
Submitted:
06 October 2024
Posted:
07 October 2024
You are already at the latest version
Abstract
The increasing incidence of hepatocellular carcinoma (HCC) together with the poor effectiveness of the available therapeutic treatments, make an early diagnosis and an effective screening of utmost relevance. Liquid biopsy represents a potential novel approach for early HCC detection and monitoring. The identification of blood markers has many desirable features among which the absence of any significant risk for the patients, the possibility to be used as screening tool and to perform multiple tests thus allowing a real-time monitoring of HCC evolution. Unfortunately, the available blood markers of HCC have several limitations mostly related to specificity and sensitivity. In this context, the employment of non-coding RNAs (ncRNAs) may represent an interesting and novel diagnostic approach. ncRNAs which includes among others, micro interfering RNAs (miRNAs), long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), regulate human gene expression via the interaction with their target mRNA. Notably, their expression can be altered in HCC thus reflecting the diseased status. In this review, we discuss some noticeable works, which describe the use of miRNAs, lncRNAs and circRNA as HCC biomarkers. Despite some open aspects related to ncRNA use, the presented works strongly support the potential effectiveness of these molecules as diagnostic/prognostic markers for HCC.
Keywords:
hepatocellular carcinoma
; miRNA
; lncRNA
; circRNA
; prognosis
; diagnosis
1. Introduction: Hepatocellular Carcinoma
The increasing incidence [1] of liver cancer together with the fact that nowadays it represents the third leading cause of cancer deaths worldwide just after lung and colorectal cancer [2], make of liver cancer a global health challenge [3]. Primary liver cancer is a malignant tumor characterized by different histological types. Hepatocellular carcinoma (HCC), the most common type of primary liver cancer, (75%-85% of primary liver cancer cases [4]) originates from hepatocytes [5]. The risk of HCC development has marked regional differences [4]: in the world, age standardized incidence/mortality are highest in Eastern Asia, South-Eastern Asia and Northern Africa [2]. HCC prognosis is very poor with a 5-year overall survival rate less than 12% [6]. More than 90% of HCC occur in the context of chronic liver disease, especially liver fibrosis [7]. In this regard, it is now known that liver fibrosis produce a “premalignant” environment [7] favoring HCC development [8]. Most of the HCC diagnoses occur in 60/70 years old subjects with men being mostly affected [9]. Alcohol consumption, hepatitis B/C infection, non-alcoholic fatty liver disease, non-alcoholic steatohepatitis, diabetes, cigarette smoking, exposure to aflatoxins are the most relevant risk factors [5].
HCC molecular signature is various depending to the etiologies. Different molecular pathways including those regulating cell cycle, DNA methylation, chromosomal stability, apoptosis, immune system and non-coding RNAs expression can be affected [10]. The molecular heterogeneity of HCC makes the identification of effective drug very challenging.
1.1. Available Therapeutic Approaches for HCC
The stage of the disease guides the choice of the most appropriated treatment. For the localized forms of HCC (single nodule), the surgical resection is indicated for patients with HCC occurring in non-cirrhotic liver or in case of limited cirrhotic liver with conserved liver functions [11]. Liver transplantation is indicated for patients bearing one nodule <5 cm or up to three nodules each <3 cm (Milan criteria [12]). However, the limited availability of healthy liver makes this option rarely applicable. For the treatment of the intermediate disease, the employment of trans-arterial chemoembolization (TACE) and trans-arterial radio-embolization (TARE) are considered when two to seven nodules, the largest of which is <7 cm in size [13] are present. TACE and TARE can also represent a bridging therapy in patients waiting for liver transplantation [14]. In the advanced stages of the disease, it is possible to consider the use of targeted therapies and immune checkpoint inhibitors [13]. Systemic therapies can be also used in patients with disease recurrence after a curative intent. First line systemic therapies based on the use of oral multi-kinase inhibitor such as sorafenib are indicated in patients with contraindication in the use of immunotherapy [13]. If no contraindication to immunotherapy exists, first line drugs such as atezolizumab and bevacizumab can be used. These are humanized monoclonal antibodies directed against the programmed cell death protein 1 ligand (PD-L1) or the vascular endothelial growth factor (VEGF-A), respectively. In particular, Atezolizumab blocks PD-L1 favoring the anti HCC effects of T-lymphocytes while bevacizumab down-regulates HCC neo-angiogenesis. Finally, for the terminal stage of HCC no anticancer treatment can be proposed, but only palliative approaches. Unfortunately, about 20/30% of patients worldwide are diagnosed at this end stage of the disease.
1.2. HCC Diagnoses
1.2.1. Instrumental Diagnoses
The early detection of HCC is crucial, as the therapeutic options available are more effective for the first stages of the disease. Ultrasound Examination (UE) [13] represents the elective diagnostic tool for HCC screening. The sensitivity range of UE is of 51%-87% while specificity is of 80-100% [15]. UE allows determining the location, size, morphology and vascular invasion of HCC. The determination of the circulating level of Alpha-fetoprotein (AFP) can be associated to UE. AFP is a serum glycoprotein produced by the fetal yolk sac and fetal liver during gestation. Whereas its level can increase in the course of HCC, other pathological conditions such as chronic hepatitis, cirrhosis, pregnancy and some germ line tumors can induce its increase. Furthermore, approximately 40% of small HCC do not secrete AFP [16]. For these reasons, its use is considered not particularly useful in terms of cost-effectiveness. However, recent data indicate that the combination of UE and AFP could rise HCC detection to 63% compared to the 45% of UE alone [17].
When the liver is difficult to be analyzed by UE, the use of contrast-computed tomography (CT) is indicated. Sensitivity and specificity of CT are of 65% and 96%, respectively [18]. However, for lesions smaller than 2 cm, sensitivity goes down to 40%. Finally, magnetic resonance imaging (MRI) [19] can be considered for HCC diagnose, even if questions arise with regard to the cost of screening and equipment availability.
1.2.2. Biochemical Diagnoses
Whereas liver biopsy is not routinely undertaken for HCC diagnoses, in the case of lesions smaller than 1-2 cm, which cannot be accurately characterized by CT/MRI, biopsy is indicated [11]. Beside this aspect, tissue biopsy allows the histological classification of HCC thus helping in defining disease prognosis. Biopsy limitations are represented by sampling bias and tumor heterogeneity, which may result in a tissue sample not fully representative of the HCC histology [20]. Moreover, liver biopsy may be associated to side effects such as pain and bleeding [21]. Notably, so far HCC has no tissue biomarker-driven therapy in contrast to other tumors such as breast and lung cancers [22].
Recently, liquid biopsy has arisen as a potential novel approach for HCC diagnoses [20]. The possibility to detect circulating HCC markers has several advantages such as the possibility of perform multiple tests thus allowing the real-time monitoring of HCC evolution. Moreover, liquid biopsy has substantially no relevant side effects in comparison to liver biopsy. So far, however, the employment of liquid biopsy is not an integral part of the standard clinical practice. This is mainly due to concerns dealing with sensitivity, specificity, and cost-effectiveness. Whereas we believe that sensitivity/specificity issues can be overcome by the modern omics techniques, the problem relies on the identification of a robust and specific circulating HCC marker(s). In principle, the liquid biopsy has the potential to help in the identification of minimal residual disease, treatment selection and the monitoring of therapeutic responses. Unfortunately, so far, the only serum makers for HCC detection has been AFP, as above discussed. Novel HCC markers under testing are the fucosylated fraction of AFP (AFP-L3) and des-gamma carboxyprothrombin (DCP) [23]. The combined use of AFP-L3, AFP and DCP with patient gender and age resulted in an improvement in sensitivity for HCC detection but an increase in false-positive results.
Novel circulating biomarkers under investigation for HCC detection are represented by circulating tumor cells (CTC), cell-free DNA (CFD) and non-coding RNAs (ncRNAs). The detection of CTC was one of the first approach in the liquid biopsy. The concept is that CTC can migrate from the tumor of origin/metastasis to the vasculature: therefore, their circulating number is somewhat proportional to the tumor burden. While they may be not indicated in the detection of early HCC (reduced number of CTC), they may be suitable to monitor HCC recurrence and treatment response [24]. CFD are represented by a large pool of double-stranded DNA fragments associated with nucleosomes and circulating in the blood. Typically, CFD have been studied for their quantity, integrity and copy number alterations [25]; more recently, they have been studied with regard to the methylation pattern and mutational signature. In this regard, different epigenetic changes in CFD of patients with HCC has been defined [20], possibly contributing to improved diagnostic specificity. ncRNAs are the topic of this review and will be presented extensively in the following sections.
1.2.3. Considerations about HCC Diagnoses/Monitoring
As above discussed, many different approaches are available for HCC diagnosis/monitoring; however, basically all of them have limitations which may delay HCC identification and/or make the monitoring not as effective as we would like it to be. The liquid biopsy represents a minimally invasive technique with a relevant diagnostic potential for HCC but also for other pathological conditions. Unfortunately, for HCC diagnoses, a robust diagnostic/prognostic marker has not yet identified. In this regard, it is possible that rather than a single marker, multiple biomarkers together need to be analyzed for an optimal result. Moreover, it is possible that a multi-parametric approach, which includes biomarkers and different patient features, will be necessary for a more effective output compared to the biomarkers alone. In this contest, we believe that ncRNAs may substantially contribute to reach the goal of a specific and sensitive HCC diagnostic/monitoring approach.
2. Non-Coding RNAs
To date a significant number of different non-coding RNAs (ncRNAs) are recognized [26]. In addition to the well-known ribosomal RNAs (rRNAs) and transfer RNAs (tRNAs), other types have been identified in the last years [27]. These include micro interfering RNAs (miRNAs), long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), heterogeneous nuclear RNAs (hnRNAs), PIWI-interacting RNAs (piRNAs), small nuclear RNAs (snRNAs) and small nucleolar RNAs (snoRNAs). NcRNAs regulate human gene expression via the interaction with their target mRNA. In addition, they also interact each other resulting in a very complex system able to control human gene expression. So far, the most studied ncRNAs in relation to human pathologies and in particular to HCC are miRNAs, lncRNAs and circRNAs, all recognized as essential regulators of several biological processes in health and disease.
2.1. miRNAs
miRNAs, firstly recognized as potential regulators of human gene expression in 2001 [28], are short double-stranded non-coding RNAs. To date, more than 2000 human mature miRNAs have been annotated in the miRNA archive miRBase [29].
miRNAs originate in the cell nucleus (Figure 1) from a long precursor named primary miRNA (pri-miRNA) derived from coding/non coding RNAs [30].
Following the processing by the enzyme Drosha [31], the pri-miRNA is converted into the so called pre-miRNA which in turn is exported to the cytoplasm by Exportin 5 (Exp5). Here, the DICER enzyme [32] generates a double stranded RNA (mature miRNA) of approximately 22 nucleotides bearing 2 nt 3’ overhangs [33]. The antisense strand of the mature miRNA is then up-taken by the enzymatic complex RISC (RNA-induced silencing complexes) [32] allowing the recognition of the target mRNA via a complementary base-pairing approach. A perfect match of the first 2-8 nucleotides, from the 5’ end of the antisense strand with the target, is required [34] for efficient translation inhibition. When a full complementarity occurs among all nucleotides, the degradation of the target RNA takes place. The above mechanism of interaction, occurring at the 3’UTR of the target mRNA, is considered the classical-mechanism of action. However, miRNAs, which can interact with the 5’ UTR or with the coding region of an mRNA molecule, have been also described [35]. Notably, the biological effect of miRNAs is not limited the down regulation of gene expression as in some cases they can promote gene expression via direct/indirect mechanisms [36].
Together the above considerations witness the biological complexity of miRNAs, which is further amplified by the fact that as a single miRNA can target several different mRNAs and many miRNAs regulate the same mRNA [37]. This promiscuous modality of action is further amplified by the fact that miRNAs regulate and are regulated by lncRNAs/circRNAs (see also below). Moreover, it is now becoming evident that miRNA can circulate into the blood embedded into extracellular vesicles acting as hormones as below reassumed [38]. The role of miRNAs in the pathogenesis of HCC as well as their potential values of novel biomarkers/therapeutic targets is nowadays well known [39,40,41,42].
2.2. Long Non-Coding RNA
LncRNAs are single-stranded RNA molecule with a length set longer than 200 nucleotides [43]. The first lncRNAs described in eukaryotes dates back to the beginning of 1990s [44]. Nowadays, the NONCODE database reports 173112 lncRNA transcripts [45].
LncRNA synthesis (Figure 2) is comparable to that of mRNAs: indeed, they are transcribed by RNA polymerase II (Pol II) and are often spliced, capped and poly-adenylated.
