
Submitted:
02 October 2024
Posted:
03 October 2024
You are already at the latest version
Abstract
Plant-associated yeasts can also be part of soils’ and pollinating-insects’ microbiomes, where they play significant ecological roles. Indeed, in previous studies, we have shown that yeasts may help pollinators find nectar, which is crucial for their nutrition and for the reproduction of many angiosperms, and, in the soil, can act as plant-growth promoters. Given the importance of yeasts for plant development, in this review, we first delve deep into the biochemical processes behind the ecological role of these microorganisms in soil, insects, and in direct association with plants. Based on this premise, we discuss the influence of this relationship on agricultural production and the possible effects of pesticide use on the yeast microbiota. Finally, we address the most relevant studies in the literature that support the potential of these microorganisms (either indigenous or genetically engineered) as bioremediation agents for soils and foods contaminated by pesticides. Our review indicates that yeasts can be satisfactorily employed in organic agriculture to increase plant growth or bioremediate contaminated soil or food.
Keywords:
soil yeast
; bioremediation
; organic agriculture
; plant-growth promoter
; indol-acetic acid
; genetic engineering
; 2
; 4-D
; glyphosate
; whole-cell yeast biocatalists
; Saccharomyces cerevisiae
1. Introduction
With the increase in food production resulting from population growth, the use of chemicals in agriculture seems to be an inevitable practice. However, despite the benefits brought to crops in terms of productivity, the use of agrochemicals generates a series of environmental concerns, especially considering their permanence in the soil and food [1].
For the destination of the residual load of these compounds, the soil's nutrient composition and classification must be considered. Pesticides can alter the different environments' pH, moisture, organic matter content, and microbial communities. On the other hand, the environment microbiota can include bacteria and fungi that are often capable of playing important roles in the biodegradation process of these residues [2].
Yeasts are among the microorganisms associated with plants that can be affected to a greater or lesser extent by the use of pesticides. These unicellular fungi play critical ecological roles, either as natural plant defenders against pathogens, plant growth promoters, or producers of volatile organic compounds (VOCs) that attract pollinators to flowers. Yeasts also play a prominent role in nutrient cycling (biogeochemical cycles) and contribute significantly to the digestive and immune systems of herbivores, pollinivores, and nectarivores [3,4].
In the following sections, we address the biochemical processes involved in the ecological roles of yeasts that benefit plant development, the harmful effects of pesticides on these microorganisms, and how their cells can act as agrochemical degraders.
2. The Contribution of Soil Yeasts to Plant Health
For hundreds of millions of years, yeasts have evolved and conquered their place in the ecological balance, having as their natural habitat the most diverse environments [3]. In these habitats, these microorganisms are not inert; on the contrary, they are active and passive agents, carrying out various biological activities and composing a mix of biochemical reactions with diverse effects on the environments they colonize (Figure 1) [5].
One of their most challenging habitats is the soil, which dwells several microbial species that biochemically interact and alter the environment's physical, chemical, and biological characteristics [6]. The surface layers of the soil concentrate most of the organic matter and biological activity of this system [7]. Consequently, the greatest diversity of yeast species is found in this environment [8,9].
During the battle to maintain life, these microorganisms perform ecological services that help other living organisms survive, including stimulating their development [10]. In fact, yeasts can act as plant growth promoters (Table 1).
Yeasts play a prominent role in nutrient cycling, releasing enzymes into the environment capable of catalyzing the transformation of molecules, especially making them smaller and, thus, more easily used by other organisms. In this context, it is worth mentioning nitrogen as one of the elements made available in the environment through the action of yeasts. The proteolytic enzymes secreted by yeasts increase the availability of essential elements to other microorganisms and plants. In addition to the source of nitrogen, the amount of carbon in the environment is also essential for the growth and development of other species. Xylanases, amylases, pectinases, and cellulases are examples of enzymes capable of making this element available in the environment [11].
Phosphorus-based nutrients are some of the most required by plant species. Their presence in the soil directly impacts plant growth [13]. This nutrient is often found in forms that are not available to plants; however, yeasts such as Pichia kudriavzevii and Issatchenkia terricola was found to provide this important element in forms that are better assimilated by plants, generating an increase of 80.31% and 50.90%, respectively, in the growth of mung bean roots [12].
An increase in phosphorus solubilization has also been observed from the activity of other yeast species, such as Cryptococcus flavus and Candida railenensis, which, when inoculated into the corn rhizosphere, caused an increase in root growth of 53% and 34%, respectively. This increase was attributed to the plant's greater phosphorus absorption, especially when the yeasts were inoculated in consortia with mycorrhizal fungi. Therefore, a synergistic activity between the species can also be observed, relating a greater development of the aerial part of the crop (26%) to an increase (20 to 29%) in phosphorus absorption by the plants [13].
In addition to the greater availability of nutrients, yeasts also promote plant growth by other means, such as the production and release of compounds related to plant growth stimulation, especially indol-acetic acid (IAA). Yeasts of the species Meyerozyma guilliermondii, Candida zemplinina, Candida pimensis, Lachancea lanzarotensis, Rhodotorula mucilaginosa [10], Pichia kudriavzevii, Issatchenkia terricola [12], Kazachstania rupicola, Rhodosporidium diabovatum, and Saccharomyces cerevisiae [14] have already been reported as good IAA producers. The yeast metabolic pathways involved in the production of this important plant-growth promoter are summarized in Figure 2.
IAA is a phytohormone of the auxin class, responsible for stimulating apical and lateral growth in plants through cell elongation [8]. The presence of this phytohormone in plant roots allows for greater development of their root area, increasing the area of contact with the soil and, consequently, increasing the capacity of the plants to intercept and absorb nutrients in the soil [14], thus increasing their ability to compete with pathogenic organisms in the soil [12].
As already mentioned, soil is a habitat for several species of microorganisms, especially in its most superficial area. It is true that phytopathogenic microorganisms are also easily found in this environment [21], but the presence of wild yeast strains also exerts pressure on the survival and establishment of these organisms, as they perform antagonistic activities to them [22], either by competing for physical space [23], for nutrients [12,23,24], or even secreting extracellular enzymes such as β-glucanases and chitinases [22,24] that actively participate in plant protection [12,25], as exemplified in Table 2.
The species Papiliotrema (Cryptococcus) laurentii has already been reported to inhibit the pathogen Pythium ultimum in vitro and in vivo. This inhibition was due to the high lytic activity exerted by the production of large amounts of β-1,3-glucanase by the tested strain [26]. Strains of Rhodotorula minuta, Candida azyma, and Aureobasidium pullulans showed a high antagonistic effect in vivo on Geotrichum citri-aurantii, a fungus that causes citrus sour rot. In this case, the three yeasts exerted this effect through three mechanisms simultaneously: killer activity, competition for nutrients, and production of extracellular enzymes such as β-1,3-glucanase (R. minuta) and chitinases (R. minuta and C. azyma), which are capable of breaking down the cell wall of pathogens [27]. Therefore, yeasts do not use just one mode of action to tackle their antagonists; commonly, several mechanisms come into action at the same time, exerting deleterious effects on pathogens [22,23]. Another species that also demonstrated antifungal potential against different pathogens was the yeast Pseudozyma graminicola, which made the survival of fungal cells unfeasible. This yeast produced a glycolipid containing cellobiose in the saccharide portion that acted as a fungicide [28].
The beneficial action of yeasts can also occur more discreetly, with these microorganisms acting as external sensors of plants, signaling the presence of pathogens. In this context, yeasts act by releasing compounds such as salicylic acid (SA), jasmonic acid (JA), and ethylene (ET). These substances induce the plant to activate its defense system, producing secondary compounds that act by repelling or hindering the attack of pathogens (Figure 1). This prevents or reduces damage to the plant. This phenomenon is known as induced systemic resistance (ISR) [28,29].
When challenged by the bacterium Xanthomonas axonopodis, Pseudozyma churashimaensis cells inoculated into pepper leaves (Capsicum annuum) caused a 4.5-fold increase in the expression of resistance inducers such as SA and JA. At the same time, ET production was 15 times higher than in plants not inoculated with the yeast [30]. It has also been reported that the presence of the yeast Meyerozyma (Pichia) guilliermondii in peach fruits resulted in increased activity of the enzymes glucanase and polyphenol oxidase, in addition to increased SA production, activating plant defense against the presence of the pathogens Rhizopus stolonifer (which causes soft rot and Penicillium expansum (which causes blue mold) [31].
Finally, soil composition is also affected by the presence of yeasts, as they can provide carbohydrates to the environment, as is the case of the yeasts Rhodotorula glutinis and Rhodotorula acheniorum, which were reported to provide mannose, galactose, glucose, and xylose [32,33]. In this way, yeasts also influence the soil's physical structure, making it more stable, maintaining the proportion between micro and macropores, and balancing spaces with air and satisfactory water storage for plants.
3. The Ecological Importance of Yeasts Beyond Soil
Above ground, yeasts can act in several ways. Among them, perhaps the most important and best known is their role in the pollination process of angiosperms. Because they are present in floral nectaries, they ferment nectar, releasing volatile organic compounds (VOCs) capable of attracting pollinating insects, thus creating a triple symbiosis: (a) the insect is attracted to a food source, (b) the plant benefits from the pollination, continuing its propagation, and (c) the microorganism gets access to other places not yet inhabited by it, or even survives within the gastric tract during periods when there is no flowering [3].
In the gastrointestinal tract of insects, yeasts also play an important ecological role. Once internalized, they interact with the microorganisms present there and act as probiotics. Thus, they help these invertebrates by making nutrients available (breaking down polymers, so that their monomers are more easily absorbed) or acting in their defense, producing antimicrobial substances and/or preventing the growth of pathogenic species within the guts [12,34,35,36].
Fruit fly larvae (Drosophila melanogaster) with their gastric system inoculated with Saccharomyces cerevisiae showed faster growth and better development, culminating in larger insects when compared to non-inoculated ones [37]. Similar to the effect on fly larvae, the presence of the yeast Yarrowia lipolytica in the digestive system of beetles of the species Nicrophorus vespilloides facilitated the metabolism of proteins and lipids, given the secretion, by the yeast cells, of proteases, lipases, and enzymes linked to the β-oxidation of fatty acids [34].
