
Submitted:
03 October 2024
Posted:
04 October 2024
You are already at the latest version
Abstract
Abnormal platelet functions are associated with human morbidity and mortality. Maintaining physiological platelet function is critical to hemostasis and preventing platelet-associated diseases such as cardiovascular disease, cancer metastasis, immune disorders, hypertension, diabetes, sickle cell disease, inflammatory bowel disease, sepsis, rheumatoid arthritis, myeloproliferative disease, and Alzheimer’s disease. Platelets become hyperactive in obesity, diabetes, a sedentary lifestyle, hypertension, pollution, and smokers. Platelets, upon activation, can trawl leukocytes and progenitor cells to the vascular injury sites. Platelets also release various pro-inflammatory, anti‐inflammatory, and angiogenic factors and shed microparticles in the circulation. The challenge for therapeutic intervention in these diseases will be identifying factors that preferentially block specific targets involved in platelets' complex contribution to inflammation or tumor progression while leaving their hemostatic function at least partially intact. Since antiplatelet drugs such as aspirin are not recommended as primary preventives, it is essential to use alternative safe platelet inhibitors. Potent natural antiplatelet factors were investigated, including n-3 fats, polyphenols, and other bioactive compounds. This review describes the anti-platelet bioactive compounds in food that can prevent platelet hyperactivity and thus may prevent several platelet-mediated diseases, including cardiovascular disease.
Keywords:
Platelet function
; Functional food
; Bioactive compounds
; CVD
; Tomato extract
; Kiwiffruit extract Fruitflow®
; Polyphenols
; n-3 fatty acids
; Hypertension
; Kiwifruit extract
; pla
Introduction
Maintaining the physiological function of blood platelets is crucial for overall hemostasis and several physiological processes [1,2]. Reduced activation response of blood platelets can lead to excessive bleeding, whereas hyper-activation of platelets contributes to thrombotic complications. Platelet hyperactivity is associated with several processes, such as inflammatory and immune responses, angiogenesis, atherosclerosis, regeneration of liver tissue, and cancer metastasis[3,4,5,6]. Platelet hyperactivity is related to cardiovascular disease (CVD) and many other human diseases, such as cancer, renal diseases, Alzheimer’s disease, and microorganism infections. However, the mechanisms are not well known [4,6,7,8]. The Framingham Heart Study revealed that persistent platelet hyperactivity is also associated with future arterial and venous thrombosis [9]. Individuals with diabetes, a sedentary lifestyle, obesity, and insulin resistance have platelet hyperactivity, which contributes to atherosclerotic plaque development and other platelet-mediated diseases [10,11,12]. Figure 1 depicts the association of platelet hyperactivity with CVD. Platelet hyperactivity is not only associated with CVD and stroke because platelet inhibitors slow down also cancer progression and related morbidities [13]. Platelets play critical roles in inflammation and immune responses [14,15] by release of IL-6 and NFkB[16], circulating platelet microparticles [17,18], and CRP levels [15,19]. The hyperinflammatory status can compromise the body’s response to external stresses and predispose to diabetes or atherosclerosis.
Beyond the hemostatic process, platelets are involved in many processes through complicated interactions with many cells. Aspirin is the most used antiplatelet therapy for the secondary prevention of CVD [20] , even though it causes several serious side effects. There are several reasons, including the fact that 25-30% of people are aspirin resistant and the side effects, making aspirin unsuitable for the primary prevention of CVD [21]. Aspirin prophylaxis is not recommended in individuals without CVD, as per the European Guidelines on CVD prevention [22].
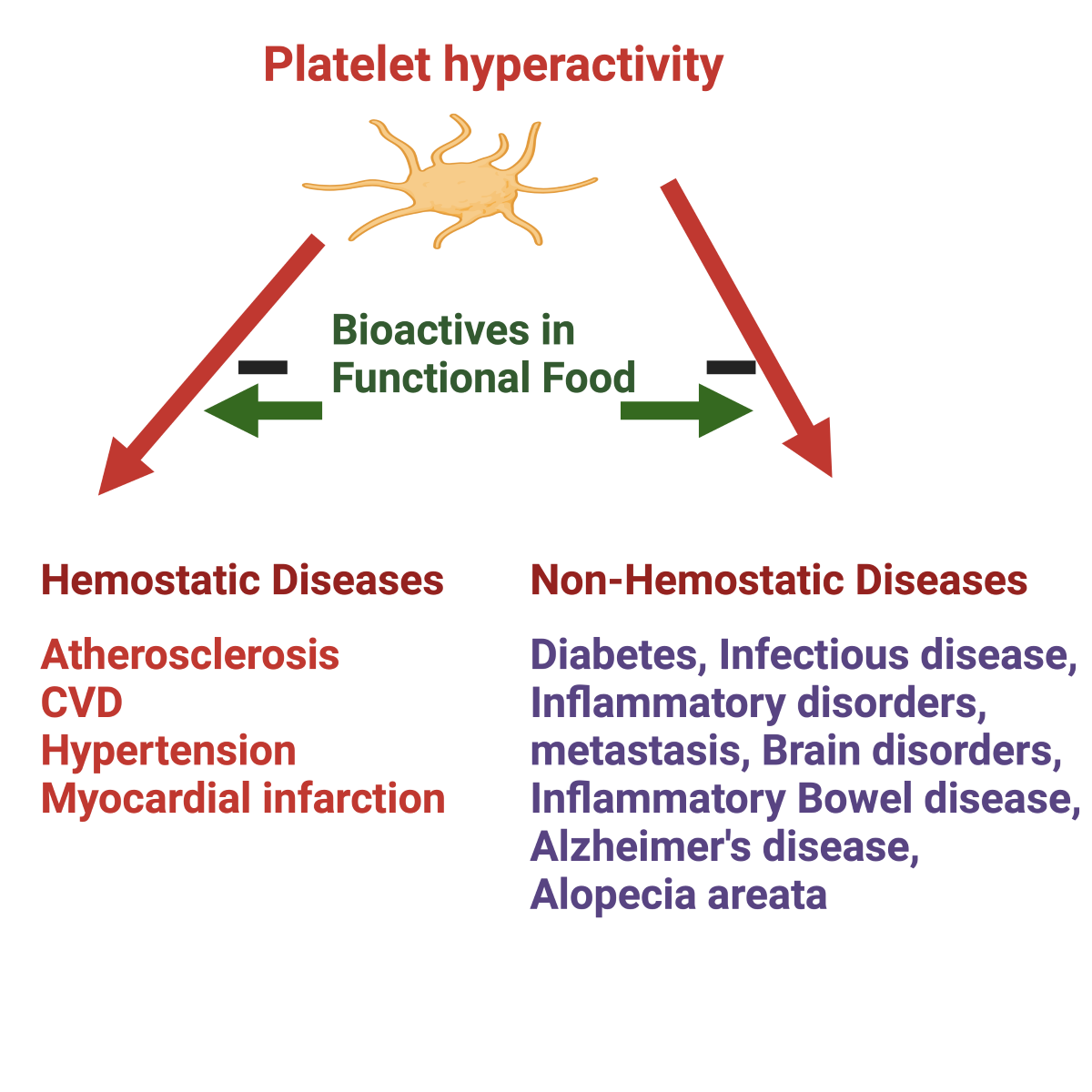
Maintaining regular platelet activity is critical in preventing platelet-associated diseases such as CVD, cancer metastasis, immune disorders, hypertension, diabetes mellitus, sickle cell disease, inflammatory diseases, bowel disease, rheumatoid arthritis, myeloproliferative disease, Alzheimer’s disease, the understanding the causes and mechanisms of action of the modifying factors of platelet hyperactivity are critical in preventing these diseases.
Consequently, there is growing interest in naturally occurring anti-platelet inhibitors that people can regularly consume. Many dietary antiplatelet components are identified that can reduce platelet hyperactivity without side effects [23,24,25,26,27]. Bioactive compounds of dietary origin with antiplatelet activity become operative when continued platelet hyperactivity in response to inappropriate nutritional and lifestyle factors or exposure to smoke/air pollution [28,29,30], hyperglycemia and hyperlipidemia [31], and specific exercise patterns [32]. This review highlights the development of bioactive compounds and their roles in preventing the hyperactivity of blood platelets and associated diseases. Figure 2 shows that platelet hyperactivity is related to hemostatic and non-hemostatic diseases. The relevance of platelet hyperactivity in the context of many diseases, including the growing epidemic nature of CVD, has strongly suggested the use of more natural approaches in the primary prevention of degenerative diseases.
Platelet Hyperactivity in Many Pathological Conditions
Human blood platelets are involved in hemostasis and thrombosis, including inflammation, infection, immunobiology, cancer metastasis, wound repair, and angiogenesis [33]. Since platelet hyperreactivity increases in hyperlipidemia, hyperglycemia, oxidative stress, and cancers [34,35], understanding how platelets become hyperresponsive and contribute to CVD under these conditions is needed to prevent CVD [36].
Activated platelets secrete mRNA, nucleotides, enzymes, metal ions, microparticles, mitochondria, ATP, ADP, growth factors, chemokines, cytokines, protease inhibitors, adhesive glycoproteins serotonin, histamine, γ-aminobutyric acid, glutamate, epinephrine, dopamine, histamine, factor V, calcium and many others [37,38,39]. Platelet microvesicles contain multiple bioactive molecules, RNA, and proteins involved in these processes [40,41]. The release of these factors by the activated platelets changes the cell membrane architecture, including blood vessel walls, affecting the immediate milieu [24]. Platelet activation, aggregation, and the subsequent generation of an occlusive intra-arterial thrombus are essential in developing CVD and platelet-mediated diseases [42,43].
Pathological Conditions Such as Metabolic Syndrome, Insulin Resistance, Obesity, Hypertension and Tumors Enhance Platelet Hyperactivity
Metabolic syndrome (MetS) is a cluster of conditions related to abdominal obesity, insulin resistance, hyperlipidemia, hypertension, and inflammation [44]. MetS is associated with increased platelet reactivity, probably caused by associated hyperglycemia, dyslipidemia, and low-grade systemic inflammation. Platelet hyperactivity in MetS is induced by inflammation, obesity, dyslipidemia, hypertension, oxidative stress, and other metabolic disease[45]. Higher plasma levels of fibrinogen, PAI-1, thrombin, von Willebrand factor, factor VII, and other coagulation factors, as well as significant platelet aggregation, are also observed MetS [46]. The MetS components increase platelet activation by increasing calcium concentration and activating arachidonic acid,20:4n-6 (ARA) signaling pathways. Platelet hyperactivity in dyslipidemia contributes to the thrombotic risk [47]. Platelet activation in dyslipidemia is induced via classic agonist receptor signaling pathways and pattern recognition receptors[47]. Platelets are stimulated by oxidized phospholipids present in low-density lipoproteins (LDLs) via specific pattern recognition receptors [48]. CD36/fatty acid translocase is platelets' scavenger receptor for oxidized lipids [48].
Oxidative stress may promote platelet hyperactivity by reducing physiologically accessible NO in MetS. Cigarette smoking and sedentary lifestyles also increase oxidative stress and platelet hyperactivity[29]. Platelets oxidize LDL by stimulating the production of reactive oxygen species (ROS) and degrading the LDL receptors by releasing proprotein convertase subtilisin/kexin type 9. Thus, platelet dysfunction and hyperlipidemia stimulate atherogenesis[49]. Moreover, reduced antiplatelet response to aspirin and higher platelet sensitivity toward agonists are observed in MetS. Therefore, hypolipidemic drugs modulate platelet function, whereas antiplatelet drugs normalize lipid metabolism. High levels of plasma leptin in obesity cause increased platelet aggregability[50]. The overall effect of MetS and obesity contribute to platelet activation [51].
Platelet function is influenced by ROS and nitric oxide (NO) metabolism [52,53]. Platelet aggregation is associated with increased glutathione disulfide levels and oxygen utilization. Changes may influence platelet-dependent thrombus formation in platelet or vascular redox status, antioxidants, and ROS and nitrogen species [54]creation. Hyperglycemia, dyslipidemia, and insulin resistance in diabetic conditions induce platelet hyperactivity, activation of the vascular wall, inflammation, and endothelial dysfunction. Hyperglycemia also causes the overproduction of superoxide and activates protein kinase C and nuclear factor kB (NFkB) [55]. NFkB stimulates the production of several inflammatory cytokines and increases cell adhesion molecules, such as CD40 ligand (CD40L). CD40L induces the production and release of pro-inflammatory cytokines [56]. Since activated platelets express CD40L, indicating a link between hemostasis and inflammation [57].
Diabetic platelets have an increased intrinsic activation profile, whereas these platelets have reduced influence of endogenous inhibitors and aspirin. Platelet hyperactivity in diabetes mellitus may result from hypersensitivity to agonists and loss of anti-aggregatory mechanism. Even though the usefulness of antiplatelet drugs in type 2 diabetes is not supported, Diabetic patients still use low-dose aspirin or other antiplatelet drugs as a potential therapeutic therapy [58,59]. In diabetes mellitus, there is an imbalance between platelet inhibitors such as NO and prostacyclin (PGI2) and increased production of platelet-activating and vasoconstricting substances. Decreased endothelium synthesis of PGI2 [60] and reduced synthesis of NO was observed in type 2 diabetes mellitus[61]. Platelets express receptors for adhesion and increased synthesis of TxA2, thrombin, and deranged calcium metabolism in diabetes. Platelets shed more platelet microparticles (PMPs) in type 2 diabetics may play a role in inflammation and thrombosis. P-selectin-positive PMPs may aid in the recruitment and adhesion of leucocytes, and platelets initiate the atherosclerosis process.
Spontaneous platelet activation is involved in the hypertension [62,63,64]. High blood pressure also stimulates platelet activation via increasing shear force on circulating platelets. Increased blood viscosity in patients with high blood pressure might also contribute to platelet hyperactivity. Under high-pressure flow, enhanced degranulation of platelets occurs [65]. Atherosclerotic lesions induce platelet activation, possibly due to the dysfunctional endothelium and local flow disturbances [66]. Numerous metabolic and physiologic changes in platelet reactivity in hypertension. Indeed, many studies have shown that various parameters of platelet activation are normalized with the treatment of hypertension[67]. Platelets from hypertensive patients tend to form aggregates [68], increased secretion of plasma β-thromboglobulin [69], high soluble and membrane expression of P-selectin[70], and increased intracellular levels of calcium[71]. However, the association between these markers and the degree of hypertension or responses to therapy are not well known[72].
Endothelial dysfunction is associated with the reduced production of platelet inhibitors and vasodilators, NO [73]. Therefore, the reduced synthesis of NO increases hypertension and platelet hyperactivity [74]. Similarly, dysfunctional endothelium produces less PGI2 and increases platelet activity. Dysfunction of the endothelium in hypertension has a cross-talk with hyperactive platelets [67]. Similarly, increased platelet activation/aggregation in response to endothelin could contribute to thrombosis [75]. Hyperactive platelets are a critical earlier trigger for cancer-associated thrombotic events [76,77]. Interactions between platelets and tumor cells result in hyperactivity or activation of circulating blood platelets.
Modulation of Platelet Activity by Bioactive Compounds of Dietary Origin
It is evident that blood platelets play an essential role in maintaining hemostasis and, under hyperactive states, may also exert a pathophysiologic role and thus contribute to atherosclerosis and other diseases. In recognition of the roles of platelet hyperactivity in the atherosclerosis process, antiplatelet therapy has become essential in managing CVD. Indeed, antiplatelet treatment has significantly reduced the incidence of primary and secondary cardiac events in secondary prevention trials [78]. However, the combination of lipid-lowering measures, blood pressure monitoring, and administration of antiplatelet agents decreased the incidence of cardiac events and strokes by up to 80% in people above 55 years old [79]. However, aspirin’s effects are relatively weak and can cause severe gastrointestinal disturbances and bleeding problems in some individuals [80]. Given the increasing incidence of CVD and the established role of platelet activity in mediating atherosclerosis, it makes sense to identify alternate means of maintaining platelet activity that does not increase bleeding time and are safe, effective and well-tolerated. This notion prompted research designed to understand the role of dietary ingredients in modulating platelet hyperactivity – i.e., dietary antiplatelet components.