Differently from mRNAs, lncRNAs do not contain a translated open reading frame. Moreover, they are generally shorter and are expressed at lower levels compared to mRNAs. Finally, they usually contain fewer but longer exons then mRNAs, are often retained into the nucleus and tend to have more tissue-specific expression [46]. They can be transcribed in both the sense and antisense orientations from diverse genomic regions such as introns, exons, intergenic regions, pseudogenes, telomeres, centromeres, gene promoters and 3’ UTR [47].
The linear nucleotides sequence of lncRNAs determines their 3D structure, which eventually is responsible for their biological effects [48]. Based on the 3D structure, lncRNAs can bind and recruit transcription activators/repressors to the promoters of their target genes thus modulating transcription. They can bind miRNAs to complementary miRNA sequences (sponge effect), thus inhibiting miRNAs biological effects. Moreover, lncRNAs can be scaffolds to promote the formation of protein complexes thus promoting/inhibiting protein functions. All together these features witnesses lncRNAs complex and multiple biological functions also making not surprising that their altered expression and/or mutation are implicated in numerous human diseases [49]. Nowadays, the involvement of lncRNAs in HCC with regard to the pathogenesis of the disease and the possibility to use them as biomarker/target for novel molecular therapeutic approaches is well documented [40,41,42].
2.3. circRNAs
circRNAs have a circular structure constituted by a single stranded RNA covalently closed at the extremities [50]. The circular shape confers to these molecules increased stability against degradation by exonucleases compared to linear RNAs. Initially described in the early nineties of the last century, we now know that the expression of these molecules is rather wide, as over 10,000 circRNAs have been described in many living organisms including human [51].
RNA polymerase II (Pol II) [52] drives the transcription of circRNAs from precursor mRNA (pre-mRNA) (Figure 3).
CircRNAs can derive from exons, introns, exon–intron junctions or intergenic regions of the genome [53]. Interestingly, the same gene can give origin to multiple circRNAs by alternative splicing mechanism [54] and often cirRNA expression is unrelated to the expression of the host gene [55]. CircRNAs are generated in the cell nucleus by a mechanism called back-splicing that most likely requires canonical spliceosomal machinery [56]. Subsequently they are exported to the cell cytoplasm. CircRNAs length is variable ranging from 30-40 nucleotides to longer sequences [57]. While most often the expression level is low [58], circRNAs abundantly expressed have been also described [59]. To date, different biological functions have been defined for circRNAs [26]. In particular, they can: 1) bind miRNAs via complementary regions (sponge effect), thus impairing miRNA effects, 2) interact with specific mRNAs regulating their stability and/or translation, 3) undergo translation to generate small peptides, 4) interact with RNA-binding proteins acting as decoys or sponges for proteins, 5) interact with gene promoters, thus modulating gene expression. CircRNAs involvement in HCC pathogenesis and the possible role as novel biomarkers/therapeutic targets is commonly accepted [40,42].
3. Extracellular Vesicles
The observation that ncRNAs can circulate in the blood embedded into extracellular vesicles (EVs) thus acting as hormones, has busted the research in the biology of EVs. These are a heterogeneous group of lipid bilayer particles, synthesized and secreted by innumerable cell types into the extracellular environment [38]. Thus, they are able to circulate in many biological fluids including blood [60]. Based on their size, they can be classified as exosomes (30–150 nm diameter), ectosomes (50–10000 nm diameter), apoptotic bodies (1000–5000 nm diameter) and other vesicles [61]. The precise molecular mechanisms of EVs generation remains in part unclear; however, it is accepted the idea that they form either through inward-budding vesicles within the endocytic system, or through outward-budding vesicles at the plasma membrane (Figure 4).
The first demonstration of the existence of EVs dates back to 1987 [62], but the biological meaning become clearer in 1996 [63] and especially in 2007 when it became evident that EVs can carry cytoplasmic molecules such as mRNA and miRNAs [64]. Afterwards it was observed that both stromal and cancer cells can produce EVs modulating tumor progression through molecular exchange [65]. More recently, it has been shown that ncRNAs carried by EVs regulate HCC treatment response and disease progression [66]. Nowadays it is accepted that circulating exosome-derived ncRNAs like miR-21 [67], lncRNA ASMTL-AS1 [68], lncRNA TUC339 [69] and circ_0051443 [70] are strongly related to HCC disease.
4. ncRNAs Employment as Biomarkers for HCC
4.1. miRNA
miRNAs were the first class on ncRNA to be studied as potential diagnostic/prognostic markers for HCC. Among the many works published in the field, here we focus on a selection of representative papers (Table 1).
4.1.1. miR-221 and miR-18a
miR-221 is encoded on the human X chromosome in humans and regulates target genes essential for the growth of several types of cancers [71]. miR-18a is one of the most conserved and multifunctional miRNAs in the polycistronic miR-17-92 cluster and is frequently overexpressed in malignancies, such as non-small-cell lung cancer, cervical cancer and gastric cancer. Among the various recognized targets of miR-221 is the Wnt/β-catenin pathway [72], while miR-18a has been shown to regulate the expression of KLF transcription factor 4 (KLF4) and p21, promoting HCC cell proliferation and migration [73]. Both KLF4 and p21 are inhibitors of the cell cycle.
Yun et al. [74] evaluated the dysregulation of miR-221 and miR-18a in 50 formalin fixed paraffin embedded tissues obtained from surgically resected HCC. Both miR-221 and miR-18a were found to be more expressed in HCC tissues compared to adjacent noncancerous tissues by real-time qRT-PCR. In addition, the survival rate of patients with low miR-221 expression was significantly higher than that of patients with high expression patients (p=0.020). Although miR-18a expression was not correlated with survival or recurrence, the authors found that miR-18a levels were significantly associated with tumor stage and size.
Chen et al. have also suggested the potential role of miR-221 as a prognostic biomarker for HCC [75]. Significant upregulation of miR-221 expression was detected in 135 primary HCC tissues compared to non-tumor tissues by real-time qRT-PCR. Notably, in patients with multiple tumor nodules and microvascular invasion, miR-221 was upregulated compared to patients with a single tumor nodule and no microvascular invasion. In addition, miR-221 levels showed a close association with tumor stage. Kaplan-Meier survival analysis revealed that HCC patients with low miR-221 expression displayed increased disease-free survival (DFS) and overall survival (OS) than HCC patients with high miR-221 expression (long-rank test: p<0.001). Furthermore, the authors showed that tumor size, AFP protein level, microvascular invasion and miR-221 were significantly correlated with DFS and OS. Finally, a multivariate Cox analysis suggested that miR-221 was an independent indicator of poor prognosis in HCC patients.
Another evidence of miR-221 involvement in HCC derives from the work of Yousurf et al. [76]. The group identified a tree-miRNA panel, including miR-221, with improved diagnostic efficiency in differentiating HCC patients from healthy controls. The expression profile of miR-221 in the serum of 33 HCC patients and 33 controls was studied by real-time qRT-PCR and was found to be downregulated in HCC. In contrast, miR-221 expression levels in tumor specimens showed significantly higher expression compared to the peri-tumor counterparts. It would be interesting to understand the contrasting levels of miR221 in HCC tissue and in serum: unfortunately, the authors did not comment on this peculiar aspect. The ROC analysis was used to validate the discriminatory potential of significantly altered miRNAs. Among nine differentially expressed serum miRNA in HCC patients, miR-221 showed the second-best diagnostic efficacy with a rather high Area Under the Curve (AUC) value (AUC 0.786, 95% CI = 0.666-0.906, cut off <1.626; 228, sensitivity 77.14%, specificity 80.77%, p < 0.0001). The authors also observed that the diagnostic efficacy of miR-221 alone was superior to that of AFP. Importantly, it was pointed out that the best diagnostic efficacy occurred when miR-221 expression was associated to those of miR-221 and Let-7a. This observation reinforces the concept that probably the evaluation of the expression levels of a single miRNA may not be optimal for precise HCC diagnose.
Finally, a recent meta-analysis [77] found that the overexpression of miR-221 was significantly associated with poor OS (HR = 1.91, 95% CI: 1.53–2.38, p< 0.01) and DFS (HR = 2.02, 95% CI: 1.58–2.57, p<0.01), comparing the results of seven articles with 416 patients and five studies with 391 patients, respectively. Subgroup analysis for OS showed that high miR-221 expression in HCC tissue was closely associated with poor prognosis regardless of ethnic groups (Asian Vs no-Asian patients) and of the specimen used for detection (formalin-fixed, paraffin-embedded tissue or frozen tissue).
4.1.2. miR-487a
Recent studies have demonstrated that miR-487a plays a key role in cancer progression, including gastric and breast cancer. In addition, it targets the mRNA of TIA1 [78], a protein that binds the 3’ untranslated region (3’ UTR) of mRNA and the mRNA of membrane-associated guanylate kinase inverted 2 (MAGI2) [79], a protein that seems to act as a scaffold molecule at synaptic junctions. Both TIA1 and MAGI2 are involved in the TGFβ-induced epithelial-mesenchymal transition.
Chang R-M. et al. [80] studied a total of 198 cases which were randomly divided into two groups: a training cohort (132 patients) and a validation cohort (66 patients). In both groups, the authors demonstrated that miR-487a expression in HCC tissues was higher than that in adjacent non-tumorous liver tissues in 85.6% (113/132) and 81.8% (54/66) of cases in the training cohort and validation cohort, respectively. In addition, to determine whether miR-487a level correlated with the prognosis of HCC patients, cases from the training cohort and validation cohorts were divided into two additional subgroups consisting of patients whose miR-487a expression in HCC tissues was 2-fold higher than in their adjacent non-tumorous liver tissues (high expression group) and the remaining samples (low expression group). In the training cohort, miR-487a levels were directly associated with tumor size, nodule number, capsule formation, microvascular invasion, and tumor-node-metastasis classification. In contrast, in the validation cohort, miR-487a expression was only associated with tumor size, nodule number, and microvascular invasion. To understand whether miR-487a could be a prognostic factor, the OS and DFS were analyzed in both groups. The authors observed that OS and DFS were worst in the high miR-487a expression group compared to the low miR-487a expression group in both cohorts. Using univariate analysis, it was demonstrated that liver cirrhosis, nodule number, microvascular invasion, and miR-487a expression were independent risk factors for OS and DSF in the training cohorts. In the validation cohort, nodule number, microvascular invasion, tumor-node-metastasis classification and miR-487a levels were independent risk factors for OS, whereas only microvascular invasion and miR-487a expression were independent risk factors for DFS. All these evidences suggests that the upregulation of miR-487a is an independent risk factor and predictor of poor survival for HCC patients.
4.1.3. miR-33a
miR-33a is a member of the miR-33 family, highly conserved in human species. It is an intronic miRNA within the genes of sterol regulatory element-binding proteins (SREBPs) [81]; it primarily regulates the metabolism of cholesterol [82] and glucose [83]. The role of miR-33a in cancer has been described in literature, with numerous studies indicating that it exerts tumor-suppressive effects in certain cancers, including pancreatic and lung cancers [84]. Another study has demonstrated its implication in the development of chemoresistance in osteosarcoma [85]. Furthermore, miR-33a has been demonstrated to significantly increase in liver tissue in a fibrosis progression-dependent manner [86].
In 2018, Ru-Ting Xie et al. [87] analyzed 149 HCC biopsies, 36 of which were paired with para-carcinoma tissues. The authors found that patients with lower miR-33a expression had significantly poorer survival compared to those with higher miR-33a levels. miR-33a expression was significantly correlated with the presence of multiple foci; moreover, multivariate Cox analysis suggested that the combination of low miR-33a levels and multiple foci was associated with a significant decrease in OS and progression free survival (PFS). Conversely, a more recent paper reported that reduced miR-33a level was associated to increased survival probability [88]. Given the presence of conflicting data in the existing literature regarding its role and prognostic value, further investigation is required to unravel the significance of miR-33a.
4.1.4. miR-105-1
miR-105-1 represents a member of the miR-105 family, which is located within the X chromosome [89]. miR-105-1 can affect Annexin A9 (ANXA9) expression, enhancing the chemo-resistance and attenuating the cell apoptosis induced by cisplatin in cisplatin-sensitive ovarian cancer cells [90].
miR-105-1 has been previously identified as key regulator of circulation, pre-metastatic process and metastatic progression in early breast cancer [91]. Additionally, low levels of miR-105-1 correlated with poor clinical outcomes and aggressive progression in glioma [92]. In 2014, the expression of miR-105-1 was observed to be markedly downregulated in both HCC cell lines and HCC tissues [93]. The authors also showed that miR-105-1 overexpression suppressed HCC cell proliferation in vitro and in vivo by targeting FosfoInositide-3-Kinasi (PI3K)/AKT serine/threonine kinase 1 signaling pathway, thus suggesting a role as a tumor suppressor.
