
Submitted:
29 September 2024
Posted:
30 September 2024
You are already at the latest version
Abstract
A specialized microenvironment in the bone marrow, composed of stromal cells including mesenchymal stem cells (MSCs), support hematopoietic stem cells (HSCs) self-renewal and differentiation but also regulates leukemia development and progression. The reciprocal direct interaction between MSCs and CD34+ HSCs under physiological and pathological conditions has not been well characterized yet. Here, we established a direct co-culture model between MSCs and CD34+ HSCs or MSCs and acute myeloid leukemia cells (THP-1, Molm-13, and primary cells from patients) to study cell-cell communication. Following MSCs-CD34+ HSCs co-culture, the expression of adhesion markers N-cadherin and connexin 43 increased in both cell types, forming gap junction channels. Moreover, the clonogenic potential of CD34+ HSCs was increased. However, direct contact of acute myeloid leukemia cells with MSCs reduced the expression levels of connexin 43 and N-cadherin in MSCs. The impairment in gap junction formation could be ascertained to a defect in the acute myeloid leukemia derived MSCs. Interestingly, CD34+ HSCs and acute myeloid leukemia cell lines restrained MSCs from engaging into osteoblastic differentiation upon prolonged direct cell-cell contact. In conclusion, under physiological conditions, connexin 43 and N-cadherin interaction preserves stemness of both CD34+ HSCs and MSCs, a process that is compromised in acute myeloid leukemia, pointing out to the possible role of gap junctions in modulating stemness.
Keywords:
Bone marrow
; microenvironment
; CD34+ hematopoietic stem cells
; mesenchymal stem cells
; connexin 43
; N-cadherin
; gap junction
; acute myeloid leukemia
; direct contact
Introduction
Hematopoietic stem cells (HSCs) give rise to all blood cells and their progenies and reside in a special microenvironment in the bone marrow (BM) [1]. Under physiological conditions, most HSCs are quiescent, and only a small fraction of HSCs enters the cell cycle and differentiates into multipotent progenitors [2,3,4,5,6]. Intrinsic or extrinsic signals induce the differentiation of these progenitors to common lymphoid and myeloid progenitors under the so-called process of hematopoiesis [7]. In hematological malignancies, aberrations in the bone marrow microenvironment limit the regeneration and differentiation potential of HSCs by reducing their numbers and/or function [8]. Therefore, the hematopoietic system is well-organized in a hierarchical differentiation cascade, where each phase in the differentiation process requires a different combination of cytokines, chemokines, extracellular matrix (ECM) interactions and cell-cell interactions, which are offered by different stromal cell types [9]. Mesenchymal stem cells (MSCs) are the pivotal stromal cellular players of the endosteal niche. They are in intimate interaction with HSCs, maintaining their stem cell quiescence, self-renewal, apoptosis and differentiation properties through the production of cytokines, chemokines, and ECM [3]. Several studies have shown that the indirect interaction between MSCs and HSCs is maintained by the secretion of thrombopoietin (TPO), stem cell factor (SCF), and fms-like tyrosine kinase 3/fetal liver kinase-2 (FLT3/Flk2) cytokines that regulate the function of HSCs [10,11]. Moreover, MSCs possess C-X-C motif chemokine 12/ stromal-derived factor 1 (CXCL12/SDF1), SCF/c-Kit and Slit-2/Robo-4 chemokines which play a role in the trafficking and homing of HSCs [2,4,5,12,13]. In addition to the production of a large array of regulatory molecules, recent studies document the role of MSCs in affecting the function of HSCs through direct contact [7,14]. Communication of cells by cellular contacts could be mediated through gap junctions which are membrane proteins that mediate exchanges of small molecules (<1KDa), secondary messengers, and ions between various cells, for example, Ca2+, cyclic adenosine monophosphate (cAMP), and inositol triphosphate (IP3) [15]. Cell-cell communication through gap junctions between HSCs and MSCs or stromal cells in the hematopoietic microenvironment of the human system is sparse. Yet, functional connexin (Cx)-43 gap junctions exist between stromal cells and immature hematopoietic progenitor cells in humans [16], and Cx-43 and Cx-37 mRNA are expressed at low levels in human BM and cord blood hematopoietic progenitor CD34+ cells [15,17]. This provides evidence of an important regulatory pathway of hematopoiesis through gap junctions in humans. Studies have shown that cadherins are major cell adhesion molecules responsible for Ca2+-dependent cell-cell interaction [18,19]. The association between gap junctions and adherens junctions, particularly between Cx43 and N-cadherin, is required for gap junction assembly and function [20]. The role of N-cadherin in HSCs-BM niche interactions is debatable. While some studies showed that N-cadherin conditional knockout mice had no observable phenotype in HSCs maintenance and hematopoiesis [21], others demonstrated that the inhibition of N-cadherin expression reduced the anchoring of HSCs to the endosteal surface and inhibited their long-term engraftment [22].
In acute myeloid leukemia (AML), which is a heterogeneous clonal disorder characterized by an increase in the number of leucoblast cells featuring cytogenetic aberrations and recurrent somatic mutations [23,24], the role of the BM niche is well documented.
Intercellular communication with stromal cells affects AML cell proliferation, apoptosis [25] and increases their drug resistance [26], thus promoting AML relapse [27]. Weber et al. demonstrated that the direct interaction of human myeloid leukemia cell lines with BM stromal cells decreased leukemic cell differentiation [28]. Recently, a study showed that gap junction inhibition reduced chemoresistance to cytarabine, used in standard chemotherapy regimens against AML [17] in BM-MSCs. Moreover, the disruption of gap junction interaction between AML cells and stromal cells, by carbenoxolone (a gap junction inhibitor), abridged the induced-chemoresistance by MSCs, suggesting the important role of gap junctions in AML chemoresistance [28].
Although the role of soluble mediators was extensively studied in the differentiation process of HSCs and in the promotion of leukemic cells [29,30], very little is known about the role of direct communication via gap junctions in modulating these two processes. In this study, we investigated the role of MSCs in modulating HSCs differentiation, via direct interactions through N-cadherin and Cx-43 under physiological conditions. We also examined whether the cell-cell interaction between MSCs and AML cells is differentially regulated under pathological conditions such as AML.
Materials and Methods
Isolation and Phenotypic Identification of CD34+ Cells
The study protocol was approved by the Institutional Review Board of the American University of Beirut. Based on the difficulty to voluntarily obtain mobilized peripheral blood from normal donors, frozen mobilized peripheral blood from deceased leukemic patients was recuperated and used in this study. Frozen granulocyte colony-stimulating factor (G-CSF) mobilized peripheral blood of deceased leukemia patients was collected. Blood was layered on ficoll-Paque (Sigma-Aldrich, USA) density gradient and centrifuged at 400 x g for 30 minutes. Mononuclear cells were recuperated, washed, and stained with different antibodies.
FITC mouse anti-CD38, APC mouse anti-CD45 and PE anti-CD34 (BD, USA) were used for separate immunophenotyping of cells. The isotype control antibodies were either FITC mouse IgG2a or APC mouse IgG1 (BD, USA). For each staining, 10000 events were acquired by flow cytometry.
For isolation of CD34+ cells, cells with small size and granularity were selected (according to HSC size less than 10 µm [31] and further analyzed for the expression of the CD34 marker. CD34+ cells were sorted using FACS Aria SORP cell sorter (BD, USA). The purity of isolated CD34+ cells routinely ranged between 90 and 99%. The percentage of CD34+ cells varied between 0.5 and 4% among patients diagnosed for T-cell Lymphoma, Hodgkin Lymphoma, Non-Hodgkin Lymphoma, Lymphoma, Multiple Myeloma, Ewing Sarcoma and Neuroblastoma. Moreover, immunophenotypic characterization of differentiation markers indicated that these cells were an enriched population of HSCs with high self-renewing capacity since at least 50% of CD34+ cells lack the expression of CD45 and CD38 markers (Supplementary Table 1). In this study, we focused on CD34+ cells isolated from Multiple Myeloma patients, since the mutations causing the disease affect the differentiated cells and not the CD34 progenitors [32].
To isolate an enriched population of CD34+ HSCs, cells having small size and granularity were sorted from the whole MNCs population, and an enriched population of with 93% of CD34+ cells was obtained (Supplementary Figure 1A). Colony forming assay (CFU) was performed on CD34+ cells to assess their clonogenic potential. Results showed that these cells were able to form all types of colonies including Granulocyte (CFU-G), Macrophage (CFU-M), Granulocyte-macrophage (CFU-GM), Erythroid (CFU-E), Granulocyte-Erythrocyte-Macrophage-Megakaryocyte (CFU-GEMM) in methylcellulose-based medium (Supplementary Figure 1B).
