allows for intracellular RNA amplification and increased levels of protein expression. For optimal transcription the mRNA products need to contain an open reading frame encoding the antigen of interest, flanking UTRs, a 5’ cap and a poly-A tail, this results in an mRNA product that resembles naturally occurring processed mRNA molecules found in eukaryotic cell cytoplasm [40]. Exogenous mRNA is immunostimulatory itself as it can be recognised by innate immune receptors such as TLRs 3, 7 and 8 [41]. This is in theory an advantageous feature of using mRNA in vaccine technology as it may provide and adjuvant activity which can drive dendritic cell maturation and therefore elicit strong B and T-cell responses [40].
Proof of concept in poultry has been carried out in vitro using a luciferase encoding saRNA in lipid nanoparticles. Treatment of chicken organoids and cells with the saRNA-luciferase construct resulted in bioluminescent signals detectable in in vitro transfected chicken tracheal and cecal cells showing that saRNA can effectively enter and replicate in poultry cells [42]. An in vivo study has since been carried out in SPF chickens against AIV. This study showed that an mRNA-lipid nanoparticle vaccine was safe in both embryos and chicks. Furthermore, birds inoculated with the mRNA based AIV vaccine produced significantly higher antigen specific antibody titres, increased IFN-γ responses and decreased viral load in organs when challenged [43]. A significant issue for the use of mRNA vaccine technology is the cost associated with vaccine production and storage. mRNA vaccines typically require cold chain storage which is not practical in field settings.
2.7 Recombinant Viral Vectored Vaccines:
There are several recombinant viral vectors commonly used to produce poultry vaccines. These are turkey herpesvirus (HVT), fowlpox virus (FPV), adenovirus, infectious laryngotracheitis virus (ILTV), and MDV. The most commonly used of these are FPV and HVT as these are phenotypically stable, rarely transmitted horizontally, and do not revert to virulence [3,44]. HVT-based vaccines appear to confer a higher efficacy as they are not as affected by MDAs compared with FPV based vaccines [45,46]. Recombinant viral vectored vaccines can be administered in ovo or subcutaneous injection at one day old [47,48] and can also be modified to contain more than one type of antigen which broadens the protective efficacy [49].
There are several licensed recombinant viral vector vaccines against NDV (Table 1). Originally these were based on an FPV vector but more recently HVT vectors expressing the fusion or haemagglutinin-neuraminidase NDV proteins have been used. NDV itself can also be used as a virus vector with the advantages that it can be used to create bivalent vaccines. Like HVT and FPV, NDV is a natural pathogen of birds ensuring the delivery of the antigen to the target tissues eliciting both cellular and humoral immunity [12,50,51].
2.8. Nanotechnology:
Nanoparticles are defined as materials with a dimension of less than 100 nm [52]. Nanoparticles can be used as delivery systems for vaccines where the vaccine antigen can be incorporated on the surface or encapsulated within the surface of a nanoparticle. When encapsulated within the nanoparticle, the antigens can also be stabilised for uptake and processing by immune cellular system, which might otherwise be rapidly degraded by host cell proteases upon administration. [53]. The main targets for vaccination of poultry using nanoparticles technology are NDV and AIV, and vaccination against E-coli and Salmonella has also been explored [54].
Nanoparticles have therefore been found to increase the potency of vaccines. An Ag@SiO2 nanoparticle encapsulated DNA vaccine has been tested against NDV in poultry. This method of vaccination resulted in both a cell based and humoral immune response. Increased levels of serum antibody, IL-2 and IFN-γ secretion as compared to the standard DNA vaccine alone were observed when administered intranasally. This study showed that the Ag@SiO2 hollow nanoparticle is a safe and effective delivery method for NDV DNA vaccines and can also induce mucosal immunity [55].
Chitosan is a non-toxic biodegradable natural polysaccharide derived from crustacean and insect exoskeletons. Mice vaccinated with chitosan containing vaccine elicited increased levels of antigen specific antibody, antigen specific splenic CD4+ T-cells and strong delayed hypersensitivity responses [56]. In poultry, chitosan and calcium phosphate particles have both been used in the study of an NDV vaccine. Nanoparticle-based vaccines result in high antibody titres in both mucosa and blood compared to standard inactivated NDV vaccines. When challenged with a lethal dose of NDV, the chitosan-based vaccine induced better protection than the calcium phosphate-based vaccine. [57]. An oral chitosan nanoparticle vaccine was also shown to induce both cellular and humoral immunity towards Salmonella when delivered in feed and drinking water. This vaccine contained both the highly immunogenic Salmonella outer membrane proteins (OMPs) together with the flagellin (F) surface protein which acts as TLR5 antagonist in the intestines of birds. This resulted in the induction of both a cell mediated and humoral response in vaccinated birds. Analysis of the immune response post-vaccination showed increased expression of other TLRs, Th1 and Th2 cytokines in chicken immune cells, increased IgA and IgY responses and reduced pathogen load upon live bacterial challenge [58].
Altogether these studies show that nanoparticles can support both cellular and humoral protective immune responses in poultry and may allow increased stability of the vaccine to allow for administration in food and drinking water. In addition, nanotechnology is a promising field for increasing the potency of poultry vaccines as some nanoparticles can also provide and adjuvant effect (for further information see “4. The use of adjuvants in poultry vaccines) and can be used as carriers for many other novel vaccine technologies such as VLPs and liposomes for the encapsulation of mRNA vaccine technology.
2.9 Conclusions:
In conclusion, there are several platforms available to be used in poultry vaccine technology however, most commercially available vaccines use live based vaccines which are then attenuated or inactivated (Table 1). This is most likely due to the ability for live vaccines to induce robust immune responses and can often be used in combination allowing for protection against several diseases and or strains. The production of live attenuated and inactivated vaccines is also a well-established process commercially. This makes switching to new technology harder as the scale-up of manufacturing will take time and money. This may make poultry vaccines more expensive which may impact their field use.
3. Delivery Mechanisms
Effective poultry vaccination has become a challenge around the world due to the increase in demand for poultry meat and eggs leading to more and larger flock sizes. Vaccination has become more popular to carry out in the hatchery where mass vaccination can be administered in ovo at embryo day 18-19, or by spray on the day of hatching. Vaccines can also be administered individually on the day of hatch via intramuscular or subcutaneous injection and intraocular/nasally (Figure 1). Once at the farm, further vaccination can occur via spray, drinking water application, wing web as well as intramuscular or subcutaneous injection (Figure 1). Although individual vaccination of animals is thought to be more effective as all birds are vaccinated with the same known amount of antigen, it is becoming impractical due to the number of commercial chickens worldwide.
3.1 In ovo:
In ovo vaccination can be carried out via the puncturing of a small hole through the blunt end of an egg. A needle is then used to deliver the vaccine to the amniotic cavity, this technique was developed in the 1980s [59]. Today, most of the commercial egg injection machines have the ability to vaccinate either the amniotic cavity or embryo body. Vaccination does not affect the hatchability or the performance of hatched chicks. The chicks that hath from vaccinated eggs have protective immunity from hatch. In ovo vaccination is common in U.S. broiler hatcheries and its popularity is expanding around the world [59]. Vaccines administered in ovo is include MDV, IBD, HVT, ILT, NDV and coccidiosis (Table 1) [59,60]. Although still in experimental stages vaccines against AIV and Mycoplasma gallisepticum were effective when administered in ovo [61,62].
The mechanism of protection elicited from in ovo vaccination is not fully understood but both the innate and adaptive immune system are thought to be involved. Nevertheless, there are several factors that influence the efficacy of in ovo vaccination. These include site of injection, embryo development stage, aseptic control measures in hatchery, including the vaccination equipment and vaccine preparation [60]. In ovo vaccination occurs at embryonic incubation day 18, when all functional components of the immune system are developed in the chicken allowing a sufficient immune response to be generated [59]. Vaccination of embryos via in ovo injection has been proven to be safe and effective with minimal adverse effects on hatchability. The site of injection is of particular importance as when MDV vaccine was administered to the allantoic sack the vaccine was not effective, while delivery to the embryo body or amnion resulted in protection against MDV disease [63]. Determining the appropriate volume that is used in in ovo vaccination is also important with suggested volumes not exceeding 2000 μl for electrolyte solution injection and 700 μl for carbohydrate solution injections. The commercially available MDV vaccine is administered in 50 μl of solution [60].
3.2 Spray:
On the day of hatch live attenuated IBV and NDV, avian metapneumovirus and coccidiosis vaccines (Table 1) are administered as a coarse spray (droplets between 70-150 μM in hatcheries and 100-150 μM in houses) as standard procedure in hatcheries and poultry houses. Vaccines are usually diluted in distilled water to prevent any reactions with chlorine or natural salts in the water supply degrading the vaccine. Per application, spray equipment delivers 7-20 ml of diluted vaccine per 100 chicks [64]. Spray vaccination has lowered the costs associated with time and labour and is an efficient and effective method of vaccination. The size of droplets means they land predominantly in eyes and nostrils and are confined to the upper airway. The prevention of penetration in the lower respiratory tract by the vaccine virus reduces potential vaccine associated adverse effects. [9].
Vaccines administered via coarse spray result in detectable immune responses from three days post-vaccination with protective antibody titres present around 2-3 weeks post- vaccination [64].
3.3 Drinking Water:
Vaccinating via drinking water also allows for application to many birds in a short space of time. It can be used to vaccinate several poultry diseases including avian encephalomyelitis virus, chicken anaemia virus, coccidia and E. coli (Table 1). To achieve successful vaccination chemical and disinfectant residues need to be removed from the water line at least 2-days prior to vaccination, along with any other water treatment or medication [65]. To attain successful vaccination, birds are subjected to limited water availability two hours prior to the application of vaccination to drinking water in order to increase their thirst and thus likelihood of consuming the vaccine. In addition, the water containing the vaccine needs to be available for 2-4 hours to ensure all birds have a chance to consume it [64].
3.4 Subcutaneous/Intramuscular:
A subset of vaccines are recommended to be administered subcutaneously immediately after hatch. This was first introduced using live attenuated MDV and is still commonly used as a hatchery vaccine in layer birds along with FPV and reovirus vaccines (Table 1) [9,64]. Inactivated and bacterial oil emulsion vaccines are commonly administered this way in several poultry species including turkey breeder pullets, broiler breeder and layers before the onset of egg production (Table 1). In breeders, the use of inactivated vaccines is essential for the vertical transmission of antibodies to chicks for early protection, especially for protection against immunosuppressive viruses [66]. The individual handling of chickens for subcutaneous administration is, however, labour intensive but have proven effective for NDV and IBD vaccination [9].
3.5 Intraocular/Nasal Drop:
Intraocular or intranasal vaccination are delivered using a manufacture-provided dropper that delivers approximately 0.03 ml of vaccine solutions to the desired site (Figure 1). Dye can be added to the vaccine formulation to help determine which birds have been vaccinated in a given session. This method can present issues regarding cold chain storage as the vaccine can be warmed by the hand of the vaccinator decreasing vaccine efficacy. Although intraocular/nasal vaccination was popular in the early stages of large-scale poultry production it has gradually been replaced by the use of drinking water and spray vaccination. However, vaccination against NDV, IBV and avian rhinotracheitis can still administered using this technique (Table 1) [64].
3.6 Wing Web:
Wing web vaccination is commonly used in chickens and turkeys against FPV, avian encephalomyelitis, chicken infectious anaemia virus and Pasteurella, although not currently used in the United Kingdom (UK) (Table 1), wing web inoculation is used in other countries such as the USA, Vietnam and India (https://bi-animalhealth.com/poultry/all-poultry-vaccines, https://www.avac.com.vn/en/products-for-poultry/sanphamchogiacam/, https://ahd.maharashtra.gov.in/en/fpv). Here, the vaccine is applied to the wing web using an individual stabber with steel prongs containing small holes which hold the vaccine. Prongs must be submerged in the vaccine before each application (Figure 1). Seven days after vaccination the formation of small nodule scabs that can be palpated at the site of inoculation are used to confirm successful vaccination, however, protection may vary depending on the breed of bird and vaccine applied [64].
4. The Use of Adjuvants in Poultry Vaccines:
Vaccine adjuvants are used to enhance the immune response to vaccines with a lower dose per animal or to avoid the necessity for booster vaccinations. Adjuvants are often necessary to stimulate and direct both the innate and adaptive immune responses to otherwise poorly immunogenic vaccine antigens and can have more than one mechanism of action [13]. There are several adjuvants used in commercial and experimental poultry vaccines which can be categorised into two groups: immunostimulant and delivery agents. Immunostimulants include toll like receptors (TLRs), saponins and cytokines, these promote the secretion of pro inflammatory cytokines which simulate the antigen presenting cells (APCs). Delivery agents, on the other hand, help preserve the antigen conformation for correct presentation to APCs and can also provide slow release for continuous stimulation of the immune system [67].
4.1 Emulsions (Delivery Agent and Immunostimulant):
Emulsions have been utilised as adjuvant systems in animal vaccines for many years and are used frequently as they show good efficacy in the production of antibodies, are cost effective and easy to produce [67,68]. Using emulsions as an adjuvant allows the extended release of antigen and enhanced antibody production resulting in an overall increased immune response of the birds to the antigen [13]. Emulsions are formed when two immiscible liquids which when combined, form small droplets of one liquid dispersed within the other. There are three types of emulsions used in vaccine formulations: water in oil (W/O), oil in water (O/W) and water in oil in water (W/O/W) (Figure 2) [67].
