
Submitted:
27 September 2024
Posted:
30 September 2024
You are already at the latest version
Abstract
Background/Objectives: Bovine babesiosis is a vector-borne disease transmitted by ticks that causes important losses in livestock worldwide. Recent research performed on the drugs currently used to control bovine babesiosis reported several issues including drug resistance, toxicity impact, and residues in edible tissue, suggesting the need for developing novel effective therapies. The endochin-like quinolones ELQ-316 and buparvaquone (BPQ) act as cytochrome bc1 inhibitors and have been proven to be safe and efficacious against related apicomplexans, such as Plasmodium spp. and Babesia microti, without showing toxicity in mammals. The objectives of this study are investigating whether ELQ-316, BPQ and their combination treatment could be effective against Babesia bovis in an in vitro culture model, and comparing with imidocarb (ID), the routinely used drug. Methods: In vitro cultured parasites at starting 2% percentage of parasitemia (PPE) were treated with BPQ, ELQ-316, ID, and the combinations of BPQ+ ELQ-316 and ID+ ELQ-316 at drug concentrations that ranged from 25 to 1200 nM, during four consecutive days. Parasitemia levels were evaluated daily using microscopic examination. Data was compared using the independent Student's t-test and ANOVA test. Results: All drugs tested, whether used alone or in combination, significantly decreased the survival (p<0.05) of B. bovis in in vitro cultures. The combination of BPQ+ ELQ-316 had the lowest calculated inhibitory concentration 50 (IC50) 31.21 nM (IC95%: 15.06 - 68.48) values, followed by BPQ, 77.06 nM (IC95%: 70.16- 86.01), ID+ELQ316 197 nM (IC95%:129.0 - 311.2), ID 635.1 nM (IC95%: 280.9 – 2119) and ELQ316 654.9 nM (IC95%: 362.3 – 1411). Conclusion: The results reinforce the higher efficacy of BPQ at affecting B. bovis survival, and the potential synergistic effects of its combination with ELQ-316, providing a promising treatment option against B. bovis.
Keywords:
apicomplexa
; treatment
; cytocrome
; new drugs
; IC50
1. Introduction
Babesiosis, primarily caused by B. bovis and B. bigemina, is a tick-borne parasitic disease that significantly impacts the cattle industry worldwide ([1,2,3,4]. The disease can lead to acute disease and persistent infection in livestock [5], causing high morbidity and mortality in tropical and semi-tropical regions [6].
Effective control of piroplasmosis involves three primary strategies: vaccination, the use of antipiroplasm drugs, and vector control measures [7,8,9,10,11,12]. Chemotherapy is a critical tool for babesiosis control, with several drugs and combinations providing to be effective [13,14,15,16,17,18,19]. However, most, if not all of the more effective drugs currently in use have toxic side effects, accumulation of residues in the edible tissues of treated animals or generate drug-resistant parasites [20,21,22,23]. These outcomes triggered the urgent need to develop novel antipiroplasm drugs and effective new regimens [9,24,25].
Imidocarb is currently the first-line treatment for bovine babesiosis, but it is not yet approved for use in cattle in the US [26]. This drug has potent babesiacidal effects, providing clearance of parasites and prophylactic protection [7,13]. However, ID's mechanism of action is not well understood. ID has toxic effects in animals, and persistent residues in edible tissues [8,27,28,29,30].
BPQ, a second-generation hydroxynaphthoquinone antiprotozoal drug, selectively inhibits the parasite’s Qo quinone-binding site of mitochondrial cytochrome b electron transport system, leading to its lethal effect on parasites [31,32,33,34]. Although BPQ is registered in around 20 countries for treating East Coast fever and tropical theileriosis [35], its approval in other countries is limited due to residual toxicity concerns [36]
BPQ was initially developed as an anti-malarial drug and has also shown promise in treating Theileria species in previous studies [37,38,39,40,41,42,43,44]. Furthermore, it was recently reported that BPQ is significantly more effective than ID in inhibiting the growth of B. bovis in vitro, at various concentrations and time points [19].
ELQ-316 is an endochin-like quinolone (ELQ) compound that selectively inhibits the parasite’s Qi quinone-binding site of mitochondrial cytochrome bc1, and was highly effective against Plasmodium falciparum, Babesia microti and Toxoplasma gondii [45] It was also shown to be effective in in vitro treatment of Theileria spp. and Babesia spp. parasites [16] . The ELQs also demonstrated high antimalarial potency in vitro and in vivo, parasite selectivity, chemical and metabolic stability, desirable pharmacokinetics, and low mammalian cell toxicity. In addition to their antimalarial activity, compounds in the ELQ series were later found to be highly active against other Apicomplexa, for which effective treatments are urgently needed. With favorable properties and broad-spectrum activity, the ELQ compound class may yield effective, safe treatments for a range of important human and animal afflictions.
Although a single-drug treatment is preferred, the use of mitochondrial bc1 inhibitors may result in the emergence of resistant parasites associated with a mutation in the Cytb Qi or Qo sites. However, the combination of compounds with activity against both sites (Cytb Qi and Qo), like ELQ-316 and BPQ, may improve the treatment and reduce the possibility for the emergence of parasite with drug resistance [46]. This was previously demonstrated by complete and persistent clearance of parasites [47]. Therefore, BPQ and ELQ-316 may offer effective options for managing babesiosis, especially in cases where traditional treatments like imidocarb dipropionate fail, result in frequent clinical relapses, or induce toxicity or generate undesired residues in food. So far as we know, there are no previous studies regarding the effects on the survival of these combinations against B. bovis.
The aim of this study was to compare the effects of ELQ-316, BPQ, ID and the combinations of ID + ELQ-316 and BPQ + ELQ-316 on their ability to affect the B. bovis in vitro survival.
2. Materials and Methods
2.1. Compounds Tested
BPQ (98% purity) was obtained from Combi-Blocks, Inc. (San Diego, CA, USA). ID (VETRANALTM, Supelco® Buchs, Switzerland) was used as a positive control for the in vitro inhibition assays for B. bovis, using an identical protocol as BPQ described below. Purity of ID was determined to be > 98% by proton-nuclear magnetic resonance spectroscopy and High-performance liquid chromatography (HPLC), according to the certificate of analysis.
ELQ-316 was synthesized by following methods previously described by Nilsen et al. (2014), identified by proton nuclear magnetic resonance (1H NMR), and determined to be ≥95% pure by reversed-phase high-performance liquid chromatography (RP-HPLC) [48].
BPQ, ELQ-316 and ID were diluted in 100% dimethyl sulfoxide (DMSO) to prepare stock solutions, which were kept at room temperature until use. Working solutions were freshly prepared in a parasite culture medium every test day before being added to the parasite cultures.
2.2. Babesia Bovis In Vitro CULTURE
B. bovis (Texas T2Bo strain: Goff et al. 1998) were grown in long-term microaerophilic stationary-phase cultures and incubated at 37°C in an atmospheric condition of 5% CO2, 5% O2, and 90% N2, as previously described (Levy & Ristic, 1980). B. bovis were cultured in 96-well-culture plates, containing 180 µl per well of PL culture media (pH 7.2, to prepare 100 ml, = 29 ml F-12K Nutrient Mixture + L-glutamine (Life Technologies), 29 ml Stable Cell IMDM (Sigma Aldrich), 2 ml 0.5 M TAPSO (Sigma Aldrich), 0.5 ml Antibiotic Antimycotic solution 100X (Sigma Aldrich), 1 mM calcium chloride (Sigma Aldrich), 100 µl Antioxidant Supplement 1000x (Sigma Aldrich), 1 ml Insulin-Transferrin-Selenium-Ethanolamine 100x (Sigma Aldrich), 1 ml 50% Glucose (Teknova), 500 µl L-glutamine 200 mM (GIBCO) were added), supplemented with 40% bovine serum and contained a suspension of 10 % cells volume of erythrocytes (RBCs).