Co-Culture System of MSCs with HSCs or with AML Cell Lines
Normal healthy MSCs were purchased from Lonza (Switzerland) or isolated from healthy donors and AML patients. Two AML cell lines, THP-1 and Molm-13 were grown routinely in RPMI medium supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin. Three primary cells isolated from AML patients, having different genetic and cytogenetic backgrounds (Supplementary Table 2) were used in this study. MSCs were seeded (7000/cm2) in 6 well plates in DMEM low glucose medium (Sigma-Aldrich, USA). After reaching 80% confluency, cells were switched to IMDM medium 24 h prior to co-culture. Freshly isolated CD34+ cells or AML cell lines were directly co-cultured with MSCs (ratio 1:1). After 24h, two CD34+ cell populations and AML cell lines were identified: adherent and suspension fractions. HSCs or AML cell lines in suspension were collected, and then the adherent fraction was separated from MSCs by gentle PBS washes. The purity of this separation between MSCs-HSCs was verified by FACS analysis according to the size and expression of a specific mesenchymal stem cell marker CD73 [14], and showing that the HSCs adherent fraction is smaller than MSCs and more than 95% of these small cells do not express CD73 mesenchymal marker (supplementary Figure 1C).
Colony Forming Cell Assay
CD34+ cells (1 x 103) were cultured for 15 days in IMDM medium. Colonies of erythroid progenitors (CFU-E and BFU-E), granulocyte-macrophage progenitors (CFU-GM, CFU-G, and CFU-M), and multi-potential granulocyte-erythroid-macrophage-megakaryocyte progenitors (CFU-GEMM) were morphologically characterized, counted, and quantified by light microscopy (Zeiss, Germany).
Transcriptional Expression Analysis
Total RNA was extracted from cells using RNeasy Mini Kit (Qiagen, USA) as per manufacturer’s instructions. Briefly, 1 μg of total RNA was reverse transcribed to cDNA using RevertAid 1st Strand cDNA synthesis Kit (Thermo scientific, USA). Quantitative PCR (qPCR) was performed using the iQ SYBR GreenSupermix in a CFX96 system (Bio-Rad Laboratories, USA). Products were amplified using primers that recognize Cx-43, N-cadherin, MMP2, SDF-1, VEGF, Snail, Twist, Nanog, Oct-4, CXCR4 and GAPDH (Table 1). PCR parameters consist of a pre-cycle of 95 °C for 3 min followed by 40 cycles consisting of 95 °C for 10 sec, 52–62 °C for 30 sec, and 72 °C for 30 sec with a final extension at 72 °C for 5 min. The fluorescence quantitative cycle value (Cq) was obtained for each gene and normalized to their corresponding GAPDH in the same sample. All experiments were carried out in duplicates and independently performed three times.
Protein Expression Analysis
Western blot analysis was performed to examine the levels of Cx-43 and N-cadherin expression in MSCs and HSCs alone or in co-culture. 100 μg from protein lysate were separated in a 10% SDS-PAGE gel. Proteins were transferred onto PVDF (Bio-Rad Laboratories, USA) membrane. Then, membranes were blocked with 5% skimmed milk and 0.05% Tween 20 in PBS (TPBS). Primary mouse anti Cx-43 (1 µg/mL, Sigma-Aldrich, USA) and rabbit anti N-cadherin (1 µg/mL, life technologies, USA) antibodies were added for overnight at 4°C. Anti-GAPDH antibody (1 µg/mL, Sigma-Aldrich, USA) was used as a housekeeping control. Blots were then incubated with adequate secondary IgG antibody (anti mouse or anti rabbit). Bands were visualized using ChemiDoc MP Imaging System-Biorad (Bio-Rad Laboratories, USA). The intensity of bands, in the linear range of intensity, was quantified using ImageJ software (U. S. National Institutes of Health, USA).
Immunofluorescence Microscopy
MSCs and CD34+ HSCs or AML cell lines were co-cultured on coverslips. Adherent CD34+ HSCs or AML cells were carefully fixed with 4% paraformaldehyde. Cells were then washed with PBS, permeabilized using Triton X-100 (0.5%) and further blocked with 5% normal goat serum (NGS) in PBS for 1h in a humidified chamber. Incubation with primary antibodies Cx-43, N-cadherin, and alkaline phosphatase (2 µg/mL) diluted in 1% NGS was performed overnight at 4°C. Cells were next washed and incubated by IgG-conjugated secondary antibodies Texas Red (anti-rabbit) and Alexa 488 (anti-mouse) (1 µg/mL) for 1h at room temperature. Cells nuclei were stained using Hoechst 33342 (Molecular probes) for 10 min in the dark. Cells were washed with PBS, mounted on slides using Prolong anti-fade kit and observed under confocal microscopy (Laser scanning confocal microscope, LSM 710, Carl Zeiss, Germany). Data analyses were performed using Zeiss Zen software.
Duo-Link In Situ Proximity Ligation Assay
MSCs grown on glass coverslips were co-cultured with or without CD34+ HSCs for 24 h at 37°C. Cells were fixed with 4% paraformaldehyde in PBS at room temperature, permeabilized with 0.1% Triton X-100 for 15 min and then blocked in 1h blocking stock. After incubation at room temperature with primary antibodies (anti-Cx-43 and anti-N-cadherin) for 1h and secondary antibody conjugated with oligonucleotide for 2h, assays were performed according to the manufacturers’ instructions (Olink Bioscience, Sweden). Slides were imaged using confocal microscopy (Laser scanning confocal microscope, LSM 710, Carl Zeiss, Germany).
Functional Assay of Adhesion and Communication
The dye transfer assay was performed using the membrane-permeable dye Calcein AM (Molecular Probes, USA). Upon entry of the dye into the cell, intracellular esterases rapidly cleave the molecule to the fluorescent membrane-impermeable but gap junction-permeable acid form. Briefly, MSCs were labeled with 2 μM of calcein in complete medium for 1 h at 37°C. Then, labeled cells were incubated in serum-free medium for 30 min at 37°C to allow any non-de-esterified dye to leave the cells and were used immediately in dye transfer experiments. Next, CD34+ HSCs or AML cell lines were seeded on top of labeled monolayers of MSCs for 3 h at 37°C. Finally, suspension cells were removed, and adherent cells were washed and detached by trypsinization. Both cell populations were fixed in 4% formaldehyde and analyzed by flow cytometry (BD, USA).
Osteogenic Differentiation Engagement
MSCs were seeded in 6 well plates (70 x 103/well) or 12 well plates (35 x 103/well) until reaching 80% confluency. Cells were either co-cultured with CD34+ HSCs or AML cell lines for 24 h, 21 days or were left in medium throughout the experiment. Osteogenic engagement medium was added to the above conditions consisting of IMDM supplemented with 250 nM dexamethasone (Sigma-Aldrich, USA). MSCs were treated for 21 days, and the medium was changed every 3 days. After 21 days, cells were harvested for the assessment of alkaline phosphatase levels by immunofluorescence and alizarin red staining.
Alizarin Red Staining
MSCs were seeded into 12 well plates and cultured in the presence/absence of CD34+ HSCs/AML cell lines (THP-1 and MOLM-13) or dexamethasone for 24 h or 21 days. Cells were washed with PBS and fixed with 4% paraformaldehyde for 20 min. Cells were stained using the Alizarin Red S reagent (40 mM) for 40 min and then washed with H2O. Images were taken using a light microscopy.
Results
Reciprocal Direct Interaction between MSCs and CD34+ HSCs
Mesenchymal stem cells are precursors of the BM cellular components. Therefore, MSCs monolayer interacting with CD34+ HSCs provide a simple and suitable in vitro model system for the stem cell niche [14,33]. The reciprocal interaction of MSCs with CD34+ HSCs was assessed in a co-culture system which reflects direct cell-cell contact between the two cell types (Figure 1A). CD34+ HSCs were directly co-cultured with MSCs at 1:1 ratio [14,33]. After 24 h, an (~80%) population of CD34+ HSCs adhered to the monolayer of MSCs (adherent fraction), whilst some cells (~20%) stayed in suspension (suspension fraction). The expression of cellular adhesion and communication markers was assessed by qPCR in CD34+ HSCs (adherent vs. suspension fractions) as well as in MSCs (alone vs. MSC+CD34+ HSCs). After 24 h of direct co-culture, the expression of stemness markers such as Oct-4 and Nanog exhibited a decrease of 28.89 ± 2.95% (p=0.0006) and 27.71 ± 7.04% (p=0.0171), respectively, in MSCs, suggesting that the sub-population of CD34+ HSCs affect their self-renew properties and may induce MSCs differentiation. Furthermore, the expression of adhesion markers specifically N-cadherin harbor 17.21 ± 2.39% (p=0.0020) of significant increase in MSCs (Figure 1B). The up-regulation in the expression of Cx-43 (66.25 ± 13.70, p<0.0001) and N-cadherin (173 ± 36.52, p<0.0001) was further confirmed at the protein level (Figure 1C). It is noteworthy that these two proteins were among the most expressed proteins in MSCs (Supplementary Figure 2).
