
Submitted:
03 October 2024
Posted:
07 October 2024
You are already at the latest version
Abstract
The incorporation of unconventional feed ingredients in animal husbandry aims to improve nutritional efficiency and animal health. Bamboo, known for its rich mineral, antioxidant, and dietary fiber content, represents a potential alternative feed ingredient. This study explores the effects of fermented bamboo powder (FBP) on growth performance, antioxidant status, intestinal morphology, and expression of nutrient transporter genes in broiler chickens. 600 healthy 1-day-old chicks were randomly divided into two groups, each with 10 replicates of 30 chicks. The control group was provided with a basal diet. The experimental group was given the basal diet with supplemented 1.0 and 2.0 g/kg FBP for two different stages: phase Ⅰ (1-22 days) and phase Ⅱ (23-45 days). The results indicated a significant improvement in growth performance of the group fed with fermented bamboo powder compared to the control group (P < 0.05). More-over, there was a significant effect on the weight of organs and muscles when compared to the control group (P < 0.05). Moreover, there was a significant effect on triglyceride levels (P < 0.05), with a positive effect on urea and creatinine levels (P < 0.05), as well as a significant improvement in the levels of antioxidant enzymes (MDA, SOD, and CAT) compared to the control group (P < 0.01). In contrast, fermented bamboo powder significantly enhanced the surface area available for nutrient absorption in the intestine (duodenum, jejunum, and ileum) by increasing the villus height, intestinal lumen depth, and villus area (P < 0.01). FBP increased the expression levels of genes involved in the transport of nutrients from the intestinal regions (duodenum, jejunum, and ileum) namely GLUT1, GLUT2, CAT1, CAT2, PEPT1, PEPT2, and LAT1, compared to control. The upregulation of nutrient transporter genes further facilitated nutrient absorption, contributing to the observed improvements in growth and body weight. These findings endorse FBP as a viable feed additive in broiler diets to enhance health and performance.
Keywords:
fermented bamboo powder
; growth performance
; intestinal morphology
; nutrient transporter genes
; broilers
1. Introduction
Bamboo, a globally prevalent perennial grass with a woody texture, belongs to the Gramineae family and Bambuseae subfamily. As the leading producer, China contributes significantly to the global bamboo supply, offering large quantities at low costs [1]. which account for nearly 3% of the world’s forest area [2]. Renowned for its rapid growth and high production potential, bamboo stands out as a crucial renewable resource with a broad geographical distribution [3]. Bamboo composition is enriched with various physiologically active compounds, predominantly flavonoids, polysaccharides, phenolic acids which has significant effects on chicken [4,5]. Additionally, bamboo is rich in insoluble dietary fiber (IDF) [6]. Flavonoids in bamboo are known to mitigate intestinal inflammation and modulate immune activities within the gut [7]. Phenolic components in bamboo are recognized for their ability to reduce fatty liver risks while also providing antioxidant and antibacterial benefits [8,9,10]. certain substances may be more concentrated and bioavailable in fermented bamboo powder than in unfermented bamboo powder due to the fermentation process which potentially enhance the growth of lactic acid bacteria and improve substrate fermentability [11].Furthermore, the fermented bamboo powder had the following percentages in its as-fed composition: dry matter (89.80%), crude protein (2.28%), ether extract (0.18%), acid detergent fiber (47.36%), calcium (0.12%), total phosphorus (0.02%), and gross energy (16.85 MJ/kg) [12,13].
Nutrient absorption in the small intestinal epithelium is intricately linked to various transporters, including glucose transporters (GLUT1 and GLUT2) for glucose, cationic amino acid transporters (CAT1 and CAT2) for amino acids, Na+-dependent neutral/cationic amino acid exchangers (y+L amino acid transporter-1 and y+L amino acid transporter-2) for amino acids, and peptide transporters (PepT1 and PepT2) for peptides [14,15].
Our recent works have also showed that FBP activates gut odorant receptors, and promotes intestinal health and growth performance of Broiler Chickens [16]. Despite existing studies on fermented bamboo powder (FBP) usage in chickens and pigs, information on the mechanisms by which FBP affects these animals remains scarce. This study seeks to fill this gap by examining the effects of various FBP supplementation levels on growth performance, serum biochemical markers, antioxidant status, intestinal morphology, and nutrient transporter gene expression in broiler chickens.
2. Materials and Methods
2.1. Diets, Animals, and Experimental Design
The experimental protocols were authorized by the Animal Care and Use Committee of Nanjing Agricultural University, Nanjing, China, with permission Numbers: (Permit Number: SYXK (Su) 2022–0031. The basal and diet was obtained from Jiangsu Yancheng Xiling Agricultural Science and Technology Co., Ltd. (Nanjing, China). The basal diet was formulated in accordance with NRC (1994) recommendations for nutrition requirements which outlines the nutrient requirements for yellow chickens as shown in Table 1. FBP was obtained from Zhejiang Muyi Xiangzhu Biotechnology Co. Ltd, China, and core technology from the Research Institute of Global 3E, Kyoto, Japan. The ingredients of FBP are listed in Table 2.
A group of 600 dwarf yellow-feathered broiler (DYB) chickens, aged 1 day and with an average body weight of 52.16 ± 0.26 gm were used for this experiment and administered normal immunization regimens and provided with full access to food and water for a period of 45 days. Chicks were divided into two groups: the control group and the FBP group. The control group received the basal diet throughout the entire trial. On the other hand, the FBP group was given the basal diet supplemented with 1.0 g/kg FBP from day 1 to day 22 (phase Ⅰ) and then the supplementation was increased to 2.0 g/kg FBP from day 23 to day 45 (phase Ⅱ). Each group is comprised of 10 replicates with each replicate consisting of 30 chickens reared on ground bedding.
2.2. Sampling and Data Collection
During the experiment, measurements of body weights (BW) were taken on three specific days: day 1, day 22, and day 45. Additionally, average daily gain (ADG) and feed consumption were recorded and subsequently calculated on a pen basis. The calculation of the average daily feed intake (ADFI) per bird involved dividing the total feed consumption for the pen for the full trial period by the number of days in that period. The feed conversion ratio (FCR), also known as the gain-to-feed ratio was determined on a pen basis by analyzing the body weight and feed consumption data. In each repetition of two groups, two broilers that had undergone a 12-hour fasting period were randomly chosen and their weights were measured on the 45th day. The venous blood sample was obtained and subjected to centrifugation at a speed of 3000 (rpm) for 10 minutes. The serum samples that were acquired were kept at a temperature of -20℃. The chosen chickens were slaughtered to assess the slaughter performance by measuring various parameters such as eviscerated yield, breast and thigh weights, as well as the weights of internal organs including the heart, spleen, kidneys, and liver. Tissues from the small intestine (duodenum, jejunum, and ileum) were collected for subsequent hematoxylin and eosin (HE) examination and RNA extraction.
2.3. HE Staining of Tissue Sections
Tissues from the small intestine (duodenum, jejunum, and ileum) were stored in 75% ethanol at room temperature for subsequent HE examination. Intestine tissues were sliced to 3–4 mm thick and fixed in 10% neutral buffered formalin then dehydrated by soaking in a graded series of ethanol, cleared in xylene, and embedded in paraffin. The paraffin blocks were sectioned with a microtome at (5–7μm) thickness and stained with HE staining (Beijing Solarbio Science &Technology Co. Ltd) to examine their morphology study. Stained sections were examined using Nikon eclipse E200 microscope (Nikon Instruments, shanghai Co. Ltd). The values of villus height (VH), crypt depth (CD), and villus area for intestinal samples were measured 5 times from different villus and crypts per section from each broiler using the ImageJ software 1.54d (USA) and Villus surface area was calculated using the formula: [17]
villus area = 2π × (average villus width/2) × villus height.
