
Submitted:
30 September 2024
Posted:
01 October 2024
You are already at the latest version
Abstract
The diploid and autotetraploid of the eight tartary buckwheat cultivars were used to make a karyotype study by the conventional squash method to establish a reference for polyploid breeding. In this study, the eight cultivars exhibited similar karyotypic features between diploids and autotetraploids. The chromosome number in diploids was found to be 2n=2x=16, while 2n=4x=32 in autotetraploids. T6, T9, T18, T22, T27 and T36 possessed one pair of SAT chromosomes, whereas T25 and T39 had two pairs of SAT chromosomes. Compared with diploids, the total number of chromosomes and SAT chromosomes doubled in autotetraploids. However, the karyotype of autotetraploids remained unchanged in all cases which belonged to the 2A type. The karyotype of the same crop with different ploidy in diploid and autotetraploid buckwheat is the same. These eight cultivars' karyotype and chromosome number of autotetraploids are reported for the first time.
Keywords:
tartary buckwheat
; diploid
; autotraploid
; karyotype analysis
1. Introduction
Buckwheat belongs to Fagopyrum of Polygonaceous. Sweet buckwheat (F.esculentum Moench) and tartary buckwheat [F.tataricum (L.) Gaertn] are the two main cultivars for grain[1].
With the continuous improvement of material and cultural living standards, people pay more and more attention to healthy food and diet therapy. Tartary buckwheat is a crop that integrates nutrition, health care, and medical treatment. People have partiality for buckwheat food products. This demand has been increasing.
While Stevens reported the numbers of buckwheat chromosomes at first as early as 1912[2], there are a few reports of buckwheat chromosomes at home and abroad. The buckwheat chromosomes are small and belong to small chromosomes, and the cytological observation is difficult, so most research work is still at the level of chromosome numbers [3].
To date, the karyotype analysis of buckwheat has been limited to a few buckwheat species[3-15], namely: F.tartaricum, F.esculentum, F.megaspartanium, F.pilus, F.zuogongense, F.crispatifolium, F.gracilipes, F.densovillosum, F.cymosum, F.qiangcai, F.wenchuanense, F.urophyllum, F. leptopodum var. grossii.
Previous studies have primarily focused on karyotype analysis of diploid buckwheat and wild buckwheat, but the autotetraploid tartary buckwheat has not been reported. This study aims to investigate the number and karyotype of mitotic chromosomes in root tips of eight species of diploid and autotetraploid tartary buckwheat to provide valuable insights into genetic breeding strategies in buckwheat.
2. Materials and Methods
2.1. Experimental Materials
The diploid tartary buckwheat used in this experiment was provided by the Buckwheat Industrial Technology Research Center of Guizhou Normal University. The autotetraploids were obtained in the Lab Center by the author using the method of chromosome doubling of buckwheat. The materials used to study the diploids and autotetraploids have been summarized in Table 1.
2.2. Experimental Design
Seeds of diploid and autotetraploid tartary buckwheat were germinated on moist filter papers in petri dishes at room temperature in the dark. Once the root length of the seeds reached approximately 1.5cm, the root tips were collected at a length of 2mm, pretreated in saturated α-bromonaphthalene for 5h at 25℃, and taken out to fix in Carnoy’s Fixative SolutionⅠ for 12h for synchronization purposes. These were extracted, washed 2X with distilled water, and fixed in 0.075M KCl for 1h. After washing KCl in distilled water for 10min, the samples were incubated in an enzyme solution of 4% enzyme mixture (pectinase and cellulase) at 37℃ for 5h. The macerated tissues were transferred onto alcohol-cleaned glass slides, absorbed the residual enzyme solution, rinsed 3X with distilled water, and immersed in distilled water for 3h. Following enzymatic hydrolysis, the specimen was centred on the prepared slide, and the remaining distilled water was removed by washing with Carnoy’s Fixative SolutionⅠ. It was meticulously crushed with the tip of forceps to eliminate any impurities, followed by adding a fixative around the crushed area for concentration purposes. It was gently heated over an alcohol lamp flame until almost dry to extinguish the flame. Once completely dried, it was carefully placed on a white porcelain plate using toothpicks and slowly immersed in a 3% Giemsa dyeing solution with pH 6.8 for 1h. Thorough rinsing with tap water was performed until no traces of blue liquid were observed beneath the film, followed by rinsing 3X with distilled water and air-drying. The prepared sample was examined under an Olympus BX51-DP70 scientific research microscope.