The communication and adhesion markers Cx-43 and N-cadherin were 88.64 ± 45.39%, p=0.1226 and 88.46 ± 30.52%, p= 0.0442 up-regulated in the adherent fraction of HSCs, respectively (Figure 1D). Interestingly, the mRNA expression of CXCR4 implicated in homing to the bone marrow and in HSC quiescence was 95.5 ± 18.7% (p= 0.0070) upregulated in HSC adherent fraction as well as the angiogenic gene VEGF (44.43 ± 9.21%, p= 0.0085 of increase). CFU assay was performed to test the ability of CD34+ HSCs to undergo terminal differentiation in cytokine-supplemented methylcellulose medium. The number of colonies formed by the adherent fraction of CD34+ HSCs was increased when compared to CD34+ HSCs cultured alone (Figure 1E). Interestingly, the percentage of CD45+ cells (a differentiation marker in mature lymphohematopoietic cells) was more pronounced (~4 folds) in HSCs’ suspension fraction (66.6%) when compared to the adherent fraction (13.6%) (Figure 1F), thus suggesting that the communication between MSCs and CD34+ HSCs conserves the stemness of CD34+ HSCs.
Connexin 43 and N-cadherin interaction is the major player in cell-cell communication through gap junctional complex.
After 24 h of direct co-culture, Cx-43 and N-cadherin were up-regulated in both MSCs and CD34+ HSCs, suggesting that cell-cell communication might be mediated through gap junctional complexes. To investigate the interaction between Cx-43 and N-cadherin, The expression and localization of these two proteins in CD34+ HSCs and MSCs was examined using immunofluorescence microscopy (Figure 2A). Interestingly, Cx-43 and N-cadherin were detected and specifically localized at the cell membrane level, in the zone of contact between adjacent MSCs and CD34+ HSCs. The interaction between Cx-43 and N-cadherin was further asserted using the proximity ligation assay in MSCs cultured with CD34+ HSCs (Figure 2B). These findings are consistent with the observed co-localization of Cx-43 and N-cadherin proteins by immunofluorescence and the co-immunoprecipitation assay performed with both proteins (Supplementary Figure 2B).
Whether Cx-43 and N-cadherin assembly form a functional gap junctional complex was then investigated using the Calcein dye transfer assay. Briefly, MSCs were labeled with Calcein-AM dye and then co-cultured for 3h with unlabeled CD34+ HSCs. Dye transfer between both cell types was evaluated by flow cytometry and mean fluorescence intensity (MFI) was quantified. Interestingly, a shift in MFI was noticed only in the HSCs’ adherent fraction (Figure 2C), demonstrating that cell-cell communication between MSCs and HSCs is mediated through functional gap junctional complexes, which are formed as early as 3 h.
Based on the aforementioned results, the expression of stemness markers was decreased in MSCs while the clonogenic potential of CD34+ HSCs was increased, suggesting that CD34+ HSCs self-renew properties were affected and that MSCs differentiation may be hindered. To investigate whether the differentiation of MSCs is affected by direct communication with CD34+ HSCs, an engagement into osteoblastic differentiation is carried out on MSCs having direct contact with HSCs for 24 h or 21 days using dexamethasone. Alkaline phosphatase protein (ALP), an osteoblastic marker, was expressed only in MSCs that were in contact with CD34+ HSCs for a short period of time (24 h). A longer co-culture period for 21 days, resulted in failure of MSCs to be engaged into differentiation. These results were confirmed by Alizarin red staining (Figure 3). MSCs were more engaged into osteoblastic differentiation, only after removal of CD34+ HSCs. Hence, the sustained intercellular communication between HSCs and MSCs seems to conserve the stemness of MSCs.
Direct Cell-Cell Communication Is Decreased in Acute Myeloid Leukemia.
We previously showed that AML primary cells and cell lines have lower expression of Cx-43 [17]. We interrogated whether the communication of MSCs is modulated under pathological conditions using the same co-culture model of MSCs in direct contact with two AML cell lines: THP-1 and Molm-13 (Figure 4A). Our results demonstrated that the mRNA expression of Cx-43 and N-cadherin were significantly decreased in MSCs after 24 h of co-culture with THP-1 (58% for Cx-43, p=0.0028 and 55% for N-cadherin, p=0.0003) and Molm-13 (61% for Cx-43, p=0.0138 and 70% for N-cadherin, p<0.0001) (Figure 4B). These results were further confirmed at the protein levels, where the expression of Cx-43 decreased to 43.7 ± 18.55% (p=0.0781) in THP-1 and to 40.59 ± 16.64% (p= 0.0926) in Molm-13 as well as the expression of N-cadherin decreased to 64.79 ± 6.87% (p=0.0007) in THP-1 and to 84 ± 3.095% (p=0.0001) in Molm-13 cell lines. Furthermore, using the Calcein dye transfer assay, we demonstrated that the MFI shift was evident in the adherent fraction of THP-1 and Molm-13 after co-culture (Figure 4C). Consequently, despite a lower number of forming junctional complexes between AML cell lines and MSCs, the functionality is preserved. Finally, in order to assess the differentiation potential of MSCs following their direct contact with AML cell lines, we co-cultured THP-1 and Molm-13 for 24 h or 21 days, in the presence of dexamethasone, a compound known for inducing osteoblastic differentiation. Our results showed that MSCs lose their differentiation ability, only when the communication with AML cell lines persists for a longer period (21 days) (Figure 4D).
To further assess the regulation of cell-cell communication in leukemia patients, we performed a co-culture of MSCs with AML primary cells isolated from 3 different patients. Consistent with the obtained results with AML cell lines, Cx-43 and N-cadherin protein levels were diminished in MSCs to 35.31 ± 11.37%, p=0.0360 and 37.87 ± 13.71%, p=0.0507, respectively. These results suggest a reduced formation of gap junction, at least with Cx-43 as a junctional partner (Figure 4E).
Finally, to check whether BM-MSCs are causing a lower gap junction assembly, we interchanged the source of MSCs, either from two healthy individuals (one from adult and one from infant) or from two AML patients and performed the co-culture with CD34+ HSC (isolated from healthy individuals) and CD34+ AML (isolated from AML patients). Following co-culture, the expression levels of Cx-43 and N-cadherin proteins were more pronounced in healthy MSCs in comparison to AML-MSCs, regardless of whether the source of CD34+ was from normal or AML patients (Figure 3F and supplementary Figure 2C). These results suggest that the gap junction formation is reduced, probably due to a pathological defect, mainly in MSCs and not CD34+ cells.
Discussion
The cellular players of the hematopoietic niche direct self-renewal and differentiation of HSCs under physiological conditions. Using an in vitro co-culture model, we showed that the direct contact between CD34+ HSCs and MSCs is mediated by junctional complexes, including Cx-43 and N-cadherin proteins as main players. We also demonstrated that the stemness steady-state situation of MSCs is preserved through direct communication with CD34+ HSCs, which determines the fate of both cell types. Under pathological conditions such as AML, we validated that the gap junction formation is reduced due to a decrease in Cx-43 and N-cadherin expressions. We showed that MSCs isolated from AML patients could be the source of gap junction impairment.
The microenvironment regulates the fate of HSC through direct or indirect communication, enabling the generation of all the blood cells. Few studies addressed the role of direct communication in affecting the properties of MSCs and HSCs [7,14]. The interaction between both cell types regulates the hematopoietic process on two different levels. First, the adhesion receptors and their ligands, expressed on the surface of both cell types, mediate either MSCs-HSCs or HSCs-ECM adhesion. These receptors and ligands function as signaling molecules capable of altering the proliferative behavior of HSCs [14]. Second, the direct intercellular communication via gap junctions could allow the exchange of low molecular weight regulatory molecules between these cells [2,5,14,34]. We showed that under normal physiological conditions, MSCs directly communicate with CD34+ HSCs and thus modulate the fate of both counterparts. We also confirmed that the expression of Cx-43 and N-cadherin was up-regulated following the direct interaction of MSCs and CD34+ HSCs. The co-localization of Cx-43 and N-cadherin highly suggests that the cell-cell communication between MSCs and CD34+ HSCs is mediated through junctional complexes. In agreement with our study, it was previously demonstrated that functional Cx-43-mediated direct gap junctional intercellular communication between BM stromal cells and between stromal and hematopoietic cells is required for hematopoiesis in vitro and in vivo [35,36,37]. In fact, Cx-43 was shown to have a protective role on HSCs during hematopoietic regeneration where Cx-43 mediated the transfer of reactive oxygen species from HSCs to the hematopoietic microenvironment thus preventing their quiescence/senescence [38].