Indeed, extensive in vitro and preclinical research demonstrates the efficacy of various food components in reducing platelet hyperactivity. However, there are only a small number of well-designed studies consistently demonstrating an antiplatelet benefit of only a few select dietary ingredients in healthy human subjects with measurable clinical outcomes. These include fish and fish oil, flavonoids in cocoa, and garlic. Therefore, this review's objectives are two-fold: summarize the available clinical science to substantiate the antiplatelet efficacy of these ingredients and highlight recent and emerging clinical evidence for a new dietary antiplatelet component derived from fruits, vegetables, and fish oils. The antiplatelet benefits of these ingredients on humans, along with clearly defined outcomes related to the primary prevention of vascular events, namely platelet function, are discussed. Therefore, there is an increased interest in identifying bioactive compounds from edible sources with beneficial health effects, efficiency, and fewer adverse effects [4,24,81].
N-3 Fatty Acids
Since the first cross-cultural epidemiological studies in the 1970s, the CVD preventive role of n-3 long-chain polyunsaturated fatty acids (LCPUFAs), in particular docosahexaenoic acid,22:6n-3 (DHA), and eicosapentaenoic acid,20:5n-3 (EPA) emerged [82]. Fish oil supplementation reduced platelet aggregation[83]. Essential fatty acids, linoleic acid,18:2n-6 (LA), and alpha-linolenic acid,18:3n-3 (ALA), in turn, compete for desaturase and elongase enzymes for their metabolism to LCPUFAs. A higher dietary intake of ALA compared to LA leads to an enhanced synthesis of EPA and DHA metabolites, resulting in a higher production of TxA3 than the pro-aggregatory ARA-derived TxA2. Prostaglandin H2 is a precursor of other prostaglandins and thromboxane [84]. Anti-platelet mechanisms of action by DHA and EPA are thought to be mediated via inhibition of phospholipase A2 activity, which would diminish the liberation of ARA from platelet plasma membrane phospholipids; inhibition of platelet cyclooxygenase 1 (COX-1), which would decrease the conversion of ARA to TxA2; competition with ARA for platelet COX-1 and formation of less active TxA3, or by alteration of platelet membrane fluidity state; and inhibition of TxA2 induced platelet aggregation and decreased receptor affinity for TxA2[85].
N-3 LCPUFAs have gained considerable attention for their ability to improve cardiovascular health and prognosis. In contrast, the n-6 fatty acids such as linoleic acid,18:2n-6 and ARA may increase platelet aggregation. Both EPA and DHA are incorporated into platelet membrane phospholipids replacing ARA, thus decreasing platelet aggregation via decreased production of ARA-derived platelet-aggregating eicosanoids [23,86]. EPA also competes with ARA for COX, reducing ARA-derived metabolites. Thus, EPA minimizes the synthesis of the pro-aggregatory ARA-derived TXA2[87,88]. EPA and DHA also alter membrane fluidity, regulating receptor expression and thrombin generation [89,90,91]. Resolvins produced from both EPA (resolvins E) and DHA (resolvins D) reduce and inhibit thromboxane-induced platelet aggregation [92]. Oxylipins (11-HDHA and 14-HDHA) derived from DHA also inhibited protein kinase A-mediated platelet activation and regulated agonist-induced platelet aggregation [93]. DHA and its 12-LOX-derived oxylipins also modulated collagen-induced platelet aggregation.
Several studies such as in vitro, clinical trial, observational, epidemiological and animal studies investigated the impact of seafood consumption and n-3 fatty acids on CVD. Human clinical studies showed that n-3 LCPUFAs impart antiplatelet effects. Fischer et al. (1983) [94] showed that EPA supplementation at 4 gm per day through cod liver oil in eight healthy adult males for 25 days reduced platelet aggregation. In contrast, Gibney and Bolton-Smith (1988) demonstrated that consumption of a low dose of EPA (2.25 gm) plus DHA (1.35 gm) per day did not affect platelet aggregation in response to collagen or low levels of ADP in eight healthy adult males for six weeks. However, there was a significant increase in platelet aggregation when higher levels of ADP were used.
Additionally, platelet TxB2 production was affected [95]. In contrast to other studies, the lack of DHA and EPA effect on platelet function may be due to the low dose of the n-3 fatty acids used in this study. In a randomized, double-blind study by Li and Steiner (1991), 3, 6, or 9 gm EPA per day supplementation to fifteen healthy adults for three weeks reduced platelet adhesion to fibrinogen within the first week. Under low shear rates conditions, the 3 and 6 gm per day inhibited platelet adhesion by 81.7% and 85.5%, respectively, and the 9gm dose inhibited rates of platelet adhesion at a similar level to the 3-gm dose. In contrast, platelet adhesion was maximally inhibited at high shear rates by the 3 gm of EPA per day [96].
Pirich and colleagues (1991) also demonstrated that supplementing twenty healthy adult subjects with EPA (216 mg) plus DHA (140 mg) per day for six weeks reduced TxB2 synthesis and significantly increased platelet survival time [97]. Indeed, the levels of DHA and EPA used were remarkably low, considering the positive benefits observed. However, this study's fish oil supplementation regimen also provided 390 mg of gamma-linolenic acid,18:3n-6. So, whether the effects observed above were due to the combination of DHA and EPA or the three fatty acids is unclear.
In 2002, Svaneborg and colleagues showed that 10 gm of fish oil per day to eighteen healthy adults significantly decreased TxA2 production and showed no change in the bleeding time [98]. In a more extensive randomized, double-blind study, Eschen et al. (2004) reported that EPA(3gm) and DHA (2.9gm) per day to sixty healthy adults for 12 weeks significantly decreased plasma P-selectin levels, used as an index of platelet activation [99]. Consuming n-3 fatty acids for four weeks reduced platelet integrin (GPIIa/IIIb) activation, levels of fibrinogen, and factor V, but no effect on coagulation in healthy subjects [100]. In 2008, Din and colleagues showed that 500 gm per day of mackerel consumption for four weeks reduced platelet-monocyte aggregation by ~35%, which returned to baseline after cessation of fish consumption [101]. However, Eschen et al. (2004) found no change in soluble P-selectin levels [99]. More recently, an open-label 4-week sequential study showed that EPA (1.86 gm) and DHA (1.5 gm) per day to thirty healthy adults decreased GPIIa/IIIb activation and significantly reduced the expression of P-selectin[102]. Vericel and colleagues (1999) demonstrated that a low dose of EPA (30 mg) plus DHA (150 mg) per day for 42 days resulted in a reduction of platelet aggregation but not significantly [103]. These data contrast those published by Croset et al. (1990) [104] and Driss et al. (1984)[105]. Herein, both investigators showed that low supplementation of EPA in elderly subjects significantly decreased platelet aggregation. In the study by Croset et al. (1990)[104], healthy elderly subjects were provided with the EPA at (1984), which provided 150 mg EPA per day for one month[105]. The discrepancy in the findings between the Vericel et al. study (1999) and that of Driss and colleagues (1984) and Croset and colleagues (1990) is most likely due to the level of EPA supplementation. In 2009, Guillot and coworkers[106] demonstrated that 400 and 800 mg DHA per day for two weeks to twelve healthy elderly subjects significantly reduced platelet reactivity and TxA2 production. Still, this effect was reversed when 1600 mg DHA per day was consumed. DHA intake at 200 mg/day did not affect platelet reactivity or TxA2 production. The lack of an effect of very high intakes of DHA and EPA in healthy elderly subjects on platelet activity is consistent with findings from Larson et al. (2008), who showed no inhibition of platelet aggregation with 4 gm of n-3 LCPUFAs per day for 4 weeks[102]. Most studies support the antiplatelet effect of marine fish or oils in healthy human subjects; although the dose used appears to be key in dictating a benefit, low and high doses do not always show a benefit.
Indeed, a meta-analysis of 15 randomized controlled trials in humans has confirmed that n-3 LCPUFAs inhibit platelet aggregation [88]. EPA has also reduced P-selectin, oxidized LDL (ox-LDL) antibodies, and glycoprotein IIb/IIIa expression on the platelet membranes [107]. Consuming 6.6 g of n-3 LCPUFAs reduced serum P-selectin expression, which may help to decrease platelet activity [99]. However, the increase in the n-6/n-3 ratio has shifted the balance into a pro-aggregatory state.
The data on the primary and secondary prevention of CVD using n-3 LCPUFAs are controversial. The latest meta-analysis suggested that regular intake of fish oil may be a risk factor for atrial fibrillation and stroke among the general population. However, it may be beneficial in certain CVD cases [108]. The published data support the use of n-3 LCPUFAs in reducing the risk of CVD or CVD-related death. The expert guidelines consistently recommend consuming at least 250 mg/day of n-3 LCPUFAs or at least two servings/week of oily fish for the general population [109]. However, the beneficial role of n-3 LCPUFAs in CVD is still not sure [110,111,112]. The clinical trials showed limited or no effect on cardiovascular health, which may be the result of heterogeneous populations, background n-3 LCPUFAs status, and consumption discrepancies [113].
However, it is considered appropriate for all individuals to consume n-3 LCPUFA daily as there are some benefits associated with CVD risk reduction and no adverse effects at the recommended levels. Epidemiological and intervention studies showed that dietary intakes between 0.4 g and 1.8 g/day of n-3 LCPUFAs are optimum for cardiovascular health. Several systematic reviews, meta-analyses, and experts examined the role of n-3 LCPUFAs on the cardiovascular system [109,114,115,116,117,118,119,120]. Expert recommendations generally support the cardiovascular health benefit of n-3 LCPUFAs with a recommendation for a daily intake of 500 mg as DHA and EPA or 1–2 servings of seafood per week. The US Dietary Advisory Committee recommends at least two servings of seafood per week. American Heart Association recommends one to two seafood meals per week to prevent the risk of congestive heart failure, coronary heart disease, ischemic stroke, and sudden cardiac death [116].
Metanalyses of several randomized trials indicated that n-LCPUFAs positively affect CVD outcomes for primary and secondary prevention. Marine n-3 LCPUFAs also improve CVD risk factors, including blood pressure, plasma lipids, and inflammation; however, many physicians do not recommend n-3 fatty acids, mainly due to controversial results in randomized trials.
Plant Polyphenols
Plant polyphenols protect the cardiovascular system by reducing platelet hyperactivity and oxidative stress [121]. Polyphenols can impart several health benefits, including protection from atherosclerosis, thrombosis, inflammation, and platelet hyperactivity [121,122,123,124]. The amount of polyphenols varies in plant-based foods, including fruits, chocolate, beverages, vegetables, and grains[125]. All polyphenols have the basic structure of an aromatic ring with a hydroxyl group. Polyphenols are mainly two groups, flavonoids, and nonflavonoids, despite numerous classes based on their carbon skeleton, phenol rings, and structural elements [125]. Flavonoids are further classified into flavonols, isoflavones, flavanones, anthocyanins, and flav-3-ols, whereas non-flavonoids consist of phenolic acids, hydrolyzable tannins, and stilbenes.
There is accumulating epidemiological and clinical evidence demonstrating that flavanol-rich foods can positively influence hemostasis through mechanisms that directly or indirectly affect platelet function and, in this way, ultimately decrease the risk for hypertension and stroke [126,127]. Plant-derived foods and beverages, such as red wine, tea, grapes, grape juice, cocoa, and chocolate, are typically flavonol-rich. Hamed et al. (2008) proposed that flavonoids can competitively inhibit binding to platelet-derived growth factor and TxA2 receptors, negating the effects of ARA - and collagen-induced platelet aggregation [128]. In addition to their antioxidant activity, flavonoids inhibit platelet lipoxygenase or reduce phospholipase C activity [129].
Most clinical studies were conducted with cocoa because cocoa products contain more flavonoids than teas and wines [130,131]. Wright et al. [132] demonstrated that functional groups of the flavonoid skeleton regulated the polyphenol inhibition of collagen-induced aggregation. The flavonoid aglycone increases the hydroxyl groups on the A and B rings, decreasing flavonoid activity. In contrast, the presence of O-methyl groups can increase activity. On the other hand, a hydroxyl group at position C3 can increase flavonoid activity [133]. In a study of 30 healthy adults, Rein et al. (2000) demonstrated that a cocoa beverage containing 18.75 g procyanidin and a total epicatechin content of 897 mg inhibited platelet aggregation and suppressed membrane GPIIa/IIIb expression and reduced the shedding of PMPs. PMPs are hemostatically active, phospholipid-rich microvesicles formed by activated platelets [134]. Using the same level of flavonoids in another study of sixteen healthy young men, Pearson and coworkers (2002) showed no effect on GPIIa/IIIb activity or P-selectin expression. Heptinstall et al. (2006) evaluated the impact of different amounts of cocoa-containing flavonoids in twelve healthy volunteers[126]. Study participants consumed 80 mg, 300 mg, 600 mg, or 900 mg cocoa flavanols. Cocoa drinks containing 600 or 900 mg of cocoa flavanols significantly inhibited platelet aggregation [126].
Flavonols are present in red wine, tea, grapes, grape juice, cocoa, and chocolates. Hermann and colleagues (2006) showed 40 gm of dark chocolate consumption by 20 healthy adult male smokers significantly improved flow-mediated dilation and decreased platelet adhesion[131]. Hamed and coworkers also evaluated the benefits of cocoa flavanols through dark chocolate consumption [128]. Consumption of dark chocolate (100 g) for one week (~70% cocoa; 700 mg flavonoids) significantly decreased GPIIa/IIIb expression [128]. Both the studies mentioned here are consistent with the findings of Innes et al. (2003) [135]. Consuming cocoa-related products in moderate amounts inhibited platelet aggregation [136].
Several reviews documented the impact of polyphenols on platelet function. Using different agonists, polyphenols' antithrombotic properties are determined by measuring their inhibitory effect on whole blood or platelet-rich plasma aggregation [23,26,137,138,139]. Table 1 summarizes the impact of the consumption of polyphenols on platelet aggregation ex vivo. Cocoa is a rich flavonoid source that modulates platelet activity, including catechins, proanthocyanins, anthocyanins, and flavonol glycosides [140]. Cocoa supplementation inhibited platelet aggregation when consumed acutely or chronically [141].
Olive leaf extract contains various polyphenols and flavones that inhibit platelet aggregation and ATP release [142]. The olive leaf polyphenols inhibit oxidative stress-induced platelet activation via antioxidant activity [143]. Polyphenol reduces degranulation by scavenging H2O2, a substrate involved in the COX-1 activation. Nevertheless, polyphenols can inhibit aggregation and secretion by inhibiting intracellular pathways in platelets.
Chlorogenic acid present in fruits inhibits collagen- and ADP-induced platelet aggregation [144]. Polyphenols dose-dependently inhibit platelet aggregation and ATP release mediated via the purinergic receptor- and GPVI pathways [144]. In vitro studies showed that ellagic acid modulates platelet intracellular signaling pathways involving PLC/PKC pathways.
Zhou et al. [145] reported that anthocyanin cyanidin-3-glucoside (C3G) significantly attenuated thrombin-induced platelet activation and ATP secretion in mice fed with high-fat diets. C3G also inhibited thrombin and collagen-induced platelet aggregation [146]. Other low bioavailable polyphenols also had antiplatelet activity [147].
Hydroxytyrosol (HT) acetate and aspirin have similar anti-aggregatory activity [148]. However, HT-acetate had higher anti-aggregation activity than HT. A more significant anti-aggregation impact of aspirin and HT acetate was observed in whole blood aggregation induced by ADP or collagen compared with aggregation in platelet-rich plasma, indicating an alternative aggregation pathway involving leucocytes, erythrocytes, platelets, and other plasma proteins. Aspirin and the two phenolic compounds increase NO production in leucocytes, thus contributing to the anti-aggregation effect. NO can also affect platelet aggregation by modulating cAMP, cGMP, and TxA2 synthesis. Propolis inhibited ADP-induced platelet aggregation via the P2Y12 pathway [149] by decreasing the expression of the fibrinogen receptor (GPIIb/IIIa) [150].
Gallic acid inhibits ADP-induced platelet aggregation via decreasing intracellular Ca2+ levels, reducing phosphorylation and P-selectin secretion [151] Olas et al. [152] demonstrated that grape seed extracts (gallic acid, falvan-3-ols, and proanthocyanins) inhibited thrombin- and thrombin receptor-activation peptide (TRAP)-induced platelet activation and PMPs formation more than resveratrol [153]. Furthermore, resveratrol and its analog isorhapontigenin inhibited ADP-induced platelet aggregation [154]. Polyphenols inhibit the COX-1 and P2Y12 pathways and may also attenuate PMP generation in activated platelets [155]. Several polyphenols in grape seed extracts may interact with different pathways involved in platelet function.