2.3. In Vitro Growth Inhibitory Assays
The in vitro inhibitory efficacies of BPQ, ELQ-316 and ID against the survival of the T2Bo B. bovis strain were evaluated with a starting percentage of parasitized erythrocytes (PPE) of 2%. B. bovis was grown, as described above, in culture media containing different concentrations of BPQ, ELQ-316, ID, and the combinations (BPQ + ELQ-316 and ID + ELQ-316) at 25, 50, 75, 100, 150, 200, 300, 600 and 1200 nM concentrations, diluted in 100% DMSO. Parasites cultured in the presence of DMSO (1 µl) and the absence of drug compounds were used as a positive control for parasite growth. Extra wells containing uninfected bovine RBC were prepared and used as negative controls. Parasite cultures were fed daily with fresh culture medium (180 µl/well) containing the respective drug concentration. The experiments were carried out in triplicate wells for each tested concentration and control, over 96 h (4 days). PPE was monitored daily by counting parasites in Hema 3 Stat Pack (Fisher Scientific) stained thin blood smears (GBS). Before the daily change of the media, the supernatant media (180 µl) of each well was collected, and the bottom with the RBC layer was gently mixed; 1 µl of sample was taken from each well to make a thin smear, and the number of infected red blood cells was counted by microscopical examination of 5000 erythrocytes in each slide. Morphological appearance was also observed. Drug responsiveness of the parasites was measured as percent parasitemia after every 24 h exposure to each concentration of drug until 96 h.
2.4. Post-Treatment Survival
At 96 h after the first treatment, fresh medium without drug was replaced in all the culture wells, and 10 μl of fresh RBCs were added. The same procedure was done for the next five days to determine the post-treatment survival. Quantitative and qualitative parasitemia was determined by microscopic examination of Hema 3 Stat Pack (Fisher Scientific) stained thin blood smears.
2.5. Statistical Analysis
Levels of parasitemia were counted every day and the percentage of survival was calculated. Normality was assessed by Shapiro Wilk test, and media comparison between BPQ and BPQ+ ELQ-316, and ID and ID+ ELQ-316 combinations were done through independent Student's t-test. Comparison of the mean percentage of survival B. bovis against the three different drug treatments (BPQ, ELQ-316 and ID) was performed using a One-Way ANOVA test.
The doses of a drug that produces 50% inhibition (IC50) relative to the control population, and the maximal (100%) inhibitory concentration (IC100) were estimated for BPQ, ELQ-316, ID and the combinations, at concentrations ranging from 25 to 1200 nM, in 24 h, 48 h, 72 h, and 96 h post-incubation. Total inhibitory concentrations (IC100) were determined as the drug doses needed to reduce parasite growth to the same level observed in non-infected erythrocytes. (approximately 0.1%). The survival curves were compared using Log-rank (Mantel-Cox) test. The level of significance was set at <0.05. GraphPad Prism ver. 7 software for Windows (Graphpad Software Inc., San Diego, CA, USA) was used for the statistical analysis.
3. Results
3.1. Comparative In Vitro Survival Effects of BPQ, ID, ELQ-316, BPQ+ ELQ-316 and ID+ ELQ-316 Combinations on B. bovis
Parasite survival levels in 96 h of in vitro cultures in the presence of BPQ, ELQ-316, ID, and the combination BPQ + ELQ-316 and ID + ELQ-316, after starting with 2% PPE were depicted in Figure 1. The differences in the mean percentages of survival between BPQ, ID and ELQ-316 were statistically significant at all treatments except for 50 nM drug concentration. Between 75 to 300 nM, BPQ had a significantly higher effect than ID and ELQ-316 in eliminating the parasite under tested experimental conditions. In contrast, ID and ELQ-316 had similar parasiticidal kinetics but lower speed of kill at same drug concentrations. ELQ-316 had significantly higher effect in eliminating parasites in vitro than ID at 600 nM, but similar to BPQ at 1200 nM (p< 0.01).
While BPQ alone caused a gradual decrease in survival, its combination with ELQ-316 dramatically decresed parasite survival starting at 50 nM and higher concentrations (Table 1). However, the survival curve appeared to follow a sigmoid shape, with a plateau between 50 to 1200 nM (Figure 1). The data presented in Figure A1 (Appendix A) also depicts the different behavior of BPQ, BPQ+ ELQ-316 and ELQ-316 throughout time, demonstrating the clear improvement through the combination treatment when compared to treatment with ELQ-316 alone.
Comparisons between BPQ alone and the combination of BPQ+ELQ-316 are shown in Table 1. The statistically significant differences in the mean percentages of survival between two groups at 50 to 150 nM were observed through the high rate of parasite killing after the combination treatment of BPQ+ELQ316 in comparison to BPQ alone.
The calculated survival percentage values were very small in terms of parasitemia (less than 0.05%) and the data could be experimentally biased because the differences between them were minimal. Consequently, the significant difference found at 600 nM was not emphasized in the test because there was no relevant difference in the parasite counts between the two drugs beyond 150 nM.
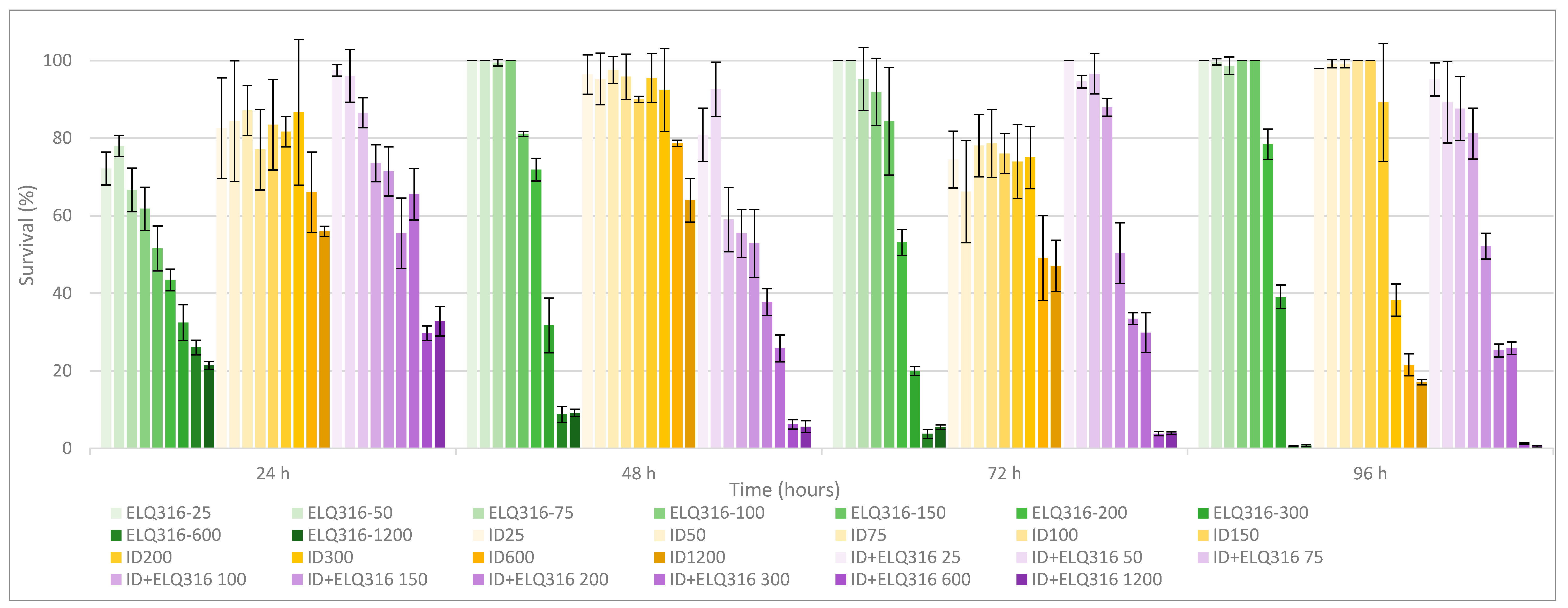
Comparative parasite survival studies demonstrated that the combination ELQ316+ID also had improved the effects of both drugs separately (Table 2), but the improvement was more gradual and became statistically significant at only higher concentrations of both drugs (100 nM to 600 nM), in comparison to the combination with BPQ. At 100 nM and higher concentrations, this combination was significatively more effective than ID alone, showing less than 1% of parasite survival al 1200 nM treatment. Similar results were observed between treatments with ELQ-316 alone and the combination ELQ316+ID, resulting in the most significant decrease in parasites survival at 600 nM (Table 2). The data in Figure A2 (Appendix A) present the different survival percentage of ID, ELQ316+ID and ELQ316 throughout time, showing the clear improvement through combination treatment in comparison to ID alone.