In plants, yeasts can also develop endophytic symbiosis, establishing themselves in intercellular spaces. In this case, microorganisms help the plant absorb nutrients such as iron, phosphorus, and zinc [38]. In this association, yeasts also increase the resistance of plants to pathogens, producing glycosylhydrolases (as already mentioned) and VOCs that act antagonistically to other microorganisms [22]. As an example, in tomato plants, Fernandez-San Millan et al. [39] demonstrated that the yeast Wickerhamomyces anomalus had a relevant antagonistic effect against pathogenic species such as Fusarium oxysporum, which causes fusarium wilt, and Verticillium dahliae, which causes verticillium wilt in tomatoes.
This antagonistic effect is not only seen in endophytic yeasts. Species present in floral nectaries compete with pathogenic organisms for physical space and nutrients found there [3]. In this microenvironment, they also produce and release substances with antimicrobial properties. It is interesting to note that this action not only protects plants but also pollinators, as it prevents the establishment of pathogens and their subsequent attack on insects that visit flowers [5].
Therefore, it is possible to infer that these yeasts' presence in agricultural environments can also provide these various ecological services, benefiting soil, plants, and the insects that pollinate them, enabling the balance between biodiversity and plant production. The alteration of the yeast population can cause deficiencies in the pollination of plant species and in the maintenance of insect life [5]. At the same time, the loss of yeast biodiversity can lead to a greater occurrence of attacks by pathogenic organisms on plants and invertebrates [36].
4. Impacts of Pesticides on Yeast Microbiota
After the Second World War, synthetic molecules began to be used in agricultural production fields in increasing quantities to protect and maintain crop health [40]. However, the action of these pesticides is not restricted to target organisms; they are mostly broad-spectrum products, also affecting non-target organisms and harming local biodiversity [41].
The repeated use of products with the same mechanism of action causes the development of resistance by target organisms; that is, these pathogens end up not being affected by the chemicals, resulting in the obsolescence of the molecules and the death of beneficial organisms, leaving the crop increasingly exposed to attack by resistant pathogens [42].
Many of the pesticides used in the field have systemic action, which means that they can penetrate the leaf tissue, passing through the plants' vascular tissues and reaching parts of the plants that were not directly exposed to the product, such as the floral nectary [41]. The presence of these molecules in such locations alters the biological dynamics of the yeasts present there, causing indirect problems related to pollinators' attraction, reduced crop productivity, and reduced food availability [5].
An example of this undesirable effect occurs with fungicides, which are used extensively in conventional agriculture to reduce the population of disease-causing fungi. However, these pesticides also cause a reduction in the population of non-target organisms [43]. Many of these affected organisms are desirable, especially in the post-harvest process, so that their loss causes problems not only to local biodiversity but also in the production chain that depends on the action of these organisms [44].
The impacts caused by these practices affect the industry, especially the fermented beverage sector. The residual persistence of molecules such as penconazole, benomyl [45], and pyrimethanil in grapes (in this case, the raw material for wine production) is widely recognized. Once present in grape must, pyrimethanil impairs the fermentation process and alters the rhythm of beverage production. This molecule inhibits the growth of the wild yeast Hanseniaspora uvarum, giving space for greater growth of the yeast S. cerevisiae, thus anticipating its participation in the anaerobic fermentation of the must, which affects the organoleptic characteristics of the product [46].
Similarly, the cell growth of yeasts present in wheat grains is highly affected by the use of fungicides, directly impacting the balance among native microorganisms, such as Aureobasidium pullulans, Candida albicans, Candida sake, Debaryomyces hansenii, Candida famata, Metschnikowia (Candida) pulcherrima, and Rhodotorula glutinis, which are present on the external and internal part of the grains. This compromises the rheological and organoleptic characteristics of bakery products produced from wheat grains with the presence of native yeasts [44].
Besides fungicides, other molecules widely used in the global agricultural environment also impact yeasts, whether they are at recommended concentrations or even in residues below the limit permitted by law [47]. In 2022, around 2,000 tons of herbicides were used in agriculture worldwide. Brazil was the country that used such synthetic products the most in the same year, consuming almost 500 tons of herbicides in its agricultural area [40].
Products based on the molecule 2,4-dichlorophenoxyacetic acid (2,4-D) are extremely useful in conventional agriculture because they are selective for plants, causing damage only to dicotyledonous [48]. However, 2,4-D has harmful effects beyond the Kingdom Plantae. It has a high affinity with the cell membrane, which facilitates its entry into yeast cells and causes their inviability [47]. Yeast cells exposed to this molecule, even at doses recommended for crops, have their intracellular pH acidified. This also triggers oxidative stress in the cells, which inhibits cell growth and increases their latency period [47,49]. The oxidative stress intensity varies according to the amount of the active ingredient (AI) administered. Yeast cells exposed to increasing doses of AI showed the formation of hydroxyl radicals; during the adaptation period, S. cerevisiae exposed to the herbicide showed an increase in the enzymatic activity of cytosolic catalase (Cttp1), CuZn-superoxide (Sod1p), glutathione-dithiols, and glutathione reductoxins (Grx1p and Grx2p), evidencing an increase in the antioxidant defense system in yeast cells [50].
Another herbicide widely used in conventional agriculture, glyphosate also demonstrates a highly inhibitory effect on yeast cell growth [51], as it inhibits the enzyme 5-enolpyruvylshikimate-3-phosphate synthase (EPSP synthase), making the shikimate pathway unfeasible [48]. Consequently, this last compound accumulates, preventing the synthesis of the aromatic amino acids tryptophan, phenylalanine, and tyrosine, precursors of compounds essential to yeast cells [52].
Faced with the adversities humans impose on rural-environment-dwelling yeasts, these microorganisms naturally seek to adapt and coexist with the presence of the stress source, thus developing resistance to the active ingredients [50]. The consecutive and indiscriminate use of these products in agriculture leads to an artificial selection of resistant organisms [52]. This has been demonstrated by Barney et al. [51], who compared the glyphosate effect on yeast strains isolated from agricultural environments before the commercial launch of glyphosate with the glyphosate effect on strains isolated after the commercialization and use of the herbicide. Strains never exposed to glyphosate demonstrated greater susceptibility to the product, while strains previously exposed to the herbicide demonstrated greater resistance.
In addition to the active ingredients, other substances, called “inerts”, are added to the commercial product to improve the effect of the AI present therein. However, these inerts are also responsible for negative effects on yeast growth [53]. The deleterious effect of inerts was demonstrated in an environment where yeasts were not dependent on the shikimate pathway to carry out cellular respiration; however, in this case, the inhibition performed by the commercial product occurred equally [54].
5. Yeasts as Potential Bioremediators
As previously mentioned, yeasts not only play several roles in the industry but also serve several essential ecological functions. From this point on, we also highlight the potential of these microorganisms in bioremediation, which is a crucial process for developing sustainable agriculture [55]. Bioremediation can be seen as the removal and degradation of chemical contaminants with variable chemical structures and, therefore, requires specific biochemical processes for their degradation [56].
The chemical structures of the compounds discussed below are presented in Figure 3. Taking 2,4-D as our starting point, it is important to highlight that the constant application of this herbicide generates major toxicological problems for the environment and resistance in weeds. A study revealed that, in addition to resistant plants, yeasts may also be resistant and adapted to 2,4-D, having acquired a plasma membrane that is not disrupted by the herbicide. The cells of these yeasts grow in the presence of different concentrations of the herbicide. These adapted yeasts display an increase in the proportion of saturated and monounsaturated fatty acids in their membranes. Yeasts do that by decreasing the expression of the gene responsible for encoding a fatty acid desaturase. As a result, the entry of the herbicide into the cell is hindered, allowing the correct maintenance of cellular activities [47,57]. Increased expression of other genes linked to maintaining cell integrity has also been detected as a tolerance strategy in yeast. Upregulation of genes such as MTL1, ROM1, and MKK2, involved in signaling stress in the cell wall, and genes linked to the synthesis of chitin (a component of the cell wall), such as SHC1 and ECM38, supports the importance of seeking to maintain and restore cell wall integrity in tolerance to 2,4-D [58].
The degradation of 2,4-D involves different metabolic pathways. While most bacteria degrade 2,4-D by initial cleavage of the side chain, resulting in simpler compounds, fungi predominantly utilize hydroxylation of the aromatic ring. Monooxygenase enzymes, specifically hydroxylases, introduce hydroxyl groups (–OH) into the aromatic ring, forming metabolites such as 2,4-dichloro-5-hydroxyphenoxyacetic acid and 2,5-dichloro-4-hydroxyphenoxyacetic acid. Furthermore, hydroxylation of 2-chlorophenoxyacetic acid (2-CPA) results in 2-chloro-4-hydroxyphenoxyacetic and 2-hydroxyphenoxyacetic acids, indicating dichlorination processes and replacement of chlorine by hydroxyl groups. These enzymatic processes and their resulting metabolites are crucial to understanding and optimizing the bioremediation of herbicide-contaminated environments [29,60].
As the most widely used herbicide in conventional agriculture, glyphosate also deserves attention (see its structure in Figure 3). As previously mentioned, glyphosate, by inhibiting the EPSP synthase enzyme, interrupts the synthesis of amino acids that are precursors of important compounds, such as alkaloids, flavonoids, and benzoic acids. Microbial degradation of glyphosate in the soil can occur through two main routes. The first involves the conversion of glyphosate into sarcosine by the action of the bacteria Agrobacterium radiobacter and Enterobacter aerogenes, using the enzyme C-P lyase. Then, sarcosine can be metabolized by other microorganisms, including yeasts. The second-most common route transforms glyphosate into amino-methyl phosphonic acid (AMPA) [61]. In this case, though, glyoxylate is also one of the products, being thus the preference route in the Kingdom Fungi [62].
In addition to herbicides, studies on the biodegradation of other agrochemicals, such as insecticides, which are widely used to combat insect pests, are also relevant. In this context, carbofuran is also worth mentioning (see its structure in Figure 3). It is a broad-spectrum agrochemical belonging to the carbamate class, used as an acaricide, insecticide, and nematicide. Commercially introduced in 1967, its specific use increased rapidly in the following years. The half-life of this pesticide in soil varies between thirty and one hundred and twenty days, depending on the soil type. In sandy soils, the half-life is approximately thirty days. In clayey soils, the half-life is intermediate, around forty days. In muddy soils, the half-life of carbofuran is approximately eighty days [63].