Trimethyl-N-oxide (TMAO) is involved in the pathology of various diseases, including cancer and CVD [156]. TMA is synthesized by the gut microbiota using dietary choline and L-carnitine as substrate; then, it is carried into the liver and converted to TMAO by flavin-containing monooxygenase 3 [156]. The conversion of choline and carnitine to TMA depends on gut microbiota diversity [156]. Therefore, gut dysbiosis increases high plasma TMAO levels and thus may ultimately result in atherosclerosis and CVD. TMAO induces platelet hyperreactivity and dyslipidemia and enhances platelet-mediated diseases such as atherosclerosis, insulin resistance, CVD, and other diseases [157,158]. TMAO enhances sub-maximal stimulus-dependent platelet activation by agonists through increased Ca2+ release [157]. Gut dysbiosis promotes atherosclerosis by increasing the production of TMAO [159,160,161]. Gut metagenome analysis showed a relatively lower abundance of Roseburia and Eubacterium, while TMA-producing Collinsella was higher in CVD patients [162]. Targeting the gut microbiota and its metabolites can effectively treat and prevent CVD by modulating TMAO levels [163,164]. Several polyphenol-rich products lower plasma TMAO levels both in animals and humans [165,166,167]. Chen et al. [165] first showed that resveratrol decreases atherosclerosis via reduced production of TMAO synthesis via remodeling gut microbiota. This observation was corroborated in rodents [166]. A study involving 20 normal-weight subjects showed a significant decrease in serum TMAO after four weeks of supplementation with 300 mg of polyphenol-rich grape pomace [167]. The polyphenol-rich extract tomato extracts lowered plasma TMAO in obese adults via modulation of gut microbiota[168].
Water-Soluble Extracts from Tomato and Kiwifruit: The Latest Dietary Antiplatelet Regimes
Hyperlipidemia, hypertension, and hyperactivity of blood platelets are critical contributors to the pathogenesis of CVD [169]. Hyperactive platelets, as observed in diabetes mellitus, insulin resistance, obesity, sedentary life, and smoking, contribute to the development of CVD[170,171,172,173,174,175]. Hyperactivity of platelets contributes to the progression of atherosclerotic plaque and other diseases [86,176]. Therefore, there is a need to decrease platelet hyperactivity through safe, effective, and practical approaches to prevent the risk of CVD and other platelet-mediated diseases. Consumption of fruits and vegetables protects against CVD [177,178,179]. The bioactive compounds in fruits and vegetables may protect the cardiovascular system via different mechanisms, such as favorably modulating oxidative stress, plasma lipid levels, hypertension, normalizing platelet hyperactivity, and other CVD risk factors[25,180]. For example, plant flavonoids inhibit cyclic nucleotide phosphodiesterase enzyme and platelet TxA2 synthesis, the main mechanisms responsible for inhibiting platelet activation.
Consequently, these bioactive components in fruits may reduce more than one CVD risk factor, such as platelet hyperactivity [23,181]. Such nutritional antiplatelet regimes were identified in the water-soluble extract in tomatoes and kiwifruits[27,78,182]. The aqueous extract of tomatoes, also known as Fruitflow® has undergone the most extensive preclinical and clinical testing to demonstrate its efficacy in inhibiting platelet hyperactivity. Recent studies showed that tamarillo, horned melon (kiwano), and raspberry extracts inhibited ADP-induced platelet aggregation [183].
Tomato
Epidemiological studies demonstrated that consumption of tomato and tomato products are associated with a reduced risk of CVD [184,185,186]. Tomatoes contain several compounds that might affect plasma lipids and platelet aggregation. The aqueous extract of tomato and kiwifruit inhibited platelet aggregation by 75-80%, whereas apple and pear had very little activity (2-5%) [25,180,182]. These tomato anti-platelet compounds had a molecular mass of less than 1000Da and were highly water soluble and stable to boiling[27]. After removing sugars, the isolated active tomato extract accounted for 4% of the aqueous tomato extract dry matter, later named Fruitflow®. Sugar-free aqueous tomato (later named Fruitflow® extract) showed potent inhibition of platelet aggregation in vitro [187]. Fruitflow® extract had three categories of compounds: nucleosides, simple phenolic derivatives, and flavonoid derivatives.
Though the mechanisms are not yet well known, however, Fruitflow® inhibits the GPIIb/IIIa activation step [187]. Fruitflow® unalter basal platelet cyclic AMP levels in vitro studies. In addition, Fruitflow® decreased the expression of P-selectin in the platelet membranes in response to ADP in whole blood [78,187]. Fruitflow® can strongly affect the size and longevity of platelet aggregates. Fruitflow® was also found to affect tissue factor (TF) binding to activated platelets, at least in part due to effects on P-selectin. These results demonstrate that the actions of Fruitflow® were consistent and mediated partly through polyphenols [180,188]. Effects on TF binding suggested that sugar-free aqueous tomato extract components could significantly impact some aspects of coagulation, such as thrombin generation. Fruitflow® inhibits platelet granule secretion by suppressing the Src-PLCγ2-PKC mediated granule secretory pathway. Platelet granule secretion occurs when collagen and thrombin bind to its receptor GPVI and PARS, and that causes activation of the downstream signaling pathway [189]. In this pathway, sarcoma, a tyrosine-protein kinase (Src) family member such as Lyn, Fyn, and Src, is first activated by phosphorylation, activated Syk, then phosphorylate and activate LAT, which in turn causes phosphorylation of an adaptor protein PLCγ2 in which protein kinase C (PKC) bind leading to the activation of downstream effector, which induces platelet granule secretion. one study reported that Fruitflow® have the ability in alteration of generation of different interleukins such as IL-1β, IL-6, IL-10 and IL-12 and chemokines such as CCL2/MCP-1, CCL3/MIP-1α, CCL5/RANTES, CXCL8/IL-8, CXCL10/IP-10 of blood leukocytes[189]. Fruitflow® interacts with immune cells and endothelial cells[190]. It distinctly modulates the production of inflammatory modulators in a context-specific way and in extenso presumably in different body compartments.
Consequently, Fruitflow® may beneficially enhance and attenuate inflammatory processes during acute and chronic inflammation. Fruitflow® modulates cytokines, chemokines, and adhesion molecules that produce cell trafficking, activation, and differentiation in different immune system compartments. However, the modulatory roles of Fruitflow® in immune and inflammation response in platelets are yet to be known. Fruitflow® is expected to have similar effects in platelets as observed with different immune cells.
The Fruitflow® prevents activation of integrin αIIbß3 (GPIIb/IIIa) [78]. Fruitflow® also inhibited platelet aggregation and P-selectin expression in collagen-stimulated human platelets[191]. Proteomics study demonstrated that compared with the Fruitflow®-treated collagen-stimulated platelets, only collagen-stimulated platelets, 60 proteins, were upregulated, and 10 proteins were downregulated. Additionally, 66 phosphorylated peptides were upregulated, whereas 37 were downregulated. Proteomic studies also demonstrated Fruitflow® also strongly affected proteins was protein disulfide isomerase (PDI), an oxidoreductase that catalyzes disulfide bond formation and isomerization. In platelets, blocking PDI with inhibitory antibodies inhibits several platelet activation pathways, including aggregation, secretion, and fibrinogen binding [192,193]. Glycosides related to quercetin in Fruitflow® demonstrated that they interact with PDI in this way [194,195]. The interaction of polyphenols with PDI suggested a possible mechanism by which Fruitflow® ingredients inhibit different platelet activation pathways.
Fruitflow® inhibited platelet function, possibly via suppression of Akt, GSK3β, p38 MAPK, and Hsp27 phosphorylation in collagen-induced platelet aggregation. The inhibition of platelet aggregation and modification of platelet proteins in collagen-stimulated platelets by Fruitflow® suggest that it can provide health benefits for people at risk of platelet hyperactivity-related thrombosis. The figure describes the mechanisms of Fruitflow®'s actions on the platelet activation pathway.
Human Trials and Animal Experiments Using Fruitflow®
Several human trials were conducted using Fruitflow® to determine the ex vivo and the onset time of an acute anti-platelet effect after consumption [24,187]. An acute lowering of platelet aggregation response to ADP and collagen was observed three hours after consuming Fruitflow®. The range of onset times was from one and a half hours to three hours after Fruitflow® consumption. On average, these studies have shown an inhibition of the platelet response to ADP of approximately 17 - 25%, and platelet aggregability returned to baseline level 18 h after consumption of a single dose (150mg) of Fruitflow® [196]. A study involving 93 men and women showed that men responded more than women, and people with higher risk factors for CVD were more responsive than others [197]. Fruitflow® ingredients affect many aspects of platelet function, including thrombin generation. However, clotting time showed no significant increases from baseline levels in all intervention studies[24].
A human trial demonstrated that the efficacious range for Fruitflow® lies between 75 mg and 300 mg [198]. A single dose of Fruitflow® was compared with 75mg aspirin, either as a single dose or taken continuously for one week, and the platelet aggregability response was measured ex vivo [199]. The effects of a single dose of Fruitflow® were similar to a 75mg dose of aspirin in terms of antiplatelet action, effects on thromboxane synthesis, and time to form a primary hemostatic clot in a study involving 47 healthy subjects [199]. Unlike aspirin, Fruitflow®’s effects were not cumulative as its effects did not irreversibly disable platelet signaling pathways[199]. The antiplatelet effects of aspirin in healthy subjects are incredibly heterogeneous, with some subjects experiencing a massive increase in time to form a primary hemostatic clot while others respond poorly. The more moderate effects of Fruitflow® were related to the reversibility of its antiplatelet action, rendering its use as primary prevention of CVD, in contrast to aspirin at any dosage.
Both Fruitflow® and aspirin affected 26 platelet proteins involved in platelet structure and function, coagulation, platelet secretory proteins, fibrinogen beta chain 5, Ras-related proteins, redox system proteins, and HSP70s. As described earlier, the platelet protein disulfide isomerase was affected by Fruitflow®, which regulates αIIbβ3 integrin activation.
Four weeks of supplementation of Fruitflow® decreased platelet aggregation and granule secretion in healthy Chinese middle-aged and older individuals [200]. After a 2-week washout period, the platelet suppressive effect of Fruitflow® had disappeared. After four weeks of Fruitflow® treatment in the obese subgroup (n = 14), significant declines in aspirin reaction units by 8.6% and in P2Y12 reaction units (PRU) by 7.5% were observed [201]. Patients with arterial hypertension and obesity may potentially benefit from consuming Fruitflow® [201].
The supplementation of Fruitflow® for four weeks in strenuous exercise rats reduced platelet hyperreactivity [202]. Fruitflow® treatment also reduced black tail length, blood flow pulse index, and vascular resistance index, ameliorating microcirculation perfusion in a rat thrombosis model. Fruitflow® inhibited platelet aggregation induced by shear flow and alleviated the blood flow and microcirculation abnormities.
EFSA authorized Fruitflow® for daily consumption in 2009[24]. The first Article 13 claim based on newly developed evidence or proprietary data (a particular category under Article 13(5)) was approved for Fruitflow® in 2009[24]. The approved claim was based on the eight human studies (seven proprietaries) and seven non-human studies (three proprietaries) conducted with Fruitflow®.
Fruitflow® differs fundamentally from aspirin or other antiplatelet drugs in that its effects are reversible. This very significant property of Fruitflow® makes it suitable for use by the general population as a dietary functional ingredient, while antiplatelet drugs cannot be used.
Kiwifruit and Its Anti-Platelet Factors
Kiwifruit contains substantial quantities of several bioactive polyphenols [203]. The familiar green kiwifruit, Actinidia deliciosa has been investigated for bioactivity, especially its effects on the risk of CVD [25,204,205,206]. As in tomato extract, the aqueous extract of kiwifruits has maximum anti-platelet factors, which was unrelated to the fruits' antioxidant potential[180]. Daily consumption of 2 or 3 kiwifruits reduced ex vivo platelet aggregation response, blood pressure, and plasma lipids [25,204,205,206]. The diverse activities of the aqueous extract of kiwifruit, such as anti-platelet, plasma triglycerides lowering, and anti-angiotensin converting enzyme (ACE) activity, were shown [25,207]. The aqueous kiwifruit extract (KFE) inhibited ADP and collagen-induced platelet aggregation.
In contrast, KFE was not that effective against ARA-induced platelet aggregation. KFE-mediated inhibition of platelet aggregation may not involve TxA2 pathway. The anti-platelet compounds in kiwifruit are water soluble and heat stable, and their molecular mass is less than 1000 Da[25]. KFE had glucose (8.9 ± 0.4 mg/ml), fructose (9.9 ± 0.5 mg/ml) and sucrose (2.3 ± 0.2 mg/ml)[182]. The inhibition of platelet aggregation by sugar-free KFE was concomitantly associated with inhibition of TxB2 synthesis and inhibited PF4 release in a dose-dependent manner. KFE at 1.68mg/ml inhibited ADP- and collagen-induced TxB2 synthesis by 91% and 81% [208].
Human Trials
Several human intervention trials were conducted to investigate the ex vivo effects of whole kiwifruits and KFE on platelet aggregation, ACE, and blood pressure [25,209,210,211]. In one human trial involving 30 healthy volunteers aged 20–51 years [25] , consuming two or three kiwifruits daily for 28 days reduced platelet aggregation ex vivo. Consuming two kiwifruit daily inhibited platelet aggregation induced by ADP significantly (18% in case of 4 μM ADP and 15% in case of 8 μM ADP) compared with those at day 0 (p < 0.05). A similar reduction in platelet aggregation was observed in response to collagen. Plasma triglyceride levels were significantly lowered on day 28. The total plasma cholesterol, LDL, and HDL levels were unchanged from days 0 to 28 in both groups [25].
Another human trial showed that consuming one kiwifruit daily for four weeks reduced whole-blood platelet aggregation in healthy volunteers[209]. The effects of consumption of 3 kiwifruit for eight weeks on blood pressure, plasma lipids, and whole-blood aggregation were investigated in a randomized, controlled trial in male smokers (aged 44–74 years) [211]. In the kiwifruit group, reductions of 10 mm Hg in systolic blood pressure and 9 mm Hg in diastolic blood pressure were observed[211]. Additionally, a 15% reduction in whole-blood aggregation and an 11% reduction in ACE activity were observed in the kiwifruit group[211]. Although only blood pressure was decreased in hypertensives, no effects on other parameters were observed in the antioxidant-rich diet group. These diverse activities of kiwifruits, such as anti-platelet and anti-ACE, were also demonstrated in differently processed aqueous extracts of kiwifruits[25,207]. Not only green kiwifruits, consuming one gold kiwifruit daily for four weeks also reduced whole-blood platelet aggregation and plasma triglycerides in healthy volunteers[209]. Consuming 2 mg of KFE in 10 g margarine inhibited ex vivo platelet aggregation by 12.7% two hours after consumption by healthy volunteers (n = 9)[182]. Another randomized, double-blinded, and placebo-controlled cross-over study in 50 subjects showed that KFE reduced systolic blood pressure by 5 mM of Hg after consuming KFE for a week (unpublished data).
Conclusions
Many studies have explored the relationship between dietary components, platelet function, and CVD risk factors. Several anti-platelet regimes have been developed, such as polyphenols, n-3 fatty acids, and fruits and vegetables. The overall data is positive for nutritional antiplatelets (such as fish and fish oil, polyphenols, Fruitflow, and KFE) to prevent CVD. However, platelet hyperactivity is also associated with many non-hemostatic diseases. Therefore, the impacts of these anti-platelet dietary factors on these diseases must be further investigated. The next question is, which of these nutritional antiplatelets is superior in its efficacy as an antiplatelet ingredient? At present, the Fruitflow® is the most studied potent anti-platelet functional food. Fruitflow®, developed from tomato-containing bioavailable cardioprotective compounds, can benefit people vulnerable to developing CVD.
Beyond efficacy, one must also consider the practical aspects of consuming such dietary antiplatelets daily. For example, dosing, product form, commercial viability, and cost should also be considered. Finally, reliable markers of platelet activity and blood flow and the techniques thereof need to be further studied.
Platelets become hyperactive in obesity, diabetes, a sedentary lifestyle, or hypertension, and in people who smoke. Platelet hyperreactivity is not merely an inevitable adverse effect of conditions but also a substantial contributing factor that exacerbates most human pathologies [23,173,212]. Recent studies indicate that platelet hyperactivity is not just a biomarker of adverse effects but a condition that requires management. Modulating platelet reactivity towards collagen, ADP, and plasma triglyceride levels by functional foods could be of potential prophylactic and therapeutic benefit in preventing and halting pathologic processes.
An array of extensive basic, mechanistic, compositional, and several human trials testify to the cardio-protective benefits of Fruitflow® [25,180,188,213]. In addition to Fruitflow®, Kiwifruits have great potential for maintaining normal platelet activity.