3.2. Drugs Potency
The mean and range of IC50 were calculated to compare drug potency on the growth of in vitro B. bovis T2Bo strain cultures. The combination of BPQ+ ELQ316 had the lower IC50 and was significantly different (p<0.05) than the IC50 of all of the drugs and combinations tested (Table 3). BPQ had the second lower IC50 but was almost two-fold lower IC50 when compared to the combination of BPQ+ELQ316. No significant differences were observed between the IC50 of ID and ELQ316, but significatively differences were found against the IC50 of these drugs and ID+ELQ316 combination (p<0.05).
3.3. Time and Concentration of Drugs to Reach 0% Survival after Treatment
The ability pf parasites to survive in in vitro cultures after drug treatments of different lengths and doses for each drug and combination, were presented in Table 4. Different treatments with BPQ, BPQ+ELQ316, ELQ316 and ID+ELQ316 treated parasites were no longer viable, reaching 0% survival at different doses and times post-incubation without drug(s). BPQ seemed to be superior to ELQ316 and ID in terms of time and concentration required to reach 0% survival. The parasites were no longer viable after 1 and 2 days of culture without BPQ in cultures treated previously with 150 nM and 200 nM, respectively. BPQ + ELQ316 combination had a superior effect than ELQ316 + ID on reaching the time to attain 0% survival. BPQ + ELQ316 combination had a superior effect than ID+ELQ316 on reaching the time to attain 0% survival. The parasites were no longer viable after 1, 2 and 3 days in BPQ + ELQ316-free media, in cultures that were treated previously with drug concentrations above 200 nM, 100 nM and 50 nM, respectively. Regarding the combination of ID+ELQ316, no parasites were found after 1 day in ID+ELQ316-free media, in cultures treated previously with concentration 600 nM, and after 4 days with concentrations 300 nM. In cultures treated previously with ID, 0% survival was reached after 4 days in ID-free media, with drug concentrations of 300 nM. No parasites were found after 1 day in ELQ316-free media cultures treated previously with drug concentrations 600 nM. At 1200 nM, there was no significant differences in timing to reach 0% survival among different drugs in free-drugs treatment cultures.
BPQ seemed to be superior to ELQ-316 and ID in terms of time and concentration required to reach 0% survival. The parasites were no longer viable after 1 and 2 days in BPQ-free media in cultures that were treated previously with drug concentrations above 200 nM and 150 nM, respectively. The combination BPQ + ELQ-316 seems to have a superior effect to ELQ-316 + ID on reaching the time to attain 0% survival. After BPQ+ELQ316 combination treatment, parasites were no longer viable after 1, 2 and 3 days of incubation in drug-free culture media from lower concentrations (from 200 nM, 100 nM and 50 nM, respectively). Regarding ID, 0% survival could be demonstrated only after 4 days of culture using drug-free media when cultures were previously treated at a concentration of 300 nM or higher. No parasites were found after 1 day without ELQ316 treatment at concentration 600 nM. With the combination ID+ELQ316, no parasites were found after 1 day without treatment at concentration 600 nM and after 4 days at concentrations 300 nM. At 1200 nM, there was no difference in the time to reach 0% survival among drugs.
4. Discussion
The shortage of approved and available drugs in most countries for the effective control of bovine babesiosis underscores the need to discover new, effective, and safer chemotherapeutic alternatives. Moreover, it is particularly important to develop alternative treatments with different mechanisms of action that can be combined to achieve radical cure and to slow down or possibly eliminate the emergence of resistance. The success of treatments depends mainly on early diagnosis, disease severity, dosage and timing of drug treatment, virulence and phenotypic/genotypic characteristics of the parasite strains involved, [50]), some of the factors that we tested in this study in vitro with different drugs and combinations.
Herein, we compared the effects of ID, BPQ and ELQ-316, and the combinations of ID+ ELQ316 and BPQ+ELQ316, on the B. bovis posttreatment survival using in vitro cultures. Findings in this study demonstrated that the combination of BPQ+ELQ 316 acted faster than each of the drugs alone, suggesting that the effects of both drugs can be enhanced by combination treatment. This finding is similar to the report on treating B. microti infected immunodeficient mice by Lawres, et al., 2016; increased efficacy of the combination of atovaquone and ELQ-316 was demonstrated with a limited capacity to generate mutations at both the Qo and the Qi sites when exposed to simultaneous drug pressure [47]. Although data in the current study shows that ELQ316 alone does not have better effects killing the parasites than BPQ at lower concentrations, a significant improvement was obtained upon combining both drugs. In addition, we found that ELQ316 can improve BPQ effects decreasing the parasites post-treatment survival from 50 nM and was similar to BPQ alone from 150 nM. As reported previously by Lawres, et la., 2016, through the radical cure of experimental B. microti in immunodeficient mice, the superior efficacy of the combination of atovaquone (ATV) and ELQ-316 in eliminating Babesia infection is a result of synergism between those drugs or a prolonged half-life of one or both compounds. Moreover, BPQ and ELQ316 combination therapy, acting on different targets of the ubiquinone-cytochrome bc1 complex, is predicted to completely block the electron transport chain through complex III, through a synergism against mitochondrial function, which is essential for parasite survival. Importantly, it was previously demonstrated that ELQ316 was superior to ATV when administered orally to eliminate T. gondii infections inside and outside the brain [45,51]. McConell, et al., 2018, in comparing treatments for toxoplasmosis suggested that the lower IC50 values of ELQs against T. gondii compared to P. falciparum may reflect differences in biologic sensitivity to cytochrome bc1 inhibition, structural differences between T. gondii and P. falciparum cyt bc1, additional targets in T. gondii, or differences in the characteristics of the assays despite a high degree of homology between the T. gondii and P. falciparum cyt bc1. If T. gondii does have greater sensitivity to cyt bc1 inhibition, it may be due T. gondii’s use of cyt bc1 for oxidative phosphorylation, while P. falciparum primarily relies on cyt bc1 for pyrimidine biosynthesis during its erythrocytic cycle [52]. The same can be applied on Babesia parasites; while it is hypothesized that T. gondii could readily cease replication and convert to a bradyzoite form because of cyt bc1 inhibition [53], the erythrocytic stages are not known to convert to a metabolically similar quiescent form. It is suggested that parasite specific structural features of cyt b contribute to susceptibility to the different ELQs [45].
BPQ potency was found with 77.06 nM (IC95%: 70.16- 86.01 nM) at 96 h post-treatment, when treated against 2% starting PPE. This value was lower than those reported by Nugraha et. al., 2019, (BPQ IC50: 135 +/- 41 nM) after 96 h of B. bovis culture at 1% starting PPE[54] , but was comparable to those reported previously by Cardillo, et al., 2024 in B. bovis using a lower starting PPE of 1% (IC50: 50 nM). In agreement with Cardillo et al, 2024, these results show a starting PPE-dependent effect, at least in in vitro culture model [19]. It remains to be determined, however, whether this observation can be applied to natural infections with fluctuating parasitemia.
ELQ316 and ID showed similar inhibitory effects in all concentrations until 300 nM at 96 h post-infection, and they consistently had comparable potencies (ID: 654,9 nM and 635.1 nM and, respectively). The mechanism of action of ID is not clearly understood (Todorovic et al. 1973), but the possible modes of action that have been proposed are related to different targets than ELQ316 [57]. ELQ316 potency value against B. bovis parasites found in our study was much higher than those reported against B. duncani (IC50 = 136.6 1 nM) [58], B. bovis (IC50=0.07 nM) [16], Besnoitia besnoiti (IC50= 7.97 nM) (Eberhard et al., 2020), and Toxoplasma gondii (IC50= 0.66 and 0.35 nM) by Alday, et al., (2017) and (IC50= 0.007 nM) [51] against the same parasite. Also, according to our study, ELQ316 alone from 600 nM concentration, greatly decreased the parasite survival. Furthermore, ELQ316 also significatively improved ID effect on the parasites post treatment survival, when used in combination with 100 nM, resulting in the reduction of almost one third of its potency (ID+ELQ316 IC50: 197 nM). These findings agree with those of Lawres, et al., 2016 [47], who reported that ELQs may prevent recrudescence in immunodeficient mice experimentally infected with B. microti [59,60].
