
Submitted:
30 September 2024
Posted:
03 October 2024
You are already at the latest version
Abstract
The bioherbicidal potential of four essential oils (EOs) from Mediterranean species of Thymus sp. pl.: Thymus algeriensis Boiss. et Reut., T. ciliatus Desf. subspecies coloratus (Boiss. et Reut.) Batt., T. vulgaris L., ecotype Fasano and T. vulgaris L. var. Varico was investigated to identify new sources of biomolecules. In vivo assays were carried out to assess the effect of the EOs on pre and post-emergence of two weed species: Lolium perenne L. and Amaranthus retroflexus L.. Four concentrations of EO emulsions (500, 750, 1000 and 2000 µl/100 mL) were applied. All the EOs expressed an important bioherbicidal activity against the target species. A total inhibition of seed germination was displayed at 2000 µl whereas a potent effec-tiveness was achieved even at 500 µl. On the contrary, for post-emergence experiments, L. perenne showed resistance to the EOs while A. retroflexus was highly sensitive, arguing a selectivity of the EOs. Bioassays were also performed on the model plant Arabidopsis thaliana (L.) Heynh, for a better understanding of the structural modifications induced by the EOs. The root system of EO-treated A. thaliana seedlings was highly damaged.
Keywords:
Thyme
; bioherbicidal activity
; pre and post emergence
; A. thaliana (L) Heynh
; structural modification
1. Introduction
Chemical inputs especially pesticides, are among the most applied agrochemicals for pest control. Nevertheless, the repeated use of the same synthetic products led to an in-creased risk of the emergence of pest resistance and ecological hazards, which raises a new problem for global environment preservation [1,2]. This potential risk effect on global health, associated with the overuse of agrochemicals, has encouraged researchers to search for eco-friendly alternatives for pest control [3].
Among the long list of threats to crop production, weeds are the most damaging be-cause they compete for resources such as light, water, and nutrients and act as reservoirs of pests and pathogens [4,5]. In addition, in these last decades, weeds have developed resistance mechanisms to herbicides: target-site resistance, which involve gene mutations that affect herbicide protein targets, and non-target-site resistance, involving complex interactions of genes from large gene families responsible for reduced herbicide absorption, translocation, degradation, and sequestration. These resistance mechanisms can combine at the individual level, demonstrating the important adaptability of weeds to selection pressures [6].
In that regard and in the perspective of promoting integrated weed management (IWM), research on bio-herbicides has greatly increased [7]. The use of bioherbicides is presented as one of the most advantageous tools for IWM, demonstrating an increased target specificity, rapid degradation with maximum residue limits, and less restrictive-ness [7,8]. Thus, bioherbicides are developed, especially those of plant origin [9,10].The search for potential bioherbicides is typically based on the chemical ecology approach, in other words, on the allelopathic potential of plants. Plants can produce phytotoxic metabolites, called allelochemicals, to compete with other species [11,12]; the mode of action of these secondary compounds seems to be similar to synthetic herbicides. In this perspective, allelochemicals can act by altering seed germination or interfering with seedlings’ establishment [13]. Given their ecological role in natural environments, these secondary metabolites could be considered promising molecules to be employed for the production of new classes of plant-based herbicides [14,15].
In this context, essential oils (EOs) are widely studied as potential candidates for new-generation and eco-friendly herbicides for weed management, due to their important phytotoxicity. EOs demonstrated their potential as promising bioherbicides, thanks to their ability to inhibit seed germination and seedling growth, causing oxidative stress and disrupting photosynthesis and respiration mechanisms [16,17,18]. EOs are chemically composed of terpenes, which are derived from the universal C5 precursor isopentenyl di-phosphate (IPP) and its allylic isomer dimethylallyl diphosphate, which form a large diversity of compounds derived from the branched C5 skeleton of isoprene, including vola-tile monoterpenes (C10) and sesquiterpenes (C15) [19]. [20] explained that this diversity is generated by the modification of a common scaffold structure, to which diversity is added. Since the differential modification of common backbone structures can change and/or modify the biological activity, the new compounds can demonstrate a potential new biological activity [21,22,23,24]. In this sense, some of these secondary metabolites demonstrated a strong herbicidal potential against a large range of weed species and also microbial and insecticidal activities.
The Mediterranean area is one of the richest hotspots of biodiversity in the world with 24.000 species. Almost 2000 plants have been recorded, among them Thymus genus [25]. For this species, the biological effectiveness of its EOs has been demonstrated, especially thanks to their large chemotype diversity.
A large diversity of species belonging to this genus is present in different environments in Algeria and Italy, therefore this study focuses on the evaluation of the herbicidal activity of thyme EOs of two Algerian endemic species, harvested from natural environments, an Italian local variety and a Swiss commercial hybrid, cultivated in Italy.
2. Results
2.1. Herbicidal Activity of the EOs against Target Weeds
2.1.1. Pre-Emergence Assay
The herbicidal effect of the different EOs of thyme species on seed germination of L. perenne and A. retroflexus is presented in Figure 1. All the treatments inhibited or reduced seed germination of both target weed species in a dose-response manner and depending on the EO. At the concentration of 2000 μL an effective herbicidal potential for L. perenne was expressed by all the tested EOs, inducing a total inhibition of the germination. At 1000, no statistical differences were found between the EOs; all of them significantly inhibited seed germination, reducing it by over 90%. A similar results was obtained using the concentration of 750 μL for T1, T2 and T4, while T3 seemed less effective, even if, also at the lowest concentration (500 μL) the reduction of seed germination was between 83% (T1) and 77,5% (T3) (Figure 1a).
T2 expressed the strongest herbicidal effect on A. retroflexus, where a total inhibition of seed germination was induced at 2000 and 1000 μL. At 750 and 500 μL, it was extremely reduced at a percentage of 14.2 and 17.5, respectively, while the germination of the negative control was 91.7%. T3 demonstrated a total inhibition at 2000 μL and significantly slowed it down at the other concentrations (2.5, 10.0 and 13.3 % respectively). Contrary to what was observed for L. perenne, T1 and T4 expressed a less significant effectiveness in comparison with T2 and T3. An important reduction of seed germination was noticed for the other concentrations (Figure 1b). However, it is still important that a total inhibition was achieved at 2000 μL and a strong reduction in seed germination of about 80% was noted for the other concentration.
In summary, T1 and T4 expressed the higher herbicidal effect against L. perenne while T2 and T3 were more effective against A. retroflexus.
2.1.2. Post-Emergence Assay
Regarding the herbicidal effect on target species seedlings, A. retroflexus was more sensitive than L. perenne to thyme EOs treatments. L. perenne seedlings were affected by the treatments only at the highest concentration (2000 µl), where the mortality of all the plantlets was observed. For the other concentrations, all the seedlings were able to survive after the exposure to the treatments. On the contrary, A. retroflexus strongly reacted to the treatments from the first application. It displayed wilting symptoms few hours after the exposure to the EOs, inducing to the mortality of the plantlets in less than 24 hours. The fresh weight of A. retroflexus biomass subjected to the treatments was consistent with the number of surviving seedlings. This demonstrate the high sensitivity of A. retroflexus plantlets to the treatments (Figure 2).
The potent herbicidal effect of all the EOs on A. retroflexus, as compared to the negative control, was well demonstrated by T2 and T3. At the highest concentration (2000 µl), all the seedlings died, while the number of surviving ones was considerably reduced at 1000 µl. The same effectiveness was observed at 750 and 500 µl where the number of surviving plantlets was importantly reduced. T4 provoked the mortality of all the plantlets at the concentration of 2000 µl. The other concentrations induced an important mortality of the seedlings, where the number of the surviving ones did not exceed 10 plantlets for the lowest concentration (500 µl). T1, even if it expressed the less important herbicidal effect in comparison to the other EOs, reduced considerably the number of surviving plantlets. As for the other treatments, it induced a total mortality of the seedlings at 2000 µl, and drastically decreased it to less than 13 plantlets for 750 µl, which was the less effective dose (Figure 3a). The results of the fresh biomass of A. retroflexus are correlated with those of the number of surviving plants (Figure 3b).
Furthermore, it is important to note that as for the surviving plantlets, there were no significant differences in the herbicidal effect of the treatments on L. perenne plant biomass in comparison to the negative control; only the highest concentration (2000 µl) exhibited an effective effect of the treatments on the seedlings. For all the treatments, the biomass was null at 2000 µl and insignificant at 1000, 750 and 500 µl.
2.3. Phytotoxic activity of EOs on Arabidopsis thaliana
The mode of action of the studied EOs was evaluated on the model plant A. thaliana. The EOs treatments did not significantly affected A. thaliana germination; however, the root growth and development of the EO-treated seedlings were significantly affected. After 14 days of growth, EO-treated seedlings were visualized under a stereomicroscope to evaluate the morphological changes induced by the EOs on the seedlings. Several anomalies were observed depending on the EO. Roots were either shorter, thinner, with visible proliferation of adventitious roots, with important condensation or without root hair in comparison to that of control (Figure 4). Furthermore, EOs reduced the number and length of the root hairs. Some A. thaliana seedlings even presented a loss of gravitropism.
The microscopic observation of roots with Evans Blue staining showed a clear membrane damage induced by the treatments which is in accordance with what was reported by [26,29], who established that Evans Blue only penetrate and color cells that have lost their membrane integrity. In this sense, all the EO-treated roots were blue stained with different intensity. This means that the effect was also correlated with the EO. T2 showed the most intense blue color arguing an important damage to the cell membrane integrity, while T1, T3 and T4 displayed low to moderate blue stained roots mostly in the root cap (Figure 5).
The histochemical detection of lipid peroxidation of the roots induced by the EOs was performed using Schiff’s reagent. This reagent reacts with aldehydes from the lipid peroxidation inducing to a magenta coloration of the tissues. The microscopic observations showed, in this sense, magenta stained EO-treated roots, demonstrating a membrane degradation. However, as for Evans Blue staining, the intensity of the red color differed according to the EO applied; EO-treated roots with T2 displayed the most intense coloration, followed by T1, which indicates an important lipid peroxidation. T3 and T4 treated roots showed low coloration but with evident torsions and structural modifications (Figure 6).