W/O emulsions involves the dispersion of water droplets in a continuous oil phase. Here antigens are captured in the water phase surrounded by a continuous oil phase. Therefore, the antigens are slowly released upon the breakdown of the oil after injection (Figure. 2a). This preserves the antigen from fast clearance by the host therefore increasing the time allowed for immune cell recruitment and antigen processing [68,69]. This is probably the most well-known type of emulsion adjuvant and the prototype is known as Freund’s adjuvant, created in 1937. Freund’s adjuvant is based on a paraffin oil with heat killed and dried mycobacteria (complete) or without mycobacteria (incomplete) and was found to be efficient at inducing high antibody titre [67,70]. However, Freund’s adjuvant also resulted in high levels of adverse effects so is no longer used. More successful commercial W/O emulsions under the product name Montanide™ Incomplete SEPPIC Adjuvants (ISA), have been developed with lower adverse effects but comparable increase in efficacy and are used in veterinary vaccines [67,71]. For example, W/O Montanide™ ISA has been successfully used as an adjuvant in vaccines against NDV and coccidiosis in chickens. The use of Montanide™ ISA as an adjuvant resulted in increased antibody titres against NDV compared to standard W/O emulsions as well as 100% protection when challenged [72]. Montanide™ ISA was used in the preparation of vaccines against Eimeria acervuline, one the causative agents of avian coccidiosis disease. When challenged, Montanide™ ISA adjuvanted vaccines resulted in increased weight gain and reduced parasite shedding compared to non-adjuvanted vaccines [73].
O/W emulsions are formed by the dispersion of oil droplets in the aqueous phase where the antigen is contained (Figure. 2b). The oil droplets help chemokine-driven immune cell recruitment and differentiation of dendritic cells and macrophages. There are several commercially available O/W adjuvants developed by Montanide™ ISA and Emulsigen ™. O/W adjuvants have been successfully used against equine and swine influenza suggesting they can be used as adjuvants in vaccines against viruses in animals but have not been tested in poultry [67]. O/W emulsion have been used in humans and veterinary vaccines. In humans, a subunit O/W MF59-adjuvanted vaccines against influenza H5N1 virus was shown to induce antibodies that are also cross reactive in conjunction with antigen dose sparing [74,75]. In cattle, swine and guineapigs, vaccines against foot-and-mouth disease were adjuvanted with commercially available O/W emulsion. These vaccines were more tolerant to long term storage and antibody levels were detectable for at least 6 months following vaccination [76].
In W/O/W emulsions, oil droplets containing internal water droplets are dispersed in a continuous water phase (Figure. 2c). This has been suggested to provide a fast release of antigens from the external water phase coupled with the prolonged release due to the internal water droplets. Therefore W/O/W may be able to facilitate both the quick and continuous stimulation of immune cells, however, few W/O/W adjuvants are available on the market, these are Montanide™ ISA 201 VG and 206 VG [67]. In poultry, vaccines against Borrelia anserina, the bacteria responsible for fowl spirochaetosis, have been studied using the commercially available Montanide™ ISA 206 VG emulsion. Chickens vaccinated with Montanide™ ISA 206 VG adjuvanted vaccines produced significantly higher antibody titres up to 42 days post vaccination whereas the non-adjuvanted vaccine saw a decrease in antibody titre from 28 days post vaccination [77]. This suggests that W/O/W emulsions can be used in poultry vaccine to increase efficacy.
4.2 Toll Like Receptors (TLRs) (Immunostimulatory):
Toll like receptors (TLRs) are membrane anchored receptors that are expressed in macrophages and dendritic cells and initiate host defence mechanisms upon recognising specific components of microbes. Once TLRs are activated they have an important role in the proinflammatory response and contribute to the production of antigen-specific immunity by inducing the expression of cytokines involved in T-cell differentiation and so induce cell mediated responses which in turn supports the humoral immune response [67,78]. There are 10 known TLRs in chickens and the use of TLRs as adjuvants in vaccines has shown promising results in several animal models including poultry (Figure 3) [78,79].
Formalin-inactivated H5N2 vaccine adjuvanted with the TLR5 ligand Salmonella flagellin resulted in enhanced titres of AIV specific IgA antibody in vaccinated birds compared to the inactivated H5N2 vaccine alone. The increase in IgA antibodies makes this adjuvant of particular interest for use against pathogens that infect via mucosal tissues [80]. Ligands targeted by CpG oligodinucleotides (CpG-ODN) (TLR21) and Bacillus subtilis spores were also shown to enhance host immunity against AIV with increased levers of specific IgA seen in the respiratory tract and serum IgG. Intranasal administration also resulted in increased expression of proinflammatory cytokines in the trachea and nasal cavity [81]. Polyinosinic-polycytidylic acid (poly I:C) induces a TLR3 response as poly I:C acts as a dsRNA analogue that can induce a type I IFN response. Poly I:C has been added to chicken embryo cells before exposure the NDV and avian reovirus [82]. Poly I:C treatment reduced plaque formation and induced an antiviral state against both viruses. Also, in NDV vaccines CpG use increased serum IgG and IgA and T-cell proliferation when NDV vaccines were administered intranasally showing it can be used as an adjuvant to stimulate mucosal immunity [83]. The use of TLR ligand adjuvanted vaccines has also been trialled with bacterial vaccines in poultry. Used in vaccines against Salmonella enteritidis CpG-ODN, which targets TLR21, increased responses in chicken innate immune cells and resulted in a significant reduction in organ invasion by Salmonella enteritidis and subsequent associated mortality in challenge studies [84].
TLR adjuvants can reduce AIV shedding after prophylactic treatment with different TLRs, however, this response varied depending on the ligand and route of administration used [85]. An alternative method to deliver TLR ligands as vaccine adjuvants is by encoding ligand sequences within a plasmid. CpG encoded plasmids administered with the inactivated H5N2 vaccine in chickens induced significantly higher levels of IFN-y responses, and splenocytes expressed increased levels of TLR3 and TLR7 transcripts [86]. Encoding TLR adjuvants into plasmids offers a more cost effective and large-scale production opportunity [87].
Therefore, TLRs can stimulate both a cell mediated and humoral response increasing the potency of vaccines in poultry in experimental trials. However, concerns have been raised about the induction of unwanted side-effects especially the induction of a strong inflammatory response when using TLRs as adjuvants in vaccines [78].
4.3 Cytokines as Adjuvants (Immunostimulatory):
The induction of cytokines is a key mechanism of action for adjuvants thus, the direct administration of cytokines could be used as an adjuvant to enhance vaccine responses. Additionally, cytokines could be used to directly bias host immune responses such as towards Th1 or Th2 cells. Methods for the administration of cytokines include encapsulation with liposomes, biodegradable polymers, conjugation with vaccine antigens, and cloning into genetic vaccine platforms. Several chicken cytokines have already been recognised to have antiviral and immunomodulatory properties [87].
Cytokines such as IFN-α, β and γ as well as IL-1β have been studied as vaccine adjuvants in chickens. The combined administration of chicken IFN-α, γ and IL-1β expressed in recombinant E-coli induced an increased antibody response against the tetanus toxin however, there was no improvement in antibody response when recombinant chicken IFNs were co-administered with inactivated IBV vaccines [88].
Chicken INF-α has been widely studied as an immunomodulator and adjuvant in poultry vaccines [82]. Chicken INF-α produced in a prokaryotic expression system was evaluated in combination with the inactivated H9N2 AIV vaccine. This resulted in increased Th1 and antibody responses as compared with the H9N2 vaccine alone [89]. Birds vaccinated with chicken IFN-α co-expressed with NDV in a FPV vector did not lose weight when challenged compared to birds that received the NDV vaccine alone. This also suggests that chicken INF-α treatment prevents disease due to its antiviral properties whilst still maintaining its vaccination potential as protective responses were still observed up to 60 days post vaccination when challenged [90].
IL-2 plays a significant role in the activation of T-cells and NK cells. Adding recombinant IL-2 to an inactivated H5N2 AIV vaccine resulted in increased levels of IgG and IgA and increased levels of mast cells in the respiratory tract of chicks when administered intranasally [91]. H3N2 AIV that contained embedded IL-2 was used to generate an inactivated AIV vaccine where virus particles also expressed surface IL-2. This vaccine induced increased systemic antibody levels compared to a conventional inactivated H3N2 AIV vaccine [92].
Plasmids expressing IL-15 have been used as genetic adjuvants in combination with a H5 AIV DNA based vaccine. H5 AIV DNA vaccine containing IL-15 plasmid resulted in increased blood CD8+ T-cells and increased serum HI titres when administered at 14-days old compared to H5 AIV plasmid alone. Immune responses were seen in 1-day old vaccinated chicks though these were considerably lower. This shows that an IL-15 adjuvant increases both antibody and cell mediated responses, which may result in a potential increase in the protective effect against AIV infection [93]. Together this shows that cytokines have potential as adjuvants in poultry vaccines.
5. Targeting the Antigen to Antigen Presenting Cells:
Antigen presenting cells (APCs) are a crucial part of the innate immune system and play an essential role in initiating the adaptive immune response. APCs, such as dendritic cells (DCs), can induce the activation and clonal expansion of T-cells therefore, targeting APCs via vaccination can help induce effective and long-term T-cell based immune responses. The uptake of antigens by APCs using antigen targeting has been developed for human use to increase potency of vaccines and immunotherapy treatments over the last two decades especially to tackle cancer and autoimmune conditions [94].
There is a large choice in methods of targeting antigen presenting cells which result in the stimulation of different immune responses and therefore affect vaccine induced protection. Factors affecting the immune response to APC targeted vaccines include the choice of antigen, receptor, adjuvant, carrier, and administration route. Different combinations of these factors allow for the customisation of vaccines for protection in different species and against different pathogens [94]. Targeting antigens to APCs using a ligand or antibody-based approach has been shown to increase vaccine efficacy in mammals and birds, with five validated APC targets in birds (Figure 4).
5.1 Ligand Based Targeting of APCs:
APCs express pattern recognition receptors (PRRs) on their surface and intracellular compartments which bind to natural ligands commonly expresses on microbes e.g., flagellin in bacteria, single stranded RNA in viruses, also known as pathogen associated molecular patterns (PAMPs). The conjugation of antigen to a PRR ligand results in simultaneous antigen processing and stimulation to occur in the same DC [95]. In ligand-based targeting vaccines, different PRRs can be selected to increase the desired immune response. These responses can include the stimulation of different families of T-cells, or site-specific immune responses in disease specific organs e.g., increase in mucosal immunity. [8,95,96].
In ligand-based APC targeting vaccines, the antigen is conjugated to a specific PRR which results in an antigen-TLR ligand conjugate vaccine. This is then recognised by the associated TLR on an APC and ensures antigen is processed and then presented via the APC (Figure 5). This increases vaccine potency as it increases antigen presentation and cellular stimulation resulting in improved associated humoral and cellular immunity [8]
There are four PRR families, TLRs, retinoic acid-inducible gene 1-like helicase receptors (RLRs), oligomerisation domain like receptors (NLRs), and C-type lectin receptors (CLRs). TLRs are the most studied in antigen-PRR ligand conjugate vaccines in mammals [95]. In poultry, TLR ligands have been studied and used as adjuvants in vaccine development and have shown to be successful. Therefore, these are promising candidates for ligands to use as APC targets (See 4.2 Toll Like Receptors (TLRs)) [8].
5.2 Antibody Based Targeting of APCs:
Antibody targeted vaccines use the targeting properties of antibodies to deliver cargo, in this case antigens, to APCs. This method has been applied to a wide diversity of antigens, as it can accommodate entire proteins or smaller disease specific peptides. Using antibodies targeted to specific APC receptors conjugated to antigen decreases the chance of non-specific antigen uptake by non-immune cells and thus increasing the amount of antigen reaching the APC [8]. Previously, antigens have been chemically conjugated to APC binding antibodies but recently this can be achieved genetically via direct Fab-fragment linkage, including in the tail of the C-terminus of the antibody heavy chain or included in the loops of the constant domain with the APC-specific V-regions on the recombinant antibody like molecule [20]. Antibody based targeting of APCs can also be carried out by the conjugation of antigens to specific monoclonal antibodies (mAbs) selected for APC surface molecules or via genetic engineering. In this case, the antigen is fused to antibody fragments e.g., scFv or fragment antigen binding regions (Fabs), for specific APC receptors (Figure 6) [8]. Most research into to antibody-based targeting of APC vaccines has focused primarily on receptors such as C-type lectin receptors (CLRs), integrins and Fc receptors. These receptors can also be found on avian APCs with CD11c present on immature DCs, a member of the β2 integrin family, DEC205 present on mature DCs and is a CLR [8] and CD83 a member of the immunoglobulin superfamily and therefore has potential to act as an Fc receptor as all known Fc receptors also belong to this family [97,98].
DEC205 and CD11c have been used in antigen-scFv vaccines against AIV in poultry. This method targets avian DCs and resulted in faster and higher neutralising antibody production compared to standard non-targeted vaccines. However, CD11c targeted vaccines induced increased levels of proinflammatory cytokines whereas DEC205 targeted vaccines did not. [99]. The ScFv technology targeting chicken CD83 with H9 antigen was also found to induce high levels of antibodies in chicks and vaccination was not affected by the presence of MDAs [100]. This method of vaccination also provided protection when vaccinated animals were challenged in studies with H9N2 virus [101].
In addition, mAb designed to mimic CD40L which targets CD40 has also been trialled in poultry. CD40 is expressed by chicken macrophages, DCs, B-cells and monocytes. The use of a monoclonal antibody targeting CD40 has also been previously shown to increase antigen specific IgY responses. When administered orally via the GI tract, significant IgA titres were induced in tracheal mucosa from only one dose of vaccine [102]. Using chicken CD40L has also been studied in poultry vaccination. Eimeria tenalla (E. tenalla) causes avian coccidiosis and is usually controlled by drug treatment, however, resistant strains are occurring. E. tenalla immune mapped protein 1 (EtIMP1) was shown to be immunogenic. Therefore, a recombinant EtIMP1-CD40L fusion protein was created and used to immunise chickens and induced stronger IgA and IFN-γ responses than EtIMP1 protein alone [103], demonstrating that the CD40/CD40L family can be used effectively for APC targeted vaccines in poultry.