The yeasts Candida tropicalis and Trichosporon cutaneum have already been tested for tolerance and degradation capacity to carbofuran and glyphosate. Both species showed good growth in medium containing glyphosate as the sole carbon source. The yeast C. tropicalis degraded 76% of the initial glyphosate in 192 hours, standing out for its biodegradation efficiency. Both yeasts also grew normally in rich medium (YEPD) with carbofuran, but the growth of T. cutaneum slowed significantly in synthetic minimal medium (YNB without amino acids) containing carbofuran above 0.3 g/L. In any case, this yeast demonstrated almost complete biodegradation of carbofuran in 192 hours, with the detection of intermediate metabolites such as carbofuran-7-phenol and pyruvate during cultivation [56].
Another widely used herbicide is pendimethalin (see its structural formula in Figure 3). Its unique combination of atoms allows it to be easily adsorbed by the soil but very difficult to desorb. However, fortunately, some fungi can oxidize its amine groups and its benzene ring thanks to their oxygenases and peroxidases, such as pendimethalin monooxygenase and pendimethalin peroxidase. These oxidative processes make this molecular structure nontoxic and more easily degraded by the soil microbial community. The degradation of pendimethalin can also be caused by microbial esterases such as pendimethalin hydrolase [64]. Regarding this herbicide, the yeast Clavispora lusitaniae deserves special mention. Han et al. [65] demonstrated that the ability of this yeast to degrade pendimethalin is inversely correlated with the medium's pH: the yeast's efficiency increases as the pH of the liquid medium decreases.
Yeast has also been successful in degrading the herbicide atrazine (see its structural formula in Figure 3), a member of the triazine class. A Pichia kudriavzevii strain was shown to degrade this herbicide both in a liquid medium and in the soil. The study demonstrated that the strain Atz-EN-01 was able to completely degrade atrazine in 7 days, with a degradation constant of 31% per day, following the first-order kinetic model. The half-life of the process was 2.2 days under optimum conditions of pH 7, 30°C, inoculum size of 3% (v/v), and agitation of 120 rpm. The degradation products identified were hydroxyatrazine, N-isopropylammelide, and cyanic acid, with the enzyme atrazine chlorohydrolase exhibiting maximum activity during degradation [66].
It is interesting to note that some yeast species, unlike most living beings, have the ability to switch between respiration and fermentation regardless of the presence of oxygen. This switching depends on the availability of a carbon source and/or the need imposed by the environment. This makes them less susceptible to agrochemicals of the dinitrophenol class, which are recognized as uncoupling agents of oxidative phosphorylation and, therefore, inhibit the generation of ATP after the respiratory chain. Thus, the resistance of yeasts to these toxic conditions represents a significant advantage in the biodegradation process of dinitrophenols, as detailed in the study by Marius et al. [67]. These researchers analyzed the use of yeasts for biodegradation, specifically in solutions of agrochemicals based on dinitrophenol. The results showed that, after treatment with yeasts, there was a decrease in the toxicity of the remaining solutions, which were then used in wheat seed germination experiments.
When considering the possibility of using yeasts as bioremediators, the origin of the microbial cells or the cost involved in producing cells to treat a large contaminated area may be questioned. However, an alternative to overcome this potential problem may be found in the study by Szpyrka et al. [2]. These authors tested three commercial yeast strains (of the species Saccharomyces cerevisiae, Yarrowia lipolytica, and Debaryomyces hansenii) against four herbicides and demonstrated that yeasts such as those (primarily used for industrial purposes) can also carry out additional tasks. Among the pesticides tested, fluazifop-P-butyl was the most efficiently degraded by the yeasts (up to 71.2% degradation after four days in the presence of the microbial cells), followed by metribuzin (20%), propyzamide (13.4%) and pendimethalin (5.3%). Moreover, when shell pea (Pisum sativum L.) seeds were planted in the same soils, yeasts also contributed to plant development: they increased plant growth by 22% and the germination capacity of the seeds by 30%. Therefore, it is possible to assume that, after their use in industry, yeast cell biomasses could still play an additional role, degrading compounds harmful to the environment and stimulating plant growth and germination.
In the face of the several examples we mentioned above, one can say that the scientific community has already accumulated enough knowledge to put into practice the use of yeasts as pesticide-degradation machines. However, the successful experiences with yeasts on bioremediation go beyond the employment of wild yeasts. In the following section, we highlight other important advances that have already been obtained with genetically modified strains.
6. Engineering Yeasts for Pesticide Degradation
The first eukaryote to have its genome sequenced was a yeast — the strain S. cerevisiae S288c. This demonstrates the scientific community's interest in these fast-growing microorganisms, which can be cultivated in simple and inexpensive culture media. In fact, yeasts have been successfully used for decades in many molecular studies and as biofactories generated from gene editing techniques [68]. Using genetic engineering tools, yeasts can, therefore, be modified to perform numerous functions, such as removing toxic waste. In this context, they can act as biocatalysts, hosting different degradation enzymes in their cells, or as biosensors to detect the presence of toxic waste in a given environment. Furthermore, although the triumphant story of yeast genetic engineering is already over 40 years old, more recently, with the development of CRISPR (clustered regularly interspaced short palindromic repeats), the functional capacity of yeasts took a new great leap, further increasing the potential of these microorganisms as bioremediators of areas affected by pesticides [69].
One successful example is related to the CYP72A18 gene, present in rice, which encodes an enzyme of the cytochrome P450 superfamily that catalyzes the (ω-1)-hydroxylation of the herbicide pelargonic acid, reducing its toxicity in the environment. Through genetic engineering, this gene was heterologously expressed in yeast cells, which then began to degrade this pesticide [70]. Still in the context of cytochrome, even human isoforms of P450 have been tested in yeast with the aim of degrading agrochemicals. In one of these tests, the CYP 1A1 and CYP 1A2 isoforms showed expressive results against the herbicides chlortoluron and atrazine. These enzymes were tested alone and fused to the NADPH-cytochrome P450 oxidoreductase of the transformed yeast strain. Interestingly, after fusion, enzymatic activities were increased [71].
Other genes encoding enzymes related to cytochrome P450 have also been expressed in yeast to induce herbicide degradation. This is the case of the monooxygenase CYP71A12 from Arabidopsis thaliana in yeast, which has shown promise in metabolizing the herbicide pyrazoxyfen. Hayashi et al. [72] showed that this enzyme, when expressed in S. cerevisiae, catalyzes the transformation of this agrochemical into less toxic metabolites that are more easily degradable by other microorganisms in the environment. They result from N-demethylation reactions on the pyrazole ring and hydroxylation on the dichlorobenzene ring of pyrazoxyfen. N-demethylation removes a methyl group (CH3) from the pyrazole ring, while hydroxylation adds a hydroxyl group (OH) to the dichlorobenzene ring.
As in the cases above, it is interesting to note that enzymes from plants of agronomic interest can be expressed in yeast to mitigate the toxic effects of pesticides on the environment. For glyphosate, for example, it was found that the overexpression of two glutathione-S-transferases from a tea plant (Camellia sinensis) in S. cerevisiae allowed the cells to grow efficiently even at a concentration of 1 g/L of the herbicide [73]. Given the advantages of using yeast as biofactories (as pointed out before), the heterologous production of these enzymes in these microbial cells can substantially increase their use as catalysts for the degradation of agrochemicals.
In the context of engineered yeasts for bioremediation, a successful strategy is the expression of degradation enzymes on these microorganisms' cell surfaces (Figure 4). This allows the yeasts to degrade compounds with no need to internalize them, which is particularly efficient in the case of substances with high molecular weight. The so-called whole-cell yeast biocatalysts also prove to be efficient by removing the transportation barrier across the plasma membrane and dismissing any enzyme preparation and purification steps, thereby reducing the cost. Besides, with the cell-surface display strategy, biocatalyst recycling for processes is also increased [69,74,75].
The cell-surface strategy has been notedly studied for the degradation of organophosphorus pesticides. Takayama et al. [76] succeeded in expressing up to 14x104 molecules of an organophosphorus hydrolase (OPH) from Flavobacterium sp. on the cell surface of S. cerevisiae. To anchor this enzyme to the yeast surface, the authors fused an α-agglutinin with a glycosylphosphatidylinositol (GPI) signal sequence to the OPH’s C-terminal region, allowing the cells to efficiently hydrolyze paraoxon. The same research group has also expressed this OPH in S. cerevisiae through the Flo1p anchor system. In this case, the authors attached the anchor protein to OPH’s N-terminal region and achieved eight times higher OPH activity in comparison to the GPI approach [77]. In any case, though, they found significantly higher activities with these yeast-surface display methods than with a similar strategy that employed bacterial cells instead [76,77]. Indeed, as eukaryotes, yeasts harbor more sophisticated pathways for secreting or displaying proteins on their surfaces than bacteria (prokaryotes) [75].
7. Concluding Remarks: Use of Yeasts from a Sustainable Agriculture Perspective
Yeasts have been used in food and beverage production since the Neolithic Revolution. Thus, these microorganisms have been undergoing selection and adaptation within the agricultural environment for millennia [68]. Furthermore, as we pointed out in this review, yeasts can benefit the health and development of plants in several ways, either by defending them from pathogens or stimulating their growth. Therefore, these microorganisms could be preferentially used instead of filamentous fungi or bacteria in an organic or agroecological agriculture context. Also, in this scenario, we can include yeasts' potential for bioremediation of areas previously contaminated by conventional pesticides. In this case, yeasts would initially play a “cleaning” role and then promote the cultivation of new crops from an organic perspective.
Fortunately, this premise has been gaining strength in recent years. Through the bibliometric search we conducted in the Scopus database, we observed that the number of studies on the use of yeast in agriculture (including the role of bioremediation) has accelerated significantly in the last twenty years (Figure 5). This search used the terms “yeast AND agriculture AND bioremediation” to identify articles published between 2004-2024.