It is worthy to mention also that structural modifications have been observed on EO-treated roots. A. thaliana control roots demonstrated a normal growth with homogenous and symmetric cell rows and layers corresponding to the different root tissues in the division, similar thickness along their entire length and growing straight according to gravitropism, while EO-treated roots presented an evident and significant tissue disruption. The calyptra was thicker and the cell rows were disorganized and hardly distinguishable. Torsions with zig-zag development were also observed in the elongation zone; and in some samples the growth patterns were completely altered, showing toxic effects, such as contraction phenomena. In some EO-treated seedlings, the columella, including the statocytes, was destructed; this could explain the loss of the gravity perception of the roots.
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
3. Discussion
The results obtained from the greenhouse experiments demonstrated an important herbicidal activity of the studied thyme EOs. All the EOs showed efficacy in negatively affecting seed germination of both weed test species at the highest concentration and drastically reducing it at lower concentrations. However, this effectiveness was selective on seedlings growth. L. perenne demonstrated a resistance where treatments were effective only at the highest concentration. On the contrary, A. retroflexus was highly sensitive to the application of the EOs at all concentrations. Globally, these results are highly consistent with those reported by the literature. Indeed, the herbicidal potential of Thymus sp. pl. EOs were investigated by several researchers who demonstrated significant efficacy in inhibiting seed germination and seedling growth of indicator test and weed species [24,27,28,29,30,31,32,33].
The negative impact of EOs on seed germination and seedling growth could be associated with physiological disturbances of the treated plant species. Secondary metabolites affect the vigor of the seeds by acting directly in the processes of cell and membrane degradation [34,35], which according to [36], disturb the leakage of electrolytes and provoke the loss of selectivity of the membrane. This disturbance can inhibit the synthesis of ATP and increase the production of reactive oxygen species, causing the rupture of membranes and leading to cell necrosis and germination reduction. The effect of these thyme EOs can also be responsible for the decrease of the weed seeds' vigor. Indeed, the seed vigor is related to the integrity of the cell membrane system; and when the seeds are immersed in the treatments, cytoplasmic solutes are released in the medium, proportionally to the state of disorganization in which the membranes are found [36].
The inhibition and/or the slowdown of seedling growth by thyme EOs, in turn, could be associated with the disturbance of photosynthesis inducing to an oxidative stress [23,37]. [38] argue that the inhibition of plant growth by EOs may occur via the generation of reactive oxygen species (ROS), subsequent oxidative stress, characterized primarily by markers specifically, and increase of malondialdehyde and proline concentrations. For [39,40,41] the exposure to EO causes oxidative stress in recipient species by activating several signaling pathways, causing metabolic damage. This could explain the results obtained in the present study. Plant tissues can become oxidized and reactive oxygen species (ROS), primarily hydrogen peroxide (H2O2), can accumulate and cause irreversible damage to plasma membranes when defense mechanisms are inefficient. The ROS causes damage on proteins, DNA, lipids, cell membranes and photosynthetic pigments, and finally leads to plant mortality [41].
In this sense and addressing plant defense mechanisms, L. perenne seedlings displayed a significant resistance while A. retrofluxus expressed a potent sensitivity to the EO treatments. These results coincide with those of [42] on D. viscosa. It is suggested that there is a selectivity in the action of bioactive molecules regarding the target plant species and their types, whether monocots or dicots [43,44,45]. [46] suggested that the seedlings release bioactive molecules in the medium that might contrast the effect of the applied treatments. [47] abounded in this sense and demonstrated that the effectiveness of the treatments depended on the targeted weed species and the type of EOs. In this regard, L. perenne is a notable resistant weed species to herbicides where resistance to 14 mechanisms of action (MOA) has been confirmed in this species [48]. According to [49], this is due to the high genetic diversity of Lolium spp., an obligate outcrossing mating system and a recurrent selection with a herbicide might lead to metabolic cross-resistance to other herbicides even from different MOA [49,50]. The results of this study revealed the high selectivity of thyme EOs and this could be interesting for the development of selective bio-herbicides.
In that regard, it has been reported that monoterpene-rich EOs significantly affect weed germination and growth rather than sesquiterpene-rich EOs [16,51], which sup-ports our findings. The tested thyme EOs used in this study contain an important percentage of monoterpenes, particularly, α-pinene, thymol, carvacrol, o-cymene and linalool. Oxygenated monoterpenes (essentially thymol, carvacrol, and p-cymene) are known to have potent phytotoxic effects, resulting in the inhibition of α-amylase, suppression of seed water uptake, disturbance of mitochondrial respiration, as well as interference with cell division processes [52,53]. Furthermore, others identified compounds in the thyme EOs, such as p-cymene, γ-terpinene, and linalool have been confirmed to possess phytotoxic effects against many weed species [54,55,56]. However, it is important to highlight that the phytotoxic potential of EOs can be attributed to antagonistic effects due to interactions among EO constituents, even those present at low concentrations [57]. In addition, the molecular structure of each component can be responsible for the specific MOA [58].
The in vitro experiments on the model plant A. thaliana were performed to better understand the alterations caused by thyme EOs. Morphological and structural damages were observed, mainly on the root part of the seedlings. The EO-treated seedlings presented, among others, a zig-zag and torsion growth of the roots and a destruction of the calyptra. These observations of the alterations of A. thaliana exposed to different EOs have been wildly reported in plants exposed to allelochemicals. [59], in their study of the effect of carvacrol on A. thaliana, established that this secondary metabolite caused shorter and thinner roots of the treated seedlings. The roots were also characterized by torsion and zig-zag development, reduction of the number and length of root hairs at the transition zone between hypocotyl and epicotyl, an increased number of ectopic roots close to the root tip and roots growing in different directions related to the loss of gravitropism. According to [19,60], these morphological and structural modifications and alterations could be attributed to the related to microtubule condensation and disorganization, the strong increment in ethylene production and unbalanced auxin content; [61] reached the same results. Treated seedlings with norharmane, an indol alkaloid, presented an inhibition of the growth with the formation of adventitious roots. This phenomenon is a phytohormone-driven process, particularly auxin, responsible for the formation of adventitious roots [62]. Therefore, the morphology of seedlings exposed to, for example, T. algeriensis EO (T1), which have short roots with more adventitious roots and root hairs than the control, suggests that the EO could be responsible for an alteration in auxin synthesis or polar auxin transport (PAT) [61]. This latter suggests that EO affects polar auxin transport (PAT) by reducing the number of PIN2, PIN3 and PIN7 proteins. This could provoke the accumulation of auxin in several root areas, without reaching the root cap, resulting in the adventitious root formation. This alteration in transport also has a negative effect on microtubules, causing inhibitory processes linked to cell elongation. Moreover, ethylene stimulates acropetal auxin transport in the plant roots, which is associated with a reduction of lateral root formation in a PIN3 and PIN7-dependent way [63]. [64] established the synergistic role played by auxin and ethylene in inhibiting lateral root initiation. At high concentrations, ethylene inhibits the ability of pericycle cells to initiate new lateral root primordia. These results are in accordance with those obtained in this study. EOs might increase endogenous ethylene concentration in the EO-treated seedlings roots leading to the inhibition of the emergence of lateral roots, as per T. vulgaris cultivar Varico (T3) EO-treated seedlings.
Furthermore, the root morphology in A. thaliana is regulated by ethylene and auxin, agonistically or antagonistically [65,66]. Our results suggest that the ultrastructure alteration, probably due to auxin and/or ethylene accumulation, and the macroscopic effects observed on root structure are linked. Moreover, thyme EOs, probably through the stimulation of auxin production, could interfere with microtubule dynamics by acting as a stabilizer of the tubulin, modifying the proper orientation of microtubules, which are essential components of cell cytoskeleton. Indeed, they are involved in vital processes, including cell division, growth and differentiation. These activities are highly dependent on the dynamic balance between the different subunits that form the microtubules [67]. When this balance is altered, the orientation of microtubules in the cell is affected, as well as all the cellular processes depending on them. The observations made in this study stipulate the clear evidence of altered microtubule organization, including the disruption of mitotic and cortical microtubules and by inducing ultra-structural cell malformations, thus modifications on root morphology through altering the cross-talk between auxin and ethylene. The results obtained from the experiments on A. thaliana, point out that the mode of action of thyme EOs could be probably related to the alteration of the hormonal balance and bio-chemical reactions, inducing morphological damages and generating the accumulation of ROS as a side product in recipient plants.
4. Materials and Methods
4.1. Plant material and EOs Extraction and Characterization
Aerial part of plant samples of four Thymus species: T. algeriensis Boiss. et Reut., T. ciliatus Desf. subsp. coloratus (Boiss. et Reut.) Batt., T. vulgaris L. cultivar Varico 3 and T. vulgaris L: ecotype Fasano, referred in the document, respectively, as T1, T2, T3 and T4, were collected at full flowering stage during May and June 2022 from the Chrea National Park and Hammam Melouane (Blida, Algeria) and Policoro (Basilicata, Italy) [68]. EOs were extracted by hydro-distillation and identified by gas chromatography coupled with mass spectrometry in a previous study [68]. The chemical composition of the EOs is presented in Table 1. EOs were stored at 4°C until their use in the biological activity.
4.2. Bioherbicidal Activity of the EOs against Target Weeds
Two weed species Lolium perenne L. and Amaranthus retroflexus L. were used to assess the herbicidal activity of the EOs on germination and early growth under in vivo conditions. Seeds were purchased from Weberseeds Company and stored at 4 °C for use during the experiments.