Although the precise cellular and molecular composition of endosteal HSC niches is not fully described, the close proximity of MSCs-HSCs forms a reticular network that decides the fate of both cell types. It has been shown that the expansion and maintenance of HSCs ex vivo requires the presence of MSCs feeder layer that can also preserve HSCs stemness [39]. Moreover, in the BM niche, a cooperative regulation among cytokine signals and adhesion molecules is required for the maintenance of HSCs. Our results demonstrated that following direct contact of MSCs- CD34+ HSCs, not only the clonogenic potential of HSCs was increased, but also the expression of stemness markers in MSCs was decreased. This finding points to the reciprocal benefits of both cells provided by direct cell-cell contact. In the same context, we proved that the cellular junctional complex delays MSCs osteoblastic differentiation. Despite the presence of dexamethasone as an exogenous signal, MSCs failed to be engaged into osteoblasts when they are directly and sustainably communicating with CD34+ HSCs. The functional relevance of the crosstalk between MSCs-HSCs in the differentiation process is that it allows the exchange of signaling molecules that hinder MSCs differentiation and impede the transition of HSCs from dormant to active state. Therefore, direct cell-cell communication might constitute an additional “stem warden” that will further preserve the stemness capacity of both cell types. However, in the presence of external signals, such as ROS, CXCL12, VEGF, GM-CSF and CXCL8, the junctional complexes will be dissociated to facilitate the mobilization of HSCs into the circulation and their differentiation into multipotent progenitor cells [9]. Consequently, free MSCs will be engaged into osteogenic/adipogenic differentiation cascade, mainly under the control of BMP2 and BMP6 (bone morphogenetic proteins) [40], β-catenin dependent/independent Wnt signaling, Hedgehog, NELL1 (NEL-like protein 1, protein kinase C-binding protein) and IGF signaling [41].
The genesis of a leukemic permissive niche was shown to be controlled by various alterations in cell adhesion and cytokine signaling of BM-derived stromal cells, specifically MSCs [42,43]. In this study, we showed that under pathological conditions, gap junction communication through Cx-43 and N-cadherin between MSCs and AML cell lines was still persistent but to a lower extent. These results are in agreement with our previous results confirming that leukemic cells isolated from AML patients lose their Cx-43 profile in comparison to normal BM-CD34+ cells and overexpress numerous connexins, especially Cx-25 [17]. These results suggest that Cx-25 (or any other connexins) could be more involved in gap junctional formation than Cx-43 under leukemic conditions. Additional investigations are needed to confirm the involvement of Cx-25 in our co-culture model. In line with our data, carbenoxolone (CBX), a gap junction disruptor, shows antileukemic effects against AML cell lines by inducing their apoptosis without affecting normal BM-CD34+ cells. Furthermore, CBX improves animal survival through preventing leukemic infiltration to the liver and the spleen of mice developing an aggressive AML [17].
Tumor microenvironment plays a major role in cancer progression by providing nutrients and survival signals to tumor cells [44]. This microenvironment protects cancer cells from normal immune responses and therapeutic treatment agents [45]. We demonstrated that a direct co-culture between normal MSCs and AML cell lines, such as chemo-resistant THP-1 and chemo-sensitive Molm-13, or AML primary cells isolated from patients, induce downregulation of Cx-43 and N-cadherin junctional complexes without affecting their interaction. Moreover, the differentiation ability of MSCs was abolished after a long nesting time with AML cell lines, further pointing to the role of communication in the regulation of leukemic cell proliferation and chemoresistance [46]. In the same context, previous studies showed that Cx-43-junctional activity increased multiple myeloma cell survival and chemoresistance when in contact with stromal cells [47].
In conclusion, we propose that under physiological conditions, MSCs-CD34+ HSCs direct communication preserves the stemness state of both cells via gap junctions, a mechanism that is probably altered in a leukemic microenvironment. The switch to use other connexins in the cell-cell communication might be another pathway undertaken for leukemogenesis.
Author Contribution
R.S. and T.M.K. performed experiments, analyzed data, and drafted the manuscript; F.K. performed and analyzed experiments; J.E.S., I.Z. provided resources to the study; HEH and K.Z. completed a critical revision of the article; V.D. performed experiments; P.M. provided resources to the study and completed a critical revision of the article; M.E.S. conceptualized, designed and supervised the work. All authors read and approved the manuscript.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board of the American University of Beirut.
Acknowledgments
The authors thank Dr. Layal El-Hajjar and Dr. Hussein Chhouri for their assistance in experimental techniques. This work is supported by the MPP and URB from the American University of Beirut and the Lebanese National Council for Scientific Research (CNRS).
Conflicts of Interest
Authors declare no competing interest.
References
- Yin, T.; Li, L. The Stem Cell Niches in Bone. J Clin Invest 2006, 116, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, H.; Schanz, U.; Manz, M.G. Ex Vivo Expansion of Hematopoietic Stem Cells: Mission Accomplished? Swiss Med Wkly 2011, 141, w13316. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Nilsson, S.K. Bone, Microenvironment and Hematopoiesis. Curr Opin Hematol 2012, 19, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Phinney, D.G.; Prockop, D.J. Concise Review: Mesenchymal Stem/Multipotent Stromal Cells: The State of Transdifferentiation and Modes of Tissue Repair--Current Views. Stem Cells 2007, 25, 2896–2902. [Google Scholar] [CrossRef]
- Pang, W.W.; Price, E.A.; Sahoo, D.; Beerman, I.; Maloney, W.J.; Rossi, D.J.; Schrier, S.L.; Weissman, I.L. Human Bone Marrow Hematopoietic Stem Cells Are Increased in Frequency and Myeloid-Biased with Age. Proc Natl Acad Sci U S A 2011, 108, 20012–20017. [Google Scholar] [CrossRef]
