
Submitted:
01 October 2024
Posted:
01 October 2024
You are already at the latest version
Abstract
Cardiovascular disease (CVD) is the primary cause of mortality globally with a multifactorial etiology that involves epigenetics. Chromobox 3 (CBX3), the major isoform of heterochromatin protein 1, is involved in intricate epigenetic mechanisms affecting congestive heart failure. In patients with CVD affected by lung cancer risk, CBX3 exerts a sophisticated mechanism of action, suppressing the proliferation, migration, and formation of neointima in vascular smooth muscle cells (VSMCs) by affecting the Notch3 pathway, indicating a potential protective function against vascular remodeling and atherosclerosis. However, the broader impact of CBX3 on endothelial function, as well as its effects on monocyte/macrophage and lymphocyte infiltration and function within the arterial wall, remain poorly understood. Since very little is known so far, more definite research would be needed to reveal the fine mechanisms of CBX3 action, along with its relationship in molecular processes and prospects as a biomarker. Specifically, CBX3 biological features could be examined to gain a greater insight into CVD risks. This review outlines the role of CBX3 in mechanisms associated with CVD and feasibility for optimizing pre-existing therapy and developing new therapeutic strategies based on personalized medi-cine.
Keywords:
CBX3
; cardiovascular disease
; epigenetics
; vascular smooth muscle cells
; vascular remodeling
; inflammation
; pathway
; chromatin
; treatment(s)
1. Introduction
Cardiovascular diseases (CVDs) affect both developed and developing countries where the nature of heart disease is the primary cause of death among adult individuals, launching the cardiovascular health challenge universally without geographical, social or economic limitations (Figure 1). CVD accounts for approximately 610,000 deaths each year in the United States, and this represents an impressive example suggesting the enormity of the problem within a single country [1]. Furthermore, the global burden of CVDs is worsened by their major role in causing not only mortality but also significant morbidity, disability and loss of productivity, making them a major health care emergency around the world [2]. For all these reasons there is an urgent need for “holistic” strategies that address the multiple challenges posed by CVD, from prevention and management to mitigation of its broader socio-economic impacts. The number of deaths due to congestive heart failure is increasing quite notable by the day, with statistics from the World Health Organization indicating an increase from 12.1 million in 1990 to an alarming 20.5 million in 2021 [3]. Such increase serves as a valid indicator relating to the mounting burden that CVD would impose on global healthcare systems and economies, with annual mortality rates expected to reach 23.6 million by 2030 [4]. This information is a bitter reminder of the continuous issue of CVD, not only as the top killer worldwide but also because of the large loss in health and life years it causes [5,6]. The ongoing increase of mortality from CVD, even after the implementation of knowledge and available treatments, it is worthy to be further investigated from different perspectives that determine its high incidence rates and the contributions of epigenetics marks in the development and progression of these conditions [7].
A multicausal approach to the pathogenesis of heart failure also underscores its intricate development and the multiple risk factors involved [8]. Infectious agents show a direct effect on heart health, and may indirectly cause serious cardiovascular complications [2,9]. Conditions such as ischemic heart disease and hypertension are not isolated risk factors but are part of a broader spectrum of problems that contribute to the overall risk of developing CVD. Hypertension greatly increases the risk of chronic heart failure [10] , with hypertensive individuals facing up to three times the risk of their normotensive counterparts [11]. These insights collectively outline an etiology of CVD as a complex interplay of infectious, genetic, and lifestyle factors, each contributing to increased risk of developing various forms of heart disease. The orchestration between genetics and epigenetics in influencing CVD has attracted considerable interest, particularly the role of chromobox CBX proteins in these mechanisms. Specifically, CBX3, a chromatin remodeling protein, acts as a crucial player in CVD through a mechanism based on epigenetic coordination of signaling pathways. CBX3 plays a role in suppressing the proliferation, migration, and neointima formation of vascular smooth muscle cells (VSMCs), primarily by affecting the Notch3 pathway, suggesting a potential protective role against vascular remodeling and atherosclerosis [12] Further, CBX3 is implicated in the regulation of angiogenesis and may have effects on restenosis after angioplasty [13]. However, the current scientific literature does not provide a full picture to address the role of CBX3 in these diseases, indicating that more advanced and accurate studies should be developed to obtain a deeper insight into the function of this protein. This review aims to fully discussing the role of CBX3 as potential therapeutic targets in CVD, where vascular remodeling up to epigenetic regulations are under consideration, along with related pathways.
2. The Multifaceted Role and Expression of CBX3 in CVD
CBX3 influences gene expression through epigenetic mechanisms [14] across multiple aspects, involving various subfragments and their distinct activities in different cellular contexts. Notably, a study showed that CBX3 subfragments exhibit anti-silencing activities in pluripotent versus differentiated cells [15] ; this argues that the differentiation stage of the cell is important for determining efficacy in gene regulation by CBX3 and viceversa [16]. This is very clear from the loss of anti-silencing activity in differentiated cells by subfragments such as CBX3 (1-339) and CBX3 (340-508) [15], which indicates that these have become nonfunctional upon cellular differentiation. This loss of function is probably related to the large-scale chromatin remodeling processes that occur as cells leave pluripotency, which could impede retroviral transgene expression. In addition, ChIP-seq analyses have revealed the co-localization of CBX3 with H3K9me3 in the promoter regions of important regulatory genes, such as SMAD-specific E3 ubiquitin protein ligases 1 and 2 (SMURF1 and SMURF2), in addition to reiterating the role of CBX3 in chromatin dynamics and gene silencing [17]. Aside from its direct effects on VSMCs, CBX3 plays a role in the broader epigenetic changes related to cardiovascular disease [18,19]. Epigenetic modifications, such as DNA methylation, histone acetylation, and miRNA expression, have been a major cause of the onset and development of CVDs [20]. The protein encoded by the CBX3 gene binds to DNA forming constitutive heterochromatin which is critical to stabilize chromosomes and gene expression regulation as well as other nuclear processes[21,22]. CBX3 can influence these epigenetic mechanisms, thus modulating the expression of genes important for cardiovascular health as SLC9A1, SLC1A5, TNRC6C [20]. The protein is engaged in epigenetic regulation through interactions with methylated histone marks including H3K9, H1K26, G9aK185 peptides. This is indicative of a mechanism where CBX3 acts on chromatin structure and gene expression [23]. Inhibition of CBX3 enhances cellular reprogramming, which means its contribution to genome-wide chromatin dynamics is significant [24]. Further, the binding of CBX3 to DNA and its association with heterochromatin suggest a potential involvement in regulating gene transcription and chromatin structure, which are critical factors in cardiovascular health [25]. Targeting CBX3 in CD8+ T cells results in enhanced transcription initiation and chromatin remodeling. This underscores potential interactions between CBX3 and other chromatin modifiers in influencing CVD progression and outcome [26]. For example, the epigenetic mechanism mediated by CBX3-PRC2 interaction is involved in the regulation of endothelial cell function, angiogenesis, and vascular inflammation, all of which are crucial processes in the pathogenesis of CVD [27]. CBX3 is reported to modulate the expression of several miRNAs implicated in the cardiovascular function and disease [28,29]. CBX3, for instance, was demonstrated to repress the expression of miR-21, a pro-fibrotic and pro-inflammatory miRNA, in cardiac fibroblasts, therefore diminishing cardiac fibrosis and remodeling in response to stress [30]. Likewise, CBX3 was identified as a regulator for miR-126, an endothelial cell-specific miRNA which is highly important for maintaining vascular integrity and induction of angiogenesis factors [31,32]. The binding of CBX3 on specific target genes in neural progenitor cells (NPC) results in the upregulation of cardiac lineage development crucial genes including TNNT3, TBX20, TBX3, HAND1 as well as platelet derived growth factor receptor alpha [16]. Knocking down CBX3 interestingly showed a different effect on NPC differentiation causing the down-regulation of neural genes, such us SOX1, while simultaneously up-regulating mesodermal genes such as WNT4, particularly those associated with circulatory system development [16]. The association of reduced CDK8 levels and enrichment of genes related to nervous system development following to CBX3 silencing reflects an intricate regulation newwork among CBX3 and pathways regulating gene expression beyond cardiovascular health. Loss of CBX3 also results in reduced recruitment of CDK8 on genes related to the development of the cardiovascular system, indicating a direct effect in this regulation [33]. Intriguingly, epigenetic modifications under the control of CBX3 could also alter these circulatory disease-related genes implicated in lipid metabolism, inflammation, and endothelial function which are major pathogenic factors for CVD. Examples demonstrating the impact of lipid-associated CpGs on metabolic genes, including carnitine-palmitoyl transferase and ATP Binding Cassette Subfamily G Member 1 (ABCG1), from studies like the Framingham Heart Study, illustrate the complexity portrayed between epigenetic modifications with CVD [34]. One of the key genetic factors that would drive the expression of CBX3 in CVD would be genotype effect on DNA methylation at specific loci, such as the ABCG1 locus [35,36]. A notable example is actually related to changes in methylation at cg27243685 within the ABCG1 locus, being linked to higher levels of triglycerides and higher risk for new-onset coronary heart disease, as well as having been associated previously with a history of prevalent myocardial infarction [37,38]. Additionally, minor allele presence in the intronic variant rs4148086 is related to increased methylation at cg27243685, leading to ABCG1 low expression in blood [39] . The co-occurrence of these genotypic and epigenetic variants reveals the complexity of gene regulation in CVD, where small genomic regions control any gene modulation through changes in DNA methylation. The full understanding of such tight-regulated mechanisms is necessary for targeted intervention able to influence risk and progression of CVD. Interesetingly, GATA4 is one of the transcription factors that controls cardiac genes and can be influenced by chromatin-modifying proteins [40]. When considering the altered CBX3 activity and its contribution to specific cardiovascular conditions, it is important to consider ApoC3 expression and downstream effects on cardiovascular health. ApoC3 overexpression is often associated with progressive atherogenesis. Atherogenesis is a condition of plaque accumulation within arterial walls, resulting with severe clinical complications including restenosis of the artery post-surgery [41]. This suggests that modified CBX3 activity, which might be affect ApoC3 expression, could be inherently related to atherosclerosis development. Induction of inflammation represent one of the major causes through which CBX3 contributes to CVD , and it is a crucial step in the development of atherosclerosi [42]. Exacerbating the risk of atherosclerosis, thus is an independent effect of traditional cardiovascular risk factors like hypertension and hyperlipidemia [43]. Besides, CBX3 is involved in metabolic regulation indicating its connection with type 2 diabetes mellitus (DM), which is a major risk factor for CVD. Studies on the genetics level have shown that individuals with type 2 DM are those likely to have overlapping genetic predispositions with subjects that are prone to cardiovascular conditions, as well as revealing the phenotypic interconnected nature of these diseases [44]. Future studies investigating CBX3 and its applications in the onset and management of CVD will be needed to further development personal treatment approaches and diagnostic tools that could help to improve cardiovascular profiles. Table 1 lists the epigenetic modifications regulated by CBX3 and their association with CVD.