2.3. Chromosome Index Determination
According to the unified standard[16], the statistics of the number of chromosomes in more than 30 cells and more than 5 cells with clearly visible chromosomes in the middle division were selected for karyotype analysis.
The mitotic phase of dispersed plant chromosomes was placed under a microscope to obtain clear images. The formula for calculating karyotype parameters is as follows[3]:
Each chromosome was named according to the Four-Point Four-Region System Naming Rule[17]. The buckwheat chromosome karyotypes were classified according to Stebbins's (1971) classification criteria.
2.4. Statistical Analysis
The analysis and measurement of the images were made using Image J software. The morphology, karyograms and idiograms were carried using Excel 2016, Word 2016 and Photoshop CS6.
3. Results
3.1. Karyotype Analysis of Diploid Chromosomes in Root Tip during the Mitotic Stage
According to Table 2, Table 3 and Figure 1, it can be concluded that the number of the 8 diploid chromosomes were 2n = 2x = 16, and the karyotype formula were 2n = 2x = 16 = 12m + 4sm. The karyotype of this species consists exclusively of metacentric (m) and submetacentric (sm) chromosomes. In addition, there were differences in the number and location of SAT chromosomes in different diploids. The T6, T9, T18, T22, T27 and T36 had one pair of SAT chromosomes while two pairs were found in T25 and T39. In T6, T25, T27, and T39, SAT was located on the sm chromosome, while the other 4 varieties were located on the m chromosome.
The relative length of the 8 diploid chromosomes (T6, T9, T18, T22, T25, T27, T36 and T39) were from 10.63% to 14.29%, from 9.17% to 15.79%, from 9.54% to 15.14%, from 9.65% to 15.73%, from 5.90% to 17.06%, from 6.55% to 15.38%, from 9.49% to 14.59%, from 8.83% to 15.90%, respectively. There were obvious differences in the relative length range.
According to Stebbins’(1971) classification standard, the karyotype analysis of 8 diploid chromosomes showed that they all belonged to the 2A type.
3.2. Karyotype Analysis of Autotetraploid in Root Tip During the Mitotic Stage
According to Table 2 and Table 3 and Figure 1, it can be inferred that the eight autotetraploid chromosomes were 2n = 4x = 32, and their karyotype formula were 2n = 4x = 32 = 24m + 8sm. The types of metacentric (m) and submetacentric(sm) remained unchanged. Furthermore, variations were observed in the quantity and location of SAT among different autotetraploids, resembling those found in diploids. Specifically, the autotetraploids(T6*, T9*, T18*, T22*, T27*, and T36*) possessed two pairs of SAT chromosomes each, while four pairs were present in both T25* and T39*.
The relative length ranges of the 8 autotetraploid chromosomes (T6*, T9*, T18*, T22*, T25*, T27*, T36* and T39*) were from 9.60% to 15.72%, from 9.57% to 14.83%, from 7.93% to 16.68%, from 9.44% to 15.25%, from 6.04% to 19.02%, from 8.05% to 15.25%, from 7.94% to 20.30%, from 9.01% to 14.76%, which showed that there were also significant differences in the relative length ranges.
According to Stebbins’ (1971) classification standard, the karyotype analysis of the eight autotetraploid chromosomes showed that they all belonged to 2A type.
The difference between diploid and autotetraploid in Karyotype analysis
As can be seen from Table 2, after diploid doubling, the number of chromosomes and SATs of eight cultivars doubled, but no changes in the types of chromosomes and karyotype were observed.
Therefore, it can be inferred that these eight autotetraploids were homologous.
4. Discussion
4.1. Karyotype of Tartary Buckwheat
In this study, the number of the eight diploid and autotetraploid chromosomes were 2n = 2x =16 and 2n = 4x = 32, respectively. In addition, this study found that one pair of SAT chromosomes co-existed with two pairs in root tip cells and each of the six diploids (T6, T9, T18, T22, T27, T36) possessed one pair of SAT chromosomes, which was consistent with the results reported by Zhang et al. [13] and Wang et al.[11]. However, two diploids (T25, T39) consistently exhibited the presence of two pairs of SAT chromosomes, aligning with previous findings reported by Zhu et al. [15], He et al.[5], Lei et al. [6], Chen [3], Yang et al. [12], and Sheng [8]. Whereas six autotetraploids (T6*, T9*, T18*, T22*, T27*, T36*) had two pairs of SAT chromosomes, but the T25* and T39* had four pairs of SAT chromosomes. The autotetraploid SAT chromosomes and their number are reported for the first time.