ID has been traditionally used with a moderate efficacy and the occurrence of side effects in treating cattle babesiosis. BPQ recently showed to be more effective and faster in killing the parasite than ID in in vitro culture assays [8,27,28]. However, the in vivo effects of these two drugs and long-acting activity could be related to their persistence in cattle tissues for long duration [61]. One of the limitations of this in vitro study is whether the combination of babesicidal drugs lacking antagonistic effects, can be a possible way to slow down the emergence of drug resistance; preferably, if the drugs can inhibit the same target receptors at different sites, or if they have different mechanisms of action. On the other hand, if these combination treatments could improve both therapeutic potency and efficacy. This could be either by acting synergistically in the animal model, and achieving stronger therapeutic effects and/or decreasing the required dose, thereby reducing side effects, and the risk of developing drug resistance in the parasites [62]. A previous work performed in the B. bovis-related parasite B. microti, demonstrated that genetic alterations in the Qi binding site of the cytochrome bc1 complex (Cytb) is associated with resistance to ELQ-316 [63]. In addition, Silva et al., 2020, reported full conservation of the two canonical Qo and Qi binding sites of Cytb of the Cytb genes of B. bovis, B. bigemina, B. caballi and T. equi together with the B. microti Cytb. Thus, the combination of ELQ316 with BPQ, with action against Qi and Q0 biding sites of the cytochrome bc1 complex respectively, can potentially help to slow down the onset of resistance, or perhaps completely avoid that risk.
When tested using a starting PPE of 2 %, the parasites were no longer viable after 1 and 2 days of culture without BPQ in cultures treated previously with 200 nM and 150 nM. These results were identical to those previously obtained by Cardillo, et al., 2024. Regarding ID, 0% survival was demonstrated after 4 days of culture without drug following the use of 300 nM. The report by Cardillo et al., 2024, didn’t follow the cultures without ID more than 2 days, but in their study, 0% survival was demonstrated at ID 300 nM in cultures using 0.2% starting PPE, but 1% PPE level of parasites were still present in cultures after 2 days without drug. In cultures treated with 600 nM of ELQ316 and the combination of ID+ELQ316, no parasites were found after 1 day of culture without drug. The same was observed in cultures treated with ID+ELQ316 at 300 nM, but after 4 days without drug. Like our previous report [19], a residual effect against parasites was found after treatment in all drugs and combinations tested. This suggests a persistent effect, likely related to the extent of the impairment or metabolic change that is dependent on the concentrations of each drug used. This is important to be considered for the intermittent dose regimen in animals [64].
We found a survival effect depending on drug doses and times of exposure. We observed a decrease in survival upon longer exposures, and lack of further survival in parasites treated with increased concentrations of drugs. Except for the starting PPE, dose and time-dependence, the effects of the drugs tested herein are mostly in agreement with previous studies on B. bovis using endochin-like quinolones, tulathromycin drugs [14,16], BPQ [19] and artemisinin [65].
Overall, the findings in this study suggest that the combination of BPQ+ELQ316 is a promising candidate for in vivo testing as a new babesiacidal regimen. This is supported by several lines of evidence in our study: 1] The potency of BPQ+ELQ316 was 31.21 (IC 95%: 15.06 - 68.48). 2] BPQ+ELQ316 was 100% babesicidal after 1 day in culture after the last treatment with 200 nM concentration; 3] No parasite survival was found after 2 and 3 days of incubation without replacing the drug during daily media changes at doses 150 nM and 50 nM; 3] ELQ316 highly improved BPQ performance killing the parasites faster at lower concentration doses than all of the other drugs and combinations tested.
5. Conclusions
In summary, all drugs alone or in combinations can significantly affect (p<0.05) B. bovis survival. However, the results presented in current study reinforce the superiority of BPQ in killing B. bovis, and the potential synergy of its combination treatment with ELQ-316. Our data demonstrate that ELQ-316 in combination with BPQ is highly effective in eliminating parasitemia without relapse after drug discontinuation in vitro for 5 days. The combination treatment of BPQ+ELQ-316 is much more effective than ID +ELQ-316 suggesting increased synergism when both combined drugs target the same pathway. This combination treatment is a promising treatment option that may eliminate clinical B. bovis and needs to be tested in the animal model.
Author Contributions
NC: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Project administration, Writing–original draft, Writing–review and editing, Software. NV: Supervision, Writing–review and editing, Conceptualization, Data curation, Formal Analysis, Software. PL: Methodology, Writing–review and editing, Supervision. JD: Conceptualization, Supervision, Writing–review and editing, Resources, Validation. MR: Conceptualization, Supervision, Validation, Writing–review and editing. MU: Supervision, Writing–review and editing. CC: Supervision, Writing–review. CS: Supervision, Writing–review and editing, Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Validation, Visualization, Writing–original draft.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was supported the United States Department of Agriculture (ARS-USDA CRIS 2090- 32000-040-000-D), This project was also supported with funds from the United States Department of Veterans Affairs, Veterans Health Administration, Office of Research and Development Program Award number i01 BX003312 (MR). MR is a recipient of a VA Research Career Scientist Award (14S-RCS001). Research reported in this publication was also supported by the US National Institutes of Health under award numbers R01AI100569 and R01AI141412 (MR) and by the U.S. Department of Defense Peer Reviewed Medical Research Program (PR181134) (MR). This work was also funded by VA Merit Review Award BX004522 to JD from the US Department of Veterans Affairs Biomedical Laboratory Research and Development.
Institutional Review Board Statement
The animal study was approved by the IACUC University of Idaho #2020-42, bovine and equine bleeding. The study was conducted in accordance with the local legislation and institutional requirements.
Data Availability Statement
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
Acknowledgments
NC is a fellow of the Research Participation Program administered by the Oak Ridge Institute for Science and Technology (ORISE).
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Appendix A
Figure A1.
B. bovis T2Bo strain in vitro culture survival (starting PPE 2%) at 24, 48, 72 and 96 h, after daily addition of different concentrations of ELQ-316 (green gradients bars), BPQ (blue gradient bars) and BPQ+ELQ-316 combination (orange gradient bars). Error bars indicate the standard deviation of the means (n= 3 experiments) for each ELQ-316, BPQ and BPQ+ELQ-316 concentration (nM) tested.
Figure A1.
B. bovis T2Bo strain in vitro culture survival (starting PPE 2%) at 24, 48, 72 and 96 h, after daily addition of different concentrations of ELQ-316 (green gradients bars), BPQ (blue gradient bars) and BPQ+ELQ-316 combination (orange gradient bars). Error bars indicate the standard deviation of the means (n= 3 experiments) for each ELQ-316, BPQ and BPQ+ELQ-316 concentration (nM) tested.