2.4. Serum Biochemical Indexes and Antioxidant Enzymes
The levels of total protein, glucose, cholesterol, triglyceride, high-density lipoprotein cholesterol (HDL), and low-density lipoprotein cholesterol (LDL) in sera were assessed in accordance with the guidelines provided by the respective kits (Nanjing Angle Gene Co. Ltd, Nanjing, China). In this study, serum samples were utilized for the purpose of assessing the activity of antioxidant enzymes. Specifically, the total activity of superoxide dismutase (SOD), the concentrations of malondialdehyde (MDA), glutathione peroxidase (GSH-PX), HemeOxygenase HO-1, and catalase (CAT) were measured using commercial kits provided by Nanjing Angle Gene Co. Ltd in Nanjing, China.
2.5. RNA Extraction, and Quantitative Real-Time PCR
2.5.1. RNA Isolation and cDNA Synthesis
The small intestine (duodenum, jejunum, and ileum) samples were collected and placed in liquid nitrogen, then stored at −80 °C until analysis. The total RNA was extracted using (Total animal tissue/cells RNAExtraction kit(TSP413), Qingke.) following the manufacturer’s protocol. The integrity and quality of RNA was verified using agarose gel electrophoresis. The RNA concentration was determined using (Epoch microplate spectrophotometer, Agilent Technologies, Inc.). The RNA sample was reverse transcribed to cDNA using (SynScript®Ⅲ RT SuperMix for qPCR reverse transcription kit), then stored at −20 °C until analysis.
2.5.2. Q-PCR (Quantitative Real-Time Polymerase Chain Reaction)
Primer sequences were used to amplify the selected genes as shown in Table 3. The primer sequences were verified using NCBI primer blast. The cDNA product obtained by reverse transcription was diluted 3 times and used as a qPCR template, it was amplified by ArtiCanCEO SYBR qPCR MixTSE401,(Qingke), following the manufacturer’s recommendations using ABI QuantStudio real-time PCR system, Thermo Fisher Scientific Inc. The amplification protocol was: initial denaturation at 95 °C for 5 min, followed by 40 cycles at 95 °C for 15 s, annealing temperature at 60°C for 20 s, and extension at 72ºC for 20 s. Dissociation curves were analyzed to verify that only one peak was found for each specific melting temperature, thus showing that the PCR products were amplified specifically. All samples were tested in duplicates. Target gene Ct values were first normalized against the Ct values of the housekeeping gene (β actin), then used to calculate the relative gene expression levels (fold change).
2.6. Statistical Analysis
Observed data were subjected to statistically analyze by statistical analysis system software SPSS V.26. Significant differences among the means were determined by independent-samples t test at P < 0.05. The data are presented as means ± standard error. For gene expression, all samples were assayed in triplicate and data were normalized to the reference gene (β-actin). Relative gene expression was calculated and analyzed by the 2^-△△Ct.
3. Results
3.1. Growth Performance
The results of the overall growth performance indicated the positive impacts of including FBP in broiler diets. The findings shown in Table 4. provided a concise overview of the impact of dietary supplementation with FBP on the BW of broiler chicks across two separate growth periods with two additional levels from FBP 1.0 and 2.0 g/kg (P < 0.001), wherein gradual levels of FBP were administered. It is evident that the birds had a similar initial body weight at the starting point of the trial in both the control and FBP groups.
3.2. Carcass and Organ Weight
The post-slaughter analysis revealed significant increases in the breast weight, thigh weight, liver weight, and kidney of chickens fed FBP supplementation diet compared to the control group (P < 0.05). These improvements indicated that the inclusion of FBP in the diet positively influenced carcass characteristics and organ weights. However, there were no significant differences observed between the control and treatment groups of eviscerated yield (P > 0.05). The data analysis demonstrated an interaction between the supplemented feed and the organs, highlighting the consistent increase in carcass weight and organ weights with the inclusion of FBP in the diet. This interaction emphasizes the potential benefits of FBP in enhancing carcass quality and organ development in chickens as shown in Figure 1.
3.3. Intestinal Morphology
The findings showed that the inclusion of FBP in the diet enhanced the structural features of the gut (duodenum, jejunum, and ileum) on day 45 in comparison to the control group. The villi in the duodenum exhibited greater height in the FBP group, as compared to the control group. This difference was shown to be statistically significant. Additionally, the FBP group displayed deeper crypts in comparison to the control group. Nevertheless, there was no notable disparity observed in the ratio of villus height and depth of crypts, as well as the villi area, between the control group and the group administered with FBP. The addition of FBP had a positive impact on the depth of the crypts and the height of the villi in the jejunum, with a significant difference (P < 0.01). However, the treatment group did not have a significant effect on the ratio of villi with crypts and the villi area. Concerning the morphology of the ileum, the addition of FBP had a significant effect on the height of the villi, the depth of crypts, and the area of the villi in the ileum. However, the correlation between the development of crypts in the ileum and the treatment with FBP was not statistically significant. The results mentioned above are presented in Table 5, Figure 2. Furthermore, the inclusion of FBP substantially enhanced the height of the villi in the duodenum, jejunum, and ileum, as well as increasing the depth of the crypts and the area of the villi when compared to the control group.
3.4. Serum Biochemical Index
The results illustrated the impact of FBP on the blood biochemical analysis of broiler chickens as shown in Table 6. The results indicated that including FBP in the diet led to a decrease in serum levels of cholesterol (Chol), glucose (GLU), and low-density lipoprotein (LDL). Furthermore, it was noted that the incorporation of FBP resulted in elevated levels of high-density lipoprotein (HDL) and total protein (TP) in comparison to the control group. Conversely, the use of FBP had a notable effect on the concentration levels of triglycerides (TG) in the serum (P < 0.05). Conversely, FBP had a notable impact on liver function by reducing the serum aspartate transaminase (AST) to alanine transaminase (ALT) ratio (P < 0.05). Furthermore, the FBP resulted in a notable reduction in urea concentration, with a significant difference (P < 0.05), while no significant change was seen in creatinine concentration (P > 0.05).
3.5. Antioxidant Status
The results in Table 7 showed the influence of FBP on serum antioxidant markers. The activities of SOD and CAT were found to be considerably increased compared to the control group (P<0.01). There were no notable disparities seen in the levels of GSH-PX content between the control group and the FBP group. Conversely, the level of MDA demonstrated a noteworthy reduction (P = 0.01) in the FBP group when compared to the control group.
3.6. Effects of FBP on Relative mRNA Expression Levels of Nutrient Transporter Genes in Intestinal
The mRNA expression levels of seven genes was investigated in feed FBP addition in in the small intestine (duodenum, jejunum, and ileum) as presented in Figure 3. the results showed that The FBP diet significantly upregulated CAT1, CAT2, PepT2, LAT1 in the intestinal parts of the birds compared with expression of the genes in the CON diet group (P < 0.05). While there is no responses to FBP diet were observed for other genes investigated, namely GLUT1, GLUT2 and PepT1 compared to the control. On the other hand, the mRNA expression level of GLUT upregulated by FBP diet in jejunum.