The trend observed in Figure 5 reflects the growing concern for sustainability and the pursuit of alternatives that reduce the negative environmental impacts of conventional agriculture. In this quest, yeasts deserve a prominent place, either due to the innate potential of the species found in nature or the biotechnological potential behind the genetic engineering of these microorganisms. In this review, these potentials were systematized with the aim of awakening the scientific community's interest in developing processes that can put into practice what was discussed here. It is imperative to pay attention to the damage that agrochemical use inflicts on the environmental microbiota and how this can impact wildlife as a whole. At the same time, it should be noted that the microorganisms we need to preserve may help us solve the problems humanity has caused. In this two-way street, new studies on yeast physiological processes are highly desired.
Author Contributions
Conceptualization, S.L.A.J.; writing—original draft preparation, E.J.P.P., D.S., C.G.O., A.F.C., A.A.S; writing—review and editing, L.C.C., A.J.M., H.T., S.L.A.J.; supervision, H.T., S.L.A.J.; project administration, H.T., S.L.A.J.. All authors have read and agreed to the published version of the manuscript.
Funding
This work is part of the National Institute of Science and Technology (INCT) “Yeasts: Biodiversity, preservation, and biotechnological innovation”. It is supported by grants and fellowships from the Brazilian National Council for Scientific and Technological Development (CNPq, grant numbers 302484/2022-1, 150719/2023-0, and 308830/2023-7), the Brazilian Coordination for the Improvement of Higher Education Personnel (CAPES), the Research and Innovation Funding Agency of the State of Santa Catarina (FAPESC, grant number 2023TR000234), the Research Support Foundation of Rio Grande do Sul (FAPERGS, grant number 22/2551-0000397-4), and the Research Promotion Program from the Federal University of Fronteira Sul (UFFS, grant number PES-2023-0349).
Data Availability Statement
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Silva, V.; Mol, H.G.J.; Zomer, P.; Tienstra, M.; Ritsema, C.J.; Geissen, V. Pesticide Residues in European Agricultural Soils – A Hidden Reality Unfolded. Science of The Total Environment 2019, 653, 1532–1545. [CrossRef]
- Szpyrka, E.; Książek-Trela, P.; Bielak, E.; Słowik-Borowiec, M. The Influence of Commercial Yeast Preparations on the Degradation of Herbicide Mixtures in the Soil and the Effect on the Shell Pea (Pisum Sativum L.) Cultivation. J Soil Sci Plant Nutr 2024, 24, 2509–2519. [CrossRef]
- Fenner, E.D.; Scapini, T.; da Costa Diniz, M.; Giehl, A.; Treichel, H.; Álvarez-Pérez, S.; Alves, S.L. Nature’s Most Fruitful Threesome: The Relationship between Yeasts, Insects, and Angiosperms. Journal of Fungi 2022, 8, 984. [CrossRef]
- Giehl, A.; dos Santos, A.A.; Cadamuro, R.D.; Tadioto, V.; Guterres, I.Z.; Prá Zuchi, I.D.; Minussi, G. do A.; Fongaro, G.; Silva, I.T.; Alves, S.L. Biochemical and Biotechnological Insights into Fungus-Plant Interactions for Enhanced Sustainable Agricultural and Industrial Processes. Plants 2023, 12, 2688. [CrossRef]
- Schaeffer, R.N.; Mei, Y.Z.; Andicoechea, J.; Manson, J.S.; Irwin, R.E. Consequences of a Nectar Yeast for Pollinator Preference and Performance. Funct Ecol 2017, 31, 613–621. [CrossRef]
- Anderson, T.-H. Microbial Eco-Physiological Indicators to Asses Soil Quality. Agric Ecosyst Environ 2003, 98, 285–293. [CrossRef]
- Santos, H.G. dos. Sistema Brasileiro de Classificação de Solos; Embrapa, 2018; ISBN 9788570358004. Available online: https://www.embrapa.br/busca-de-publicacoes/-/publicacao/1094003/sistema-brasileiro-de-classificacao-de-solos (accessed on 10/09/2024).
- Moreira, G.A.M. Diversidade e Ecologia de Leveduras em Solos Brasileiros; Tese de doutorado apresentada ao Programa de Pós-Graduação de Biologia Microbiana do Instituto de Ciências Biológicas, Universidade de Brasília – UnB. 2019.
- Álvarez-Pérez, S. Ecology: Yeasts on Their Natural Environment. In Yeasts: From Nature to Bioprocesses; Bentaham Science, 2022; pp. 27–57.
- Fernandez-San Millan, A.; Farran, I.; Larraya, L.; Ancin, M.; Arregui, L.M.; Veramendi, J. Plant Growth-Promoting Traits of Yeasts Isolated from Spanish Vineyards: Benefits for Seedling Development. Microbiol Res 2020, 237, 126480. [CrossRef]
- Gomes, F.C.O.; Safar, S.V.B.; Marques, A.R.; Medeiros, A.O.; Santos, A.R.O.; Carvalho, C.; Lachance, M.-A.; Sampaio, J.P.; Rosa, C.A. The Diversity and Extracellular Enzymatic Activities of Yeasts Isolated from Water Tanks of Vriesea Minarum, an Endangered Bromeliad Species in Brazil, and the Description of Occultifur Brasiliensis f.a., Sp. Nov. Antonie Van Leeuwenhoek 2015, 107, 597–611. [CrossRef]
- Bright, J.P.; Karunanadham, K.; Maheshwari, H.S.; Karuppiah, E.A.A.; Thankappan, S.; Nataraj, R.; Pandian, D.; Ameen, F.; Poczai, P.; Sayyed, R.Z. Seed-Borne Probiotic Yeasts Foster Plant Growth and Elicit Health Protection in Black Gram (Vigna Mungo L.). Sustainability 2022, 14, 4618. [CrossRef]
- Sarabia, M.; Jakobsen, I.; Grønlund, M.; Carreon-Abud, Y.; Larsen, J. Rhizosphere Yeasts Improve P Uptake of a Maize Arbuscular Mycorrhizal Association. Applied Soil Ecology 2018, 125, 18–25. [CrossRef]
- Marques, A.R.; Resende, A.A.; Gomes, F.C.O.; Santos, A.R.O.; Rosa, C.A.; Duarte, A.A.; de Lemos-Filho, J.P.; dos Santos, V.L. Plant Growth–Promoting Traits of Yeasts Isolated from the Tank Bromeliad Vriesea Minarum L.B. Smith and the Effectiveness of Carlosrosaea Vrieseae for Promoting Bromeliad Growth. Brazilian Journal of Microbiology 2021, 52, 1417–1429. [CrossRef]
- Braus, G.H. Aromatic Amino Acid Biosynthesis in the Yeast Saccharomyces Cerevisiae: A Model System for the Regulation of a Eukaryotic Biosynthetic Pathway. Microbiol Rev 1991, 55, 349–370. [CrossRef]
- Gómez-Arreaza, A.; Acosta, H.; Quiñones, W.; Concepción, J.L.; Michels, P.A.M.; Avilán, L. Extracellular Functions of Glycolytic Enzymes of Parasites: Unpredicted Use of Ancient Proteins. Mol Biochem Parasitol 2014, 193, 75–81. [CrossRef]
- Lee, J.W.; Yook, S.; Koh, H.; Rao, C. V; Jin, Y.-S. Engineering Xylose Metabolism in Yeasts to Produce Biofuels and Chemicals. Curr Opin Biotechnol 2021, 67, 15–25. [CrossRef]
- Rao, R.P.; Hunter, A.; Kashpur, O.; Normanly, J. Aberrant Synthesis of Indole-3-Acetic Acid in Saccharomyces Cerevisiae Triggers Morphogenic Transition, a Virulence Trait of Pathogenic Fungi. Genetics 2010, 185, 211–220. [CrossRef]
- Toyn, J.H.; Gunyuzlu, P.; White, W.H.; Thompson, L.A.; Hollis, G.F. A Counterselection for the Tryptophan Pathway in Yeast: 5-Fluoroanthranilic Acid Resistance. Yeast 2000, 16, 553–560. [CrossRef]
- Tzin, V.; Galili, G. The Biosynthetic Pathways for Shikimate and Aromatic Amino Acids in Arabidopsis Thaliana. Arabidopsis Book 2010, 8, e0132. [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The Rhizosphere Microbiome: Significance of Plant Beneficial, Plant Pathogenic, and Human Pathogenic Microorganisms. FEMS Microbiol Rev 2013, 37, 634–663. [CrossRef]
- Khunnamwong, P.; Lertwattanasakul, N.; Jindamorakot, S.; Suwannarach, N.; Matsui, K.; Limtong, S. Evaluation of Antagonistic Activity and Mechanisms of Endophytic Yeasts against Pathogenic Fungi Causing Economic Crop Diseases. Folia Microbiol (Praha) 2020, 65, 573–590. [CrossRef]
- Chen, O.; Yi, L.; Deng, L.; Ruan, C.; Zeng, K. Screening Antagonistic Yeasts against Citrus Green Mold and the Possible Biocontrol Mechanisms of Pichia Galeiformis (BAF03). J Sci Food Agric 2020, 100, 3812–3821. [CrossRef]
- Cabañas, C.M.; Hernández, A.; Martínez, A.; Tejero, P.; Vázquez-Hernández, M.; Martín, A.; Ruiz-Moyano, S. Control of Penicillium Glabrum by Indigenous Antagonistic Yeast from Vineyards. Foods 2020, 9, 1864. [CrossRef]