In terms of the karyotype formulas, the T9, T18, T22 and T36 were consistent with those reported by Zhang et al.[13]. The T25 and T39 were consistent with the karyotypes reported by Chen [3]and Sheng [8]. The T6 and T27 were consistent with those reported by Wang et al.[11]. This study has not seen the results reported by Lei et al.[6], namely 2n = 2x = 16 = 10m + 6sm(4SAT) and Yang et al.[12], namely, 2n = 2x = 16 = 14m(4SAT) + 2sm. The karyotype formulas of the eight autotetraploids (T6*, T9*, T18*, T22*, T25*, T27*, T36*) are reported for the first time.
The number of the eight diploid and autotetraploid chromosomes were consistent with previous research result. However, there were differences in the number and location of SAT and karyotype formulas. It indicated genetic diversity at the chromosome level among different germplasm of tartary buckwheat. There were significant differences in the climate and soil environments of different regions, and regional plants have been adapting to environmental conditions for a long time. It may also be due to the accumulation of subtle changes such as gene deletion, inversion, translocation, and insertion during the recombination process of chromosomes, resulting in changes in the development of chromosome morphology and structure.
4.2. Karyotype Changes after Chromosome Doubling
The karyotype characteristics of the same crop with different ploidy are similar, for example, papaya [18], broccoli [19], Solanaceae [20], non-heading Chinese cabbage [21], small fruit watermelons[22], goji berries [23].
This study has further confirmed that the homologous tetraploid obtained by doubling the diploid of tartary buckwheat exhibited symmetrical karyotype characteristics with the corresponding diploid, and there are no changes in chromosomes and karyotype types. It is consistent with previous research results. Namely, the karyotype characteristics of the same crop with different ploidy are consistent.
Author Contributions
Formal analysis, L.L., X.W. and F.C.; funding acquisition, F.C. and Y. Z.; software, L.L., P.Z. and X.W.; visualization, L.L. X.W. and F. C.; writing—original draft, L.L. and X.W.; writing—review and editing, L.L. and F.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received funding from the Special Fund Project for scientists in buckwheat breeding position of National Modern Agricultural Industrial Technology System (No. CARS-07-A5); Yunnan Science and Technology Major Project and Key Research and Development Program (No.202202AE090020); the Science and Technology Project of Bijie City (Bike Joint-[2023] No. 21), the Science and Technology Planning Project of Guizhou Province (QianKeHeZhiCheng-[2024]YiBan112).
Data Availability Statement
The data is contained within the article.
Acknowledgments
We appreciate and thank the anonymous reviewers for providing helpful comments that led to an overall improvement in the manuscript. We also thank the Journal Editor Board for their help and patience throughout the review process.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lin, R.F. The Development and Utilization of Tartary Buckwheat Resources. In Proceedings of the Proceedings of the 9 International Symposium on Buckwheat, 2004, 252-258.
- Morris, M.R. Cytogenetic studies on buckwheat. J. Hered. 1951, 42, 85-89. [CrossRef]
- Chen, Q.F. Karyotype analysis of five Fagopyrum species native to China. Guihaia 2001, 21, 107-110. [CrossRef]
- Du, X.C.; M. Y.; Liu,P.; Xu,G. D.Karyotype Analysis of Two Buckwheat Variety Chromosomes. Subtrop. Plant Sci. 2005, 34, 36-38.
- He, F.F. Karyotype analysis of buckwheat. J. Southwest Agric. Univ. 1992, 14, 522-523.
- Lei, B.J.; R. Z. Studies on the karyotype of chromosomes of buckwheats. J. Sichuan Univ. 2000, 37, 142-143.
- Liu, J.L.; Tang, Y.; Shao,J. R.; Luo,Q.; Sun,J. X. Karyotypic Studies of Two Wild Buckwheat Species in the Fagopyrum Mill. J. Northwest Botany 2009, 29, 1798-1803.