The herbicidal potential of thyme EOs against the test weed species was estimated in dose–response bioassays on seed germination (pre-emergence) and seedling early growth (post-emergence), under greenhouse conditions. Emulsions of T1, T2, T3 and T4 were prepared using 2% Tween® 20 (Sigma- Aldrich, Milan, Italy) as an emulsifying agent, according to [69]. Four concentrations (500, 750, 1000 and 2000 µl/100 ml of 2% of Tween® 20 solution) were applied. Water and a pelargonic acid based bio-herbicide (Vithal, 51.9% w/w pelargonic acid) were considered, respectively, as negative and positive controls. The concentration of the positive control was prepared as indicated in the label of the product.
Peat was used as substrate in these experiments with the purpose to evaluate the behavior of the test species in response to the treatments in controlled conditions. Trays of 21x16 cm were fitted with 2 cm drainage layer of perlite and 700 g of peat. All trays were irrigated and brought to their optimal water holding capacity and left to leach the excess of water.
4.2.1. Pre-emergence assay
Fifty (50) seeds of L. perenne L. and A. retroflexus L. were sown in each tray, then 10 ml of each treatment was spread using a micro-sprayer to guarantee its homogeneity. The number of germinated seeds was counted after fifteen (15) days and seed germination rate was calculated according to the following formula [70]:
where: GP: germination percent; n: number of germinated seeds at final count; N: total number of seeds.
Trials were conducted in a complete randomized design with three replicates. The trays were placed in the greenhouse at 22-25°C and 50 to 60% of humidity and were monitored daily for irrigation.
4.2.2. Post-Emergence Assay
For this test, pre-cultivation of the weed species was required. Seeds were allowed to germinate in nursery trays (72 holes) until the emergence of two true leaves. Then, fifty (50) seedlings of L. perenne and A. retroflexus were transplanted in trays already prepared as described previously. The seedlings received 10 ml of each treatment once they reached the phenological stage “three true leaves”, corresponding to 12–13 BBCH (Biologische Bundesanstalt, Bundessortenamt and Chemical industry) scale for L. perenne and “three to four true leaves”, corresponding to 13–14 BBCH scale for A. retroflexus. The EOs emulsions were applied using a micro-sprayer as contact treatments. The treatments were applied until the appearance of wilting symptoms on the plantlets.
The trays were placed in the greenhouse at 22-25°C and 50 to 60% of humidity and were monitored daily for irrigation.
All the treatments and controls were replicated three times in a complete randomized design.
At the end of the experiment, the number of surviving plantlets and fresh weight were determined. The fresh biomass was measured using a weighing scale on the same day.
4.3. Phytotoxic Activity of EOs on Arabidopsis thaliana (L.) Heynh
To evaluate the physiological damages caused by the EOs on the target weed species, assays on Arabidopsis thaliana were conducted. Seeds of A. thaliana (L.) Heynh Columbia ecotype (Col-0) were surface sterilized with ethanol (50%) and NaOCl (0.5%) for 3 min each and washed three times with sterilized distilled water, then stored in agar (0.1%) at 4 °C for 72 h to favor vernalization. Meanwhile, Petri dishes (90 mm) with agar medium (0.8% w/v) enriched with 0.44% Murashige-Skoog growth medium and supplemented with 1% sucrose [26,59] were prepared.
In order to determine the phytotoxic effect of the studied EOs on the growth of A. thaliana, emulsions were prepared with EOs at the concentration of 1000 µM using 0.1% EtOH (v:v) as solvent and added to the growth medium. Control plates included 0.1% EtOH. Afterward, twenty seeds were sown in each Petri dish under sterile conditions. All the Perti dishes were kept vertically in growth chamber at 22°C, with a photoperiod of 8 h light/16 h darkness, and a relative humidity of 55% for 14 days [59].
Each treatment and control was prepared in triplicate. The morphology was analyzed at the 14th day using a magnifier (Nikon SMZ 1500) and light microscope (Leica DMLB100, Milan, Italy). Root structure, thickness, morphology and abundance of root hairs of EOs-treated and control radicles of A. thaliana were evaluated using a magnifier (Nikon SMZ 1500) equipped with Nikon DXM 1200 Digital Camera (Tokyo, Japan). At the same time, Schiff’s staining method was used as histochemical indicator of membrane damage due to membrane peroxidation, while, Evans blue staining method was followed for the determination of cell viability and root plasma membrane integrity [71]. Schiff’s reagent (Carlo Erba Reagents s.r.l.) was prepared and applied to three roots from each replicate in test tubes and left to react for 20 min. After this, the reagent was removed from the tubes and root samples were rinsed with a sulfite solution to retain the staining color (0.5% w/v; K2S2O5 or Na2S2O5 in 0.05 M HCl). The solution of Evan blue (Carlo Erba reagents s.r.l.) (10 mg/mL) was placed in test tubes containing three roots for each replicate for 15 min. After this, the stain was removed from the tubes and the roots were rinsed three times with distilled water. Afterward, three roots were taken from different replicates per treatment. The terminal apex of the roots was cut and photographed under a microscope (Leica DMLB100, Milan, Italy) at 200x magnification equipped with Nikon DXM 1200 Digital Camera (Tokyo, Japan).
4.4. Statistical Analysis
Data obtained were statistically analyzed using Minitab® version 19.2020.1 (Minitab software, State College, Pennsylvania, USA). Analysis of the variance (one-way ANOVA) of the seed germination percentage, number of surviving plantlets and fresh biomass of the two target weed species was performed to assess the herbicidal activity of the treatments. Difference among means was run using Tukey’s test. Statistical significance was accepted when the probability of the result assuming the null hypothesis (p) was less than 0.05 (p<0.05) [72].
5. Conclusions
The present study demonstrated that all Thymus sp. pl. EOs tested exhibited a significant herbicidal potential against L. perenne and A. retroflexus. Thyme EOs were strongly effective as pre-emergence treatments by either totally inhibiting or drastically slowing down seed germination. This effectiveness was also obtained as a post-emergence treatment against A. retroflexus. On the contrary, L. perenne expressed a resistance reaction. This achievement argues a selectivity of the herbicidal potential of thyme EOs. As for the A. thaliana experiment, the results revealed a likely hormonal balance alteration.
Taken together, these results demonstrated that thyme EOs altered the hormonal balance of the recipient weed species, inducing a possible increase of both auxin and ethylene content, and stimulated oxidative damage. Both hormones and ROS could alter the nor-mal plant growth interfering with cell division and cytokinesis. The discovery of natural molecules that present in thyme EOs, characterized by a potent herbicidal potential is important for the development of a selective new bio-chemicals able to overcome the use of synthetic inputs. However, due to their chemical instability, EOs can be easily degraded, a way to protect them could be the nano-encapsulation as it might protect them and enhance their stability against environmental conditions; future studies on the preparation of bio-nanostructured systems to encapsulate thyme EOs would allow not only to investigate and better understand thyme EOs’ effects on recipient plant metabolism but could be interesting to enhance their stability while preserving their efficacy as bio-herbicide.
Author Contributions
Author Contributions: Conceptualization, G.D.M., C.R..; data curation, R.B., S.M.-M., M.P.A., C.R. and G.D.M.; investigation, R.B., S.M.-M., M.P.A., C.R. and G.D.M.; project administration, G.D.M. and G.J.C.; supervision, R.B., S.M.-M., G.J.C., M.P.A., C.R. and G.D.M.; validation R.B., S.M.-M., M.P.A., C.R. and G.D.M.; writing—original draft preparation, R.B. G.J.C., and S.M.-M.; writing—review and editing, R.B., C.R. and G.D.M. All authors have read and agreed to the published version of the manuscript. on.
Funding
This study was carried out within the Agritech National Research Center and received funding from the European Union Next-GenerationEU (PIANO NAZIONALE DI RIPRESA E RESILIENZA (PNRR)—MISSIONE 4 COMPONENTE 2, INVESTIMENTO 1.4—D.D. 1032 17/06/2022, CN00000022). This manuscript reflects only the authors’ views and opinions, neither the European Union nor the European Commission can be considered responsible for them.
Data Availability Statement
The original contributions presented in this study are included in the article, and further inquiries can be directed to the corresponding author.