- Abdel-Kareem, M.Marwa.; Zohri, A.-N.A.; Nasr, S.A.E.E. Novel Marine Yeast Strains as Plant Growth-Promoting Agents Improve Defense in Wheat (Triticum Aestivum) against Fusarium Oxysporum. Journal of Plant Diseases and Protection 2021, 128, 973–988. [CrossRef]
- Lara-Capistran, L.; Zulueta-Rodriguez, R.; Castellanos-Cervantes, T.; Reyes-Perez, J.J.; Preciado-Rangel, P.; Hernandez-Montiel, L.G. Efficiency of Marine Bacteria and Yeasts on the Biocontrol Activity of Pythium Ultimum in Ancho-Type Pepper Seedlings. Agronomy 2020, 10, 408. [CrossRef]
- Ferraz, L.P.; Cunha, T. da; da Silva, A.C.; Kupper, K.C. Biocontrol Ability and Putative Mode of Action of Yeasts against Geotrichum Citri-Aurantii in Citrus Fruit. Microbiol Res 2016, 188–189, 72–79. [CrossRef]
- Golubev, W.I.; Kulakovskaya, T. V.; Shashkov, A.S.; Kulakovskaya, E. V.; Golubev, N. V. Antifungal Cellobiose Lipid Secreted by the Epiphytic Yeast Pseudozyma Graminicola. Microbiology (N Y) 2008, 77, 171–175. [CrossRef]
- Silva, T.M.; Stets, M.I.; Mazzetto, A.M.; Andrade, F.D.; Pileggi, S.A. V.; Fávero, P.R.; Cantú, M.D.; Carrilho, E.; Carneiro, P.I.B.; Pileggi, M. Degradation of 2,4-D Herbicide by Microorganisms Isolated from Brazilian Contaminated Soil. Brazilian Journal of Microbiology 2007, 38, 522–525. [CrossRef]
- Lee, G.; Lee, S.-H.; Kim, K.M.; Ryu, C.-M. Foliar Application of the Leaf-Colonizing Yeast Pseudozyma Churashimaensis Elicits Systemic Defense of Pepper against Bacterial and Viral Pathogens. Sci Rep 2017, 7, 39432. [CrossRef]
- Zhao, Y.; Li, Y.; Zhang, B. Induced Resistance in Peach Fruit as Treated by Pichia Guilliermondii and Their Possible Mechanism. Int J Food Prop 2020, 23, 34–51. [CrossRef]
- Simova, E.D.; Frengova, G.I.; Beshkova, D.M. Synthesis of Mannose-Rich Exopolysaccharide by Rhodotorula Glutinis 16P Co-Cultured with Yeast or Bacteria. Zeitschrift für Naturforschung C 2000, 55, 540–545. [CrossRef]
- Grigorova, D.; Simova, E.; Pavlova, K.; Frengova, G.; Beshkova, D. Polysaccharides Production by Yeast in Whey Ultrafiltrate. Biotechnology & Biotechnological Equipment 1994, 8, 31–37. [CrossRef]
- Vogel, H.; Shukla, S.P.; Engl, T.; Weiss, B.; Fischer, R.; Steiger, S.; Heckel, D.G.; Kaltenpoth, M.; Vilcinskas, A. The Digestive and Defensive Basis of Carcass Utilization by the Burying Beetle and Its Microbiota. Nat Commun 2017, 8, 15186. [CrossRef]
- Varotto Boccazzi, I.; Ottoboni, M.; Martin, E.; Comandatore, F.; Vallone, L.; Spranghers, T.; Eeckhout, M.; Mereghetti, V.; Pinotti, L.; Epis, S. A Survey of the Mycobiota Associated with Larvae of the Black Soldier Fly (Hermetia Illucens) Reared for Feed Production. PLoS One 2017, 12, e0182533. [CrossRef]
- Al Naggar, Y.; Singavarapu, B.; Paxton, R.J.; Wubet, T. Bees under Interactive Stressors: The Novel Insecticides Flupyradifurone and Sulfoxaflor along with the Fungicide Azoxystrobin Disrupt the Gut Microbiota of Honey Bees and Increase Opportunistic Bacterial Pathogens. Science of The Total Environment 2022, 849, 157941. [CrossRef]
- Qiao, H.; Keesey, I.W.; Hansson, B.S.; Knaden, M. Gut Microbiota Affects Development and Olfactory Behavior in Drosophila Melanogaster. Journal of Experimental Biology 2019. [CrossRef]
- de Lima Targino, H.M.; Silva, V.S.L.; Escobar, I.E.C.; Ribeiro, P.R. de A.; Gava, C.A.T.; Fernandes-Júnior, P.I. Maize-Associated Meyerozyma from the Brazilian Semiarid Region Are Effective Plant Growth-Promoting Yeasts. Rhizosphere 2022, 22, 100538. [CrossRef]
- Fernandez-San Millan, A.; Larraya, L.; Farran, I.; Ancin, M.; Veramendi, J. Successful Biocontrol of Major Postharvest and Soil-Borne Plant Pathogenic Fungi by Antagonistic Yeasts. Biological Control 2021, 160, 104683. [CrossRef]
- FAO. FAOSAT. Available online: https://www.fao.org/faostat/en/#home (accessed on 15/09/2024).
- Goulart, A.C.P.; Nunes, J.C. da S. Evolução e Cenário Atual Do Tratamento de Sementes de Soja Com Fungicidas No Brasil. Revista Cultivar:Embrapa 2021.
- International Herbicide-Resistant Weed Database Most Recent Cases of Herbicide Resistant Weeds Entered into the Database.
- Guo, L.; Li, R.; Chen, W.; Dong, F.; Zheng, Y.; Li, Y. The Interaction Effects of Pesticides with Saccharomyces Cerevisiae and Their Fate during Wine-Making Process. Chemosphere 2023, 328, 138577. [CrossRef]
- Wachowska, U.; Irzykowski, W.; Jędryczka, M. Agrochemicals: Effect on Genetic Resistance in Yeasts Colonizing Winter Wheat Kernels. Ecotoxicol Environ Saf 2018, 162, 77–84. [CrossRef]
- Jawich, D.; Hilan, C.; Saliba, R.; Lteif, R.; Strehaiano, P. Effects of Some Pesticides on Two Yeast Strains <Em>Saccharomyces Cerevisiae</Em> and <Em>Metschnikowia Pulcherrima</Em> OENO One 2005, 39, 67. [CrossRef]
- Čuš, F.; Raspor, P. The Effect of Pyrimethanil on the Growth of Wine Yeasts. Lett Appl Microbiol 2008, 47, 54–59. [CrossRef]
- Viegas, C.A.; Cabral, M.G.; Teixeira, M.C.; Neumann, G.; Heipieper, H.J.; Sá-Correia, I. Yeast Adaptation to 2,4-Dichlorophenoxyacetic Acid Involves Increased Membrane Fatty Acid Saturation Degree and Decreased OLE1 Transcription. Biochem Biophys Res Commun 2005, 330, 271–278. [CrossRef]
- Marchi, G.; Carvalho, E.; Tadeu, S.M.; Guimarães, G. Documentos 227 Herbicidas: Mecanismos de Ação e Uso Empresa Brasileira de Pesquisa Agropecuária Embrapa Cerrados Ministério Da Agricultura, Pecuária e Abastecimento. 2008.
- Cabral, M.G.; Viegas, C.A.; Teixeira, M.C.; Sá-Correia, I. Toxicity of Chlorinated Phenoxyacetic Acid Herbicides in the Experimental Eukaryotic Model Saccharomyces Cerevisiae: Role of PH and of Growth Phase and Size of the Yeast Cell Population. Chemosphere 2003, 51, 47–54. [CrossRef]
- Teixeira, M.C.; Telo, J.P.; Duarte, N.F.; Sá-Correia, I. The Herbicide 2,4-Dichlorophenoxyacetic Acid Induces the Generation of Free-Radicals and Associated Oxidative Stress Responses in Yeast. Biochem Biophys Res Commun 2004, 324, 1101–1107. [CrossRef]
- Barney, J.B.; Winans, M.J.; Blackwood, C.B.; Pupo, A.; Gallagher, J.E.G. The Yeast Atlas of Appalachia: Species and Phenotypic Diversity of Herbicide Resistance in Wild Yeast. Diversity (Basel) 2020, 12, 139. [CrossRef]
- Van Bruggen, A.H.C.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G. Environmental and Health Effects of the Herbicide Glyphosate. Science of The Total Environment 2018, 616–617, 255–268. [CrossRef]
- Braconi, D.; Possenti, S.; Laschi, M.; Geminiani, M.; Lusini, P.; Bernardini, G.; Santucci, A. Oxidative Damage Mediated by Herbicides on Yeast Cells. J Agric Food Chem 2008, 56, 3836–3845. [CrossRef]
- Ravishankar, A.; Cumming, J.R.; Gallagher, J.E.G. Mitochondrial Metabolism Is Central for Response and Resistance of Saccharomyces Cerevisiae to Exposure to a Glyphosate-Based Herbicide. Environmental Pollution 2020, 262, 114359. [CrossRef]
- Lescano, M.; Fussoni, N.; Vidal, E.; Zalazar, C. Biodegradation of Pesticide-Contaminated Wastewaters from a Formulation Plant Employing a Pilot Scale Biobed. Science of The Total Environment 2022, 807, 150758. [CrossRef]
- Stoyanova, K.; Gerginova, M.; Peneva, N.; Dincheva, I.; Alexieva, Z. Biodegradation and Utilization of the Pesticides Glyphosate and Carbofuran by Two Yeast Strains. Processes 2023, 11, 3343. [CrossRef]
- Gil, F.N.; Gonçalves, A.C.; Becker, J.D.; Viegas, C.A. Comparative Analysis of Transcriptomic Responses to Sub-Lethal Levels of Six Environmentally Relevant Pesticides in Saccharomyces Cerevisiae. Ecotoxicology 2018, 27, 871–889. [CrossRef]
- Teixeira, M.C.; Fernandes, A.R.; Mira, N.P.; Becker, J.D.; Sá-Correia, I. Early Transcriptional Response of Saccharomyces Cerevisiae to Stress Imposed by the Herbicide 2,4-Dichlorophenoxyacetic Acid. FEMS Yeast Res 2006, 6, 230–248. [CrossRef]
- 59. National Library of Medicine Pub Chem: Explore Chemistry. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 15/09/2024).