An APC targeted multivalent vaccine has been developed against type 1 and type 2 Bovine Viral Diarrhoea Virus. This vaccine contained a single chain antibody directed to an MHC class II antigen epitope which proved effective in challenge studies in reducing clinical signs of disease and decrease in detection of virus in vaccinated calves [104]. Therefore, the ability to develop multivalent vaccines using APC technology in veterinary species is possible but has not been trialled in poultry [8,104].
Altogether these studies show that APC targeting vaccines can be administered orally which may help to stimulate immune responses in disease specific tissues. However, the choice of receptor that is being targeted needs to be considered as these can result in different degrees of immune response and therefore may affect their efficacy.
6. Probiotics:
Probiotics are live microorganisms which confer a health benefit through improvements of the intestinal microbial balance when adequate amounts are administered. The mode of action of probiotics is not fully understood but has been shown to be beneficial in both humans and farm animals, including chickens [105,106,107].
The gastrointestinal tract (GIT) microbiota plays an important role in the pathogenesis of viruses. A large number of viral infections occur via mucosal tissues, such as the GIT and respiratory system, where microbes encounter commensal microbiota on the surfaces of the tissues. GIT microbiota is essential for maintaining basal levels of type 1 IFN levels which is a critical antiviral response, and it has also been shown that the GIT microbiota help the host immune responses to different pathogens [87,108]. In poultry the innate immune response to antigens in the GIT is mediated via the caecal tonsils and Peyer’s patches, also referred to as GIT-associated lymphoid tissues (GALTS) (Casteleyn et al., 2010). The mechanisms of GIT microbiota mediated immunity involves PRRs, TLRs and nod like receptors (NLRs) [87].
Lactic acid producing bacteria are already used in the meat and dairy industries; these are Bifidobacterium, Lactococcus and Lactobacillus, some strains of which have probiotic properties. Via mechanisms currently unknown, lactic acid producing bacteria confer improved immune responses as well as other benefits to the host in several species including chickens [87,105,107]. The effect of lactic acid-producing bacteria has also been shown reduce enteric diseases in farms by competitive exclusion mechanisms against pathogenic bacteria, and by the production of molecules with bactericidal functions [110]. The importance of the chicken GIT microbiome and its effect on the development of immune responses to viruses is becoming increasingly evident. Interactions between lactobacilli and chicken macrophages have been shown to induce anti-AIV responses including increase in Nitric oxide (NO) and IFN-γ production as well as the expression of the co-stimulatory molecule CD40 [111]. However, probiotic treatment needs to be tailored to the type of vaccination given. It has been shown that Lactobacillus spp. enhanced IFN-γ production and increased HAI titres but only in chickens vaccinated with CpG adjuvanted whole inactivated H9N2 vaccine, while there was no effect on the immune response in birds vaccinated with non-adjuvanted whole inactivated H9N2 vaccine [112]. This suggests that the mechanisms behind the effects of probiotic treatments still need to be elucidated.
Bacillus subtilis (B. subtilis) has also been investigated as a mechanism to improve immune responses to AIV vaccination. B. subtilis had previously been used as a probiotic treatment in humans and other animal models. The adjuvant effect of B. subtilis was investigated on an inactivated H9N2 AIV vaccine in chickens. B. subtilis increased the number of B-cells and innate cells in splenocytes as well as expression of proinflammatory cytokines such as IL-6 and IL-1β. Restimulation of splenocytes also showed that the adjuvanted vaccine increased the levels of H9N2 specific CD4+ and CD8+ T-cell proliferation in chickens as compared to H9N2 vaccination alone [113].
Vaccination responses against MDV with the HVT vector were also increased with the use of probiotics. The administration of four Lactobacillus species on embryonic day 18 and chicks on days 1 to 4 post hatch increased the expression of MHC-II on spleen macrophages and B-cells and decreased spleen T-regulatory (Treg) cells. Levels of IFN-α and IFN-β were also increased in probiotic treated chicks as well as reduced tumour incidence [114].
Commercially available pre- and/or postbiotic treatments have also been assessed for their effect on the live attenuated Salmonella Enterititidis vaccine. In this study the protection of broiler chickens from challenge with Salmonella Enterititidis was investigated after pre- and/or probiotics were administered via drinking water. This study found that the treatment of pre- or postbiotics at the same time as the live Salmonella vaccine reduced mortality rates, vaccine associated negative growth performance, faecal shedding, and ability to detect the bacteria in chicken organs such as heart, liver spleen and cecum [115].
The method of probiotic administration needs to be considered as there is opposing findings in the literature. One group showed that probiotics given via feed induced higher HI anti-AIV antibodies compared to when given in drinking water [116], whereas a different experiment included probiotics within the vaccination formula itself and showed that this method increased the potency of AIV vaccination [113] therefore, the most effective method of probiotic administration to increase vaccine potency still needs to be optimised. There is some evidence of the improvement of vaccine responses in poultry in response to probiotic administration however, probiotic usage still needs further study.
7. Enhancement of Vaccine Efficacy by the Incorporation of Multiple Antigens:
In poultry, there are several multivalent vaccines already in use (Table 1). These include tetravalent vaccines e.g. Avian pneumovirus, IBV and NDV (Table 1 - VM 08327/3020) quadrivalent vaccines e.g. avian pneumovirus, EDS, IBV and NDV (Table 1 – VM 08327/5025) and penta- and octavalent vaccines against different strains of Eimeria parasites (Table 1 VM 01708/5101, 01708/4572). Multivalent vaccines are preferred in the commercial poultry industry as they reduce time and labour costs and stress caused by manual handling during the vaccination process. However, in multivalent vaccines, potency can be affected by the inclusion of multiple antigens. Interactions between responses to different antigens and the components that make up the multivalent vaccine. These could lead to inappropriate immune responses due to epitope suppression and/or antigenic competition. The number of antigens and potency therefore needs to be balanced to develop an effective multivalent vaccine [117,118].
mRNA technology has allowed for the development of a multivalent nucleoside-modified mRNA lipid nanoparticle vaccine encoding mRNA for 20 independent HA antigens of influenza A (18) and influenza B (2) subtypes. This multivalent vaccine elicited high levels of antibodies that were cross-reactive, providing protection from matched and mismatched influenza challenge in both mice and ferrets [119]. A single H9 HA mRNA vaccine has been trialled in chickens and shown to be safe and effective therefore there is the potential that a similar multivalent mRNA AIV vaccine could be trialled and prove effective in poultry [43].
8. Conclusions:
There is a need to increase the potency of avian vaccines as vaccination is an essential part of the toolkit to decrease infection and transmission of disease in poultry. A large amount of research has been carried out investigating methods to increase the immune response to vaccination against several avian diseases, however progress towards the use of these novel formulations commercially is limited. Further research into the long-term protection elicited by these new technologies also needs to be determined in the field, especially for the vaccination of layers and breeder flocks.
Improvements have been made in traditional adjuvants such as in adjuvants based on oil in water emulsion, which have shown to increase vaccine potency and are one of the most used adjuvants in commercial poultry vaccines. These are well tolerated and induce increased immune responses compared their nonadjuvanted counterparts.
However, progress towards the more regular use of novel adjuvants in commercial poultry vaccines is very limited. This is most likely due to the need to optimise and standardise not only the delivery mechanisms but the manufacturing procedures. There is still a worry that novel adjuvants may produce unacceptable side effects in the vaccinated birds, this is especially a concern with the use of immunomodulatory adjuvants, such as cytokines and TLRs.
Another issue associated with the wide use of novel technologies is the lack of infrastructure that is needed to scale up the production of new vaccine technologies and adjuvants if the technology is going to be used widely at a low cost. Increased infrastructure for the production of vaccines and adjuvants using novel technologies will be needed in order to maintain a stable vaccine supply and decrease the overall vaccine cost making it commercially viable for use in poultry.
Author Contributions
O.B.H. wrote the original draft and reviewed and edited the manuscript. M.I. and C.R. contributed to concept development, designed the structure of the review, edited figures, reviewed and contributed to the scientific content, supervised, and proofread.
Funding
This work was funded by BBSRC (grant numbers BBS/E/PI/230001C, BBS/E/PI/23NB0003, BB/T013087/1, BB/W003325/1) and the GCRF One Health Poultry Hub (grant number BB/S011269/1). O.B.H. is funded by the Pathogens and Host Defences Doctoral Training Partnership. C.R. is a Jenner Investigator and is supported by the Equal Opportunities Foundation (Hong Kong), the Braithwaite Family Foundation, and the Bill & Melinda Gates Foundation.
Acknowledgments
O.B.H. would like to acknowledge the encouragement and support of all members of the Avian Influenza and Newcastle Disease group at the Pirbright Institute for writing this review.
Conflicts of Interest
CR has performed paid consultancy for Guidepoint. CR is a contributor to the intellectual property licenced by Oxford University Innovation to AstraZeneca limited to COVID-19 applications. The other authors declare no conflicts of interest.
References
- FAO. MEAT MARKET REVIEW: Emerging Trends and Outlook 2022. Available online: https://www.fao.org/markets-and-trade/publications/detail/en/c/1620239/ (accessed on 19 December 2023).
- OECD-FAO Agricultural Outlook 2020-2029; OECD-FAO Agricultural Outlook; OECD, 2020; ISBN 9789264317673.
- Ravikumar, R.; Chan, J.; Prabakaran, M. Vaccines against Major Poultry Viral Diseases: Strategies to Improve the Breadth and Protective Efficacy. Viruses 2022, 14, 1195. [Google Scholar] [CrossRef] [PubMed]
- Charkhkar, S.; Bashizade, M.; Sotoudehnejad, M.; Ghodrati, M.; Bulbuli, F.; Akbarein, H. The Evaluation and Importance of Newcastle Disease’s Economic Loss in Commercial Layer Poultry. Journal of Poultry Sciences and Avian Diseases 2024, 2, 1–4. [Google Scholar] [CrossRef]
- Scharff, R.L. Food Attribution and Economic Cost Estimates for Meat- And Poultry-Related Illnesses. J Food Prot 2020, 83, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Dejyong, T.; Chanachai, K.; Prarakamawongsa, T.; Kongkaew, W.; Thiptara, A.; Songserm, T.; Rukkwamsuk, T.; TagoPacheco, D.; Phimpraphai, W. Economic and Value Chain Analysis to Support an Investigation and Risk Mitigation Efforts on Marek’s Disease in Layers in the Southern Part of Thailand. Vet World 2023, 16, 35–45. [Google Scholar] [CrossRef]
- Gashaw, M. A Review on Avian Influenza and Its Economic and Public Health Impact. Int J Vet Sci Technol 2020, 4, 15–027. [Google Scholar]
- Shrestha, A.; Sadeyen, J.-R.; Iqbal, M. Enhancing Protective Efficacy of Poultry Vaccines through Targeted Delivery of Antigens to Antigen-Presenting Cells. Vaccines (Basel) 2018, 6, 75. [Google Scholar] [CrossRef]
- Abdul-Cader, M.S.; Palomino-Tapia, V.; Amarasinghe, A.; Ahmed-Hassan, H.; De Silva Senapathi, U.; Abdul-Careem, M.F. Hatchery Vaccination Against Poultry Viral Diseases: Potential Mechanisms and Limitations. Viral Immunol 2018, 31, 23–33. [Google Scholar] [CrossRef]
- Hofacre, C.L.; Singer, R.S.; Johnson, T.J. Antimicrobial Therapy (Including Resistance). In Diseases of Poultry; 2013; pp. 40–43.
- Swayne, D.E. Impact of Vaccines and Vaccination on Global Control of Avian Influenza. Avian Dis 2012, 56, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Romanutti, C.; Keller, L.; Zanetti, F.A. Current Status of Virus-Vectored Vaccines against Pathogens That Affect Poultry. Vaccine 2020, 38, 6990–7001. [Google Scholar] [CrossRef] [PubMed]
- Swayne, D.E.; Kapczynski, D.R. Vaccines and Vaccination for Avian Influenza in Poultry. In Avian Influenza; 2017; pp. 378–434.
- Veterinary Medicines Directorate. Available online: https://www.vmd.defra.gov.uk/productinformationdatabase (accessed on 17 June 2024).