Acknowledgments
The authors wish to acknowledge A. Tagarelli for her technical support and G. N. Basile for his valuable contribution.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Oyekunle, J.A.O.; Adekunle, A.S.; Adewole, A.M. Determination of organochlorine pesticide residues in some evaporated milk samples in Nigeria using gas chromatography-mass spectrometry. Chem. Afr. 2021, 4, 349–366. [Google Scholar] [CrossRef]
- Fahed, C.; Hayar, S. Conventional vs. organic cucumber production in Lebanon: Risk assessment of the recommended agrochemicals on consumer health and the environment. Chem. Afr, 4. [CrossRef]
- Oliveira, A.F.; Costa Junior, L.M.; Lima, A.S.; Silva, C.R.; Ribeiro, M.N.S.; Mesquista, J.W.C.; Rocha, C.Q.; Tangerina, M.M.P.; Vilegas, W. Anthelmintic activity of plant extracts from Brazilian savanna. Vet. Parasitol. 2017, 236, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Fried, G.; Chauvel, B.; Reynaud, P.; Sache, I. Decreases in crop production by non-native weeds, pests and pathogens. In Impact of Biological Invasions on Ecosystem Services; Vilà, M., Hulme, P.E., Eds.; Springer: Cham, Switzerland, 2017; pp. 83–101. [Google Scholar]
- Byron, M.; Treadwell, D.; Dittmar, P. Weeds as reservoirs of plant pathogens affecting economically important crops. EDIS 2019, 5, 7–7. [Google Scholar] [CrossRef]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of evolved herbicide resistance. J. Biol. Chem. 2020, 295(30), 10307–10330. [Google Scholar] [CrossRef]
- Cordeau, S.; Triolet, M.; Wayman, S.; Steinberg, C.; Guillemin, J.P. Bioherbicides: Dead in the water? A review of the existing products for integrated weed management. Crop Prot. 2016, 87, 44–49. [Google Scholar] [CrossRef]
- Olson, S. An analysis of the biopesticide market now and where it is going. Outlooks Pest. Manag. 2015, 26, 203–206. [Google Scholar] [CrossRef]
- Hasan, M.; Ahmad-Hamdani, M.S.; Rosli, A.M.; Hamdan, H. Bioherbicides: An eco-friendly tool for sustainable weed management. Plants 2021, 10, 1212. [Google Scholar] [CrossRef]
- De Souza Barros, V.M.; Pedrosa, J.L.F.; Gonçalves, D.R.; de Medeiros, F.C.L.; Carvalho, G.R.; Gonçalves, A.H.; Teixeira, P.V.V.Q. Herbicides of biological origin: A review. J. Hortic. Sci. Biotechnol. 2021, 96, 288–296. [Google Scholar] [CrossRef]
- Vurro, M.; Boari, A.; Casella, F.; Zonno, M.C. Fungal phytotoxins in sustainable weed management. Curr. Med. Chem. 2018, 25, 268–286. [Google Scholar] [CrossRef]
- Palanivel, H.; Tilaye, G.; Belliathan, S.K.; Benor, S.; Abera, S.; Kamaraj, M. Allelochemicals as natural herbicides for sustainable agriculture to promote a cleaner environment. In strategies and tools for pollutant mitigation; Aravind, J., Kamaraj, M., Prashanthi Devi, M., Rajakumar, S., Eds.; Springer: Cham, Switzerland, 2021; pp. 93–116. [Google Scholar]
- Hierro, J.L.; Callaway, R.M. The ecological importance of allelopathy. Annu. Rev. Ecol. Evol. Syst. 2021, 52, 25–45. [Google Scholar] [CrossRef]
- Araniti, F.; Mancuso, R.; Lupini, A.; Giofre, S.V.; Sunseri, F.; Gabriele, B.; Abenavoli, M. R. Phytotoxic potential and biological activity of three synthetic coumarin derivatives as new natural-like herbicides. Molecules 2015, 20(10), 17883–17902. [Google Scholar] [CrossRef]
- Berestetskiy, A. Modern approaches for the development of new herbicides based on natural compounds. Plants 2023, 12(2), 234. [Google Scholar] [CrossRef]
- Verdeguer, M.; Sanchez-Moreiras, A.M.; Araniti, F. Phytotoxic effects and mechanism of action of essential oils and terpenoids. Plants 9, 1–48. [CrossRef] [PubMed]
- Werrie, P.Y.; Durenne, B.; Delaplace, P.; Fauconnier, M.L. Phytotoxicity of essential oils: opportunities and constraints for the development of biopesticides. A review. Foods 2020, 9(9), 1291. [Google Scholar] [CrossRef] [PubMed]
- Pouresmaeil, M.; Nojadeh, M. S.; Movafeghi, A.; Maggi, F. Exploring the bio-control efficacy of Artemisia fragrans essential oil on the perennial weed Convolvulus arvensis: Inhibitory effects on the photosynthetic machinery and induction of oxidative stress. Ind. Crops Prod. 2020, 155, 112785. [Google Scholar] [CrossRef]
- Araniti, F.; Graña, E.; Krasuska, U.; Bogatek, R.; Reigosa, M.J.; Abenavoli, M.R.; Sanchez-Moreiras, A.M. Loss of gravitropism in farnesene-treated Arabidopsis is due to microtubule malformations related to hormonal and ROS unbalance. PloS one 2016, 11(8), e0160202. [Google Scholar] [CrossRef]
- Trindade, H.; Pedro, L. G.; Figueiredo, A. C.; Barroso, J. G. Chemotypes and terpene synthase genes in Thymus genus: State of the art. Ind. Crops Prod. 2018, 124, 530–547. [Google Scholar] [CrossRef]
- Kong, Q.; Zhou, L.; Wang, X.; Luo, S.; Li, J.; Xiao, H.; Zhang, X.; Xiang, T.; Feng, S.; Chen, T.; Yuan, M. Chemical composition and allelopathic effect of essential oil of Litsea pungens. Agronomy 2021, 11(6), 1115. [Google Scholar] [CrossRef]
- Casella, F.; Vurro, M.; Valerio, F.; Perrino, E.V.; Mezzapesa, G.N.; Boari, A. Phytotoxic effects of essential oils from six Lamiaceae species. Agronomy 2023, 13(1), 257. [Google Scholar] [CrossRef]
- Li, J.; Chen, H.; Guo, C.; Chen, Q.; Zhao, T.; Chen, X.; Du, Y.; Du, H.; Miao, Y.; Liu, D. Artemisia argyi essential oil exerts herbicidal activity by inhibiting photosynthesis and causing oxidative damage. Ind. Crops Prod. 2023, 194, 116258. [Google Scholar] [CrossRef]
- Miloudi, S.; Abbad, I.; Soulaimani, B.; Ferradous, A.; Abbad, A. Optimization of herbicidal activity of essential oil mixtures from Satureja alpina, Thymus satureioides and Myrtus communis on seed germination and post-emergence growth of Amaranthus retroflexus L. Crop Prot. 2024, 106642. [Google Scholar] [CrossRef]
- Vargas, P. The Mediterranean floristic region: high diversity of plants and vegetation types. In: Goldestein M.I. and Dellasala D.A. (eds). Encyclopedia of the World's Biomes. Netherlands: Elsevier Science, 2020, pp. 602-616.
- Díaz-Tielas, C.; Graña, E.; Sotelo, T.; Reigosa, M.J.; Sánchez-Moreiras, A.M. The natural compound trans-chalcone induces programmed cell death in Arabidopsis thaliana roots. Plant. Cell Environ. 2012, 35, 1500–1517. [Google Scholar] [CrossRef] [PubMed]
- Benchaa, S.; Hazzit, M.; Zermane, N.; Abdelkrim, H. Chemical composition and herbicidal activity of essential oils from two Labiatae species from Algeria. J Essent. Oil Res. 2019, 31(4), 335–346. [Google Scholar] [CrossRef]
- Saleh, I.; Abd-ElGawad, A.; El Gendy, A.N.; Aty, A.A.; Mohamed, T.; Kassem, H.; Aldorsi, F.; Elshamy, A.; Hegazy, M.E.F. Phytotoxic and antimicrobial activities of Teucrium polium and Thymus decussatus essential oils extracted using hydrodistillation and microwave-assisted techniques. Plants 2020, 9, 716. [Google Scholar] [CrossRef]
- Ghasemi, G.; Alirezalu, A.; Ghosta, Y.; Jarrahi, A.; Safavi, S.A.; Abbas-Mohammadi, M.; Barba, F.J.; Munekata, P.E.; Domínguez, R.; Lorenzo, J.M. Composition, antifungal, phytotoxic, and insecticidal activities of Thymus kotschyanus essential oil. Molecules 2020, 25(5), 1152. [Google Scholar] [CrossRef]
- Portuguez-García, M.P.; Agüero-Alvarado, R.; González-Lutz, M.I. Herbicidal activity of three natural products on four weed species. Agronomía Mesoamericana.
- Zhou, S.; Han, C.; Zhang, C.; Kuchkarova, N.; Wei, C.; Zhang, C.; Shao, H. Allelopathic, phytotoxic, and insecticidal effects of Thymus proximus Serg. essential oil and its major constituents. Front. Plant Sci. 2021, 12, 689875. [Google Scholar] [CrossRef]
- Vaiciulyte, V.; Loziene, K.; Svediene, J.; Raudoniene, V.; Paskevicius, A. α-Terpinyl acetate: occurrence in essential oils bearing Thymus pulegioides, phytotoxicity, and antimicrobial effects. Molecules 2021, 26, 26,1065–1571. [Google Scholar] [CrossRef]
- Elghobashy, R.M.; El-Darier, S.M.; Atia, A.M.; Zakaria, M. Allelopathic potential of aqueous extracts and essential Oils of Rosmarinus officinalis L. and Thymus vulgaris L. J. Soil Sci. Plant Nutr. 2024, 24(1), 700–715. [Google Scholar] [CrossRef]
- Qian, H.; Xu, X.; Chen, W.; Jiang, H.; Jin, Y.; Liu, W.; Fu, Z. Allelochemical stress causes oxidative damage and inhibition of photosynthesis in Chlorella vulgaris. Chemosphere. [CrossRef]
- Silva, V.S.; Cândido, A.C.S.; Muller, C.; Laura, V.A.; Faccenda, O.; Hess, S.E.S.C.; Peres, M.T.L.P. Potencial ftotóxico de Dicranopteris fexuosa (Schrad.) Underw. (Gleicheniaceae). Acta. Bot. Bras.
