
Submitted:
05 October 2024
Posted:
07 October 2024
You are already at the latest version
Abstract
In this study, Portulaca oleracea (purslane) was investigated as a potential antioxidant supplement in cooked sausages, with a focus on its effects on lipid oxidation, fatty acid composition, and antioxidant activity. The fatty acid profile of the sausages enriched with 1.2% of purslane powder, revealed a 1.3-fold increase in alpha-linolenic acid (ALA), an essential omega-3 fatty acid. Improved oxidative stability during refrigerated storage was observed, with peroxide values of 10.9 meq/kg in the sausages with purslane by day 10, compared to 12.5 meq/kg in the control sausages. Thiobarbituric acid (TBA) values, reflecting lipid peroxidation, were also significantly lower in the sausages with purslane. The antioxidant capacity was significantly enhanced in the sausages containing purslane, which demonstrated a Ferric Reducing Antioxidant Power (FRAP) of 13.5 mg GAE/g, whereas the control sausages showed undetectable FRAP levels. Additionally, the DPPH radical scavenging activity in the sausages with purslane was 21.70%, compared to 13.73% in the control. These findings suggest that purslane not only improves the nutritional profile of meat products by increasing beneficial fatty acids but also provides substantial protection against oxidative spoilage. Portulaca oleracea offers a promising natural alternative to synthetic antioxidants, enhancing both the shelf life and quality of processed meats.
Keywords:
Purslane
; Sausages
; Lipid oxidation
; Fatty acid composition
; Thiobarbituric acid
; Ferric Reducing Antioxidant Power
; DPPH radical scavenging activity
; Shelf life
1. Introduction
Cardiovascular diseases (CVDs) remain one of the predominant causes of death worldwide, emphasizing the urgent need for effective preventive measures within contemporary healthcare systems. According to the World Health Organization, CVDs are responsible for approximately 17.9 million deaths annually, representing about 32% of global mortality rates [1]. Hyperlipidemia, marked by elevated concentrations of lipids in the bloodstream, is widely acknowledged as a major contributor to the development of cardiovascular complications [2]. Targeting serum cholesterol reduction, a critical element of blood lipids, is a well-established method for minimizing cardiovascular risk and managing related metabolic disorders, such as diabetes and obesity [3]. Extensive clinical trials and observational studies have repeatedly confirmed that lifestyle modifications, notably the integration of regular physical activity and tailored dietary practices, play a pivotal role in optimizing lipid profiles. These strategies are associated with decreased levels of total cholesterol, low-density lipoprotein cholesterol (LDL-C), triglycerides (TG), and an elevation in high-density lipoprotein cholesterol (HDL-C) [4].
Approximately a decade ago, the primary interventions for reducing plasma LDL-C were predominantly limited to dietary adjustments and pharmacological treatments, with statins being the cornerstone of lipid-lowering therapy [5]. Although statins have proven highly effective in lowering LDL-C and reducing cardiovascular events, their long-term use has been linked to adverse effects, including myopathy and a heightened risk of diabetes [6]. Consequently, there has been a growing interest within the scientific community in exploring nutraceuticals and functional foods as alternative approaches for managing hyperlipidemia with a better safety profile [7]. This shift has been particularly noticeable in Mediterranean regions, where research into plant-derived bioactive compounds has gained momentum due to their potential cardiovascular benefits [8]. Functional foods, which offer health-promoting properties beyond basic nutritional value, have thus garnered significant attention for their potential to support both preventive and therapeutic strategies in cardiovascular care.
The rising incidence of non-communicable diseases (NCDs), particularly cardiovascular disorders, has significantly amplified the need for natural and sustainable alternatives to traditional pharmacotherapies. Among these, omega-3 fatty acids have garnered considerable scientific attention due to their well-documented cardioprotective effects. Extensive epidemiological research has revealed that omega-3 supplementation can lead to a notable decrease in cardiovascular events, such as mortality, non-fatal myocardial infarction, and non-fatal stroke [9]. Specifically, daily consumption of 1.5 to 3 grams of alpha-linolenic acid (ALA) has been associated with a marked reduction in cardiovascular-related mortality [10]. While dietary intake of omega-3 remains the ideal approach, supplementation serves as a viable solution for populations with limited access to these crucial fatty acids through their regular diet [11].
In this regard, Portulaca oleracea (commonly referred to as purslane) has attracted significant attention as a potential functional food. Belonging to the Portulacaceae family, which includes over 120 species predominantly composed of succulent herbs and shrubs, Portulaca oleracea stands out for its remarkable adaptability and resilience in adverse environmental conditions, such as saline and nutrient-deficient soils. This makes it a promising and sustainable crop, particularly in regions grappling with the effects of climate change and soil degradation [12]. Additionally, its high seed viability and competitive growth advantage enhance its suitability as a functional plant with substantial nutritional and therapeutic potential [13].
Historically, Portulaca oleracea has been utilized both as a dietary component and in traditional medicinal practices, primarily due to its abundant profile of bioactive compounds [14]. These bioactives include a wide array of flavonoids, alkaloids, organic acids, vitamins, minerals, sterols, and essential fatty acids, contributing to its considerable nutritional and medicinal value [15]. High concentration of alpha-linolenic acid (ALA), an essential omega-3 fatty acid comprising an 18-carbon chain with three cis-double bonds, is of particular interest. Alpha-linolenic acid (ALA) is essential for maintaining human health, as the body is unable to produce it naturally. Therefore, it must be sourced from dietary intake. ALA is fundamental to processes related to human development, growth, and the prevention of various diseases [16]. Furthermore, ALA serves as a precursor to longer-chain omega-3 fatty acids, such as eicosapentaenoic acid (EPA), docosapentaenoic acid (DPA), and docosahexaenoic acid (DHA), although the conversion rate is quite limited and inefficient [17]. Nevertheless, these long-chain omega-3 fatty acids are renowned for their antioxidant and anti-inflammatory properties, conferring numerous health benefits [18].
The abundant concentration of long-chain polyunsaturated fatty acids (PUFAs) in Portulaca oleracea positions it as a noteworthy alternative source of these critical nutrients, with promising implications for managing hyperlipidemia [19]. An increasing body of research validates the effectiveness of Portulaca oleracea supplementation in modulating lipid profiles, showing marked reductions in total cholesterol, LDL-C, and triglycerides, alongside elevations in HDL-C [20,21]. Furthermore, the bioactive constituents of this plant have exhibited beneficial influences on glucose regulation, highlighting its potential therapeutic value for individuals managing diabetes [22].