In 2017, Yu-Shui Ma et al.[94] confirmed the above observation, demonstrating, through in silico and real-time qRT-PCR analysis, the reduced miR-105-1 expression in HCC liver biopsy compared to non-tumor liver tissues. Furthermore, the authors showed that the reduced expression was associated with HCC progression and poor prognosis. Performing overexpression and silencing assays, it was demonstrated that nuclear receptor coactivator 1 (NCOA1) was a direct target of miR-105-1. NCOA1 is a transcriptional coregulatory protein recruited to DNA promoter sites where it acylates histones, making downstream DNA more accessible to transcription. The authors observed that NCOA1 was upregulated in HCC tissue compared to normal liver and that its upregulation was related to a poorer OS rate. Thus, the reduced expression of miR-105-1 in HCC determines the increased level of NCOA1 with a consequent pathological transcriptional activation.
4.1.5. miR-138-5p
The human miR-138 family consists of miR-138-3p and miR-138-5p, which are located on chromosomes 3p21.32 and 16q13, respectively. Recent reports have demonstrated that miR-138-5p exerts a repressive effect on the proliferation of endothelial progenitor cells by inhibiting hypoxia-inducible factor-1α (HIF-1α) and vascular endothelial growth factor A (VEGFA), both master regulators of angiogenesis and tumor vascular mimicry [95]. In 2022, Liu et al. [96] demonstrated that not only miR-138-5p exhibited lower expression in HCC tissues compared to para-tumor tissues, but also that low levels of miR-138-5p were significantly associated with high vascular mimicry density and high levels of HIF-1α and VEGFA. Furthermore, the authors discovered a negative correlation between miR-138-5 expression and HIF-1α mRNA levels. The Kaplan–Meier analysis of survival performed on a dataset from “The Cancer Genome Atlas” (TCGA), revealed that low miR138-5p expression and high HIF-1α/VEGFA mRNA levels were associated with a poor prognosis in patients with HCC.
Furthermore, in 2022, the Jiang’s research group [97] demonstrated that the non-coding circular RNA circ-TLK1 functions as an oncogene in HCC progression, partly through the inhibition (sponge effect) of miR-138-5p. Notably, the expression of miR-138-5p was increased in the HCC cell line HepG2 following the knockdown of circ-TLK1; in contrast, the overexpression of circ-TLK1 resulted in a reduction of miR-138-5p levels in the HCC cell line HuH7. These observations contributed to demonstrate the functional relation between circ-TLK1 and miR-138-5p. A comparative analysis of circ-TLK1 expression profile in 87 specimens from HCC patients revealed a significant upregulation of circ-TLK1 in HCC tissues compared to adjacent non-tumor tissues. Moreover, high intra-tumor circ-TLK1 expression appeared to be closely correlated with larger tumor size (P = 0.030), advanced TNM stage (P = 0.018), and vascular invasion (P = 0.005). Additionally, lower levels of miR-138-5p were observed in HCC tissues compared to normal tissue. Notably, miR-138-5p expression was negatively correlated with that of circ-TLK1 in HCC. This is the first demonstration that circ-TLK1 is highly expressed in HCC specimens and that it acts as an oncogene in HCC progression, partly through competitive binding to miR-138-5p. This, in turn, relieves the inhibitory effects of miR-138-5p on its target SRY-related HMG-box (SOX4). SOX4 belongs to a family of transcription factors whose overexpression favors early HCC recurrence. [98]
In 2020, Xiao Xi J. et al.[99] investigated the impact of miR-138/SOX4 axis on Anillin, an actin-binding protein that plays a pivotal role in regulating cytokinesis during the cell cycle and it is thus involved in tumorigenesis progression. In vitro assays demonstrated that both SOX4 and Anillin mRNA expression were significantly reduced by miR-138 upregulation in HCC cell lines. Furthermore, the re-introduction of SOX4 into HCC cells resulted in the increase of Anillin levels, even in the presence of miR-138 upregulation. The authors investigated the expression profile of Anillin in cancer through an analysis of the TCGA database and found that Anillin is commonly highly expressed in the majority of solid human tumors, including HCC. In order to gain further insight into the expression profile of Anillin, the researchers conducted a detailed evaluation of its expression in 87 HCC tissues, compared to their adjacent non-tumor tissues. In the tumor tissues, approximately 67.8% (59/87) of samples exhibited high expression of Anillin, whereas in non-tumor tissues, high expression of Anillin was observed only in a small portion of cases (25.3%; 22/87). The remaining cases displayed markedly low Anillin expression in comparison to the tumor tissue specimens. The DFS and the OS in a dataset of 361 HCC patients revealed a significant correlation between higher Anillin levels and poor prognosis, as well as high mortality. Furthermore, DFS and OS rates were found to be markedly dismal in patients with high Anillin expression.
4.1.6. miR-21
miR-21 is one of the most extensively researched oncogenic microRNA, with a multitude of studies conducted on its role in various cancers. It is located within the Vacuole Membrane Protein 1 (VMP1) locus on chromosome 17 [100].
miR-21 has been identified as a particularly significant factor in HCC, being upregulated in tumor tissue and its overexpression has been associated with poor OS in HCC patients [101]. In particular, it has been shown that paired tissue (HCC and normal liver tissue), serum and serum exosomes sequencing indicated the positive correlation of miR-21 between serum exosomes and HCC tissue, suggesting that miR-21 is exported from HCC tissue to circulation via exosomes.
In 2023, Stechele et al. [102], investigated the potential value of circulating miR-21 as therapy response marker for HCC patients treated with high-dose rate brachytherapy. The researchers demonstrated that an increase in the plasma of miR-21 level two days after ablative therapy was associated with a poor response to therapy and a shorter time to systemic progression. These data are consistent with the results of previous studies demonstrating a correlation between high miR-21 expression in HCC tissue and poor prognosis [103]. In particular, miR-21 levels were measured in 166 samples of surgically resected HCC nodules. The output of the investigation indicated that high miR-21 expression represents an independent prognostic factor for both short OS (hazard ratio 2.36) and DFS (hazard ratio 2.02).
Another study [104] has indicated that the circulating miR-21 and miR-130b may serve as potential tumor biomarkers for the diagnosis of liver cancer. The combined detection of serum miR-130b and miR-21 has been demonstrated to be an effective approach for the diagnosis of HCC showing a sensitivity of 92.16% and an accuracy rate of 77.51%. The evidence presented was obtained through a comparative analysis of results obtained from serum samples and tumor tissues from 46 patients with HCC and 55 healthy volunteers. The level of miR-21 and miR-130b were found to be upregulated in both serum level and tumor tissues when compared to the corresponding control group. Spearman’s rank analysis revealed a highly significant positive correlation between serum and tumor tissue levels. Comparing the serum expression profile of miR-21 e miR-130b in HCC patients before and after surgical treatment, the authors observed a marked decrease in the expression level of both miRNAs two weeks after surgery, compared to the levels observed before surgical treatment. Kaplan–Meier analysis indicated a significant difference in OS and PFS between the high serum miR-130b/Mir-21 expression group and the low expression group. Moreover, the authors identified a correlation between the circulating levels of both miR-21 and miR-130b and tumor capsular infiltration, HCC stage and distant metastasis.
4.1.7. miR-122
In humans, miR-122 is encoded at a single genomic locus on chromosome 18 and its transcription is regulated by hepatocyte nuclear factor 4 alpha (HNF4α) [105]. miR-122 is highly-expressed in the liver and it has been demonstrated to regulate a range of physiological and pathological conditions, including lipid metabolism, the response to drug administration/viral infection, alcoholic hepatic injury and the formation of fibrosis [106]. Previous research has shown that miR-122 expression is downregulated in HCC tissues compared to the normal liver tissues and that it functions as a tumor suppressor during the process of hepato-carcinogenesis [107].
In 2020, a meta-analysis was conducted to evaluate the diagnostic performance of serum miR-122 in HCC patients compared to a control group [108]. A comparison of data from 13 studies, which included 920 HCC patients and 1217 healthy controls, revealed that serum miR-122 exhibited a moderate diagnostic accuracy for the discrimination of HCC (AUC=0.82). Further the analysis indicated that serum miR-122 level was an acceptable discriminator of HCC patients from healthy individuals or those with HBV or HCV (AUC=0.87) infection. However, the discrimination between cirrhotic and HCC patients was less effective (AUC=0.74). Taken together, the data indicated that the measurement of circulating miR-122 potentially offers moderate diagnostic efficacy for the discrimination of HCC patients from healthy individuals or from those affected by HBV or HCV.
In 2022, Fang Y. et al.[109] conducted a case-control study, which included 100 healthy controls and 100 patients with early-stage HCC. The aim was to evaluate the diagnostic efficacy of a profile of differentially expressed miRNAs, including miR-122. The results demonstrated a markedly increased expression level of circulating miR-16 and miR-122 in the serum of early-stage HCC patients compared to the control group. More in details, the AUC value of miR-16 was 0.798, with a sensitivity of 91% and a specificity of 58% (95% CI 0.738–0.858, p < 0.001), whereas the AUC value of miR-122 was 0.759, with a sensitivity of 83% and a specificity of 64% (95% CI 0.690–0.827, p < 0.001). The author also evaluated the diagnostic power of the combination of the two miRNAs in comparison to AFP. The AUC value for the combination of miR-16 and miR-122 increased up to 0.803, although with a lower sensitivity (58%) but a higher specificity (84%) (95% CI 0.745–0.861, p < 0.001). The AUC value of AFP was less than that observed for the miRNAs combination (0.716), with a sensitivity of 64% and a specificity of 84% (95% CI 0.641–0.791, p < 0.001). The combination of AFP with either miR-16 or miR-122 not only increased the AUC value (to 0.859 and 0.825, respectively) but also the specificity (to 87% and 79%, respectively). Finally, the combination of AFP with both miRNAs, yielded the highest AUC value (0.862), accompanied by a sensitivity of 76% and a specificity of 80% (95% CI 0.813–0.911, p < 0.001). Thus, it can be concluded that these circulating miRNAs have significant value in the early screening and diagnosis of HCC when combined with AFP.
In 2021, an Italian clinical study [110] compared the levels of circulating miR-21 and miR-122 in healthy subjects and HCC patients. The aim was to evaluate the role of these miRNAs as predictors of PFS in a group of Caucasian HCC patients treated by drug-eluting beads trans-arterial chemoembolization (DEB-TACE). Serum miR-122 levels demonstrated a progressive increase in HCC patients compared to controls (p=0.02). The increase was associated with the underlying etiology, in particular with virus-related liver disease. ROC curves were used to identify optimal prognostic cut-offs for serum miR-21 and miR-221 levels, ensuring maximal sensitivity and specificity. In contrast to miR-21, miR-122 levels immediately before DEB-TACE treatment proved to be a predictive factor for PFS. Indeed, patients with miR-122 levels below the defined cut-off exhibited a significantly longer median PFS than that observed in the comparison group (5.6 months [1.4–9.7] vs 5 months [1.8–3.2]). Notably, in a mouse model of HCC a novel interconnection between miR-122 and hypoxia-induced pathways has been identified HCC [111]. In this regard, a correlation analysis was conducted between miR-21, miR-122, and HIF-1α in HCC patients undergoing DEB-TACE treatment. As a result of this investigation, a positive correlation between miR-21 and HIF-1α in HCC patients was observed both pre- and post- surgical treatment. In contrast, no significant associations with miR-122 were identified.
4.1.8. miR 139-5p
miR-139-5p is located within the sequence of the phosphodiesterase 2A (PDE2A) gene [112]. Among the various recognized targets for this miRNA there is Rho-associated coiled-coil kinase2 (ROCK2) [113]. ROCK2 promotes cell proliferation and migration in many tissues including the liver, via the interaction with multiple targets such as of cyclin D1 [114] and matrix metalloproteinase 2 (MMP-2) [115], while reducing the levels of p27Kip1[114]. In HCC miR-139-5p down regulation is related to the methylation of its promoter, a phenomenon occurring in hepato-carcinogenesis. Since a decade, it is known that miR-139-5p is down-regulated in HCC and that its low expression relates to poor prognosis [116]. Additionally, reduced expression level relates to vein invasion and HCC stage. More recently [117] it has been observed that high tissue expression of miR-139-5p (Hazard ratio: 0.72, P<0.001), is associated with good prognosis in HCC patient. We observed [118] in vitro and in vivo models of HCC that the reactivation of miR-139-5p expression via the use of demethylating drugs, can reduce HCC cells proliferation and migration, thus contrasting HCC growth. Taken together the above observations strengthen the rational to consider miR-139-5p as a promising HCC diagnostic/prognostic marker and possible also a therapeutic target [42].