It is now recognized that 20–30% of persons experience the so-called aspirin-resistance syndrome, in which the expected antiplatelet effects are not observed [214]. This finding indicates an advantage of functional foods’ broad antiplatelet activity profile over single-target drugs such as aspirin. Dietary antiplatelet functional foods help provide suitably gentle and safe yet efficacious therapies to improve public health in response to various health challenges involving platelet hyperactivity.
Funding
This research received no external funding.
Acknowledgments
The author is grateful to the Faculty of Medicine, University of Oslo.
Conflicts of Interest
The author patented anti-platelet and anti-hypertensive factors in tomatoes (Fruitflow®) and Kiwifruit extract.
Abbreviations: CVD, Cardiovascular Disease; EPA, Eicosapentaenoic acid,20:5n-3; ARA, Arachidonic acid,20:4n-6; DHA, docosahexaenoic, 22:6n-3; EFSA, European Food Safety Authority; PDI, Protein disulphide isomerase; TF; Tissue factor, PRP, platelet rich plasma; TxA2, thromboxane A2
References
- Ho-Tin-Noe, B.; Boulaftali, Y.; Camerer, E. Platelets and vascular integrity: How platelets prevent bleeding in inflammation. Blood 2018, 131, 277–288. [Google Scholar] [CrossRef]
- Davi, G.; Patrono, C. Platelet activation and atherothrombosis. N. Engl. J. Med. 2007, 357, 2482–2494. [Google Scholar] [CrossRef]
- Xu, X.R.; Zhang, D.; Oswald, B.E.; Carrim, N.; Wang, X.; Hou, Y.; Zhang, Q.; Lavalle, C.; McKeown, T.; Marshall, A.H.; et al. Platelets are versatile cells: New discoveries in hemostasis, thrombosis, immune responses, tumor metastasis and beyond. Crit. Rev. Clin. Lab. Sci. 2016, 53, 409–430. [Google Scholar] [CrossRef]
- Das, D.; Adhikary, S.; Das, R.K.; Banerjee, A.; Radhakrishnan, A.K.; Paul, S.; Pathak, S.; Duttaroy, A.K. Bioactive food components and their inhibitory actions in multiple platelet pathways. J. Food Biochem. 2022, 46, e14476. [Google Scholar] [CrossRef]
- O'Kennedy, N.; Duss, R.; Duttaroy, A.K. Dietary Antiplatelets: A New Perspective on the Health Benefits of the Water-Soluble Tomato Concentrate Fruitflow((R)). Nutrients 2021, 13, 2184. [Google Scholar] [CrossRef]
- Das, D.; Shruthi, N.R.; Banerjee, A.; Jothimani, G.; Duttaroy, A.K.; Pathak, S. Endothelial dysfunction, platelet hyperactivity, hypertension, and the metabolic syndrome: Molecular insights and combating strategies. Front. Nutr. 2023, 10, 1221438. [Google Scholar] [CrossRef]
- Xu, X.R.; Yousef, G.M.; Ni, H. Cancer and platelet crosstalk: Opportunities and challenges for aspirin and other antiplatelet agents. Blood 2018, 131, 1777–1789. [Google Scholar] [CrossRef]
- Koupenova, M.; Corkrey, H.A.; Vitseva, O.; Manni, G.; Pang, C.J.; Clancy, L.; Yao, C.; Rade, J.; Levy, D.; Wang, J.P.; et al. The role of platelets in mediating a response to human influenza infection. Nat. Commun. 2019, 10, 1780. [Google Scholar] [CrossRef]
- Puurunen, M.K.; Hwang, S.J.; Larson, M.G.; Vasan, R.S.; O'Donnell, C.J.; Tofler, G.; Johnson, A.D. ADP Platelet Hyperreactivity Predicts Cardiovascular Disease in the FHS (Framingham Heart Study). J. Am. Heart Assoc. 2018, 7, 8522. [Google Scholar] [CrossRef]
- Natarajan, A.; Zaman, A.G.; Marshall, S.M. Platelet hyperactivity in type 2 diabetes: Role of antiplatelet agents. Diab Vasc. Dis. Res. 2008, 5, 138–144. [Google Scholar] [CrossRef]
- Massberg, S.; Brand, K.; Gruner, S.; Page, S.; Muller, E.; Muller, I.; Bergmeier, W.; Richter, T.; Lorenz, M.; Konrad, I.; et al. A critical role of platelet adhesion in the initiation of atherosclerotic lesion formation. J. Exp. Med. 2002, 196, 887–896. [Google Scholar] [CrossRef]
- Kaplan, Z.S.; Jackson, S.P. The role of platelets in atherothrombosis. Hematol. Am. Soc. Hematol. Educ. Program. 2011, 2011, 51–61. [Google Scholar] [CrossRef]
- Lagadinou, M.; Onisor, M.O.; Rigas, A.; Musetescu, D.V.; Gkentzi, D.; Assimakopoulos, S.F.; Panos, G.; Marangos, M. Antimicrobial Properties on Non-Antibiotic Drugs in the Era of Increased Bacterial Resistance. Antibiotics 2020, 9, 107. [Google Scholar] [CrossRef]
- McDonald, B.; Dunbar, M. Platelets and Intravascular Immunity: Guardians of the Vascular Space During Bloodstream Infections and Sepsis. Front. Immunol. 2019, 10, 2400. [Google Scholar] [CrossRef]
- Koupenova, M.; Clancy, L.; Corkrey, H.A.; Freedman, J.E. Circulating Platelets as Mediators of Immunity, Inflammation, and Thrombosis. Circ. Res. 2018, 122, 337–351. [Google Scholar] [CrossRef]
- Thomas, M.R.; Storey, R.F. The role of platelets in inflammation. Thromb. Haemost. 2015, 114, 449–458. [Google Scholar] [CrossRef]
- Varon, D.; Shai, E. Platelets and their microparticles as key players in pathophysiological responses. J. Thromb. Haemost. 2015, 13 (Suppl. S1), S40–S46. [Google Scholar] [CrossRef]
- Le Blanc, J.; Lordkipanidze, M. Platelet Function in Aging. Front. Cardiovasc. Med. 2019, 6, 109. [Google Scholar] [CrossRef]
- Eisenhardt, S.U.; Habersberger, J.; Peter, K. Monomeric C-reactive protein generation on activated platelets: The missing link between inflammation and atherothrombotic risk. Trends Cardiovasc. Med. 2009, 19, 232–237. [Google Scholar] [CrossRef]
- Muller, I.; Massberg, S.; Zierhut, W.; Binz, C.; Schuster, A.; Rudiger-von Hoch, S.; Braun, S.; Gawaz, M. Effects of aspirin and clopidogrel versus oral anticoagulation on platelet function and on coagulation in patients with nonvalvular atrial fibrillation (CLAFIB). Pathophysiol. Haemost. Thromb. 2002, 32, 16–24. [Google Scholar] [CrossRef]
- Cai, G.; Zhou, W.; Lu, Y.; Chen, P.; Lu, Z.; Fu, Y. Aspirin resistance and other aspirin-related concerns. Neurol. Sci. 2016, 37, 181–189. [Google Scholar] [CrossRef]
- Piepoli, M.F.; Hoes, A.W.; Agewall, S.; Albus, C.; Brotons, C.; Catapano, A.L.; Cooney, M.T.; Corra, U.; Cosyns, B.; Deaton, C.; et al. 2016 European Guidelines on cardiovascular disease prevention in clinical practice: The Sixth Joint Task Force of the European Society of Cardiology and Other Societies on Cardiovascular Disease Prevention in Clinical Practice (constituted by representatives of 10 societies and by invited experts)Developed with the special contribution of the European Association for Cardiovascular Prevention & Rehabilitation (EACPR). Eur. Heart J. 2016, 37, 2315–2381. [Google Scholar] [CrossRef]
- Dutta-Roy, A.K. Dietary components and human platelet activity. Platelets 2002, 13, 67–75. [Google Scholar] [CrossRef]
- O'Kennedy, N.; Raederstorff, D.; Duttaroy, A.K. Fruitflow((R)): The first European Food Safety Authority-approved natural cardio-protective functional ingredient. Eur. J. Nutr. 2017, 56, 461–482. [Google Scholar] [CrossRef]
- Duttaroy, A.K.; Jorgensen, A. Effects of kiwi fruit consumption on platelet aggregation and plasma lipids in healthy human volunteers. Platelets 2004, 15, 287–292. [Google Scholar] [CrossRef]
- Dutta-Roy, A.K.; Gordon, M.J.; Kelly, C.; Hunter, K.; Crosbie, L.; Knight-Carpentar, T.; Williams, B.C. Inhibitory effect of Ginkgo biloba extract on human platelet aggregation. Platelets 1999, 10, 298–305. [Google Scholar] [CrossRef]
- Dutta-Roy, A.K.; Crosbie, L.; Gordon, M.J. Effects of tomato extract on human platelet aggregation in vitro. Platelets 2001, 12, 218–227. [Google Scholar]
- Lowery, C.L., 3rd; Elliott, C.; Cooper, A.; Hadden, C.; Sonon, R.N.; Azadi, P.; Williams, D.K.; Marsh, J.D.; Woulfe, D.S.; Kilic, F. Cigarette Smoking-Associated Alterations in Serotonin/Adrenalin Signaling Pathways of Platelets. J. Am. Heart Assoc. 2017, 6, 5465. [Google Scholar] [CrossRef]
- Ishida, M.; Sakai, C.; Kobayashi, Y.; Ishida, T. Cigarette Smoking and Atherosclerotic Cardiovascular Disease. J. Atheroscler. Thromb. 2024, 31, 189–200. [Google Scholar] [CrossRef]
- Robertson, S.; Miller, M.R. Ambient air pollution and thrombosis. Part. Fibre Toxicol. 2018, 15, 1. [Google Scholar] [CrossRef]
- Zhang, X.; McGeoch, S.C.; Johnstone, A.M.; Holtrop, G.; Sneddon, A.A.; MacRury, S.M.; Megson, I.L.; Pearson, D.W.; Abraham, P.; De Roos, B.; et al. Platelet-derived microparticle count and surface molecule expression differ between subjects with and without type 2 diabetes, independently of obesity status. J. Thromb. Thrombolysis 2014, 37, 455–463. [Google Scholar] [CrossRef]
- Huskens, D.; Roest, M.; Remijn, J.A.; Konings, J.; Kremers, R.M.; Bloemen, S.; Schurgers, E.; Selmeczi, A.; Kelchtermans, H.; van Meel, R.; et al. Strenuous exercise induces a hyperreactive rebalanced haemostatic state that is more pronounced in men. Thromb. Haemost. 2016, 115, 1109–1119. [Google Scholar] [CrossRef]
- Chaudhary, P.K.; Kim, S.; Kim, S. An Insight into Recent Advances on Platelet Function in Health and Disease. Int. J. Mol. Sci. 2022, 23, 6022. [Google Scholar] [CrossRef]
- Podrez, E.A.; Byzova, T.V.; Febbraio, M.; Salomon, R.G.; Ma, Y.; Valiyaveettil, M.; Poliakov, E.; Sun, M.; Finton, P.J.; Curtis, B.R.; et al. Platelet CD36 links hyperlipidemia, oxidant stress and a prothrombotic phenotype. Nat. Med. 2007, 13, 1086–1095. [Google Scholar] [CrossRef]
- Chen, K.; Febbraio, M.; Li, W.; Silverstein, R.L. A specific CD36-dependent signaling pathway is required for platelet activation by oxidized low-density lipoprotein. Circ. Res. 2008, 102, 1512–1519. [Google Scholar] [CrossRef]
- Jennings, L.K. Mechanisms of platelet activation: Need for new strategies to protect against platelet-mediated atherothrombosis. Thromb. Haemost. 2009, 102, 248–257. [Google Scholar] [CrossRef]
- Yeung, J.; Li, W.; Holinstat, M. Platelet Signaling and Disease: Targeted Therapy for Thrombosis and Other Related Diseases. Pharmacol. Rev. 2018, 70, 526–548. [Google Scholar] [CrossRef]
- Leiter, O.; Walker, T.L. Platelets in Neurodegenerative Conditions-Friend or Foe? Front. Immunol. 2020, 11, 747. [Google Scholar] [CrossRef]
- Rendu, F.; Brohard-Bohn, B. The platelet release reaction: granules' constituents, secretion and functions. Platelets 2001, 12, 261–273. [Google Scholar] [CrossRef]
- Janowska-Wieczorek, A.; Majka, M.; Kijowski, J.; Baj-Krzyworzeka, M.; Reca, R.; Turner, A.R.; Ratajczak, J.; Emerson, S.G.; Kowalska, M.A.; Ratajczak, M.Z. Platelet-derived microparticles bind to hematopoietic stem/progenitor cells and enhance their engraftment. Blood 2001, 98, 3143–3149. [Google Scholar] [CrossRef]
- Heijnen, H.F.; Schiel, A.E.; Fijnheer, R.; Geuze, H.J.; Sixma, J.J. Activated platelets release two types of membrane vesicles: Microvesicles by surface shedding and exosomes derived from exocytosis of multivesicular bodies and alpha-granules. Blood 1999, 94, 3791–3799. [Google Scholar] [CrossRef]
- Frossard, M.; Fuchs, I.; Leitner, J.M.; Hsieh, K.; Vlcek, M.; Losert, H.; Domanovits, H.; Schreiber, W.; Laggner, A.N.; Jilma, B. Platelet function predicts myocardial damage in patients with acute myocardial infarction. Circulation 2004, 110, 1392–1397. [Google Scholar] [CrossRef]
- Tantry, U.S.; Bonello, L.; Aradi, D.; Price, M.J.; Jeong, Y.H.; Angiolillo, D.J.; Stone, G.W.; Curzen, N.; Geisler, T.; Ten Berg, J.; et al. Consensus and update on the definition of on-treatment platelet reactivity to adenosine diphosphate associated with ischemia and bleeding. J. Am. Coll. Cardiol. 2013, 62, 2261–2273. [Google Scholar] [CrossRef]
- Arnoldussen, I.A.C.; Witkamp, R.F. Effects of Nutrients on Platelet Function: A Modifiable Link between Metabolic Syndrome and Neurodegeneration? Biomolecules 2021, 11, 1455. [Google Scholar] [CrossRef]
- Barale, C.; Russo, I. Influence of Cardiometabolic Risk Factors on Platelet Function. Int. J. Mol. Sci. 2020, 21, 623. [Google Scholar] [CrossRef]
- Russo, I. The prothrombotic tendency in metabolic syndrome: Focus on the potential mechanisms involved in impaired haemostasis and fibrinolytic balance. Scientifica 2012, 2012, 525374. [Google Scholar] [CrossRef]
- Yang, M.; Kholmukhamedov, A. Platelet reactivity in dyslipidemia: Atherothrombotic signaling and therapeutic implications. Rev. Cardiovasc. Med. 2021, 22, 67–81. [Google Scholar] [CrossRef]
- Dutta-Roy, A.K.; Gordon, M.J.; Campbell, F.M.; Crosbie, L.C. Arachidonic acid uptake by human platelets is mediated by CD36. Platelets 1996, 7, 291–295. [Google Scholar] [CrossRef]
- Li, N. Platelets as an inter-player between hyperlipidaemia and atherosclerosis. J. Intern. Med. 2024, 296, 39–52. [Google Scholar] [CrossRef]
- Elbatarny, H.S.; Maurice, D.H. Leptin-mediated activation of human platelets: Involvement of a leptin receptor and phosphodiesterase 3A-containing cellular signaling complex. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E695–E702. [Google Scholar] [CrossRef]
- Konstantinides, S.; Schafer, K.; Koschnick, S.; Loskutoff, D.J. Leptin-dependent platelet aggregation and arterial thrombosis suggests a mechanism for atherothrombotic disease in obesity. J. Clin. Investig. 2001, 108, 1533–1540. [Google Scholar] [CrossRef]
- Bladowski, M.; Gawrys, J.; Gajecki, D.; Szahidewicz-Krupska, E.; Sawicz-Bladowska, A.; Doroszko, A. Role of the Platelets and Nitric Oxide Biotransformation in Ischemic Stroke: A Translative Review from Bench to Bedside. Oxid. Med. Cell Longev. 2020, 2020, 2979260. [Google Scholar] [CrossRef]
- Wang, Q.; Zennadi, R. Oxidative Stress and Thrombosis during Aging: The Roles of Oxidative Stress in RBCs in Venous Thrombosis. Int. J. Mol. Sci. 2020, 21, 4259. [Google Scholar] [CrossRef]
- Freedman, J.E. Oxidative stress and platelets. Arterioscler. Thromb. Vasc. Biol. 2008, 28, s11–s16. [Google Scholar] [CrossRef]
- Ceriello, A. Postprandial hyperglycemia and diabetes complications: Is it time to treat? Diabetes 2005, 54, 1–7. [Google Scholar] [CrossRef]
- Andre, P.; Nannizzi-Alaimo, L.; Prasad, S.K.; Phillips, D.R. Platelet-derived CD40L: The switch-hitting player of cardiovascular disease. Circulation 2002, 106, 896–899. [Google Scholar] [CrossRef]