In light of the growing consumer interest in functional foods and dietary supplements enriched with omega-3 fatty acids, Portulaca oleracea emerges as a viable option for addressing both nutrient insufficiencies and chronic diseases. Its integration into processed foods, particularly sausages, presents an opportunity to significantly elevate the nutritional profile of such products while providing cardiovascular health benefits [23]. This approach is in line with the modern shift towards healthier, more functional food alternatives, as consumers increasingly gravitate away from traditional processed foods, often criticized for their high levels of saturated fats and sodium.
In a study by Zidan et al. (2014), the impact of lyophilized aqueous extracts from Portulaca oleracea on serum HDL and lecithin levels was investigated using a rat model subjected to a cholesterol-enriched diet [14]. The results demonstrated that supplementation with purslane not only alleviated triglyceridemia and cholesterolemia but also enhanced reverse cholesterol transport, indicating its potential anti-atherogenic properties [24]. Integrating Portulaca oleracea into common dietary staples presents a novel approach for boosting the intake of cholesterol-lowering bioactive compounds, thereby supporting healthier lipid profiles and promoting cardiovascular health [25].
Portulaca oleracea is also a rich source of bioactive compounds, including alkaloids, catecholamines, organic acids, anthocyanins, and flavonoids [26,27]. These compounds possess significant antioxidant, anti-inflammatory, and antimicrobial activities, which not only offer health benefits but also improve food preservation [28]. The antioxidant capabilities of these compounds may also be crucial in reducing oxidative stress within processed foods, thereby prolonging shelf life and improving product safety [29].
Consequently, this study aimed to assess the chemical composition of meat products enriched with Portulaca and to investigate its viability as an alternative or complementary ingredient for the food industry, especially in the creation of functional foods. The outcomes of this research could facilitate the broader incorporation of Portulaca into contemporary dietary approaches, meeting both nutritional needs and health-related challenges.
2. Materials and Methods
2.1. Materials
The sausages were produced using standard methods and processes such as grinding the meat, mixing it with spices and additives, stuffing it into casings, and cooking. The purslane extract powder was added during the grinding process in an amount of 1.2% of the total sausage mixture mass. This amount was selected because our pilot study has been shown that a higher amount of purslane powder negatively affects the organoleptic properties of the final product (imparting a bitter taste). The thermal processing was carried out until the internal temperature of the sausage reached 72˚C. The samples were produced at the experimental facility of Saken Seifullin Kazakh Agro Technical Research University.
2.2. Determination of Fatty Acid Composition
Methyl esters of fatty acids were analyzed using an Agilent 7890 gas chromatograph (Agilent Technologies, USA) equipped with a flame ionization detector and an HP-Innowax capillary column (60 m x 0.32 mm x 0.5 µm) under a nitrogen flow. The temperature gradient was set from 100 to 260 °C at a rate of 10 °C/min. A 1 µL sample was injected, with a split ratio of 1:100, and the detector temperature was maintained between 250 and 300 °C. A Supelco No. 47885U standard mixture of fatty acid methyl esters (C6 to C24) was used as a reference, with automatic data processing. Quantitative analysis of fatty acids was conducted using the internal standard method.
2.3. Determination of Color Characteristics
The color properties of the samples were analyzed with a Konica Minolta CM-2300d spectrophotometer. Prior to the measurements, the spectrophotometer underwent both zero and white calibration, following the manufacturer's guidelines. Color characteristics were assessed through five parallel measurements, utilizing the CIE-Lab* color system, which is widely recognized as a universal color space expressed in Cartesian coordinates. Within this system, L* denotes lightness, a* represents redness, and b* indicates yellowness, all expressed in dimensionless units. The final result for each sample was derived as the arithmetic mean of the five measurements.
To evaluate color stability under light exposure, the criterion of color stability (C) was applied. The color stability was calculated using the following formula:
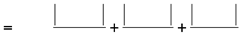
Where L1, L2 is the value of the light index before and after exposure to light;
a1, a2 – the value of the redness index before and after exposure to light;
b1, b2 – the value of the yellowness index before and after exposure to light.
To evaluate the stability of color characteristics under light exposure, the samples were exposed to an artificial light source (fluorescent incandescent lamp, with a power output of at least 40 watts). After one hour of exposure, instrumental measurements were conducted to monitor the changes in the color parameters, providing quantitative data on the extent of color alteration during the experiment.
2.4. Analysis of the Molecular Weight Distribution of Protein Fractions in the Sample Was Carried OUT Using One-Dimensional Electrophoresis
A 100 mg sample was mixed with 500 μL of lysis buffer (4.5 M urea, 2.5% β-mercaptoethanol, 1% Triton X-100, 1% ampholytes with pH 3–10) and centrifuged at 14,000 rpm for 20 min. The supernatant was carefully separated, and an equal volume of protein buffer was added to the mixture. The protein buffer was prepared by combining 1 mL of 10% sodium dodecyl sulfate (SDS), 250 μL of concentrated β-mercaptoethanol, 625 μL of 0.5 M Tris-HCl, 1.5 g of urea, and bromophenol blue as a dark stain. The total volume of the solution was adjusted to 5 mL with distilled water. Following this, the samples were subjected to heating in a boiling water bath for 5 min. Protein visualization was achieved using Coomassie G-250 staining, with the staining solution consisting of 10% acetic acid, 25% isopropanol, and 0.05% Coomassie G-250. To eliminate excess unbound dye, 10% acetic acid was used as a washing solution. For densitometric analysis, one-dimensional electrophoresis gels in their wet state were utilized. High-resolution digital images were captured using a Bio-5000 Plus scanner (Serva, Germany) at 600 ppi in 2D-RGB mode. The obtained digital images were subsequently refined using graphic editing software for further analysis.
2.5. Determination of Ferric Reducing Antioxidant Power (FRAP) and Antioxidant Activity Using the 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Assay
Ferric Reducing Antioxidant Power (FRAP) Assay: The FRAP assay was conducted using BHT and α-tocopherol as standard antioxidants. To perform the assay, 1 mL of the extract at varying dilutions was combined with 2.5 mL of phosphate buffer (0.1 M, pH 6.6) and 2.5 mL of potassium ferricyanide solution (1%, w/v). The mixture was incubated at 50°C for 20 minutes. Following incubation, 2.5 mL of trichloroacetic acid solution (10%, w/v) was added. From this mixture, 2.5 mL was withdrawn and combined with 2.5 mL of deionized water and 0.5 mL of ferric chloride solution (0.1%, w/v). The solution was allowed to stand for 30 minutes before the absorbance was measured at 700 nm. The FRAP values were expressed as milligrams of gallic acid equivalents (GAE) per gram of dry extract (mg GAE/g).