- Kumar, A.; Trefault, N.; Olaniran, A.O. Microbial Degradation of 2,4-Dichlorophenoxyacetic Acid: Insight into the Enzymes and Catabolic Genes Involved, Their Regulation and Biotechnological Implications. Crit Rev Microbiol 2014, 1–15. [CrossRef]
- Patriarcheas, D.; Momtareen, T.; Gallagher, J.E.G. Yeast of Eden: Microbial Resistance to Glyphosate from a Yeast Perspective. Curr Genet 2023, 69, 203–212. [CrossRef]
- Chen, Y.; Chen, W.-J.; Huang, Y.; Li, J.; Zhong, J.; Zhang, W.; Zou, Y.; Mishra, S.; Bhatt, P.; Chen, S. Insights into the Microbial Degradation and Resistance Mechanisms of Glyphosate. Environ Res 2022, 215, 114153. [CrossRef]
- Ariffin, F.; Rahman, S.A. Biodegradation of Carbofuran; A Review. Journal of Environmental Microbiology and Toxicology 2020, 8, 50–57. [CrossRef]
- Kocárek, M.; Artikov, H.; Vorisek, K.; Boruvka, L. Pendimethalin Degradation in Soil and Its Interaction with Soil Microorganisms. Soil and Water Research 2016, 11, 213–219. [CrossRef]
- Han, Y.; Tang, Z.; Bao, H.; Wu, D.; Deng, X.; Guo, G.; Ye, B.-C.; Dai, B. Degradation of Pendimethalin by the Yeast YC2 and Determination of Its Two Main Metabolites. RSC Adv 2019, 9, 491–497. [CrossRef]
- Abigail, E.A.; Abdul Salam, J.; Das, N. Atrazine Degradation in Liquid Culture and Soil by a Novel Yeast Pichia Kudriavzevii Strain Atz-EN-01 and Its Potential Application for Bioremediation. J Appl Pharm Sci 2013, 3, 35–43. [CrossRef]
- Zaharia, M.; Jurcoane, S.; Maftei, D.; Pui, A.; Dumitras-Hutanu, C.A.; Gradinaru, R. Yeast Biodegradation of Some Pesticide Dinitrophenols. Rom Biotechnol Lett 2013, 18, 8151.
- Alves Jr, S.L.; Treichel, H.; Basso, T.O.; Stambuk, B.U. Are Yeasts “Humanity’s Best Friends”? In Yeasts: From Nature to Bioprocesses; Bentaham Science, 2022; pp. 431–458.
- Bobate, S.; Bokade, P.; Bajaj, A. Engineered Yeasts as Biocatalysts for Pesticide Degradation. In Advances in Yeast Biotechnology for Biofuels and Sustainability; Elsevier, 2023; pp. 449–474.
- Imaishi, H.; Matumoto, S. Isolation and Functional Characterization in Yeast of CYP72A18, a Rice Cytochrome P450 That Catalyzes (ω-1)-Hydroxylation of the Herbicide Pelargonic Acid. Pestic Biochem Physiol 2007, 88, 71–77. [CrossRef]
- Shiota, N.; Kodama, S.; Inui, H.; Ohkawa, H. Expression of Human Cytochromes P450 1A1 and P450 1A2 as Fused Enzymes with Yeast NADPH-Cytochrome P450 Oxidoreductase in Transgenic Tobacco Plants. Biosci Biotechnol Biochem 2000, 64, 2025–2033. [CrossRef]
- Hayashi, E.; Fuzimoto, K.; Imaishi, H. Expression of Arabidopsis Thaliana Cytochrome P450 Monooxygenase, CYP71A12, in Yeast Catalyzes the Metabolism of Herbicide Pyrazoxyfen. Plant Biotechnology 2007, 24, 393–396. [CrossRef]
- Mi, H.; Zhou, Q.; Li, G.; Tao, Y.; Wang, A.; Wang, P.; Yang, T.; Zhu, J.; Li, Y.; Wei, C.; et al. Molecular Responses Reveal That Two Glutathione S-Transferase CsGSTU8s Contribute to Detoxification of Glyphosate in Tea Plants (Camellia Sinensis). Int J Biol Macromol 2024, 277, 134304. [CrossRef]
- Kuroda, K.; Ueda, M. Cell Surface Engineering of Yeast for Applications in White Biotechnology. Biotechnol Lett 2011, 33, 1–9. [CrossRef]
- Ye, M.; Ye, Y.; Du, Z.; Chen, G. Cell-Surface Engineering of Yeasts for Whole-Cell Biocatalysts. Bioprocess Biosyst Eng 2021, 44, 1003–1019. [CrossRef]
- Takayama, K.; Suye, S. -i.; Kuroda, K.; Ueda, M.; Kitaguchi, T.; Tsuchiyama, K.; Fukuda, T.; Chen, W.; Mulchandani, A. Surface Display of Organophosphorus Hydrolase on Saccharomyces Cerevisiae. Biotechnol Prog 2006, 22, 939–943. [CrossRef]
- Fukuda, T.; Tsuchiyama, K.; Makishima, H.; Takayama, K.; Mulchandani, A.; Kuroda, K.; Ueda, M.; Suye, S. Improvement in Organophosphorus Hydrolase Activity of Cell Surface-Engineered Yeast Strain Using Flo1p Anchor System. Biotechnol Lett 2010, 32, 655–659. [CrossRef]
Figure 1.
Examples of ecological services performed by yeasts. AS, Salicylic acid; ET, ethylene; JA, Jasmonic acid.
Figure 1.
Examples of ecological services performed by yeasts. AS, Salicylic acid; ET, ethylene; JA, Jasmonic acid.
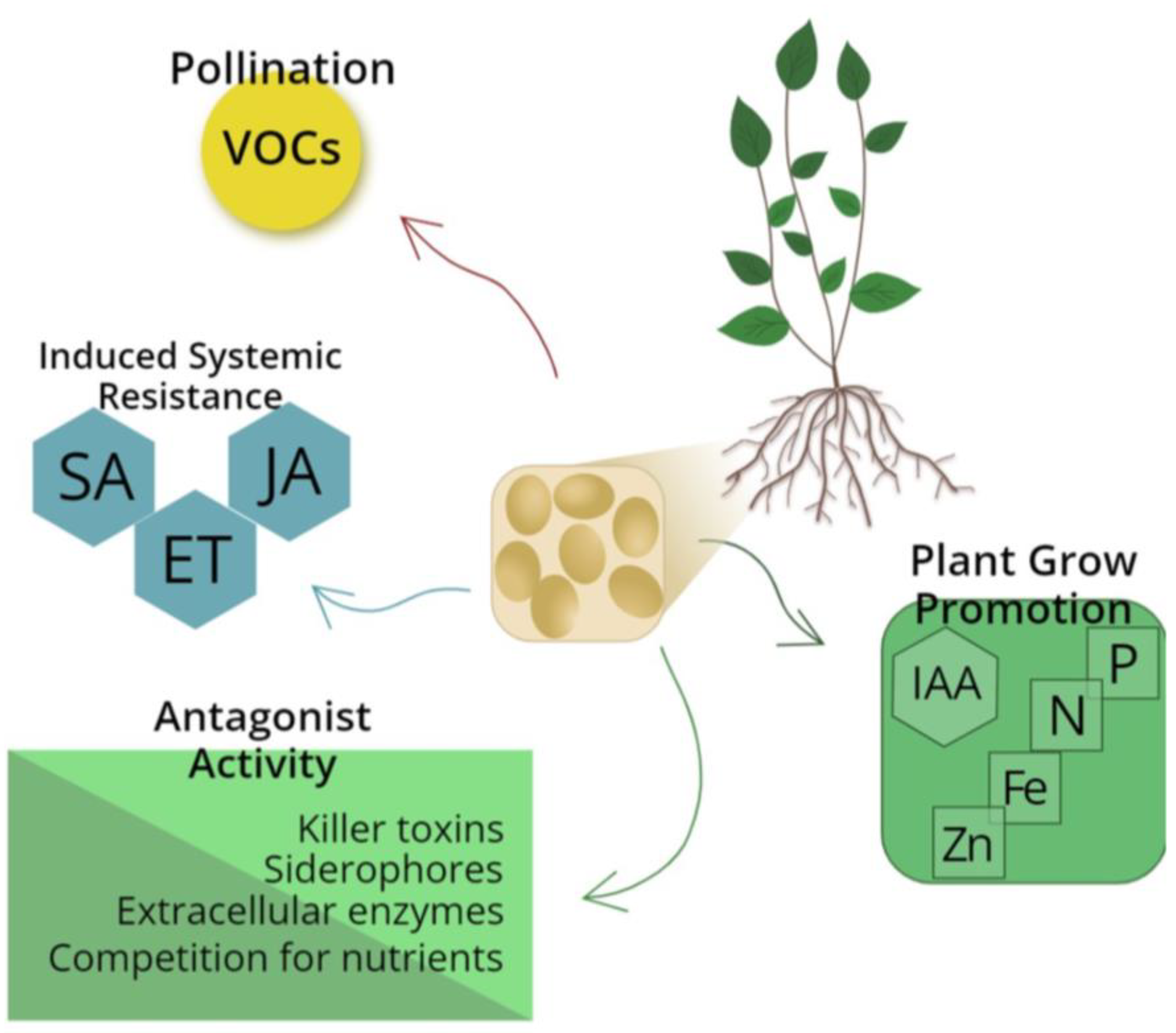
Figure 2.
Metabolic pathways for Indole-Acetic Acid (IAA) production by yeast cells from sugars (glucose and xylose). Glycolysis, Pentose-Phosphate Pathway (PPP), and Shikimate Pathway were almost entirely condensed because they are classic (well-known) metabolic pathways. The enzymes (and their isoenzymes) involved in each reaction are represented by their respective three-letter codes followed by numbers: TRP2 - Anthranilate synthase component 1, TRP4 - Anthranilate phosphoribosyltransferase, TRP1 - N-(5'-phosphoribosyl)anthranilate isomerase, TRP3 - Indole-3-glycerol-phosphate synthase, TRP5 - Tryptophan synthase, ARO8 - Aromatic/aminoadipate aminotransferase 1, ARO9 - Aromatic amino acid aminotransferase 2, PDC1 - Pyruvate decarboxylase isozyme 1, PDC5 - Pyruvate decarboxylase isozyme 2, PDC6 - Pyruvate decarboxylase isozyme 3, ALD2 - Aldehyde Dehydrogenase, ALD3 - Aldehyde Dehydrogenase. CdRP stands for 1-(2-carboxyphenylamino)-1-deoxy-D-ribulose 5-phosphate. Sources: [15,16,17,18,19,20].
Figure 2.