- De Oliveira Roberto, C.E.; Pinheiro, P.F.; de Assis Alves, T.; da Silva, J.A.; Praça-Fontes, M.M.; Soares, T.C.B. Phytogenotoxicity of thymol and semisynthetic thymoxyacetic acid in pre/post emergence of model plants and weeds. Environ. Sci. Pol. Res. 2023, 30(13), 38955–38969. [Google Scholar] [CrossRef]
- Hasan, M.; Mokhtar, A.S.; Mahmud, K.; Berahim, Z.; Rosli, A.M.; Hamdan, H.; Motmainna, M.; Ahmad-Hamdani, M.S. Physiological and biochemical responses of selected weed and crop species to the plant-based bioherbicide WeedLock. Sci. Rep. 2022, 12(1), 19602. [Google Scholar] [CrossRef]
- Amri, I.; Hamrouni, L.; Hanana, M.; Gargouri, S.; Fezzani, T.; Jamoussi, B. Chemical composition, physico-chemical properties, antifungal and herbicidal activities of Pinus halepensis Miller essential oils. Bio. Agri. Hort. 2013, 29(2), 91–106. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Rodríguez-Serrano, M.; Corpas, F.J.; Gomez, M.D.; Del Río, L.A.; Sandalio, L.M. Cadmium-induced subcellular accumulation of O2 and H2O2 in pea leaves. Plant Cell Environ. 2004, 27, 1122–1134. [Google Scholar] [CrossRef]
- Silva, E.R.; Lazarotto, D.C.; Schwambach, J.; Overbeck, G.E.; Soares, G.L.G. Phytotoxic effects of extract and essential oil of Eucalyptus saligna (Myrtaceae) leaf litter on grassland species. Austr. J. Bot. 2017, 65, 172–182. [Google Scholar] [CrossRef]
- Bruxel, F.; Schneider, C.E.; Gastmann, J.; Orlandi, C.R.; Gastmann, R.; Hoehne, L.; Soares, G.L.G.; Ethur, E.M.; Sperotto, R.A.; de Freitas, E.M. Phytotoxicity of Hesperozygis ringens (Benth.) Epling essential oil on Eragrostis plana Nees. Flora. [CrossRef]
- Omezzine, F.; Rinez, A.; Ladhari, A.; Farooq, M.; Haouala, R. Allelopathic potential of Inula viscosa against crops and weeds. Inter. J. Agri. Bio. 2011, 13(6), 841–849. [Google Scholar]
- Tuyen, P.T.; Xuan, T.D.; Tu Anh, T.T.; Mai Van, T.; Ahmad, A.; Elzaawely, A.A.; Khanh, T.D. Weed suppressing potential and isolation of potent plant growth inhibitors from Castanea crenata Sieb. et Zucc. Molecules 2018, 23(2), 345. [Google Scholar] [CrossRef]
- Lebedev, V.G.; Krutovsky, K.V. ; Shestibratov; K.A. …Fell upas sits; the hydra-tree of death; or the phytotoxicity of trees. Molecules, 1636. [Google Scholar] [CrossRef]
- Jmii, G.; Molinillo, J. M.; Zorrilla, J. G.; Haouala, R. Allelopathic activity of Thapsia garganica L. leaves on lettuce and weeds; and identification of the active principles. S. Afr. J. Bot. [CrossRef]
- Rial, C.; Gómez, E.; Varela, R.M.; Molinillo, J. M.; Macías, F.A. Ecological relevance of the major allelochemicals in Lycopersicon esculentum roots and exudates. J. Agri. Food Chem. 2018, 66(18), 4638–4644. [Google Scholar] [CrossRef]
- Jouini, A.; Verdeguer, M.; Pinton, S.; Araniti, F.; Palazzolo, E.; Badalucco, L.; Laudicina, V. A. Potential effects of essential oils extracted from Mediterranean aromatic plants on target weeds and soil microorganisms. Plants 2020, 9(10), 1289. [Google Scholar] [CrossRef]
- Heap, I. The international survey of herbicide resistant weeds. https://www.weedscience.org/Pages/Species.aspx 2024. (accessed: ). 30 June.
- Matzrafi, M.; Preston, C.; Brunharo, C. A. Evolutionary drivers of agricultural adaptation in Lolium spp. Pest Manag. Sci. 2021, 77(5), 2209–2218. [Google Scholar] [CrossRef]
- Neve, P.; Powles, S. High survival frequencies at low herbicide use rates in populations of Lolium rigidum result in rapid evolution of herbicide resistance. Heredity. [CrossRef]
- Amri, I.; Khammassi, M.; Ben Ayed, R. ; Khedhri; S.; Mansour, M.; Ben Kochti, O.; Pieracci, Y.; Flamini, G.; Mabrouk, Y.; Gargouri, S.; Hanana, M.; Hamrouni, L. Essential oils and biological activities of Eucalyptus falcata.; E. sideroxylon and E. citriodora growing in Tunisia. Plants. [CrossRef]
- Somala, N.; Laosinwattana, C.; Teerarak, M. Formulation process; physical stability and herbicidal activities of Cymbopogon nardus essential oil-based nanoemulsion. Sci. Rep. 2022, 12, 1–13. [Google Scholar] [CrossRef]
- Amato, G.; Caputo, L.; Francolino, R.; Martino, M.; Feo, V.; De Martino, L. Origanum heracleoticum essential oils: chemical composition; phytotoxic and Alpha amylase inhibitory activities. Plants 2023, 12, 866. [Google Scholar] [CrossRef]
- De Assis Alves, T.; Pinheiro, P.F.; Praça-Fontes, M.M.; Andrade-Vieira, L.F.; Corrêa, K.B.; de Assis Alves, T.; da Cruz, F.A.; Lacerda Júnior, V.; Ferreira, A.; Soares, T.C.B. Toxicity of thymol; carvacrol and their respective phenoxyacetic acids in Lactuca sativa and Sorghum bicolor. Ind. Crops Prod. 2018, 114, 59–67. [Google Scholar] [CrossRef]
- Bozhuyuk, A.U. Herbicidal activity and chemical composition of two essential oils on seed germinations and seedling growths of three weed species. J. Essent. Oil Bearing Plants 2020, 23, 821–831. [Google Scholar] [CrossRef]
- Khammassi, M.; Polito, F.; Kochti, O.; Kouki, H.; Souihi, M.; Khedhri, S.; Hamrouni, L.; Mabrouk, Y.; Amri, I.; De Feo, V. Investigation on chemical composition; antioxidant; antifungal and herbicidal activities of volatile constituents from Deverra tortuosa (desf.). Plants, 2556. [Google Scholar] [CrossRef]
- Xianfei, X.; Xiaoqiang, C.; Shunying, Z.; Guolin, Z. Chemical composition; and antimicrobial activity of essential oils of Chaenomeles speciosa from China. Food Chem. 2007, 100, 1312–1315. [Google Scholar] [CrossRef]
- Lins, L.; Dal Maso, S.; Foncoux, B.; Kamili, A.; Laurin, Y.; Genva, M.; Jijakli, M.H.; De Clerck, C.; Fauconnier, M.L.; Deleu, M. Insights into the relationships between herbicide activities; molecular structure and membrane interaction of cinnamon and citronella essential oils components. Int. J. Mol. Sci. 2019, 20, 4007. [Google Scholar] [CrossRef]
- Verdeguer, M.; Torres-Pagan, N.; Muñoz, M.; Jouini, A.; García-Plasencia, S.; Chinchilla, P.; Berbegal, M.; Salamone, A.; Agnello, S.; Carrubba, A.; Cabeiras-Freijanes, L. Herbicidal activity of Thymbra capitata (L.) Cav. essential oil. Molecules, 2832. [Google Scholar] [CrossRef]
- Blume, Y.B.; Krasylenko, Y.A.; Yemets, A.I. Effects of phytohormones on the cytoskeleton of the plant cell. Russ. J. Plant. Physiol. 2012, 59, 515–529. [Google Scholar] [CrossRef]
- López-González, D.; Costas-Gil, A.; Reigosa, M.J.; Araniti, F.; Sánchez-Moreiras, A.M. A natural indole alkaloid; norharmane; affects PIN expression patterns and compromises root growth in Arabidopsis thaliana. Plant. Physiol. Biochem. 2020, 151, 378–390. [Google Scholar] [CrossRef]
- Lakehal, A.; Bellini, C. Control of adventitious root formation: insights into synergistic and antagonistic hormonal interactions. Physiol. Plantarum 2019, 165, 90–100. [Google Scholar] [CrossRef]
- Lewis, D.R.; Negi, S.; Sukumar, P.; Muday, G.K. Ethylene inhibits lateral root development; increases IAA transport and expression of PIN3 and PIN7 auxin efflux carriers. Development, 3485. [Google Scholar] [CrossRef]
- Ivanchenko, M.G.; Muday, G.K.; Dubrovsky, J.G. Ethylene–auxin interactions regulate lateral root initiation and emergence in Arabidopsis thaliana. Plant J. 2008, 55(2), 335–47. [Google Scholar] [CrossRef]
- Swarup, R.; Parry, G.; Graham, N.; Allen, T.; Bennett, M. Auxin cross-talk: integration of signalling pathways to control plant development. Auxin Molecular Biology: Springer. 2002 p. 411–26.
- Alonso, J.M.; Stepanova, A.N.; Solano, R.; Wisman, E.; Ferrari, S.; Ausubel, F.M.; Ecker, J.R. Five components of the ethylene-response pathway identified in a screen for weak ethylene-insensitive mutants in Arabidopsis. Proceed. Nat. Acad. Sci. 2003, 100(5), 2992–2997. [Google Scholar] [CrossRef]
- Ellis, J.R.; Taylor, R.; Hussey, P.J. Molecular modeling indicates that two chemically distinct classes of anti-mitotic herbicide bind to the same receptor site (s). Plant Physio. 1994, 105(1), 15–18. [Google Scholar] [CrossRef]
- Boukhalfa, R.; Ruta, C.; Messgo-Moumene, S.; Calabrese, G.J.; Argentieri, M.P.; De Mastro, G. Valorization of Mediterranean species of thyme for the formulation of bio-herbicides. Agronomy 2024, 14(9), 2077. [Google Scholar] [CrossRef]
- Abd-El Gawad, A.M.; El Gendy, A.E.-N.G.; Assaeed, A.M.; Al-Rowaily, S.L.; Alharth, A.S.; Mohamed, T.A.; Nassar, M.I.; Dewir, Y.H.; Elshamy, A.I. Phytotoxic effects of plant essential oils: A systematic review and structure-activity relationship based on chemometric analyses. Plants. [CrossRef]
- Islam, A.M.; Kato-Noguchi, H. Phytotoxic activity of Ocimum tenuiflorum extracts on germination and seedling growth of different plant species. Sci. World J. 2014, 2014(1), 676242. [Google Scholar] [CrossRef] [PubMed]
- El Mahdi, J.; Tarraf, W.; Ruta, C.; Piscitelli, L.; Aly, A.; De Mastro, G. Bio-herbicidal potential of the essential oils from different Rosmarinus officinalis L. chemotypes in laboratory assays. Agronomy 2020, 10(6), 775. [Google Scholar] [CrossRef]
- Shedden, K. Generalized Linear Models; Creative Commons Attribution Share Alike 3.0 License; Department of Statistics, University of Michigan: Ann Arbor, MI, USA, 2015; 35p. [Google Scholar]
Figure 1.