DPPH Radical Scavenging Activity Analysis: For the DPPH assay, 2 mL of a DPPH solution (0.1 mg/mL in methanol) was mixed with 2 mL of sample solutions at a concentration of 200 μg/mL. All analyses were performed in triplicate. The reaction mixture was shaken and incubated in the dark at room temperature for 30 min. After incubation, the absorbance was measured at 517 nm against a blank. Ascorbic acid was prepared as a positive control in a similar manner, with the antioxidant solution being replaced accordingly.
3. Results and Discussion
3.1. Determination of Fatty Acid Composition
Lipid oxidation in meat products frequently results in a decline in consumer quality, affecting key attributes such as flavor, aroma, color, texture, and both nutritional and biological value, while also reducing the shelf life. Furthermore, lipid oxidation products pose significant health risks due to their mutagenic, carcinogenic, and cytotoxic effects on humans [30]. The progression of oxidative processes in fats, influenced by the type of fat and storage conditions, can lead to a noticeable degradation in the organoleptic properties of the product. This deterioration is further exacerbated by alterations in the fats during storage, which subsequently diminishes the nutritional value of the meat. The formation of carbonyl compounds during fat oxidation also promotes carbonyl-amine reactions, altering the color of dried meat and adversely impacting overall quality. These oxidative reactions are accelerated by increased temperature, light exposure, and the presence of catalysts such as meat pigments. Lipid oxidation can proceed through both enzymatic and non-enzymatic pathways, with the rate of oxidation largely governed by the degree of unsaturation in fatty acids. This rate escalates with the number of double bonds present in the fatty acid structure, making polyunsaturated fats particularly susceptible to oxidative degradation. Given the above, the primary interest was in evaluating the total amount of fatty acids in the sausages with purslane to understand oxidation processes, taking into account the quantity of unsaturated fatty acids. Table 1 presents the main fatty acid composition of the studied samples.
From the 36 fatty acids analyzed, only 22 could be quantified. The sausages with purslane exhibited a slightly higher proportion of saturated fatty acids compared to the control sample—29.0% versus 28.6%. Although the difference in saturated fatty acid content is marginal, at just 0.4%, it is noteworthy that the control sample contained two additional fatty acids that were absent in the sausages with purslane. Similarly, there was virtually no difference in the levels of monounsaturated fatty acids between the samples. Both the control and sausages with purslane had nearly identical amounts—38.4% and 38.3%, respectively. When examining individual monounsaturated fatty acids, no significant variations were observed, with the exception of gondoic acid, which was present at 0.5% in the sausages with purslane and 1% in the control sample, and erucic acid, which was detected at 0.3% in the control but was not detected in the sausages with purslane. The most pronounced difference between the samples emerged in the content of omega-3 fatty acids. The incorporation of purslane into the product led to a nearly 1.3-fold increase in linolenic acid levels. The levels of omega-6 fatty acids, much like the monounsaturated fatty acids, showed minimal variation between the control and experimental samples—31.6% and 30.8%, respectively. It is also important to highlight that no trans fatty acid isomers (such as elaidic and linolelaidic acids) were detected in either sample.
When analyzing the total content of unsaturated fatty acids in the control sausage sample, it was observed that their overall level was marginally, yet noticeably, higher compared to the sausages containing Portulaca oleracea (purslane). This disparity could have implications for the shelf life and the susceptibility of fats to oxidative spoilage during storage, as unsaturated fatty acids, characterized by double or triple bonds between adjacent carbon atoms, are more prone to oxidation. In contrast, the inclusion of purslane in the sausage formulation appeared to exert a stabilizing effect on the oxidation process, as evidenced by the evaluation of oxidative spoilage indices, which are detailed later in the report. Additionally, sausages with purslane exhibited higher levels of stearic acid, a saturated fatty acid. Currently, synthetic antioxidants are extensively employed in the meat industry to prevent the oxidation of lipids and proteins. However, oxidative reactions during the production, retail, and storage phases of meat and meat products induce undesirable physicochemical alterations and off-flavors, detrimentally impacting product quality. These changes often lead to consumer dissatisfaction and potential economic losses. One common strategy to address this issue has been the use of synthetic antioxidants. However, with rising consumer awareness of health concerns and a growing preference for natural additives, the demand for natural alternatives to synthetic antioxidants has intensified [31]. The objective of this study was to assess the protective effects of Portulaca oleracea extract against lipid and protein oxidation in meat during refrigerated storage over a 10-day period. The progression of oxidative spoilage in cooked products is presented in Table 2, Table 3 and Table 4.
It can be concluded that the accumulation of peroxide values occurs at a relatively fast rate, likely driven by autoxidation. Autoxidation refers to the reaction between oxygen and unsaturated lipids, leading to the formation of hydroperoxides, which subsequently undergo further transformations. Since the accumulation of hydroperoxides is measured by the peroxide value, their levels initially rise and then begin to decrease, as was observed in the sausages containing Portulaca oleracea starting from day 10 of storage. On day 8, the peroxide value in the sausages with purslane reached 10.1 meq/kg, and by day 10, it had only slightly increased to 10.9 meq/kg. This is in contrast to the control samples, where the oxidation and accumulation of peroxides were more pronounced, with values rising from 8.6 meq/kg on day 8 to 12.5 meq/kg on day 10. This difference may be attributed to the antioxidant properties of purslane, which appears to have a stabilizing effect, or possibly due to a reduction in oxidation-initiating compounds during the first six days of storage. It is worth noting that, after day 6, the rate of peroxide accumulation slowed significantly.
The extent of oxidative degradation in the fat fraction of the sausages during refrigerated storage was further assessed through the thiobarbituric acid (TBA) value. The progression and severity of oxidative changes in the fat phase of meat products are positively correlated with the TBA value, which quantifies the presence of malondialdehyde. The relative stability of malondialdehyde allows for an accurate evaluation of the accumulation of secondary oxidation products. The formation of malondialdehyde is regarded as one of the adverse outcomes of lipid peroxidation, stemming from the breakdown of polyunsaturated fatty acids under the influence of free radicals, a process often associated with the development of undesirable off-flavors [32]. Data on the malondialdehyde content in the sausage formulations under study are presented in Table 3.
It was determined that during sausage storage, a gradual increase in TBA values was observed across all tested sausage formulations. The accumulation of malondialdehyde, relative to its initial concentration, resulted in a fourfold increase in the control sausages by the 10th day of storage, while sausages containing Portulaca oleracea (purslane) showed a 3.7-fold increase over the same period. Despite this rise, the absolute TBA values for all tested products remained below 0.5 mg/kg, indicating that the lipid fraction remained relatively stable throughout the storage period. It is important to note that TBA concentrations exceeding 0.5 mg/kg indicate the onset of oxidation, while values surpassing 1.0 mg/kg are considered indicative of significant oxidative degradation [32]. The accumulation dynamics of the acid value (AV) in fats demonstrated noticeable signs of oxidative changes only after 8 days of storage. However, the rate of AV increase in the experimental sausages containing purslane was lower compared to the control samples. Interestingly, until the 6th day of storage, the AV in sausages with purslane was slightly higher than in the control, but after this point, the rate of fat spoilage accelerated significantly in the samples without purslane. Based on the analysis of these experimental findings, it can be concluded that the inclusion of Portulaca oleracea extract in sausage formulations has a notable impact on both the rate and extent of lipid oxidation, demonstrating its potential as an effective antioxidant in meat products.