3. CBX3 Impacts Proliferation, Migration, and Formation of Neointima Via Notch3 Pathway
The Notch3 pathway can be considered as a probable area of research in CVDs, opening new prospects for therapeutic interventions [45]. Since explorations on targeting components of the Notch pathway for cardiovascular conditions are preliminary, the opportunity for new treatment modalities in this area is immeasurable. The collaboration between the specialist in CVD and researchers is important to further determine the complexity of the Notch pathway and its impact on the management of the disease. The pivotal role of the Notch pathway in embryonic development and in the maintenance of the cardiovascular system underscores its importance in the pathophysiology of different cardiac diseases [46]. The fact that the Notch pathway is involved in common pediatric and adult cardiac diseases stresses its importance as a therapeutic target for a broad spectrum of CVDs [47]. Moreover, the Notch pathway control of vascular cell growth and survival, maintenance of endothelial cell precursor recruitment for endothelial repair and the transient reactivation in cardiomyocytes after myocardial infarction alludes to its multifaceted contribution to heart health and disease advancement [48]. These results indicate that modulation of the Notch pathway could enhance cardiomyocyte survival, promote angiogenesis, and sustain the regenerative potential of the injured myocardium, providing a novel means of augmenting myocardial adaptation to stress and salvaging myocardium damage. The role of CBX3 in the regulation of VSMC behavior affects proliferation, migration, and neointimal formation, which are fundamental to the pathophysiology of vascular injury and disease. CBX3 is mainly associate with gene expression silencing, however it can function also as a regulator supporting transcriptional activation and RNA processing modulation, reflecting its epigenetic flexibility. This versatility is important in VSMC behavior i.e especially in response to vascular injury where the CBX3 interaction with the Notch3 pathway reveals a specific mechanism of action. By acting on Notch3-dependent VSMC proliferation, migration, and anti-apoptotic programs, CBX3 directly governs the evolution of vascular injury (Figure 2) and resulting adverse arterial remodeling, which underlies most of heart disease vast morbidity and mortality [16].
CBX3 further promotes neointimal inhibition through transcriptional repression of Notch3 signaling in VSMCs, therefore modulating key VSMC phenotypes as proliferation, migration, and apoptosis as well as expression of contractile genes [49]. This is based on binding to the promoter region of smooth muscle myosin heavy chain, calponin, and smooth muscle α-actin 2 and facilitating diphosphorylation diaphanous homolog 1/activation serum response factor (SRF) or myocardin that leads to an increased expression of contractile proteins [50]. In addition to this, a reduced expression of collagen genes COL1A1 and COL4A1 drives VSMCs into a more contractile differentiated phenotype[51]. It was detected that CBX3 expression was markedly enhanced in the neointima of rat-injured carotid arteries, interestingly, its silencing decreased neointimal hyperplasia and vascular remodeling in a mouse model of carotid artery injury, indicating that CBX3 might be a promising therapeutic target for restenosis. Moreover, the effect of CBX3 on VSMC collagen production and apoptosis could influence VSMCs vital aspects during vascular remodeling [52]. The Notch signaling pathway plays a critical role in the regulation of vascular morphogenesis throughout development and maintenance of vascular cell homeostasis. It is also involved in vascular repair following injury, relying to some degree on the regulation of the VSMC phenotype [53]. Activation of the Notch3 signaling pathway was found to induce the transcription of genes associated with cell cycle regulation, anti-apoptosis, and stemness-related characteristics [54]. Another study, by collecting tissue and plasma samples from healthy adults and patients with atherosclerosis, showed that tRNA-derived stress-induced RNA from glycine tRNA with the anticodon GCC (tiRNA-Gly-GCC) is upregulated in synthetic VSMCs and atherosclerotic tissues [55,56]. The level of tiRNA-Gly-GCC, a mediator of human aortic smooth muscle cell phenotypic switching, is elevated when human aortic smooth muscle cells (SMCs) undergo a phenotypic transition, both in vivo and in vitro. Hindering the activity of tiRNA-Gly-GCC by targeting CBX3 was shown to promote retention of the myogenic phenotype, decrease the proliferation and migration rate of human aortic SMCs, and decrease neointima formation following vascular injury [57]. CBX3 and SRF are considered important in the differentiation of SMCs from stem cells and functional pathways of cardiovascular development [58]. As regards the chromo shadow domain of CBX3, a specific set of four amino acids (165 to 168) that are responsible for the interaction between CBX3 and Dia-1, is involved in the differentiation of SMCs induced by CBX3 [58]. CBX3 was also identified to be involved in the expression of SMC-specific genes by regulating the recruitment of SRF to these gene promoters in a process heavily dependent on interactions between CBX3, Dia-1, and SRF [59]. The molecual details of how CBX3 affects cell proliferation and migration point the fact that its regulatory effects are quite sophisticated and require more detailed explorations toward therapeutic interventions targeting CBX3 and the Notch3 pathway.
4. CBX3 is Associated with Lung Cancer Risk in CVD Patients
The interplay CVD and lung cancer is intricate. Shared risk factors, such as smoking, hypertension, diabetes, old age, and obesity, create a common soil for the progression of both diseases [60,61]. The involvement of these risk factors at the molecular level is confirmed by the effect of CVD on the regulation of genes associated with cancer. As evidenced, being overexpressed in different types of cancers, CBX3 is markedly related with bad prognosis. CBX3, encoding HP1γ, is centrally involved in cancer progression and patient outcomes; thus, it can be a putative prognostic marker in different tumor types. Further, the common risk factors and molecular pathways support a bidirectional relationship among the regulators of the cardiopulmonary axis (see Figure 3) where one condition would worsen the other present one [62,63]. Considering these interconnections, there arises an urgent need for interrelated management strategies dealing with both CVD and lung cancer at the same time for the improvement of patient outcomes. This approach is related not only to clinical interventions but to a more profound knowledge of molecular mechanisms that underlie the development of these diseases, for example, the involvement of CBX3. One of the common pathways by which CBX3 may affect both conditions involves its role in inflammation and cellular senescence [64]. Inflammation is a well-established factor in the pathogenesis of both lung cancer and CVD since it can promote cellular mutations (and atherosclerosis). Moreover, additional evidence has demonstrated that individuals with self-reported CVD (including myocardial infarction, angina pectoris, or stroke) have a higher occurrence of lung cancer most notably seen in those who are former or current smokers [65]. This indicates that the inflammatory milieu related to CVD might be favorable for the development of lung cancer.
In addition, epidemiological studies developed over a more extended period from Nordic and other European countries have produced evidence that the history of CVD increases the risk of subsequent lung cancer long after the first cancer events have transpired[66]. The association indicates the potential shared channels in view, for example, of both diseases manifesting through oxidative stress and endothelial dysfunction among others. Comprehension of these interlinking pathways is important in terms of novel integrated therapeutic strategies where both conditions can be addressed concurrently for better patient outcomes[67,68]. Increases in proinflammatory factors, such as high-sensitivity C-reactive protein and interleukin-6, are independent predictors of cardiovascular risk underscoring the importance of inflammatory pathways in the progression of CVDs[69,70]. Furthermore, the CRP/IL-6/IL-1 inflammatory axis has been recognized as a therapeutic pathway with anti-inflammatory interventions. For example, the canakinumab has shown promising result in reducing cardiovascular events and it potentially might have oncologic benefits[71,72]. This common inflammatory mechanism underlines the need for combined treatment approaches that would address cardiac and oncologic disease at the same time. CBX3 binding is especially noted to be associated with increased acetylation of histones H3 and H4, which are important markers of active transcription and have been tied to the induction of cellular senescence [24]. This epigenetic modification can bring about a change in patterns of gene expression resulting in phenotypes that favor cardiovascular cell aging. Moreover, CBX3 role in cellular proliferation and cycle progression has been well described in cancer cells; here, its overexpression leads to support for growth as part of uncontrolled proliferation (bad prognosis). The same mechanisms by which CBX3 drives cancer proliferation could also contribute toward loss of regulation regarding cell cycle checkpoints in cardiovascular cells at an early age, hence senescence [73]. Besides, the silencing of CBX3 has been demonstrated to elicit cell cycle arrest at G0/G1 phase and higher early apoptosis underpinning its prospective contribution to orchestrating cellular homeostasis and warding off senescence [74]. This indicates that CBX3 might have a role in coordinating cellular stress plus maintenance of RNA integrity, which are key prerequisites for cellular longevity. Furter, alterations of metabolism in malignant cells as well as those in cardiac cells under stress make a considerable contribution toward their survival and progression for both diseases [75,76]. These metabolic derangements provide a basis on which to construct, de novo, comprehensive therapeutic strategies designed to mitigate the impact of both pathologies. In addition, altered angiogenesis, a hallmark of both failing hearts and tumors, represents another shared pathway (in this case) that can be targeted for therapeutic intervention [77,78]. Focusing on these shared hallmarks including inflammation, metabolic stress and angiogenesis, applied research may be more effective by developing multi-faceted treatment strategies that consider interactions between CVD and cancer as a comorbidity. Optimizing cardiovascular health and, at the same time, combating systemic inflammation using targeted anti-inflammatory and metabolic therapy, carries great promise in terms of improving the prognosis of oncologic patients with concurrent diseases that are drivers of mortality. Although its direct involvement in suppressing lung cancer proliferation and reducing CVD risk via the c-Met/AKT/mTOR pathway, known for its role in glioma, has not been explicitly established; the implication of CBX3 in these critical signaling pathways suggests a potential mechanism through which CBX3 may exert its influence in the context of lung cancer and associated CVD risk[79]. This multifaceted nature of CBX3 function underscores its relevance to the pathophysiology of lung cancer and putative impact on cardiovascular health, thus deserving further exploration for therapeutic potential. Tobacco smoking is a major risk factor for both heart disease and lung cancer, underlining the role lifestyle choices play alongside genetic predispositions in disease development. [80]. This not only holds much promise in increasing survival rates among patients with more advanced stages of lung cancer but also holds an opportunity to mitigate risk related to cardiopulmonary morbidity, a very important issue given the high mortality associated with heart disease [81].