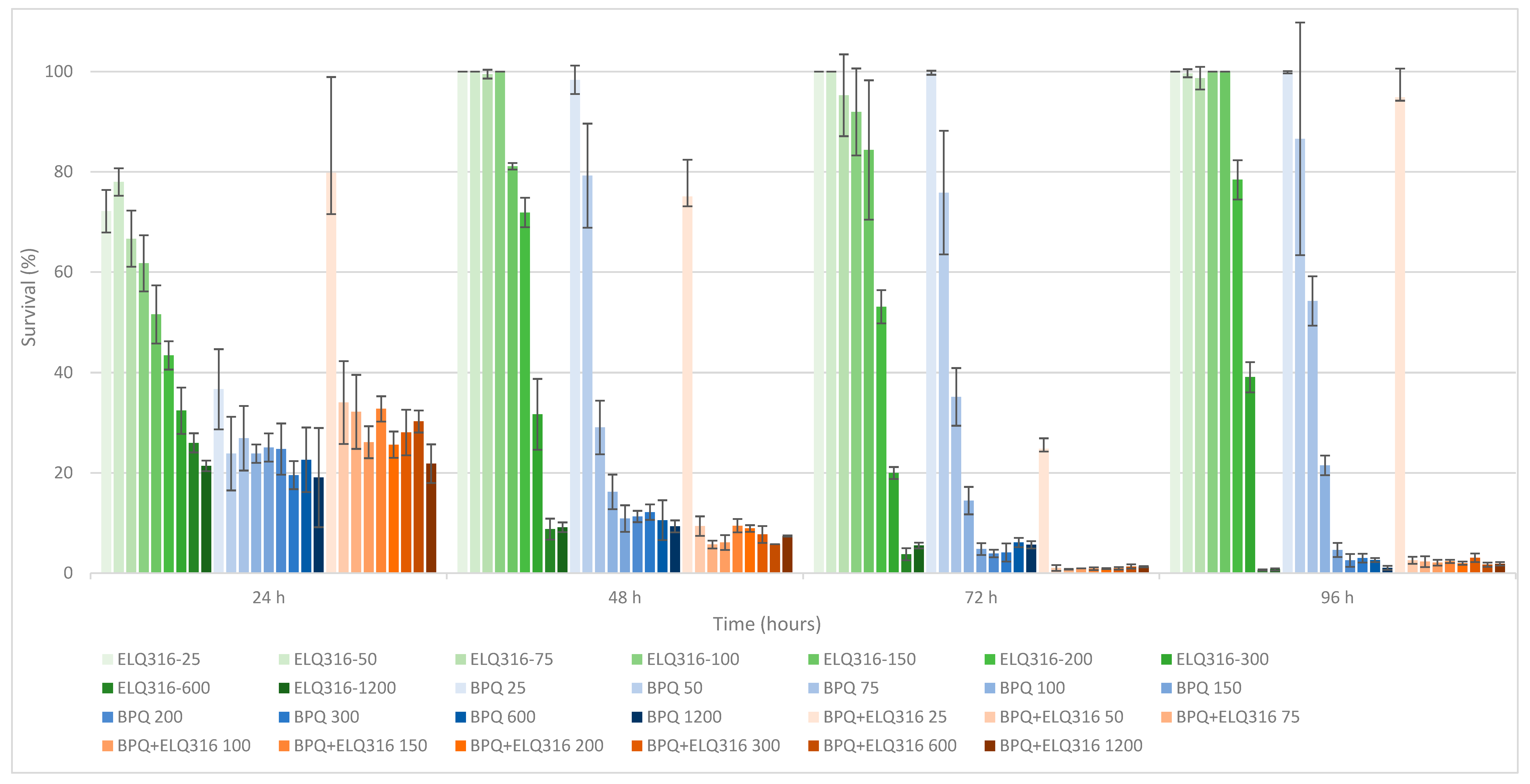
Figure A2.
B. bovis T2Bo strain in vitro culture survival (starting PPE 2%) at 24, 48, 72 and 96 h, after daily addition of different concentrations of ELQ-316 (green gradients bars), ID (blue gradient bars) and ID+ELQ-316 combination (pink gradient bars). Error bars indicate the standard deviation of the means (n= 3 experiments) for each ELQ316, ID and ID+ELQ-316 concentration (nM) tested.
Figure A2.
B. bovis T2Bo strain in vitro culture survival (starting PPE 2%) at 24, 48, 72 and 96 h, after daily addition of different concentrations of ELQ-316 (green gradients bars), ID (blue gradient bars) and ID+ELQ-316 combination (pink gradient bars). Error bars indicate the standard deviation of the means (n= 3 experiments) for each ELQ316, ID and ID+ELQ-316 concentration (nM) tested.
References
- Pipano, E.; Hadani, A. Control of Bovine Babesiosis. In Malaria and Babesiosis. Ristic, M. Ambroise-Thomas, P. Kreier, J.P.; 1984; pp. 263–303.
- Bock, R.; Jackson, L.; De Vos, A.; Jorgensen, W. Babesiosis of Cattle. Parasitology 2004, 129, S247–S269. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.S.; Sengupta, P.P.; Paramanandham, K.; Suresh, K.P.; Chamuah, J.K.; Rudramurthy, G.R.; Roy, P. Bovine Babesiosis: An Insight into the Global Perspective on the Disease Distribution by Systematic Review and Meta-Analysis. Vet Parasitol 2020, 283, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Asrar, R.; Farhan, H.R.; Sultan, D.; Ahmad, M.; Hassan, S.; Kalim, F.; Shakoor, A.; Muhammad Taimoor Ihsan, H.; Shahab, A.; Ali, W.; et al. Review Article Bovine Babesiosis; Review on Its Global Prevalence and Anticipatory Control for One Health. Continental Vet J 2022, 2, 42–49. [Google Scholar]
- Smith, R.D.; Evans, D.E.; Martins, J.R.; Ceresér, V.H.; Correa, B.L.; Cardozo, C.P.H.; Solari, M.A.; Nari, A. Babesiosis (Babesis Bovis) Stability in Unstable Environments. Ann N Y Acad Sci 2000, 916, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Zintl, A.; Mcgrath, G.; O’grady, L.; Fanning, J.; Downing, K.; Roche, D.; Casey, M.; Gray, J.S. Changing Incidence of Bovine Babesiosis in Ireland. Ir Vet J 2014, 67, 1–14. [Google Scholar] [CrossRef]
- Todorovic, R.A.; Viscaino, O.G.; Gonzalez, E.F.; Adams, L.G. Chemoprophylaxis (Imidocarb) against Babesia Bigemina and Babesia Argentina Infections. Am J Vet Res 1973, 34, 1153–1161. [Google Scholar]
- Vial, H.J.; Gorenflot, A. Chemotherapy against Babesiosis. Vet Parasitol 2006, 138, 147–160. [Google Scholar] [CrossRef]
- Almazan, C.; Tipacamu, G.A.; Rodriguez, S.; Mosqueda, J.; Perez De Leon, A. Immunological Control of Ticks and Tick-Borne Diseases That Impact Cattle Health and Production. Frontiers in Bioscience 2018, 23, 1535–1551. [Google Scholar] [CrossRef]
- Alvarez, J.A.; Rojas, C.; Figueroa, J.V. An Overview of Current Knowledge on in Vitro Babesia Cultivation for Production of Live Attenuated Vaccines for Bovine Babesiosis in Mexico. Front Vet Sci 2020, 7, 1–12. [Google Scholar] [CrossRef]
- Azhar, M.; Gadahi, J.A.; Bhutto, B.; Tunio, S.; Vistro, W.A.; Tunio, H.; Bhutto, S.; Ram, T. Babesiosis: Current Status and Future Perspectives in Pakistan and Chemotherapy Used in Livestock and Pet Animals. Heliyon 2023, e17172, 1–16. [Google Scholar] [CrossRef]
- Mosqueda, J.; Olvera-Ramírez, A.; Aguilar-Tipacamú, G.; Cantó, G.J. Current Advances in Detection and Treatment of Babesiosis; 2012; Vol. 19;
- Kuttler, K.L.; Aliu, Y.O. Chemotherapy of Babesiosis. In Malaria and Babesiosis. Ristic, M., Kreier, J.P., Eds. Academic Press: New York, NY, USA; 1984; pp. 151–172.