- Swayne, D.E. Avian Influenza Vaccines and Therapies for Poultry. Comp Immunol Microbiol Infect Dis 2009, 32, 351–363. [Google Scholar] [CrossRef]
- Jang, H.; Elaish, M.; Mahesh, K.C.; Abundo, M.C.; Ghorbani, A.; Ngunjiri, J.M.; Lee, C.W. Efficacy and Synergy of Live-Attenuated and Inactivated Influenza Vaccines in Young Chickens. PLoS One 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Brokstad, K.; Cox, R. Influenza Vaccination Strategies: Comparing Inactivated and Live Attenuated Influenza Vaccines. Vaccines (Basel) 2015, 3, 373–389. [Google Scholar] [CrossRef]
- Peek, H.W.; Landman, W.J.M. Coccidiosis in Poultry: Anticoccidial Products, Vaccines and Other Prevention Strategies. Veterinary Quarterly 2011, 31, 143–161. [Google Scholar] [CrossRef]
- Steel, J.; Lowen, A.C.; Pena, L.; Angel, M.; Solórzano, A.; Albrecht, R.; Perez, D.R.; García-Sastre, A.; Palese, P. Live Attenuated Influenza Viruses Containing NS1 Truncations as Vaccine Candidates against H5N1 Highly Pathogenic Avian Influenza. J Virol 2009, 83, 1742–1753. [Google Scholar] [CrossRef]
- Grødeland, G.; Fossum, E.; Bogen, B. Polarizing T and B Cell Responses by APC-Targeted Subunit Vaccines. Front Immunol 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Meunier, M.; Chemaly, M.; Dory, D. DNA Vaccination of Poultry: The Current Status in 2015. Vaccine 2016, 34, 202–211. [Google Scholar] [CrossRef]
- Lauer, K.B.; Borrow, R.; Blanchard, T.J. Multivalent and Multipathogen Viral Vector Vaccines. Clinical and Vaccine Immunology 2017, 24. [Google Scholar] [CrossRef]
- Aida, V.; Pliasas, V.C.; Neasham, P.J.; North, J.F.; McWhorter, K.L.; Glover, S.R.; Kyriakis, C.S. Novel Vaccine Technologies in Veterinary Medicine: A Herald to Human Medicine Vaccines. Front Vet Sci 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Harper, D.M.; Franco, E.L.; Wheeler, C.; Ferris, D.G.; Jenkins, D.; Schuind, A.; Zahaf, T.; Innis, B.; Naud, P.; De Carvalho, N.S.; et al. Efficacy of a Bivalent L1 Virus-like Particle Vaccine in Prevention of Infection with Human Papillomavirus Types 16 and 18 in Young Women: A Randomised Controlled Trial. Lancet 2004, 364, 1757–1765. [Google Scholar] [CrossRef]
- Keating, G.M.; Noble, S. Recombinant Hepatitis B Vaccine (Engerix-B): A Review of Its Immunogenicity and Protective Efficacy against Hepatitis B. Drugs 2003, 63, 1021–1051. [Google Scholar] [CrossRef]
- Zepeda-Cervantes, J.; Ramírez-Jarquín, J.O.; Vaca, L. Interaction Between Virus-Like Particles (VLPs) and Pattern Recognition Receptors (PRRs) From Dendritic Cells (DCs): Toward Better Engineering of VLPs. Front Immunol 2020, 11, 1100. [Google Scholar] [CrossRef] [PubMed]
- Mejía-Méndez, J.L.; Vazquez-Duhalt, R.; Hernández, L.R.; Sánchez-Arreola, E.; Bach, H. Virus-like Particles: Fundamentals and Biomedical Applications. Int J Mol Sci 2022, 23, 8579. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Qian, J.; Qin, L.; Li, J.; Xue, C.; Ding, J.; Wang, W.; Ding, W.; Yin, R.; Jin, N.; et al. Chimeric Newcastle Disease Virus-like Particles Containing DC-Binding Peptide-Fused Haemagglutinin Protect Chickens from Virulent Newcastle Disease Virus and H9N2 Avian Influenza Virus Challenge. Virol Sin 2020, 35, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Firouzamandi, M.; Helan, J.A.; Moeini, H.; Soleimanian, A.; Khatemeh, S.; Hosseini, S.D. Developing a Vaccine against Velogenic Sub-Genotype Seven of Newcastle Disease Virus Based on Virus-like Particles. AMB Express 2023, 13, 114. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.Y.; Park, J.K.; Lee, D.H.; Yuk, S.S.; Kwon, J.H.; Lee, S.W.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Chimeric Bivalent Virus-like Particle Vaccine for H5N1 HPAI and ND Confers Protection Against a Lethal Challenge in Chickens and Allows a Strategy of Differentiating Infected from Vaccinated Animals (DIVA). PLoS One 2016, 11. [Google Scholar] [CrossRef]
- Yan, F.; Zhao, Y.; Hu, Y.; Qiu, J.; Lei, W.; Ji, W.; Li, X.; Wu, Q.; Shi, X.; Li, Z. Protection of Chickens against Infectious Bronchitis Virus with a Multivalent DNA Vaccine and Boosting with an Inactivated Vaccine. J Vet Sci 2013, 14, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Faurez, F.; Dory, D.; Le Moigne, V.; Gravier, R.; Jestin, A. Biosafety of DNA Vaccines: New Generation of DNA Vectors and Current Knowledge on the Fate of Plasmids after Injection. Vaccine 2010, 28, 3888–3895. [Google Scholar] [CrossRef]
- Robinson, H.L.; Hunt, L.A.; Webster, R.G. Protection against a Lethal Influenza Virus Challenge by Immunization with a Haemagglutinin-Expressing Plasmid DNA. Vaccine 1993, 11. [Google Scholar] [CrossRef] [PubMed]
- Bublot, M. Poultry Vaccine Technology Platforms. Avian Dis 2023, 67. [Google Scholar] [CrossRef]
- Jazayeri, S.D.; Poh, C.L. Recent Advances in Delivery of Veterinary DNA Vaccines against Avian Pathogens. Vet Res 2019, 50, 78. [Google Scholar] [CrossRef]
- Martinon, F.; Krishnan, S.; Lenzen, G.; Magné, R.; Gomard, E.; Guillet, J. -G; Lévy, J. -P; Meulien, P. Induction of Virus-specific Cytotoxic T Lymphocytes in Vivo by Liposome-entrapped MRNA. Eur J Immunol 1993, 23. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.; Liu, X.; Li, M.; Zhang, Z.; Song, L.; Zhu, B.; Wu, X.; Liu, J.; Zhao, D.; Li, Y. Advances in COVID-19 MRNA Vaccine Development. Signal Transduct Target Ther 2022, 7, 94. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of Pseudouridine into MRNA Yields Superior Nonimmunogenic Vector with Increased Translational Capacity and Biological Stability. Molecular Therapy 2008, 16, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, K.J.; Webber, M.J.; Anderson, D.G. Materials for Non-Viral Intracellular Delivery of Messenger RNA Therapeutics. Journal of Controlled Release 2016, 240, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. MRNA Vaccines — a New Era in Vaccinology. Nat Rev Drug Discov 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Chen, N.; Xia, P.; Li, S.; Zhang, T.; Wang, T.T.; Zhu, J. RNA Sensors of the Innate Immune System and Their Detection of Pathogens. IUBMB Life 2017, 69, 297–304. [Google Scholar] [CrossRef]
- Snoeck, J.; Chiers, K.; Tam, Y.; Sanders, N.N.; Garmyn, A. Evaluation of a Self-Amplifying MRNA Reporter Vaccine in Explant Models of Broiler Chickens. Poult Sci 2023, 102, 103078. [Google Scholar] [CrossRef]
- Xu, S.; Zhang, B.; Yao, J.; Ruan, W. A New H9 Influenza Virus MRNA Vaccine Elicits Robust Protective Immunity against Infection. Vaccine 2023, 41, 2905–2913. [Google Scholar] [CrossRef]
- Hein, R.; Koopman, R.; García, M.; Armour, N.; Dunn, J.R.; Barbosa, T.; Martinez, A. Review of Poultry Recombinant Vector Vaccines. Avian Dis 2021, 65. [Google Scholar] [CrossRef]
- Swayne, D.E.; Beck, J.R.; Kinney, N. Failure of a Recombinant Fowl Poxvirus Vaccine Containing an Avian Influenza Hemagglutinin Gene to Provide Consistent Protection against Influenza in Chickens Preimmunized with a Fowl Pox Vaccine. Avian Dis 2000, 44, 132. [Google Scholar] [CrossRef]
- Vagnozzi, A.; Zavala, G.; Riblet, S.M.; Mundt, A.; García, M. Protection Induced by Commercially Available Live-Attenuated and Recombinant Viral Vector Vaccines against Infectious Laryngotracheitis Virus in Broiler Chickens. Avian Pathology 2012, 41, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hamid, H.S.; Ellakany, H.F.; Elbestawy, A.R.; Setta, A. The Combined Use of RHVT-H5 and RHVT-F Vector Vaccines in the Hatchery Enhances Immunity against Highly Pathogenic Avian Influenza H5N1 and Velogenic Newcastle Disease Viral Infections in Commercial Chickens. Poultry Science Journal 2018, 6, 165–171. [Google Scholar] [CrossRef]
- Williams, C.J.; Hopkins, B.A. Field Evaluation of the Accuracy of Vaccine Deposition by Two Different Commercially Available in Ovo Injection Systems. Poult Sci 2011, 90, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Gergen, L.; Cook, S.; Ledesma, B.; Cress, W.; Higuchi, D.; Counts, D.; Cruz-Coy, J.; Crouch, C.; Davis, P.; Tarpey, I.; et al. A Double Recombinant Herpes Virus of Turkeys for the Protection of Chickens against Newcastle, Infectious Laryngotracheitis and Marek’s Diseases. Avian Pathology 2019, 48, 45–56. [Google Scholar] [CrossRef]
- Huang, Z.; Elankumaran, S.; Panda, A.; Samal, S. Recombinant Newcastle Disease Virus as a Vaccine Vector. Poult Sci 2003, 82, 899–906. [Google Scholar] [CrossRef]
- Zhao, H.; Peeters, B.P.H. Recombinant Newcastle Disease Virus as a Viral Vector: Effect of Genomic Location of Foreign Gene on Gene Expression and Virus Replication. J Gen Virol 2003, 84, 781–788. [Google Scholar] [CrossRef]
- Lozano, D.; Larraga, V.; Vallet-Regí, M.; Manzano, M. An Overview of the Use of Nanoparticles in Vaccine Development. Nanomaterials 2023, 13, 1828. [Google Scholar] [CrossRef]
- Gregory, A.E.; Titball, R.; Williamson, D. Vaccine Delivery Using Nanoparticles. Front Cell Infect Microbiol 2013, 4. [Google Scholar] [CrossRef]
- Calderon-Nieva, D.; Goonewardene, K.B.; Gomis, S.; Foldvari, M. Veterinary Vaccine Nanotechnology: Pulmonary and Nasal Delivery in Livestock Animals. Drug Deliv Transl Res 2017, 7, 558–570. [Google Scholar] [CrossRef]
- Zhao, K.; Rong, G.; Hao, Y.; Yu, L.; Kang, H.; Wang, X.; Wang, X.; Jin, Z.; Ren, Z.; Li, Z. IgA Response and Protection Following Nasal Vaccination of Chickens with Newcastle Disease Virus DNA Vaccine Nanoencapsulated with Ag@SiO2 Hollow Nanoparticles. Sci Rep 2016, 6, 25720. [Google Scholar] [CrossRef]
- Zaharoff, D.A.; Rogers, C.J.; Hance, K.W.; Schlom, J.; Greiner, J.W. Chitosan Solution Enhances Both Humoral and Cell-Mediated Immune Responses to Subcutaneous Vaccination. Vaccine 2007, 25, 2085–2094. [Google Scholar] [CrossRef] [PubMed]
- Volkova, M.A.; Irza, A. V.; Chvala, I.A.; Frolov, S.F.; Drygin, V. V.; Kapczynski, D.R. Adjuvant Effects of Chitosan and Calcium Phosphate Particles in an Inactivated Newcastle Disease Vaccine. Avian Dis 2014, 58, 46–52. [Google Scholar] [CrossRef]
- Renu, S.; Han, Y.; Dhakal, S.; Lakshmanappa, Y.S.; Ghimire, S.; Feliciano-Ruiz, N.; Senapati, S.; Narasimhan, B.; Selvaraj, R.; Renukaradhya, G.J. Chitosan-Adjuvanted Salmonella Subunit Nanoparticle Vaccine for Poultry Delivered through Drinking Water and Feed. Carbohydr Polym 2020, 243, 116434. [Google Scholar] [CrossRef] [PubMed]
- Schat, K.A.; Kaspers, B.; Kaiser, P. Avian Immunology; Elsevier, 2014; ISBN 9780123969651.
- Peebles, E.D. In Ovo Applications in Poultry: A Review. Poult Sci 2018, 97, 2322–2338. [Google Scholar] [CrossRef]
- Toro, H.; Tang, D.C.; Suarez, D.L.; Sylte, M.J.; Pfeiffer, J.; Van Kampen, K.R. Protective Avian Influenza in Ovo Vaccination with Non-Replicating Human Adenovirus Vector. Vaccine 2007, 25, 2886–2891. [Google Scholar] [CrossRef] [PubMed]
- Elliott, K.E.C.; Branton, S.L.; Evans, J.D.; Gerard, P.D.; Peebles, E.D. Layer Chicken Embryo Survival to Hatch When Administered an in Ovo Vaccination of Strain F Mycoplasma Gallisepticum and Locations of Bacteria Prevalence in the Newly Hatched Chick. Poult Sci 2017, 96, 3879–3884. [Google Scholar] [CrossRef] [PubMed]
- Wakenell, P.S.; Bryan, T.; Schaeffer, J.; Avakian, A.; Williams, C.; Whitfill, C. Effect of in Ovo Vaccine Delivery Route on Herpesvirus of Turkeys/SB-1 Efficacy and Viremia. Avian Dis 2002, 46, 274–280. [Google Scholar] [CrossRef]
- de Wit, J.J.; Montiel, E. Practical Aspects of Poultry Vaccination. In Avian Immunology; Elsevier, 2021; pp. 469–488 ISBN 9780128190715.