5. Dysregulation of CBX3 in the Immune Response and Its Impact in Heart Disease
Recent studies indicated that the effect of CBX3 on immune cell function in different tumor types is pleiotropic, playing a considerable role in regulating immune cell infiltration and behavior [82,83,84]. The expression of CBX3 is remarkably correlated to immune cell infiltration levels for most immune cells, indicating CBX3 as an important regulator that modulates the immune microenvironment in tumors. In particular, the expression of CBX3 has been found to correlate positively with T helper, T memory, Tγδ and Th2 cells in the tumors, suggesting that CBX3, maybe also in another setting, supports either activity or recruitment of these specific subsets of immune cells [85] Evidence relating to CBX3 involvement in the development of heart disease, particularly through its effects on monocyte/macrophage and lymphocyte accumulation and activity within the arterial wall, is starting to become a major area of interest [86]. CBX3 has been identified as a regulator altering the function of CD8+ T effector memory cells through induction of a sustained increase of LEF-1 and IL-21R [87,88]. These results indicate that the expression of CBX3 is a key regulator of immune cell dynamics and may offer novel treatment strategies for heart disease. All this suggests that CBX3 could play a very complex role in modulating local immune responses and structural integrity within blood vessels that impacts on endothelial function driving CVD progression [89]. Interestingly, investigation of the correlative transcription factors and miRNAs for genes bound by CBX3 will provide a more detailed insight into these mechanisms [90,91]. This interaction network reaches the immune system, where CBX3 influences immune-related genes, including immune checkpoints and chemokines, and thus it is related to modulation of immune response [92]. These interactions emphasize the role of CBX3 in cellular processes as well as disease states in CVD and the need for more research toward potential therapeutic interventions targeting CBX3-related pathways. The interaction of CBX3 with genes related to immune responses, including those that participate in activation and inhibition of immunity, indicates that it might regulate inflammation, one of the important pathways for atherogenesis [93]. Inflammation is a key player in the pathophysiology of atherosclerosis, as it contributes to the destabilization of plaques and subsequent cardiovascular events. Based on these preliminary studies, CBX3 could be a suitable therapeutic target of atherosclerosis in high-risk individuals, including cancer patients treated with immune checkpoint inhibitors (ICIs)[94,95]. Especially in combinatorial therapy regimens, ICIs are associated with a 3-fold higher risk for atherosclerotic cardiovascular events, including myocardial infarction, coronary revascularization, and ischemic stroke, and with a >3-fold higher rate of aortic plaque progression due to several mechanisms, including chronic inflammation and overexpression of TGF-β/Suppressor of Mothers against Decapentaplegic (SMAD) signaling [96,97]. Given that CBX3 is reported to enhance pancreatic cancer progression by inhibiting SMURF2 and promoting the activation of TGF-β signaling [17], we speculate that selective inhibition of CBX3 could also be beneficial for the stabilization of atherosclerotic plaque in high-risk patients through TGF-β signaling pathway downregulation. Moreover, CBX3 role in atherosclerotic plaque biology extends to the regulation of foam cell formation, which occurs when macrophages engulf oxidized low-density lipoprotein [98]. By affecting these critical cellular events, CBX3 might contribute to plaque stability and the potential for plaque rupture, which are pivotal in the clinical manifestations of atherosclerosis. One of the critical functions of CBX3 is its ability to prevent the activation of the NLR (nucleotide-binding leucine-rich repeat receptors) family pyrin domain containing 3 (NLRP3) inflammasome, a crucial component in the innate immune response, particularly in the presence of cholesterol crystals, which are often abundant in atherosclerotic lesions [99]. By reducing the release of IL-1β, CBX3 directly attenuates inflammatory signaling within cardiovascular tissues, thereby potentially limiting the extent of inflammation and tissue damage [100]. Furthermore, CBX3 also impedes the vesicular secretion of various inflammatory signals, which is another critical mechanism by which it helps to control the inflammatory environment in these tissues [101]. This multiple approach to inflammation management underscores the complex interplay between the different cellular and molecular processes that CBX3 regulates, highlighting its potential as a therapeutic target for CVD characterized by chronic inflammation. Recent research has highlighted the multifaceted role of CBX3, particularly regarding its interaction with other key immune mediators. One significant interaction involves Coenzyme Q (CoQ), a crucial molecule in cellular energy production and antioxidant defense, with ABCG1 (e.g. microRNA-33), which facilitate the process crucial for macrophage reverse cholesterol transport in CVD[102,103]. This transport mechanism is vital as it helps to clear cholesterol from macrophages in atherosclerotic plaques, reducing the risk of plaque formation and subsequent cardiovascular events[104]. Furthermore, the chemokine CX3CL1, enriched in atherosclerotic plaques, plays a pivotal role by attracting CX3CR1 positive NK cells, subsets of T-cells, and mast cells [105,106]. This chemokine-receptor interaction is critical because it modulates the inflammatory response within the plaques, which can either exacerbate or mitigate disease progression[107,108]. Table 2 lists the mechanisms immune-regulated by CBX3 and their correlation with CVD. Addressing interactions with immune pathways could lead to novel strategies for preventing and treating atherosclerosis and other cardiovascular conditions. The specific pathways through which CBX3 impacts these processes are not yet well defined. This emphasizes the need for future work that will deconvolute how CBX3 is playing a regulatory role in endothelial function and immune cell infiltration during CVD contexts.
6. The Potential Therapeutic Benefits of Targeting CBX3 in CVDs
Epigenetic pathway has a strong involvement in CVD providing alternative therapeutic avenue for interventions [109] . Specifically, targeting CBX3 has shown promising results to reducing the progression of CVD [110]. Thus, therapeutic strategies modulating CBX3 expression may offer a new way of managing the progression of CVD and decreasing related risks. The interplay between genetic and environmental factors in CVD also points up the importance of exploring epigenetic pathways for therapeutic interventions [34,111]. Clinical trials and research investigating CBX3-targeted therapies are introducing the possible advantages of epigenetic interventions in CVD management. For example, BET-onMACE Phase 3 clinical trial may reveal cardiovascular efficacy and safety data on apa-betalone, which means such epigenetic-based therapies can be used in further clinical applications for patients with CVDs [112]. In addition, research on histone deacetylases and acetyltransferases bring to the fore their significance as principal epigenetic regulators in heart health, underlining the importance of the therapy targeting these enzymes [113]. CBX3-targeting therapeutic strategies have shown promise in augmenting myocardial repair and enhancing cardiac function after infarction. One way of doing this is by modulating CBX3 activity to reduce adverse remodeling and fibrosis, therefore maintaining myocardial structure and function [114]. Targeted therapies could involve small molecule inhibitors or gene-silencing approaches such as RNA interference to reduce CBX3 expression. Or alternatively, based on the patient-specific context, improve strategies able to increase CBX3 expression. Interestingly, these approaches may be combined with regenerative strategies like stem cell transplantation to promote tissue repair and regeneration making CBX3 a key therapeutic target for fine-tuning the detrimental effects of myocardial infarction and enhancing recovery of the heart. Clinical trials targeting CBX3 in myocardial infarction would be essential for translation of these strategies into routine clinical practice. Unravelling the complex interplay of epigenetic modifications with cardiovascular health, it is necessary to improve diagnostic precision, prognostic potency, and therapeutic achievability within cardiology [115]. The exploration of epigenetic biomarkers and targets is currently shifting the paradigm to precision medicine in cardiovascular care that will allow evidence-based reviews on innovative and targeted interventions for managing CVD [116] . Therefore, examining the therapeutic potential of CBX3 in CVD could reveal new treatment approaches that might bring about a transformation in the field of cardiovascular medicine.
7. Conclusions and Perspectives
The area of CVD epigenetics is a newly emerging area of research that focuses on under-standing the impact of epigenetic modifications on the development and progression of CVD [117]. In this review, we describe the epigenetic factor CBX3 in the context of cardiovascular health to provide valuable insights into mechanism through which epigenetic modifications influence disease susceptibility and progression thus paving the way for innovative approaches in CVD management. CBX3 inhibits VSMC proliferation and migration to prevent exaggerated neointima development after vascular injury, yet it interacts with Dia-1, Notch3, SRF: critical regulators of VSMC phenotype and differentiation. Inhibition of VSMCs viability and collagen over deposition are the effects of its overexpression [118]. Taken together, these findings suggest that in normal conditions, CBX3 might act to preclude pathogenic vascular remodeling. The development of advanced technologies, including those of NGS applications, would make it possible to realize further research aimed at elucidating the complex molecular mechanisms related to the functional activity of CBX3; this type of study would be particularly useful for lung cancer associated with CVD, giving a more personalized molecular signature in patients presenting both risk factors. The fine CBX3 action mechanisms need distinct multilevel analysis on the molecular and cellular side. For instance, the identification of 50 genes as a CBX3-binding protein identified by the STRING software opens new possibilities to elucidate how CBX3 interacts with other genes within cellular pathways [14]. Furthermore, the mention of CBX3 in studies focusing on other chromatin regulators in heart disease solidifies its potential importance as a major player in epigenetic modification of CVD. This is further line of evidence underpinning the major contributory role for this gene in cardiac abnormalities. Based on the rhetoric surrounding heart disease, recent findings have further amplified the focus on those molecular underpinnings that could uncover new treatments. The revelations on CBX3 involvement portray a potential target for developed therapies, emphasizing the need for more specific research on its role in the cardiovascular model, which fits into larger research trajectories trying to decipher molecular roads to CVD, a key development in prevention and treatment strategies [119]. Because of the critical impact of CVDs on global mortality, finding effective biomarkers for early detection and personalized treatment approaches has become very important. As the cardiovascular medicine field moves toward more personalized and precise treatments, having identified biomarkers, such as CBX3, becomes very crucial. It is likely that personal immunotherapy approaches and combination therapies will represent future directions for CVD treatment [120,121]. However, the demographics and medication-based traditional methods of risk estimation still leave some at-risk individuals undetected[122,123] The alarming statistics presented only underscore the urgent need for comprehensive strategies to address challenges (multifaceted) of CVDs from prevention to management and mitigation of socio-economic impacts [124]. This gives grounds that strategies towards CBX3 may be very promising in terms of therapy to reduce adverse vascular remodeling, which is one of the key unmet challenges for effective treatment in cardiovascular medicine. Collectively, we believe our review can be helpful in getting insight into very complex mosaic puzzles of CVDs, which could be solved by a continues exploration of molecular pathways and potential therapeutic targets in order to advance prevention and treatment strategies in combatting health collapse.