- Silva, M.G.; Villarino, N.F.; Knowles, D.P.; Suarez, C.E. Assessment of Draxxin® (Tulathromycin) as an Inhibitor of in Vitro Growth of Babesia Bovis, Babesia Bigemina and Theileria Equi. Int J Parasitol Drugs Drug Resist 2018, 8, 265–270. [Google Scholar] [CrossRef]
- Sears, K.; Knowles, D.; Dinkel, K.; Mshelia, P.W.; Onzere, C.; Silva, M.; Fry, L. Imidocarb Dipropionate Lacks Efficacy against Theileria Haneyi and Fails to Consistently Clear Theileria Equi in Horses Co-Infected with T. Haneyi. Pathogens 2020, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.G.; Bastos, R.G.; Stone Doggett, J.; Riscoe, M.K.; Pou, S.; Winter, R.; Dodean, R.A.; Nilsen, A.; Suarez, C.E. Endochin-like Quinolone-300 and ELQ-316 Inhibit Babesia Bovis, B. Bigemina, B. Caballi and Theileria Equi. Parasit Vectors 2020, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Aguado-Martinez, A.; Manser, V.; Balmer, V.; Winzer, P.; Ritler, D.; Hostettler, I.; Solís, D.A.; Ortega-Mora, L.; Hemphill, A. Buparvaquone Is Active against Neospora Caninum in Vitro and in Experimentally Infected Mice. Int J Parasitol Drugs Drug Resist 2015, 5, 16–25. [Google Scholar] [CrossRef]
- Rizk, M.A.; El-Sayed, S.A.E.-S.; Igarashi, I. Diminazene Aceturate and Imidocarb Dipropionate-Based Combination Therapy for Babesiosis – A New Paradigm. Ticks Tick Borne Dis 2023, 14, 102145. [Google Scholar] [CrossRef] [PubMed]
- Cardillo, N.M.; Lacy, P.A.; Villarino, N.F.; Doggett, J.S.; Riscoe, M.K.; Bastos, R.G.; Laughery, J.M.; Ueti, M.W.; Suarez, C.E. Comparative Efficacy of Buparvaquone and Imidocarb in Inhibiting the in Vitro Growth of Babesia Bovis. Front Pharmacol 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Belloli, C.; Crescenzo, G.; Lai, O.; Carofiglio, V.; Marang, O.; Ormas, P. Pharmacokinetics of Imidocarb Dipropionate in Horses after Intramuscular Administration. Equine Vet J 2002, 34, 625–629. [Google Scholar] [CrossRef]
- Mdachi, R.E.; Murilla, G.A.; Omukuba, J.N.; Cagnolati, V. Disposition of Diminazene Aceturate (Berenil®) in Trypanosome-Infected Pregnant and Lactating Cows. Vet Parasitol 1995, 58, 215–225. [Google Scholar] [CrossRef]
- Hwang, S.-J.; Yamasaki, M.; Nakamura, K.; Sasaki, N.; Murakami, M.; Kumara, B.; Rajapakshage, W.; Ohta, H.; Maede, Y.; Takiguchi, M. Development and Characterization of a Strain of Babesia Gibsoni Resistant to Diminazene Aceturate In Vitro. J. Vet. Med. Sci 2010, 72, 765–771. [Google Scholar] [CrossRef]
- Tuntasuvan, D.; Jarabrum, W.; Viseshakul, N.; Mohkaew, K.; Borisutsuwan, S.; Theeraphan, A.; Kongkanjana, N. Chemotherapy of Surra in Horses and Mules with Diminazene Aceturate. Vet Parasitol 2003, 110, 227–233. [Google Scholar] [CrossRef]
- Pérez de León, A.A.; Strickman, D.A.; Knowles, D.P.; Fish, D.; Thacker, E.; de la Fuente, J.; Krause, P.J.; Wikel, S.K.; Miller, R.S.; Wagner, G.G.; et al. One Health Approach to Identify Research Needs in Bovine and Human Babesioses: Workshop Report. Parasit Vectors 2010, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pérez de Leon, A.A.; Teel, P.D.; Auclair, A.N.; Messenger, M.T.; Guerrero, F.D.; Schuster, G.; Miller, R.J. Integrated Strategy for Sustainable Cattle Fever Tick Eradication in USA Is Required to Mitigate the Impact of Global Change. Front Physiol 2012, 3, 1–17. [Google Scholar] [CrossRef] [PubMed]
- CVM- FDA FOI Summary for Original Approval of IMIZOL - NADA 141-071; 1997.
- Todorovic, R.A.; Viscaino, O.G.; Gonzalez, E.F.; Adams, L.G. Chemoprophylaxis (Imidocarb) against Babesia Bigemina and Babesia Argentina Infections. Am J Vet Res 1973, 34, 1153–1161. [Google Scholar] [PubMed]
- Adams, L.; Corrier, D.; Williams, J. A Study of the Toxicity of Imidocarb Dipropionate in Cattle. Res Vet Sci. 1980, 28, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Yu, N.; Liu, C.; Han, M.; Wang, H.; Chen, X.; Kang, J.; Li, X.; Liu, Y. Residue Depletion of Imidocarb in Bovine Tissues by UPLC-MS/MS. Animals 2023, 13. [Google Scholar] [CrossRef]
- Coldham, N.G.; Moore, A.S.; Sivapathasundaram, S.; Sauer, M.J. Lmidocarb Depletion from Cattle Liver and Mechanism of Retention in Isolated Bovine Hepatocytes*t; 1994; Vol. 119;
- Fray, M.; Pudney, M. Site of Action of the Antimalarial Hydroxynaphthoquinone,2- [Trans-4-(40chlorophenyl)Cyclohexyl]-3-Hydroxy-1,4,-Naphthoquinone (566c80). Biochem. Pharmacol. 1992, 40, 914–919. [Google Scholar] [CrossRef]
- Hudson, A.T.; Randall, A.W.; Fry, M.; Ginger, C.D.; Hill, B.; Latter, V.S.; McHardy, N.; Williams, R.B. Novel Anti-Malarial Hydroxynpahthoquinones with Potent Broad Spectrum Anti-Protozoal Activity. Parasitology 1985, 45–55. [Google Scholar] [CrossRef]
- Sharifiyazdi, H.; Namazi, F.; Oryan, A.; Shahriari, R.; Razavi, M. Point Mutations in the Theileria Annulata Cytochrome b Gene Is Associated with Buparvaquone Treatment Failure. Vet Parasitol 2012, 187, 431–435. [Google Scholar] [CrossRef]
- Mhadhbi, M.; Chaouch, M.; Ajroud, K.; Darghouth, M.A.; BenAbderrazak, S. Sequence Polymorphism of Cytochrome b Gene in Theileria Annulata Tunisian Isolates and Its Association with Buparvaquone Treatment Failure. PLoS One 2015, 10, 1–11. [Google Scholar] [CrossRef]
- Carter, P. Assessment of the Efficacy of Buparvaquone for the Treatment of “benign” Bovine Theileriosis. Tick Fever Centre Department of Employment, Economic Development and Innovation. 2011, 1–12.
- Müller, J.; Aguado-Martínez, A.; Manser, V.; Wong, H.N.; Haynes, R.K.; Hemphill, A. Repurposing of Antiparasitic Drugs: The Hydroxy-Naphthoquinone Buparvaquone Inhibits Vertical Transmission in the Pregnant Neosporosis Mouse Model. Vet Res 2016, 47, 1–8. [Google Scholar] [CrossRef]
- Muraguri, G.R.; Ngumi, P.N.; Wesonga, D.; Ndungu, S.G.; Wanjohi, J.M.; Bang, K.; Fox, A.; Dunne, J.; McHardy, N. Clinical Efficacy and Plasma Concentrations of Two Formulations of Buparvaquone in Cattle Infected with East Coast Fever (Theileria Parva Infection). Res Vet Sci 2006, 81, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Carter, P. Animal Health Assessment of the Efficacy of Buparvaquone for the Treatment of “benign” Bovine Theileriosis; 2011. ISBN 9781741916591.
- Bailey, G. Buparvaquone Tissue Residue Study; 2013. ISBN 9781925045475.