Metabolic pathways for Indole-Acetic Acid (IAA) production by yeast cells from sugars (glucose and xylose). Glycolysis, Pentose-Phosphate Pathway (PPP), and Shikimate Pathway were almost entirely condensed because they are classic (well-known) metabolic pathways. The enzymes (and their isoenzymes) involved in each reaction are represented by their respective three-letter codes followed by numbers: TRP2 - Anthranilate synthase component 1, TRP4 - Anthranilate phosphoribosyltransferase, TRP1 - N-(5'-phosphoribosyl)anthranilate isomerase, TRP3 - Indole-3-glycerol-phosphate synthase, TRP5 - Tryptophan synthase, ARO8 - Aromatic/aminoadipate aminotransferase 1, ARO9 - Aromatic amino acid aminotransferase 2, PDC1 - Pyruvate decarboxylase isozyme 1, PDC5 - Pyruvate decarboxylase isozyme 2, PDC6 - Pyruvate decarboxylase isozyme 3, ALD2 - Aldehyde Dehydrogenase, ALD3 - Aldehyde Dehydrogenase. CdRP stands for 1-(2-carboxyphenylamino)-1-deoxy-D-ribulose 5-phosphate. Sources: [15,16,17,18,19,20].
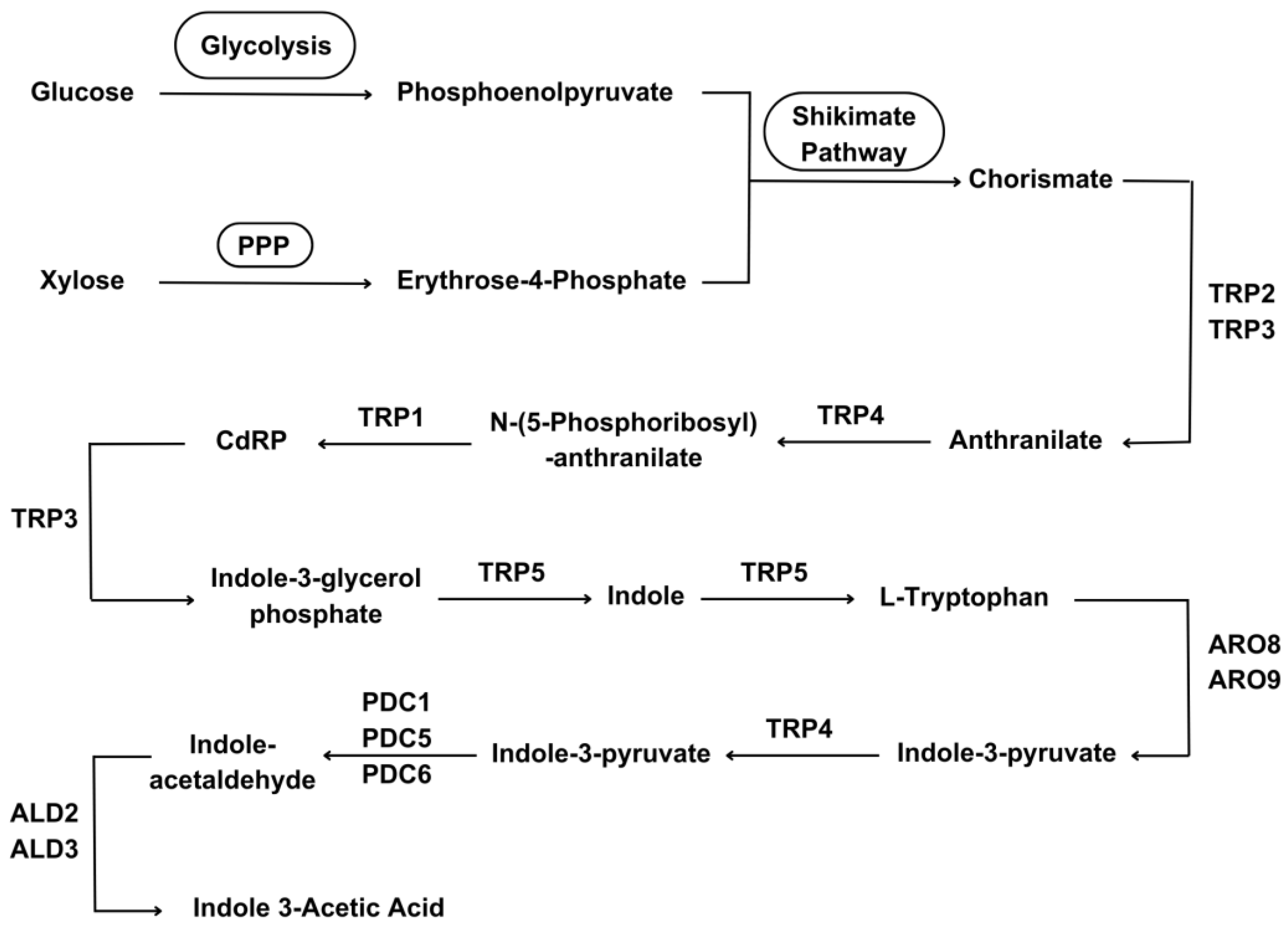
Figure 3.
Chemical structures of (a) 2,4-Dichlorophenoxyacetic acid (2,4-D); (b) 2,2-dimethyl-3H-1-benzofuran-7-yl) N-methylcarbamate (Carbofuran); (c) 2-(phosphonomethylamino) acetic acid (Glyphosate); (d) 3,4-dimethyl-2,6-dinitro-N-pentan-3-ylaniline (Pendimethalin); and, (e) 6-chloro-4-N-ethyl-2-N-propan-2-yl-1,3,5-triazine-2,4-diamine (Atrazine). Source: [59].
Figure 3.
Chemical structures of (a) 2,4-Dichlorophenoxyacetic acid (2,4-D); (b) 2,2-dimethyl-3H-1-benzofuran-7-yl) N-methylcarbamate (Carbofuran); (c) 2-(phosphonomethylamino) acetic acid (Glyphosate); (d) 3,4-dimethyl-2,6-dinitro-N-pentan-3-ylaniline (Pendimethalin); and, (e) 6-chloro-4-N-ethyl-2-N-propan-2-yl-1,3,5-triazine-2,4-diamine (Atrazine). Source: [59].
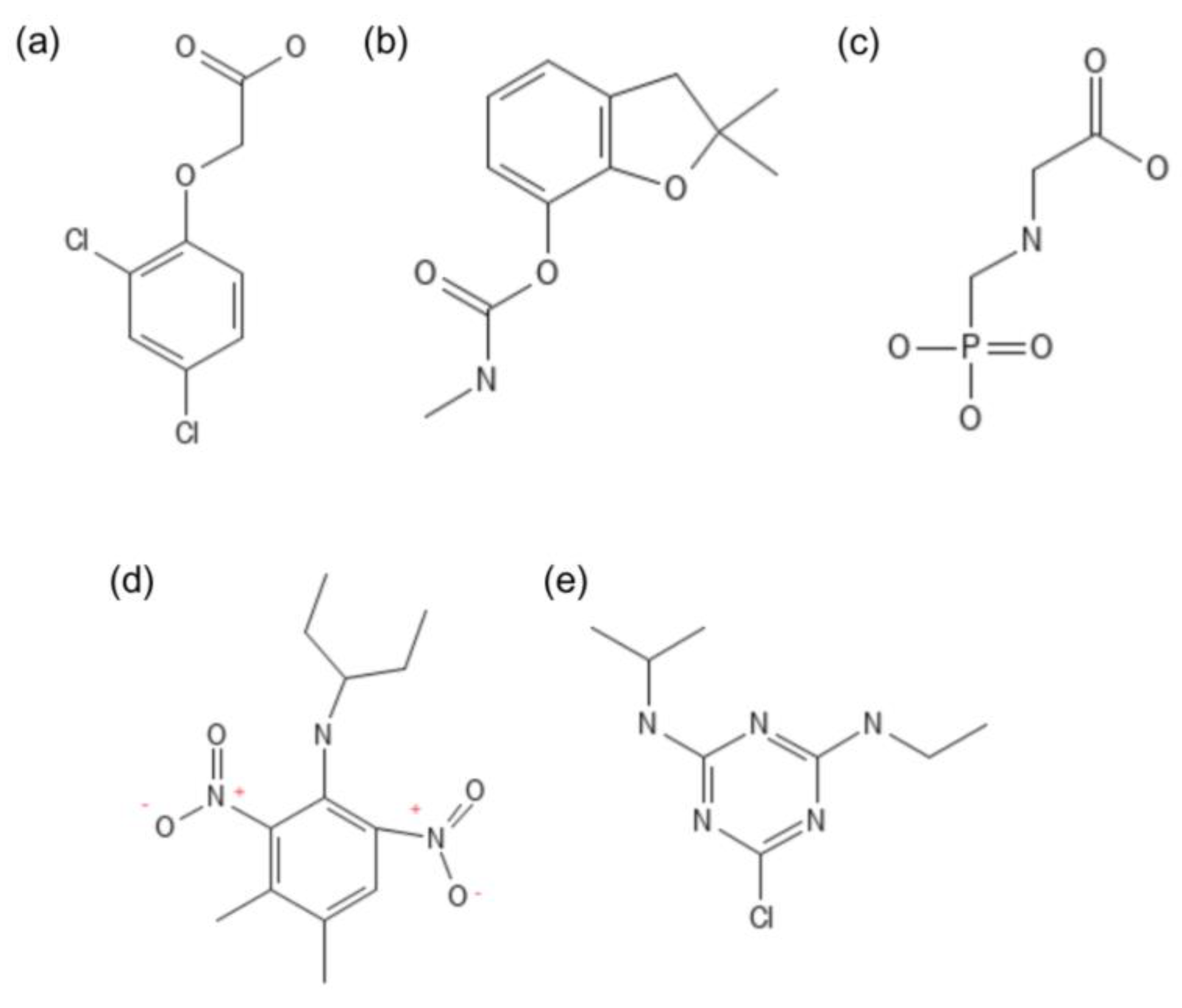
Figure 4.