Author Contributions
M.C. was responsible for the conceptualization, design of the study and image processing. L.A. supervised the study. M.A.W. and B.G. wrote and edited the manuscript. A.N., N.D.G., V.Q., and N.M. revised the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by PNRR-MAD-2022-12376723; PNRR-CN3, National Centre for Gene Therapy and Drugs Based on RNA Technology, cod: CN000000041. PNRR Project AN-THEM (AdvaNced Technologies for Human-cEntred Medicine) CUP: B53C22006540001. EPI- MET Fondo Crescita Sostenibile – Accordi per l’Innovazione D.M. 31.12.2021, D.D. 18.03.2022 no. 34; no. project F/310034/03/X56 (VANVITELLI). MUR PRIN/PNRR2022 P2022KMP9K (MC); P2022F3YRF(LA); Bando di Ateneo per il finanziamento di progetti di ricerca fondamentale ed ap-plicata dedicato ai giovani Ricercatori D.R. no. 834 del 30/09/2022: IDEA (CUP: B63C22001470005). VALERE: EPInhibitDRUGre (CUP B66J20000680005). VALERE 2020 “NETWINS” D.R. no. 138 of 17/02/2020. NDG was supported by PON Ricerca e Innovazione 2014–2020 Linea 1, AIM (AIM1859703); M.A.W. and B.G. are PhD students in translational medicine.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Acknowledgments
The figures in this review article were created with BioRender.com.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ABCG1 ATP Binding Cassette Subfamily G Member 1 |
| CBX chromobox |
| CoQ coenzyme Q |
| CVD cardiovascular disease |
| DM diabetes mellitus |
| ICIs immune checkpoint inhibitors |
| MS metabolic syndrome |
| NLR nucleotide-binding leucine-rich repeat receptors |
| NLRP3 NLR family pyrin domain containing 3 |
| NPC Neural progenitor cells |
| SRF serum response factor |
| SMURF1 SMAD specific E3 ubiquitin protein ligase 1 |
| SMURF2 SMAD specific E3 ubiquitin protein ligase 2 |
| SMCs smooth muscle cells |
| SMAD TGF-β/Suppressor of Mothers against Decapentaplegic |
| tiRNA-Gly-GCC tRNA with the anticodon GCC |
| VSMCs vascular smooth muscle cells |
References
- Brown, J.C.; Gerhardt, T.E.; Kwon, E. Risk Factors for Coronary Artery Disease; 2024.
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990-2019: Update From the GBD 2019 Study. J Am Coll Cardiol 2020, 76, 2982–3021. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, M.; Perel, P.; Taylor, S.; Kabudula, C.; Bixby, H.; Gaziano, T.A.; McGhie, D.V.; Mwangi, J.; Pervan, B.; Narula, J.; et al. The Heart of the World. Glob Heart 2024, 19, 11. [Google Scholar] [CrossRef] [PubMed]
- Bin, W.; Le, Z.; Mubarik, S.; Fu, G.; Wang, Y.; Nawsherwan. Prediction of Cardiovascular Diseases Mortality- and Disability-Adjusted Life-Years Attributed to Modifiable Dietary Risk Factors from 1990 to 2030 among East Asian Countries and the World. Front Nutr 2022, 9, 898978. [Google Scholar] [CrossRef]
- Gooding, H.C.; Gidding, S.S.; Moran, A.E.; Redmond, N.; Allen, N.B.; Bacha, F.; Burns, T.L.; Catov, J.M.; Grandner, M.A.; Harris, K.M.; et al. Challenges and Opportunities for the Prevention and Treatment of Cardiovascular Disease Among Young Adults: Report From a National Heart, Lung, and Blood Institute Working Group. J Am Heart Assoc 2020, 9, e016115. [Google Scholar] [CrossRef]
- Ullah, A.; Kumar, M.; Sayyar, M.; Sapna, F.; John, C.; Memon, S.; Qureshi, K.; Agbo, E.C.; Ariri, H.I.; Chukwu, E.J.; et al. Revolutionizing Cardiac Care: A Comprehensive Narrative Review of Cardiac Rehabilitation and the Evolution of Cardiovascular Medicine. Cureus 2023, 15, e46469. [Google Scholar] [CrossRef]
- Ordovás, J.M.; Smith, C.E. Epigenetics and Cardiovascular Disease. Nat Rev Cardiol 2010, 7, 510–519. [Google Scholar] [CrossRef]
- Elendu, C.; Amaechi, D.C.; Elendu, T.C.; Fiemotonghan, B.-E.; Okoye, O.K.; Agu-Ben, C.M.; Onyekweli, S.O.; Amapu, D.A.; Ikpegbu, R.; Asekhauno, M.; et al. A Comprehensive Review of Heart Failure: Unraveling the Etiology, Decoding Pathophysiological Mechanisms, Navigating Diagnostic Modalities, Exploring Pharmacological Interventions, Advocating Lifestyle Modifications, and Charting the Horizon of Emerging Therapies in the Complex Landscape of Chronic Cardiac Dysfunction. Medicine 2024, 103, e36895. [Google Scholar] [CrossRef]
- Crowson, C.S.; Liao, K.P.; Davis, J.M.; Solomon, D.H.; Matteson, E.L.; Knutson, K.L.; Hlatky, M.A.; Gabriel, S.E. Rheumatoid Arthritis and Cardiovascular Disease. Am Heart J 2013, 166, 622–628. [Google Scholar] [CrossRef]
- Fuchs, F.D.; Whelton, P.K. High Blood Pressure and Cardiovascular Disease. Hypertension 2020, 75, 285–292. [Google Scholar] [CrossRef]
- Kannel, W.B. Progression of Uncontrolled Hypertension and Implications for Managing Its Sequelae. Manag Care 2003, 12, 26–33. [Google Scholar]
- Zhang, C.; Chen, D.; Maguire, E.M.; He, S.; Chen, J.; An, W.; Yang, M.; Afzal, T.A.; Luong, L.A.; Zhang, L.; et al. Cbx3 Inhibits Vascular Smooth Muscle Cell Proliferation, Migration, and Neointima Formation. Cardiovasc Res 2018, 114, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.W.; Roubin, G.S.; King, S.B. Restenosis after Coronary Angioplasty. Potential Biologic Determinants and Role of Intimal Hyperplasia. Circulation 1989, 79, 1374–1387. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Chen, P.; Fan, L.; Sun, B. Comprehensive Pan-Cancer Analysis on CBX3 as a Prognostic and Immunological Biomarker. BMC Med Genomics 2022, 15, 29. [Google Scholar] [CrossRef]
- Kunkiel, J.; Gödecke, N.; Ackermann, M.; Hoffmann, D.; Schambach, A.; Lachmann, N.; Wirth, D.; Moritz, T. The CpG-Sites of the CBX3 Ubiquitous Chromatin Opening Element Are Critical Structural Determinants for the Anti-Silencing Function. Sci Rep 2017, 7, 7919. [Google Scholar] [CrossRef]
- Huang, C.; Su, T.; Xue, Y.; Cheng, C.; Lay, F.D.; McKee, R.A.; Li, M.; Vashisht, A.; Wohlschlegel, J.; Novitch, B.G.; et al. Cbx3 Maintains Lineage Specificity during Neural Differentiation. Genes Dev 2017, 31, 241–246. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, H.; Ren, D.; Sun, Y.; Guo, F.; Cai, H.; Zhou, C.; Zhou, Y.; Jin, X.; Wu, H. CBX3 Regulated By YBX1 Promotes Smoking-Induced Pancreatic Cancer Progression via Inhibiting SMURF2 Expression. Int J Biol Sci 2022, 18, 3484–3497. [Google Scholar] [CrossRef]
- Koseler, A.; Ma, F.; Kilic, I.D.; Morselli, M.; Kilic, O.; Pellegrini, M. Genome-Wide DNA Methylation Profiling of Blood from Monozygotic Twins Discordant for Myocardial Infarction. In Vivo 2020, 34, 361–367. [Google Scholar] [CrossRef]
- Muka, T.; Koromani, F.; Portilla, E.; O’Connor, A.; Bramer, W.M.; Troup, J.; Chowdhury, R.; Dehghan, A.; Franco, O.H. The Role of Epigenetic Modifications in Cardiovascular Disease: A Systematic Review. Int J Cardiol 2016, 212, 174–183. [Google Scholar] [CrossRef]
- Lorenzen, J.M.; Martino, F.; Thum, T. Epigenetic Modifications in Cardiovascular Disease. Basic Res Cardiol 2012, 107, 245. [Google Scholar] [CrossRef]
- Yabe, K.; Kamio, A.; Oya, S.; Kakutani, T.; Hirayama, M.; Tanaka, Y.; Inagaki, S. H3K9 Methylation Regulates Heterochromatin Silencing through Incoherent Feedforward Loops. Sci Adv 2024, 10, eadn4149. [Google Scholar] [CrossRef]
- Ninova, M.; Fejes Tóth, K.; Aravin, A.A. The Control of Gene Expression and Cell Identity by H3K9 Trimethylation. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Ouyang, H.; Amaya, M.F.; Ravichandran, M.; Loppnau, P.; Min, J.; Zang, J. Structural Basis of the Chromodomain of Cbx3 Bound to Methylated Peptides from Histone H1 and G9a. PLoS One 2012, 7, e35376. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, R.; Gonzales-Cope, M.; Chronis, C.; Bonora, G.; McKee, R.; Huang, C.; Patel, S.; Lopez, D.; Mishra, N.; Pellegrini, M.; et al. Proteomic and Genomic Approaches Reveal Critical Functions of H3K9 Methylation and Heterochromatin Protein-1γ in Reprogramming to Pluripotency. Nat Cell Biol 2013, 15, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.A.; Vondriska, T.M. Clinical Epigenomics for Cardiovascular Disease: Diagnostics and Therapies. J Mol Cell Cardiol 2021, 154, 97–105. [Google Scholar] [CrossRef]