- Gentry, R.F.; Braune, M.O. Prevention of Virus Inactivation During Drinking Water Vaccination of Poultry. Poult Sci 1972, 51, 1450–1456. [Google Scholar] [CrossRef]
- Gharaibeh, S.; Mahmoud, K.; Al-Natour, M. Field Evaluation of Maternal Antibody Transfer to a Group of Pathogens in Meat-Type Chickens. Poult Sci 2008, 87, 1550–1555. [Google Scholar] [CrossRef]
- Burakova, Y.; Madera, R.; McVey, S.; Schlup, J.R.; Shi, J. Adjuvants for Animal Vaccines. Viral Immunol 2018, 31, 11–22. [Google Scholar] [CrossRef]
- Cox, J.C.; Coulter, A.R. Adjuvants - A Classification and Review of Their Modes of Action. Vaccine 1997, 15, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Herbert, W.J. The Mode of Action of Mineral-Oil Emulsion Adjuvants on Antibody Production in Mice. Immunology 1968, 14, 301–318. [Google Scholar] [PubMed]
- Freund, J.; Casals, J.; Hosmer, E.P. Sensitization and Antibody Formation after Injection of Tubercle Bacilli and Paraffin Oil. Proceedings of the Society for Experimental Biology and Medicine 1937, 37. [Google Scholar] [CrossRef]
- Leenaars, P.P.A.M.; Hendriksen, C.F.M.; Angulo, A.F.; Koedam, M.A.; Claassen, E. Evaluation of Several Adjuvants as Alternatives to the Use of Freund’s Adjuvant in Rabbits. Vet Immunol Immunopathol 1994, 40. [Google Scholar] [CrossRef]
- Arous, J. Ben; Deville, S.; Pal, J.K.; Baksi, S.; Bertrand, F.; Dupuis, L. Reduction of Newcastle Disease Vaccine Dose Using a Novel Adjuvant for Cellular Immune Response in Poultry. Procedia Vaccinol 2013, 7, 28–33. [Google Scholar] [CrossRef]
- Jang, S.I.; Lillehoj, H.S.; Lee, S.H.; Lee, K.W.; Park, M.S.; Bauchan, G.R.; Lillehoj, E.P.; Bertrand, F.; Dupuis, L.; Deville, S. Immunoenhancing Effects of MontanideTM ISA Oil-Based Adjuvants on Recombinant Coccidia Antigen Vaccination against Eimeria Acervulina Infection. Vet Parasitol 2010, 172, 221–228. [Google Scholar] [CrossRef]
- Chada, K.E.; Forshee, R.; Golding, H.; Anderson, S.; Yang, H. A Systematic Review and Meta-Analysis of Cross-Reactivity of Antibodies Induced by Oil-in-Water Emulsion Adjuvanted Influenza H5N1 Virus Monovalent Vaccines. Vaccine 2017, 35, 3162–3170. [Google Scholar] [CrossRef] [PubMed]
- Banzhoff, A.; Gasparini, R.; Laghi-Pasini, F.; Staniscia, T.; Durando, P.; Montomoli, E.; Capecchi, P.; di Giovanni, P.; Sticchi, L.; Gentile, C.; et al. MF59®-Adjuvanted H5N1 Vaccine Induces Immunologic Memory and Heterotypic Antibody Responses in Non-Elderly and Elderly Adults. PLoS One 2009, 4, e4384. [Google Scholar] [CrossRef]
- Barnett, P.V.; Pullen, L.; Williams, L.; Doel, T.R. International Bank for Foot-and-Mouth Disease Vaccine: Assessment of Montanide ISA 25 and ISA 206, Two Commercially Available Oil Adjuvants. Vaccine 1996, 14, 1187–1198. [Google Scholar] [CrossRef]
- Aslam, B.; Hussain, I.; Mahmood, M.S.; Khan, A. Preparation and Evaluation of Montanide ISA 206 Adjuvanted Bacterin of Borrelia Anserina in Laying Chickens. Journal of Applied Poultry Research 2013, 22, 196–203. [Google Scholar] [CrossRef]
- Gupta, S.K.; Deb, R.; Dey, S.; Chellappa, M.M. Toll-like Receptor-Based Adjuvants: Enhancing the Immune Response to Vaccines against Infectious Diseases of Chicken. Expert Rev Vaccines 2014, 13, 909–925. [Google Scholar] [CrossRef]
- Temperley, N.D.; Berlin, S.; Paton, I.R.; Griffin, D.K.; Burt, D.W. Evolution of the Chicken Toll-like Receptor Gene Family: A Story of Gene Gain and Gene Loss. BMC Genomics 2008, 9, 62. [Google Scholar] [CrossRef]
- Chaung, H.C.; Cheng, L.T.; Hung, L.H.; Tsai, P.C.; Skountzou, I.; Wang, B.; Compans, R.W.; Lien, Y.Y. Salmonella Flagellin Enhances Mucosal Immunity of Avian Influenza Vaccine in Chickens. Vet Microbiol 2012, 157, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Fu, J.; Kang, H.; Lin, J.; Yu, Q.; Yang, Q. Comparison of 3 Kinds of Toll-like Receptor Ligands for Inactivated Avian H5N1 Influenza Virus Intranasal Immunization in Chicken. Poult Sci 2013, 92, 2651–2660. [Google Scholar] [CrossRef] [PubMed]
- Bhadouriya, S.; Sharma, B.K.; Kakker, N.K.; Chhabra, R. Toll like Receptors and Cytokines as Immunostimulatory Adjuvants in Poultry Vaccines: Current Status and Future Trends. Worlds Poult Sci J 2019, 75, 417–427. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, M.; Li, J.; Cao, T.; Tian, X.; Zhou, F. Enhancement of Mucosal Immune Responses by Intranasal Co-Delivery of Newcastle Disease Vaccine plus CpG Oligonucleotide in SPF Chickens in Vivo. Res Vet Sci 2008, 85, 495–502. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Lowry, V.K.; Swaggerty, C.L.; Ferro, P.J.; Kogut, M.H. In Vitro Activation of Chicken Leukocytes and in Vivo Protection against Salmonella Enteritidis Organ Invasion and Peritoneal S. Enteritidis Infection-Induced Mortality in Neonatal Chickens by Immunostimulatory CpG Oligodeoxynucleotide. FEMS Immunol Med Microbiol 2005, 43, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Barjesteh, N.; Shojadoost, B.; Brisbin, J.T.; Emam, M.; Hodgins, D.C.; Nagy, É.; Sharif, S. Reduction of Avian Influenza Virus Shedding by Administration of Toll-like Receptor Ligands to Chickens. Vaccine 2015, 33, 4843–4849. [Google Scholar] [CrossRef]
- Hung, L.-H.; Tsai, P.-C.; Wang, C.-H.; Li, S.-L.; Huang, C.-C.; Lien, Y.-Y.; Chaung, H.-C. Immunoadjuvant Efficacy of Plasmids with Multiple Copies of a CpG Motif Coadministrated with Avian Influenza Vaccine in Chickens. Vaccine 2011, 29, 4668–4675. [Google Scholar] [CrossRef]
- Alqazlan, N.; Astill, J.; Raj, S.; Sharif, S. Strategies for Enhancing Immunity against Avian Influenza Virus in Chickens: A Review. Avian Pathology 2022, 51, 211–235. [Google Scholar] [CrossRef]
- Schijns, V.E.C.J.; Weining, K.C.; Nuijten, P.; Rijke, E.O.; Staeheli, P. Immunoadjuvant Activities of E. Coli- and Plasmid-Expressed Recombinant Chicken IFN-α/β, IFN-γ and IL-1β in 1-Day- and 3-Week-Old Chickens. Vaccine 2000, 18, 2147–2154. [Google Scholar] [CrossRef]
- Gan, L.; Tian, Y.; Zhao, Y.; Shan, X.; Zhou, W.; Xia, B.-B.; Chen, J.; Wang, M.-L.; Zhao, J. Enhancing Immunogenicity and Protective Efficacy of Inactivated Avian Influenza H9N2vaccine with Recombinant Chicken IFN-α in Chicken. Vet Microbiol 2019, 234, 77–82. [Google Scholar] [CrossRef]
- Karaca, K.; Sharma, J.M.; Winslow, B.J.; Junker, D.E.; Reddy, S.; Cochran, M.; McMillen, J. Recombinant Fowlpox Viruses Coexpressing Chicken Type I IFN and Newcastle Disease Virus HN and F Genes: Influence of IFN on Protective Efficacy and Humoral Responses of Chickens Following in Ovo or Post-Hatch Administration of Recombinant Viruses. Vaccine 1998, 16, 1496–1503. [Google Scholar] [CrossRef]
- Xiaowen, Z.; Qinghua, Y.; Xiaofei, Z.; Qian, Y. Co-Administration of Inactivated Avian Influenza Virus with CpG or RIL-2 Strongly Enhances the Local Immune Response after Intranasal Immunization in Chicken. Vaccine 2009, 27, 5628–5632. [Google Scholar] [CrossRef]
- Yang, Y.; Leggat, D.; Herbert, A.; Roberts, P.C.; Sundick, R.S. A Novel Method to Incorporate Bioactive Cytokines as Adjuvants on the Surface of Virus Particles. Journal of Interferon and Cytokine Research 2009, 29, 9–22. [Google Scholar] [CrossRef]
- Lim, K.-L.; Jazayeri, S.D.; Yeap, S.K.; Alitheen, N.B.M.; Bejo, M.H.; Ideris, A.; Omar, A.R. Co-Administration of Avian Influenza Virus H5 Plasmid DNA with Chicken IL-15 and IL-18 Enhanced Chickens Immune Responses. BMC Vet Res 2012, 8. [Google Scholar] [CrossRef]
- Melgoza-González, E.A.; Bustamante-Córdova, L.; Hernández, J. Recent Advances in Antigen Targeting to Antigen-Presenting Cells in Veterinary Medicine. Front Immunol 2023, 14. [Google Scholar] [CrossRef]
- Chen, P.; Liu, X.; Sun, Y.; Zhou, P.; Wang, Y.; Zhang, Y. Dendritic Cell Targeted Vaccines: Recent Progresses and Challenges. Hum Vaccin Immunother 2016, 12, 612–622. [Google Scholar] [CrossRef]
- Longet, S.; Lundahl, M.L.E.; Lavelle, E.C. Targeted Strategies for Mucosal Vaccination. Bioconjug Chem 2018, 29, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ju, X.; Silveira, P.A.; Abadir, E.; Hsu, W.-H.; Hart, D.N.J.; Clark, G.J. CD83: Activation Marker for Antigen Presenting Cells and Its Therapeutic Potential. Front Immunol 2019, 10. [Google Scholar] [CrossRef]
- Sun, P. Structural Recognition of Immunoglobulins by Fcγ Receptors. In Antibody Fc: Linking adaptive and innate immunity; 2014; pp. 131–144.