- Wilkie, G.M.; Kirvar, E.; Thomas, E.M.; Sparagano, O.; Brown, C.G.D. Stage-Specific Activity in Vitro on the Theileria Infection Process of Serum from Calves Treated Prophylactically with Buparvaquone. Vet Parasitol 1998, 80, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Goud, S.K.; Vijayakumar, K.; Davis, J.; Tresamol, P.V.; Ravishankar, C.; Devada, K. Efficacy of Different Treatment Regimens against Oriental Theileriosis in Naturally Infected Cattle. Indian J. Vet. Med 2021, 40, 14–19. [Google Scholar]
- Brasseur, P.; Lecoublet, S.; Kapel, N.; Favennec, L.; Ballet, J.J. In Vitro Evaluation of Drug Susceptibilities of Babesia Divergens Isolates. Antimicrob Agents Chemother 1998, 42, 818. [Google Scholar] [CrossRef]
- Hasheminasab, S.S.; Moradi, P.; Wright, I. A Four Year Epidemiological and Chemotherapy Survey of Babesiosis and Theileriosis, and Tick Vectors in Sheep, Cattle and Goats in Dehgolan, Iran. Ann Parasitol 2018, 64, 43–48. [Google Scholar] [CrossRef]
- Shah, N.D.; Bhikane, A.U.; Jadhav, R.K.; Chavhan, S.G.; Mohan, A. Therapeutic Management of Babesiosis Alone and Its Mixed Infection with Theileriosis in Cattle. Indian J. Vet. Med. Vol. 2019, 39, 26–31. [Google Scholar]
- McConnell, E.V.; Bruzual, I.; Pou, S.; Winter, R.; Dodean, R.A.; Smilkstein, M.J.; Krollenbrock, A.; Nilsen, A.; Zakharov, L.N.; Riscoe, M.K.; et al. Targeted Structure–Activity Analysis of Endochin-like Quinolones Reveals Potent Qi and Qo Site Inhibitors of Toxoplasma Gondii and Plasmodium Falciparum Cytochrome Bc 1 and Identifies ELQ-400 as a Remarkably Effective Compound against Acute Experimental Toxoplasmosis. ACS Infect Dis 2018, 4, 1574–1584. [Google Scholar] [CrossRef]
- Fisher, N.; Meunier, B.; Biagini, G.A. The Cytochrome Bc1 Complex as an Antipathogenic Target. FEBS Lett 2020, 594, 2935–2952. [Google Scholar] [CrossRef]
- Lawres, L.A.; Garg, A.; Kumar, V.; Bruzual, I.; Forquer, I.P.; Renard, I.; Virji, A.Z.; Boulard, P.; Rodriguez, E.X.; Allen, A.J.; et al. Radical Cure of Experimental Babesiosis in Immunodeficient Mice Using a Combination of an Endochin-like Quinolone and Atovaquone. Journal of Experimental Medicine 2016, 213, 1307–1318. [Google Scholar] [CrossRef]
- Nilsen, A.; Miley, G.P.; Forquer, I.P.; Mather, M.W.; Katneni, K.; Li, Y.; Pou, S.; Pershing, A.M.; Stickles, A.M.; Ryan, E.; et al. Discovery, Synthesis, and Optimization of Antimalarial 4(1 H)-Quinolone-3-Diarylethers. J Med Chem 2014, 57, 3818–3834. [Google Scholar] [CrossRef]
- Levy, M.G.; Ristic, M. Babesia Bovis: Continuous Cultivation in a Microaerophilous Stationary Phase Culture. Science (1979) 1980, 207, 1218–1220. [Google Scholar] [CrossRef] [PubMed]
- Mehlhorn, H. Babesiacidal Drugs. In Encyclopedia of Parasitology; Springer Berlin Heidelberg, 2016; pp. 1–11.
- Doggett, J.S.; Nilsen, A.; Forquer, I.; Wegmann, K.W.; Jones-Brando, L.; Yolken, R.H.; Bordón, C.; Charman, S.A.; Katneni, K.; Schultz, T.; et al. Endochin-like Quinolones Are Highly Efficacious against Acute and Latent Experimental Toxoplasmosis. Proc Natl Acad Sci U S A 2012, 109, 15936–15941. [Google Scholar] [CrossRef] [PubMed]
- Painter, H.J.; Morrisey, J.M.; Mather, M.W.; Vaidya, A.B. Specific Role of Mitochondrial Electron Transport in Blood-Stage Plasmodium Falciparum. [CrossRef]
- Tomavo, S.; Boothroydt, J.C. Interconnection between Organellar Functions, Development and Drug Resistance in the Protozoan Parasite, Toxoplasma Gondii; 1995; Vol. 25;
- Nugraha, A.B.; Tuvshintulga, B.; Guswanto, A.; Tayebwa, D.S.; Rizk, M.A.; Gantuya, S.; El-Saber Batiha, G.; Beshbishy, A.M.; Sivakumar, T.; Yokoyama, N.; et al. Screening the Medicines for Malaria Venture Pathogen Box against Piroplasm Parasites. Int J Parasitol Drugs Drug Resist 2019, 10, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Yu, N.; Liu, C.; Han, M.; Wang, H.; Chen, X.; Kang, J.; Li, X.; Liu, Y. Residue Depletion of Imidocarb in Bovine Tissues by UPLC-MS/MS. Animals 2023, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Coldham, N.G.; Moore, A.S.; Sivapathasundaram, S.; Sauer, M.J. Lmidocarb Depletion from Cattle Liver and Mechanism of Retention in Isolated Bovine Hepatocytes. Analyst 1994, 119, 2549–2552. [Google Scholar] [CrossRef]
- Hudson, A.T.; Randall, A.W.; Fry, M.; Ginger, C.D.; Hill, B.; Latter, V.S.; McHardy, N.; Williams, R.B. Novel Anti-Malarial Hydroxynpahthoquinones with Potent Broad Spectrum Anti-Protozoal Activity. Parasitology 1985, 90, 45–55. [Google Scholar] [CrossRef]
- Chiu, J.E.; Renard, I.; Pal, A.C.; Singh, P.; Vydyam, P.; Thekkiniath, J.; Kumar, M.; Gihaz, S.; Pou, S.; Winter, R.W.; et al. Effective Therapy Targeting Cytochrome Bc1prevents Babesia Erythrocytic Development and Protects from Lethal Infection. Antimicrob Agents Chemother 2021, 65, 1–10. [Google Scholar] [CrossRef]
- Nilsen, A.; Lacrue, A.N.; White, K.L.; Forquer, I.P.; Cross, R.M.; Marfurt, J.; Mather, M.W.; Delves, M.J.; Shackleford, D.M.; Saenz, F.E.; et al. Quinolone-3-Diarylethers: A New Class of Drugs for a New Era of Malaria Eradication. [CrossRef]
- Miley, G.P.; Pou, S.; Winter, R.; Nilsen, A.; Li, Y.; Kelly, J.X.; Stickles, A.M.; Mather, M.W.; Forquer, I.P.; Pershing, A.M.; et al. ELQ-300 Prodrugs for Enhanced Delivery and Single-Dose Cure of Malaria. Antimicrob Agents Chemother 2015, 59, 5555–5560. [Google Scholar] [CrossRef]
- McDougall, S.; Hillerton, J.E.; Pegram, D. Concentrations of Buparvaquone in Milk and Tissue of Dairy Cows. N Z Vet J 2016, 64, 318–323. [Google Scholar] [CrossRef]
- Tallarida, R.J. Drug Synergism and Dose-Effect Data Analysis. Drug Synergism and Dose-Effect Data Analysis. [CrossRef]
- Winter, R.; Kelly, J.X.; Smilkstein, M.J.; Hinrichs, D.; Koop, D.R.; Riscoe, M.K. Optimization of Endochin-like Quinolones for Antimalarial Activity. [CrossRef]
- Proma, F.H.; Shourav, M.K.; Choi, J. Post-Antibiotic Effect of Ampicillin and Levofloxacin to Escherichia Coli and Staphylococcus Aureus Based on Microscopic Imaging Analysis. Antibiotics 2020, 9, 1–13. [Google Scholar] [CrossRef]
- Mazuz, M.L.; Golenser, J.; Fish, L.; Haynes, R.K.; Wollkomirsky, R.; Leibovich, B.; Shkap, V. Artemisone Inhibits in Vitro and in Vivo Propagation of Babesia Bovis and B. Bigemina Parasites. Exp Parasitol 2013, 135, 690–694. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Comparative survival kinetics (mean %) of in vitro B. bovis Texas strain culture during 96 h incubation in the presence of different concentrations of BPQ, ID, ELQ-316, BPQ+ELQ-316 and ID+ELQ-316 combinations, after starting at 2% percentage of parasitemia (PPE). Error bars indicate the standard deviation of the means (n= 3 experiments) for each drug and combinations tested.
Figure 1.
Comparative survival kinetics (mean %) of in vitro B. bovis Texas strain culture during 96 h incubation in the presence of different concentrations of BPQ, ID, ELQ-316, BPQ+ELQ-316 and ID+ELQ-316 combinations, after starting at 2% percentage of parasitemia (PPE). Error bars indicate the standard deviation of the means (n= 3 experiments) for each drug and combinations tested.