4. Discussion
Previous research conducted on broilers has demonstrated that the inclusion of insoluble fiber in their diet can effectively enhance their growth [18,19]. The growth performance of broilers is influenced by various aspects, such as the type of fiber resources utilized [20], the level of fiber addition and the particle size of the fiber [21], and the specific feeding stage [22]. Bamboo a rapidly growing perennial plant, is recognized for its high nutritional fiber content in the form of bamboo powder. Consequently, it has been regarded as a significant substitute feedstock in animal production [23]. Nevertheless, further investigation is warranted to examine the utilization of FBP in the context of poultry farming. The present investigation that FBP has significantly increased on growth performance, specifically ADFI, across different growth stages when gradually incorporating diets supplemented with FBP. Furthermore, the inclusion of FBP in the diet showed significanlty improvement in FCR and a significant increase in (ADG) and (BWG) during growth stages 1-22 and 23-45. These improvements were observed with a gradual increase in FBP levels by 1% and 2% compared to the basic diet. This finding coincides with the results of a recent study which indicated that the inclusion of 1% micronized bamboo powder in broiler diets can enhance their growth performance [24,25]. In a comparable direction, a further study conducted by researchers found that no negative influences in the growth performance, such as ADG and ADFI, for growing–fattening pigs fed the FBP addition diets [13]. The findings of this study suggest that FBP has the potential to serve as a viable feed element in the production of chickens.
Serum biochemical measures are commonly employed in nutritional assessments to evaluate the quality of test feedstuffs or additives and provide insights into the physiological, metabolic activity of animals [26]. The present investigation demonstrated that the inclusion of FBP had beneficial results on a lot of serum biochemical indicators. Triglycerides and cholesterol are distinct lipid molecules that move inside the body as constituents of lipoproteins. Coronary heart disease is a condition that arises from the accumulation of cholesterol on the walls of arteries, resulting in the formation of plaque and subsequent constriction of the arteries. This narrowing leads to reduced blood flow to the heart. An excessive presence of triglycerides and cholesterol has been linked to the development of coronary heart disease. Monocytes internalize the low-density lipoprotein (LDL) particles and attach to the endothelial cells lining the coronary artery. Subsequently, these monocytes undergo differentiation into macrophages and subsequently amass within the artery, leading to the formation of a "fatty streak" that ultimately progresses into an atherosclerotic lesion [27,28]. In the present investigation, it was observed that the levels of blood triglycerides exhibited a consistent and statistically significant increase in chickens that were fed the control diet, as compared to those that were provided with the FBP Diet. Conversely, no significant differences were seen in terms of cholesterol and LDL concentration. The decrease in serum metabolites seen in broilers that were fed diets supplemented with FBP has revealed an additional health advantage associated with the use of FBP as a dietary supplement in broiler chickens. These findings are consistent with previous research that has addressed the beneficial impact of a BLE diet on serum biochemical markers in broiler chickens [29], as well as the effects of a diet containing Fermented Bamboo Shoot Processing Waste FPSBW on serum triglycerides and cholesterol in pigs [11].
Nevertheless, the addition of FBP resulted in a greater increase in serum HDL levels, although this difference was not statistically significant. Consequently, the addition of FBP may lead to higher HDL and IgA levels, thereby improving the health status of broiler chickens and providing protection against illnesses. The elevated levels of HDL in bamboo products may be attributed to their high concentrations of flavonoids, organic acids, phenolic compounds, and polysaccharides, which have been found to regulate lipid metabolism [10,30].
Previous research demonstrated that the inclusion of bamboo shoot shell fiber in the diet improved fat metabolism disorders in mice [31]. This dietary intervention resulted in reduced serum cholesterol, triglyceride, and low-density lipoprotein cholesterol levels while increasing the levels of high-density lipoprotein cholesterol. Conversely, the levels of serum total protein and urea nitrogen are commonly considered markers of protein synthesis and metabolism, factors that are associated with the growth performance of broiler chickens. The observed elevations in serum total protein levels suggest an accelerated synthesis and absorption of protein. Serum urea nitrogen is a result of the breakdown of proteins. Therefore, a reduction in serum urea nitrogen suggests an increase in the synthesis of proteins from amino acids [32]. The concentration of serum urea nitrogen is considered to be a significant determinant of the quality of protein in the diet. So the reduction in serum urea nitrogen levels indicates the better protein quality of fermented feed, as it demonstrates a more efficient utilization of amino acids for tissue protein synthesis. However, The concentration of glucose in the serum serves as a reliable measure of the availability of energy [33]. The current study observed a notable decrease in serum urea and glucose levels in chickens that were administered the FBP, in comparison to the control group. Nevertheless, there was no discernible effect on the overall protein concentration. This conclusion is consistent with the results published in previous research. [24].
The antioxidant parameters observed in serum serve as indicators of the organisms’ antioxidant capacity. Antioxidant enzymes, such as superoxide dismutase SOD, catalase CAT, and glutathione peroxidase GSH-Px, work in collaboration to effectively remove free radicals and uphold cellular equilibrium. MDA is considered to be a lipid peroxidation byproduct that serves as an indicator of the level of oxidative stress. The findings of our study indicate that the inclusion of FBP in the diet resulted in a significant enhancement in the activities of SOD and CAT in the serum, as well as an improvement in GSH-Px activity, as compared to the control group. Furthermore, the concentration of MDA in the serum was shown to be significantly decreased in the FBP group compared to the control group. It is reasonable to hypothesize that the flavonoids present in FBP play a significant role in enhancing the activities of antioxidant enzymes and preventing the formation of thiobarbituric acid-reactive substances. Previous research has elucidated that the antioxidant properties of flavonoids in biological systems are attributed to their ability to transfer electron free radicals, chelate metal catalysts, activate antioxidant enzymes, and inhibit oxidases [34]. Furthermore, other studies have demonstrated that the supplementation of broilers with plant extracts containing flavonoids, polyphenols, and polysaccharides, such as Ginkgo biloba leaf extract [35] and Artemisia annua extract [36], can augment the capacity for scavenging free radicals.. Based on the findings of our current study, it is recommended that the inclusion of FBP in the diet of broiler chickens resulted in enhanced antioxidant capacity. These outcomes agree with prior research indicating that the administration of BLE contributed to improvements in the oxidative status of broiler chickens [37].
The majority of nutrient absorption takes place in the small intestine. Longer intestinal villi are associated with an increase in the absorptive surface of the intestine while lowering the height of intestinal villi results in a reduction in the absorption of nutrients. Increased height of villus corresponds to improved nutrient digestion and absorption [38]. The inclusion of a diet supplemented BLE has been found to have positive effects on biogenesis of mitochondria in the small intestine of broiler chickens [39]. Our investigation revealed that the administration of FBP enhanced gut development by augmenting the height and area of the intestinal villi. The observed augmentation in both villus height and area of the intestine in this study can be attributed to the antimicrobial, antioxidant, and anti-inflammatory properties of bamboo by increasing the number of pathogenic bacteria while stimulating the growth of beneficial bacteria in the intestine, leading to reduced inflammatory processes at the intestinal mucosa [13,40]. These improvements in gut structure and function allow better utilization of the available dietary nutrients, including protein and minerals, which subsequently drive the improvements in feed efficiency.