- Kopp, H.-G.; Avecilla, S.T.; Hooper, A.T.; Rafii, S. The Bone Marrow Vascular Niche: Home of HSC Differentiation and Mobilization. Physiology (Bethesda) 2005, 20, 349–356. [Google Scholar] [CrossRef]
- Dorshkind, K. Regulation of Hemopoiesis by Bone Marrow Stromal Cells and Their Products. Annu Rev Immunol 1990, 8, 111–137. [Google Scholar] [CrossRef]
- Schepers, K.; Campbell, T.B.; Passegué, E. Normal and Leukemic Stem Cell Niches: Insights and Therapeutic Opportunities. Cell Stem Cell 2015, 16, 254–267. [Google Scholar] [CrossRef]
- Mendelson, A.; Frenette, P.S. Hematopoietic Stem Cell Niche Maintenance during Homeostasis and Regeneration. Nat Med 2014, 20, 833–846. [Google Scholar] [CrossRef]
- Jeltsch, K.S.; Radke, T.F.; Laufs, S.; Giordano, F.A.; Allgayer, H.; Wenz, F.; Zeller, W.J.; Kögler, G.; Fruehauf, S.; Maier, P. Unrestricted Somatic Stem Cells: Interaction with CD34+ Cells in Vitro and in Vivo, Expression of Homing Genes and Exclusion of Tumorigenic Potential. Cytotherapy 2011, 13, 357–365. [Google Scholar] [CrossRef]
- Mishima, S.; Nagai, A.; Abdullah, S.; Matsuda, C.; Taketani, T.; Kumakura, S.; Shibata, H.; Ishikura, H.; Kim, S.U.; Masuda, J. Effective Ex Vivo Expansion of Hematopoietic Stem Cells Using Osteoblast-Differentiated Mesenchymal Stem Cells Is CXCL12 Dependent. Eur J Haematol 2010, 84, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Lévesque, J.-P.; Helwani, F.M.; Winkler, I.G. The Endosteal “osteoblastic” Niche and Its Role in Hematopoietic Stem Cell Homing and Mobilization. Leukemia 2010, 24, 1979–1992. [Google Scholar] [CrossRef] [PubMed]
- London, N.R.; Li, D.Y. Robo4-Dependent Slit Signaling Stabilizes the Vasculature during Pathologic Angiogenesis and Cytokine Storm. Curr Opin Hematol 2011, 18, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Jing, D.; Fonseca, A.-V.; Alakel, N.; Fierro, F.A.; Muller, K.; Bornhauser, M.; Ehninger, G.; Corbeil, D.; Ordemann, R. Hematopoietic Stem Cells in Co-Culture with Mesenchymal Stromal Cells--Modeling the Niche Compartments in Vitro. Haematologica 2010, 95, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Willecke, K.; Eiberger, J.; Degen, J.; Eckardt, D.; Romualdi, A.; Güldenagel, M.; Deutsch, U.; Söhl, G. Structural and Functional Diversity of Connexin Genes in the Mouse and Human Genome. Biol Chem 2002, 383, 725–737. [Google Scholar] [CrossRef]
- Dürig, J.; Rosenthal, C.; Halfmeyer, K.; Wiemann, M.; Novotny, J.; Bingmann, D.; Dührsen, U.; Schirrmacher, K. Intercellular Communication between Bone Marrow Stromal Cells and CD34+ Haematopoietic Progenitor Cells Is Mediated by Connexin 43-Type Gap Junctions. Br J Haematol 2000, 111, 416–425. [Google Scholar]
- Kouzi, F.; Zibara, K.; Bourgeais, J.; Picou, F.; Gallay, N.; Brossaud, J.; Dakik, H.; Roux, B.; Hamard, S.; Le Nail, L.-R.; Hleihel, R.; Foucault, A.; Ravalet, N.; Rouleux-Bonnin, F.; Gouilleux, F.; Mazurier, F.; Bene, M.C.; Akl, H.; Gyan, E.; Domenech, J.; El-Sabban, M.; Herault, O. Disruption of Gap Junctions Attenuates Acute Myeloid Leukemia Chemoresistance Induced by Bone Marrow Mesenchymal Stromal Cells. Oncogene 2020, 39, 1198–1212. [Google Scholar] [CrossRef]
- Maître, J.-L.; Heisenberg, C.-P. Three Functions of Cadherins in Cell Adhesion. Curr Biol 2013, 23, R626–33. [Google Scholar] [CrossRef]
- Alimperti, S.; Andreadis, S.T. CDH2 and CDH11 Act as Regulators of Stem Cell Fate Decisions. Stem Cell Res 2015, 14, 270–282. [Google Scholar] [CrossRef]
- Dbouk, H.A.; Mroue, R.M.; El-Sabban, M.E.; Talhouk, R.S. Connexins: A Myriad of Functions Extending beyond Assembly of Gap Junction Channels. Cell Commun Signal 2009, 7, 4. [Google Scholar] [CrossRef]
- Kiel, M.J.; Acar, M.; Radice, G.L.; Morrison, S.J. Hematopoietic Stem Cells Do Not Depend on N-Cadherin to Regulate Their Maintenance. Cell Stem Cell 2009, 4, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, K.; Arai, F.; Yoshihara, H.; Iwasaki, H.; Nakamura, Y.; Gomei, Y.; Suda, T. Knockdown of N-Cadherin Suppresses the Long-Term Engraftment of Hematopoietic Stem Cells. Blood 2010, 116, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Stölzel, F.; Mohr, B.; Kramer, M.; Oelschlägel, U.; Bochtler, T.; Berdel, W.E.; Kaufmann, M.; Baldus, C.D.; Schäfer-Eckart, K.; Stuhlmann, R.; Einsele, H.; Krause, S.W.; Serve, H.; Hänel, M.; Herbst, R.; Neubauer, A.; Sohlbach, K.; Mayer, J.; Middeke, J.M.; Platzbecker, U.; Schaich, M.; Krämer, A.; Röllig, C.; Schetelig, J.; Bornhäuser, M.; Ehninger, G. Karyotype Complexity and Prognosis in Acute Myeloid Leukemia. Blood Cancer J 2016, 6, e386. [Google Scholar] [CrossRef]
- Döhner, H.; Weisdorf, D.J.; Bloomfield, C.D. Acute Myeloid Leukemia. N Engl J Med 2015, 373, 1136–1152. [Google Scholar] [CrossRef]
- Bendall, L.J.; Daniel, A.; Kortlepel, K.; Gottlieb, D.J. Bone Marrow Adherent Layers Inhibit Apoptosis of Acute Myeloid Leukemia Cells. Exp Hematol 1994, 22, 1252–1260. [Google Scholar]
- Konopleva, M.; Konoplev, S.; Hu, W.; Zaritskey, A.Y.; Afanasiev, B.V.; Andreeff, M. Stromal Cells Prevent Apoptosis of AML Cells by Up-Regulation of Anti-Apoptotic Proteins. Leukemia 2002, 16, 1713–1724. [Google Scholar] [CrossRef]
- Wang, A.; Zhong, H. Roles of the Bone Marrow Niche in Hematopoiesis, Leukemogenesis, and Chemotherapy Resistance in Acute Myeloid Leukemia. Hematology 2018, 23, 729–739. [Google Scholar] [CrossRef]
- Weber, M.C.; Tykocinski, M.L. Bone Marrow Stromal Cell Blockade of Human Leukemic Cell Differentiation. Blood 1994, 83, 2221–2229. [Google Scholar] [CrossRef]
- Morrison, S.J.; Scadden, D.T. The Bone Marrow Niche for Haematopoietic Stem Cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef]
- Nilsson, S.K.; Johnston, H.M.; Whitty, G.A.; Williams, B.; Webb, R.J.; Denhardt, D.T.; Bertoncello, I.; Bendall, L.J.; Simmons, P.J.; Haylock, D.N. Osteopontin, a Key Component of the Hematopoietic Stem Cell Niche and Regulator of Primitive Hematopoietic Progenitor Cells. Blood 2005, 106, 1232–1239. [Google Scholar] [CrossRef]
- Shariatmadar, S.; Sharma, S.; Cabana, R.; Powell, S.; Ruiz, P.; Krishan, A. Electronic Volume of CD34 Positive Cells from Peripheral Blood Apheresis Samples. Cytometry B Clin Cytom 2008, 74, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Vescio, R.A.; Hong, C.H.; Cao, J.; Kim, A.; Schiller, G.J.; Lichtenstein, A.K.; Berenson, R.J.; Berenson, J.R. The Hematopoietic Stem Cell Antigen, CD34, Is Not Expressed on the Malignant Cells in Multiple Myeloma. Blood 1994, 84, 3283–3290. [Google Scholar] [CrossRef] [PubMed]