- Kolbus, D.; Ljungcrantz, I.; Andersson, L.; Hedblad, B.; Fredrikson, G.N.; Björkbacka, H.; Nilsson, J. Association between CD8+ T-Cell Subsets and Cardiovascular Disease. J Intern Med 2013, 274, 41–51. [Google Scholar] [CrossRef]
- Russell-Hallinan, A.; Watson, C.J.; O’Dwyer, D.; Grieve, D.J.; O’Neill, K.M. Epigenetic Regulation of Endothelial Cell Function by Nucleic Acid Methylation in Cardiac Homeostasis and Disease. Cardiovasc Drugs Ther 2021, 35, 1025–1044. [Google Scholar] [CrossRef]
- Lozano-Velasco, E.; Inácio, J.M.; Sousa, I.; Guimarães, A.R.; Franco, D.; Moura, G.; Belo, J.A. MiRNAs in Heart Development and Disease. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Ha, T.-Y. MicroRNAs in Human Diseases: From Cancer to Cardiovascular Disease. Immune Netw 2011, 11, 135–154. [Google Scholar] [CrossRef]
- Nonaka, C.K.V.; Sampaio, G.L.; Silva, K.N.; Khouri, R.; Macedo, C.T.; Rogatto, S.R.; Ribeiro Dos Santos, R.; Souza, B.S. de F.; Soares, M.B.P.; Chagas Translational Research Consortium. Therapeutic MiR-21 Silencing Reduces Cardiac Fibrosis and Modulates Inflammatory Response in Chronic Chagas Disease. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Mierzejewski, B.; Ciemerych, M.A.; Streminska, W.; Janczyk-Ilach, K.; Brzoska, E. MiRNA-126a Plays Important Role in Myoblast and Endothelial Cell Interaction. Sci Rep 2023, 13, 15046. [Google Scholar] [CrossRef]
- Arderiu, G.; Peña, E.; Civit-Urgell, A.; Badimon, L. Endothelium-Released Microvesicles Transport MiR-126 That Induces Proangiogenic Reprogramming in Monocytes. Front Immunol 2022, 13, 836662. [Google Scholar] [CrossRef]
- Chen, J.; Lin, Y.; Zheng, S.; Chen, Q.; Tang, S.; Zhong, X. CBX3 Promotes Clear Cell Renal Carcinoma through PI3K/AKT Activation and Aberrant Immunity. J Transl Med 2023, 21, 600. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Teng, X.; Luo, C.; Kong, L. Mechanisms and Advances of Epigenetic Regulation in Cardiovascular Disease. Frontiers in Bioscience-Landmark 2024, 29, 205. [Google Scholar] [CrossRef] [PubMed]
- van der Laan, S.W.; Siemelink, M.A.; Haitjema, S.; Foroughi Asl, H.; Perisic, L.; Mokry, M.; van Setten, J.; Malik, R.; Dichgans, M.; Worrall, B.B.; et al. Genetic Susceptibility Loci for Cardiovascular Disease and Their Impact on Atherosclerotic Plaques. Circ Genom Precis Med 2018, 11, e002115. [Google Scholar] [CrossRef]
- Palou-Márquez, G.; Subirana, I.; Nonell, L.; Fernández-Sanlés, A.; Elosua, R. DNA Methylation and Gene Expression Integration in Cardiovascular Disease. Clin Epigenetics 2021, 13, 75. [Google Scholar] [CrossRef]
- Miroshnikova, V. V; Panteleeva, A.A.; Pobozheva, I.A.; Razgildina, N.D.; Polyakova, E.A.; Markov, A. V; Belyaeva, O.D.; Berkovich, O.A.; Baranova, E.I.; Nazarenko, M.S.; et al. ABCA1 and ABCG1 DNA Methylation in Epicardial Adipose Tissue of Patients with Coronary Artery Disease. BMC Cardiovasc Disord 2021, 21, 566. [Google Scholar] [CrossRef]
- Jin, J.; Zhao, X.; Zhu, C.; Li, M.; Wang, J.; Fan, Y.; Liu, C.; Shen, C.; Yang, R. Hypomethylation of ABCG1 in Peripheral Blood as a Potential Marker for the Detection of Coronary Heart Disease. Clin Epigenetics 2023, 15, 120. [Google Scholar] [CrossRef]
- Hedman, Å.K.; Mendelson, M.M.; Marioni, R.E.; Gustafsson, S.; Joehanes, R.; Irvin, M.R.; Zhi, D.; Sandling, J.K.; Yao, C.; Liu, C.; et al. Epigenetic Patterns in Blood Associated With Lipid Traits Predict Incident Coronary Heart Disease Events and Are Enriched for Results From Genome-Wide Association Studies. Circ Cardiovasc Genet 2017, 10, e001487. [Google Scholar] [CrossRef]
- Bauer, A.J.; Martin, K.A. Coordinating Regulation of Gene Expression in Cardiovascular Disease: Interactions between Chromatin Modifiers and Transcription Factors. Front Cardiovasc Med 2017, 4, 19. [Google Scholar] [CrossRef]
- Cao, G.; Xuan, X.; Zhang, R.; Hu, J.; Dong, H. Gene Therapy for Cardiovascular Disease: Basic Research and Clinical Prospects. Front Cardiovasc Med 2021, 8, 760140. [Google Scholar] [CrossRef]
- Alfaddagh, A.; Martin, S.S.; Leucker, T.M.; Michos, E.D.; Blaha, M.J.; Lowenstein, C.J.; Jones, S.R.; Toth, P.P. Inflammation and Cardiovascular Disease: From Mechanisms to Therapeutics. Am J Prev Cardiol 2020, 4, 100130. [Google Scholar] [CrossRef] [PubMed]
- Bairey Merz, C.N.; Dwyer, J.; Nordstrom, C.K.; Walton, K.G.; Salerno, J.W.; Schneider, R.H. Psychosocial Stress and Cardiovascular Disease: Pathophysiological Links. Behavioral medicine (Washington, D.C.) 2002, 27, 141–147. [Google Scholar] [CrossRef]
- De Rosa, S.; Arcidiacono, B.; Chiefari, E.; Brunetti, A.; Indolfi, C.; Foti, D.P. Type 2 Diabetes Mellitus and Cardiovascular Disease: Genetic and Epigenetic Links. Front Endocrinol (Lausanne) 2018, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Malka, K.; Liaw, L. NOTCH3 as a Modulator of Vascular Disease: A Target in Elastin Deficiency and Arterial Pathologies. J Clin Invest 2022, 132. [Google Scholar] [CrossRef]
- Rusanescu, G.; Weissleder, R.; Aikawa, E. Notch Signaling in Cardiovascular Disease and Calcification. Curr Cardiol Rev 2008, 4, 148–156. [Google Scholar] [CrossRef]
- Gomez, A.H.; Joshi, S.; Yang, Y.; Tune, J.D.; Zhao, M.-T.; Yang, H. Bioengineering Systems for Modulating Notch Signaling in Cardiovascular Development, Disease, and Regeneration. J Cardiovasc Dev Dis 2021, 8. [Google Scholar] [CrossRef]
- Kachanova, O.; Lobov, A.; Malashicheva, A. The Role of the Notch Signaling Pathway in Recovery of Cardiac Function after Myocardial Infarction. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Baeten, J.T.; Lilly, B. Notch Signaling in Vascular Smooth Muscle Cells. Adv Pharmacol 2017, 78, 351–382. [Google Scholar] [CrossRef]
- Jensen, L.F.; Bentzon, J.F.; Albarrán-Juárez, J. The Phenotypic Responses of Vascular Smooth Muscle Cells Exposed to Mechanical Cues. Cells 2021, 10. [Google Scholar] [CrossRef]
- Steffensen, L.B.; Rasmussen, L.M. A Role for Collagen Type IV in Cardiovascular Disease? Am J Physiol Heart Circ Physiol 2018, 315, H610–H625. [Google Scholar] [CrossRef]
- Jiang, Y.; Qian, H.-Y. Transcription Factors: Key Regulatory Targets of Vascular Smooth Muscle Cell in Atherosclerosis. Mol Med 2023, 29, 2. [Google Scholar] [CrossRef]
- Alva, J.A.; Iruela-Arispe, M.L. Notch Signaling in Vascular Morphogenesis. Curr Opin Hematol 2004, 11, 278–283. [Google Scholar] [CrossRef]
- Xiu, M.; Wang, Y.; Li, B.; Wang, X.; Xiao, F.; Chen, S.; Zhang, L.; Zhou, B.; Hua, F. The Role of Notch3 Signaling in Cancer Stemness and Chemoresistance: Molecular Mechanisms and Targeting Strategies. Front Mol Biosci 2021, 8, 694141. [Google Scholar] [CrossRef]
- Wang, K.; Liu, C.-Y.; Fang, B.; Li, B.; Li, Y.-H.; Xia, Q.-Q.; Zhao, Y.; Cheng, X.-L.; Yang, S.-M.; Zhang, M.-H.; et al. The Function and Therapeutic Potential of Transfer RNA-Derived Small RNAs in Cardiovascular Diseases: A Review. Pharmacol Res 2024, 206, 107279. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, K.; Zhao, C.; Liu, N.; Wang, Z.; Yang, W.; Cheng, Z.; Zhou, L.; Wang, K. The Function of TRNA-Derived Small RNAs in Cardiovascular Diseases. Mol Ther Nucleic Acids 2024, 35, 102114. [Google Scholar] [CrossRef]
- Rong, Z.; Li, F.; Zhang, R.; Niu, S.; Di, X.; Ni, L.; Liu, C. Inhibition of TiRNA-Gly-GCC Ameliorates Neointimal Formation via CBX3-Mediated VSMCs Phenotypic Switching. Front Cardiovasc Med 2023, 10, 1030635. [Google Scholar] [CrossRef]
- Xiao, Q.; Wang, G.; Yin, X.; Luo, Z.; Margariti, A.; Zeng, L.; Mayr, M.; Ye, S.; Xu, Q. Chromobox Protein Homolog 3 Is Essential for Stem Cell Differentiation to Smooth Muscles in Vitro and in Embryonic Arteriogenesis. Arterioscler Thromb Vasc Biol 2011, 31, 1842–1852. [Google Scholar] [CrossRef]
- Dorn, T.; Kornherr, J.; Parrotta, E.I.; Zawada, D.; Ayetey, H.; Santamaria, G.; Iop, L.; Mastantuono, E.; Sinnecker, D.; Goedel, A.; et al. Interplay of Cell-Cell Contacts and RhoA/MRTF-A Signaling Regulates Cardiomyocyte Identity. EMBO J 2018, 37. [Google Scholar] [CrossRef]