Effect of the different treatments and doses on seed germination of L. perenne (a) and A. retroflexus (b) after 15 days exposure to a 500- 2000 μl/100 mL range of concentrations of the four tested EOs. T1: T. algeriensis, T2: T. ciliatus subsp. coloratus, T3: T. vulgaris cultivar Varico 3, T4: T. vulgaris ecotype Fasano. Data are means of three replicates. At each dose, column with different letters are significantly different (p ≤ 0.05, Tukey’s test).
Figure 1.
Effect of the different treatments and doses on seed germination of L. perenne (a) and A. retroflexus (b) after 15 days exposure to a 500- 2000 μl/100 mL range of concentrations of the four tested EOs. T1: T. algeriensis, T2: T. ciliatus subsp. coloratus, T3: T. vulgaris cultivar Varico 3, T4: T. vulgaris ecotype Fasano. Data are means of three replicates. At each dose, column with different letters are significantly different (p ≤ 0.05, Tukey’s test).
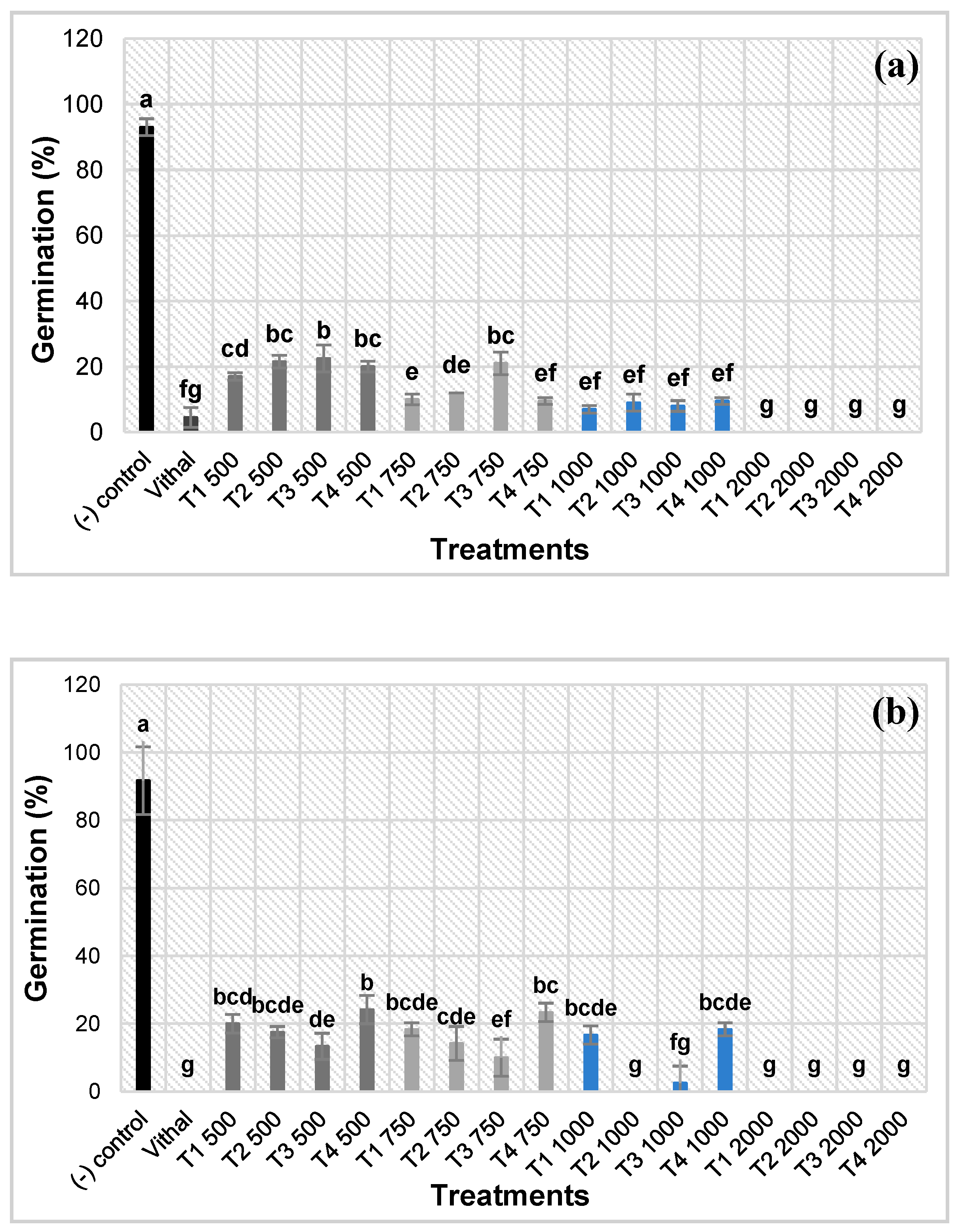
Figure 2.
Effect of the treatments on target weed species plantlets. (a): L. perenne seedling treated with EOs at 1000 µl, (b): A. retroflexus seedling treated with EOs at 1000 µl, (c): L. perenne seedling treated with EOs at 2000 µl.
Figure 2.
Effect of the treatments on target weed species plantlets. (a): L. perenne seedling treated with EOs at 1000 µl, (b): A. retroflexus seedling treated with EOs at 1000 µl, (c): L. perenne seedling treated with EOs at 2000 µl.

Figure 3.
Number of surviving (a) and fresh biomass (b) of controls and treated A. retroflexus plants, 24 hours after exposure to the different treatments and doses (500- 2000 μl/100 mL range of concentrations) of the four tested EOs. T1: T. algeriensis, T2: T. ciliatus subsp. coloratus, T3: T. vulgaris cultivar Varico 3, T4: T. vulgaris ecotype Fasano. Data are means of three replicates. At each dose, column with different letters are significantly different (p ≤ 0.05, Tukey’s test).