- Sheng, M.Y. Physical mapping of the 45S and 5SrDNA of cultivated buckwheat. J. Plant Genet. Resour. 2013, 14, 317-321. [CrossRef]
- Shi, J.Q.; Li,Y. Q.; Zhang,Z. W.; Wu,B.; Wang,A. H. Analysis of karyotypes and evolutionary features of wild buckwheat species of buckwheat. J. Plant Genet. Resour. 2016, 17, 455-460.
- Suresh, N.; Tamaki, H.; Junichi,W.; Kazumi,T.; Hiroko,K.; Shigeru,S. Karyotype analysis of buckwheat using atomic force microscopy. Microsc. Microanal. 2011, 17, 572-577. [CrossRef]
- Wang, J.S.; Chai, Y.;Zhao,X.T.; JI,W. Q. Karyotype Analysis of Chinese Buckwheat Cultivars. Acta Bot. Boreali-Occident. Sin. 2005, 25, 1114-1117.
- Yang, X.Y.; Wu, Z. F.; Chen,H.; Shao,J. R.; Wu,Q. Karyotype and genetic relationship based on RAPD markers of six wild buckwheat species (Fagopyrum spp.) from southwest of China. Genet. Resour. Crop Evol. 2010, 57, 649-655. [CrossRef]
- Zhang, H.Z.; Guan, Z. X.; Liu,X. Y.; Liu,Y. H. . Karyotype analysis of Fagopyrum exculetum and F. tataricum. J. Inn. Mong Agric. Univ. 2000, 21, 69-74.
- Zhou, M.L.; Bai, D. Q.; Tang,Y.; Zhu, X. M.; Shao,J. R. Genetic diversity of four new species related to southwestern Sichuan buckwheats as revealed by karyotype, ISSR and allozyme characterization. Plant Syst. Evol. 2012, 298, 751-759. [CrossRef]
- Zhu, F.S.; Lin,R. F.; Li,Y. Q.; Niu,D. K. The initial report of studies on different types of chromosomes of Buckwheat. Chin. J. Cell Biol. 1984, 6, 130-131.
- Li, M.X.; Chen, R.Y. A suggestion on the standardization of karyotype analysis in plants. J. Wuhan Bot. Res. 1985, 3, 297-302.
- Levan, A.; Fredya, K.; Sandeberg, A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201-220. [CrossRef]
- Wang, W.X.; Li, C. Y.; Xiang,S. Q.; Liang,G. L. . Karyotypes Analysis and 45S rDNA-FISH of the Tetraploid and Diploid in Carica papaya L. Acta Horticulturae Sinica 2007, 34, 345-348.
- Zhang, H.M.; Zhang, S. N.;Yu,X. H.;Hou,X. L. Karyotype Analysis of Autotetraploid Broccoli. Acta Bot. Boreali-Occident. Sin. 2010, 30, 63-67.
- Melo, C.A.F.; Martins, M.I.G.; Oliveira, M.B.M.; Benko-Iseppon, A.M.; Carvalho, R. Karyotype analysis for diploid and polyploid species of the Solanum L. Plant Syst. Evol. 2011, 293, 227-235. [CrossRef]
- Zheng, J.S.; Zhang, S. N.; Sun,C. Z.; Wang,J. J.; Hou,X. L. Karyotype analysis of diploid and autotetraploid non-heading Chinese cabbage. J. Nanjing Agric. Univ. 2012, 35, 131-134.
- Yuan, J.M.; Dang, X. M.; Zhan,Y. F.; Li,Y. R.; Dan,Z.; Su,Y. L.; Mu,W. F.; Yang,C. K. . Karyotype analysis of diploid and autotetraploid mini-watermelon. Nor. Horticul. 2013, 40-43.
- Gao, F.H.; He, L. J.; Chen,H. J.; Liu,J. W.; Shan,Y. M.; Wang,W. J.; Yu,L. X. . Chromosome doubling and polyploid karyotype analysis of wild Lycium ruthenium. Mol. Plant Breed 2020, 18, 7522-7529.
Figure 1.
The morphology, karyograms and idiograms of the eight diploid and autotetraploid chromosomes c. The morphology, karyograms, and idiograms of the diploid (T6, T9, T18, T22, T25, T27, T36, T39) and autotetraploid (T6*, T9*, T18*, T22*, T25*, T27*, T36*, T39*) chromosomes are represented by A, B ,C and D,E, F, respectively.