4.2. LncRNAs
LncRNAs have been associated to human pathologies in general and to HCC in particular. For this type of molecules, less information about their biological behavior are known compared to miRNA. Despite this, nowadays they are actively investigated as potential diagnostic molecules in HCC. Among the many works published in the field so far, here we focussed on a selection we believe can be representative (Table 2).
4.2.1. LncRNAs HULC and MALATI
Initially identified as a highly upregulated transcript in the blood/cancer tissue of HCC patients [119], HULC is a 482 lncRNA encoded by a gene located on locus 6p24.3. Its aberrant expression has been associated with HCC development and progression [119]. HULC acts as a molecular sponge for miR-2001-3p, miR-186, and miR-107, thereby increasing the levels of multiple oncogenes, including ZEB1, HMGA2, and E2F1, respectively [120]. A recent meta-analysis published by Lumkul et al. [121], demonstrated that the serum levels of HULC, HOTARI and UCA1 are increased in HCC patients. Moreover, they exhibited excellent discriminatory capabilities between HCC patients and patients with other forms of liver disease (AUC=86%, 95% CI, 83–89%). Notably, the detection of serum HULC showed the best performance, compared to HOTAIR or UCA1.
In 2018, Wang et al. [122] evaluated the role of several genetic polymorphisms of HULC in affecting cancer susceptibility and clinical outcomes. The researchers identified a specific polymorphism, rs1041279 that was associated with a 1.41-fold increased HCC risk in 521 patients when compared to 817 sex- and age-matched controls. However, no significant association was observed between HULC polymorphisms and OS. In the same years, Sonohara et al. [123] investigated the prognostic value of lncRNA HULC and MALAT1 (see also below) following tumor surgical resection. In comparison to the control group, the expression levels of HULC were found to be more elevated in HCC samples compared to normal tissue; MALAT1 expression did not differ between the control and cancerous tissues. A positive correlation was observed between the expression levels of MALAT1 and both capsular formation and tumor size. Both MALAT1 and HULC expression were found to be negatively correlated with the level of AFP. Stratifying the patients by HULC levels, those with increased HULC expression exhibited a tendency towards improved relapse-free survival, although the difference was not statistically significant. Preliminary HULC analysis with Gene Expression Omnibus (GEO) and in silico analysis showed that increased HULC expression correlates with a better prognosis in HCC [124]. All together, these findings suggest that the increased expression levels of HULC in HCC tissue may serve as a promising prognostic biomarker for HCC.
MALAT1, initially described in non-small cell lung cancer in 2003 [125], is an 8.5kb lncRNA located on 11q13 locus. It acts as a proto-oncogene involved in the modulation of several molecular signaling pathways related to cell proliferation, death, migration, invasion, immunity, angiogenesis and tumorigenicity. A recent case-control study published by Golam et.al in 2024 [126], evaluated the diagnostic potential of MALAT1 in HCC in comparison to the routinely used diagnostic biomarkers. The MALAT1 blood levels were found to be significantly upregulated in both the HCV patient cohort and the HCC/HCV patient cohort in comparison to the healthy control group. Additionally, the MALAT1 expression level was increased in the HCV group compared to the HCC/HCV group. This observation may indicate that MALATI is more specific for HCV diagnoses rather than for HCC detection. A ROC analysis was performed to assess the diagnostic efficacy and identify the optimal cut-off value for MALAT1 and the routinely employed biomarker AFP. Unfortunately, MALAT1 exhibited lower accuracy and specificity than AFP.
In 2023, Liao et al. [127] published a study based on multi-omics analysis and real-time qRT-PCR validation to investigate the prognostic value of MALAT1. 368 patients with HCC were included in the analysis, based on data from The Cancer Genome Atlas (TCGA) liver hepatocellular carcinoma (TCGA-LIHC) database. The authors identified five protein-coding genes (AGO2, HNRNPC, EZH2, SFPQ, and SRSF1) associated with worse survival when highly expressed. In addition, the authors showed that MALATI is overexpressed in HCC tissue compared to non-tumor tissue. It is thus possible that MALATI contributes to HCC by interacting with AGO2, HNRNPC, EZH2, SFPQ, and SRSF1. In Kaplan-Meier analysis, patients with increased MALAT1 expression showed shorter PFS (p = 0.033) and OS (p = 0.023) compared to patients with low expression. MALAT1 expression not only positively correlated with more than 20 mutations in genes related to HCC progression but also with epigenetic modification. In particular, the authors found that MALAT1-associated methylation was related to poor survival of HCC patients. This suggests that MALAT1 may be involved in the progression and prognosis of HCC through its DNA methylation effect.
4.2.2. LncRNAs Linc00152
In 2015, Linc00152, an 828 bp lncRNA located at chromosome 2p11.2, was first identified as a potential marker for HCC diagnosis [128]. In this study conducted by Li et al., it was observed that the levels of Linc00152 and HULC were significantly upregulated in plasma samples obtained from HCC patients who had undergone hepatic resection compared to the plasma samples obtained from healthy controls. Furthermore, the levels of Linc00152 were found to be also significantly correlated with differentiation grade, tumor size and HCC stage. The ROC curve analysis of the two lncRNAs demonstrated the substantial diagnostic value of HULC and Linc00152 for HCC diagnosis, with AUC values of 0.78 and 0.85, respectively. The combination of HULC and Linc00152 resulted in an increased area under ROC curve value up to 0.87, thereby demonstrating the highest ability for discrimination between HCC patients and controls. Subsequently, in vitro studies were conducted to further elucidate the effects of Linc00152, which demonstrated that this lncRNA plays a pivotal role in regulating cell proliferation, clonogenicity, and migration [129].
In 2020, Wang B. and colleagues [130] investigated the potential diagnostic role of lncRNA NRADI and Linc00152 in HCC. The study cohort comprised 63 patients with primary HCC who had undergone surgical resection. The authors observed that NRAD1 was expressed in 47.6% (30 of 63) of HCC patients, whereas Linc00152 was expressed in 71.4% (45 of 63) of HCC patients. Furthermore, the levels of the two lncRNA were significantly higher in HCC samples than in non-tumor tissues. The Overexpression of NRAD1 and LincC00152 was found to be associated with a reduction in both OS and PFS rates. The univariable analysis demonstrated that microvascular invasion, tumor size and high expression levels of NRAD1 and Linc00152 were significantly associated with the prognosis of patients with HCC. No data were provided regarding the plasma or serum levels of these two lncRNA. Taken together these data suggest that the overexpression of lncRNA NRAD1 and LINC00152 represents an independent risk factor associated with the prognosis of HCC patients.
In 2023, Shehab-Eldeen et al. [131] demonstrated that the serum expression levels of Linc00152 and UCA1 were significantly higher in patients with HCC than in patients with liver cirrhosis or healthy subjects. The ROC curve analysis indicated that Linc00152 and UCA1 exhibited high sensitivity (81.7% and 83.3% respectively) and specificity (63.3% and 83.3%, respectively) for HCC, thereby establishing their efficacy as diagnostic biomarkers. Moreover, the combination of the two lncRNAs with AFP resulted in a significant increase in the accuracy of HCC diagnosis, with an AUC area of 0.99, a sensitivity of 100% and a specificity of 95%. These findings collectively indicate the potential utility of serum Linc00152 and UCA1 as non-invasive biomarkers for HCC.
4.2.3. lncRNA HOTAIR
HOTAIR (homeobox transcript antisense intergenic RNA) is located in the Homeobox C gene cluster a spliced and poly-adenylated RNA with 2,158 nucleotides and 6 exons. It originates from the transcription of antisense strand of HoxC gene, situated on chromosome 12q13.13 [132]. HOTAIR has been demonstrated to exhibit excellent discriminatory capabilities between patients with HCC and patients with other forms of liver disease. Additionally, it is involved in tumorigenesis, metastasis, and drug resistance in various cancer types, including HCC. Ishibashi et al. [133] published one of the earliest studies that investigated the clinical significance of HOTAIR expression in HCC in 2013. By staging patients according to their HOTAIR expression levels in HCC tissue, the authors demonstrated that patients with high HOTAIR expression exhibited a markedly poorer prognosis, reduced OS and a significantly larger tumor size than those with low HOTAIR expression. In a study published in 2021, Han and colleagues [134] reached similar conclusions regarding the role of HOTAIR in advanced HCC and its correlation with the use of Sunitinib as a chemotherapy agent. 60 patients with a diagnosis of advanced HCC who had not received any form of treatment before Sunitinib treatment, were enrolled in the study. The authors demonstrated that the expression of HOTAIR was significantly higher in tumor tissues in comparison to that observed in adjacent non-tumor tissues. Furthermore, HOTAIR levels in peripheral blood of HCC patients were higher than those observed in healthy subjects. Moreover, a positive correlation was identified between HOTAIR levels in tumor tissue and in peripheral blood of HCC patients. From a prognostic perspective, patients with low levels of HOTAIR in tumor tissues displayed significantly prolonged survival (13.4 vs. 9.5 months) and PFS (8.4 vs. 6.2 months) compared to those with high expression levels. Similarly, patients with low HOTAIR levels in peripheral blood demonstrated significantly higher OS (12.8 vs. 9.1 months) and PFS (8.9 vs. 6.4 months) compared to those with high expression. Patients with low expression in both tumor tissue and peripheral blood exhibited the most promising OS and PFS outcomes, with median OS and PFS times of 14.3 and 10.6 months, respectively, compared to the remaining patient cohort. The results of COX regression analysis indicated that the levels of HOTAIR in tumor tissue and peripheral blood were independent predictive factors of OS and PFS in patients with advanced HCC who were treated with Sunitinib.
In 2022, a further study conducted by Lou et al. [135] provided additional evidence to support the potential role of serum HOTAIR as marker for the early diagnosis and for the prognosis in HCC patients. A total of 61 patients diagnosed with HCC and a control group comprising both patients with liver cirrhosis and healthy subjects were included. Serum levels of HOTAIR, as well as BRM and ICR, were all found to be significantly increased in patients with tumors in comparison to both patients with liver cirrhosis and healthy controls. Serum HOTAIR levels were found to be closely associated with HCC stage, metastasis, vascular invasion and tumor size. A notable reduction in the serum levels of HOTAIR and ICR was detected between the assessments conducted during the preoperative visit and one week after tumor resection. The ROC curve analysis demonstrated that the three lncRNAs alone showed higher accuracy for identifying HCC compared to AFP. Notably, HOTAIR showed the highest AUC (0.991 with 96.7% of sensitivity and 95% of specificity) to distinguish HCC patients from healthy controls. The COX regression analysis demonstrated that the serum levels of HOTAIR, BRM and ICR were significantly correlated with HCC prognosis. In conclusion, the data collectively indicates that the serum levels of these three lncRNAs could serve as a panel of markers for early HCC diagnosis and possible for monitoring therapeutic effects.
A recent study published in 2024, further investigated the role of HOTARI as a potential biomarker for HCC. The meta-analysis conducted by Wen et al. [136] included eight studies comprising 399 subjects. The statistical analysis revealed a significant correlation between high HOTAIR levels and advanced HCC stage (OR=1.45; 95% CI [1.02, 2.05]; p=0.038), distant metastasis (OR=2.51; 95% CI [1.02, 6.14]; p=0.044), and poor tumor differentiation (OR=1.59; 95% CI [1.21, 2.10]; p=0.001). No correlations were found between HOTARI expression levels and age, gender, tumor number, tumor size, lymph node metastasis or AFP level. Among the eight studies, only three identified a negative correlation between OS and HOTARI expression levels. Patients have been classified into two distinct groups based on their expression levels of HOTARI. The findings revealed that patients in the high-level HOTAIR group exhibited a poor OS rate (HR=1.69; 95% CI [1.14, 2.51]; p=0.009), in comparison to the low-level HOTAIR group. Furthermore, a significant negative association was observed between HOTAIR expression and RFS (HR = 1.89; 95% CI [1.37, 2.59]; p=0.0001). In conclusion, the data suggest that HOTAIR may serve as a promising biomarker for evaluating HCC prognosis.
4.3. CircRNAs
CircRNAs are the last class of ncRNA described in this review to be associated to HCC and its diagnosis. Like lncRNAs, also for this class of molecules we still need to acquire more knowledge about their biology. Despite this, their potential as HCC biomarker is under active investigation. Among the many works published in the field so far, in the present review, we focus on some representative examples (Table 3).