Figure 3.
Number of surviving (a) and fresh biomass (b) of controls and treated A. retroflexus plants, 24 hours after exposure to the different treatments and doses (500- 2000 μl/100 mL range of concentrations) of the four tested EOs. T1: T. algeriensis, T2: T. ciliatus subsp. coloratus, T3: T. vulgaris cultivar Varico 3, T4: T. vulgaris ecotype Fasano. Data are means of three replicates. At each dose, column with different letters are significantly different (p ≤ 0.05, Tukey’s test).
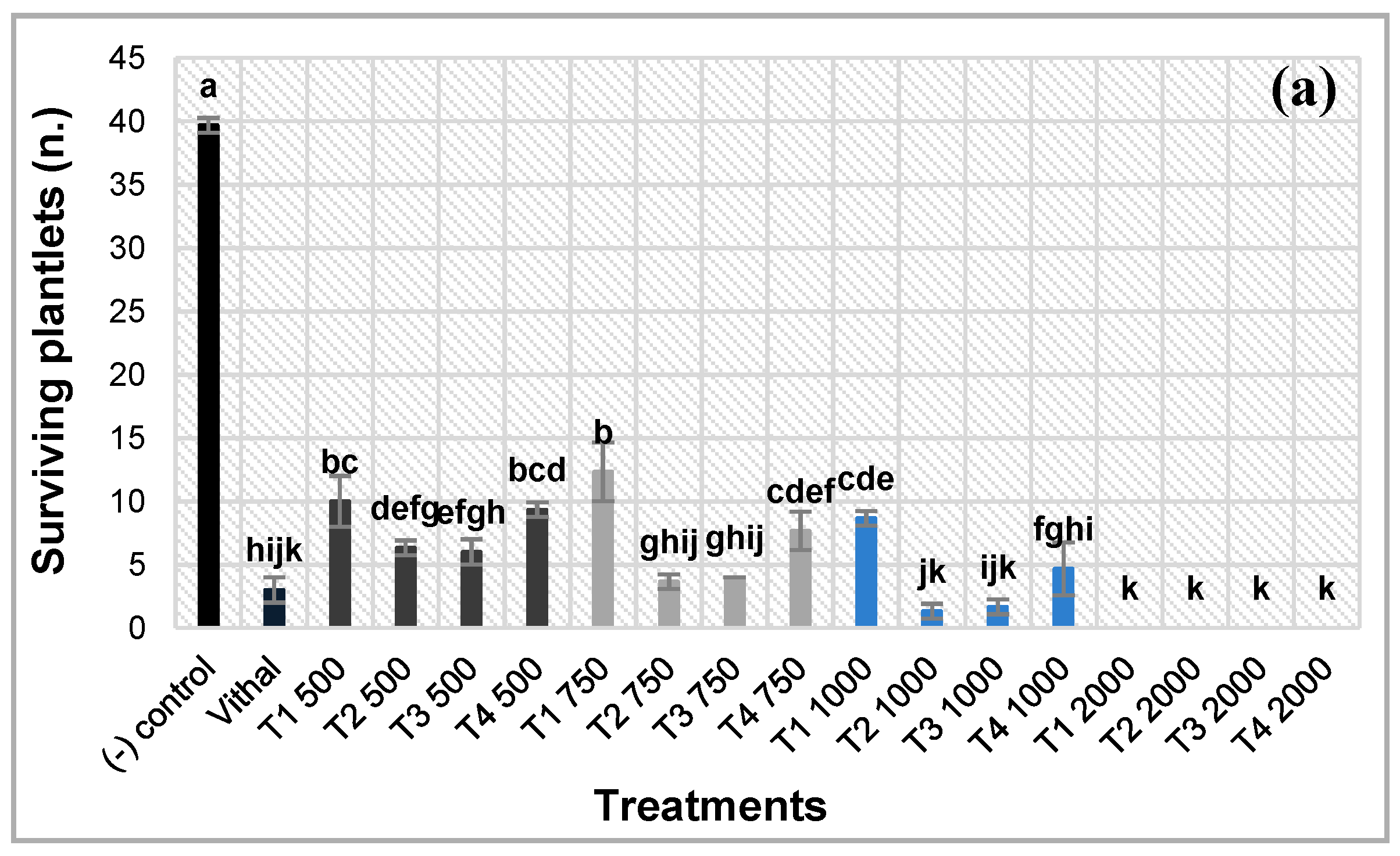
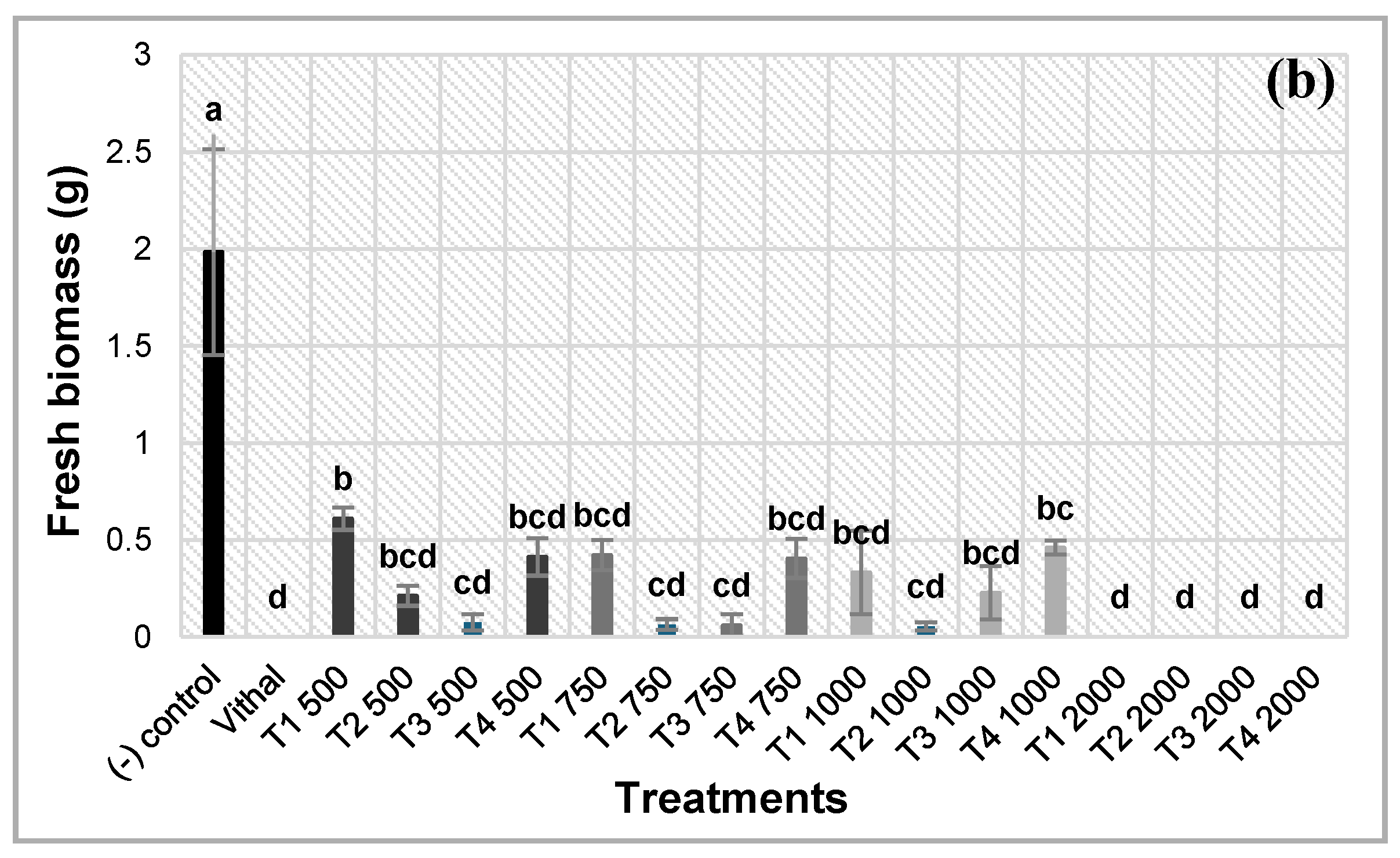
Figure 4.
Images of Arabidopsis thaliana seedlings control and treated with thyme EOs. (a, b): control; (c, d): short roots and condensate root hair belonging to seedlings treated with T1; (e, f): absence of lateral and root hair of seedlings treated with T3.
Figure 4.
Images of Arabidopsis thaliana seedlings control and treated with thyme EOs. (a, b): control; (c, d): short roots and condensate root hair belonging to seedlings treated with T1; (e, f): absence of lateral and root hair of seedlings treated with T3.
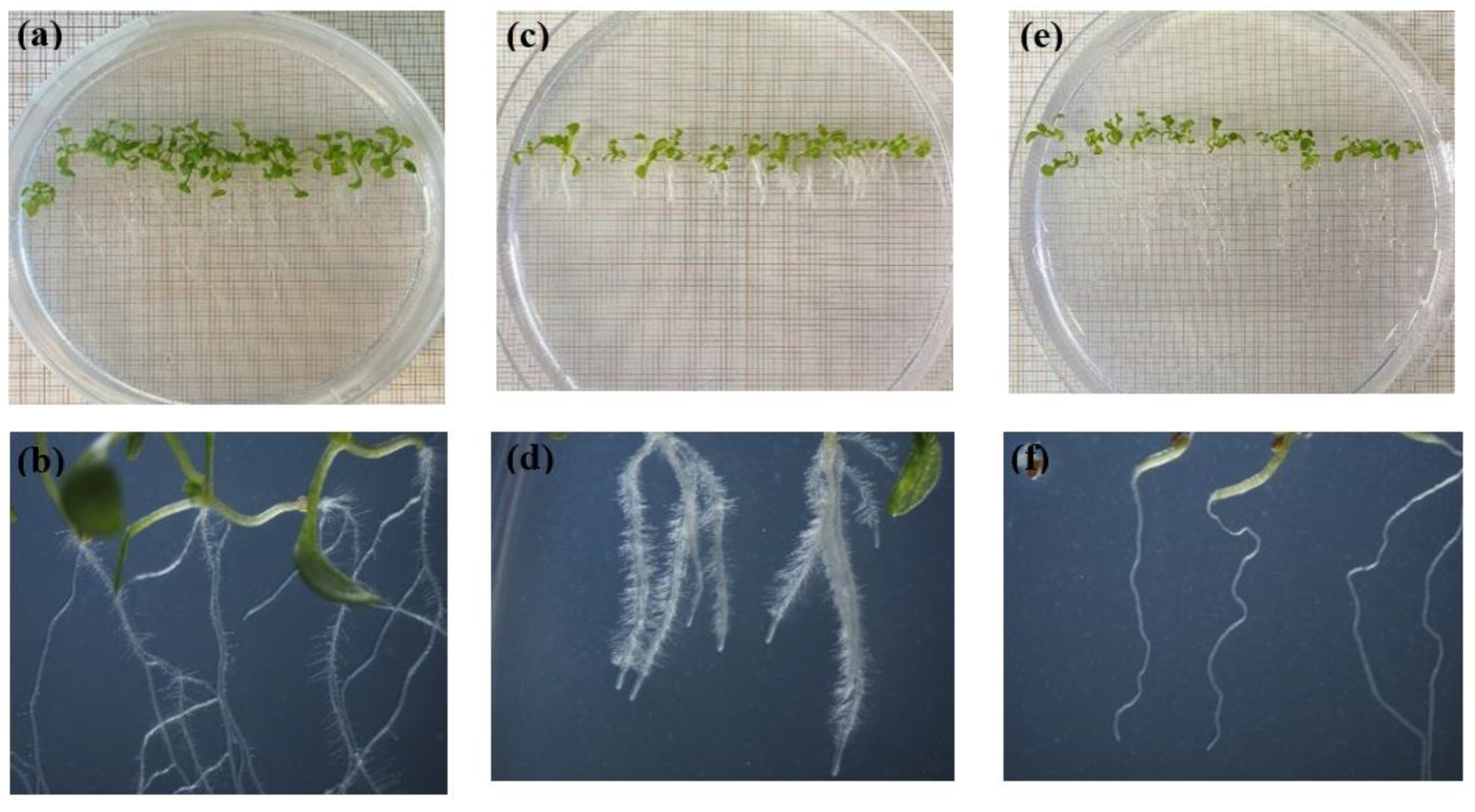
Figure 5.
Effect of thyme EOs on the loss of membrane integrity of apical root longitudinal sections of A. thaliana stained with Evans Blue reagent visualized under light microscopy. (a): control, (b): EO-treated roots with T1; (c): EO-treated roots with T2. (d): EO-treated roots with T3 and (e): EO-treated roots with T4. Images taken at 10 and 20X magnification.
Figure 5.
Effect of thyme EOs on the loss of membrane integrity of apical root longitudinal sections of A. thaliana stained with Evans Blue reagent visualized under light microscopy. (a): control, (b): EO-treated roots with T1; (c): EO-treated roots with T2. (d): EO-treated roots with T3 and (e): EO-treated roots with T4. Images taken at 10 and 20X magnification.