Yeast-cell surface display strategy. Through genetic engineering, heterologous degradation enzymes can be displayed at the yeast cell wall when attached to a carrier protein with a signal peptide and an anchor system (either GPI or Flo1p — see the text for additional details). In this way, pesticides can be transformed into less-toxic compounds outside the yeast cells.
Figure 4.
Yeast-cell surface display strategy. Through genetic engineering, heterologous degradation enzymes can be displayed at the yeast cell wall when attached to a carrier protein with a signal peptide and an anchor system (either GPI or Flo1p — see the text for additional details). In this way, pesticides can be transformed into less-toxic compounds outside the yeast cells.
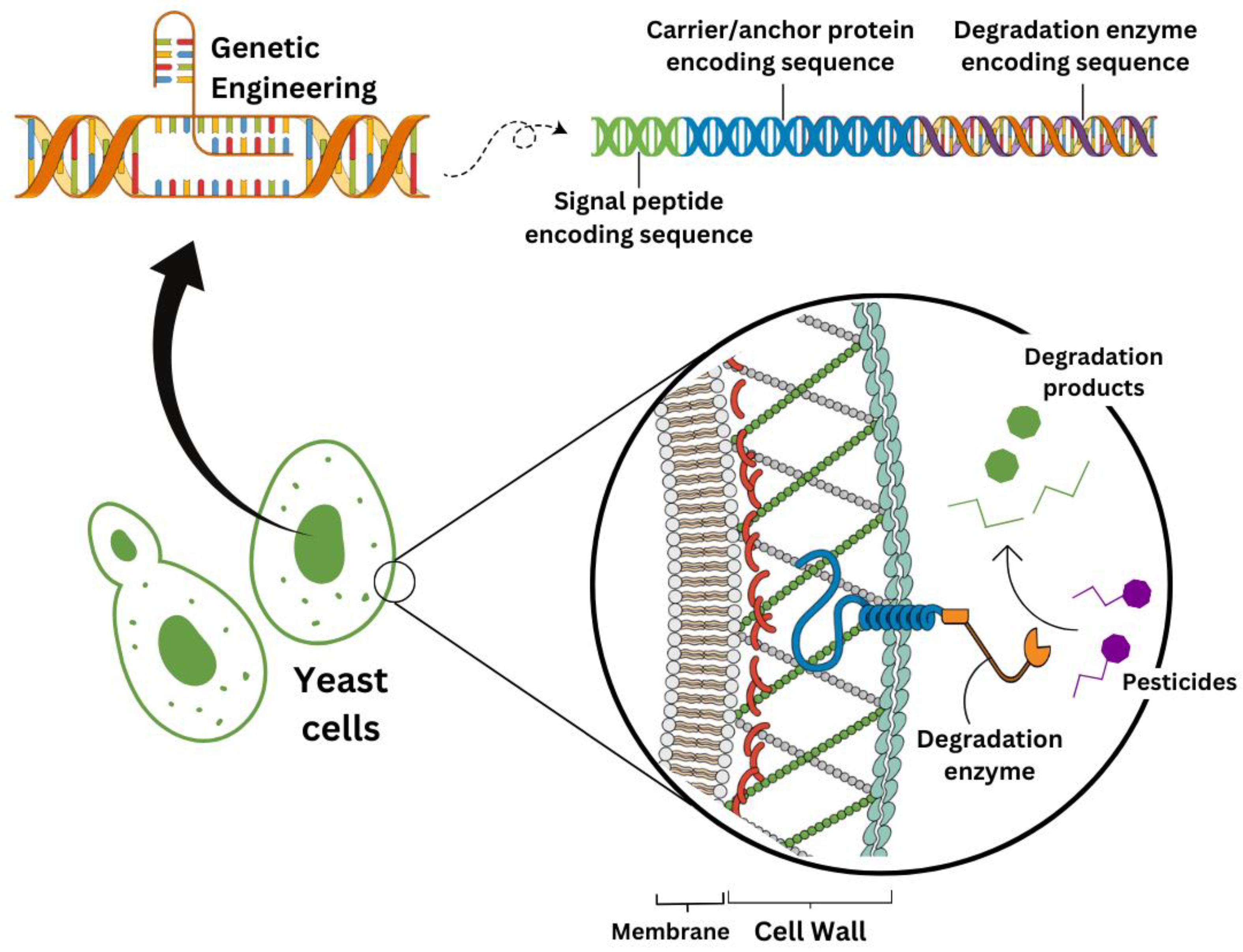
Figure 5.
Scientific production related to yeasts in agriculture in the years 2004-2024. The search terms used were “yeasts AND agriculture AND bioremediation”.
Figure 5.
Scientific production related to yeasts in agriculture in the years 2004-2024. The search terms used were “yeasts AND agriculture AND bioremediation”.
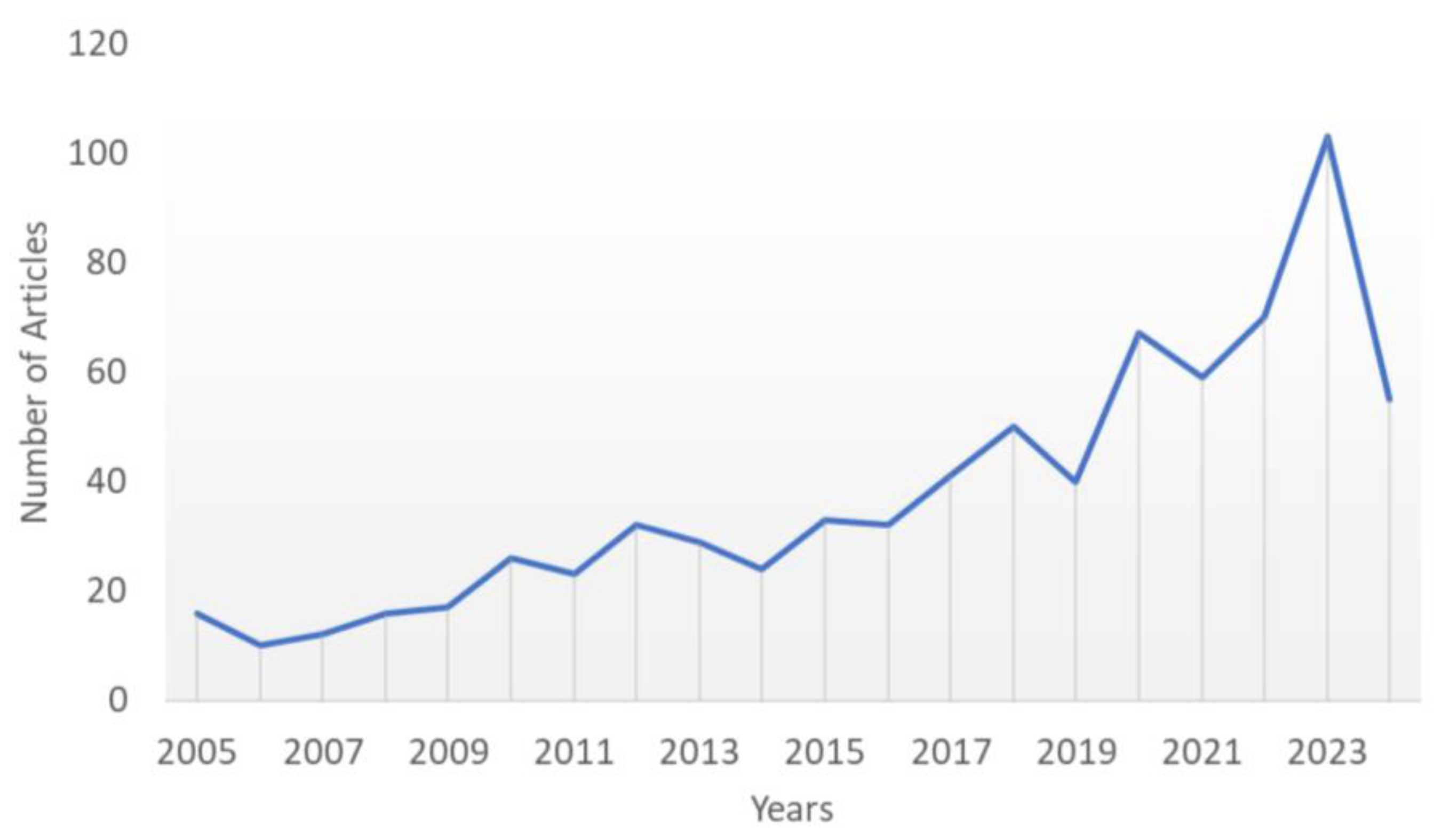

Table 1.
Physiological processes through which yeasts promote plant growth.
| Yeast Species | Physiological Process | Reference |
| Aureobasidium pullulans, Myriangiales sp., Occultifur brasiliensis, Candida silvae, Cryptococcus podzolicus | Nitrogen and Carbon availability | [11] |
| Pichia kudriavzevii, Issatchenkia terricola | Phosphorus availability, IAA1 production | [12] |
| Cryptococcus flavus, Candida railenensis | Phosphorus availability | [13] |
| Meyerozyma guilliermondii, Candida zemplinina, Candida pimensis, Lachancea lanzarotensis, Rhodotorula mucilaginosa | IAA1 production | [10] |
| Kazachstania rupicola, Rhodosporidium diabovatum, Saccharomyces cerevisiae | IAA1 production | [14] |
1Indol-acetic acid.
Table 2.
Antagonistic activities exerted by yeasts.
| Yeast Species | Pathogen | Yeast Antagonist Action | Reference |
| Papiliotrema laurentii | Pythium ultimum | β-1,3-glucanase production | [26] |
| Rhodotorula minuta, Candida azyma, Aureobasidium pullulans | Georichum citri-aurantii | Competition for nutrients, β-1,3-glucanase, Chitinase, Killer activity | [27] |
| Wickerhamomyces anomalus | Rhizoctonia solani, Curvularia lunata, Fusarium moniliforme | Production of VOCs, β-1,3-glucanase, and chitinase | [22] |
| Pichia galeiformis | Penicillium digitatum | Competition for space and nutrients, VOCs production | [23] |
| Pseudozyma graminicola | Bullera hannae, Cryptococcus nemorosus, Dacrymyces stillatus, Neovossia setariae, Sporobolomyces singularis | Cellobiose-lipid production | [28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