- Singh, P. MSC and HSPC Coculture: Mimicking Ex Vivo Bone Marrow Niche. Methods Mol Biol 2023, 2567, 181–189. [Google Scholar] [PubMed]
- Li, T.; Wu, Y. Paracrine Molecules of Mesenchymal Stem Cells for Hematopoietic Stem Cell Niche. Bone Marrow Res 2011, 2011, 353878. [Google Scholar] [CrossRef]
- Hurtado, S.P.; Balduino, A.; Bôdi, E.C.A.; El-Cheikh, M.C.; Campos de Carvalho, A.C.; Borojevic, R. Connexin Expression and Gap-Junction-Mediated Cell Interactions in an in Vitro Model of Haemopoietic Stroma. Cell Tissue Res 2004, 316, 65–76. [Google Scholar] [CrossRef]
- Cancelas, J.A.; Koevoet, W.L.; de Koning, A.E.; Mayen, A.E.; Rombouts, E.J.; Ploemacher, R.E. Connexin-43 Gap Junctions Are Involved in Multiconnexin-Expressing Stromal Support of Hemopoietic Progenitors and Stem Cells. Blood 2000, 96, 498–505. [Google Scholar] [CrossRef]
- Montecino-Rodriguez, E.; Dorshkind, K. Regulation of Hematopoiesis by Gap Junction-Mediated Intercellular Communication. J Leukoc Biol 2001, 70, 341–347. [Google Scholar] [CrossRef]
- Taniguchi Ishikawa, E.; Gonzalez-Nieto, D.; Ghiaur, G.; Dunn, S.K.; Ficker, A.M.; Murali, B.; Madhu, M.; Gutstein, D.E.; Fishman, G.I.; Barrio, L.C.; Cancelas, J.A. Connexin-43 Prevents Hematopoietic Stem Cell Senescence through Transfer of Reactive Oxygen Species to Bone Marrow Stromal Cells. Proc Natl Acad Sci U S A 2012, 109, 9071–9076. [Google Scholar] [CrossRef]
- Perucca, S.; Di Palma, A.; Piccaluga, P.P.; Gemelli, C.; Zoratti, E.; Bassi, G.; Giacopuzzi, E.; Lojacono, A.; Borsani, G.; Tagliafico, E.; Scupoli, M.T.; Bernardi, S.; Zanaglio, C.; Cattina, F.; Cancelli, V.; Malagola, M.; Krampera, M.; Marini, M.; Almici, C.; Ferrari, S.; Russo, D. Mesenchymal Stromal Cells (MSCs) Induce Ex Vivo Proliferation and Erythroid Commitment of Cord Blood Haematopoietic Stem Cells (CB-CD34+ Cells). PLoS One 2017, 12, e0172430. [Google Scholar] [CrossRef]
- Beederman, M.; Lamplot, J.D.; Nan, G.; Wang, J.; Liu, X.; Yin, L.; Li, R.; Shui, W.; Zhang, H.; Kim, S.H.; Zhang, W.; Zhang, J.; Kong, Y.; Denduluri, S.; Rogers, M.R.; Pratt, A.; Haydon, R.C.; Luu, H.H.; Angeles, J.; Shi, L.L.; He, T.-C. BMP Signaling in Mesenchymal Stem Cell Differentiation and Bone Formation. J Biomed Sci Eng 2013, 6, 32–52. [Google Scholar] [CrossRef]
- James, A.W. Review of Signaling Pathways Governing MSC Osteogenic and Adipogenic Differentiation. Scientifica (Cairo) 2013, 2013, 684736. [Google Scholar] [CrossRef] [PubMed]
- Corradi, G.; Baldazzi, C.; Očadlíková, D.; Marconi, G.; Parisi, S.; Testoni, N.; Finelli, C.; Cavo, M.; Curti, A.; Ciciarello, M. Mesenchymal Stromal Cells from Myelodysplastic and Acute Myeloid Leukemia Patients Display in Vitro Reduced Proliferative Potential and Similar Capacity to Support Leukemia Cell Survival. Stem Cell Res Ther 2018, 9, 271. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Ross, C.; Srivastava, S. Ally to Adversary: Mesenchymal Stem Cells and Their Transformation in Leukaemia. Cancer Cell Int 2019, 19, 139. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Fan, X.; Houghton, J. Tumor Microenvironment: The Role of the Tumor Stroma in Cancer. J Cell Biochem 2007, 101, 805–815. [Google Scholar] [CrossRef]
- Trédan, O.; Galmarini, C.M.; Patel, K.; Tannock, I.F. Drug Resistance and the Solid Tumor Microenvironment. J Natl Cancer Inst 2007, 99, 1441–1454. [Google Scholar] [CrossRef]
- Paraguassú-Braga, F.H.; Borojevic, R.; Bouzas, L.F.; Barcinski, M.A.; Bonomo, A. Bone Marrow Stroma Inhibits Proliferation and Apoptosis in Leukemic Cells through Gap Junction-Mediated Cell Communication. Cell Death Differ 2003, 10, 1101–1108. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Zhang, L.; Zhou, D.; Sun, Y.; Wang, P.; Ju, S.; Chen, P.; Li, J.; Fu, J. Impact of Connexin 43 Coupling on Survival and Migration of Multiple Myeloma Cells. Arch Med Sci 2017, 13, 1335–1346. [Google Scholar] [CrossRef]
Figure 1.
Direct co-culture of MSCs and CD34+ HSCs. A. Schematic representation of the direct co-culture system between MSCs and CD34+ HSCs. Representative images of HSCs alone, MSCs alone and MSCs+CD34+ HSCs after direct co-culture. Scale bar, 100µm. B. Histogram representing mRNA expression levels of adhesion and communication markers in MSCs following direct co-culture, assessed by qPCR. C. Western blot of Cx-43 and N-cadherin expression in MSCs after 24 h of direct co-culture with CD34+ HSCs. Results are represented as normalized expression to GAPDH in three independent experiments ± SEM. D. Expression of adhesion/communication markers and VEGF, CXCR4 in CD34+ HSCs following direct co-culture, assessed by qPCR. Results are represented as normalized expression of GAPDH in three independent experiments. E. Clonogenic potential of CD34+ HSCs, expressed as the number of colonies after direct co-culture with MSCs. F. Cell profiling of CD34+ HSCs after co-culture, showing the percentage of CD38+ and CD45+ cells in both suspension and adherent fractions.
Figure 1.
Direct co-culture of MSCs and CD34+ HSCs. A. Schematic representation of the direct co-culture system between MSCs and CD34+ HSCs. Representative images of HSCs alone, MSCs alone and MSCs+CD34+ HSCs after direct co-culture. Scale bar, 100µm. B. Histogram representing mRNA expression levels of adhesion and communication markers in MSCs following direct co-culture, assessed by qPCR. C. Western blot of Cx-43 and N-cadherin expression in MSCs after 24 h of direct co-culture with CD34+ HSCs. Results are represented as normalized expression to GAPDH in three independent experiments ± SEM. D. Expression of adhesion/communication markers and VEGF, CXCR4 in CD34+ HSCs following direct co-culture, assessed by qPCR. Results are represented as normalized expression of GAPDH in three independent experiments. E. Clonogenic potential of CD34+ HSCs, expressed as the number of colonies after direct co-culture with MSCs. F. Cell profiling of CD34+ HSCs after co-culture, showing the percentage of CD38+ and CD45+ cells in both suspension and adherent fractions.
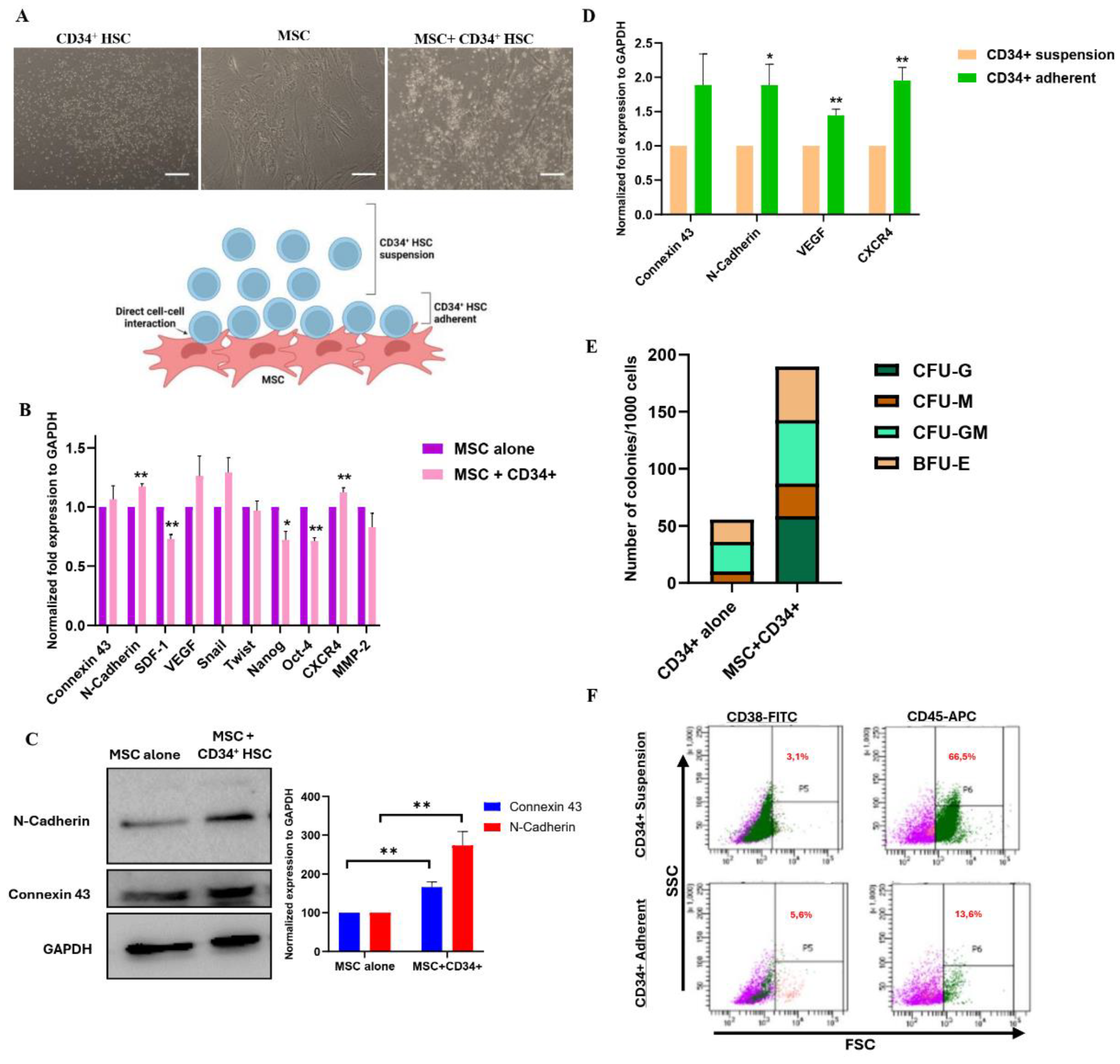
Figure 2.