- Shrestha, A.; Sadeyen, J.-R.; Lukosaityte, D.; Chang, P.; Van Hulten, M.; Iqbal, M. Targeting Haemagglutinin Antigen of Avian Influenza Virus to Chicken Immune Cell Receptors Dec205 and CD11c Induces Differential Immune-Potentiating Responses. Vaccines (Basel) 2021, 9, 784. [Google Scholar] [CrossRef]
- Shrestha, A.; Meeuws, R.; Sadeyen, J.-R.; Chang, P.; Van Hulten, M.; Iqbal, M. Haemagglutinin Antigen Selectively Targeted to Chicken CD83 Overcomes Interference from Maternally Derived Antibodies in Chickens. NPJ Vaccines 2022, 7, 33. [Google Scholar] [CrossRef]
- Shrestha, A.; Sadeyen, J.-R.; Lukosaityte, D.; Chang, P.; Smith, A.; Van Hulten, M.; Iqbal, M. Selectively Targeting Haemagglutinin Antigen to Chicken CD83 Receptor Induces Faster and Stronger Immunity against Avian Influenza. NPJ Vaccines 2021, 6, 90. [Google Scholar] [CrossRef]
- Chou, W.K.; Chen, C.H.; Vuong, C.N.; Abi-Ghanem, D.; Waghela, S.D.; Mwangi, W.; Bielke, L.R.; Hargis, B.M.; Berghman, L.R. Significant Mucosal SIgA Production after a Single Oral or Parenteral Administration Using in Vivo CD40 Targeting in the Chicken. Res Vet Sci 2016, 108, 112–115. [Google Scholar] [CrossRef]
- Yin, G.; Lin, Q.; Qiu, J.; Qin, M.; Tang, X.; Suo, X.; Huang, Z.; Liu, X. Immunogenicity and Protective Efficacy of an Eimeria Vaccine Candidate Based on Eimeria Tenella Immune Mapped Protein 1 and Chicken CD40 Ligand. Vet Parasitol 2015, 210, 19–24. [Google Scholar] [CrossRef]
- Pecora, A.; Malacari, D.A.; Perez Aguirreburualde, M.S.; Bellido, D.; Nuñez, M.C.; Dus Santos, M.J.; Escribano, J.M.; Wigdorovitz, A. Development of an APC-Targeted Multivalent E2-Based Vaccine against Bovine Viral Diarrhea Virus Types 1 and 2. Vaccine 2015, 33, 5163–5171. [Google Scholar] [CrossRef]
- Brisbin, J.T.; Gong, J.; Orouji, S.; Esufali, J.; Mallick, A.I.; Parvizi, P.; Shewen, P.E.; Sharif, S. Oral Treatment of Chickens with Lactobacilli Influences Elicitation of Immune Responses. Clinical and Vaccine Immunology 2011, 18, 1447–1455. [Google Scholar] [CrossRef]
- Callaway, T.R.; Edrington, T.S.; Anderson, R.C.; Harvey, R.B.; Genovese, K.J.; Kennedy, C.N.; Venn, D.W.; Nisbet, D.J. Probiotics, Prebiotics and Competitive Exclusion for Prophylaxis against Bacterial Disease. Anim Health Res Rev 2008, 9, 217–225. [Google Scholar] [CrossRef]
- Brisbin, J.T.; Gong, J.; Parvizi, P.; Sharif, S. Effects of Lactobacilli on Cytokine Expression by Chicken Spleen and Cecal Tonsil Cells. Clinical and Vaccine Immunology 2010, 17, 1337–1343. [Google Scholar] [CrossRef]
- Schaupp, L.; Muth, S.; Rogell, L.; Kofoed-Branzk, M.; Melchior, F.; Lienenklaus, S.; Ganal-Vonarburg, S.C.; Klein, M.; Guendel, F.; Hain, T.; et al. Microbiota-Induced Type I Interferons Instruct a Poised Basal State of Dendritic Cells. Cell 2020, 181, 1080–1096. [Google Scholar] [CrossRef] [PubMed]
- Casteleyn, C.; Doom, M.; Lambrechts, E.; Van den Broeck, W.; Simoens, P.; Cornillie, P. Locations of Gut-Associated Lymphoid Tissue in the 3-Month-Old Chicken: A Review. Avian Pathol 2010, 39, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, D.; Kolożyn-Krajewska, D. Food-Origin Lactic Acid Bacteria May Exhibit Probiotic Properties: Review. Biomed Res Int 2018, 2018, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Shojadoost, B.; Kulkarni, R.R.; Brisbin, J.T.; Quinteiro-Filho, W.; Alkie, T.N.; Sharif, S. Interactions between Lactobacilli and Chicken Macrophages Induce Antiviral Responses against Avian Influenza Virus. Res Vet Sci 2019, 125, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Alqazlan, N.; Astill, J.; Taha-Abdelaziz, K.; Nagy, É.; Bridle, B.; Sharif, S. Probiotic Lactobacilli Enhance Immunogenicity of an Inactivated H9N2 Influenza Virus Vaccine in Chickens. Viral Immunol 2021, 34, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Kye, Y.-C.; Park, S.-M.; Shim, B.-S.; Yoo, S.; Hwang, E.; Kim, H.; Kim, S.-J.; Han, S.H.; Park, T.S.; et al. Bacillus Subtilis Spores as Adjuvants against Avian Influenza H9N2 Induce Antigen-Specific Antibody and T Cell Responses in White Leghorn Chickens. Vet Res 2020, 51, 68. [Google Scholar] [CrossRef]
- Bavananthasivam, J.; Alizadeh, M.; Astill, J.; Alqazlan, N.; Matsuyama-Kato, A.; Shojadoost, B.; Taha-Abdelaziz, K.; Sharif, S. Effects of Administration of Probiotic Lactobacilli on Immunity Conferred by the Herpesvirus of Turkeys Vaccine against Challenge with a Very Virulent Marek’s Disease Virus in Chickens. Vaccine 2021, 39, 2424–2433. [Google Scholar] [CrossRef]
- El-Shall, N.A.; Awad, A.M.; El-Hack, M.E.A.; Naiel, M.A.E.; Othman, S.I.; Allam, A.A.; Sedeik, M.E. The Simultaneous Administration of a Probiotic or Prebiotic with Live Salmonella Vaccine Improves Growth Performance and Reduces Fecal Shedding of the Bacterium in Salmonella-Challenged Broilers. Animals 2020, 10. [Google Scholar] [CrossRef]
- Yousaf, A.; Anwar, H. Effect of Gutcare TM and Enterogermina ® on Humoral Response of Avian Influenza Immunization in Broilers. International Journal of Veterinary Sciences and Animal Husbandry 2019, 4, 19–23. [Google Scholar]
- Wu, P.; Lu, J.; Feng, L.; Wu, H.; Zhang, X.; Mei, M.; Hou, J.; Liu, X.; Tang, Y. Antigen-Sparing and Enhanced Efficacy of Multivalent Vaccines Adjuvanted with Immunopotentiators in Chickens. Front Microbiol 2017, 8. [Google Scholar] [CrossRef]
- Lauer, K.B.; Borrow, R.; Blanchard, T.J. Multivalent and Multipathogen Viral Vector Vaccines. Clinical and Vaccine Immunology 2017, 24. [Google Scholar] [CrossRef]
- Arevalo, C.P.; Bolton, M.J.; Le Sage, V.; Ye, N.; Furey, C.; Muramatsu, H.; Alameh, M.G.; Pardi, N.; Drapeau, E.M.; Parkhouse, K.; et al. A Multivalent Nucleoside-Modified MRNA Vaccine against All Known Influenza Virus Subtypes. Science (1979) 2022, 378, 899–904. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Illustration of different methods of vaccination in poultry. Methods used in the poultry industry for vaccinating chickens. Created with BioRender.com.
Figure 1.
Illustration of different methods of vaccination in poultry. Methods used in the poultry industry for vaccinating chickens. Created with BioRender.com.
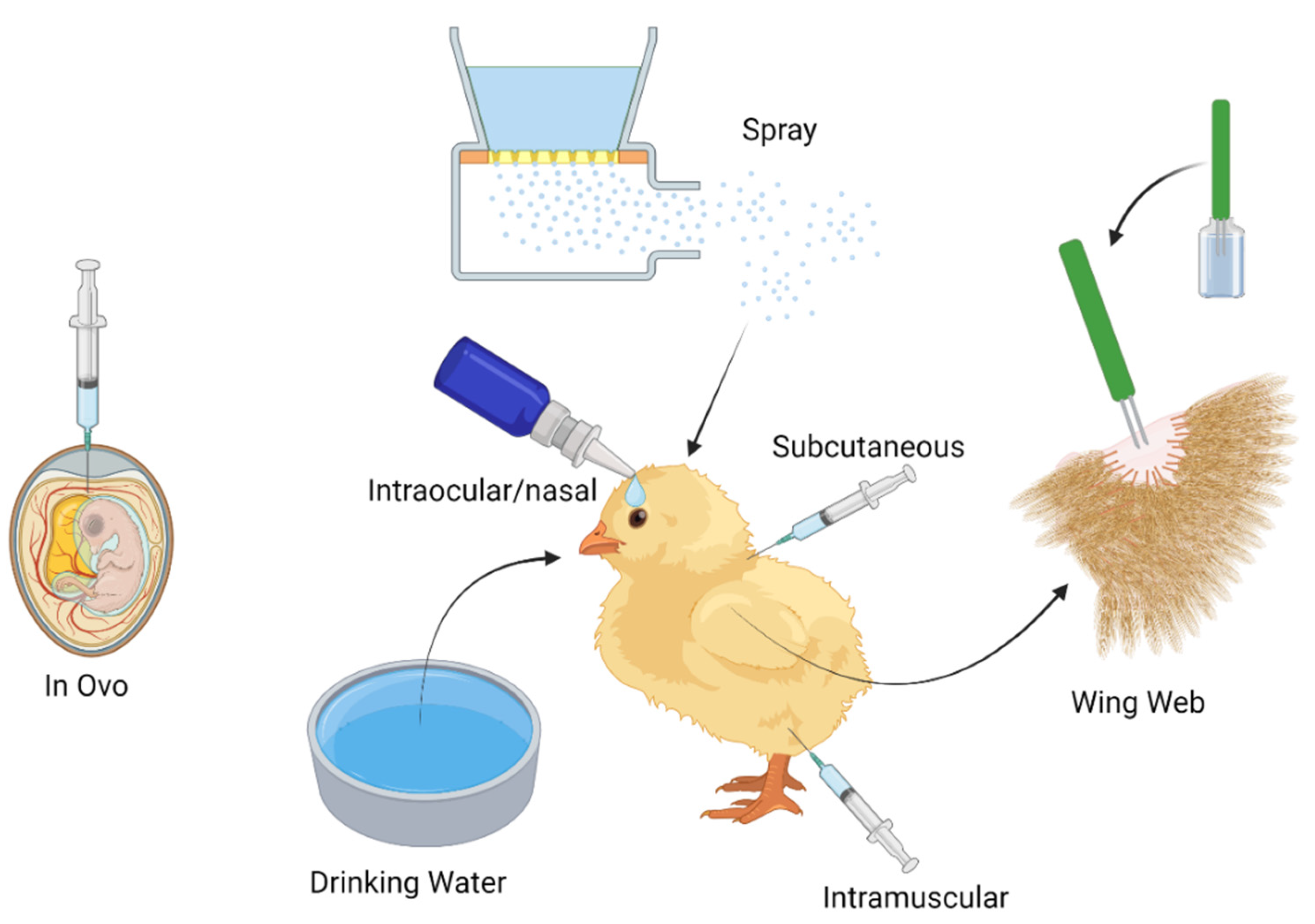
Figure 2.
Illustration of different types of oil emulsions. (a) Water in oil emulsion (W/O). This method traps antigen (Ag) inside water droplets. This protects the structure of the Ag, allowing for slow antigen release due to the breakdown of oil. This prevents rapid clearance of the antigen which allows for increased immune cell recruitment and antigen processing. (b) Oil in water (O/W) emulsion. Here the oil helps attract monocytes and chemokines to the injection site. This promotes the differentiation of monocytes into dendritic cells (DCs) and stimulates a strong cellular immune response. (c) Water in oil in water (W/O/W). Here antigen is in both the oil and water phases, so antigen is both up taken quickly from the water phase but also slowly release from internal water phase. The breakdown of the oil also helps with the attraction of monocytes and chemokines to help induce a strong cellular immunity. Created with BioRender.com.
Figure 2.
Illustration of different types of oil emulsions. (a) Water in oil emulsion (W/O). This method traps antigen (Ag) inside water droplets. This protects the structure of the Ag, allowing for slow antigen release due to the breakdown of oil. This prevents rapid clearance of the antigen which allows for increased immune cell recruitment and antigen processing. (b) Oil in water (O/W) emulsion. Here the oil helps attract monocytes and chemokines to the injection site. This promotes the differentiation of monocytes into dendritic cells (DCs) and stimulates a strong cellular immune response. (c) Water in oil in water (W/O/W). Here antigen is in both the oil and water phases, so antigen is both up taken quickly from the water phase but also slowly release from internal water phase. The breakdown of the oil also helps with the attraction of monocytes and chemokines to help induce a strong cellular immunity. Created with BioRender.com.
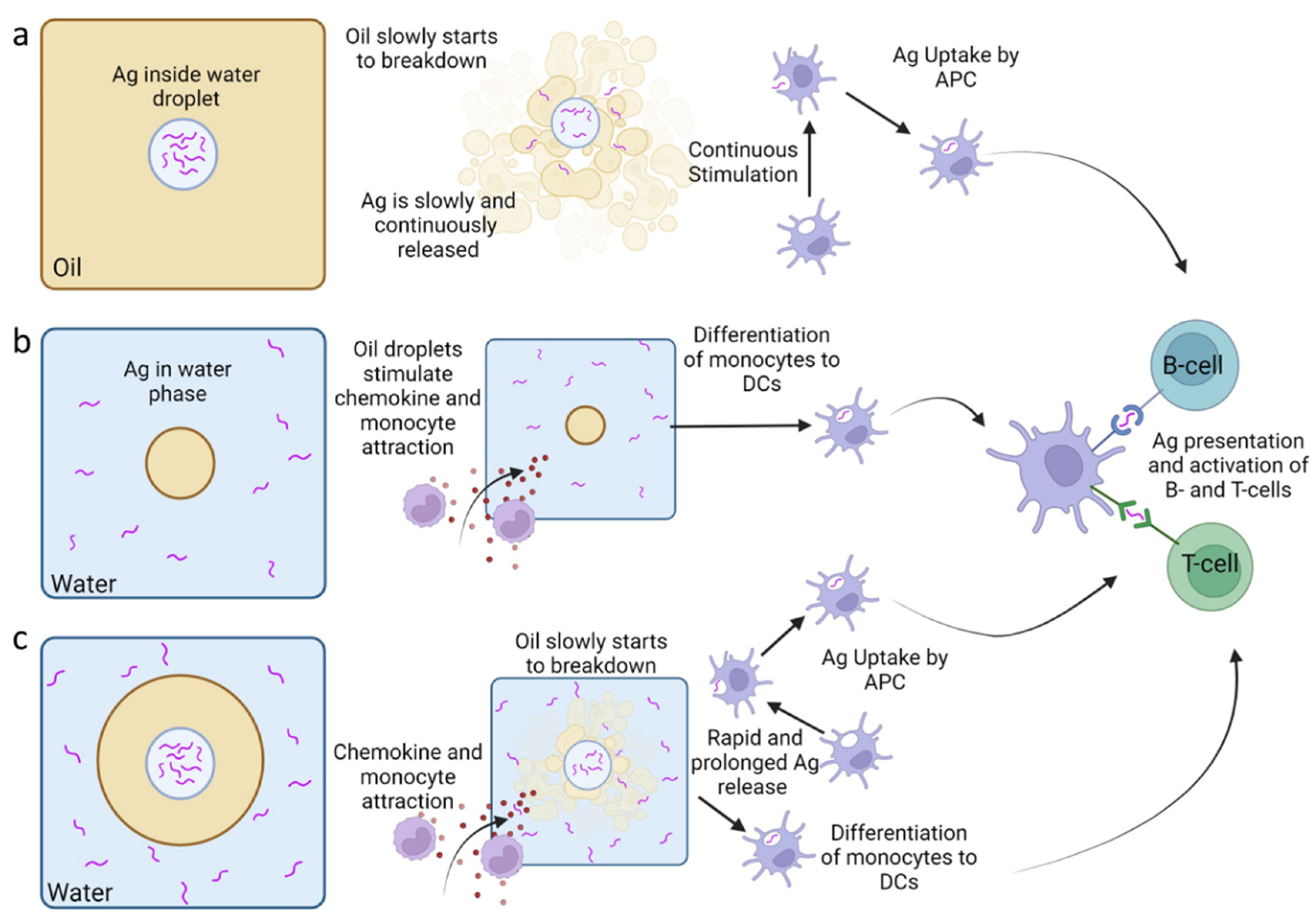
Figure 3.