4.3.1. Circ_0001445/ cSMARCA5
Despite an increasing number of studies in recent years have demonstrated the pivotal role of circRNA in regulating cellular functions and metabolism in cancer [137], only a limited number of circRNAs has a significant value in terms of HCC diagnosis and prognosis. One of the circRNAs correlated with poor prognosis is circ_0001445 (also named cSMARCA5). It is derived from exons 15 and 16 of the SWI/SNF-related, matrix-associated, actin-dependent regulator of chromatin, subfamily a, member 5 (SMARCA5) gene. Circ_0001445 acts as a sponge for miR-17-3p and miR-181b-5p, which regulate the levels of the tissue inhibitor of metalloproteinase-3 (TIMP3) implicated in matrix remodeling. In 2018, Zhang et al. [138] analyzed circ_0001445 level in 73 pairs of HCC and adjacent non-tumor tissues observing that circ_0001445 expression was significantly lower in tumor tissues compared to adjacent non-tumor tissues. Furthermore, its expression inversely correlated with the number of HCC foci. In HCC cells cultured in vitro, overexpression of circ_0001445 promoted apoptosis and inhibit proliferation, migration and invasion indicating a tumor suppressor effect. The authors also analyzed circ_0001445 levels in the plasma of HCC (104), cirrhosis (57), HBV (44) patients and healthy subjects (52). It was observed that circ_0001445 levels were markedly reduced in HCC patients in comparison to healthy controls, cirrhosis patients or HBV patients. Notably, the levels of circ_0001445 in HCC patients were significantly lower than in cirrhotic and HBV patients. These findings suggest that circ_0001445 may be a useful tool to discriminate HCC patients from cirrhotic/HBV patients. ROC curve analysis was performed to assess the diagnostic value of circ_0001445 for the discrimination of HCC. It is noteworthy that circ-RNA circ_0001445 resulted to be a more effective diagnostic marker compared to AFP.
In 2020, Xu et al.[139] corroborated the results of circ_0001445 down-regulation previously reported. The authors showed that its overexpression impairs HCC progression by down regulating the epithelial-mesenchymal transition (EMT) process and glycolysis in HCC cells in vitro. This phenotypic effect was due to the down regulation of the miR-942-5p/ALX4 (Aristaless-like homeobox 4) axis. In particular, circ_0001445 promoted ALX4 expression by targeting miR-942-5p. Notably, ALX4 is a transcription factor known to have anti HCC effects [140].
In 2020, Wang et al. [141] investigated the correlation between multiple circRNA, including circ_0001445, and the clinic-pathological characteristics of HCC patients. The authors demonstrated that reduced blood levels of circ_0001445 correlated with increased HCC malignancy as indicated by the presence of satellite nodules and frequent vascular invasion. These findings fully support the concept of the prognostic value for this circRNA.
In contrast to the above results, a recent control-case study [142], has indicated that circ_0001445 exhibits higher expression in the specimens of 106 patients with HCC compared to adjacent non-tumor tissue. Furthermore, the authors demonstrated that high expression of circ_0001445 in HCC patients represented a risk factor for 3-year OS (Hazard ratio=1.798, 95%CI: 1.165~3.231, p=0.0321). The results of the multivariate Cox analysis revealed that elevated circ_0001445 expression levels were associated with the development of portal vein tumor thrombus (PVTT), tumor size exceeding 5 cm, poor histopathologic grading and the presence of multiple tumor foci. These discrepant results indicate the necessity to clarify the role of circ_0001445 as a biomarker for HCC.
4.3.2. circ_0001649
circ_0001649 is a transcript product of the SHPRH (SNF2 histone linker PHD RING helicase) gene located on chromosome 6 [143]. SHPRH is a ubiquitously expressed protein that contains motifs characteristics of several DNA repair proteins, transcription factors, and helicases. Qin et al. [144] published one of the first studies on this circRNA in 2016. The authors evaluated the expression of circ_0001649 in 89 HCC samples and their paired adjacent liver tissues by qRT-PCR, showing a significant downregulation in HCC tissue. They also observed that circ_0001649 expression was negatively associated with tumor emboli formation and tumor size. Moreover, in vitro silencing in HCC cell showed that the mRNA levels of MMP (matrix methallo proteinases) 9, 10, and 13 were significantly increased after circ_0001649 knockdown. Notably, both the presence of tumor emboli and high levels of MMPs are known to correlate strongly with the invasive and metastatic properties of HCC. ROC curve analysis was performed to evaluate the prognostic power of circ_0001649. The AUC of circ_0001649 was 0.63. with a sensitivity and a specificity of 81% and 69%, respectively. These data suggest that circ_0001649 has the potential to be used as a novel biomarker for HCC.
In 2018, Zhang et al. [145] confirmed the potential biomarker role of circ_0001649 in HCC by analyzing the correlation between its expression level and HCC prognosis. This study was conducted on 77 tumor samples and their paired adjacent liver tissues. The results of the survival analysis indicated that patients with low expression levels of circ_0001649 exhibited the poorest OS rates. The univariate analysis revealed that circ_0001649, HCC stage and AST (transaminase) had a statistically significant impact on the OS of HCC patients. Furthermore, the multivariate analysis demonstrated that the expression levels of circ_0001649 and HCC stage were the independent prognostic factors for OS in HCC patients. Notably, the multivariate analysis also indicated that circ_0001649 levels were an independent prognostic factor for HCC patients.
4.3.4. circ_0003570
circ_0003570 is a highly conserved transcript product belonging to the family of the sequence similarity 53member B (FAM53B) gene, located on chromosome 10. In 2018, Fu et al. [146], demonstrated that circ_0003570 is downregulated in HCC cell lines and tissues. The authors conducted an analysis on 107 pairs of tumor tissues and adjacent non-tumor tissues obtained from patients with HCC. Firstly, they evaluated the expression level of circ_0003570 in cancerous tissues and in HCC cell lines observing that cancerous tissues as well as tumor cell lines exhibited a lower expression level of circ_0003570 compared to adjacent non-tumor tissues and human non-tumorigenic hepatic cell line, respectively. Moreover, the authors observed a progressive and statistically significant decrease in the expression levels of circ_0003570, from chronic hepatitis (14 cases) to fibrosis (63 cases) and HCC. This suggests that the expression levels of this circRNA may undergo dynamic changes in response to different liver disease. Finally, the researchers investigated the correlation between circ_0003570 levels and the HCC clinical pathological features. Significant correlations were observed between cicr_0003570 levels and tumor size, differentiation, microvascular invasion, HCC stage and serum AFP levels. The ROC curve analysis was performed to assess the potential diagnostic effectiveness of this circRNA, demonstrating an AUC of 0.70 with a sensitivity of 45% and a specificity of 86.8%. When the analysis was restricted to cirrhotic patients, the sensitivity increased to 69.7%, while the AUC and specificity remained comparable to those observed in the HCC patient group. These findings collectively indicate that circ_0003570 is significantly associated with the clinical and pathological characteristic of HCC patients and thus may serve as potential biomarker for differentiating between various liver pathologies.
In 2022, Zhang et al. [147] demonstrated that circ_0003570 down regulates the growth, migration and invasion of HCC cells; in a xenograft mouse model of HCC the authors also observed that circ_0003570 overexpression results in smaller tumor volume/weight compared to controls. At the molecular level, it was shown that circ_0003570 binds miR-182-5p thus regulating the repression of the downstream target gene StAR-related lipid transfer domain protein 13 (STARD13). STARD13 is implicated in HCC as it is down regulated in the less differentiated HCC forms.
The study conducted by Jang et al. [148] further corroborated the notion of circ_0003570 down-regulation in HCC. The clinical study included 162 HCC patients for which both tumor and adjacent non-tumor tissues were collected. The authors confirmed the reduced expression of circ_0003570 in HCC tissues vs paired non-cancerous tissues. Subsequently, patients were stratified according to the levels of circ_0003570, and classified as high or low-expression groups. Low circ_003570 were observed to be positively associated with larger tumors size (> 5 cm), vessel invasion, advanced HCC stage and higher AFP serum level. The survival analysis revealed a significant association between the OS of HCC patients and the expression levels of circ_0003570. In particular, the cumulative 1-, 2-, and 4-year OS rates in the low-expression group were 40.0%, 31.1%, 24.4%, respectively. In contrast, the OS rates increased up to 68.1%, 62.7% and 40%, respectively, in the high-expression group. The univariate analysis demonstrated that the significant predictors of OS were high circ_0003570 levels, the presence of multiple tumors, an AFP level higher than 200 ng/mL, chronic hepatitis B and curative treatment. The multivariate analysis identified high circ_0003570 levels and curative treatment as independent prognostic factors for OS. Similarly, a significant difference in PFS was observed between patients stratified according to their circ_0003570 expression levels. The cumulative 1-, 2-, and 4-year PFS rates in the low-expression group were 31.1%, 22.2%, and 15.6%, respectively. In contrast, the PFS rates increased up to 60.5%, 43.4% and 26.3%, respectively, in the high-expression group. The univariate analysis demonstrated that the prognostic factor for PFS were high circ_0003570 level, multiple tumors, AFP level higher than 200 ng/mL and curative treatment. The multivariate analysis identified as independent prognostic factors for PFS high circ_0003570 expression and curative treatment. Together these data support a significant correlation between circ_0003570 expression level with the clinical and pathological characteristics of HCC, and with the survival and progression of HCC patients.
Kang et al. further substantiated the prognostic value of circ_0003570 with regard to OS and PFS in HCC patients in 2023 [149]. The authors enrolled 86 patients with HVB-related HCC, who were divided into high- and low-expression groups, based on the median expression levels of the circRNA. The Kaplan–Meier survival analysis revealed that patients with low circ_0003570 expression displayed significantly reduced OS. The cumulative 1-, 3-, and 5-year OS rates were 76.7%, 58.1%, and 51.2%, respectively, for patients with high expression levels, in comparison with 46.5%, 30.2%, and 30.2% for those with low expression levels. The multivariate analysis identified advanced HCC stage, the presence of nodal involvement, metastasis, and sarcopenia as significant risk factors for OS in patients with HBV-related HCC. Similarly, the PFS rate was found to be significantly affected in patients with low expression levels compared to those with high expression levels. The cumulative 1-, 3-, and 5-year PFS rates were 39%, 21.8%, and 21.8%, respectively, for patients with low expression levels, in comparison to 71.4%, 34.3%, and 21.8% for those with high expression levels. The multivariate analysis demonstrated that advanced HCC stage and decompensated liver cirrhosis were significant risk factors for PSF in patients with HBV-HCC. Collectively, these data indicate that circ_0003570 expression levels associate with favorable clinical and pathological features, as well as improved OS and PFS in patients with HBV-HCC. It remains to be clarified whether this observation holds also for HCC induced by predisposing conditions other than HBV.
4.3.5. Other circRNAs
In 2019,circ_0028502 and circ_0076251, were identified as potential biomarkers for diagnosis and OS in HCC patients [150]. Circ_0028502 is transcribed from Solute Carrier Family 8-member B1 (SLC24A6) located on chromosome 12, while circ_0076251 is transcribed from Zinc Finger AN1-type containing 3 (ZFAND3) located on chromosome 6. The study was conducted in 100 paired samples of HCC and adjacent liver tissues obtained from patients who had undergone surgical treatment. The authors demonstrated that both circRNAs were downregulated in HCC tissues compared to normal tissues. Furthermore, they observed a correlation between the expression level of circ_0028502 and HCC stage; the levels of circ_0076251 were instead related to the presence of comorbidities, specifically type 2 diabetes mellitus and the presence of hepatitis B surface antigen (HbsAg). By extending the study population to include patients with chronic hepatitis (14) and liver fibrosis (56), the authors observed that both circRNA were differentially expressed in relation to liver disease, with a progressive and significant decrease from chronic hepatitis to fibrosis and to HCC. To evaluate the diagnostic efficacy of circ_0028502 and circ_0076251, ROC curves analyses were performed. The AUC for discriminating HCC from liver fibrosis and chronic hepatitis for circ_0028502 was 0.675, with a specificity of 72.1% and a sensitivity of 58%. Circ_0076251 demonstrated a higher AUC value (0.738) and sensitivity (64%) compared to circ_0028502, while maintaining a comparable specificity (71.3%). The combined diagnostic value of the two circRNAs was also evaluated, demonstrating a higher efficiency (AUC=0.754) compared to the use of the two circRNA independently. The Kaplan-Meier curve revealed that the survival rate differed depending on the expression levels of circ_0076251. Specifically, patients with low circ_0076251 levels of exhibited a poorer OS rate in comparison with those with high expression levels.