Immunofluorescence of Cx-43 and N-cadherin in MSCs and CD34+ HSCs. A. Single staining of Cx-43 (red, upper panel) and N-cadherin (green, lower panel) following direct co-culture. Scale bar, 5 µm (upper panel) and 10 µm (lower panel). B. Cx-43 and N-cadherin interaction in MSCs and HSCs after direct co-culture as detected by Duo-Link assay. The dots (red) are representatives of the close proximity of the two proteins of interest. Nuclei were stained with Hoechst 33342 dye (blue). Scale bar, 10 µm (upper panel) and 2 µm (lower panel). C. Representative flow cytometry graph showing the shift in MFI following co-culture of unlabeled HSCs with Calcein-labeled MSCs.
Figure 2.
Immunofluorescence of Cx-43 and N-cadherin in MSCs and CD34+ HSCs. A. Single staining of Cx-43 (red, upper panel) and N-cadherin (green, lower panel) following direct co-culture. Scale bar, 5 µm (upper panel) and 10 µm (lower panel). B. Cx-43 and N-cadherin interaction in MSCs and HSCs after direct co-culture as detected by Duo-Link assay. The dots (red) are representatives of the close proximity of the two proteins of interest. Nuclei were stained with Hoechst 33342 dye (blue). Scale bar, 10 µm (upper panel) and 2 µm (lower panel). C. Representative flow cytometry graph showing the shift in MFI following co-culture of unlabeled HSCs with Calcein-labeled MSCs.
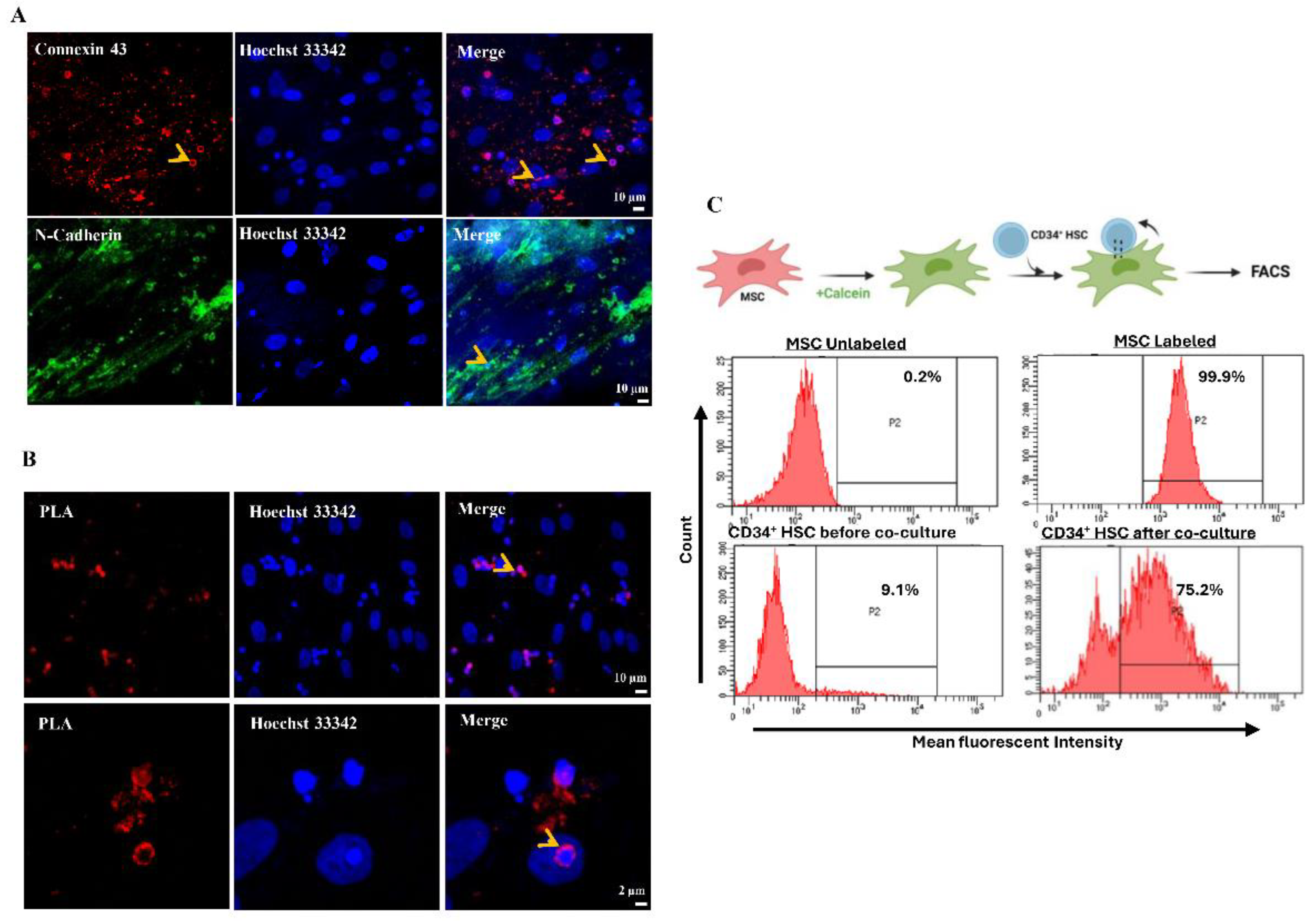
Figure 3.
Osteoblastic differentiation of MSCs after co-culture with HSCs for 24 h or 21 days. Left panel: Expression of ALP protein in MSCs detected by immunofluorescence. Right panel: Alizarin red staining of MSCs visualized by light microscopy. Scale bar, 20 µm.
Figure 3.
Osteoblastic differentiation of MSCs after co-culture with HSCs for 24 h or 21 days. Left panel: Expression of ALP protein in MSCs detected by immunofluorescence. Right panel: Alizarin red staining of MSCs visualized by light microscopy. Scale bar, 20 µm.
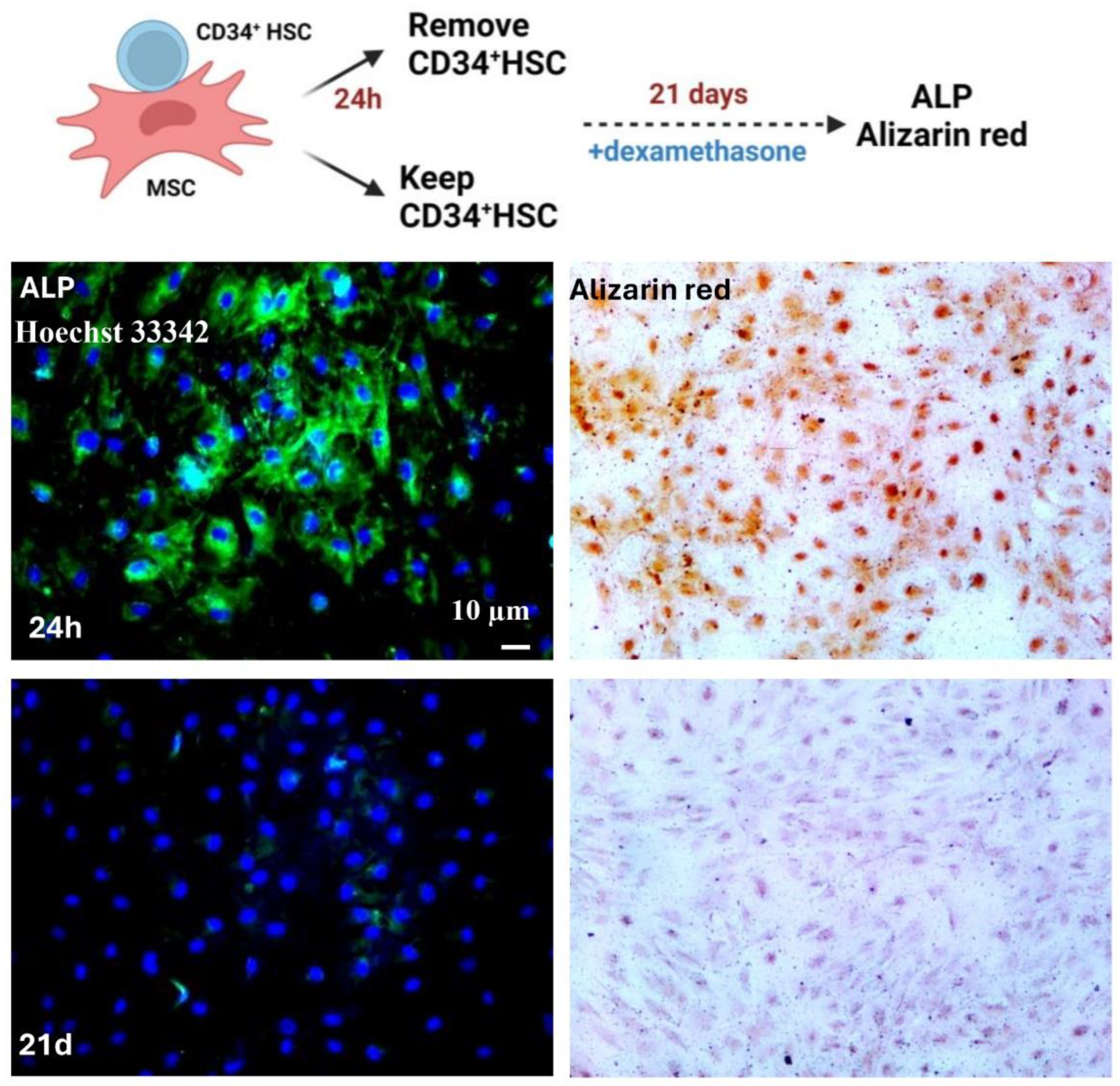
Figure 4.