Overview of 10 known chicken TLRs. TLRs are anchored membrane receptor proteins that are expressed on APC. They recognise conserved components on microbes and once activated result in a proinflammatory response and induce the expression of cytokines involved in immune cell differentiation. This helps the induction of cell mediated and humoral immunity. IFN = interferon, TLR = toll like receptor, LPS = lipopolysaccharide. Created with BioRender.com.
Figure 3.
Overview of 10 known chicken TLRs. TLRs are anchored membrane receptor proteins that are expressed on APC. They recognise conserved components on microbes and once activated result in a proinflammatory response and induce the expression of cytokines involved in immune cell differentiation. This helps the induction of cell mediated and humoral immunity. IFN = interferon, TLR = toll like receptor, LPS = lipopolysaccharide. Created with BioRender.com.
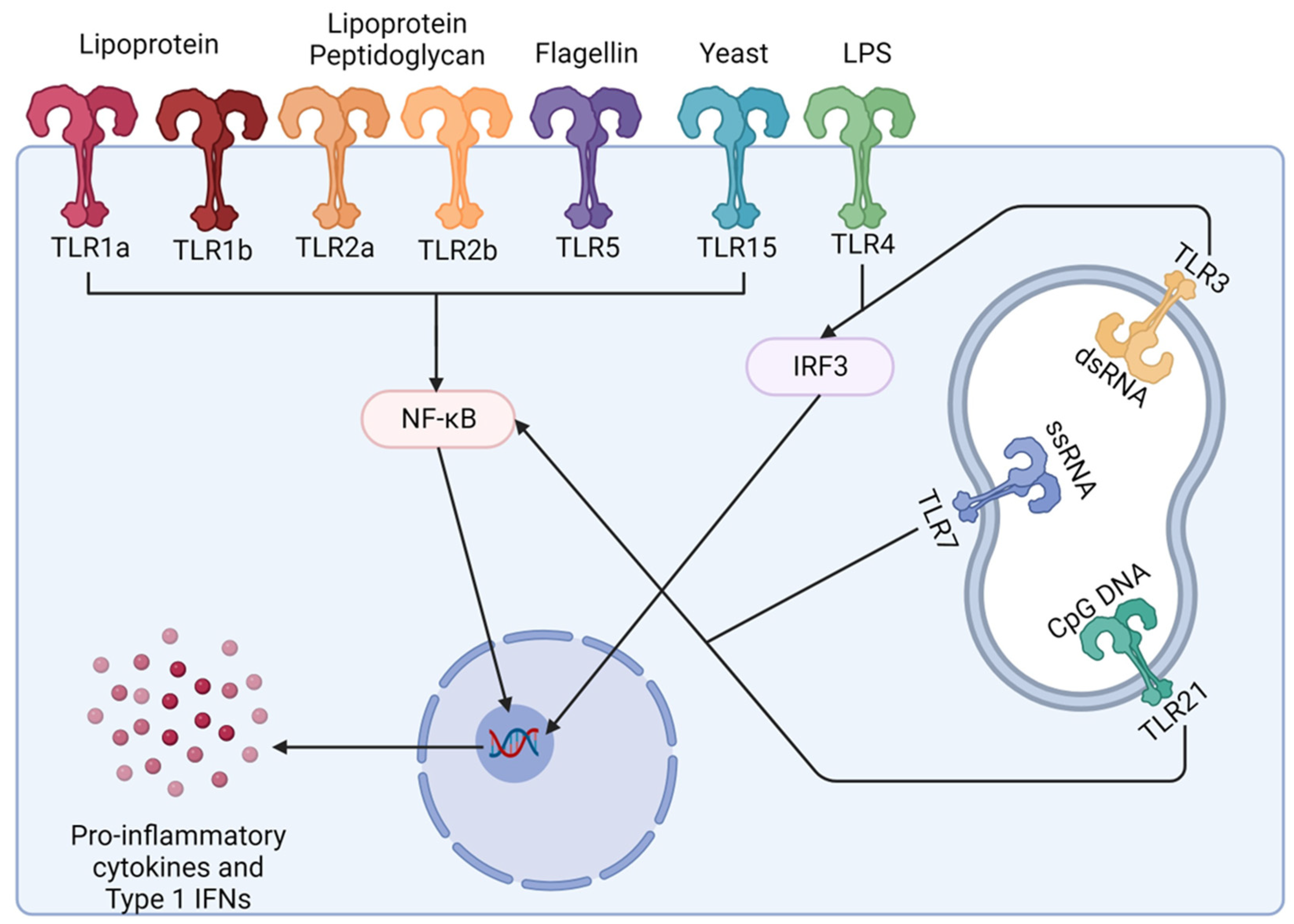
Figure 4.
Validated APC targets in chickens. Antigen presenting cell targets that have been validated in poultry vaccines. CD11c = β-integrin molecule, shown to be on mammalian dendritic cells, neutrophils, macrophages, and NK cells. In chickens CD11c has been found on dendritic cells and macrophages but has not been well studied. DC-pep = peptides that can recognise dendritic cells from a larger leukocyte population. CD40 = co-stimulatory cell surface receptor from the tumour necrosis factor family. In humans CD40 is expressed on dendritic cells, monocytes, macrophages, and B-cells. In chickens CD40 has been detected on monocytes, macrophages DCs, and B-cells. CD83 = immunoglobulin molecule which in mammals is used as an activation molecule. In mammals CD83 has been found on activated NK cells, monocytes, macrophages, neutrophils, dendritic cells and B-cells. In chickens, CD83 has been found on dendritic cells but has not been well studied. DEC-205 = endocytic receptor which in mammals is expressed on DCs, macrophages, B and T-cells. In chickens, DEC-205 has been detected on DCs, B-cells, T-cells but has not been well studied. Figure created with BioRender.com.
Figure 4.
Validated APC targets in chickens. Antigen presenting cell targets that have been validated in poultry vaccines. CD11c = β-integrin molecule, shown to be on mammalian dendritic cells, neutrophils, macrophages, and NK cells. In chickens CD11c has been found on dendritic cells and macrophages but has not been well studied. DC-pep = peptides that can recognise dendritic cells from a larger leukocyte population. CD40 = co-stimulatory cell surface receptor from the tumour necrosis factor family. In humans CD40 is expressed on dendritic cells, monocytes, macrophages, and B-cells. In chickens CD40 has been detected on monocytes, macrophages DCs, and B-cells. CD83 = immunoglobulin molecule which in mammals is used as an activation molecule. In mammals CD83 has been found on activated NK cells, monocytes, macrophages, neutrophils, dendritic cells and B-cells. In chickens, CD83 has been found on dendritic cells but has not been well studied. DEC-205 = endocytic receptor which in mammals is expressed on DCs, macrophages, B and T-cells. In chickens, DEC-205 has been detected on DCs, B-cells, T-cells but has not been well studied. Figure created with BioRender.com.
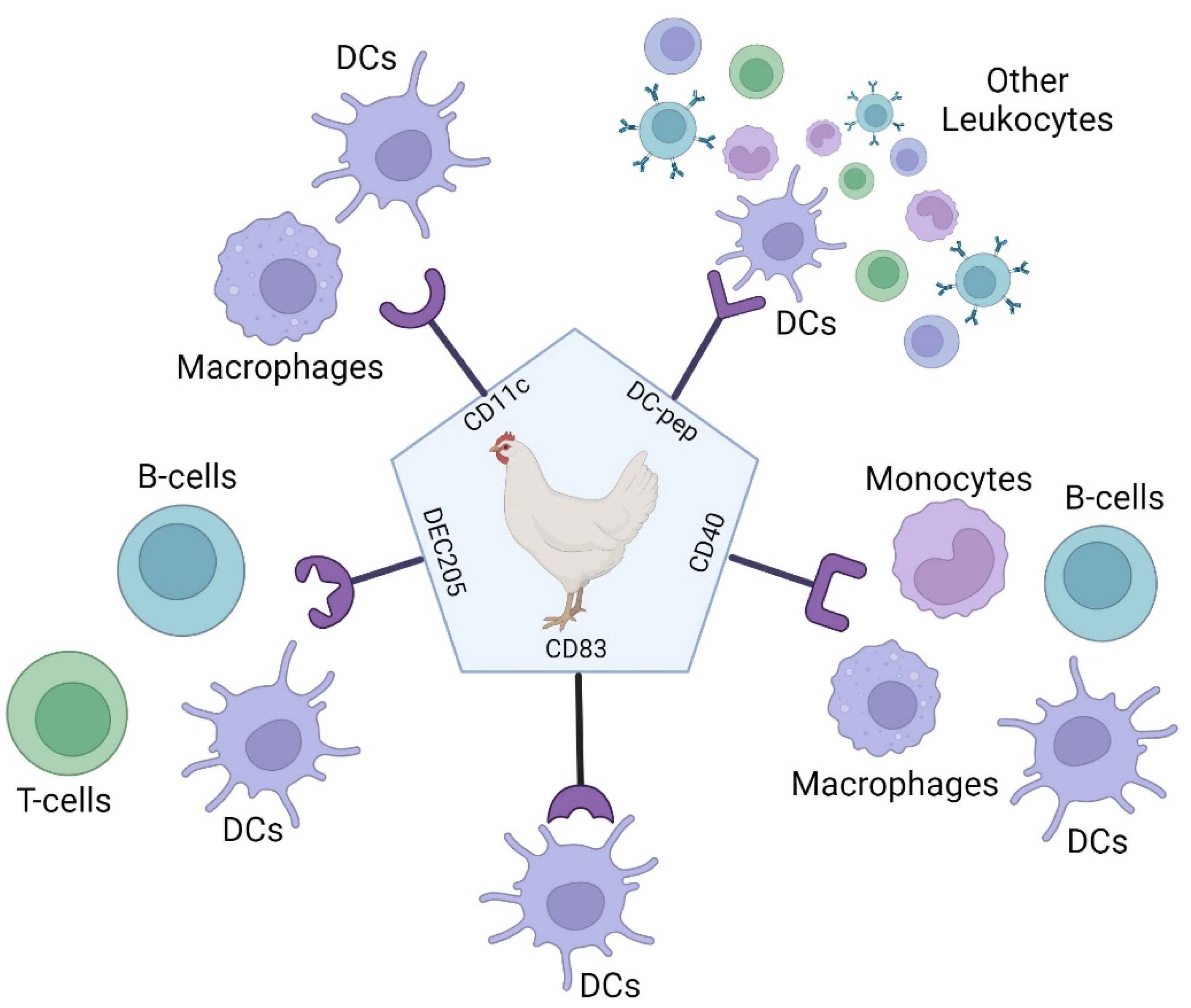
Figure 5.
Illustration of ligand-based targeting of APCs. PRR ligands can be chemically or recombinantly bound to antigens. TLR ligands are most used for antigen targeting studies as these enable both the targeting of antigen to APC and have adjuvant properties. The antigen-TLR conjugate vaccine binds to its receptor on the APC, this induces receptor mediated endocytosis. This induces APC maturation and allows for T-cell presentation via either MHC-I or MHC-II pathways. Presentation MHC-I presentation of antigen to CD8 T-cells results in the development of a population of antigen specific cytotoxic T-cells which are capable of lysing infected cells. MHC-II presentation of antigen to CD4 T-cells which present antigen to B-cells and results in the development of antigen specific antibody production. Created with BioRender.com.
Figure 5.
Illustration of ligand-based targeting of APCs. PRR ligands can be chemically or recombinantly bound to antigens. TLR ligands are most used for antigen targeting studies as these enable both the targeting of antigen to APC and have adjuvant properties. The antigen-TLR conjugate vaccine binds to its receptor on the APC, this induces receptor mediated endocytosis. This induces APC maturation and allows for T-cell presentation via either MHC-I or MHC-II pathways. Presentation MHC-I presentation of antigen to CD8 T-cells results in the development of a population of antigen specific cytotoxic T-cells which are capable of lysing infected cells. MHC-II presentation of antigen to CD4 T-cells which present antigen to B-cells and results in the development of antigen specific antibody production. Created with BioRender.com.