CircDLC1 derives from exons 14, 15 and 16 of the DLC1 gene. In 2021, Liu et al. [151] demonstrated that circDLC1 was markedly downregulated in HCC tissues and that this was significantly correlated with prognosis. The Kaplan-Meier’s survival curves revealed a positively correlations between low circDLC1 levels and poor OS and RFS. In vitro and in vivo assays demonstrated that circDLC1 inhibits metastasis in hepatoma cells by interacting with the RNA-binding protein HuR and reducing the stability of matrix metalloproteinases 1 (MMP1) mRNAs.
Another circRNA that has been identified as a potential biomarker for HCC progression is circ_0066659. Circ_0066659 is a 1254bp length RNA derived from the transmembrane protein 45A (TMEM45A) gene located on chromosome 3. In 2020, Zhang et al. [152] investigated the circRNA profiling in HCC samples and cell lines and selected circ_0066659 among the circRNAs found to be significantly upregulated. The study included paired HCC tissues and adjacent normal tissues obtained from patients with HCC. ROC curve analysis was employed to access the diagnostic value of circ_0066659, demonstrating a high accuracy (AUC=0.888). Furthermore, among patients exhibiting disparate circ_0066659 expression levels, HCC patients with low circ_0066659 expression displayed a prolonged OS rate compared to those with high circ_0066659 expression, as evidenced by Kaplan-Meier survival curve analysis. Moreover, high expression levels of circ_0066659 was significantly correlated with tumor size, HCC stage and vascular invasion. By in vitro analysis, the authors confirmed the oncogenic role of cicr_0066659 showing its ability to promote cell growth. The authors demonstrated that Circ_0066659 mechanically sponges miR-665 expression, resulting in the upregulation of insulin growth factor 2 (IGF2) whose overexpression accelerates formation of liver tumors and HCC progression in mouse [153].
In 2018, Chen et al. identified circ_0128298 as a potential biomarker in the diagnosis and prognosis of HCC [154]. The study, which considered 78 HCC and para-tumor tissues collected from patients with HCC who underwent surgery, demonstrated that circ_0128298 is significantly upregulated in HCC tissues compared to adjacent non-tumor tissues. The increased levels of circ_0128298 were found to be associated with the presence of vascular cancer embolus, lymphatic metastasis and organ metastasis. The ROC curve was employed to investigate the diagnostic value of circ_0128298 in differentiating between HCC tissues and adjacent non-tumor tissues, demonstrating a moderate accuracy (AUC=0.668) and sensitivity (67.4%) but a high specificity (80.5%). The univariate analysis demonstrated that gender, AFP levels, intrahepatic metastasis, organ metastasis and circ_0128298 expression levels, were independent factors in HCC. Furthermore, the multivariate COX regression analysis indicated that AFP levels and circ_0128298 expression levels were the prognostic factors predicting poor survival among HCC patients. Finally, HCC patients who exhibited lower expression levels of circ_0128298 demonstrated prolonged OS compared to patients with higher levels of circ_0128298. These findings collectively suggest that circ_0128298 may serve as a predictive biomarker for HCC occurrence and prognosis. However, further research is required to confirm the diagnostic efficacy of circ_0128298 and to gain a deeper understanding of the molecular and biological effects of circ_0128298 in HCC.
5. Conclusion
The increasing incidence of liver cancer together with the poor effectiveness of the therapeutic treatments especially for the advanced forms of the disease, make an early diagnosis/effective screening of the disease very urgent. The instrumental diagnostic tools despite being of great utility can be hardly used as screening methods. In this context, liquid biopsy has arisen as a potential novel approach for HCC diagnosis. The identification of blood markers has many amenable features among which the absence of any risk for patients, the possibility to be used as screening approach and the possibility to perform multiple tests allowing the real-time monitoring of HCC evolution. Unfortunately, the available blood marker of HCC markers, i.e., AFP, has several limitations as discussed at the beginning of the present review. In this context, the employment of ncRNAs may represent an interesting and novel diagnostic approach. Here we have presented some noticeable examples of the many works so far published in the field. All together, these works strongly support the potential effectiveness of ncRNA detection as diagnostic/prognostic markers for HCC. An evident advantage over AFP level determination is because for ncRNA detection we can use real time qRT-PCR and/or digital droplet PCR. These are techniques significantly more sensitive than those used to detect the AFP protein. Thus, it is reasonable to predict that ncRNA detection may be more precocious than AFP with obvious advantages.
Despite this, some considerations must be done on the use of ncRNA as HCC markers. First, it is not yet clear whether it is more appropriate the quantification of ncRNAs directly isolated from plasma or from extracellular vesicles (EVs) obtained from plasma. In EVs ncRNAs are protected from blood nucleases degradation, thus their presence and abundance may be enriched over ncRNAs freely circulating in the blood. Furthermore, it is not clear which ncRNAs circulate as naked molecules in the blood. Notably, none of the papers mentioned in this review specifically addressed this aspect. The second aspect deals with the relation between circulating ncRNAs and HCC tissue ncRNAs. In many of the papers mentioned here, the authors limited the ncRNA quantification in HCC tissue without checking blood levels. It is somehow acceptable that variations in the tissue levels may be reflected in the blood amount; however this is not necessarily the rule as for example Yousurf et al. [76] reported that miR-221 in the serum was down regulated while in HCC tissue it was up regulated. Third, being the nature of miRNAs and perhaps that of lncRNAs/circRNAs very promiscuous as multiple ncRNAs regulate the same mRNA and an mRNA is regulated by many ncRNAs, it is possible that a signature of multiple ncRNA is more appropriate to identify instead of a single ncRNA. In this regard, it is now well known that different type of tumors associates with the dysregulation of the same ncRNA. Thus, for the specific identification of a given tumor, the quantification of multiple ncRNAs may be required. Fourth, to be considered a valuable marker of HCC, there must be concordant data in the literature about a given ncRNA; this is however not always the case as for example circ_0001445 has been shown to be down regulated in HCC in some works [138,139,140,141] and up regulated in another [142] (see section 4.3 Circ-non-coding RNAs/Circ_0001445/ cSMARCA5).
In conclusion, despite further studies are necessary to better characterize the biology of ncRNA in general and in relation to HCC, we believe that ncRNA may represent attractive and novel markers for the early diagnoses of HCC and possible for its monitoring.
Author Contributions
Conceptualization, F.T., G.G., B.D.; methodology, D.T, A.B., M.G., C.G, B.D.; software, F.T., C.G.; data curation, D.T, A.B., M.G., C.G, B.D; writing—original draft preparation, F.T. and G.G,; writing—review and editing, D.T, A.B., M.G., C.G, B.D.; visualization, F.T., C.G.; supervision., G.G., B.D.; project administration, G.G.; funding acquisition, G.G. All authors have read and agreed to the published version of the manuscript.”..
Funding
We wish to thank the Italian Ministry of Foreign Affairs and International Cooperation (MAECI), grant number VN21GR01 and the “Lega Italiana Lotta Contro I Tumori” (project “Evaluation of a novel antifibrotic and antitumor molecule for hepatocellular carcinoma”) for the support.
Conflicts of Interest
The authors declare no conflicts of interest.” The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Dasgupta, P.; Henshaw,C.; Youlden,D.R.; Clark,P.J.; Aitken,J.F.; Baade,P.D. Global Trends in Incidence Rates of Primary Adult Liver Cancers: A Systematic Review and Meta-Analysis. Front Oncol. 2020, 10, 171. [CrossRef]
- Rumgay, H.; Arnold,M.; Ferlay,J.; Lesi,O.; Cabasag,C.J.; Vignat,J.; Laversanne,M.; McGlynn,K.A.; Soerjomataram,I. Global burden of primary liver cancer in 2020 and predictions to 2040. J Hepatol. 2022, 77, 1598-1606. [CrossRef]
- Siegel, R. L.; Miller,K.D.; Fuchs,H.E.; Jemal,A. Cancer statistics, 2022. CA Cancer J Clin. 2022, 72, 7-33.
- Rumgay, H.; Ferlay,J.; de,M.C.; Georges,D.; Ibrahim,A.S.; Zheng,R.; Wei,W.; Lemmens,V.E.P.P.; Soerjomataram,I. Global, regional and national burden of primary liver cancer by subtype. Eur. J Cancer. 2022, 161, 108-118. [CrossRef]
- Llovet, J. M.; Kelley,R.K.; Villanueva,A.; Singal,A.G.; Pikarsky,E.; Roayaie,S.; Lencioni,R.; Koike,K.; Zucman-Rossi,J.; Finn,R.S. Hepatocellular carcinoma. Nat. Rev Dis. Primers. 2021, 7, 6.
- Knox, J. J.; Cleary,S.P.; Dawson,L.A. Localized and systemic approaches to treating hepatocellular carcinoma. J. Clin. Oncol. 2015, 33, 1835-1844. [CrossRef]
- Affo, S.; Yu,L.X.; Schwabe,R.F. The Role of Cancer-Associated Fibroblasts and Fibrosis in Liver Cancer. Annu. Rev Pathol. 2017, 12, 153-186. [CrossRef]
- Schwabe, R. F. and Luedde,T. Apoptosis and necroptosis in the liver: a matter of life and death. Nat. Rev Gastroenterol. Hepatol. 2018, 15, 738-752. [CrossRef]
- Petrick, J. L.; Florio,A.A.; Znaor,A.; Ruggieri,D.; Laversanne,M.; Alvarez,C.S.; Ferlay,J.; Valery,P.C.; Bray,F.; McGlynn,K.A. International trends in hepatocellular carcinoma incidence, 1978-2012. Int. J Cancer. 2020, 147, 317-330. [CrossRef]
- Chidambaranathan-Reghupaty, S.; Fisher,P.B.; Sarkar,D. Hepatocellular carcinoma (HCC): Epidemiology, etiology and molecular classification. Adv. Cancer Res. 2021, 149, 1-61.
- EASL Clinical Practice Guidelines: Management of hepatocellular carcinoma. J Hepatol. 2018, 69, 182-236.
- Mazzaferro, V.; Regalia,E.; Doci,R.; Andreola,S.; Pulvirenti,A.; Bozzetti,F.; Montalto,F.; Ammatuna,M.; Morabito,A.; Gennari,L. Liver transplantation for the treatment of small hepatocellular carcinomas in patients with cirrhosis. N. Engl. J Med. 1996, 334, 693-699. [CrossRef]
- Ducreux, M.; Abou-Alfa,G.K.; Bekaii-Saab,T.; Berlin,J.; Cervantes,A.; de,B.T.; Eng,C.; Galle,P.; Gill,S.; Gruenberger,T.; Haustermans,K.; Lamarca,A.; Laurent-Puig,P.; Llovet,J.M.; Lordick,F.; Macarulla,T.; Mukherji,D.; Muro,K.; Obermannova,R.; O’Connor,J.M.; O’Reilly,E.M.; Osterlund,P.; Philip,P.; Prager,G.; Ruiz-Garcia,E.; Sangro,B.; Seufferlein,T.; Tabernero,J.; Verslype,C.; Wasan,H.; Van,C.E. The management of hepatocellular carcinoma. Current expert opinion and recommendations derived from the 24th ESMO/World Congress on Gastrointestinal Cancer, Barcelona, 2022. ESMO. Open. 2023, 8, 101567.