Figure 1.
The morphology, karyograms and idiograms of the eight diploid and autotetraploid chromosomes c. The morphology, karyograms, and idiograms of the diploid (T6, T9, T18, T22, T25, T27, T36, T39) and autotetraploid (T6*, T9*, T18*, T22*, T25*, T27*, T36*, T39*) chromosomes are represented by A, B ,C and D,E, F, respectively.
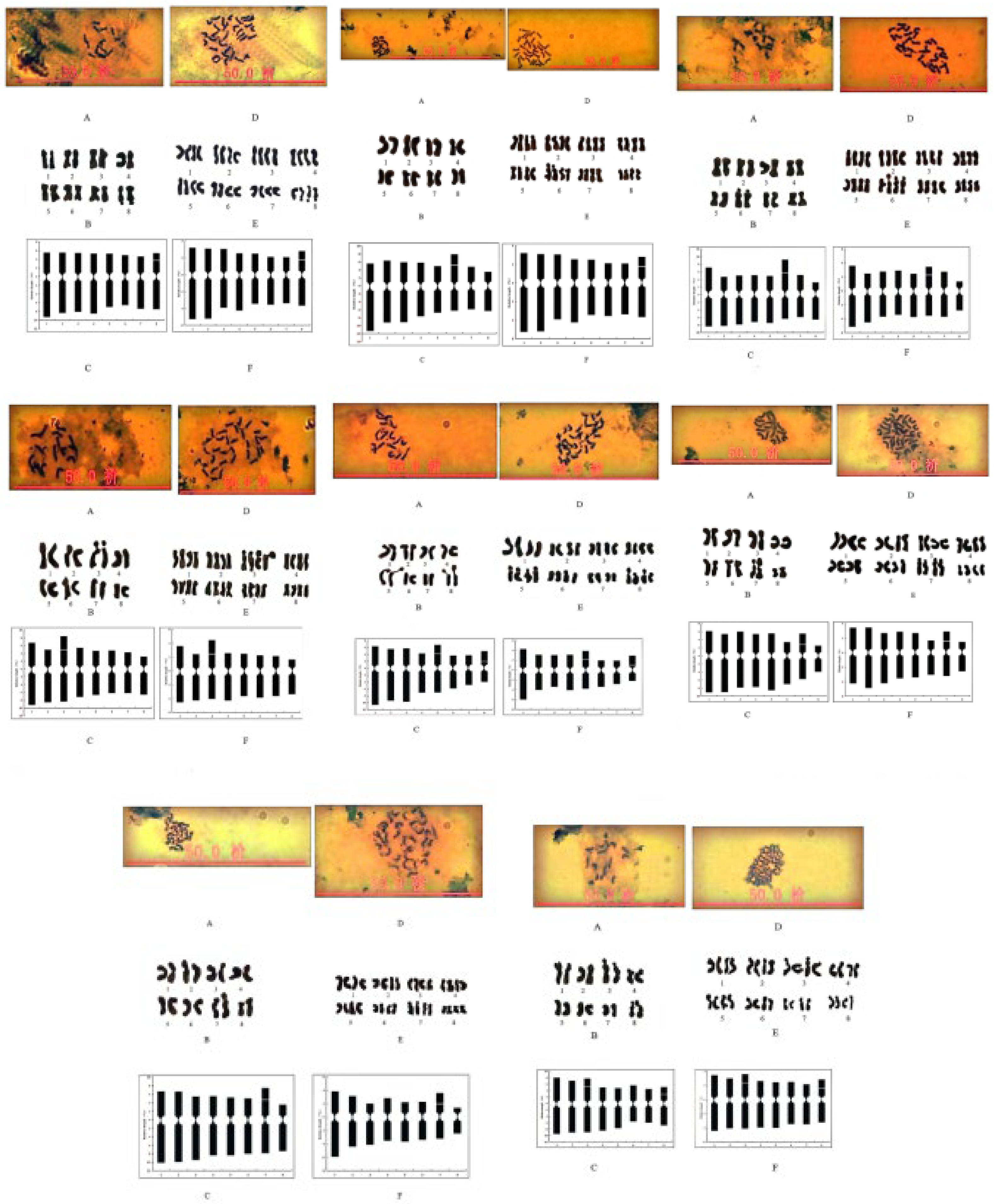

Table 1.