Direct co-culture of MSCs and AML cells. A. Schematic representation of the direct co-culture system between MSCs and THP-1 and Molm-13. Scale bar, 100µm. B. mRNA and protein expression levels of Cx-43 and N-cadherin in MSCs following direct co-culture with AML cell lines, assessed by qPCR and western blot, respectively. C. Representative flow cytometry graphs showing the shift in MFI following co-culture of unlabeled AML cell lines (THP-1 and Molm-13) with Calcein-labeled MSCs. D. Fluorescent and light microscopy images of MSCs co-cultured with AML cell lines for ALP expression and Alizarin Red staining, respectively. Scale bar, 20 µm. E. Expression of Cx-43 and N-Cadherin in MSCs following direct co-culture with AML primary cells (3 patients), assessed by western blot analysis. F. mRNA expression levels of Cx-43 and N-cadherin in MSCs of healthy and AML patients following direct co-culture with AML cell lines, CD34+ N from healthy individuals and CD34+ AML from patients.
Figure 4.
Direct co-culture of MSCs and AML cells. A. Schematic representation of the direct co-culture system between MSCs and THP-1 and Molm-13. Scale bar, 100µm. B. mRNA and protein expression levels of Cx-43 and N-cadherin in MSCs following direct co-culture with AML cell lines, assessed by qPCR and western blot, respectively. C. Representative flow cytometry graphs showing the shift in MFI following co-culture of unlabeled AML cell lines (THP-1 and Molm-13) with Calcein-labeled MSCs. D. Fluorescent and light microscopy images of MSCs co-cultured with AML cell lines for ALP expression and Alizarin Red staining, respectively. Scale bar, 20 µm. E. Expression of Cx-43 and N-Cadherin in MSCs following direct co-culture with AML primary cells (3 patients), assessed by western blot analysis. F. mRNA expression levels of Cx-43 and N-cadherin in MSCs of healthy and AML patients following direct co-culture with AML cell lines, CD34+ N from healthy individuals and CD34+ AML from patients.
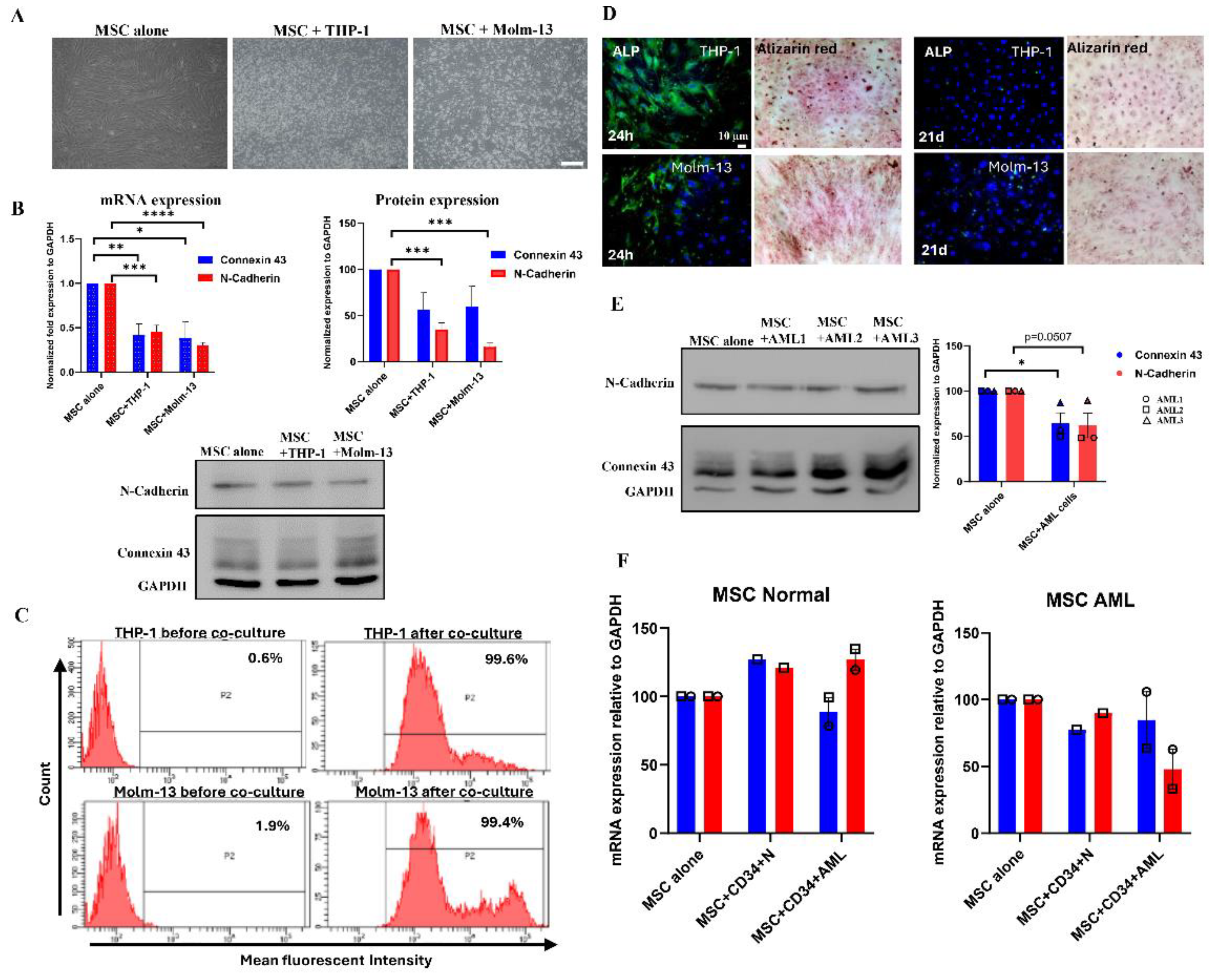

Table 1.
List of human primers used for qPCR.
| Genes | Sequence |
|---|---|
| Connexin 43 | F: GGA AGA TGG GCT CAT GAA AA R: GCA AAG GCC TGT AAC ACC AT |
| N-cadherin | F: GGT GGA GGA GAA GAA GAC CAG R: GGC ATC AGG CTC CAC AGT |
| MMP2 | F: TTG ACG GTA AGG ACG GAC TC R: ACT TGC AGT ACT CCC CAT CG |
| SDF-1 | F: GCC CGT CAG CCT GAG CTA CA R: TTC TTC AGC CGG CGT ACA ATC T |
| VEGF | F: AGG CCC ACA GGG ATT TTC TT R: ATC AAA CCT CAC CAA GGC CA |
| Snail | F: CTT CCA GCA GCC CTA CGA C R: CGG TGG GGT TGA GGA TCT |
| Twist | F: AGC TAC GCC TTC TCG GTC T R: CCT TCT CTG GAA ACA ATG ACA TC |
| Nanog | F: CAG AAG GCC TCA GCA CCT AC R: ATT GTT CCA GGT CTG GTT GC |
| Oct-4 | F: CAG TGC CCG AAA CCC ACA C R: GGA GGAC CCA GCA GCC TCA AA |
| CXCR4 | F: CCT CCT GCT GAC TAT TCC CGA R: GGA ACA CAA CCA CCC ACA AGT |
| GAPDH | F: TGG TGC TCA GTG TAG CCC AG R: GGA CCT TGA CCT GCC GTC TAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