- Wang, C.; Lu, D.; Cronin-Fenton, D.; Huang, C.; Liew, Z.; Wei, D.; Qin, G.; Yu, Y.; Li, J. Cardiovascular Disease and Risk of Lung Cancer Incidence and Mortality: A Nationwide Matched Cohort Study. Front Oncol 2022, 12, 950971. [Google Scholar] [CrossRef]
- de Jesus, M.; Chanda, A.; Grabauskas, T.; Kumar, M.; Kim, A.S. Cardiovascular Disease and Lung Cancer. Front Oncol 2024, 14, 1258991. [Google Scholar] [CrossRef]
- Chianca, M.; Panichella, G.; Fabiani, I.; Giannoni, A.; L’Abbate, S.; Aimo, A.; Del Franco, A.; Vergaro, G.; Grigoratos, C.; Castiglione, V.; et al. Bidirectional Relationship Between Cancer and Heart Failure: Insights on Circulating Biomarkers. Front Cardiovasc Med 2022, 9, 936654. [Google Scholar] [CrossRef] [PubMed]
- Koene, R.J.; Prizment, A.E.; Blaes, A.; Konety, S.H. Shared Risk Factors in Cardiovascular Disease and Cancer. Circulation 2016, 133, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Chen, Z.; Shen, W.; Huang, G.; Sedivy, J.M.; Wang, H.; Ju, Z. Inflammation, Epigenetics, and Metabolism Converge to Cell Senescence and Ageing: The Regulation and Intervention. Signal Transduct Target Ther 2021, 6, 245. [Google Scholar] [CrossRef]
- Hatlen, P.; Langhammer, A.; Carlsen, S.M.; Salvesen, Ø.; Amundsen, T. Self-Reported Cardiovascular Disease and the Risk of Lung Cancer, the HUNT Study. Journal of Thoracic Oncology 2014, 9, 940–946. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, L.; Shi, S.; He, H.; Shen, Q.; Wang, H.; Qin, S.; Chang, J.; Zhong, R. Bidirectional Association Between Cardiovascular Disease and Lung Cancer in a Prospective Cohort Study. J Thorac Oncol 2024, 19, 80–93. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and Tumor Progression: Signaling Pathways and Targeted Intervention. Signal Transduct Target Ther 2021, 6, 263. [Google Scholar] [CrossRef]
- Chehelgerdi, M.; Behdarvand Dehkordi, F.; Chehelgerdi, M.; Kabiri, H.; Salehian-Dehkordi, H.; Abdolvand, M.; Salmanizadeh, S.; Rashidi, M.; Niazmand, A.; Ahmadi, S.; et al. Exploring the Promising Potential of Induced Pluripotent Stem Cells in Cancer Research and Therapy. Mol Cancer 2023, 22, 189. [Google Scholar] [CrossRef]
- Hebib, L.; Herraiz-Adillo, Á.; Higueras-Fresnillo, S.; Berglind, D.; Daka, B.; Wennberg, P.; Hagström, E.; Lenander, C.; Ahlqvist, V.H.; Östgren, C.J.; et al. Life’s Essential 8 Is Inversely Associated with High-Sensitivity C-Reactive Protein. Sci Rep 2024, 14, 15024. [Google Scholar] [CrossRef]
- Mossmann, M.; Wainstein, M.V.; Mariani, S.; Machado, G.P.; de Araújo, G.N.; Andrades, M.; Gonçalves, S.C.; Bertoluci, M.C. Increased Serum IL-6 Is Predictive of Long-Term Cardiovascular Events in High-Risk Patients Submitted to Coronary Angiography: An Observational Study. Diabetol Metab Syndr 2022, 14, 125. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb Perspect Biol 2014, 6, a016295. [Google Scholar] [CrossRef]
- Ridker, P.M. From C-Reactive Protein to Interleukin-6 to Interleukin-1: Moving Upstream To Identify Novel Targets for Atheroprotection. Circ Res 2016, 118, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, M.; Hong, D.; Zeng, M.; Zhang, X. The Paradoxical Role of Cellular Senescence in Cancer. Front Cell Dev Biol 2021, 9, 722205. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Jat, P. Mechanisms of Cellular Senescence: Cell Cycle Arrest and Senescence Associated Secretory Phenotype. Front Cell Dev Biol 2021, 9, 645593. [Google Scholar] [CrossRef] [PubMed]
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic Reprogramming and Cancer Progression. Science 2020, 368. [Google Scholar] [CrossRef] [PubMed]
- Hammoudi, N.; Ahmed, K.B.R.; Garcia-Prieto, C.; Huang, P. Metabolic Alterations in Cancer Cells and Therapeutic Implications. Chin J Cancer 2011, 30, 508–525. [Google Scholar] [CrossRef]
- Dudley, A.C.; Griffioen, A.W. Pathological Angiogenesis: Mechanisms and Therapeutic Strategies. Angiogenesis 2023, 26, 313–347. [Google Scholar] [CrossRef]
- de Boer, R.A.; Hulot, J.-S.; Tocchetti, C.G.; Aboumsallem, J.P.; Ameri, P.; Anker, S.D.; Bauersachs, J.; Bertero, E.; Coats, A.J.S.; Čelutkienė, J.; et al. Common Mechanistic Pathways in Cancer and Heart Failure. A Scientific Roadmap on Behalf of the Translational Research Committee of the Heart Failure Association (HFA) of the European Society of Cardiology (ESC). Eur J Heart Fail 2020, 22, 2272–2289. [Google Scholar] [CrossRef]
- Ray, M.R.; Jablons, D.; He, B. Lung Cancer Therapeutics That Target Signaling Pathways: An Update. Expert Rev Respir Med 2010, 4, 631–645. [Google Scholar] [CrossRef]
- Gallucci, G.; Tartarone, A.; Lerose, R.; Lalinga, A.V.; Capobianco, A.M. Cardiovascular Risk of Smoking and Benefits of Smoking Cessation. J Thorac Dis 2020, 12, 3866–3876. [Google Scholar] [CrossRef]
- Kepka, L.; Bujko, K.; Orlowski, T.M.; Jagiello, R.; Salata, A.; Matecka-Nowak, M.; Janowski, H.; Rogowska, D. Cardiopulmonary Morbidity and Quality of Life in Non-Small Cell Lung Cancer Patients Treated with or without Postoperative Radiotherapy. Radiother Oncol 2011, 98, 238–243. [Google Scholar] [CrossRef]
- Grimaldi, A.M.; Affinito, O.; Salvatore, M.; Franzese, M. CBX Family Members in Two Major Subtypes of Renal Cell Carcinoma: A Comparative Bioinformatic Analysis. Diagnostics (Basel) 2022, 12. [Google Scholar] [CrossRef]
- Zhou, F.; Chen, L.; Lu, P.; Cao, Y.; Deng, C.; Liu, G. An Integrative Bioinformatics Investigation and Experimental Validation of Chromobox Family in Diffuse Large B-Cell Lymphoma. BMC Cancer 2023, 23, 641. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lai, C.; Luo, J.; Shi, J.; Guo, K.; Hu, J.; Mulati, Y.; Xiao, Y.; Kong, D.; Liu, C.; et al. The Predictive Significance of Chromobox Family Members in Prostate Cancer in Humans. Cell Oncol (Dordr) 2024. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Jiang, C.; Chen, D.; Wu, Y.; Lu, J.; Zhong, L.; Yao, F. Analysis of Pan-Cancer Revealed the Immunological and Prognostic Potential of CBX3 in Human Tumors. Front Med (Lausanne) 2022, 9, 869994. [Google Scholar] [CrossRef]
- Xie, L.; Chen, J.; Wang, Y.; Jin, C.; Xie, Y.; Ma, H.; Xiang, M. Emerging Roles of Macrophages in Heart Failure and Associated Treatment Approaches. Ther Adv Chronic Dis 2023, 14, 20406223231168756. [Google Scholar] [CrossRef]
- Le, P.T.; Ha, N.; Tran, N.K.; Newman, A.G.; Esselen, K.M.; Dalrymple, J.L.; Schmelz, E.M.; Bhandoola, A.; Xue, H.-H.; Singh, P.B.; et al. Targeting Cbx3/HP1γ Induces LEF-1 and IL-21R to Promote Tumor-Infiltrating CD8 T-Cell Persistence. Front Immunol 2021, 12, 738958. [Google Scholar] [CrossRef]
- Sun, M.; Ha, N.; Pham, D.-H.; Frederick, M.; Sharma, B.; Naruse, C.; Asano, M.; Pipkin, M.E.; George, R.E.; Thai, T.-H. Cbx3/HP1γ Deficiency Confers Enhanced Tumor-Killing Capacity on CD8+ T Cells. Sci Rep 2017, 7, 42888. [Google Scholar] [CrossRef]
- Jansen, F.; Li, Q.; Pfeifer, A.; Werner, N. Endothelial- and Immune Cell-Derived Extracellular Vesicles in the Regulation of Cardiovascular Health and Disease. JACC Basic Transl Sci 2017, 2, 790–807. [Google Scholar] [CrossRef]
- Li, R.; Jiang, S.; Li, W.; Hong, H.; Zhao, C.; Huang, X.; Zhang, Z.; Li, H.; Chen, H.; Bo, X. Exploration of Prognosis-Related MicroRNA and Transcription Factor Co-Regulatory Networks across Cancer Types. RNA Biol 2019, 16, 1010–1021. [Google Scholar] [CrossRef]
- Asiaee, A.; Abrams, Z.B.; Pua, H.H.; Coombes, K.R. Transcriptome Complexity Disentangled: A Regulatory Molecules Approach. bioRxiv 2024. [Google Scholar] [CrossRef]
- Qin, S.; Xie, B.; Wang, Q.; Yang, R.; Sun, J.; Hu, C.; Liu, S.; Tao, Y.; Xiao, D. New Insights into Immune Cells in Cancer Immunotherapy: From Epigenetic Modification, Metabolic Modulation to Cell Communication. MedComm (Beijing) 2024, 5, e551. [Google Scholar] [CrossRef] [PubMed]