In addition to lipid oxidation, proteins in meat products are also vulnerable to oxidative processes. One of the most commonly employed methods for assessing protein oxidation is the measurement of carbonyl derivatives. In this study, the total content of carbonyl compounds extracted from the protein matrix of both control sausages and those supplemented with Portulaca oleracea (purslane) was evaluated. This assessment was conducted only during the initial days of storage. Unfortunately, due to the lack of extended storage studies, it was not possible to monitor the accumulation of carbonyl compounds over time to fully examine purslane's role as an antioxidant and stabilizer in mitigating protein oxidation. However, it is notable that even in the early stages of storage, the control sausages displayed higher levels of carbonyl compounds compared to those with purslane supplementation. The presence of a higher proportion of unsaturated fatty acids and sulfur-containing amino acids in animal muscles leads to an elevated susceptibility of both lipids and proteins to oxidative processes. Lipid oxidation, in particular, is a critical factor impacting the quality of meat products, as it contributes to color changes, the development of off-flavors and odors, and the formation of toxic compounds, all of which pose potential risks to human health [33]. The oxidation of both lipids and proteins can lead to undesirable changes in meat products from both a technological and sensory standpoint, including discoloration, defects in texture, diminished nutritional value, and negative health implications [34].
A range of essential oils is being investigated for their strong antioxidant properties as natural alternatives to synthetic antioxidants used in the meat industry. Many of these oils are classified as Generally Recognized As Safe (GRAS) and have demonstrated beneficial effects on meat products when used alone or in combination with other essential oils, ingredients, or preservation technologies. The effectiveness of these oils depends on various factors such as concentration, potential synergistic effects, and the method of extraction. While steam distillation remains the most widely used method for extracting essential oils industrially, new extraction technologies have been developed to overcome the limitations of traditional methods, enabling the production of higher-quality essential oils [35]. Portulaca oleracea extract is known for its potent antioxidant activity, attributed to its high concentration of polyphenols, short-chain organic acids, and saccharides. As a natural antioxidant, purslane offers several advantages over synthetic antioxidants, including superior stability, non-toxicity, and medicinal properties. Additionally, it is a low-calorie food with numerous therapeutic benefits [36,37,38]. The use of dried purslane further enhances its shelf life and maintains its usability, as this form retains a greater proportion of its beneficial properties compared to extracts, whether water-based or solvent-based. This makes dried purslane a valuable ingredient for improving both the nutritional and functional properties of meat products.
3.2. Determination of Color Characteristics
In the study on stability before and after light exposure on the indicators of cooked sausages with purslane, the following values were obtained for the main color characteristics: L-lightness, a-redness, and b-yellowness. The data are presented in Table 5.
In the evaluation of color stability in meat products under the influence of thermal processing (heating to 70-72°C), the addition of chemical agents (as per the standard recipe), and during storage, the color characteristics were assessed to determine overall color stability. The results indicated a slight decrease in lightness and redness, with reductions of approximately 2.5% and 2.1% observed in the control sausages and sausages with purslane, respectively. In contrast, the yellowness values showed a slight increase, by 1.22%, after light exposure compared to the initial measurements. The stable color formation in the experimental sausages may be attributed to the inclusion of purslane, which possesses potent antioxidant properties, effectively preventing the oxidation of structural protein components that would otherwise contribute to color degradation.
3.3. Analysis of the Molecular Weight Distribution of Protein Fractions in the Sample by One-Dimensional Electrophoresis
The analysis of the molecular weight distribution of protein fractions in the samples, performed via one-dimensional electrophoresis in polyacrylamide gel, yielded the results shown in Figure 1.
A broad range of protein fractions was identified, including key structural proteins, with no significant differences observed between the control sausages and those containing Portulaca oleracea (purslane). Among the protein bands, several likely correspond to fragments of major proteins such as NADH dehydrogenase (21.7 kDa), frataxin (23.5 kDa), NADH dehydrogenase subunit K (31.1 kDa), myozenin-1 (31.6 kDa), and alpha-1D tubulin chain (50.2 kDa), which were more prominently expressed in the control sample. Additionally, other protein fractions were well-represented, including fragments of cofilin (18.5 kDa), cathepsin B (36.6 kDa), cysteine protease (39.9 kDa), RNA-binding protein (40.0 kDa), cyclin-dependent kinase (42.7 kDa), actin (42.0 kDa), serine/threonine-protein kinase (49.0 kDa), ribonuclease (64.2 kDa), heat shock protein (70.2 kDa), catenin beta-1 (85.5 kDa), spondin-1 (90.9 kDa), desmocollin 2 (100.4 kDa), histidine-rich calcium-binding protein (104.6 kDa), and supervillin (244.8 kDa). Moreover, densitometric analysis revealed two distinct proteins, unique to the purslane family, that were identified in the sausages containing purslane (Table 6). These findings suggest the potential influence of purslane on the protein composition of the product, although the overall protein profile between the two sausage types remains largely similar.
As a result of computer-assisted densitometry, two plant-associated proteins (enzymes) were identified in the sausages containing Portulaca oleracea: 4,5-DOPA dioxygenase extradiol and Maturase K.
4,5-DOPA dioxygenase extradiol plays a crucial role in the formation of betalamic acid, which functions similarly to anthocyanins in other plants, serving as a structural, chromophoric, and bioactive component of plant betalain pigments [39]. This enzyme catalyzes the oxidative cleavage of DOPA, producing 4,5-secodopa, which spontaneously cyclizes to form betalamic acid. Betalains not only serve as physiological pigments but also possess strong radical-scavenging activity [40]. These water-soluble pigments are unique to plants in the Caryophyllaceae family and are simpler to synthesize and regulate compared to anthocyanins and carotenoids, beginning from tyrosine and regulated primarily by the MYB transcription factor [41]. Functionally, betalains are analogous to anthocyanins, attracting animal pollinators and exhibiting high antioxidant and free radical-scavenging properties [42]. In addition, betalains are used commercially as food colorants and additives, further highlighting their practical value [43].