The F. tataricum accessions used as experimental materials in this study.
| Accessions (diploids) |
Spices | Sources | Accessions (autotetraploids) |
| T6 | Basu tartary buckwheat | Nala, Basu County, Tibet, China | T6* |
| T9 | WN090 | Weining, Guizhou, China | T9* |
| T18 | Wugang tartary Buckwheat | Wugang, Hunan, China | T18* |
| T22 | WN065 | Weining, Guizhou, China | T22* |
| T25 | Erjizao | Weining, Guizhou, China | T25* |
| T27 | Qianwei No.1 | Weining, Guizhou, China | T27* |
| T36 | Liuqiao No.1 | Liupanshui, Guizhou, China | T36* |
| T39 | Qianwei No.2 | Weining, Guizhou, China | T39* |
* Refers to autotetraploids obtained by chromosome doubling technique. The same below.
Table 2.
Significant characteristics of the eight diploid and autotetraploid chromosomes.
| Accessions | Karyotypic formula | A-SAT | P-SAT | Karyotpe type |
| T6 | 2n = 2x = 16 = 12m + 4sm (2SAT) | 2 | 8 | 2A |
| T6* | 2n = 4x = 32 = 24m + 8sm (4SAT) | 4 | 8 | 2A |
| T9 | 2n = 2x = 16 = 12m (2SAT) + 4sm | 2 | 6 | 2A |
| T9* | 2n = 4x = 32 = 24m(4SAT) + 8sm | 4 | 6 | 2A |
| T18 | 2n = 2x = 16 = 12m(2SAT) + 4sm | 2 | 6 | 2A |
| T18* | 2n = 4x = 32 = 24m(4SAT) + 8sm | 4 | 6 | 2A |
| T22 | 2n = 2x = 16 = 12m (2SAT) + 4sm | 2 | 3 | 2A |
| T22* | 2n = 4x = 32 = 24m(4SAT) + 8sm | 4 | 3 | 2A |
| T25 | 2n = 2x = 16 = 12m + 4sm(4SAT) | 4 | 5,8 | 2A |
| T25* | 2n = 4x = 32 = 24m + 8sm(8SAT) | 8 | 5,8 | 2A |
| T27 | 2n = 2x = 16 = 12m + 4sm(2SAT) | 2 | 7 | 2A |
| T27* | 2n = 4x = 32 = 24m + 8sm(4SAT) | 4 | 7 | 2A |
| T36 | 2n = 2x = 16 = 12m(2SAT) + 4sm | 2 | 7 | 2A |
| T36* | 2n = 4x = 32 = 24m(4SAT) + 8sm | 4 | 7 | 2A |
| T39 | 2n = 2x = 16 = 12m + 4sm(4SAT) | 4 | 3,8 | 2A |
| T39* | 2n = 4x = 32 = 24m + 8sm(8SAT) | 8 | 3,8 | 2A |
Table 3.
Karyotype parameters of the eight somatic diploid and autotetraploid chromosomes.