- Llovet, J. M.; de,B.T.; Kulik,L.; Haber,P.K.; Greten,T.F.; Meyer,T.; Lencioni,R. Locoregional therapies in the era of molecular and immune treatments for hepatocellular carcinoma. Nat. Rev Gastroenterol. Hepatol. 2021, 18, 293-313. [CrossRef]
- Hanna, R. F.; Miloushev,V.Z.; Tang,A.; Finklestone,L.A.; Brejt,S.Z.; Sandhu,R.S.; Santillan,C.S.; Wolfson,T.; Gamst,A.; Sirlin,C.B. Comparative 13-year meta-analysis of the sensitivity and positive predictive value of ultrasound, CT, and MRI for detecting hepatocellular carcinoma. Abdom. Radiol. (NY). 2016, 41, 71-90. [CrossRef]
- Chen, D. S.; Sung,J.L.; Sheu,J.C.; Lai,M.Y.; How,S.W.; Hsu,H.C.; Lee,C.S.; Wei,T.C. Serum alpha-fetoprotein in the early stage of human hepatocellular carcinoma. Gastroenterology. 1984, 86, 1404-1409. [CrossRef]
- Tzartzeva, K.; Obi,J.; Rich,N.E.; Parikh,N.D.; Marrero,J.A.; Yopp,A.; Waljee,A.K.; Singal,A.G. Surveillance Imaging and Alpha Fetoprotein for Early Detection of Hepatocellular Carcinoma in Patients With Cirrhosis: A Meta-analysis. Gastroenterology. 2018, 154, 1706-1718. [CrossRef]
- Yu, N. C.; Chaudhari,V.; Raman,S.S.; Lassman,C.; Tong,M.J.; Busuttil,R.W.; Lu,D.S. CT and MRI improve detection of hepatocellular carcinoma, compared with ultrasound alone, in patients with cirrhosis. Clin. Gastroenterol. Hepatol. 2011, 9, 161-167. [CrossRef]
- Gupta, P.; Soundararajan,R.; Patel,A.; Kumar,M.; Sharma,V.; Kalra,N. Abbreviated MRI for hepatocellular carcinoma screening: A systematic review and meta-analysis. J Hepatol. 2021, 75, 108-119. [CrossRef]
- Lehrich, B. M.; Zhang,J.; Monga,S.P.; Dhanasekaran,R. Battle of the biopsies: Role of tissue and liquid biopsy in hepatocellular carcinoma. J Hepatol. 2024, 80, 515-530. [CrossRef]
- Piccinino, F.; Sagnelli,E.; Pasquale,G.; Giusti,G. Complications following percutaneous liver biopsy. A multicentre retrospective study on 68,276 biopsies. J Hepatol. 1986, 2, 165-173. [CrossRef]
- Scott, E. C.; Baines,A.C.; Gong,Y.; Moore,R., Jr.; Pamuk,G.E.; Saber,H.; Subedee,A.; Thompson,M.D.; Xiao,W.; Pazdur,R.; Rao,V.A.; Schneider,J.; Beaver,J.A. Trends in the approval of cancer therapies by the FDA in the twenty-first century. Nat. Rev Drug Discov. 2023, 22, 625-640. [CrossRef]
- Tayob, N.; Kanwal,F.; Alsarraj,A.; Hernaez,R.; El-Serag,H.B. The Performance of AFP, AFP-3, DCP as Biomarkers for Detection of Hepatocellular Carcinoma (HCC): A Phase 3 Biomarker Study in the United States. Clin. Gastroenterol. Hepatol. 2023, 21, 415-423. [CrossRef]
- Cui, K.; Ou,Y.; Shen,Y.; Li,S.; Sun,Z. Clinical value of circulating tumor cells for the diagnosis and prognosis of hepatocellular carcinoma (HCC): A systematic review and meta-analysis. Medicine (Baltimore). 2020, 99, e22242.
- Tao, K.; Bian,Z.; Zhang,Q.; Guo,X.; Yin,C.; Wang,Y.; Zhou,K.; Wan,S.; Shi,M.; Bao,D.; Yang,C.; Xing,J. Machine learning-based genome-wide interrogation of somatic copy number aberrations in circulating tumor DNA for early detection of hepatocellular carcinoma. EBioMedicine. 2020, 56, 102811. [CrossRef]
- Nemeth, K.; Bayraktar,R.; Ferracin,M.; Calin,G.A. Non-coding RNAs in disease: from mechanisms to therapeutics. Nat. Rev Genet. 2024, 25, 211-232. [CrossRef]
- Uszczynska-Ratajczak, B.; Lagarde,J.; Frankish,A.; Guigo,R.; Johnson,R. Towards a complete map of the human long non-coding RNA transcriptome. Nat. Rev Genet. 2018, 19, 535-548. [CrossRef]
- Elbashir, S. M.; Harborth,J.; Lendeckel,W.; Yalcin,A.; Weber,K.; Tuschl,T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature. 2001, 411, 494-498. [CrossRef]
- Kozomara, A.; Birgaoanu,M.; Griffiths-Jones,S. miRBase: from microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155-D162. [CrossRef]
- Lee, Y.; Jeon,K.; Lee,J.T.; Kim,S.; Kim,V.N. MicroRNA maturation: stepwise processing and subcellular localization. EMBO J. 2002, 21, 4663-4670. [CrossRef]
- Aukerman, M. J. and Sakai,H. Regulation of flowering time and floral organ identity by a MicroRNA and its APETALA2-like target genes. Plant Cell. 2003, 15, 2730-2741. [CrossRef]
- Carthew, R. W. and Sontheimer,E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell. 2009, 136, 642-655. [CrossRef]
- Bracht, J.; Hunter,S.; Eachus,R.; Weeks,P.; Pasquinelli,A.E. Trans-splicing and polyadenylation of let-7 microRNA primary transcripts. RNA. 2004, 10, 1586-1594. [CrossRef]
- Lee, Y.; Ahn,C.; Han,J.; Choi,H.; Kim,J.; Yim,J.; Lee,J.; Provost,P.; Radmark,O.; Kim,S.; Kim,V.N. The nuclear RNase III Drosha initiates microRNA processing. Nature. 2003, 425, 415-419. [CrossRef]
- Ray, A.; Sarkar,A.; Banerjee,S.; Biswas,K. Non-Canonical Targets of MicroRNAs: Role in Transcriptional Regulation, Disease Pathogenesis and Potential for Therapeutic Targets. Microrna. 2024. [CrossRef]
- Vasudevan, S. Posttranscriptional upregulation by microRNAs. Wiley. Interdiscip. Rev RNA. 2012, 3, 311-330. [CrossRef]
- Bartel, D. P. Metazoan MicroRNAs. Cell. 2018, 173, 20-51.
- Kumar, M. A.; Baba,S.K.; Sadida,H.Q.; Marzooqi,S.A.; Jerobin,J.; Altemani,F.H.; Algehainy,N.; Alanazi,M.A.; Abou-Samra,A.B.; Kumar,R.; Al-Shabeeb Akil,A.S.; Macha,M.A.; Mir,R.; Bhat,A.A. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct. Target Ther. 2024, 9, 27. [CrossRef]
- Ladd, A. D.; Duarte,S.; Sahin,I.; Zarrinpar,A. Mechanisms of drug resistance in HCC. Hepatology. 2024, 79, 926-940. [CrossRef]
- Tian, Y.; Zhang,M.; Liu,L.X.; Wang,Z.C.; Liu,B.; Huang,Y.; Wang,X.; Ling,Y.Z.; Wang,F.; Feng,X.; Tu,Y. Exploring non-coding RNA mechanisms in hepatocellular carcinoma: implications for therapy and prognosis. Front Immunol. 2024, 15, 1400744. [CrossRef]
- Mallela, V. R.; Rajtmajerova,M.; Trailin,A.; Liska,V.; Hemminki,K.; Ambrozkiewicz,F. miRNA and lncRNA as potential tissue biomarkers in hepatocellular carcinoma. Noncoding. RNA. Res. 2024, 9, 24-32. [CrossRef]
- Jesenko, T.; Brezar,S.K.; Cemazar,M.; Biasin,A.; Tierno,D.; Scaggiante,B.; Grassi,M.; Grassi,C.; Dapas,B.; Truong,N.H.; Abrami,M.; Zanconati,F.; Bonazza,D.; Rizzolio,F.; Parisi,S.; Pastorin,G.; Grassi,G. Targeting Non-Coding RNAs for the Development of Novel Hepatocellular Carcinoma Therapeutic Approaches. Pharmaceutics. 2023, 15.
- Quinn, J. J. and Chang,H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev Genet. 2016, 17, 47-62. [CrossRef]
- Brannan, C. I.; Dees,E.C.; Ingram,R.S.; Tilghman,S.M. The product of the H19 gene may function as an RNA. Mol. Cell Biol. 1990, 10, 28-36.
- Zhao, L.; Wang,J.; Li,Y.; Song,T.; Wu,Y.; Fang,S.; Bu,D.; Li,H.; Sun,L.; Pei,D.; Zheng,Y.; Huang,J.; Xu,M.; Chen,R.; Zhao,Y.; He,S. NONCODEV6: an updated database dedicated to long non-coding RNA annotation in both animals and plants. Nucleic Acids Res. 2021, 49, D165-D171. [CrossRef]
- Derrien, T.; Johnson,R.; Bussotti,G.; Tanzer,A.; Djebali,S.; Tilgner,H.; Guernec,G.; Martin,D.; Merkel,A.; Knowles,D.G.; Lagarde,J.; Veeravalli,L.; Ruan,X.; Ruan,Y.; Lassmann,T.; Carninci,P.; Brown,J.B.; Lipovich,L.; Gonzalez,J.M.; Thomas,M.; Davis,C.A.; Shiekhattar,R.; Gingeras,T.R.; Hubbard,T.J.; Notredame,C.; Harrow,J.; Guigo,R. The GENCODE v7 catalog of human long noncoding RNAs: analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775-1789.
- Wu, H.; Yang,L.; Chen,L.L. The Diversity of Long Noncoding RNAs and Their Generation. Trends Genet. 2017, 33, 540-552. [CrossRef]
- He, J.; Huang,B.; Zhang,K.; Liu,M.; Xu,T. Long non-coding RNA in cervical cancer: From biology to therapeutic opportunity. Biomed. Pharmacother. 2020, 127, 110209. [CrossRef]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev Genet. 2011, 12, 861-874. [CrossRef]
- Chen, L. L. The biogenesis and emerging roles of circular RNAs. Nat. Rev Mol. Cell Biol. 2016, 17, 205-211. [CrossRef]
- Li, X.; Yang,L.; Chen,L.L. The Biogenesis, Functions, and Challenges of Circular RNAs. Mol. Cell. 2018, 71, 428-442. [CrossRef]
- Liang, D. and Wilusz,J.E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev. 2014, 28, 2233-2247. [CrossRef]
- Pisignano, G.; Michael,D.C.; Visal,T.H.; Pirlog,R.; Ladomery,M.; Calin,G.A. Going circular: history, present, and future of circRNAs in cancer. Oncogene. 2023, 42, 2783-2800. [CrossRef]
- Li, Z.; Huang,C.; Bao,C.; Chen,L.; Lin,M.; Wang,X.; Zhong,G.; Yu,B.; Hu,W.; Dai,L.; Zhu,P.; Chang,Z.; Wu,Q.; Zhao,Y.; Jia,Y.; Xu,P.; Liu,H.; Shan,G. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256-264. [CrossRef]
- Czubak, K.; Sedehizadeh,S.; Kozlowski,P.; Wojciechowska,M. An Overview of Circular RNAs and Their Implications in Myotonic Dystrophy. Int. J Mol. Sci. 2019, 20. [CrossRef]
- Rybak-Wolf, A.; Stottmeister,C.; Glazar,P.; Jens,M.; Pino,N.; Giusti,S.; Hanan,M.; Behm,M.; Bartok,O.; Ashwal-Fluss,R.; Herzog,M.; Schreyer,L.; Papavasileiou,P.; Ivanov,A.; Ohman,M.; Refojo,D.; Kadener,S.; Rajewsky,N. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell. 2015, 58, 870-885.
- Zhang, X. O.; Wang,H.B.; Zhang,Y.; Lu,X.; Chen,L.L.; Yang,L. Complementary sequence-mediated exon circularization. Cell. 2014, 159, 134-147. [CrossRef]
- Memczak, S.; Jens,M.; Elefsinioti,A.; Torti,F.; Krueger,J.; Rybak,A.; Maier,L.; Mackowiak,S.D.; Gregersen,L.H.; Munschauer,M.; Loewer,A.; Ziebold,U.; Landthaler,M.; Kocks,C.; le,N.F.; Rajewsky,N. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013, 495, 333-338. [CrossRef]
- Salzman, J.; Chen,R.E.; Olsen,M.N.; Wang,P.L.; Brown,P.O. Cell-type specific features of circular RNA expression. PLoS. Genet. 2013, 9, e1003777. [CrossRef]
- Guo, T.; Tang,X.H.; Gao,X.Y.; Zhou,Y.; Jin,B.; Deng,Z.Q.; Hu,Y.; Xing,X.F.; Li,Z.Y.; Ji,J.F. A liquid biopsy signature of circulating exosome-derived mRNAs, miRNAs and lncRNAs predict therapeutic efficacy to neoadjuvant chemotherapy in patients with advanced gastric cancer. Mol. Cancer. 2022, 21, 216. [CrossRef]
- Shi, M.; Jia,J.S.; Gao,G.S.; Hua,X. Advances and challenges of exosome-derived noncoding RNAs for hepatocellular carcinoma diagnosis and treatment. Biochem. Biophys. Rep. 2024, 38, 101695. [CrossRef]
- Johnstone, R. M.; Adam,M.; Hammond,J.R.; Orr,L.; Turbide,C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol. Chem. 1987, 262, 9412-9420.