Maturase K (matK) is a plastid gene found in plants and is considered one of the most variable coding genes in angiosperms. It has been proposed as a molecular "barcode" for identifying terrestrial plants [44]. Given its variability, matK could potentially serve as a marker to identify the addition of Portulaca oleracea in food products, though further research is necessary to confirm this hypothesis [45]. While plant mitochondrial genes evolve slowly and are less effective in distinguishing between plant species, matK offers a more promising alternative for plant identification [46].
The inclusion of purslane in meat products is thus a functionally justified approach, as it helps reduce oxidative lipid spoilage under various environmental conditions. This natural additive provides antioxidant protection without the need for synthetic stabilizers or chemical antioxidants in the product formulation, making it a valuable and sustainable ingredient.
3.4. Determination of Ferric Reducing Antioxidant Power (FRAP) and Antioxidant Activity Using the 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Assay
The exploration of the antioxidant potential of plant extracts holds considerable relevance for the advancement of novel, safer natural antioxidants in the food industry. Within this framework, the subsequent phase of the study concentrated on evaluating the Ferric Reducing Antioxidant Power (FRAP) and DPPH radical scavenging activity, as presented in Table 7. This analysis provides critical insights into the efficacy of the extracts in neutralizing oxidative processes, thus offering valuable contributions to the development of functional food ingredients.
The study demonstrated substantial levels of ferric reducing antioxidant power (FRAP) in the experimental sausage samples. The incorporation of Portulaca oleracea into sausage formulations appears to be advantageous in enhancing the antioxidant properties of the final product. As evidenced by the data in Table 7, the sausages with purslane exhibited a significant FRAP value of 13.5 mg GAE/g, while no measurable FRAP activity was detected in the control samples. This highlights the ability of purslane to markedly improve the product's capacity for ferric ion reduction, an important indicator of antioxidant potential. Moreover, the antioxidant activity, as evaluated by DPPH radical scavenging, also showed a pronounced improvement in the experimental sample compared to the control. The DPPH radical scavenging activity in the sausages with purslane reached 21.70%, considerably higher than the 13.73% observed in the control, confirming the enhanced ability of the purslane-enriched sausages to neutralize free radicals. Additionally, the IC50 value for DPPH radical scavenging activity was greater in the sausages with purslane (116.55 µg/mL) compared to the control (76.12 µg/mL), indicating that a higher concentration of the purslane-enriched sample was required to achieve 50% inhibition of DPPH radicals. This further underscores the conclusion that the addition of Portulaca oleracea significantly elevates the antioxidant profile of the sausages.
4. Conclusions
Our results demonstrated that Portulaca oleracea (purslane) is a valuable natural additive for enhancing the antioxidant properties and oxidative stability of cooked sausage products. The inclusion of purslane significantly increased the content of beneficial omega-3 fatty acids, particularly alpha-linolenic acid (ALA), and improved resistance to lipid oxidation, as indicated by lower peroxide and thiobarbituric acid (TBA) values during refrigerated storage. Additionally, the antioxidant capacity of purslane-enriched sausages was markedly higher, with significant improvements in both Ferric Reducing Antioxidant Power (FRAP) and DPPH radical scavenging activity. These findings suggest that purslane not only contributes to the nutritional enhancement of meat products but also offers a natural solution to reducing oxidative spoilage without the need for synthetic additives. The incorporation of Portulaca oleracea has a great potential for the development of functional foods, maintaining product quality and extending shelf life.
Author Contributions
Conceptualization, K.M. and T.T.; methodology, G.T.; validation, G.O.; formal analysis, G.Zh.; investigation, G.T. and G.Zh., V.K.; resources, K.M.; data curation, Sh.A. and A.E.; writing—original draft preparation, K.M.; writing—review and editing, G.Z.; visualization, K.M.; supervision, K.M. All authors have read and agreed to the published version of the manuscript.
Funding
“This research is funded by the Ministry of Science and Higher Education of the Republic of Kazakhstan (AP14871765).
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- World Health Organization. Cardiovascular diseases (CVDs) fact sheet. Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases- (accessed on day month year).
- Jain, K.S.; Kathiravan, M.K.; Somani, R.S.; Shishoo, C.J. The biology and chemistry of hyperlipidemia. Bioorg. Med. Chem. 2007, 15, 4674–4699. [Google Scholar] [CrossRef]