- Henn, V.; Steinbach, S.; Buchner, K.; Presek, P.; Kroczek, R.A. The inflammatory action of CD40 ligand (CD154) expressed on activated human platelets is temporally limited by coexpressed CD40. Blood 2001, 98, 1047–1054. [Google Scholar] [CrossRef]
- Miceli, G.; Basso, M.G.; Rizzo, G.; Pintus, C.; Tuttolomondo, A. The Role of the Coagulation System in Peripheral Arterial Disease: Interactions with the Arterial Wall and Its Vascular Microenvironment and Implications for Rational Therapies. Int. J. Mol. Sci. 2022, 23, 14914. [Google Scholar] [CrossRef]
- Azimova, K.; San Juan, Z.; Mukherjee, D. Cardiovascular safety profile of currently available diabetic drugs. Ochsner J. 2014, 14, 616–632. [Google Scholar]
- Johnson, M.; Harrison, H.E.; Raftery, A.T.; Elder, J.B. Vascular prostacyclin may be reduced in diabetes in man. Lancet 1979, 1, 325–326. [Google Scholar] [CrossRef]
- Rabini, R.A.; Staffolani, R.; Fumelli, P.; Mutus, B.; Curatola, G.; Mazzanti, L. Decreased nitric oxide synthase activity in platelets from IDDM and NIDDM patients. Diabetologia 1998, 41, 101–104. [Google Scholar] [CrossRef]
- Blann, A.D.; Nadar, S.; Lip, G.Y. Pharmacological modulation of platelet function in hypertension. Hypertension 2003, 42, 1–7. [Google Scholar] [CrossRef]
- Pravenec, M.; Kunes, J.; Zicha, J.; Kren, V.; Klir, P. Platelet aggregation in spontaneous hypertension: Genetic determination and correlation analysis. J. Hypertens. 1992, 10, 1453–1456. [Google Scholar] [CrossRef]
- Breddin, H.K.; Lippold, R.; Bittner, M.; Kirchmaier, C.M.; Krzywanek, H.J.; Michaelis, J. Spontaneous platelet aggregation as a predictive risk factor for vascular occlusions in healthy volunteers? Results of the HAPARG Study. Haemostatic parameters as risk factors in healthy volunteers. Atherosclerosis 1999, 144, 211–219. [Google Scholar] [CrossRef]
- Torsellini, A.; Becucci, A.; Citi, S.; Cozzolino, F.; Guidi, G.; Lombardi, V.; Vercelli, D.; Veloci, M. Effects of pressure excursions on human platelets. In vitro studies on betathromboglobulin (beta-TG) and platelet factor 4 (PF4) release and on platelet sensitivity to ADP-aggregation. Haematologica 1982, 67, 860–866. [Google Scholar]
- Lip, G.Y.; Blann, A.D.; Farooqi, I.S.; Zarifis, J.; Sagar, G.; Beevers, D.G. Sequential alterations in haemorheology, endothelial dysfunction, platelet activation and thrombogenesis in relation to prognosis following acute stroke: The West Birmingham Stroke Project. Blood Coagul. Fibrinolysis 2002, 13, 339–347. [Google Scholar] [CrossRef]
- Blann, A.D.; Naqvi, T.; Waite, M.; McCollum, C.N. von Willebrand factor and endothelial damage in essential hypertension. J. Hum. Hypertens. 1993, 7, 107–111. [Google Scholar]
- Nityanand, S.; Pande, I.; Bajpai, V.K.; Singh, L.; Chandra, M.; Singh, B.N. Platelets in essential hypertension. Thromb. Res. 1993, 72, 447–454. [Google Scholar] [CrossRef]
- Kjeldsen, S.E.; Gjesdal, K.; Eide, I.; Aakesson, I.; Amundsen, R.; Foss, O.P.; Leren, P. Increased beta-thromboglobulin in essential hypertension: Interactions between arterial plasma adrenaline, platelet function and blood lipids. Acta Med. Scand. 1983, 213, 369–373. [Google Scholar] [CrossRef]
- Spencer, C.G.; Gurney, D.; Blann, A.D.; Beevers, D.G.; Lip, G.Y.; Ascot Steering Committee, A.-S.C.O.T. Von Willebrand factor, soluble P-selectin, and target organ damage in hypertension: A substudy of the Anglo-Scandinavian Cardiac Outcomes Trial (ASCOT). Hypertension 2002, 40, 61–66. [Google Scholar] [CrossRef]
- Lechi, C.; Sinigaglia, D.; Delva, P.; Guzzo, P.; Arosio, E.; Steele, A.; Lechi, A. Platelet intracellular free Ca2+ after incubation with plasma from hypertensive patients. J. Hum. Hypertens. 1989, 3, 45–48. [Google Scholar]
- Yamanishi, J.; Sano, H.; Saito, K.; Furuta, Y.; Fukuzaki, H. Plasma concentrations of platelet-specific proteins in different stages of essential hypertension: Interactions between platelet aggregation, blood lipids and age. Thromb. Haemost. 1985, 54, 539–543. [Google Scholar] [CrossRef]
- Forte, P.; Copland, M.; Smith, L.M.; Milne, E.; Sutherland, J.; Benjamin, N. Basal nitric oxide synthesis in essential hypertension. Lancet 1997, 349, 837–842. [Google Scholar] [CrossRef]
- Camilletti, A.; Moretti, N.; Giacchetti, G.; Faloia, E.; Martarelli, D.; Mantero, F.; Mazzanti, L. Decreased nitric oxide levels and increased calcium content in platelets of hypertensive patients. Am. J. Hypertens. 2001, 14, 382–386. [Google Scholar] [CrossRef]
- Dockrell, M.E.; Walker, B.R.; Noon, J.P.; Watt, G.C.; Williams, B.C.; Webb, D.J. Platelet aggregation in young men with contrasting predisposition to high blood pressure. Am. J. Hypertens. 1999, 12, 115–119. [Google Scholar] [CrossRef]
- Li, B.; Lu, Z.; Yang, Z.; Zhang, X.; Wang, M.; Chu, T.; Wang, P.; Qi, F.; Anderson, G.J.; Jiang, E.; et al. Monitoring circulating platelet activity to predict cancer-associated thrombosis. Cell Rep. Methods 2023, 3, 100513. [Google Scholar] [CrossRef]
- Gussoni, G.; Frasson, S.; La Regina, M.; Di Micco, P.; Monreal, M.; Investigators, R. Three-month mortality rate and clinical predictors in patients with venous thromboembolism and cancer. Findings from the RIETE registry. Thromb. Res. 2013, 131, 24–30. [Google Scholar] [CrossRef]
- O'Kennedy, N.; Crosbie, L.; van Lieshout, M.; Broom, J.I.; Webb, D.J.; Duttaroy, A.K. Effects of antiplatelet components of tomato extract on platelet function in vitro and ex vivo: A time-course cannulation study in healthy humans. Am. J. Clin. Nutr. 2006, 84, 570–579. [Google Scholar] [CrossRef]
- Weisman, S.M.; Graham, D.Y. Evaluation of the benefits and risks of low-dose aspirin in the secondary prevention of cardiovascular and cerebrovascular events. Arch. Intern. Med. 2002, 162, 2197–2202. [Google Scholar] [CrossRef]
- Schror, K. Aspirin and platelets: The antiplatelet action of aspirin and its role in thrombosis treatment and prophylaxis. Semin. Thromb. Hemost. 1997, 23, 349–356. [Google Scholar] [CrossRef]
- Zaragoza, C.; Monserrat, J.; Mantecon, C.; Villaescusa, L.; Zaragoza, F.; Alvarez-Mon, M. Antiplatelet activity of flavonoid and coumarin drugs. Vasc. Pharmacol. 2016, 87, 139–149. [Google Scholar] [CrossRef]
- Wang, C.; Harris, W.S.; Chung, M.; Lichtenstein, A.H.; Balk, E.M.; Kupelnick, B.; Jordan, H.S.; Lau, J. n-3 Fatty acids from fish or fish-oil supplements, but not alpha-linolenic acid, benefit cardiovascular disease outcomes in primary- and secondary-prevention studies: A systematic review. Am. J. Clin. Nutr. 2006, 84, 5–17. [Google Scholar] [CrossRef]
- Andrioli, G.; Carletto, A.; Guarini, P.; Galvani, S.; Biasi, D.; Bellavite, P.; Corrocher, R. Differential effects of dietary supplementation with fish oil or soy lecithin on human platelet adhesion. Thromb. Haemost. 1999, 82, 1522–1527. [Google Scholar]
- Golanski, J.; Szymanska, P.; Rozalski, M. Effects of Omega-3 Polyunsaturated Fatty Acids and Their Metabolites on Haemostasis-Current Perspectives in Cardiovascular Disease. Int. J. Mol. Sci. 2021, 22, 2394. [Google Scholar] [CrossRef]
- Parent, C.A.; Lagarde, M.; Venton, D.L.; Le Breton, G.C. Selective modulation of the human platelet thromboxane A2/prostaglandin H2 receptor by eicosapentaenoic and docosahexaenoic acids in intact platelets and solubilized platelet membranes. J. Biol. Chem. 1992, 267, 6541–6547. [Google Scholar] [CrossRef]
- Dutta-Roy, A.K. Insulin mediated processes in platelets, erythrocytes and monocytes/macrophages: Effects of essential fatty acid metabolism. Prostaglandins Leukot. Essent. Fat. Acids 1994, 51, 385–399. [Google Scholar] [CrossRef]
- von Schacky, C. Prophylaxis of atherosclerosis with marine omega-3 fatty acids. A comprehensive strategy. Ann. Intern. Med. 1987, 107, 890–899. [Google Scholar] [CrossRef]
- Gao, L.G.; Cao, J.; Mao, Q.X.; Lu, X.C.; Zhou, X.L.; Fan, L. Influence of omega-3 polyunsaturated fatty acid-supplementation on platelet aggregation in humans: A meta-analysis of randomized controlled trials. Atherosclerosis 2013, 226, 328–334. [Google Scholar] [CrossRef]
- Hashimoto, M.; Hossain, S.; Shido, O. Docosahexaenoic acid but not eicosapentaenoic acid withstands dietary cholesterol-induced decreases in platelet membrane fluidity. Mol. Cell Biochem. 2006, 293, 1–8. [Google Scholar] [CrossRef]
- Larson, M.K.; Tormoen, G.W.; Weaver, L.J.; Luepke, K.J.; Patel, I.A.; Hjelmen, C.E.; Ensz, N.M.; McComas, L.S.; McCarty, O.J. Exogenous modification of platelet membranes with the omega-3 fatty acids EPA and DHA reduces platelet procoagulant activity and thrombus formation. Am. J. Physiol. Cell Physiol. 2013, 304, C273–C279. [Google Scholar] [CrossRef]
- McEwen, B.J.; Morel-Kopp, M.C.; Tofler, G.H.; Ward, C.M. The effect of omega-3 polyunsaturated fatty acids on fibrin and thrombin generation in healthy subjects and subjects with cardiovascular disease. Semin. Thromb. Hemost. 2015, 41, 315–322. [Google Scholar] [CrossRef]
- Dona, M.; Fredman, G.; Schwab, J.M.; Chiang, N.; Arita, M.; Goodarzi, A.; Cheng, G.; von Andrian, U.H.; Serhan, C.N. Resolvin E1, an EPA-derived mediator in whole blood, selectively counterregulates leukocytes and platelets. Blood 2008, 112, 848–855. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Stanger, L.; Freedman, C.J.; Standley, M.; Hoang, T.; Adili, R.; Tsai, W.C.; van Hoorebeke, C.; Holman, T.R.; Holinstat, M. DHA 12-LOX-derived oxylipins regulate platelet activation and thrombus formation through a PKA-dependent signaling pathway. J. Thromb. Haemost. 2021, 19, 839–851. [Google Scholar] [CrossRef]
- Fischer, S.; Weber, P.C. Thromboxane A3 (TXA3) is formed in human platelets after dietary eicosapentaenoic acid (C20:5 omega 3). Biochem. Biophys. Res. Commun. 1983, 116, 1091–1099. [Google Scholar] [CrossRef]
- Gibney, M.J.; Bolton-Smith, C. The effect of a dietary supplement of n-3 polyunsaturated fat on platelet lipid composition, platelet function and platelet plasma membrane fluidity in healthy volunteers. Br. J. Nutr. 1988, 60, 5–12. [Google Scholar] [CrossRef]
- Li, X.L.; Steiner, M. Dose response of dietary fish oil supplementations on platelet adhesion. Arterioscler. Thromb. 1991, 11, 39–46. [Google Scholar] [CrossRef]
- Pirich, C.; Gaszo, A.; Granegger, S.; Sinzinger, H. Effects of fish oil supplementation on platelet survival and ex vivo platelet function in hypercholesterolemic patients. Thromb. Res. 1999, 96, 219–227. [Google Scholar] [CrossRef]
- Svaneborg, N.; Kristensen, S.D.; Hansen, L.M.; Bullow, I.; Husted, S.E.; Schmidt, E.B. The acute and short-time effect of supplementation with the combination of n-3 fatty acids and acetylsalicylic acid on platelet function and plasma lipids. Thromb. Res. 2002, 105, 311–316. [Google Scholar] [CrossRef]
- Eschen, O.; Christensen, J.H.; De Caterina, R.; Schmidt, E.B. Soluble adhesion molecules in healthy subjects: A dose-response study using n-3 fatty acids. Nutr. Metab. Cardiovasc. Dis. 2004, 14, 180–185. [Google Scholar] [CrossRef]
- Vanschoonbeek, K.; Feijge, M.A.; Paquay, M.; Rosing, J.; Saris, W.; Kluft, C.; Giesen, P.L.; de Maat, M.P.; Heemskerk, J.W. Variable hypocoagulant effect of fish oil intake in humans: Modulation of fibrinogen level and thrombin generation. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1734–1740. [Google Scholar] [CrossRef]
- Din, J.N.; Harding, S.A.; Valerio, C.J.; Sarma, J.; Lyall, K.; Riemersma, R.A.; Newby, D.E.; Flapan, A.D. Dietary intervention with oil rich fish reduces platelet-monocyte aggregation in man. Atherosclerosis 2008, 197, 290–296. [Google Scholar] [CrossRef]
- Larson, M.K.; Shearer, G.C.; Ashmore, J.H.; Anderson-Daniels, J.M.; Graslie, E.L.; Tholen, J.T.; Vogelaar, J.L.; Korth, A.J.; Nareddy, V.; Sprehe, M.; et al. Omega-3 fatty acids modulate collagen signaling in human platelets. Prostaglandins Leukot. Essent. Fat. Acids 2011, 84, 93–98. [Google Scholar] [CrossRef]
- Vericel, E.; Calzada, C.; Chapuy, P.; Lagarde, M. The influence of low intake of n-3 fatty acids on platelets in elderly people. Atherosclerosis 1999, 147, 187–192. [Google Scholar] [CrossRef]
- Croset, M.; Vericel, E.; Rigaud, M.; Hanss, M.; Courpron, P.; Dechavanne, M.; Lagarde, M. Functions and tocopherol content of blood platelets from elderly people after low intake of purified eicosapentaenoic acid. Thromb. Res. 1990, 57, 1–12. [Google Scholar] [CrossRef]
- Driss, F.; Vericel, E.; Lagarde, M.; Dechavanne, M.; Darcet, P. Inhibition of platelet aggregation and thromboxane synthesis after intake of small amount of icosapentaenoic acid. Thromb. Res. 1984, 36, 389–396. [Google Scholar] [CrossRef]
- Guillot, N.; Caillet, E.; Laville, M.; Calzada, C.; Lagarde, M.; Vericel, E. Increasing intakes of the long-chain omega-3 docosahexaenoic acid: Effects on platelet functions and redox status in healthy men. FASEB J. 2009, 23, 2909–2916. [Google Scholar] [CrossRef]
- Nomura, S.; Kanazawa, S.; Fukuhara, S. Effects of eicosapentaenoic acid on platelet activation markers and cell adhesion molecules in hyperlipidemic patients with Type 2 diabetes mellitus. J. Diabetes Complicat. 2003, 17, 153–159. [Google Scholar] [CrossRef]
- Chen, G.; Qian, Z.M.; Zhang, J.; Zhang, S.; Zhang, Z.; Vaughn, M.G.; Aaron, H.E.; Wang, C.; Lip, G.Y.; Lin, H. Regular use of fish oil supplements and course of cardiovascular diseases: Prospective cohort study. BMJ Med. 2024, 3, e000451. [Google Scholar] [CrossRef]
- Van Dael, P. Role of n-3 long-chain polyunsaturated fatty acids in human nutrition and health: Review of recent studies and recommendations. Nutr. Res. Pract. 2021, 15, 137–159. [Google Scholar] [CrossRef]
- Wen, Y.T.; Dai, J.H.; Gao, Q. Effects of Omega-3 fatty acid on major cardiovascular events and mortality in patients with coronary heart disease: A meta-analysis of randomized controlled trials. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 470–475. [Google Scholar] [CrossRef]