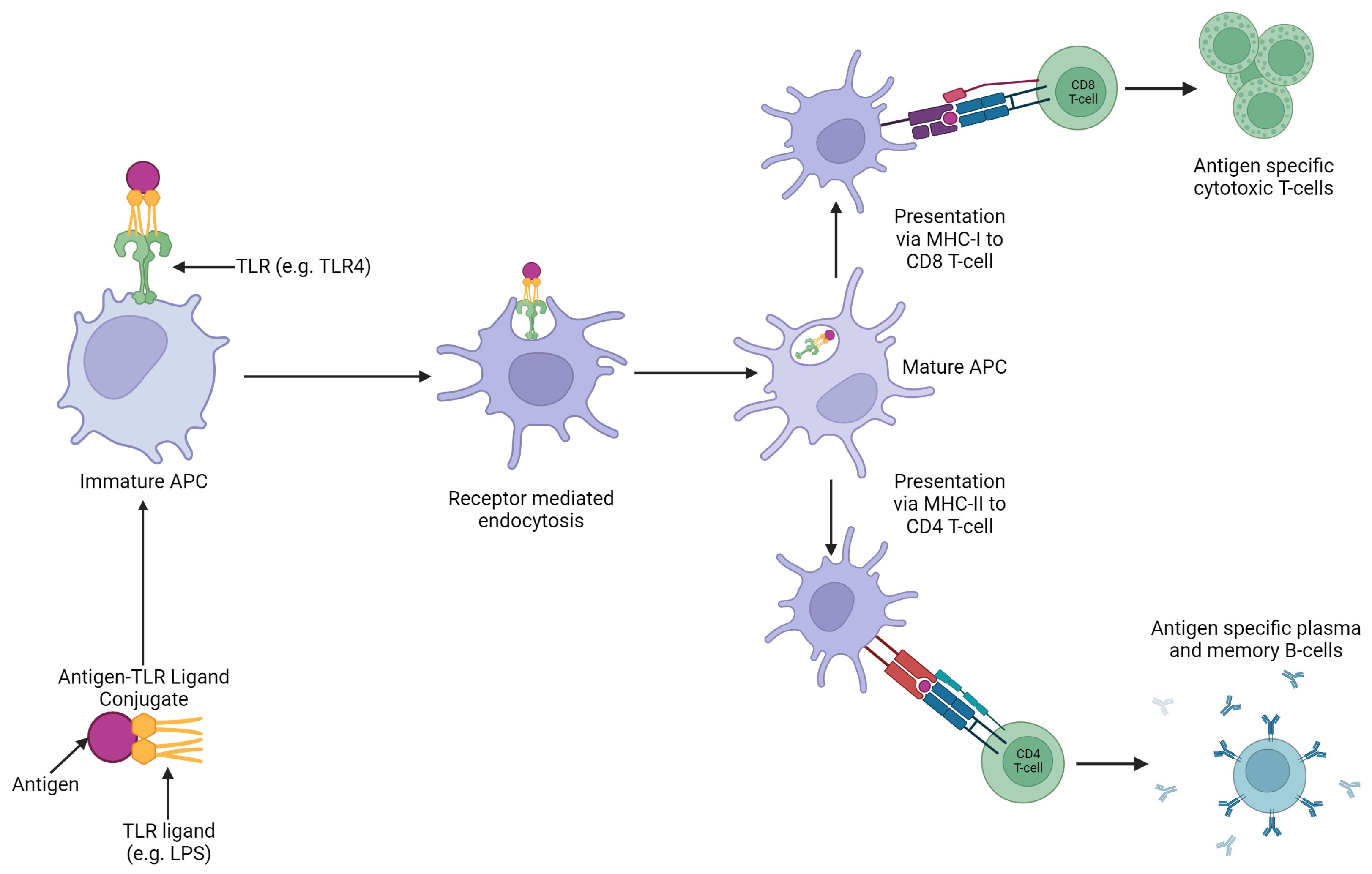
Figure 6.
Illustration of ligand-based targeting of APCs. Full antibody or antibody-based fragments can be chemically or recombinantly bound to antigens. APC specificity is determined via the use of anti-receptor antibodies which target the desired receptor on an APC. Antigen-antibody complex will bind to its targeted receptor on an APC this induces receptor mediated endocytosis. This induces APC maturation and allows for T-cell presentation via either MHC-I or MHC-II pathways. MHC-I presentation of antigen to CD8 T-cells results in the development of a population of antigen specific cytotoxic T-cells which are capable of lysing infected cells. MHC-II presentation of antigen to CD4 T-cells can activate B-cells and results in the development of antigen specific antibody production. Created with BioRender.com.
Figure 6.
Illustration of ligand-based targeting of APCs. Full antibody or antibody-based fragments can be chemically or recombinantly bound to antigens. APC specificity is determined via the use of anti-receptor antibodies which target the desired receptor on an APC. Antigen-antibody complex will bind to its targeted receptor on an APC this induces receptor mediated endocytosis. This induces APC maturation and allows for T-cell presentation via either MHC-I or MHC-II pathways. MHC-I presentation of antigen to CD8 T-cells results in the development of a population of antigen specific cytotoxic T-cells which are capable of lysing infected cells. MHC-II presentation of antigen to CD4 T-cells can activate B-cells and results in the development of antigen specific antibody production. Created with BioRender.com.
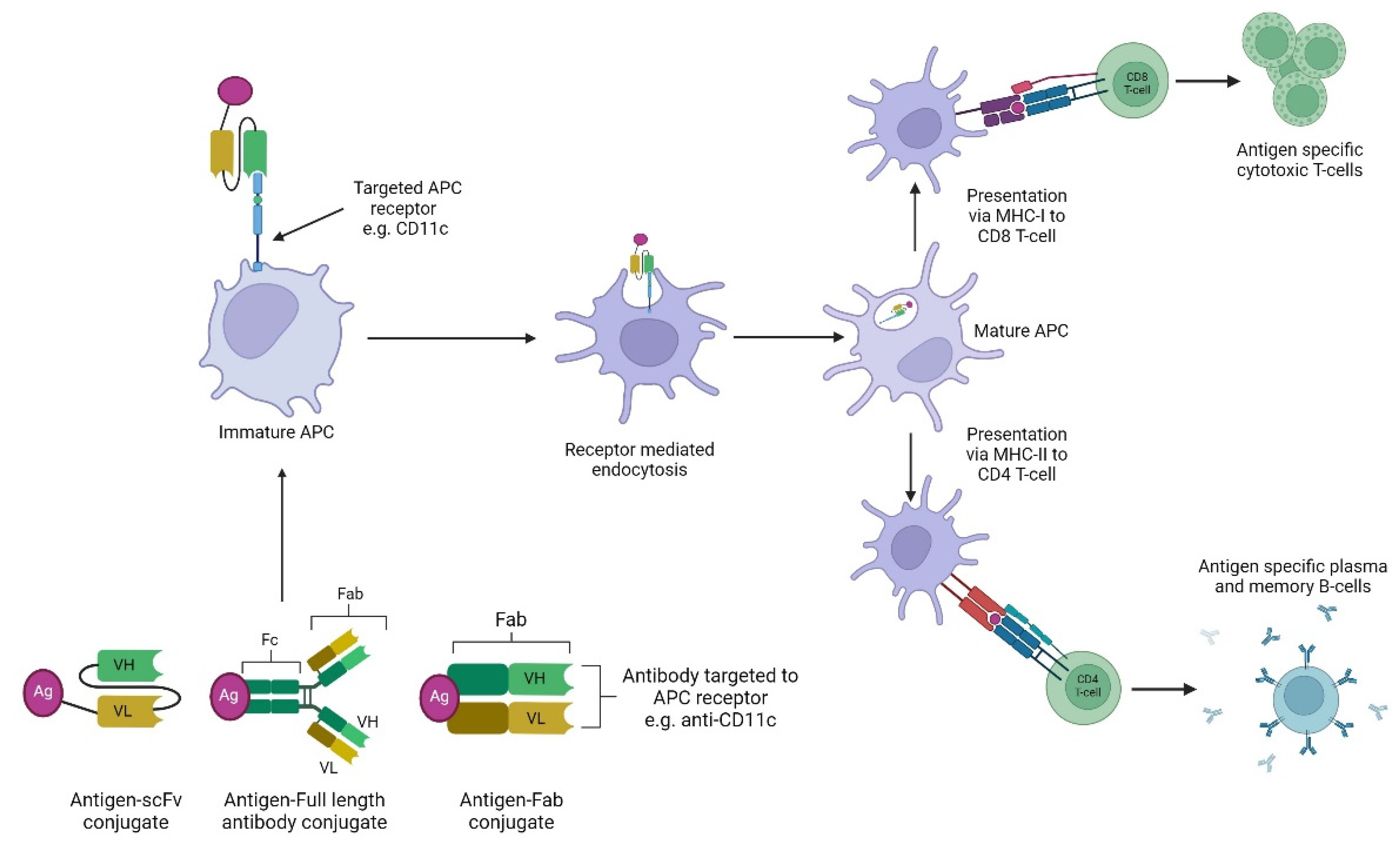

Table 1.
List of poultry vaccines licensed in the United Kingdom as per the Veterinary Medicines Directorate, GOV.UK[14] https://www.vmd.defra.gov.uk/productinformationdatabase.
Table 1.
List of poultry vaccines licensed in the United Kingdom as per the Veterinary Medicines Directorate, GOV.UK[14] https://www.vmd.defra.gov.uk/productinformationdatabase.
| Disease | Biological Agent | Type of Vaccine | Method of Vaccine Production | Delivery Method | Adjuvant Used | Veterinary Medicine (VM) Number |
| Avian encephalomyelitis virus | Virus | Live | Apathogenic strain (for adult birds) | Drinking water | None specified | 00879/5005 01708/4281 42058/5134 |
| Avian influenza Virus (H5N2) | Virus | Inactivated | Inactivation method unspecified | Subcutaneous Intramuscular |
Paraffin oil | 01708/5044 |
| Avian metapneumovirus | Virus | Live strain Live attenuated |
Apathogenic strain; Attneuated (exact method unspecified) |
Drinking water Spray Intraocular/nasal |
None specified | 15052/4086 08327/5008 42058/4112 |
| Avian reovirus | Virus | Inactivated | Formaldehyde inactivated | Intramuscular Subcutaneous |
None specified | 01708/4329 |
| Avian rhinotracheitis virus |
Virus | Live attenuated; Inactivated |
Continuous passage in tissue culture; Formaldehyde or β-propiolactone inactivated |
Intraocular/nasal Spray Intramuscular |
Paraffin oil | 08327/5022 08327/5025 01708/4505 01708/5093 01708/5083 01708/5105 08327/4134 |
| Chicken Anaemia Virus | Virus | Live; Live attenuated |
Apathogenic strain (for adult birds) | Drinking water; Intramuscular Subcutaneous |
dl-α-tocopherol acetate (emulsion) | 00879/5042 01708/4322 |
| Coccidia (Eimeria) | Parasite | Live attenuated | Attneuated (exact method unspecified) | Spray Drinking water In ovo |
None specified | 17533/5014 30282/5018 30282/4032 01708/5101 01708/4572 |
| E. coli | Bacteria | Inactivated; Live |
Genetically modified | Intramuscular Subcutaneous Spray Drinking water |
Paraffin oil | 42058/5046 01708/5085 |
| Egg drop syndrome (EDS) | Virus | Inactivated | Inactivation method unspecified | Intramuscular | Paraffin oil | 08327/5012 08327/5025 01708/4275 01708/5083 |
| Erysipelothrix rhusiopathiae | Bacteria | Inactivated | Inactivation method unspecified | Subcutaneous |
None specified | 01708/4546 |
| Herpes Virus of Turkeys (HVT) | Virus | Live recombinant; Live |
Genetic recombination |
Subcutaneous In ovo |
None specified | 15052/4089 01708/5082 01708/5040 01708/5041 01708/4289 04491/5043 15052/5001 04491/5060 |
| Infectious Bronchitis Virus (IBV) | Virus | Inactivated; Live; Live attenuated; |
Attneuated (exact method unspecified); Formaldehyde or β-propiolactone inactivated |
Spray Intraocular/nasal Drinking water Intramuscular |
Paraffin oil | 43676/4005 43676/4002 15052/5043 15052/4088 08327/5012 08327/5025 08327/4265 08327/5005 01708/4275 01708/5043 01708/4283 01708/5042 01708/4315 01708/5093 01708/5083 42058/5121 42058/4103 42058/5103 42058/5140 |
| Infectious Bursal disease (IBD) | Virus | HVT Vector expressing IBD VP2 protein; Live; Live attenuated; |
Viral Vector Attenuation in eggs; Attenuation in tissue culture |
Subcutaneous In ovo Drinking water Spray Intraocular/nasal |
Paraffin oil | 00879/3032 00879/4187 43676/5000 15052/4059 15052/4030 08327/4192 08327/4323 17533/5005 17533/5017 17533/4002 01708/5082 01708/5040 15052/4154 01708/4333 01708/4237 01708/5093 15052/5032 42058/5138 42058/5137 04491/5043 15052/5001 04491/5060 |
| Infectious Laryngotracheitis (ILT) | Virus | HVT Vector expressing ILT gD and gl glycoproteins | Viral Vector | Subcutaneous; In ovo Intraocular |
None specified | 01708/5039 01708/5082 01708/5041 42058/4106 |
| Mareks Disease Virus (MDV) | Virus | Live apathogenic; HVT Vector, Live recombinant Live attenuated |
Live, Viral vector, Genetic recombination; Attneuated (exact method unspecified) |
Subcutaneous Intramuscular In ovo |
None specified | 15052/4089 15052/4151 08327/4104 01708/4294 01708/4354 42058/4107 42058/4108 42508/4109 04491/5042 04491/5043 |
|
Mycoplasma gallisepticum; Mycoplasma synovia |
Bacteria | Live attenuated; Inactivated |
Attneuated (exact method unspecified) | Spray Subcutaneous Intraocular Intraocular/nasal |
Paraffin Oil | 43877/5001 43877/5000 42983/5000 01708/5103 01708/4607 |
| Newcastle disease (NDV) | Virus | HVT Vector expressing NDV F protein; Inactivated; Live ; Live attenuated ; Live lentogenic; |
Viral Vector, Attneuated (exact method unspecified) Formaldehyde or β-propiolactone inactivated, Lentogenic strain |
Subcutaneous Intramuscular In ovo Spray Intraocular/nasal Drinking water |
Paraffin Oil | 08327/4266 00879/5035 43676/4003 43676/4000 08327/5012 08327/5025 01708/5040 01708/5041 01708/4275 01708/4315 01708/5088 01708/4276 01708/5093 01708/5083 |
| Paramyxovirus Type 3 | Virus | Inactivated | Inactivation method unspecified | Intramuscular | Paraffin oil | 08327/4134 |
| Salmonella (enteritidis, typhimurium and infantis) | Bacteria | Live attenuated; Inactivated |
Attenuation method unspecified; Attenuation via genetic mutation, Inactivation method unspecified |
Intramuscular Drinking water |
Paraffin oil, aluminium hydroxide | 00879/4188 00879/4189 00879/5037 15052/5045 08327/4222 01708/5073 01708/5114 20634/5002 20634/5000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