In the chickens, the small intestine is the main site of nutrient absorption.The transport of nutrients from the intestine to the blood is mediated by transporters located on the surface of epithelial cells lining the villi [41]. Carbohydrates are absorbed into the enterocytes of the small intestine through the action of sugar transporters, which facilitate the diffusion of monosaccharides into the extracellular fluid and subsequently into the bloodstream [42]. Amino acids are transported either as free amino acids or as small peptides, using specific amino acid transporters or peptide transporters, respectively [43] It is known that the glucose transporters: glucose transporter-1 (GLUT1) and glucose transporter 2 (GLUT2, cationic amino acid transporters, such as CAT1 and CAT2, Na +-dependent neutral/cationic amino acid exchanger, such as y+ L amino acid transporter-1 and y+ L amino acid transporter-2, and peptide transporters, such as PepT1 and PepT2, in the small intestinetinal epithelium are closely associated with nutrient absorption capacity [14], both sugarcane bagasse (SB) and coarse particle size as dietary fibers modulate expression of genes encoding important digestive enzymes and nutrient transporters digestive system in broiler chicken [44]. In the present study, broiler performance was improved by inclusion of dietary FBP and various nutrient transporters in the duodenum, jejunum and ileum were shown to be upregulated. For instance, CAT1, a transporter mediating the bidirectional transport of cationic amino acids, in duodenum, jejunum and ileum were upregulated by FBP. Furthermore, jejunal GLUT1, responsible for Glucose transportation and PepT2, which has a minor contribution in transporting di- and tri-peptides, were upregulated by FBP. In fact, the upregulated nutrient transporters will not only improve nutrient absorption but also play a vital role in the maintenance of intestinal barrier integrity and immune response. A deficiency of amino acids, such as alanine, cysteine, serine, threonine, arginine, and lysine, has long been known to impair immune function and increase the susceptibility of animals to infectious disease.
5. Conclusions
Overall, the addition of FBP to the diet has significantly impact on on growth performance. Additionally, it had positive effects on promoting serum biochemical and antioxidant indices. This study serves as a significant milestone in comprehending the potential of FBP as a dietary fiber source to enhance poultry production, Given the beneficial properties of this plant, we highly recommend the use of FBP as a supplement in poultyr diet.
Author Contributions
Shoura Hytham: conceptualization, investigation, methodology, formal analysis, writing-original draft, and validation. Linsong Hou and Rahmani Mohammad Malyar: Methodology and formal analysis. Wei Ding, Quanwei Wei, Weisheng Zhou and Fangxiong Shi: Conceptualization, supervision, validation, project administration, funding acquisition, review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Jiangsu Provincial Seed Industry Revitalization Project (JBGS (2021)108) and the Basic Business Expenses for International Students in Nanjing Agricultural University (Z552000008).
Institutional Review Board Statement
This animal study protocol was approved by the Animal Care and Use Committee of Nanjing Agricultural University, Nanjing, China, with approval Numbers: SYXK (Su) 2022–0031.
Data Availability Statement
Not applicable
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Bian, F., et al., Bamboo - An untapped plant resource for the phytoremediation of heavy metal contaminated soils. Chemosphere, 2020. 246: p. 125750. [CrossRef]
- Qi, S., et al., Bamboo Forest Mapping in China Using the Dense Landsat 8 Image Archive and Google Earth Engine. Remote Sensing, 2022. 14(3). [CrossRef]
- Bisht, N.C.a.M.S., Bamboo: A Prospective Ingredient for Functional Food and Nutraceuticals. 10th World Bamboo Congress, Korea 2015, 2015.
- Cheng, Y., et al., Bamboo leaf: A review of traditional medicinal property, phytochemistry, pharmacology, and purification technology. J Ethnopharmacol, 2023. 306: p. 116166. [CrossRef]
- Rattanawut, J., O. Pimpa, and K.E. Yamauchi, Effects of dietary bamboo vinegar supplementation on performance, eggshell quality, ileal microflora composition, and intestinal villus morphology of laying hens in the late phase of production. Anim Sci J, 2018. 89(11): p. 1572-1580. [CrossRef]
- Nirmala, C., Bisht, M.S., Bajwa, H.K., Santosh, O.,, Bamboo: A Rich Source of Natural Antioxidants and its Applications in the Food and Pharmaceutical Industry. Trends in Food Science & Technology, 2018. [CrossRef]
- Pei, R., X. Liu, and B. Bolling, Flavonoids and gut health. Curr Opin Biotechnol, 2020. 61: p. 153-159. [CrossRef]
- Kwon, J.H., S.Y. Hwang, and J.S. Han, Bamboo (Phyllostachys bambusoides) leaf extracts inhibit adipogenesis by regulating adipogenic transcription factors and enzymes in 3T3-L1 adipocytes. Food Sci Biotechnol, 2017. 26(4): p. 1037-1044. [CrossRef]
- Van Hoyweghen, L., et al., Phenolic compounds and anti-oxidant capacity of twelve morphologically heterogeneous bamboo species. Phytochem Anal, 2012. 23(5): p. 433-43. [CrossRef]
- Rattanawut, J., et al., Effects of bamboo charcoal powder, bamboo vinegar, and their combination in laying hens on performance, egg quality, relative organ weights, and intestinal bacterial populations. Trop Anim Health Prod, 2021. 53(1): p. 83. [CrossRef]
- Huang, Y., et al., Effects of Fermented Bamboo Shoot Processing Waste on Growth Performance, Serum Parameters, and Gut Microbiota of Weaned Piglets. Animals (Basel), 2022. 12(20). [CrossRef]
- Xiao, Z., et al., Structural characterization, antioxidant and antimicrobial activity of water-soluble polysaccharides from bamboo (Phyllostachys pubescens Mazel) leaves. Int J Biol Macromol, 2020. 142: p. 432-442. [CrossRef]
- Liu, Z., et al., Effects of Fermented Bamboo Powder Supplementation on Serum Biochemical Parameters, Immune Indices, and Fecal Microbial Composition in Growing-Finishing Pigs. Animals (Basel), 2022. 12(22). [CrossRef]
- Fotiadis, D., Y. Kanai, and M. Palacin, The SLC3 and SLC7 families of amino acid transporters. Mol Aspects Med, 2013. 34(2-3): p. 139-58. [CrossRef]
- Roder, P.V., et al., The role of SGLT1 and GLUT2 in intestinal glucose transport and sensing. PLoS One, 2014. 9(2): p. e89977. [CrossRef]
- Malyar, R.M., et al., Fermented Bamboo Powder Activates Gut Odorant Receptors, and Promotes Intestinal Health and Growth Performance of Dwarf Yellow-Feathered Broiler Chickens. Poultry Science, 2024. [CrossRef]
- Nain, S., et al., Effect of metabolic efficiency and intestinal morphology on variability in n-3 polyunsaturated fatty acid enrichment of eggs. 2012. 91(4): p. 888-898.