- Kwak, S.M.; Myung, S.K.; Lee, Y.J.; Seo, H.G.; Korean Meta-analysis Study, G. Efficacy of omega-3 fatty acid supplements (eicosapentaenoic acid and docosahexaenoic acid) in the secondary prevention of cardiovascular disease: A meta-analysis of randomized, double-blind, placebo-controlled trials. Arch. Intern. Med. 2012, 172, 686–694. [Google Scholar] [CrossRef]
- Nestel, P.; Clifton, P.; Colquhoun, D.; Noakes, M.; Mori, T.A.; Sullivan, D.; Thomas, B. Indications for Omega-3 Long Chain Polyunsaturated Fatty Acid in the Prevention and Treatment of Cardiovascular Disease. Heart Lung Circ. 2015, 24, 769–779. [Google Scholar] [CrossRef]
- Meyer, B.J.; Groot, R.H.M. Effects of Omega-3 Long Chain Polyunsaturated Fatty Acid Supplementation on Cardiovascular Mortality: The Importance of the Dose of DHA. Nutrients 2017, 9, 1305. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Wu, J.H. Omega-3 fatty acids and cardiovascular disease: Effects on risk factors, molecular pathways, and clinical events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J.; American Heart Association. Nutrition, C. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef]
- Siscovick, D.S.; Barringer, T.A.; Fretts, A.M.; Wu, J.H.; Lichtenstein, A.H.; Costello, R.B.; Kris-Etherton, P.M.; Jacobson, T.A.; Engler, M.B.; Alger, H.M.; et al. Omega-3 Polyunsaturated Fatty Acid (Fish Oil) Supplementation and the Prevention of Clinical Cardiovascular Disease: A Science Advisory From the American Heart Association. Circulation 2017, 135, e867–e884. [Google Scholar] [CrossRef]
- Skulas-Ray, A.C.; Wilson, P.W.F.; Harris, W.S.; Brinton, E.A.; Kris-Etherton, P.M.; Richter, C.K.; Jacobson, T.A.; Engler, M.B.; Miller, M.; Robinson, J.G.; et al. Omega-3 Fatty Acids for the Management of Hypertriglyceridemia: A Science Advisory From the American Heart Association. Circulation 2019, 140, e673–e691. [Google Scholar] [CrossRef]
- Aung, T.; Halsey, J.; Kromhout, D.; Gerstein, H.C.; Marchioli, R.; Tavazzi, L.; Geleijnse, J.M.; Rauch, B.; Ness, A.; Galan, P.; et al. Associations of Omega-3 Fatty Acid Supplement Use With Cardiovascular Disease Risks: Meta-analysis of 10 Trials Involving 77 917 Individuals. JAMA Cardiol. 2018, 3, 225–234. [Google Scholar] [CrossRef]
- Abdelhamid, A.S.; Brown, T.J.; Brainard, J.S.; Biswas, P.; Thorpe, G.C.; Moore, H.J.; Deane, K.H.; AlAbdulghafoor, F.K.; Summerbell, C.D.; Worthington, H.V.; et al. Omega-3 fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2018, 7, CD003177. [Google Scholar] [CrossRef]
- Rimm, E.B.; Appel, L.J.; Chiuve, S.E.; Djousse, L.; Engler, M.B.; Kris-Etherton, P.M.; Mozaffarian, D.; Siscovick, D.S.; Lichtenstein, A.H.; American Heart Association Nutrition Committee of the Council on, L. ; et al. Seafood Long-Chain n-3 Polyunsaturated Fatty Acids and Cardiovascular Disease: A Science Advisory From the American Heart Association. Circulation 2018, 138, e35–e47. [Google Scholar] [CrossRef]
- Ed Nignpense, B.; Chinkwo, K.A.; Blanchard, C.L.; Santhakumar, A.B. Polyphenols: Modulators of Platelet Function and Platelet Microparticle Generation? Int. J. Mol. Sci. 2019, 21, 146. [Google Scholar] [CrossRef]
- Goszcz, K.; Duthie, G.G.; Stewart, D.; Leslie, S.J.; Megson, I.L. Bioactive polyphenols and cardiovascular disease: Chemical antagonists, pharmacological agents or xenobiotics that drive an adaptive response? Br. J. Pharmacol. 2017, 174, 1209–1225. [Google Scholar] [CrossRef]
- Arts, I.C.; Hollman, P.C. Polyphenols and disease risk in epidemiologic studies. Am. J. Clin. Nutr. 2005, 81, 317S–325S. [Google Scholar] [CrossRef]
- Khan, H.; Jawad, M.; Kamal, M.A.; Baldi, A.; Xiao, J.; Nabavi, S.M.; Daglia, M. Evidence and prospective of plant derived flavonoids as antiplatelet agents: Strong candidates to be drugs of future. Food Chem. Toxicol. 2018, 119, 355–367. [Google Scholar] [CrossRef]
- Ludovici, V.; Barthelmes, J.; Nagele, M.P.; Flammer, A.J.; Sudano, I. Polyphenols: Anti-Platelet Nutraceutical? Curr. Pharm. Des. 2018, 24, 146–157. [Google Scholar] [CrossRef]
- Heptinstall, S.; May, J.; Fox, S.; Kwik-Uribe, C.; Zhao, L. Cocoa flavanols and platelet and leukocyte function: Recent in vitro and ex vivo studies in healthy adults. J. Cardiovasc. Pharmacol. 2006, 47 (Suppl. S2), S197–S205, discussion S206–S199. [Google Scholar] [CrossRef]
- Holt, R.R.; Actis-Goretta, L.; Momma, T.Y.; Keen, C.L. Dietary flavanols and platelet reactivity. J. Cardiovasc. Pharmacol. 2006, 47 (Suppl. S2), S187–S196, discussion S206-S189. [Google Scholar] [CrossRef]
- Hamed, M.S.; Gambert, S.; Bliden, K.P.; Bailon, O.; Singla, A.; Antonino, M.J.; Hamed, F.; Tantry, U.S.; Gurbel, P.A. Dark chocolate effect on platelet activity, C-reactive protein and lipid profile: A pilot study. South. Med. J. 2008, 101, 1203–1208. [Google Scholar] [CrossRef]
- Pearson, D.A.; Holt, R.R.; Rein, D.; Paglieroni, T.; Schmitz, H.H.; Keen, C.L. Flavanols and platelet reactivity. Clin. Dev. Immunol. 2005, 12, 1–9. [Google Scholar] [CrossRef]
- Ding, E.L.; Hutfless, S.M.; Ding, X.; Girotra, S. Chocolate and prevention of cardiovascular disease: A systematic review. Nutr. Metab. (Lond) 2006, 3, 2. [Google Scholar] [CrossRef]
- Hermann, F.; Spieker, L.E.; Ruschitzka, F.; Sudano, I.; Hermann, M.; Binggeli, C.; Luscher, T.F.; Riesen, W.; Noll, G.; Corti, R. Dark chocolate improves endothelial and platelet function. Heart 2006, 92, 119–120. [Google Scholar] [CrossRef]
- Wright, B.; Moraes, L.A.; Kemp, C.F.; Mullen, W.; Crozier, A.; Lovegrove, J.A.; Gibbins, J.M. A structural basis for the inhibition of collagen-stimulated platelet function by quercetin and structurally related flavonoids. Br. J. Pharmacol. 2010, 159, 1312–1325. [Google Scholar] [CrossRef]
- Bojic, M.; Debeljak, Z.; Tomicic, M.; Medic-Saric, M.; Tomic, S. Evaluation of antiaggregatory activity of flavonoid aglycone series. Nutr. J. 2011, 10, 73. [Google Scholar] [CrossRef]
- Rein, D.; Paglieroni, T.G.; Pearson, D.A.; Wun, T.; Schmitz, H.H.; Gosselin, R.; Keen, C.L. Cocoa and wine polyphenols modulate platelet activation and function. J. Nutr. 2000, 130, 2120S–2126S. [Google Scholar] [CrossRef]
- Innes, A.J.; Kennedy, G.; McLaren, M.; Bancroft, A.J.; Belch, J.J. Dark chocolate inhibits platelet aggregation in healthy volunteers. Platelets 2003, 14, 325–327. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Remesy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef]
- Das, D.; Adhikary, S.; Das, R.K.; Banerjee, A.; Radhakrishnan, A.K.; Paul, S.; Pathak, S.; Duttaroy, A.K. Bioactive food components and their inhibitory actions in multiple platelet pathways. J. Food Biochem. 2022, 46, e14476. [Google Scholar] [CrossRef]
- Dutta-Roy, A.K.; Crosbie, L.C.; Gordon, M.J.; Campbell, F.M. Platelet membrane glycoprotein IV (CD36) is involved in arachidonic acid induced-platelet aggregation. Biochem. Soc. Trans. 1996, 24, 167S. [Google Scholar] [CrossRef]
- Pierre, S.; Crosbie, L.; Duttaroy, A.K. Inhibitory effect of aqueous extracts of some herbs on human platelet aggregation in vitro. Platelets 2005, 16, 469–473. [Google Scholar] [CrossRef]
- Rimbach, G.; Melchin, M.; Moehring, J.; Wagner, A.E. Polyphenols from cocoa and vascular health-a critical review. Int. J. Mol. Sci. 2009, 10, 4290–4309. [Google Scholar] [CrossRef]
- Ostertag, L.M.; O'Kennedy, N.; Kroon, P.A.; Duthie, G.G.; de Roos, B. Impact of dietary polyphenols on human platelet function--a critical review of controlled dietary intervention studies. Mol. Nutr. Food Res. 2010, 54, 60–81. [Google Scholar] [CrossRef]
- Singh, I.; Mok, M.; Christensen, A.M.; Turner, A.H.; Hawley, J.A. The effects of polyphenols in olive leaves on platelet function. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 127–132. [Google Scholar] [CrossRef]
- Singh, I.; Quinn, H.; Mok, M.; Southgate, R.J.; Turner, A.H.; Li, D.; Sinclair, A.J.; Hawley, J.A. The effect of exercise and training status on platelet activation: Do cocoa polyphenols play a role? Platelets 2006, 17, 361–367. [Google Scholar] [CrossRef]
- Fuentes, E.; Caballero, J.; Alarcon, M.; Rojas, A.; Palomo, I. Chlorogenic acid inhibits human platelet activation and thrombus formation. PLoS ONE 2014, 9, e90699. [Google Scholar] [CrossRef]
- Zhou, F.H.; Deng, X.J.; Chen, Y.Q.; Ya, F.L.; Zhang, X.D.; Song, F.; Li, D.; Yang, Y. Anthocyanin Cyanidin-3-Glucoside Attenuates Platelet Granule Release in Mice Fed High-Fat Diets. J. Nutr. Sci. Vitaminol. (Tokyo) 2017, 63, 237–243. [Google Scholar] [CrossRef]
- Yao, Y.; Chen, Y.; Adili, R.; McKeown, T.; Chen, P.; Zhu, G.; Li, D.; Ling, W.; Ni, H.; Yang, Y. Plant-based Food Cyanidin-3-Glucoside Modulates Human Platelet Glycoprotein VI Signaling and Inhibits Platelet Activation and Thrombus Formation. J. Nutr. 2017, 147, 1917–1925. [Google Scholar] [CrossRef]
- de la Torre, R. Bioavailability of olive oil phenolic compounds in humans. Inflammopharmacology 2008, 16, 245–247. [Google Scholar] [CrossRef]
- Correa, J.A.; Lopez-Villodres, J.A.; Asensi, R.; Espartero, J.L.; Rodriguez-Gutierez, G.; De La Cruz, J.P. Virgin olive oil polyphenol hydroxytyrosol acetate inhibits in vitro platelet aggregation in human whole blood: Comparison with hydroxytyrosol and acetylsalicylic acid. Br. J. Nutr. 2009, 101, 1157–1164. [Google Scholar] [CrossRef]
- Bojic, M.; Antolic, A.; Tomicic, M.; Debeljak, Z.; Males, Z. Propolis ethanolic extracts reduce adenosine diphosphate induced platelet aggregation determined on whole blood. Nutr. J. 2018, 17, 52. [Google Scholar] [CrossRef]
- Faggio, C.; Sureda, A.; Morabito, S.; Sanches-Silva, A.; Mocan, A.; Nabavi, S.F.; Nabavi, S.M. Flavonoids and platelet aggregation: A brief review. Eur. J. Pharmacol. 2017, 807, 91–101. [Google Scholar] [CrossRef]
- Chang, S.S.; Lee, V.S.; Tseng, Y.L.; Chang, K.C.; Chen, K.B.; Chen, Y.L.; Li, C.Y. Gallic Acid Attenuates Platelet Activation and Platelet-Leukocyte Aggregation: Involving Pathways of Akt and GSK3beta. Evid. Based Complement. Altern. Med. 2012, 2012, 683872. [Google Scholar] [CrossRef]
- Olas, B.; Wachowicz, B.; Stochmal, A.; Oleszek, W. The polyphenol-rich extract from grape seeds inhibits platelet signaling pathways triggered by both proteolytic and non-proteolytic agonists. Platelets 2012, 23, 282–289. [Google Scholar] [CrossRef]
- Shi, J.; Yu, J.; Pohorly, J.E.; Kakuda, Y. Polyphenolics in grape seeds-biochemistry and functionality. J. Med. Food 2003, 6, 291–299. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, Y.; Zou, J.; Cao, K.; Xu, Y.; Wu, J.M. Effects of red wine and wine polyphenol resveratrol on platelet aggregation in vivo and in vitro. Int. J. Mol. Med. 2002, 9, 77–79. [Google Scholar] [CrossRef]
- Bijak, M.; Saluk-Bijak, J. Flavonolignans inhibit the arachidonic acid pathway in blood platelets. BMC Complement. Altern. Med. 2017, 17, 396. [Google Scholar] [CrossRef]
- Duttaroy, A.K. Role of Gut Microbiota and Their Metabolites on Atherosclerosis, Hypertension and Human Blood Platelet Function: A Review. Nutrients 2021, 13, 144. [Google Scholar] [CrossRef]
- Zhu, W.; Gregory, J.C.; Org, E.; Buffa, J.A.; Gupta, N.; Wang, Z.; Li, L.; Fu, X.; Wu, Y.; Mehrabian, M.; et al. Gut Microbial Metabolite TMAO Enhances Platelet Hyperreactivity and Thrombosis Risk. Cell 2016, 165, 111–124. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Drosos, I.; Tavridou, A.; Kolios, G. New aspects on the metabolic role of intestinal microbiota in the development of atherosclerosis. Metabolism 2015, 64, 476–481. [Google Scholar] [CrossRef]
- Gregory, J.C.; Buffa, J.A.; Org, E.; Wang, Z.; Levison, B.S.; Zhu, W.; Wagner, M.A.; Bennett, B.J.; Li, L.; DiDonato, J.A.; et al. Transmission of atherosclerosis susceptibility with gut microbial transplantation. J. Biol. Chem. 2015, 290, 5647–5660. [Google Scholar] [CrossRef]
- Jie, Z.; Xia, H.; Zhong, S.L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The gut microbiome in atherosclerotic cardiovascular disease. Nat. Commun. 2017, 8, 845. [Google Scholar] [CrossRef]
- Karlsson, F.H.; Fak, F.; Nookaew, I.; Tremaroli, V.; Fagerberg, B.; Petranovic, D.; Backhed, F.; Nielsen, J. Symptomatic atherosclerosis is associated with an altered gut metagenome. Nat. Commun. 2012, 3, 1245. [Google Scholar] [CrossRef]
- Lau, K.; Srivatsav, V.; Rizwan, A.; Nashed, A.; Liu, R.; Shen, R.; Akhtar, M. Bridging the Gap between Gut Microbial Dysbiosis and Cardiovascular Diseases. Nutrients 2017, 9, 859. [Google Scholar] [CrossRef]
- Santisteban, M.M.; Qi, Y.; Zubcevic, J.; Kim, S.; Yang, T.; Shenoy, V.; Cole-Jeffrey, C.T.; Lobaton, G.O.; Stewart, D.C.; Rubiano, A.; et al. Hypertension-Linked Pathophysiological Alterations in the Gut. Circ. Res. 2017, 120, 312–323. [Google Scholar] [CrossRef]
- Chen, M.L.; Yi, L.; Zhang, Y.; Zhou, X.; Ran, L.; Yang, J.; Zhu, J.D.; Zhang, Q.Y.; Mi, M.T. Resveratrol Attenuates Trimethylamine-N-Oxide (TMAO)-Induced Atherosclerosis by Regulating TMAO Synthesis and Bile Acid Metabolism via Remodeling of the Gut Microbiota. mBio 2016, 7, e02210–e02215. [Google Scholar] [CrossRef]
- Lim, T.; Ryu, J.; Lee, K.; Park, S.Y.; Hwang, K.T. Protective Effects of Black Raspberry (Rubus occidentalis) Extract against Hypercholesterolemia and Hepatic Inflammation in Rats Fed High-Fat and High-Choline Diets. Nutrients 2020, 12, 1448. [Google Scholar] [CrossRef]
- Annunziata, G.; Maisto, M.; Schisano, C.; Ciampaglia, R.; Narciso, V.; Tenore, G.C.; Novellino, E. Effects of Grape Pomace Polyphenolic Extract (Taurisolo((R))) in Reducing TMAO Serum Levels in Humans: Preliminary Results from a Randomized, Placebo-Controlled, Cross-Over Study. Nutrients 2019, 11, 139. [Google Scholar] [CrossRef]
- Rehman, A.; Tyree, S.M.; Fehlbaum, S.; DunnGalvin, G.; Panagos, C.G.; Guy, B.; Patel, S.; Dinan, T.G.; Duttaroy, A.K.; Duss, R.; et al. A water-soluble tomato extract rich in secondary plant metabolites lowers trimethylamine-n-oxide and modulates gut microbiota: A randomized, double-blind, placebo-controlled cross-over study in overweight and obese adults. J. Nutr. 2023, 153, 96–105. [Google Scholar] [CrossRef]