- Mokdad, A.H.; Ballestros, K.; Echko, M.; Glenn, S.; Olsen, H.E.; Mullany, E.; Lee, A.; Khan, A.R.; Ahmadi, A.; Ferrari, A.J. The state of US health, 1990–2016: Burden of diseases, injuries, and risk factors among US states. JAMA 2018, 319, 1444–1472. [Google Scholar] [CrossRef]
- Poli, A.; Visioli, F.; Bianchi, A.; Romeo, F.; Cimaglia, P.; Mambelli, G.; Bernini, W. Nutraceuticals and functional foods for the control of plasma cholesterol levels. Pharmacol. Res. 2018, 134, 51–60. [Google Scholar] [CrossRef]
- Thompson, P.D.; Panza, G.; Zaleski, A.; Taylor, B. Statin-associated side effects. J. Am. Coll. Cardiol. 2016, 67, 2395–2410. [Google Scholar] [CrossRef]
- Baigent, C.; Keech, A.; Kearney, P.M.; Blackwell, L.; Buck, G.; Pollicino, C.; Kirby, A.; Sourjina, T.; Peto, R.; Collins, R. Efficacy and safety of cholesterol-lowering treatment: Prospective meta-analysis of data from 90,056 participants in 14 randomised trials of statins. Lancet 2005, 366, 1267–1278. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 fatty acids and cardiovascular disease: Evidence explained and mechanisms explored. Clin. Sci. 2020, 134, 277–300. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-3 fatty acids and cardiovascular disease: The epidemiological evidence. Environ. Health Prev. Med. 2004, 9, 45–51. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Lemaitre, R.N.; King, I.B.; Song, X.; Huang, H.; Sacks, F.M.; Rimm, E.B.; Wang, M.; Siscovick, D.S. Plasma phospholipid long-chain ω-3 fatty acids and total and cause-specific mortality in older adults: A cohort study. Ann. Intern. Med. 2013, 158, 515–525. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, Â.; Arampatzis, D.A.; Tsiropoulos, N.G.; Petrović, J.; Soković, M.; Barros, L.; Ferreira, I.C.F.R. Seed oil and seed oil byproducts of common purslane (Portulaca oleracea L.): A new insight to plant-based sources rich in omega-3 fatty acids. LWT 2020, 123, 109099. [Google Scholar] [CrossRef]
- Liu, L.; Howe, P.; Zhou, Y.-F.; Hocart, C.; Zhang, R. Fatty acids and β-carotene in Australian purslane (Portulaca oleracea) varieties. J. Chromatogr. A 2000, 893, 207–213. [Google Scholar] [CrossRef]
- Bhuiyan, N.H.; Murakami, K.; Adachi, T. Variation in betalain content and factors affecting the biosynthesis in Portulaca sp. “Jewel” cell cultures. Plant Biotechnol. 2002, 19, 369–376. [Google Scholar] [CrossRef]
- Gu, Y.; Leng, A.; Zhang, W.; Ying, X.; Stien, D. A novel alkaloid from Portulaca oleracea L. and its anti-inflammatory activity. Nat. Prod. Res. 2020, 34, 1–6. [Google Scholar] [CrossRef]
- Zidan, Y.; Bouderbala, S.; Djellouli, F.; Lacaille-Dubois, M.A.; Bouchenak, M. Lyophilized aqueous extract of Portulaca oleracea improves serum high-density lipoproteins and lecithin levels in cholesterol-enriched diet-fed rats. Phytomedicine 2014, 21, 1504–1508. [Google Scholar] [CrossRef]
- Simopoulos, A.P.; Norman, H.A.; Gillaspy, G.E.; Duke, J.A. Common purslane: A source of omega-3 fatty acids and antioxidants. J. Am. Coll. Nutr. 1992, 11, 374–382. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef]
- Dubois, V.; Breton, S.; Linder, M.; Fanni, J.; Parmentier, M. Fatty acid profiles of 80 vegetable oils with regard to their nutritional potential. Eur. J. Lipid Sci. Technol. 2007, 109, 710–732. [Google Scholar] [CrossRef]
- Hadi, A.; Mohammadi, H.; Hadi, Z.; Roshanravan, N.; Kafeshani, M. Cumin (Cuminum cyminum L.) is a safe approach for management of lipid parameters: A systematic review and meta-analysis of randomized controlled trials. Phytother. Res. 2018, 32, 9–16. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Yalcin, H.; Elibol, M.; Sevim, P.; Bakkal, G. The effects of purslane (Portulaca oleracea L.) on serum lipid profiles and oxidative stress in atherogenic rats. Food Res. Int. 2020, 136, 109558. [Google Scholar] [CrossRef]
- Lin, S.; Huang, X.; Cheng, H.; Yang, L.; Zhang, D.; Zhang, Q.; Shi, J.; Zhang, M. Extraction and purification of alpha-linolenic acid from Portulaca oleracea L. and its effects on lipid profiles and oxidative stress in high-fat diet rats. Nutrients 2020, 12, 1704. [Google Scholar] [CrossRef]
- González, M.; Zarzuelo, A.; Guerra, J.; Menéndez, R.; Burrell, M.A.; Risco, S. Hypoglycemic activity of Portulaca oleracea in normal and diabetic rats. J. Ethnopharmacol. 2014, 93, 461–466. [Google Scholar] [CrossRef]
- Dubois, V.; Parmentier, M.; Baudet, D.; Collinet, A.; Breton, S. Fatty acid content and antioxidant activity of Portulaca oleracea extracts. Food Chem. 2007, 104, 1801–1810. [Google Scholar] [CrossRef]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E.; Collins, K.J.; Dennison Himmelfarb, C.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA guideline for the prevention, detection, evaluation, and management of high blood pressure in adults. Hypertension 2018, 71, e13–e115. [Google Scholar] [CrossRef]