- Jimenez-Moreno, E., et al., Inclusion of insoluble fiber sources in mash or pellet diets for young broilers. 1. Effects on growth performance and water intake. Poult Sci, 2016. 95(1): p. 41-52. [CrossRef]
- Jimenez-Moreno, E., et al., Insoluble fiber sources in mash or pellets diets for young broilers. 2. Effects on gastrointestinal tract development and nutrient digestibility1. Poult Sci, 2019. 98(6): p. 2531-2547. [CrossRef]
- Pourazadi, Z., et al., Effect of particle size of insoluble fibre on growth performance, apparent ileal digestibility and caecal microbial population in broiler chickens fed barley-containing diets. Br Poult Sci, 2020. 61(6): p. 734-745. [CrossRef]
- Tejeda, O.J. and W.K. Kim, Effects of fiber type, particle size, and inclusion level on the growth performance, digestive organ growth, intestinal morphology, intestinal viscosity, and gene expression of broilers. Poult Sci, 2021. 100(10): p. 101397. [CrossRef]
- Berrocoso, J.D., et al., The effect of added oat hulls or sugar beet pulp to diets containing rapidly or slowly digestible protein sources on broiler growth performance from 0 to 36 days of age. Poult Sci, 2020. 99(12): p. 6859-6866. [CrossRef]
- Ramakrishnan, M., et al., Genetics and genomics of moso bamboo (Phyllostachys edulis): Current status, future challenges, and biotechnological opportunities toward a sustainable bamboo industry. Food and Energy Security, 2020. 9(4). [CrossRef]
- Dai, F., et al., Effects of micronized bamboo powder on growth performance, serum biochemical indexes, cecal chyme microflora and metabolism of broilers aged 1-22 days. Trop Anim Health Prod, 2022. 54(3): p. 166. [CrossRef]
- Dai, F., et al., Effects of micronised bamboo powder on growth performance, intestinal development, caecal chyme microflora and metabolic pathway of broilers aged 24–45 days. 2023. [CrossRef]
- Ogbuewu, I.P., O.O. Emenalom, and I.C. Okoli, Alternative feedstuffs and their effects on blood chemistry and haematology of rabbits and chickens: a review. Comparative Clinical Pathology, 2015. 26(2): p. 277-286. [CrossRef]
- Weingartner, O., et al., Alterations in cholesterol homeostasis are associated with coronary heart disease in patients with aortic stenosis. Coron Artery Dis, 2009. 20(6): p. 376-82. [CrossRef]
- Lee, E.J., et al., Proteasome inhibition protects against diet-induced gallstone formation through modulation of cholesterol and bile acid homeostasis. Int J Mol Med, 2018. 41(3): p. 1715-1723. [CrossRef]
- Oloruntola, O.D., et al., Neem, pawpaw and bamboo leaf meal dietary supplementation in broiler chickens: Effect on performance and health status. 2019. 43(2): p. e12723.
- Ge, Q., et al., Investigation of physicochemical properties and antioxidant activity of ultrafine bamboo leaf powder prepared by ball milling. Journal of Food Processing and Preservation, 2020. 44(7). [CrossRef]
- Luo, X., et al., Hydration properties and binding capacities of dietary fibers from bamboo shoot shell and its hypolipidemic effects in mice. Food Chem Toxicol, 2017. 109(Pt 2): p. 1003-1009. [CrossRef]
- Kim, J.S., et al., Effects of energy levels of diet and β-mannanase supplementation on growth performance, apparent total tract digestibility and blood metabolites in growing pigs. Animal Feed Science and Technology, 2013. 186(1-2): p. 64-70. [CrossRef]
- Wang, H., et al., Effects of replacing soybean meal by detoxified Jatropha curcas kernel meal in the diet of growing pigs on their growth, serum biochemical parameters and visceral organs. Animal Feed Science and Technology, 2011. 170(1-2): p. 141-146. [CrossRef]
- Kelly E. Heim, A.R.T., Dennis J. Bobilya, Flavonoid antioxidants: chemistry, metabolism and structure-activity relationships. Journal of Nutritional Biochemistry, 2002(13): p. 572–584. [CrossRef]
- Niu, Y., et al., Effect of supplemental fermented Ginkgo biloba leaves at different levels on growth performance, meat quality, and antioxidant status of breast and thigh muscles in broiler chickens. Poult Sci, 2017. 96(4): p. 869-877. [CrossRef]
- Wan, X.L., et al., Evaluation of enzymatically treated Artemisia annua L. on growth performance, meat quality, and oxidative stability of breast and thigh muscles in broilers. Poult Sci, 2017. 96(4): p. 844-850. [CrossRef]
- Shen, M., et al., Effect of Bamboo Leaf Extract on Antioxidant Status and Cholesterol Metabolism in Broiler Chickens. Animals (Basel), 2019. 9(9). [CrossRef]
- Montagne, L., J.R. Pluske, and D.J. Hampson, A review of interactions between dietary fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Animal Feed Science and Technology, 2003. 108(1-4): p. 95-117. [CrossRef]
- Xie, Z., et al., Effects of bamboo leaf extract on energy metabolism, antioxidant capacity, and biogenesis of small intestine mitochondria in broilers. 2023. 101: p. skac391. [CrossRef]
- Li, Q., et al., Retarding effect of dietary fibers from bamboo shoot (Phyllostachys edulis) in hyperlipidemic rats induced by a high-fat diet. Food Funct, 2021. 12(10): p. 4696-4706. [CrossRef]
- Kaminski, N.A. and E.A. Wong, Differential mRNA expression of nutrient transporters in male and female chickens. Poult Sci, 2018. 97(1): p. 313-318. [CrossRef]
- Wright, E.M. and E. Turk, The sodium/glucose cotransport family SLC5. Pflugers Arch, 2004. 447(5): p. 510-8. [CrossRef]
- Daniel, H., Molecular and integrative physiology of intestinal peptide transport. Annu Rev Physiol, 2004. 66: p. 361-84. [CrossRef]
- Kheravii, S.K., et al., Upregulation of genes encoding digestive enzymes and nutrient transporters in the digestive system of broiler chickens by dietary supplementation of fiber and inclusion of coarse particle size corn. BMC Genomics, 2018. 19(1): p. 208. [CrossRef]
Figure 1.
Effects of FBP on Carcass and organ weight of DYB Chickens. (a). Eviscerated Yield, (b) organ index, (c)weights of the breast, and thigh muscles (d). Weights of the kidney, liver, spleen, and heart. The mean of the variables is shown in bar graphs. Error bars show the standard error of the mean. The symbols *, **, *** are used to indicate significant differences between groups, and the different star symbols follow significantly different means (P < 0.05).
Figure 1.
Effects of FBP on Carcass and organ weight of DYB Chickens. (a). Eviscerated Yield, (b) organ index, (c)weights of the breast, and thigh muscles (d). Weights of the kidney, liver, spleen, and heart. The mean of the variables is shown in bar graphs. Error bars show the standard error of the mean. The symbols *, **, *** are used to indicate significant differences between groups, and the different star symbols follow significantly different means (P < 0.05).