- Grundy, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C., Jr.; et al. Diagnosis and management of the metabolic syndrome: An American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Circulation 2005, 112, 2735–2752. [Google Scholar] [CrossRef]
- Shimodaira, M.; Niwa, T.; Nakajima, K.; Kobayashi, M.; Hanyu, N.; Nakayama, T. Correlation between mean platelet volume and fasting plasma glucose levels in prediabetic and normoglycemic individuals. Cardiovasc. Diabetol. 2013, 12, 14. [Google Scholar] [CrossRef]
- Pamukcu, B.; Lip, G.Y.; Snezhitskiy, V.; Shantsila, E. The CD40-CD40L system in cardiovascular disease. Ann. Med. 2011, 43, 331–340. [Google Scholar] [CrossRef]
- Ferroni, P.; Basili, S.; Falco, A.; Davi, G. Platelet activation in type 2 diabetes mellitus. J. Thromb. Haemost. JTH 2004, 2, 1282–1291. [Google Scholar] [CrossRef]
- Ferroni, P.; Martini, F.; D'Alessandro, R.; Magnapera, A.; Raparelli, V.; Scarno, A.; Davi, G.; Basili, S.; Guadagni, F. In vivo platelet activation is responsible for enhanced vascular endothelial growth factor levels in hypertensive patients. Clin. Chim. Acta 2008, 388, 33–37. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, Z.; Ye, Z.; Li, Q.; Wen, J.; Tao, X.; Chen, L.; He, M.; Wang, X.; Lu, B.; et al. Lipocalin-2, glucose metabolism and chronic low-grade systemic inflammation in Chinese people. Cardiovasc. Diabetol. 2012, 11, 11. [Google Scholar] [CrossRef]
- Davi, G.; Patrono, C. Platelet activation and atherothrombosis. N. Engl. J. Med. 2007, 357, 2482–2494. [Google Scholar] [CrossRef]
- Kaplan, Z.S.; Jackson, S.P. The role of platelets in atherothrombosis. Hematol. Am. Soc. Hematol. Educ. Program. 2011, 2011, 51–61. [Google Scholar] [CrossRef]
- Slavin, J.L.; Lloyd, B. Health benefits of fruits and vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef]
- Gaziano, J.M.; Manson, J.E.; Branch, L.G.; Colditz, G.A.; Willett, W.C.; Buring, J.E. A prospective study of consumption of carotenoids in fruits and vegetables and decreased cardiovascular mortality in the elderly. Ann. Epidemiol. 1995, 5, 255–260. [Google Scholar] [CrossRef]
- Joshipura, K.J.; Ascherio, A.; Manson, J.E.; Stampfer, M.J.; Rimm, E.B.; Speizer, F.E.; Hennekens, C.H.; Spiegelman, D.; Willett, W.C. Fruit and vegetable intake in relation to risk of ischemic stroke. JAMA 1999, 282, 1233–1239. [Google Scholar] [CrossRef]
- Dutta-Roy, A.K.; Crosbie, L.; Gordon, M.J. Effects of tomato extract on human platelet aggregation in vitro. Platelets 2001, 12, 218–227. [Google Scholar] [CrossRef]
- Obarzanek, E.; Sacks, F.M.; Vollmer, W.M.; Bray, G.A.; Miller, E.R., 3rd; Lin, P.H.; Karanja, N.M.; Most-Windhauser, M.M.; Moore, T.J.; Swain, J.F.; et al. Effects on blood lipids of a blood pressure-lowering diet: The Dietary Approaches to Stop Hypertension (DASH) Trial. Am. J. Clin. Nutr. 2001, 74, 80–89. [Google Scholar] [CrossRef]
- Dizdarevic, L.L.; Biswas, D.; Uddin, M.D.; Jorgenesen, A.; Falch, E.; Bastani, N.E.; Duttaroy, A.K. Inhibitory effects of kiwifruit extract on human platelet aggregation and plasma angiotensin-converting enzyme activity. Platelets 2014, 25, 567–575. [Google Scholar] [CrossRef]
- Barin, A.; Das, R.K.; Bastani, N.E.; Iversen, P.O.; Duttaroy, A.K. Extracts of tamarillo, horned melon, and raspberry, but not extract of pear, inhibit human blood platelet aggregation: Investigating the underlying factors for their differential mechanisms. J. Funct. Foods 2023, 110, 105847. [Google Scholar] [CrossRef]
- Giovannucci, E. Tomatoes, tomato-based products, lycopene, and cancer: Review of the epidemiologic literature. J. Natl. Cancer Inst. 1999, 91, 317–331. [Google Scholar] [CrossRef]
- Giovannucci, E. A review of epidemiologic studies of tomatoes, lycopene, and prostate cancer. Exp. Biol. Med. 2002, 227, 852–859. [Google Scholar] [CrossRef]
- Giovannucci, E.; Rimm, E.B.; Liu, Y.; Stampfer, M.J.; Willett, W.C. A prospective study of tomato products, lycopene, and prostate cancer risk. J. Natl. Cancer Inst. 2002, 94, 391–398. [Google Scholar] [CrossRef]
- O'Kennedy, N.; Crosbie, L.; van Lieshout, M.; Broom, J.I.; Webb, D.J.; Duttaroy, A.K. Effects of antiplatelet components of tomato extract on platelet function in vitro and ex vivo: A time-course cannulation study in healthy humans. Am. J. Clin. Nutr. 2006, 84, 570–579. [Google Scholar] [CrossRef]
- Fuentes, E.; Castro, R.; Astudillo, L.; Carrasco, G.; Alarcon, M.; Gutierrez, M.; Palomo, I. Bioassay-Guided Isolation and HPLC Determination of Bioactive Compound That Relate to the Antiplatelet Activity (Adhesion, Secretion, and Aggregation) from Solanum lycopersicum. Evid. Based Complement. Altern. Med. 2012, 2012, 147031. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, S.; Wang, H.; Bao, L.; Wu, W.; Qi, R. Fruitflow inhibits platelet function by suppressing Akt/GSK3beta, Syk/PLCgamma2 and p38 MAPK phosphorylation in collagen-stimulated platelets. BMC Complement. Med. Ther. 2022, 22, 75. [Google Scholar] [CrossRef]
- Schwager, J.; Richard, N.; Mussler, B.; Raederstorff, D. Tomato Aqueous Extract Modulates the Inflammatory Profile of Immune Cells and Endothelial Cells. Molecules 2016, 21, 168. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, H.; Li, C.; Chen, B.; Gong, H.; Zhao, Y.; Qi, R. Water-Soluble Tomato Extract Fruitflow Alters the Phosphoproteomic Profile of Collagen-Stimulated Platelets. Front. Pharmacol. 2021, 12, 746107. [Google Scholar] [CrossRef]
- Cho, J. Protein disulfide isomerase in thrombosis and vascular inflammation. J. Thromb. Haemost. 2013, 11, 2084–2091. [Google Scholar] [CrossRef]
- Manickam, N.; Sun, X.; Li, M.; Gazitt, Y.; Essex, D.W. Protein disulphide isomerase in platelet function. Br. J. Haematol. 2008, 140, 223–229. [Google Scholar] [CrossRef]
- Jasuja, R.; Passam, F.H.; Kennedy, D.R.; Kim, S.H.; van Hessem, L.; Lin, L.; Bowley, S.R.; Joshi, S.S.; Dilks, J.R.; Furie, B.; et al. Protein disulfide isomerase inhibitors constitute a new class of antithrombotic agents. J. Clin. Invest. 2012, 122, 2104–2113. [Google Scholar] [CrossRef]
- Sheu, J.R.; Hsiao, G.; Chou, P.H.; Shen, M.Y.; Chou, D.S. Mechanisms involved in the antiplatelet activity of rutin, a glycoside of the flavonol quercetin, in human platelets. J. Agric. Food Chem. 2004, 52, 4414–4418. [Google Scholar] [CrossRef]
- O'Kennedy, N.; Crosbie, L.; Whelan, S.; Luther, V.; Horgan, G.; Broom, J.I.; Webb, D.J.; Duttaroy, A.K. Effects of tomato extract on platelet function: A double-blinded crossover study in healthy humans. Am. J. Clin. Nutr. 2006, 84, 561–569. [Google Scholar] [CrossRef]
- O'Kennedy, N.; Crosbie, L.; Whelan, S.; Luther, V.; Horgan, G.; Broom, J.I.; Webb, D.J.; Duttaroy, A.K. Effects of tomato extract on platelet function: A double-blinded crossover study in healthy humans. Am. J. Clin. Nutr. 2006, 84, 561–569. [Google Scholar] [CrossRef]
- Das, R.K.; Datta, T.; Biswas, D.; Duss, R.; O'Kennedy, N.; Duttaroy, A.K. Evaluation of the equivalence of different intakes of Fruitflow in affecting platelet aggregation and thrombin generation capacity in a randomized, double-blinded pilot study in male subjects. BMC Nutr. 2021, 7, 80. [Google Scholar] [CrossRef]
- O'Kennedy, N.; Crosbie, L.; Song, H.J.; Zhang, X.; Horgan, G.; Duttaroy, A.K. A randomised controlled trial comparing a dietary antiplatelet, the water-soluble tomato extract Fruitflow, with 75 mg aspirin in healthy subjects. Eur. J. Clin. Nutr. 2017, 71, 723–730. [Google Scholar] [CrossRef]
- Tian, Z.; Fan, D.; Li, K.; Zhao, D.; Liang, Y.; Ji, Q.; Gao, X.; Ma, X.; Zhao, Y.; Mao, Y.; et al. Four-Week Supplementation of Water-Soluble Tomato Extract Attenuates Platelet Function in Chinese Healthy Middle-Aged and Older Individuals: A Randomized, Double-Blinded, and Crossover Clinical Trial. Front. Nutr. 2022, 9, 891241. [Google Scholar] [CrossRef]
- Krasinska, B.; Osinska, A.; Osinski, M.; Krasinska, A.; Rzymski, P.; Tykarski, A.; Krasinski, Z. Standardised tomato extract as an alternative to acetylsalicylic acid in patients with primary hypertension and high cardiovascular risk - a randomised, controlled trial. Arch. Med. Sci. 2018, 14, 773–780. [Google Scholar] [CrossRef]
- Liu, L.; Xiao, S.; Wang, Y.; Wang, Y.; Liu, L.; Sun, Z.; Zhang, Q.; Yin, X.; Liao, F.; You, Y.; et al. Water-soluble tomato concentrate modulates shear-induced platelet aggregation and blood flow in vitro and in vivo. Front. Nutr. 2022, 9, 961301. [Google Scholar] [CrossRef]
- Fiorentino, A.; D'Abrosca, B.; Pacifico, S.; Mastellone, C.; Scognamiglio, M.; Monaco, P. Identification and assessment of antioxidant capacity of phytochemicals from kiwi fruits. J. Agric. Food Chem. 2009, 57, 4148–4155. [Google Scholar] [CrossRef]
- Karlsen, A.; Svendsen, M.; Seljeflot, I.; Laake, P.; Duttaroy, A.K.; Drevon, C.A.; Arnesen, H.; Tonstad, S.; Blomhoff, R. Kiwifruit decreases blood pressure and whole-blood platelet aggregation in male smokers. J. Hum. Hypertens. 2012, 27, 126–130. [Google Scholar] [CrossRef]
- Brevik, A.; Gaivao, I.; Medin, T.; Jorgenesen, A.; Piasek, A.; Elilasson, J.; Karlsen, A.; Blomhoff, R.; Veggan, T.; Duttaroy, A.K.; et al. Supplementation of a western diet with golden kiwifruits (Actinidia chinensis var.'Hort 16A':) effects on biomarkers of oxidation damage and antioxidant protection. Nutr. J. 2011, 10, 54. [Google Scholar] [CrossRef]
- Chang, W.H.; Liu, J.F. Effects of kiwifruit consumption on serum lipid profiles and antioxidative status in hyperlipidemic subjects. Int. J. Food Sci. Nutr. 2009, 60, 709–716. [Google Scholar] [CrossRef]
- Jung, K.A.; Song, T.C.; Han, D.; Kim, I.H.; Kim, Y.E.; Lee, C.H. Cardiovascular protective properties of kiwifruit extracts in vitro. Biol. Pharm. Bull. 2005, 28, 1782–1785. [Google Scholar] [CrossRef]
- Kroll, M.H.; Schafer, A.I. Biochemical mechanisms of platelet activation. Blood 1989, 74, 1181–1195. [Google Scholar] [CrossRef]
- Brevik, A.; Gaivao, I.; Medin, T.; Jorgenesen, A.; Piasek, A.; Elilasson, J.; Karlsen, A.; Blomhoff, R.; Veggan, T.; Duttaroy, A.K.; et al. Supplementation of a western diet with golden kiwifruits (Actinidia chinensis var.'Hort 16A':) effects on biomarkers of oxidation damage and antioxidant protection. Nutr. J. 2011, 10, 54. [Google Scholar] [CrossRef]
- Chang, W.H.; Liu, J.F. Effects of kiwifruit consumption on serum lipid profiles and antioxidative status in hyperlipidemic subjects. Int. J. Food Sci. Nutr. 2009, 60, 709–716. [Google Scholar] [CrossRef]
- Karlsen, A.; Svendsen, M.; Seljeflot, I.; Laake, P.; Duttaroy, A.K.; Drevon, C.A.; Arnesen, H.; Tonstad, S.; Blomhoff, R. Kiwifruit decreases blood pressure and whole-blood platelet aggregation in male smokers. J. Hum. Hypertens. 2013, 27, 126–130. [Google Scholar] [CrossRef]
- Ferroni, P.; Santilli, F.; Guadagni, F.; Basili, S.; Davi, G. Contribution of platelet-derived CD40 ligand to inflammation, thrombosis and neoangiogenesis. Curr. Med. Chem. 2007, 14, 2170–2180. [Google Scholar] [CrossRef]
- Fuentes, E.J.; Astudillo, L.A.; Gutierrez, M.I.; Contreras, S.O.; Bustamante, L.O.; Rubio, P.I.; Moore-Carrasco, R.; Alarcon, M.A.; Fuentes, J.A.; Gonzalez, D.E.; et al. Fractions of aqueous and methanolic extracts from tomato (Solanum lycopersicum L.) present platelet antiaggregant activity. Blood Coagul. Fibrinolysis Int. J. Haemost. Thromb. 2012, 23, 109–117. [Google Scholar] [CrossRef]
- Hankey, G.J.; Eikelboom, J.W. Aspirin resistance. Lancet 2006, 367, 606–617. [Google Scholar] [CrossRef]
- Ravishankar, D.; Albadawi, D.A.I.; Chaggar, V.; Patra, P.H.; Williams, H.F.; Salamah, M.; Vaiyapuri, R.; Dash, P.R.; Patel, K.; Watson, K.A.; et al. Isorhapontigenin, a resveratrol analogue selectively inhibits ADP-stimulated platelet activation. Eur. J. Pharmacol. 2019, 862, 172627. [Google Scholar] [CrossRef]
- Schumacher, E.; Vigh, E.; Molnar, V.; Kenyeres, P.; Feher, G.; Kesmarky, G.; Toth, K.; Garai, J. Thrombosis preventive potential of chicory coffee consumption: A clinical study. Phytother. Res. 2011, 25, 744–748. [Google Scholar] [CrossRef]
- Nickel, T.; Lackermair, K.; Scherr, J.; Calatzis, A.; Vogeser, M.; Hanssen, H.; Waidhauser, G.; Schonermark, U.; Methe, H.; Horster, S.; et al. Influence of High Polyphenol Beverage on Stress-Induced Platelet Activation. J. Nutr. Health Aging 2016, 20, 586–593. [Google Scholar] [CrossRef]
- Draijer, R.; de Graaf, Y.; Slettenaar, M.; de Groot, E.; Wright, C.I. Consumption of a polyphenol-rich grape-wine extract lowers ambulatory blood pressure in mildly hypertensive subjects. Nutrients 2015, 7, 3138–3153. [Google Scholar] [CrossRef]
- Ras, R.T.; Zock, P.L.; Zebregs, Y.E.; Johnston, N.R.; Webb, D.J.; Draijer, R. Effect of polyphenol-rich grape seed extract on ambulatory blood pressure in subjects with pre- and stage I hypertension. Br. J. Nutr. 2013, 110, 2234–2241. [Google Scholar] [CrossRef]
- Flammer, A.J.; Sudano, I.; Wolfrum, M.; Thomas, R.; Enseleit, F.; Periat, D.; Kaiser, P.; Hirt, A.; Hermann, M.; Serafini, M.; et al. Cardiovascular effects of flavanol-rich chocolate in patients with heart failure. Eur. Heart J. 2012, 33, 2172–2180. [Google Scholar] [CrossRef]
- Loo, B.M.; Erlund, I.; Koli, R.; Puukka, P.; Hellstrom, J.; Wahala, K.; Mattila, P.; Jula, A. Consumption of chokeberry (Aronia mitschurinii) products modestly lowered blood pressure and reduced low-grade inflammation in patients with mildly elevated blood pressure. Nutr. Res. 2016, 36, 1222–1230. [Google Scholar] [CrossRef]
- Zhang, X.; McGeoch, S.C.; Megson, I.L.; MacRury, S.M.; Johnstone, A.M.; Abraham, P.; Pearson, D.W.; de Roos, B.; Holtrop, G.; O'Kennedy, N.; et al. Oat-enriched diet reduces inflammatory status assessed by circulating cell-derived microparticle concentrations in type 2 diabetes. Mol. Nutr. Food Res. 2014, 58, 1322–1332. [Google Scholar] [CrossRef]
Figure 1.