- Chan, E.; Bailey, D.; Busta, F.; Cohen, R.; Schwarzschild, M. Clinical effects of plant-based lipids on plasma lipoproteins. Clin. Nutr. 2019, 38, 2349–2357. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, R.; Peng, D.; Zhang, M.; Zhang, H. Antioxidant activity of Portulaca oleracea polysaccharides and their influence on rice starch digestion. Antioxidants 2020, 9, 1075. [Google Scholar] [CrossRef]
- Rydlewski, B.; Nowakowski, A.; Lewandowska, K.; Kwiatkowska, M. Bioactive compounds in Portulaca oleracea seeds and their effects on oxidative stress in a food matrix. J. Agric. Food Chem. 2017, 65, 177–185. [Google Scholar] [CrossRef]
- Voulgaridou, G.P.; Anestopoulos, I.; Franco, R.; Panayiotidis, M.I.; Pappa, A. Antioxidant and anticancer properties of polyphenolic compounds in foods. Antioxid. Redox Signal. 2011, 15, 2027–2041. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Lipid oxidation and improving the oxidative stability of muscle foods. J. Food Lipids 2005, 12, 89–105. [Google Scholar] [CrossRef]
- Cheng, J.H. Lipid Oxidation in Meat. J. Nutr. Food Sci. 2016, 6, 494. [Google Scholar] [CrossRef]
- Pateiro, M. ; Essential oils as natural additives to prevent oxidation reactions in meat and meat products: A review. Food Res. Int. 2018, 113, 156–166. [Google Scholar] [CrossRef]
- Gurinovich, G.V.; Patrakova, I.S.; Kudryashov, L.S. Study of the effect of curing mixture composition on the lipid oxidation process in meat systems. Meat Systems 2018, 3, 48–1. [Google Scholar]
- Fan, X.; Liu, S.; Li, H.; He, J.; Feng, J.; Zhang, X.; Yan, H. Effects of Portulaca oleracea L. extract on lipid oxidation and color of pork meat during refrigerated storage. Meat Sci. 2019, 147, 82–90. [Google Scholar] [CrossRef]
- Estévez, M.; Luna, C. Dietary protein oxidation: A silent threat to human health? Crit. Rev. Food Sci. 2017, 57, 3781–3793. [Google Scholar] [CrossRef]
- Šojić, B. Isolation, Bioactive Potential, and Application of Essential Oils and Terpenoid-Rich Extracts as Effective Antioxidant and Antimicrobial Agents in Meat and Meat Products. Molecules 2023, 28, 2293. [Google Scholar] [CrossRef]
- Gatea, F.; Dumitra-Teodor, E.; Maria-Seciu, A.; Nagodă, E.; Lucian-Radu, G. Chemical constituents and bioactive potential of Portulaca pilosa L. vs. Portulaca oleracea L. Med. Chem. Res. 2017, 26, 1516–1527. [Google Scholar] [CrossRef]
- Zhao, R.; Meng, X.; Jia, G.; Yu, Y.; Song, B. Oral pre-administration of Purslane polysaccharides enhance immune responses to inactivated foot-and-mouth disease vaccine in mice. BMC Vet. Res. 2019, 15, 38. [Google Scholar] [CrossRef]
- Li, Y.; Hu, Y.; Shi, S.; Jiang, L. Evaluation of antioxidant and immuno-enhancing activities of Purslane polysaccharides in gastric cancer rats. Int. J. Biol. Macromol. 2014, 68, 113–116. [Google Scholar] [CrossRef]
- Gandía-Herrero, F.; García-Carmona, F. Characterization of recombinant Beta vulgaris 4,5-DOPA-extradiol-dioxygenase active in the biosynthesis of betalains. Planta 2012, 236, 91–100. [Google Scholar] [CrossRef]
- Kapadia, G.J.; Azuine, M.A.; Sridhar, R.; Okuda, Y.; Tsuruta, A.; Ichiishi, E.; Mukainake, T.; Takasaki, M.; Konoshima, T.; Nishino, H.; Tokuda, H. Chemoprevention of DMBA-induced UV-B promoted, NOR-1-induced TPA promoted skin carcinogenesis, and DEN-induced phenobarbital promoted liver tumors in mice by extract of beetroot. Pharmacol. Res. 2003, 47, 141–148. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, Y.; Long, T.; Wang, S.; Yang, J. Regulation Mechanism of Plant Pigments Biosynthesis: Anthocyanins, Carotenoids, and Betalains. Metabolites 2022, 12, 871. [Google Scholar] [CrossRef]
- Jain, G.; Gould, K.S. Are betalain pigments the functional homologues of anthocyanins in plants? Environ. Exp. Bot. 2015, 119, 48–53. [Google Scholar] [CrossRef]
- Gandia-Herrero, F.; Garcia-Carmona, F. Characterization of recombinant Beta vulgaris 4,5-DOPA-extradiol-dioxygenase active in the biosynthesis of betalains. Planta 2012, 236, 91–100. [Google Scholar] [CrossRef]
- Ahlert, D.; Piepenburg, K.; Kudla, J.; Bock, R. Evolutionary origin of a plant mitochondrial group II intron from a reverse transcriptase/maturase-encoding ancestor. J. Plant Res. 2006, 119, 363–371. [Google Scholar] [CrossRef]
- Jing, Y.; Jian-Hua, X.; Shi-Liang, C. New universal matK primers for DNA barcoding of angiosperms. J. Syst. Evol. 2011, 49, 176–181. [Google Scholar] [CrossRef]
- Chase, M.W.; Salamin, N.; Wilkinson, M.; Dunwell, J.M.; Kesanakurthi, R.P.; Haidar, N.; Savolainen, V. Land plants and DNA barcodes: Short-term and long-term goals. Evol. 2005, 360, 1889–1895. [Google Scholar] [CrossRef]
Figure 1.
One-dimensional electropherogram of sausage samples. Ст – Molecular weight standards: 250, 150, 100, 70, 50, 40, 30, 20, 15, and 10 kDa (from top to bottom). К – Control sausages. О – Sausages with purslane
Figure 1.
One-dimensional electropherogram of sausage samples. Ст – Molecular weight standards: 250, 150, 100, 70, 50, 40, 30, 20, 15, and 10 kDa (from top to bottom). К – Control sausages. О – Sausages with purslane
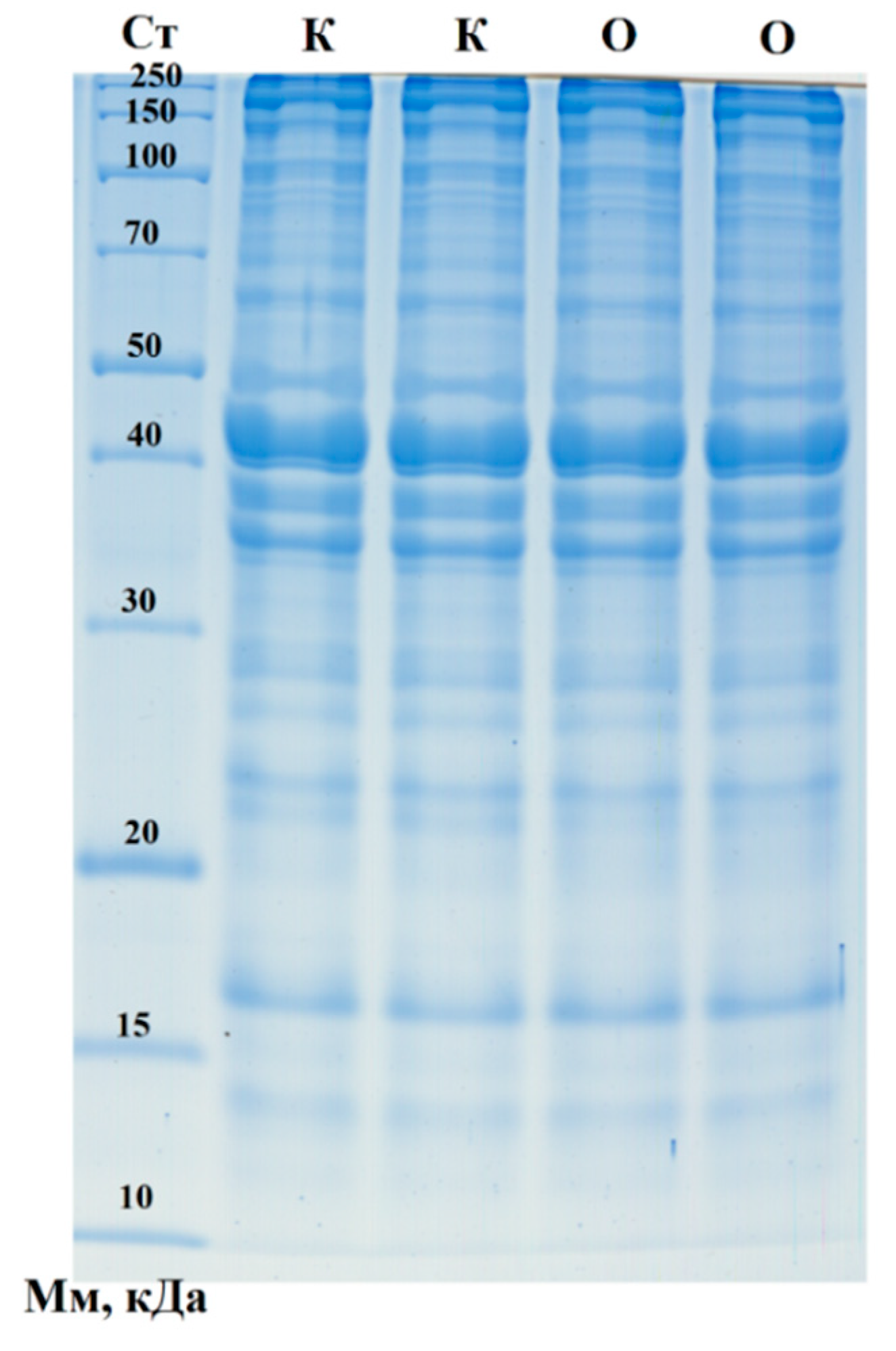

Table 1.