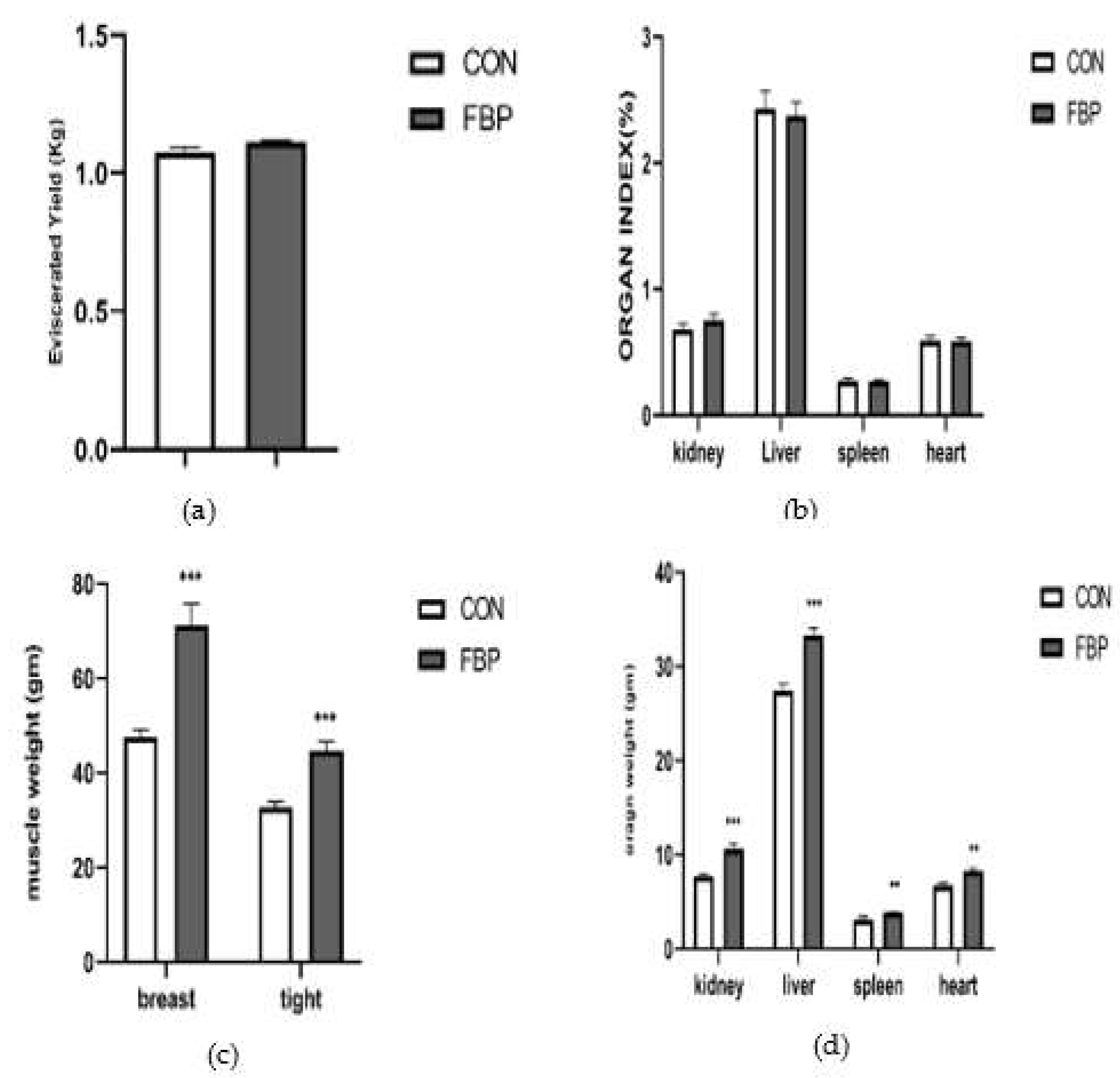
Figure 2.
Morphology of intestinal segments for control and FBP feed of DYB chickens. (a, b) duodenum CON and FBP, (c, d) jejunum CON and FBP, (e, f) ileum CON and FBP using 40x magnification with Scale bar100 µm. red arrows belong to villi height, villus width, crypt depth.
Figure 2.
Morphology of intestinal segments for control and FBP feed of DYB chickens. (a, b) duodenum CON and FBP, (c, d) jejunum CON and FBP, (e, f) ileum CON and FBP using 40x magnification with Scale bar100 µm. red arrows belong to villi height, villus width, crypt depth.
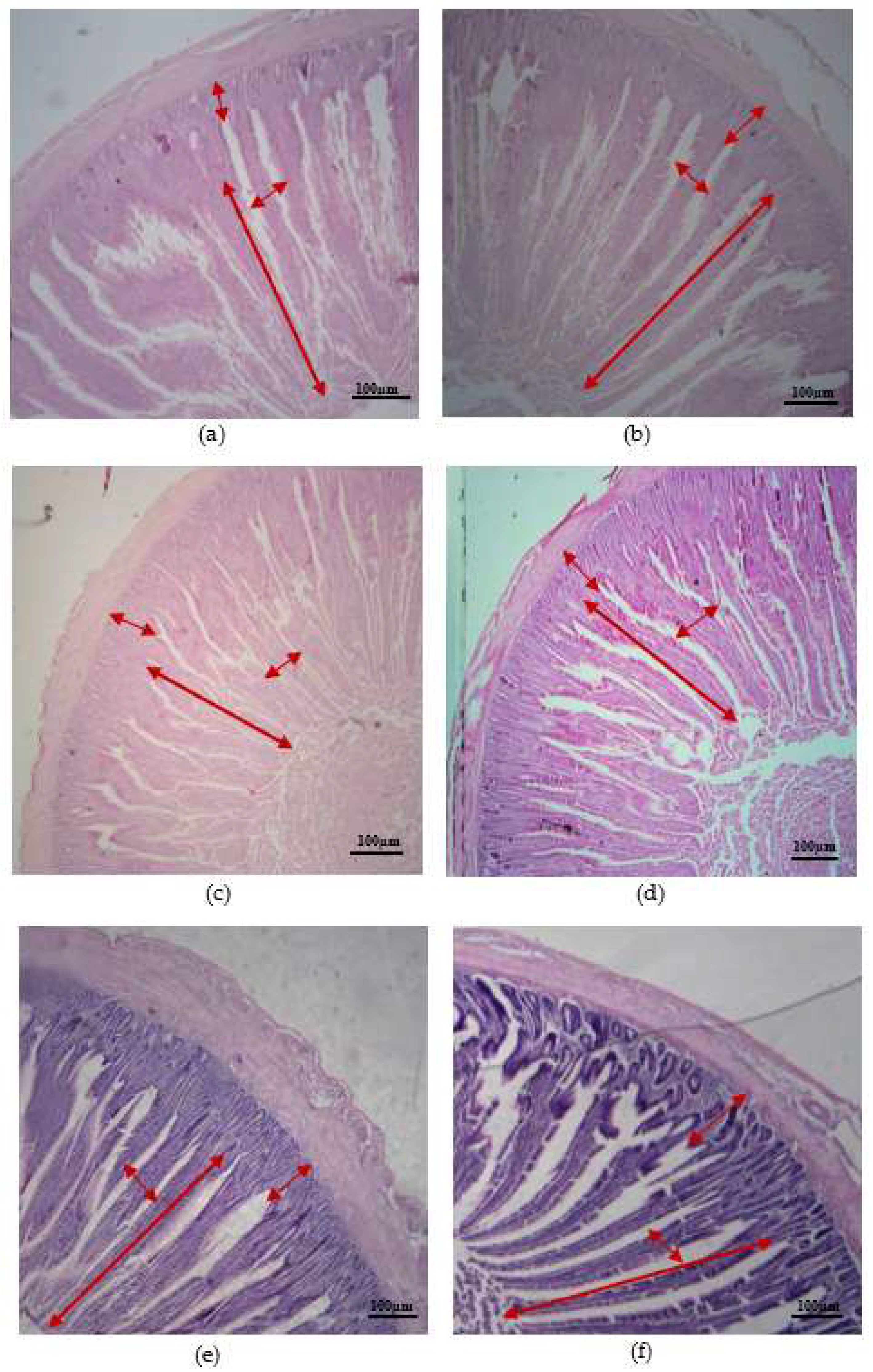
Figure 3.
The relative gene expression of mRNAs encoding (a) GLUT1,(b) GLUT2, (c) CAT1, (d) CAT2, (e) PepT1, (f) PepT2, and (g) LAT1. Expression is measured as 2^–ΔΔCt relative to expression at day 45. Data shown are means ±SEM.
Figure 3.