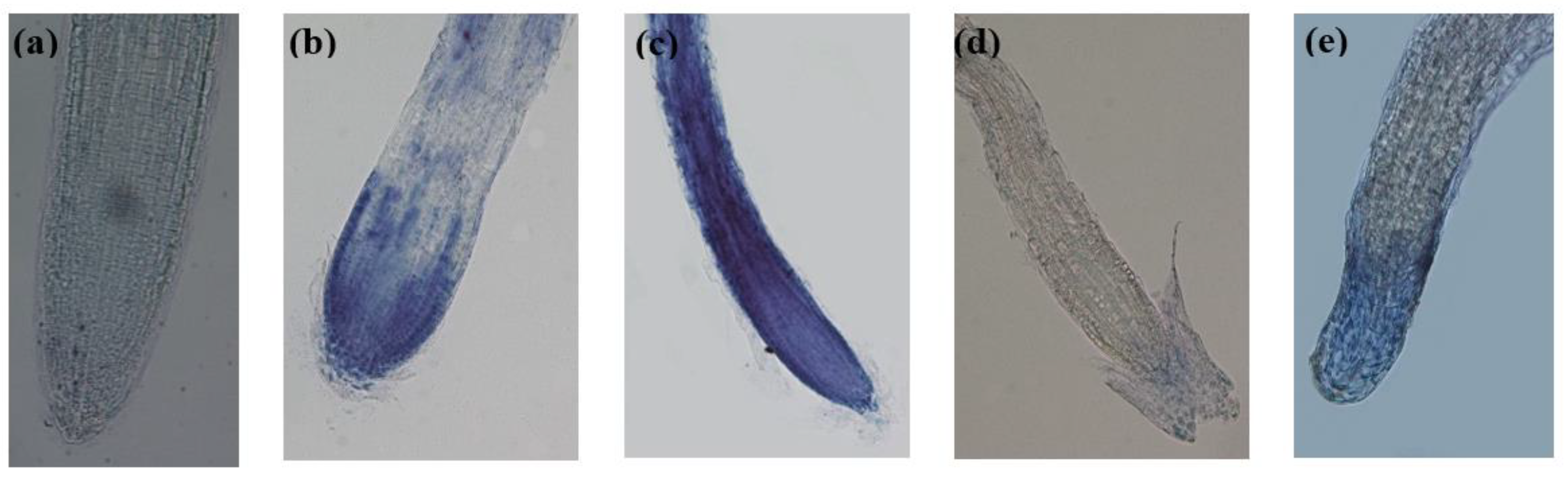
Figure 6.
Histochemical determination of lipid peroxidation in A. thaliana roots under the effect of thyme EOs by Schiff’s reagent (longitudinal sections of root apices). (a): control, (b): EO-treated roots with T1; (c): EO-treated roots with T2. (d): EO-treated roots with T3 and (e): EO-treated roots with T4. Images taken at 10 and 20X magnification.
Figure 6.
Histochemical determination of lipid peroxidation in A. thaliana roots under the effect of thyme EOs by Schiff’s reagent (longitudinal sections of root apices). (a): control, (b): EO-treated roots with T1; (c): EO-treated roots with T2. (d): EO-treated roots with T3 and (e): EO-treated roots with T4. Images taken at 10 and 20X magnification.
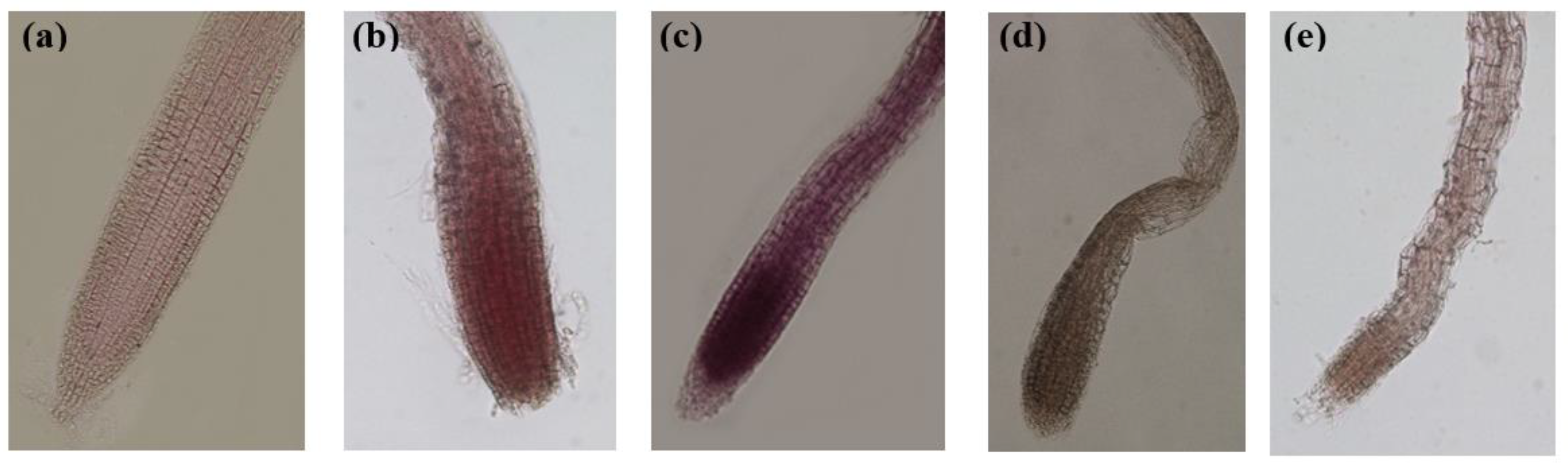

Table 1.
Chemical composition of the studied Thymus EOs.
| N° | Compound | KI1 | KI2 | T1 | T2 | T3 | T4 |
|---|---|---|---|---|---|---|---|
| 01 | santolina triene | 908 | 910 | 0.36 | -- | -- | -- |
| 02 | trycyclene | 926 | 924 | 0.47 | -- | -- | -- |
| 03 | α-thujene | 926 | 924 | -- | 0.20 | 0.48 | 0.66 |
| 04 | α-pinene | 936 | 940 | 19.73 | 0.21 | 1.03 | 1.15 |
| 05 | camphene | 954 | 950 | 8.64 | 0.29 | 0.88 | 0.93 |
| 06 | verbenene | 967 | 968 | 0.34 | -- | 0.05 | -- |
| 07 | sabinene | 975 | 975 | 0.87 | 0.05 | -- | -- |
| 08 | β-pinene | 979 | 980 | 4.37 | 0.14 | 0.21 | -- |
| 09 | myrcene | 991 | 992 | 2.77 | 0.36 | 0.52 | 1.35 |
| 10 | α-terpinene | 1017 | 1012 | 0.25 | 0.05 | 0.13 | 0.24 |
| 11 | p-cymene | 1022 | 1021 | 0.30 | 0.13 | 35.63 | 23.85 |
| 12 | limonene | 1029 | 1026 | 1.95 | 1.36 | 0.79 | 1.33 |
| 13 | 1,8 cineole | 1031 | 1028 | 2.99 | 0.16 | 3.53 | 3.30 |
| 14 | β(E)- ocymene | 1050 | 1047 | 1.17 | 0.07 | -- | -- |
| 15 | γ-terpinene | 1059 | 1058 | 0.54 | 0.98 | 2.35 | 10.36 |
| 16 | cis-sabinene hydrate | 1070 | 1067 | 0.45 | 0.10 | -- | -- |
| 17 | terpinolene | 1088 | 1088 | 0.47 | -- | 0.48 | 0.17 |
| 18 | linalool | 1096 | 1103 | 3.09 | 93.06 | 2.57 | 2.54 |
| 19 | 1-octen-3γlacetate | 1110 | 1116 | 0.43 | -- | -- | -- |
| 20 | α-campholenal | 1125 | 1125 | 0.99 | -- | -- | -- |
| 21 | camphor | 1143 | 1142 | 4.32 | 0.38 | 1.66 | 1.20 |
| 22 | trans-verbenol | 1144 | 1145 | 2.61 | -- | -- | -- |
| 23 | pinocarvone | 1162 | 1158 | 0.56 | -- | -- | -- |
| 24 | borneol | 1165 | 1166 | 11.31 | -- | 1.47 | 1.22 |
| 25 | ρ-mentha-1,5 dien-8-ol | 1170 | 1174 | 1.72 | -- | 1.76 | 1.65 |
| 26 | terpin-4-ol | 1177 | 1182 | 0.76 | -- | -- | -- |
| 27 | α-terpineol | 1189 | 1187 | 0.96 | -- | -- | -- |
| 28 | myrtenal | 1193 | 1190 | 0.90 | -- | -- | -- |
| 29 | verbenone | 1204 | 1215 | 0.73 | -- | -- | -- |
| 30 | isobonyl formate | 1233 | 1243 | 0.33 | -- | -- | -- |
| 31 | thymol methyl ester | 1235 | 1234 | -- | -- | 1.97 | 1.48 |
| 32 | linalool acetate | 1257 | 1262 | 3.96 | -- | -- | -- |
| 33 | isobornyl acetate | 1285 | 1289 | 5.28 | -- | -- | -- |
| 34 | thymol | 1290 | 1293 | -- | 0.55 | 20.35 | 21.77 |
| 35 | carvacrol | 1298 | 1300 | -- | -- | 11,76 | 18.15 |
| 36 | trans-carvyl acetate | 1328 | 1337 | 0.11 | -- | -- | -- |
| 37 | α-terpenyl acetate | 1350 | 1353 | 13.21 | -- | -- | -- |
| 38 | α-copaene | 1376 | 1370 | 0.49 | -- | -- | -- |
| 39 | β-bourbonene | 1384 | 1376 | 0.22 | -- | -- | -- |
| 40 | β-cedrene | 1418 | 1404 | 0.51 | 1.98 | 7.69 | 6.56 |
| 41 | germacrene | 1480 | 1475 | 1.58 | -- | -- | -- |
| 42 | Δ-cadinene | 1524 | 1530 | 0.26 | -- | -- | -- |
| 43 | caryophyllene oxide | 1581 | 1573 | -- | -- | 5.57 | 2.09 |
| Monoterpene hydrocarbons | 42.23 | 3.77 | 42.85 | 40.04 | |||
| Oxygen-containing monoterpenes | 54.71 | 94.18 | 43.89 | 51.31 | |||
| Sesquiterpene hydrocarbons | 3.06 | 1.98 | 7.69 | 6.56 | |||
| Oxygen-containing sesquiterpenes | - | - | 5.57 | 2.09 | |||
| Others | - | - | - | - | |||
| KI: Kovats index; 1: literature; 2: calculated | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