| Chromosomal code(Ploidy) | Accessions | ||||||||||||||||||||||||
| T6 | T9 | T18 | T22 | T25 | T27 | T36 | T39 | ||||||||||||||||||
| RL | AR | PC | RL | AR | PC | RL | AR | PC | RL | AR | PC | RL | AR | PC | RL | AR | PC | RL | AR | PC | RL | AR | PC | ||
| 1(2x) | 14.29 | 1.67 | m | 15.79 | 2.01 | sm | 15.14 | 1.17 | m | 15.73 | 1.36 | m | 17.06 | 1.66 | m | 15.38 | 1.46 | m | 14.59 | 1.50 | m | 15.90 | 1.19 | m | |
| 2(2x) | 13.37 | 1.51 | m | 14.48 | 1.44 | m | 12.60 | 1.74 | sm | 13.32 | 1.71 | sm | 15.18 | 1.68 | m | 14.57 | 1.65 | m | 14.37 | 1.49 | m | 14.69 | 1.27 | m | |
| 3(2x) | 13.03 | 1.48 | m | 14.03 | 1.53 | m | 12.58 | 1.55 | m | 13.06 | 1.69 | m# | 15.71 | 1.67 | m | 14.03 | 1.30 | m | 12.99 | 1.71 | sm | 12.85 | 1.74 | sm# | |
| 4(2x) | 13.32 | 1.59 | m | 12.55 | 1.30 | m | 12.00 | 1.40 | m | 12.45 | 1.30 | m | 11.55 | 1.64 | m | 13.30 | 1.41 | m | 12.10 | 1.50 | m | 12.21 | 1.70 | m | |
| 5(2x) | 11.83 | 1.31 | m | 10.89 | 1.36 | m | 12.11 | 1.51 | m | 11.19 | 1.42 | m | 11.25 | 1.71 | sm# | 14.24 | 1.52 | m | 11.83 | 1.58 | m | 11.32 | 1.62 | m | |
| 6(2x) | 11.10 | 1.32 | m | 10.55 | 1.22 | m# | 11.75 | 1.12 | m# | 10.81 | 1.29 | m | 9.68 | 1.58 | m | 10.54 | 2.02 | sm | 11.33 | 1.57 | m | 10.24 | 1.00 | m | |
| 7(2x) | 10.63 | 2.02 | sm | 9.93 | 1.22 | m | 10.81 | 1.14 | m | 10.33 | 1.43 | m | 8.74 | 1.29 | m | 8.56 | 2.07 | sm# | 10.83 | 1.68 | m# | 9.57 | 1.33 | m | |
| 8(2x) | 10.75 | 2.03 | sm# | 9.17 | 1.77 | sm | 9.54 | 2.01 | sm | 9.65 | 2.03 | sm | 5.90 | 2.08 | sm# | 6.55 | 1.53 | m | 9.49 | 2.04 | sm | 8.83 | 2.36 | sm# | |
| 1(4x) | 15.72 | 1.63 | m | 14.83 | 2.05 | sm | 16.68 | 1.42 | m | 15.25 | 1.26 | m | 19.02 | 1.39 | m | 15.25 | 1.22 | m | 20.30 | 1.53 | m | 14.76 | 1.27 | m | |
| 2(4x) | 15.41 | 1.66 | m | 13.27 | 1.64 | m | 13.35 | 1.80 | sm | 12.59 | 1.73 | sm | 13.43 | 1.29 | m | 16.54 | 1.39 | m | 15.69 | 1.39 | m | 13.27 | 1.34 | m | |
| 3(4x) | 12.96 | 1.26 | m | 12.93 | 1.70 | m | 12.36 | 1.32 | m | 12.12 | 1.67 | m# | 12.11 | 1.10 | m | 13.75 | 1.54 | m | 12.78 | 2.01 | sm | 12.10 | 1.75 | sm# | |
| 4(4x) | 12.63 | 1.65 | m | 12.40 | 1.49 | m | 12.13 | 1.16 | m | 12.85 | 1.65 | m | 13.51 | 1.30 | m | 12.75 | 1.21 | m | 13.20 | 1.23 | m | 12.33 | 1.50 | m | |
| 5(4x) | 11.07 | 1.36 | m | 12.14 | 1.25 | m | 11.81 | 1.51 | m | 11.52 | 1.45 | m | 11.18 | 1.72 | sm# | 12.13 | 1.25 | m | 12.20 | 1.63 | m | 12.11 | 1.63 | m | |
| 6(4x) | 10.55 | 1.66 | m | 11.76 | 1.47 | m# | 11.15 | 1.55 | m | 11.51 | 1.70 | m | 10.05 | 1.70 | m | 9.39 | 1.79 | sm | 11.81 | 1.46 | m# | 11.15 | 1.38 | m | |
| 7(4x) | 9.60 | 1.86 | sm | 11.18 | 1.61 | m | 12.35 | 1.34 | m | 10.86 | 1.67 | m | 8.91 | 1.45 | m | 9.48 | 2.01 | sm# | 10.73 | 1.63 | m | 10.60 | 1.60 | m | |
| 8(4x) | 10.05 | 2.08 | sm# | 9.57 | 1.78 | sm | 7.93 | 2.04 | sm | 9.44 | 2.02 | sm | 6.04 | 2.07 | sm# | 8.05 | 1.69 | m | 7.94 | 1.73 | sm | 9.01 | 2.02 | sm# | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