The relative gene expression of mRNAs encoding (a) GLUT1,(b) GLUT2, (c) CAT1, (d) CAT2, (e) PepT1, (f) PepT2, and (g) LAT1. Expression is measured as 2^–ΔΔCt relative to expression at day 45. Data shown are means ±SEM.
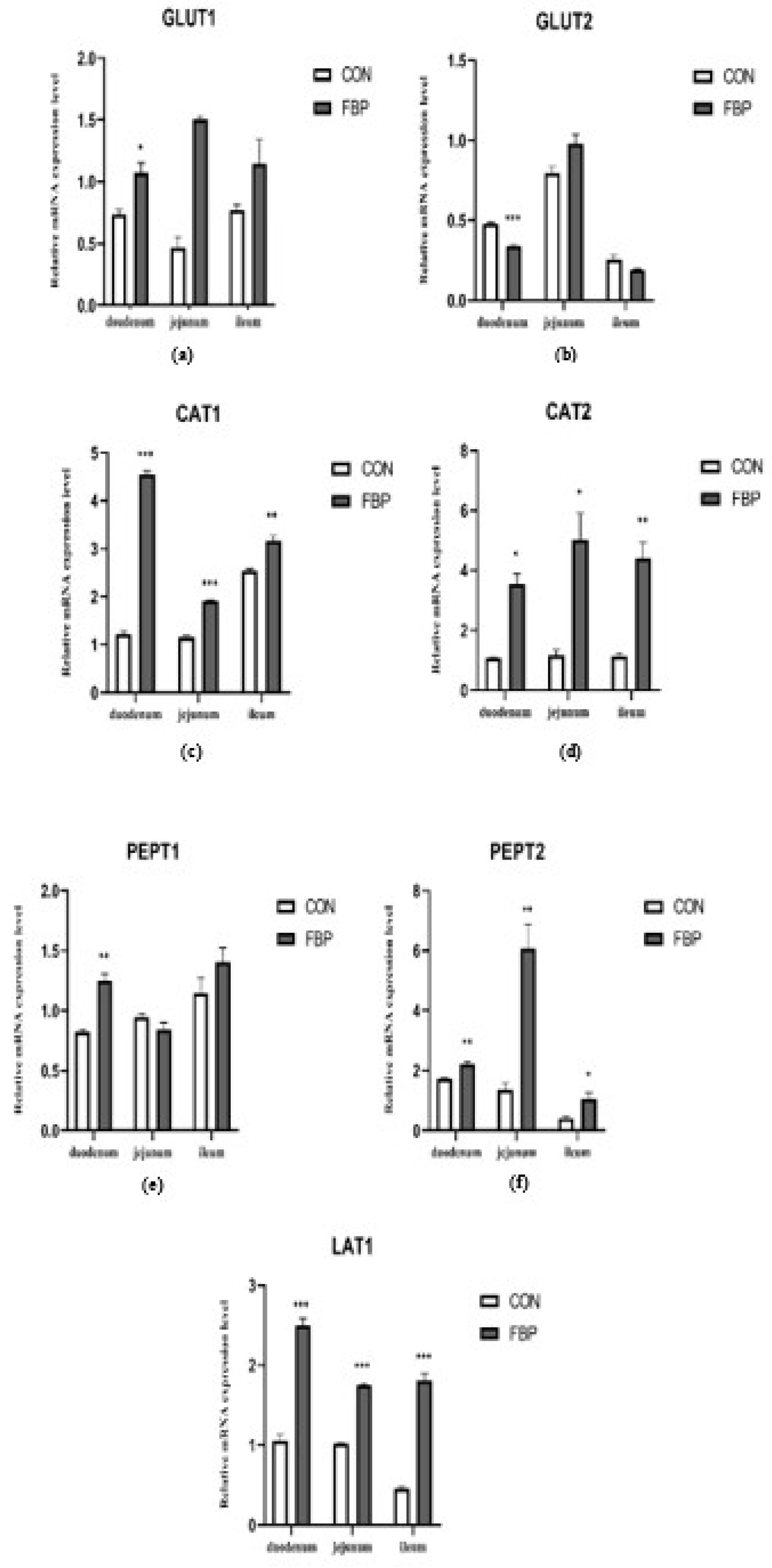

Table 1.
Ingredient composition and calculation of ingredients in the basal diet.
| Item | Starter phase 1-22 | Grower phase 23-45 |
|---|---|---|
| Corn | 406.6 | 297.3 |
| Wheat | 200 | 400 |
| Soybean meal | 223.1 | 57.3 |
| Sunflower | 30 | 50 |
| Rapeseed meal | 30 | 40 |
| Palm kernel meal | 0 | 20 |
| Corn gluten meal | 40 | 50 |
| Rice husk oil | 23.1 | 42.2 |
| Calcium bisphosphate | 14 | 10.3 |
| Limestone | 11.8 | 11.2 |
| Liquid methionine (88%) | 1.4 | 2.3 |
| Premix | 20 | 20 |
| Total | 1000 | 1000 |
| Calculation of nutrients | ||
| Metabolizable energy (MJ/kg) | 2956 | 3008 |
| Crude protein | 211.4 | 208.7 |
| Crude fat | 40.48 | 44.1 |
| Methionine | 4.86 | 7.64 |
| Lysine | 10.97 | 13.65 |
| Calcium | 9.89 | 9.62 |
| Available phosphorus | 4.85 | 5.1 |
Ingredient composition and calculated chemical analysis of the basal diets. Premix provided the following per kg of the diet: 50% Choline 5150 IU; Complex enzyme 15000 IU, L- Lysine 5500 IU; Rice bran meal 2400 IU; Tributyrin 38000 IU; Tryptophan 61000 IU; Threonine 9500 IU; Salt 560 IU; Probiotics 26000 IU; Organic mineral 1.5500 IU; Phytase 20000 IU.
Table 2.
Analyzed composition of the fermented bamboo powder (FBP).
| Item | FBP Composition (%) |
|---|---|
| Moisture | 11.21 |
| Crude protein | 17.07 |
| Coarse fiber | 17.66 |
| Crude fat | 3.48 |
| Coarse Ash content | 9.21 |
| Acid soluble protein | 7.13 |
| Acid washing lignin | 3.41 |
| Calcium (%) | 0.12 |
| Total phosphorus (%) | 0.02 |
Table 3.
Primer sequences were used for the RT-qPCR experiment.
| Gene | Reverse primer (5′-3′) | Amplicon Size (bp) |
Accession no |
|---|---|---|---|
| β-actin | F/ GCCCTCTTCCAGCCATCTTT R/CAATGGAGGGTCCGGATTCA | 107bp | NM_205518.2 |
| GLUT1 | F/ATGGGCTTCCAGTACATTGC R/TTTGTCTCCGGCACCTTGA |
110 bp | NM_205209.2 |
| GLUT2 | F/ GTTCCTGGCTGGTCTGATGG R/ TGGCGACCATGCTGACATAA |
107 bp | NM_207178.2 |
| CAT1 | F/ GCAAAGCGACTTTCCGGACT R/GCCTGTAAGAAACTCTGAGAAACC |
132 bp | NM_001398060.1 |
| CAT2 | F/TTGCTACATTGGTGGTGTCCT R/ TGAAACCAAGTGCCATCCAG |
198 bp | XM_040699004.2 |
| LAT1 | F/ TGCGTTACAAGAAGCCGGAG R/ CGATCCCGCATTCCTTTGGT |
129 bp | XM_046911929.1 |
| PepT1 | F/CCTTATCGTGGCTGGAGCAT R/TGGGCTTCAACCTCATTTGGA |
144 bp | NM_204365.2 |
| PepT2 | F/ TAGGTCATCCAACCTGCTCCT R/TGCCTGGAGGAGAAAGAACAC |
109 bp | NM_001319028.3 |
β-actin, beta-actin; GLUT1, Glucose transporter-1; GLUT2, Glucose transporter-2; CAT1, Cationic amino acid transporter-1; CAT2, Cationic amino acid transporter-2; LAT1 L-type amino acid transporter-1; PepT1, Peptide transporter-1; PepT2 Peptide transporter-2. F: forward primer; R: reverse primer.