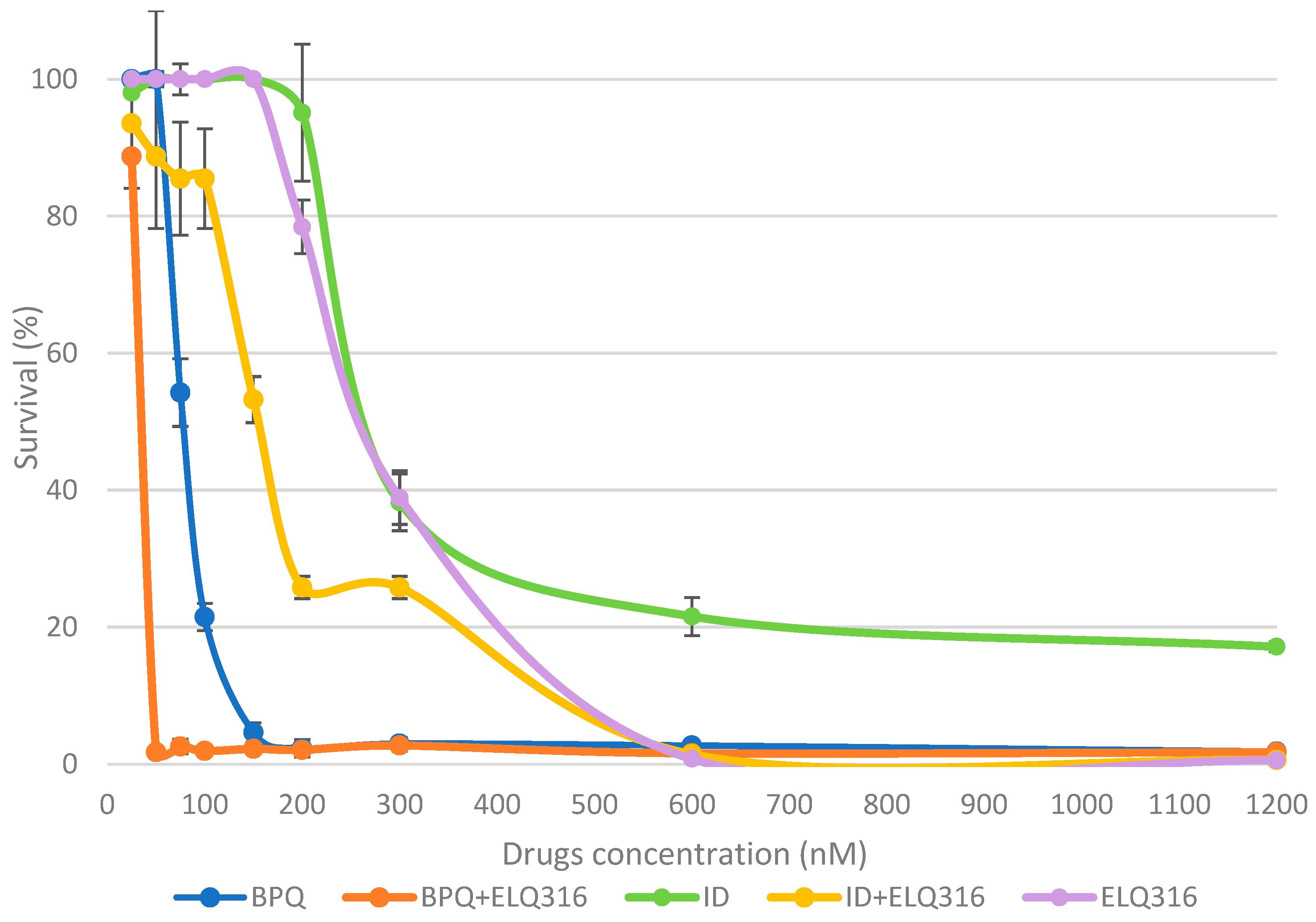

Table 1.
Comparative survival rate (%) of 96 h in vitro B. bovis Texas strain culture after starting at 2% PPE in the presence of different concentrations of BPQ and combination of BPQ+ELQ316.
Table 1.
Comparative survival rate (%) of 96 h in vitro B. bovis Texas strain culture after starting at 2% PPE in the presence of different concentrations of BPQ and combination of BPQ+ELQ316.
| Drug Treatment Concentration#break#(nM) | BPQ | BPQ+ ELQ-316 | p-value | ||
|---|---|---|---|---|---|
| Mean (%) | CI 95% | Mean (%) | CI 95% | ||
| 25 | 99.87 | (99.78 - 99.96) | 97.31 | (96.16 – 98.46) | p=0.72 |
| 50 | 86.6 | (77.56 – 95.64) | 2.9 | (2.8 - 3.00) | p<0.05 |
| 75 | 54.25 | (52.33 – 56.17) | 2.31 | (1.89 – 2.73) | p<0.01 |
| 100 | 21.5 | (20.74 – 22.26) | 2.1 | (1.87 – 2.33) | p<0.01 |
| 150 | 4.64 | (4.1 – 5.18) | 2.37 | (2.24 – 2.50) | p<0.05 |
| 200 | 2.55 | (2.05 – 3.05) | 1.99 | (1.86 - 2.12) | p=0.51 |
| 300 | 3.01 | (2.67 – 3.35) | 3.06 | (2.73 – 3.39) | p=0.94 |
| 600 | 2.62 | (2.51 – 2.73) | 1.72 | (1.60 – 1.84) | p<0.01 |
| 1200 | 1.08 | (0.88 – 1.28) | 1.88 | (1.75 - 2.01) | p=0.1 |
Note: These results were expressed as survival percentages.
Table 2.
Comparative survival rate (%) of 96 h in vitro B. bovis Texas strain culture after starting at 2% PPE in the presence of different concentrations of ID and combination of ID + ELQ-316.
Table 2.
Comparative survival rate (%) of 96 h in vitro B. bovis Texas strain culture after starting at 2% PPE in the presence of different concentrations of ID and combination of ID + ELQ-316.
| Drug Treatment Concentration (nM) | ID | ELQ316 + ID | p-value | ||
|---|---|---|---|---|---|
| Mean (%) | CI 95% | Mean (%) | CI 95% | ||
| 25 | 98.04 | N/A | 95.16 | (93.5 – 96.82) | p=0.36 |
| 50 | 99.22 | (98.90 – 99.54) | 89.25 | (85.16 – 93.34) | p=0.24 |
| 75 | 99.22 | N/A | 87.63 | (84.41 – 90.85) | p=0.13 |
| 100 | 100 | N/A | 81.18 | (79.37 – 82.99) | p<0.05 |
| 150 | 100 | N/A | 52.15 | (50.84 – 53.46) | p<0.01 |
| 200 | 89.22 | (77.98 – 100) | 25.27 | (24.81 – 25.73) | p<0.05 |
| 300 | 38.24 | (36.26 – 40.22) | 25.81 | (25.18 – 26.44) | p<0.05 |
| 600 | 21.57 | (20.25 – 22.89) | 1.33 | (1.29 - 1.37) | p<0.05 |
| 1200 | 17.16 | (16.83 – 17.49) | 0.59 | (0.49 – 0.69) | p<0.01 |
N/A not applicable: CI are not reported for this concentration’s groups because the mathematical models did not fit the data adequately.
Table 3.
Drug potency (IC50) of different drugs and their combinations on the growth of 96 h in vitro B. bovis Texas strain culture after starting at 2% PPE.
Table 3.
Drug potency (IC50) of different drugs and their combinations on the growth of 96 h in vitro B. bovis Texas strain culture after starting at 2% PPE.
| Drug | IC50 (nM) | |
|---|---|---|
| Mean | 95% CI | |
| BPQ | 77.06 | (70.16 - 86.01) |
| BPQ + ELQ-316 | 31.21 | (15.06 - 68.48) |
| ID | 635.1 | (280.9 – 2119) |
| ELQ-316 | 654.9 | (362.3 – 1411) |
| ID + ELQ-316 | 197 | (129.0 - 311.2) |
Table 4.
Range of concentrations and time in which they reach 0% survival.
| Single drugs and combination treatments | Time post-treatment without drug (h) | ||||
|---|---|---|---|---|---|
| 24 | 48 | 72 | 96 | 120 | |
| Control | N/A | ||||
| BPQ | 200 to 1200 | 150 to 1200 | |||
| BPQ + ELQ-316 | 200 to 1200 | 100 to 1200 | 50 to 1200 | ||
| ELQ-316 | 600 to 1200 | ||||
| ID+ELQ-316 | 600 to 1200 | 300 to 1200 | |||
| ID | 1200 | 300 to 1200 | |||
N/A not applicable: controls have never reach 0% survival during the time of the study (120 h).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