Hyperactivity of platelets and its impact on cardiovascular health. Platelets play an essential role in CVD, both in the pathogenesis of atherosclerosis and in the development of acute thrombotic events. Hyperactive platelets are involved in the development of atherosclerosis through different mechanisms, such as membrane shedding, growth factor secretion, and the expression of several adhesive factors. In addition, hyperactive platelets are involved in the well-known penultimate thrombotic events.
Figure 1.
Hyperactivity of platelets and its impact on cardiovascular health. Platelets play an essential role in CVD, both in the pathogenesis of atherosclerosis and in the development of acute thrombotic events. Hyperactive platelets are involved in the development of atherosclerosis through different mechanisms, such as membrane shedding, growth factor secretion, and the expression of several adhesive factors. In addition, hyperactive platelets are involved in the well-known penultimate thrombotic events.
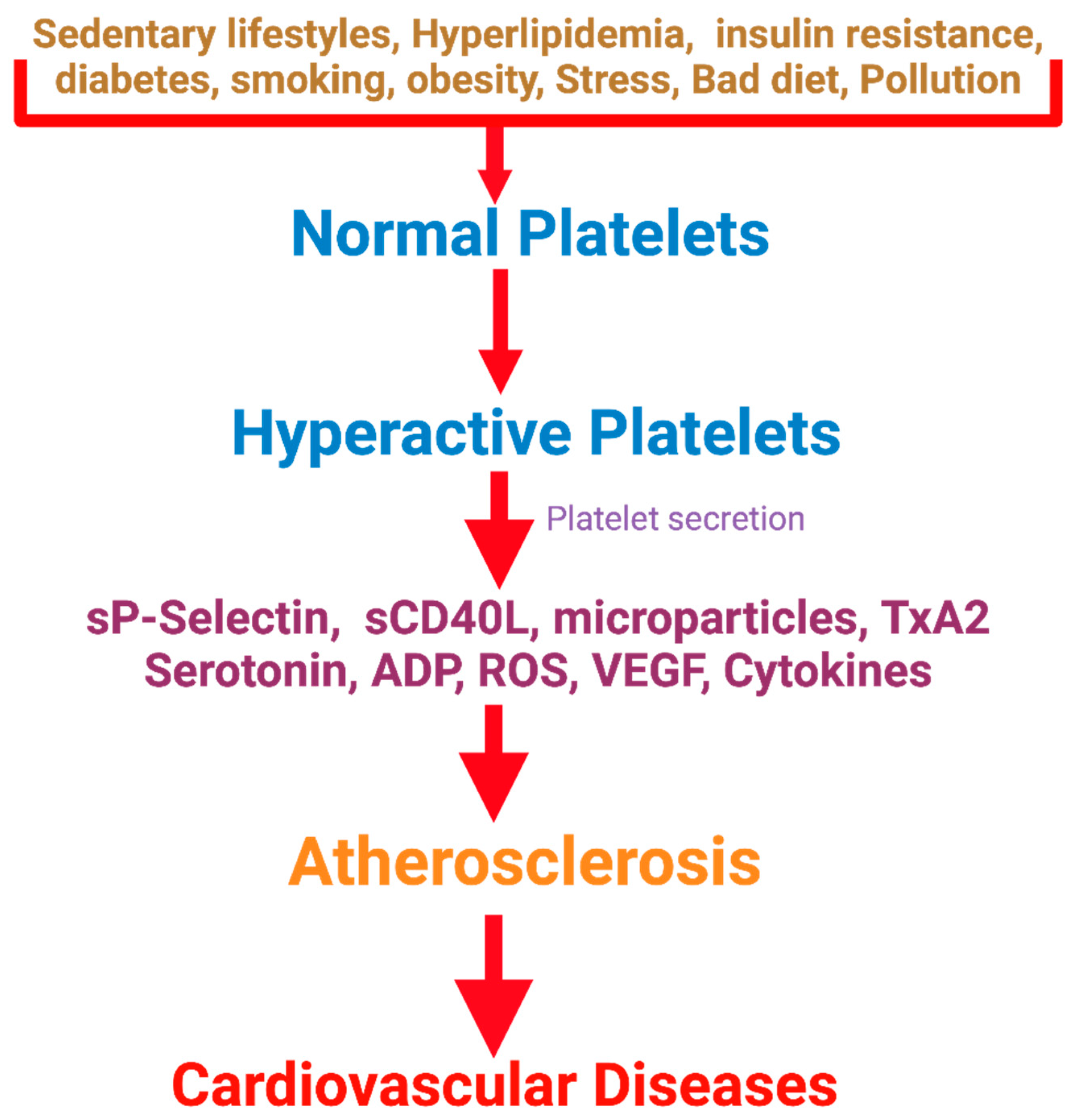
Figure 2.
Roles of Platelets in several hemostatic and non-hemostatic pathways. Platelets are versatile cells engaged in numerous pathophysiological processes, including inflammation and immunity, angiogenesis, regeneration, and carcinogenesis. They are also crucial in thrombosis and hemostasis via various molecular and cellular events. This figure introduces the emerging role of platelets in the immune system, vascular biology, tumorigenesis, and beyond.
Figure 2.
Roles of Platelets in several hemostatic and non-hemostatic pathways. Platelets are versatile cells engaged in numerous pathophysiological processes, including inflammation and immunity, angiogenesis, regeneration, and carcinogenesis. They are also crucial in thrombosis and hemostasis via various molecular and cellular events. This figure introduces the emerging role of platelets in the immune system, vascular biology, tumorigenesis, and beyond.
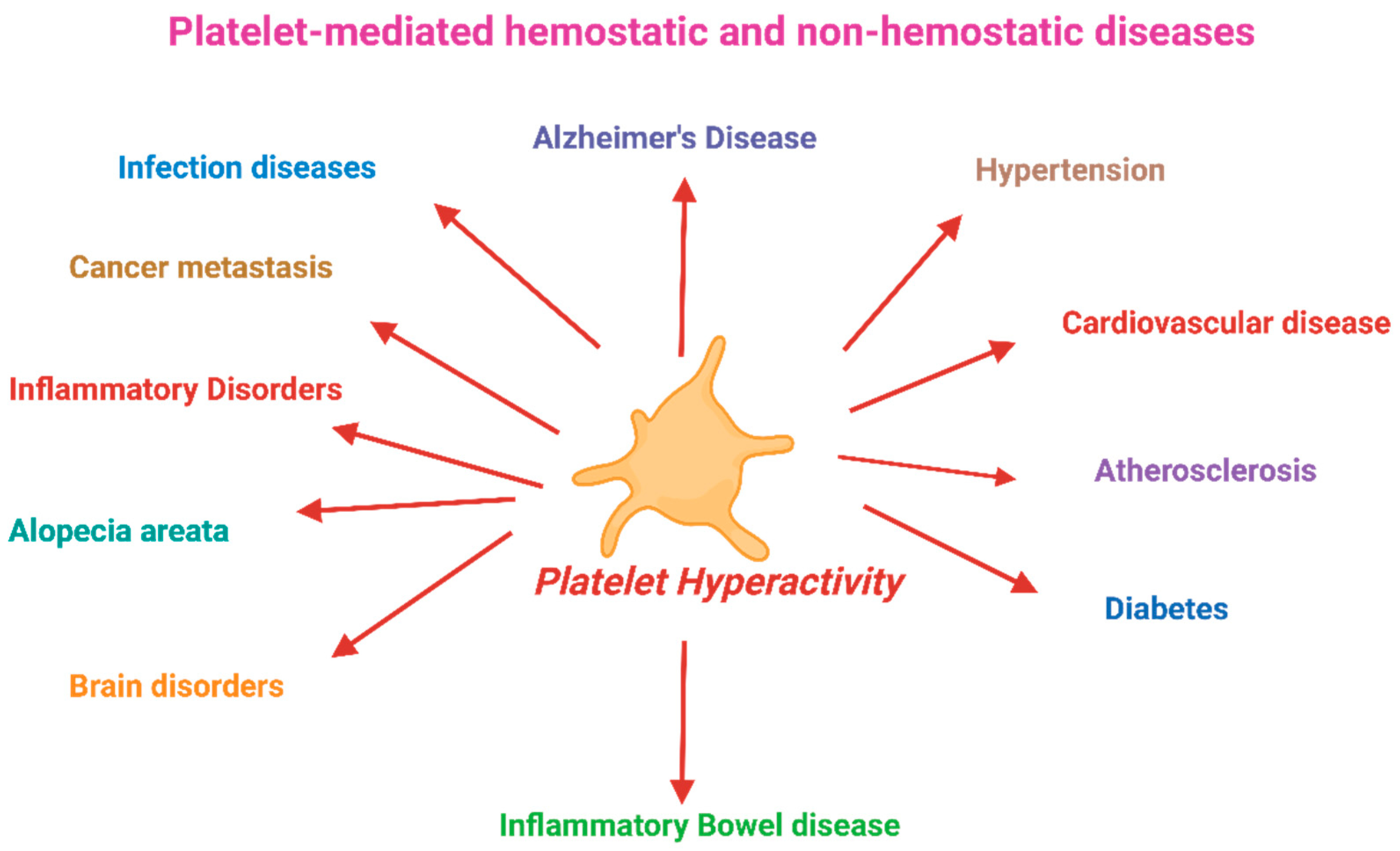

Table 1.
Ex vivo effects of polyphenols on platelet aggregation in animals and human trials.
| Source | Models | Experimental conditions | Conclusions |
|---|---|---|---|
| Chlorogenic acid | Mice (n=18) | Inhibition of in vivo thrombus formation | Inhibited thrombus formation[144] |
| Anthocyanin cyaniding-3-glucoside from purified black rice | Mice (n=60) | Mice randomly assigned to 3 groups (n=20): control group, high-fat diet, or a high-fat diet+ Anthocyanin cyaniding-3-glucoside | Decreased platelet activation, and inhibited platelet ATP release [145] |
| Red wine polyphenols (Provinols™) | Rats (n= 149) | Rats were randomly grouped as treated with or without aldosterone-salt, with or without Provinols (20 mg/kg/day) or spironolactone (30 mg/kg/day) for 4 weeks | Provinols decreased circulating levels microparticles [215]. |
| Cocoa | Healthy males (n=16) | Double-blind, crossover study. Placebo-controlled. Eight subjects in two groups (trains and untrained) randomly received placebo or cocoa polyphenol (236 mg/day) for a week and then afterwards subjected to one hour of exercise | No change in collagen induced platelet aggregation post exercise. ATP release higher post exercise in both groups. Cocoa supplementation did not normalize platelet activity after exercise[143] . |
| Chicory coffee | Healthy subjects (n=27) | Chicory coffee (300 mL) consumed every day for 1 week | Effect on platelet aggregation is variable depending on the agonists used [216]. |
| High polyphenol beverage | Healthy Athletes (n = 103) | Group 1 received a polyphenol-rich beverage, Group 2 a placebo in a randomized, double-blind study. Samples were collected three weeks before, one day before, immediately, as well as 24 h and 72 h, after a marathon run | Control group demonstrated a 2.2-fold increase in platelet aggregation after marathon completion. But there was no increase in platelet aggregation in polyphenol-rich beverage group [217]. |
| Polyphenol-rich grape wine | Untreated, mildly hypertensive subjects (n =60) | Grape juice extract; grape and wine extract each for 4 weeks including a 2-week run-in period in a double-blind placebo-controlled crossover study. | There was no effect on ADP, collagen, or epinephrine induced platelet aggregation[218]. |
| Polyphenol-rich grape seed extract | Untreated subjects with pre and stage 1 hypertension (n=35) | Double-blind, placebo-controlled, randomized, parallel-group intervention with 300 mg/day grape seed extract capsule. Eight-week duration study |
Platelet aggregation was not affected [219] |
| Flavanol-rich chocolate | Patients with congestive heart failure (n =20) | 2h after after consumption of a chocolate bar and 4 weeks of consumption (two chocolate bars/day) in a double-blind, randomized placebo-controlled trial. | Platelet adhesion significantly decreased 2 h after flavanol-rich chocolate ingestion. But no effect after 2- and 4-week supplementation[220]. |
| Chokeberry (Aronia mitschurinii) products | Patients with untreated mild hypertension (n =38) | Cold-pressed 100% chokeberry juice (300 mL/day) and oven-dried chokeberry powder (3 g/day), or placebo for 8 weeks without washout in a single blinded crossover trial for 16-week. | No change in platelet aggregation response [221] |
| Oats | Type 2 diabetes subjects (n =22) | Randomized crossover involving 8-week intervention with either oat enriched diet or a standard dietary advised diet. Pre-intervention habitual intakes were used to compare responses. | Decreased in tissue factor-activated platelets (CD142) after oat rich diet than habitual or standard advised diet. Decreased in tissue factor positive PMPs and fibrinogen-positive PMPs with oat enriched diet intervention[222] |
| Anthocyanin rich beverage | Sedentary subjects (n =21) | Queen garnet plum juice (200 mL/day) were consumed for 28 days in a double-blind placebo controlled study | Reduced ADP-, collagen- and ARA-induced platelet aggregation. Reduced P-selectin expression |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