Table 4.
Effect of dietary supplemental with FBP on performance in dwarf yellow-feathered broiler (DYB).
Table 4.
Effect of dietary supplemental with FBP on performance in dwarf yellow-feathered broiler (DYB).
| Parameters | CON | FBP | SEM | p-value |
|---|---|---|---|---|
| Initial BW(g) | 51.98 | 52.35 | 0.53 | 0.477 |
| 1-22 day(1% FBP) | ||||
| BW (g) | 424.83 | 466.88*** | 4.95 | <0.001 |
| BWG(g) | 364.53 | 448.30*** | 7.23 | <0.001 |
| ADFI (g) | 49.21 | 46.00 | 2.96 | 0.334 |
| ADG (g) | 17.36 | 21.35*** | 0.35 | <0.001 |
| FCR | 2.83 | 2.15 | 0.34 | 0.086 |
| Mortality rate % | 1.33 | 1.17 | 0.54 | 0.766 |
| 23-45 day(2%FBP) | ||||
| BW (g) | 1154.33 | 1288.67*** | 21.52 | <0.001 |
| BWG (g) | 728.33 | 813.07*** | 20.93 | <0.001 |
| ADFI (g) | 78.18 | 73.73 | 4.03 | 0.332 |
| ADG (g) | 31.67 | 35.35*** | 0.91 | <0.001 |
| FCR | 2.47 | 2.09* | 0.19 | 0.053 |
| Mortality rate % | 1.25 | 1.00 | 0.32 | 0.470 |
| 1-45 day | ||||
| BW (g) | 1154.33 | 1288.67*** | 21.52 | <0.001 |
| BWG (g) | 1141.03 | 1275.15*** | 26.05 | <0.001 |
| ADFI (g) | 70.71 | 66.57 | 3.72 | 0.330 |
| ADG (g) | 25.93 | 28.98*** | 0.59 | <0.001 |
| FCR | 2.73 | 2.30* | 0.22 | 0.054 |
| Mortality rate % | 2.58 | 2.08 | 0.23 | 0.069 |
CON, control group, fed antibiotic-free basal diet; FBP, group fed basal diet with FBP added with gradual levels. SEM: standard error of the means. The mean of the variables is shown in the same row. The symbols *, **, ***, is used to indicate significant differences between groups, and star symbols follow means that are significantly different (P < 0.05, P < 0.01, P < 0.001).
Table 5.
Effects of dietary FBP on intestinal histology of (DYB).
| Item | CON | FBP | SEM | p-value |
|---|---|---|---|---|
| duodenum | ||||
| VH(mm) | 0.58 | 0.67** | 0.02 | 0.003 |
| CD (mm) | 0.10 | 0.17 | 0.04 | 0.096 |
| VCR | 6.63 | 4.81 | 1.08 | 0.153 |
| VA (mm2) | 1.66 | 1.92 | 0.21 | 0.263 |
| jejunum | ||||
| VH (mm) | 0.46 | 0.59** | 0.03 | 0.007 |
| CD (mm) | 0.09 | 0.13** | 0.01 | 0.007 |
| VCR | 5.12 | 4.66 | 0.45 | 0.347 |
| VA (mm2) | 1.80 | 2.01 | 0.27 | 0.474 |
| ileum | ||||
| VH (mm) | 0.60 | 0.74* | 0.05 | 0.038 |
| CD (mm) | 0.34 | 0.49* | 0.04 | 0.021 |
| VCR | 1.85 | 1.52 | 0.20 | 0.142 |
| VA (mm2) | 1.55 | 2.86* | 0.50 | 0.048 |
VH, villus height; CD, crypt depth; VCR, villus height/crypt depth ratio; VA, villus area; CON, chicken fed with basal; FBP, a basal diet supplemented with fermented bamboo powder through 45 day with gradual levels. SEM: standard error of mean difference, The mean of the variables is shown in the same row, The symbols *, **, *** are used to indicate significant differences between groups, and the different star symbols follow means that are significantly different (P < 0.05, P < 0.01, P < 0.001).
Table 6.
Effect of dietary supplemental with FBP on serum biochemical indexes in DYB chicken.
| Parameters | Dietary treatment | SEM | p-value | |
|---|---|---|---|---|
| CON FBP | ||||
| TC(mmol/L) | 3.38 | 2.78 | 0.54 | 0.323 |
| TG(mmol/L) | 0.80 | 0.57* | 0.07 | 0.025 |
| GLU(mmol/L) | 7.25 | 6.55 | 0.70 | 0.079 |
| LDL(mmol/L) | 1.14 | 0.81 | 0.32 | 0.406 |
| HDL(mmol/L) | 1.87 | 2.25 | 0.16 | 0.133 |
| TP(g/L) | 23.80 | 26.35 | 1.24 | 0.108 |
| ALB(g/L) | 13.40 | 14.57 | 1.06 | 0.385 |
| GLB(g/L) | 11.63 | 12.77 | 1.57 | 0.510 |
| sCr (umol/L) | 10.73 | 8.47 | 3.14 | 0.511 |
| UREA(mmol/L) | 1.22 | 0.59 ⃰ | 0.14 | 0.011 |
| AST(U/L) | 303.30 | 259.83 | 29.87 | 0.277 |
| ALT(U/L) | 3.33 | 2.30 ⃰ | 0.31 | 0.028 |
TC: total cholesterol; TG: triglyceride; GLU: glucose; LDL: low-density lipoprotein cholesterol; HDL: high-density lipoprotein cholesterol; TP: total protein; ALB: albumin; GLB: globulin; sCr: serum creatinine; AST: aspartate transaminase; ALT: alanine transaminase; the mean of the variables is shown in the same row with significance (P < 0.05). The symbol * indicates significant differences between groups, SEM: standard error of the means.
Table 7.
Effect of dietary FBP on serum antioxidant index of (DYB) chicken.
| Item | CON | FBP | SEM | p-value |
|---|---|---|---|---|
| SOD (U/ml) | 48.79 | 122.90** | 5.28 | 0.005 |
| GSH-PX (U/mg) | 119.64 | 136.63 | 7.31 | 0.146 |
| CAT(nmol/mg prot) | 40.73 | 116.38** | 3.84 | 0.003 |
| MAD(nmol/mg prot) | 2.29 | 0.49** | 0.19 | 0.011 |
| HO-1(pg/ml) | 255.37 | 243.03 | 22.01 | 0.631 |
SOD: Superoxide dismutase; GSH-Px: Glutathione peroxidase; CAT: catalase; MDA: malondialdehyde; Heme Oxygenase HO-1. CON: basal diet, FBP group, basal diet adding gradual levels of FBP. The mean of the variables is shown in the same row with significant (P < 0.05). The symbols *, is used to indicate significant differences between groups, SEM: standard error of the means.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