- Napiórkowska-Baran, K.; Schmidt, O.; Szymczak, B.; Lubański, J.; Doligalska, A.; Bartuzi, Z. Molecular Linkage between Immune System Disorders and Atherosclerosis. Curr Issues Mol Biol 2023, 45, 8780–8815. [Google Scholar] [CrossRef] [PubMed]
- Poels, K.; Neppelenbroek, S.I.M.; Kersten, M.J.; Antoni, M.L.; Lutgens, E.; Seijkens, T.T.P. Immune Checkpoint Inhibitor Treatment and Atherosclerotic Cardiovascular Disease: An Emerging Clinical Problem. J Immunother Cancer 2021, 9. [Google Scholar] [CrossRef]
- Tan, S.; Spear, E.; Sane, N.; Chan, J.; Nelson, A.J.; Alamgeer, M.; Nerlekar, N.; Segelov, E.; Nicholls, S.J. Atherosclerotic Cardiovascular Events in Cancer Patients Treated With Immune Checkpoint Inhibitors: A Retrospective Cohort Study. Heart Lung Circ 2024, 33, 721–729. [Google Scholar] [CrossRef]
- Khan, A.; Ley, K. Immunotherapy for Atherosclerosis by Targeting Pro-Inflammatory T Cells. Cell Res 2024, 34, 467–468. [Google Scholar] [CrossRef]
- Suero-Abreu, G.A.; Zanni, M. V; Neilan, T.G. Atherosclerosis With Immune Checkpoint Inhibitor Therapy: Evidence, Diagnosis, and Management: JACC: CardioOncology State-of-the-Art Review. JACC CardioOncol 2022, 4, 598–615. [Google Scholar] [CrossRef]
- Aprotosoaie, A.C.; Costache, A.-D.; Costache, I.-I. Therapeutic Strategies and Chemoprevention of Atherosclerosis: What Do We Know and Where Do We Go? Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Rajamäki, K.; Lappalainen, J.; Öörni, K.; Välimäki, E.; Matikainen, S.; Kovanen, P.T.; Eklund, K.K. Cholesterol Crystals Activate the NLRP3 Inflammasome in Human Macrophages: A Novel Link between Cholesterol Metabolism and Inflammation. PLoS One 2010, 5, e11765. [Google Scholar] [CrossRef]
- Megha, K.B.; Joseph, X.; Akhil, V.; Mohanan, P. V Cascade of Immune Mechanism and Consequences of Inflammatory Disorders. Phytomedicine 2021, 91, 153712. [Google Scholar] [CrossRef]
- Campos, J.H.; Soares, R.P.; Ribeiro, K.; Andrade, A.C.; Batista, W.L.; Torrecilhas, A.C. Extracellular Vesicles: Role in Inflammatory Responses and Potential Uses in Vaccination in Cancer and Infectious Diseases. J Immunol Res 2015, 2015, 832057. [Google Scholar] [CrossRef]
- Sidorkiewicz, M. Is MicroRNA-33 an Appropriate Target in the Treatment of Atherosclerosis? Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Tavoosi, Z.; Moradi-Sardareh, H.; Saidijam, M.; Yadegarazari, R.; Borzuei, S.; Soltanian, A.; Goodarzi, M.T. Cholesterol Transporters ABCA1 and ABCG1 Gene Expression in Peripheral Blood Mononuclear Cells in Patients with Metabolic Syndrome. Cholesterol 2015, 2015, 682904. [Google Scholar] [CrossRef] [PubMed]
- Poznyak, A. V; Kashirskikh, D.A.; Sukhorukov, V.N.; Kalmykov, V.; Omelchenko, A. V; Orekhov, A.N. Cholesterol Transport Dysfunction and Its Involvement in Atherogenesis. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Apostolakis, S.; Spandidos, D. Chemokines and Atherosclerosis: Focus on the CX3CL1/CX3CR1 Pathway. Acta Pharmacol Sin 2013, 34, 1251–1256. [Google Scholar] [CrossRef]
- Duan, Q.; Flynn, C.; Niepel, M.; Hafner, M.; Muhlich, J.L.; Fernandez, N.F.; Rouillard, A.D.; Tan, C.M.; Chen, E.Y.; Golub, T.R.; et al. LINCS Canvas Browser: Interactive Web App to Query, Browse and Interrogate LINCS L1000 Gene Expression Signatures. Nucleic Acids Res 2014, 42, W449–60. [Google Scholar] [CrossRef]
- Russo, R.C.; Garcia, C.C.; Teixeira, M.M.; Amaral, F.A. The CXCL8/IL-8 Chemokine Family and Its Receptors in Inflammatory Diseases. Expert Rev Clin Immunol 2014, 10, 593–619. [Google Scholar] [CrossRef]
- Márquez, A.B.; van der Vorst, E.P.C.; Maas, S.L. Key Chemokine Pathways in Atherosclerosis and Their Therapeutic Potential. J Clin Med 2021, 10. [Google Scholar] [CrossRef]
- Herman, A.B.; Occean, J.R.; Sen, P. Epigenetic Dysregulation in Cardiovascular Aging and Disease. The journal of cardiovascular aging 2021, 1. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, H.; Huang, S.; Yin, L.; Wang, F.; Luo, P.; Huang, H. Epigenetic Regulation in Cardiovascular Disease: Mechanisms and Advances in Clinical Trials. Signal Transduct Target Ther 2022, 7, 200. [Google Scholar] [CrossRef]
- Sawalha, K.; Norgard, N.; López-Candales, A. Epigenetic Regulation and Its Effects on Aging and Cardiovascular Disease. Cureus 2023, 15, e39395. [Google Scholar] [CrossRef]
- Ray, K.K.; Nicholls, S.J.; Buhr, K.A.; Ginsberg, H.N.; Johansson, J.O.; Kalantar-Zadeh, K.; Kulikowski, E.; Toth, P.P.; Wong, N.; Sweeney, M.; et al. Effect of Apabetalone Added to Standard Therapy on Major Adverse Cardiovascular Events in Patients With Recent Acute Coronary Syndrome and Type 2 Diabetes: A Randomized Clinical Trial. JAMA 2020, 323, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Pagiatakis, C.; Di Mauro, V. The Emerging Role of Epigenetics in Therapeutic Targeting of Cardiomyopathies. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Garza, M.A.; Wason, E.A.; Zhang, J.Q. Cardiac Remodeling and Physical Training Post Myocardial Infarction. World J Cardiol 2015, 7, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Sopic, M.; Robinson, E.L.; Emanueli, C.; Srivastava, P.; Angione, C.; Gaetano, C.; Condorelli, G.; Martelli, F.; Pedrazzini, T.; Devaux, Y.; et al. Integration of Epigenetic Regulatory Mechanisms in Heart Failure. Basic Res Cardiol 2023, 118, 16. [Google Scholar] [CrossRef]
- Al-Hasani, K.; Mathiyalagan, P.; El-Osta, A. Epigenetics, Cardiovascular Disease, and Cellular Reprogramming. J Mol Cell Cardiol 2019, 128, 129–133. [Google Scholar] [CrossRef]
- Alghamdi, B.A.; Aljohani, I.M.; Alotaibi, B.G.; Ahmed, M.; Almazmomi, K.A.; Aloufi, S.; Alshamrani, J. Studying Epigenetics of Cardiovascular Diseases on Chip Guide. Cardiogenetics 2022, 12, 218–234. [Google Scholar] [CrossRef]
- Kapustin, A.; Tsakali, S.S.; Whitehead, M.; Chennell, G.; Wu, M.-Y.; Molenaar, C.; Kutikhin, A.; Bogdanov, L.; Sinitsky, M.; Rubina, K.; et al. Extracellular Vesicles Stimulate Smooth Muscle Cell Migration by Presenting Collagen VI. bioRxiv 2023. [Google Scholar] [CrossRef]
- Wagner, N.; Wagner, K.-D. Molecular Mechanisms of Cardiac Development and Disease. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Adhikary, S.; Pathak, S.; Palani, V.; Acar, A.; Banerjee, A.; Al-Dewik, N.I.; Essa, M.M.; Mohammed, S.G.A.A.; Qoronfleh, M.W. Current Technologies and Future Perspectives in Immunotherapy towards a Clinical Oncology Approach. Biomedicines 2024, 12. [Google Scholar] [CrossRef]
- Jyotsna, F.; Ikram, J.; Nageeta, F.; Komal, F.; Anjlee, F.; Patel, H.; Nassri, T.; Kumari, M.; Kumar, R.; Shah, S.U.; et al. Unlocking the Potential of Immunotherapy in Cardiovascular Disease: A Comprehensive Review of Applications and Future Directions. Cureus 2023. [Google Scholar] [CrossRef]
- Suzumura, E.A.; de Oliveira Ascef, B.; Maia, F.H. de A.; Bortoluzzi, A.F.R.; Domingues, S.M.; Farias, N.S.; Gabriel, F.C.; Jahn, B.; Siebert, U.; de Soarez, P.C. Methodological Guidelines and Publications of Benefit-Risk Assessment for Health Technology Assessment: A Scoping Review. BMJ Open 2024, 14, e086603. [Google Scholar] [CrossRef] [PubMed]
- Duffy, A.; Browne, F.; Connolly, M. Safeguarding Adults: A Concept Analysis. J Adv Nurs 2024. [Google Scholar] [CrossRef] [PubMed]
- Sapna, F.; Raveena, F.; Chandio, M.; Bai, K.; Sayyar, M.; Varrassi, G.; Khatri, M.; Kumar, S.; Mohamad, T. Advancements in Heart Failure Management: A Comprehensive Narrative Review of Emerging Therapies. Cureus 2023, 15, e46486. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Adapted graph showing the number of deaths due to each cause estimated annually. Red star indicates the higher number of deaths shared by cardiovascular diseases. Data source: IHME, Global Burden of Disease (2024); https://www.healthdata.org/research-analysis/gbd.
Figure 1.
Adapted graph showing the number of deaths due to each cause estimated annually. Red star indicates the higher number of deaths shared by cardiovascular diseases. Data source: IHME, Global Burden of Disease (2024); https://www.healthdata.org/research-analysis/gbd.