Fatty Acid Composition of Sausage Samples with the Addition of Purslane.
| Fatty acids | Fatty acid content | |
|---|---|---|
| Control sausages | Sausages with purslane | |
| Butyric C4:0 | below 0.1 | below 0.1 |
| Caproic C6:0 | below 0.1 | below 0.1 |
| Caprylic C8:0 | below 0.1 | below 0.1 |
| Capric C10:0 | below 0.1 | below 0.1 |
| Undecylic C11:0 | below 0.1 | below 0.1 |
| Lauric C12:0 | below 0.1 | below 0.1 |
| Tridecanoic C13:0 | below 0.1 | below 0.1 |
| Myristic C14:0 | below 0.1 | below 0.1 |
| Pentadecanoic C15:0 | below 0.1 | below 0.1 |
| Palmitic C16:0 | 19.7 | 19.2 |
| Margaric C17:0 | 0.4 | below 0.1 |
| Stearic C18:0 | 7.6 | 9.4 |
| Arachidic C20:0 | below 0.1 | below 0.1 |
| Heneicosanoic C21:0 | below 0.1 | below 0.1 |
| Behenic C22:0 | 0.6 | 0.5 |
| Tricosanoic C23:0 | 0.3 | below 0.1 |
| Lignoceric C24:0 | below 0.1 | below 0.1 |
| Myristoleic C14:1 | below 0.1 | below 0.1 |
| Cis-10-Pentadecenoic C15:1 | below 0.1 | below 0.1 |
| Palmitoleic C16:1 | 2.8 | 2.7 |
| Heptadecenoic C17:1 | 0.5 | 0.5 |
| Oleic C18:1 | 33.9 | 33.6 |
| Elaidic C18:1 | below 0.1 | below 0.1 |
| Gondoic C20:1 | 0.5 | 1.0 |
| Erucic C22:1 | 0.3 | below 0.1 |
| Nervonic C24:1 | 0.3 | 0.5 |
| Linolenic C18:3ω3 | 1.4 | 1.8 |
| Timnodonic Acid C20:5ω3 | below 0.1 | below 0.1 |
| Eicosatrienoic Acid C20:3ω3 | below 0.1 | below 0.1 |
| Docosahexaenoic C22:6ω3 | below 0.1 | below 0.1 |
| Linoleic C18:2 ω6 | 29.7 | 29.1 |
| Linolelaidic C18:2 ω6 | below 0.1 | below 0.1 |
| Dihomo-γ-linolenic Acid C20:3 ω6 | 0.3 | below 0.1 |
| Arachidonic Acid C20:4 ω6 | 0.9 | 1.0 |
| Eicosadienoic Acid C20:2 ω6 | 0.6 | 0.8 |
| Docosadienoic Acid C22:2 ω6 | below 0.1 | below 0.1 |
Table 2.
Dynamics of fat and protein oxidation (baseline) in products during storage, with the accumulation of peroxide value.
Table 2.
Dynamics of fat and protein oxidation (baseline) in products during storage, with the accumulation of peroxide value.
| Indicators | Concentration | ||
| Storage time, days | Control sausages | Sausages with purslane | |
| Peroxide number, meq/kg | 0 | 3.1±0.3 | 3.5±0.3 |
| 2 | 4.3±0.4 | 4.1±0.4 | |
| 4 | 5.0±0.5 | 4.5±0.5 | |
| 6 | 6.3±0.3 | 7.1±0.4 | |
| 8 | 8.6±0.4 | 10.1±0.5 | |
| 10 | 12.5±0.6 | 10.9±0.5 | |
| Carbonyl compounds, nmol/mg of protein | 109.86 | 105.65 | |
Table 3.
The dynamics of fat oxidation, with the accumulation of thiobarbituric number in sausages during storage.
Table 3.
The dynamics of fat oxidation, with the accumulation of thiobarbituric number in sausages during storage.
| Thiobarbituric number, storage time, days | Concentration, mgMA/kg | ||
| Control sausages | Sausages with purslane | ||
| 0 | Below 0.039 | Below 0.039 | |
| 2 | 0.047±0.005 | 0.041±0.004 | |
| 4 | 0.108±0.011 | 0.086±0.009 | |
| 6 | 0.101±0.010 | 0.094±0.009 | |
| 8 | 0.140±0.014 | 0.109±0.011 | |
| 10 | 0.202±0.020 | 0.153±0.015 | |
Table 4.
The dynamics of fat oxidation, with the accumulation of acid number in sausages during storage.
Table 4.
The dynamics of fat oxidation, with the accumulation of acid number in sausages during storage.
| Acid value, storage time, days | Concentration, mg KOH/g | |
| Control sausages | Sausages with purslane | |
| 0 | 2.0±0.2 | 2.2±0.2 |
| 2 | 2.7±0.2 | 2.9±0.2 |
| 4 | 3.2±0.2 | 3.4±0.2 |
| 6 | 3.4±0.2 | 3.6±0.3 |
| 8 | 4.0±0.2 | 3.9±0.3 |
| 10 | 4.8±0.2 | 4.2±0.3 |
Table 5.
The dynamics of fat oxidation, with the accumulation of acid number in sausages during storage.
Table 5.
The dynamics of fat oxidation, with the accumulation of acid number in sausages during storage.
| Color characteristics before exposure to light | Color characteristics after exposure to light | Color stability, % | ||||
| L-lightness | a-redness | b-yellowness | L-lightness | a-redness | b-yellowness | |
| Control sausages | ||||||
| 62.64±0.93 | 17.18±0.40 | 14.83±0.96 | 60.05±1.25 | 15.32±0.37 | 16.05±1.04 | 92.30±2.17 |
| Sausages with purslane | ||||||
| 59.01±0.63 | 6.85±0.20 | 15.62±0.42 | 56.94±1.23 | 5.51±0.37 | 16.84±0.61 | 89.68±1.74 |
Table 6.
Proteins identified as belonging to the Portulaca.
| Protein | Gene | Organism | Molecular weight, kDa |
|---|---|---|---|
| 4,5-DOPA dioxygenase estradiol | DODA | Portulaca grandiflora | 29,92 |
| Maturase K | matK | Portulaca oleracea | 60,87 |
Table 7.
Ferric Reducing Antioxidant Power (FRAP) and Antioxidant Activity (DPPH).
| Name of indicators | Results | |
| Control sausages | Sausages with purslane | |
| Ferric Reducing Antioxidant Power (FRAP), mg GAE/g | Not detected | 13.5 ± 0.05 |
| DPPH Radical Scavenging Activity, % | 13.73±0.005 | 21.70±0.01 |
| IC50 of DPPH Radical Scavenging Activity, µg/mL | 76.12±5.04 | 116.55±10.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