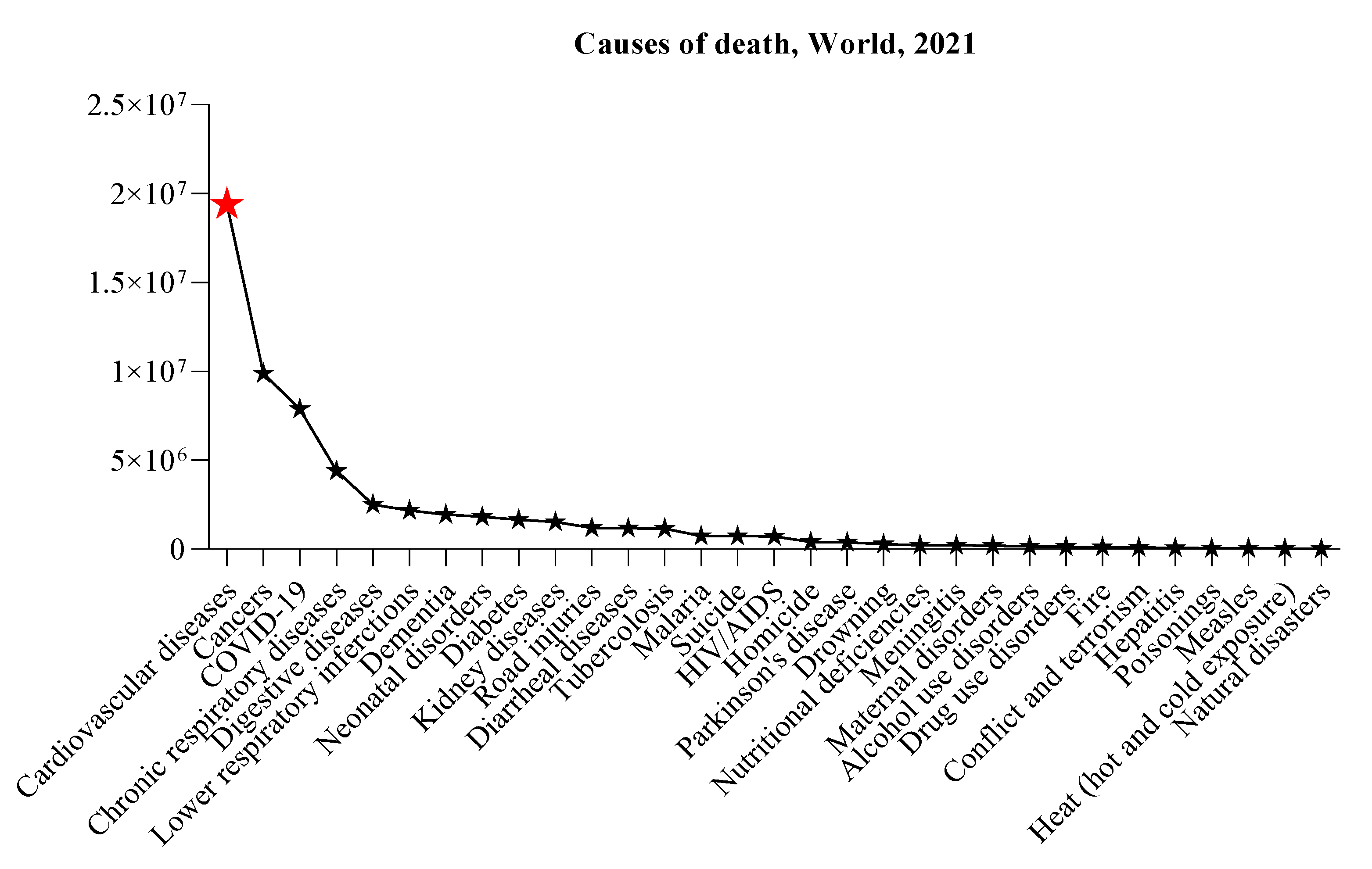
Figure 2.
Schematic representation of CBX3 roles in cardiovascular disease. The involvement of CBX3 brought by Notch3 interaction in controlling VSMC defines the contributions made by proliferation, migration and formation of neointima to pathogenesis of vascular injuries and diseases. CBX3 is involved in the regulation of angiogenesis and could have an impact in restenosis after angioplasty. Epigenetic modifications shared by CBX3 also involve vascular remodeling and homeostasis as well as regulation of angiogenesis and atherosclerosis.
Figure 2.
Schematic representation of CBX3 roles in cardiovascular disease. The involvement of CBX3 brought by Notch3 interaction in controlling VSMC defines the contributions made by proliferation, migration and formation of neointima to pathogenesis of vascular injuries and diseases. CBX3 is involved in the regulation of angiogenesis and could have an impact in restenosis after angioplasty. Epigenetic modifications shared by CBX3 also involve vascular remodeling and homeostasis as well as regulation of angiogenesis and atherosclerosis.
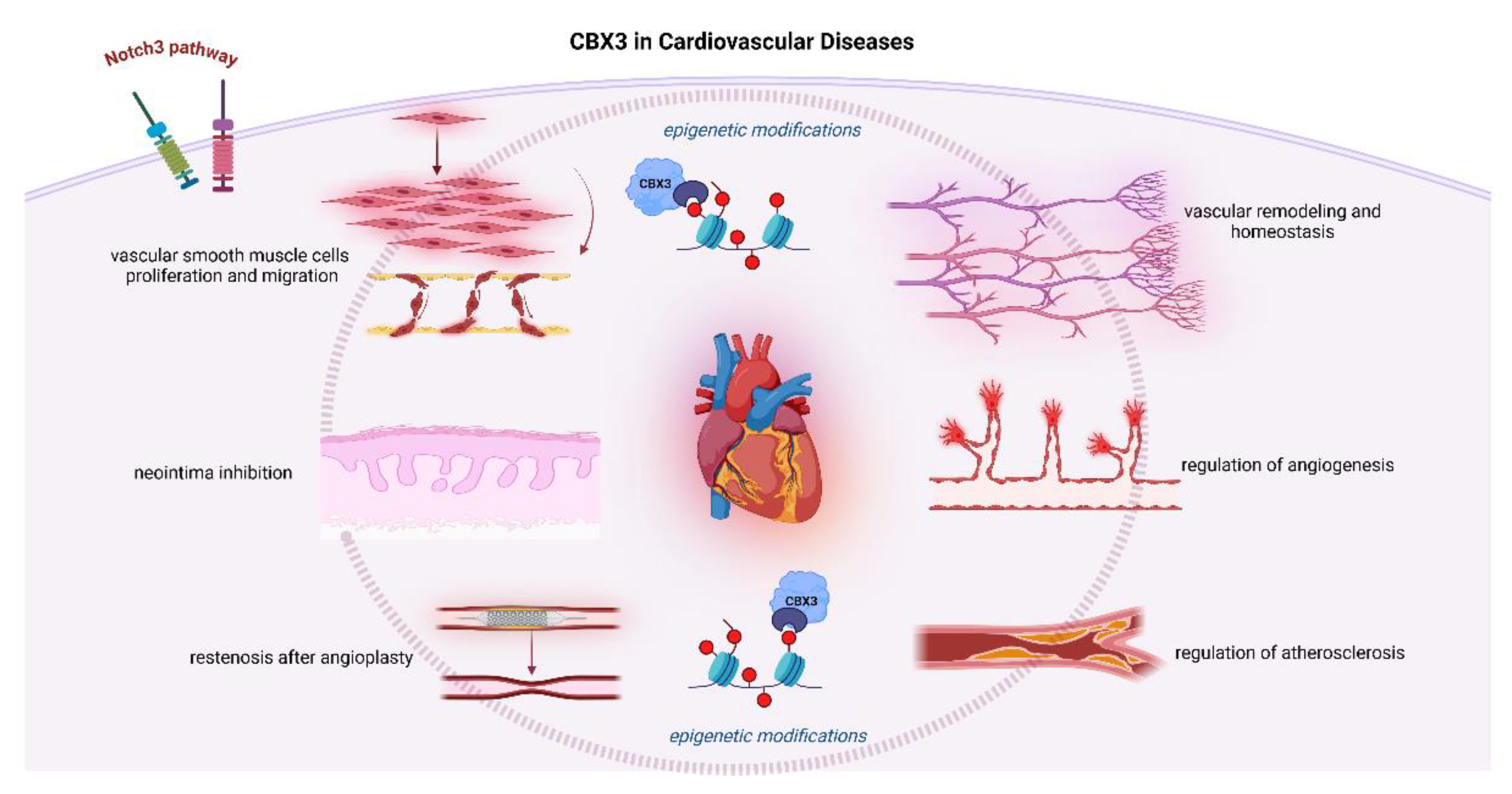
Figure 3.
The interrelated functions of CBX3 with numerous controllers of the cardiovascular and respiratory systems.
Figure 3.
The interrelated functions of CBX3 with numerous controllers of the cardiovascular and respiratory systems.

Table 1.
Epigenetic changes controlled by CBX3 and their relationship with cardiovascular disease.
| Target | CBX3-mediated epigenetic effect |
Effect on CVD | Reference |
|---|---|---|---|
| DNA | binding | regulation of gene transcription and chromatin structure for cardiovascular health | [25] |
| CD8+ T cells | interaction with chromatin modifiers | cardiovascular disease progression and outcomes | [26] |
| miR-21 | repressed expression | cardiac fibrosis attenuation and remodeling in response to stress | [30] |
| miR-126 | expression regulation | maintenance of vascular integrity and promotion angiogenesis | [31,32] |
| Tnnt3, Tbx20, Tbx3, Hand1, PDGFRA | upregulation | cardiac lineage development | [16] |
| Sox1 | downregulation | circulatory system development | [16] |
| Wnt4 | up-regulation | circulatory system development | [16] |
| Cdk8 | decreased recruitment | cardiovascular system development | [33] |
| CPT1A, ABCG1 | negative/positive directions | lipid-associated CpGs | [34] |
| ABCG1 locus | expression | increased triglyceride levels and a heightened risk of new-onset coronary heart disease | [35,36] |
| GATA4 | modulation of chromatin environment | cardiac gene regulation | [40] |
| ApoC3 | altered activity | progression of atherosclerosis | [41] |
Table 2.
Immune system regulated by CBX3 and their association with CVD.
| Target/factor | CBX3-mediated mechanism | Effect on CVD | Reference |
|---|---|---|---|
| CD8+ effector T cells | increase of LEF-1 and IL-21R | immune response | [87,88] |
| B cells | modulation of the local immune response and the structural integrity of blood vessels | progression | [89] |
| immune checkpoints and chemokines |
immune response modulation |
cellular processes and disease states |
[92] |
| Genes in immune activation and suppression |
Interaction |
development of atherosclerosis | [92] |
| SMURF2 inhibition and TGF-β signaling activation |
selective inhibition | stabilization of atherosclerotic plaque | [17] |
| NLRP3 inflammasome | prevention of the activation | atherosclerotic lesions | [99] |
| IL-1β | reduction of the release | limiting the extent of inflammation and tissue damage | [100] |
| inflammatory signals | impedes the vesicular secretion | control of the inflammatory environment |
[101] |
| CoQ |
interaction | macrophage reverse cholesterol transport | [101,102] |
| CX3CL1 | interaction | modulates the inflammatory response within the plaques | [105,106] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