- Raposo, G.; Nijman,H.W.; Stoorvogel,W.; Liejendekker,R.; Harding,C.V.; Melief,C.J.; Geuze,H.J. B lymphocytes secrete antigen-presenting vesicles. J Exp. Med. 1996, 183, 1161-1172. [CrossRef]
- Valadi, H.; Ekstrom,K.; Bossios,A.; Sjostrand,M.; Lee,J.J.; Lotvall,J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654-659. [CrossRef]
- Hood, J. L.; San,R.S.; Wickline,S.A. Exosomes released by melanoma cells prepare sentinel lymph nodes for tumor metastasis. Cancer Res. 2011, 71, 3792-3801. [CrossRef]
- Fang, T.; Lv,H.; Lv,G.; Li,T.; Wang,C.; Han,Q.; Yu,L.; Su,B.; Guo,L.; Huang,S.; Cao,D.; Tang,L.; Tang,S.; Wu,M.; Yang,W.; Wang,H. Tumor-derived exosomal miR-1247-3p induces cancer-associated fibroblast activation to foster lung metastasis of liver cancer. Nat. Commun. 2018, 9, 191. [CrossRef]
- Zhou, Y.; Ren,H.; Dai,B.; Li,J.; Shang,L.; Huang,J.; Shi,X. Hepatocellular carcinoma-derived exosomal miRNA-21 contributes to tumor progression by converting hepatocyte stellate cells to cancer-associated fibroblasts. J Exp. Clin. Cancer Res. 2018, 37, 324. [CrossRef]
- Ma, D.; Gao,X.; Liu,Z.; Lu,X.; Ju,H.; Zhang,N. Exosome-transferred long non-coding RNA ASMTL-AS1 contributes to malignant phenotypes in residual hepatocellular carcinoma after insufficient radiofrequency ablation. Cell Prolif. 2020, 53, e12795. [CrossRef]
- Li, X.; Lei,Y.; Wu,M.; Li,N. Regulation of Macrophage Activation and Polarization by HCC-Derived Exosomal lncRNA TUC339. Int. J Mol. Sci. 2018, 19. [CrossRef]
- Chen, W.; Quan,Y.; Fan,S.; Wang,H.; Liang,J.; Huang,L.; Chen,L.; Liu,Q.; He,P.; Ye,Y. Exosome-transmitted circular RNA hsa_circ_0051443 suppresses hepatocellular carcinoma progression. Cancer Lett. 2020, 475, 119-128. [CrossRef]
- Fang, Y.; Zhang,Q.; Chen,Z.; Guo,C.; Wu,J. Clinical significance and immune characteristics analysis of miR-221-3p and its key target genes related to epithelial-mesenchymal transition in breast cancer. Aging (Albany. NY). 2024, 16, 322-347. [CrossRef]
- Eslaminejad, T.; Nematollahi-Mahani,S.N.; Sargazi,M.L.; Ansari,M.; Mirzaie,V. Evaluating the effects of curcumin nano-chitosan on miR-221 and miR-222 expression and Wnt/beta-catenin pathways in MCF-7, MDA-MB-231 and SKBR3 cell lines. Diagn. Pathol. 2024, 19, 35.
- Hu, G.; Zhai,S.; Yu,S.; Huang,Z.; Gao,R. Circular RNA circRHOBTB3 is downregulated in hepatocellular carcinoma and suppresses cell proliferation by inhibiting miR-18a maturation. Infect. Agent. Cancer. 2021, 16, 48. [CrossRef]
- Yun, J. H.; Baek,M.J.; Jung,H.I. Expression of miR-221 and miR-18a in patients with hepatocellular carcinoma and its clinical significance. Korean J Clin. Oncol. 2022, 18, 17-26. [CrossRef]
- Chen, F.; Li,X.F.; Fu,D.S.; Huang,J.G.; Yang,S.E. Clinical potential of miRNA-221 as a novel prognostic biomarker for hepatocellular carcinoma. Cancer Biomark. 2017, 18, 209-214. [CrossRef]
- Yousuf, T.; Dar,S.B.; Bangri,S.A.; Choh,N.A.; Rasool,Z.; Shah,A.; Rather,R.A.; Rah,B.; Bhat,G.R.; Ali,S.; Afroze,D. Diagnostic implication of a circulating serum-based three-microRNA signature in hepatocellular carcinoma. Front Genet. 2022, 13, 929787.
- Liu, W.; Hu,K.; Zhang,F.; Lu,S.; Chen,R.; Ren,Z.; Yin,X. The prognostic significance of microRNA-221 in hepatocellular carcinoma: An updated meta-analysis. Int. J Biol. Markers. 2021, 36, 17246008211032689. [CrossRef]
- Yang, X.; Wang,M.; Lin,B.; Yao,D.; Li,J.; Tang,X.; Li,S.; Liu,Y.; Xie,R.; Yu,S. miR-487a promotes progression of gastric cancer by targeting TIA1. Biochimie. 2018, 154, 119-126.
- Luo, A.; Liu,H.; Huang,C. KLF5-induced miR-487a augments the progression of osteosarcoma cells by targeting NKX3-1 in vitro. Oncol. Lett. 2022, 24, 258.
- Chang, R. M.; Xiao,S.; Lei,X.; Yang,H.; Fang,F.; Yang,L.Y. miRNA-487a Promotes Proliferation and Metastasis in Hepatocellular Carcinoma. Clin. Cancer Res. 2017, 23, 2593-2604. [CrossRef]
- Cirera-Salinas, D.; Pauta,M.; Allen,R.M.; Salerno,A.G.; Ramirez,C.M.; Chamorro-Jorganes,A.; Wanschel,A.C.; Lasuncion,M.A.; Morales-Ruiz,M.; Suarez,Y.; Baldan,A.; Esplugues,E.; Fernandez-Hernando,C. Mir-33 regulates cell proliferation and cell cycle progression. Cell Cycle. 2012, 11, 922-933. [CrossRef]
- Najafi-Shoushtari, S. H.; Kristo,F.; Li,Y.; Shioda,T.; Cohen,D.E.; Gerszten,R.E.; Naar,A.M. MicroRNA-33 and the SREBP host genes cooperate to control cholesterol homeostasis. Science. 2010, 328, 1566-1569. [CrossRef]
- Ramirez, C. M.; Goedeke,L.; Rotllan,N.; Yoon,J.H.; Cirera-Salinas,D.; Mattison,J.A.; Suarez,Y.; de,C.R.; Gorospe,M.; Fernandez-Hernando,C. MicroRNA 33 regulates glucose metabolism. Mol. Cell Biol. 2013, 33, 2891-2902. [CrossRef]
- Zhang, T.; Han,G.; Wang,Y.; Chen,K.; Sun,Y. MicroRNA expression profiles in supraglottic carcinoma. Oncol. Rep. 2014, 31, 2029-2034. [CrossRef]
- Zhou, Y.; Huang,Z.; Wu,S.; Zang,X.; Liu,M.; Shi,J. miR-33a is up-regulated in chemoresistant osteosarcoma and promotes osteosarcoma cell resistance to cisplatin by down-regulating TWIST. J Exp. Clin. Cancer Res. 2014, 33, 12. [CrossRef]
- Huang, C. F.; Sun,C.C.; Zhao,F.; Zhang,Y.D.; Li,D.J. miR-33a levels in hepatic and serum after chronic HBV-induced fibrosis. J Gastroenterol. 2015, 50, 480-490. [CrossRef]
- Xie, R. T.; Cong,X.L.; Zhong,X.M.; Luo,P.; Yang,H.Q.; Lu,G.X.; Luo,P.; Chang,Z.Y.; Sun,R.; Wu,T.M.; Lv,Z.W.; Fu,D.; Ma,Y.S. MicroRNA-33a downregulation is associated with tumorigenesis and poor prognosis in patients with hepatocellular carcinoma. Oncol. Lett. 2018, 15, 4571-4577. [CrossRef]
- Mangiapane, G.; Pascut,D.; Dalla,E.; Antoniali,G.; Degrassi,M.; Croce,L.S.; De,S., V; Piazza,S.; Canarutto,G.; Tiribelli,C.; Tell,G. Clinical Significance of Apurinic/Apyrimidinic Endodeoxyribonuclease 1 and MicroRNA Axis in Hepatocellular Carcinoma. J Clin. Transl. Hepatol. 2023, 11, 1291-1307.
- Mourelatos, Z.; Dostie,J.; Paushkin,S.; Sharma,A.; Charroux,B.; Abel,L.; Rappsilber,J.; Mann,M.; Dreyfuss,G. miRNPs: a novel class of ribonucleoproteins containing numerous microRNAs. Genes Dev. 2002, 16, 720-728.
- Kou, X.; Ding,H.; Li,L.; Chao,H. Hsa-miR-105-1 Regulates Cisplatin-Resistance in Ovarian Carcinoma Cells by Targeting ANXA9. Anal. Cell Pathol. (Amst). 2021, 2021, 6662486.
- Zhou, W.; Fong,M.Y.; Min,Y.; Somlo,G.; Liu,L.; Palomares,M.R.; Yu,Y.; Chow,A.; O’Connor,S.T.; Chin,A.R.; Yen,Y.; Wang,Y.; Marcusson,E.G.; Chu,P.; Wu,J.; Wu,X.; Li,A.X.; Li,Z.; Gao,H.; Ren,X.; Boldin,M.P.; Lin,P.C.; Wang,S.E. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell. 2014, 25, 501-515.
- Guan, Y.; Chen,L.; Bao,Y.; Li,Z.; Cui,R.; Li,G.; Wang,Y. Identification of low miR-105 expression as a novel poor prognostic predictor for human glioma. Int. J Clin. Exp. Med. 2015, 8, 10855-10864.
- Shen, G.; Rong,X.; Zhao,J.; Yang,X.; Li,H.; Jiang,H.; Zhou,Q.; Ji,T.; Huang,S.; Zhang,J.; Jia,H. MicroRNA-105 suppresses cell proliferation and inhibits PI3K/AKT signaling in human hepatocellular carcinoma. Carcinogenesis. 2014, 35, 2748-2755. [CrossRef]
- Ma, Y. S.; Wu,T.M.; Lv,Z.W.; Lu,G.X.; Cong,X.L.; Xie,R.T.; Yang,H.Q.; Chang,Z.Y.; Sun,R.; Chai,L.; Cai,M.X.; Zhong,X.J.; Zhu,J.; Fu,D. High expression of miR-105-1 positively correlates with clinical prognosis of hepatocellular carcinoma by targeting oncogene NCOA1. Oncotarget. 2017, 8, 11896-11905. [CrossRef]
- Zhou, W.; Zhou,W.; Zeng,Q.; Xiong,J. MicroRNA-138 inhibits hypoxia-induced proliferation of endothelial progenitor cells via inhibition of HIF-1alpha-mediated MAPK and AKT signaling. Exp. Ther. Med. 2017, 13, 1017-1024.
- Liu, H.; Tang,T.; Hu,X.; Tan,W.; Zhou,P.; Zhang,H.; Liu,Y.; Chen,C.; Yang,M.; Zhou,M.; Xuan,S.; Cheng,B.; Yin,W.; Lin,J. miR-138-5p Inhibits Vascular Mimicry by Targeting the HIF-1alpha/VEGFA Pathway in Hepatocellular Carcinoma. J Immunol. Res. 2022, 2022, 7318950.
- Lu, Y.; Liu,Y.; Zhang,K.; Jiang,L. Circular RNA TLK1 Exerts Oncogenic Functions in Hepatocellular Carcinoma by Acting as a ceRNA of miR-138-5p. J Oncol. 2022, 2022, 2415836. [CrossRef]
- Zheng, J. H.; Jian,Z.X.; Jin,H.S.; Chen,S.C.; Wang,G.Y. [Expression of SOX4 gene and early recurrence of hepatocellular carcinoma: their relationship and the clinical significance]. Nan. Fang Yi. Ke. Da. Xue. Xue. Bao. 2010, 30, 818-819.
- Xiao, J. X.; Xu,W.; Fei,X.; Hao,F.; Wang,N.; Chen,Y.; Wang,J. Anillin facilitates cell proliferation and induces tumor growth of hepatocellular carcinoma via miR-138/SOX4 axis regulation. Transl. Oncol. 2020, 13, 100815.
- Jenike, A. E. and Halushka,M.K. miR-21: a non-specific biomarker of all maladies. Biomark. Res. 2021, 9, 18. [CrossRef]
- Mjelle, R.; Dima,S.O.; Bacalbasa,N.; Chawla,K.; Sorop,A.; Cucu,D.; Herlea,V.; Saetrom,P.; Popescu,I. Comprehensive transcriptomic analyses of tissue, serum, and serum exosomes from hepatocellular carcinoma patients. BMC. Cancer. 2019, 19, 1007.
- Stechele, M.; Link,H.; Hirner-Eppeneder,H.; Alunni-Fabbroni,M.; Wildgruber,M.; Salvermoser,L.; Corradini,S.; Schinner,R.; Ben,K.N.; Rossler,D.; Galun,E.; Goldberg,S.N.; Ricke,J.; Kazmierczak,P.M. Circulating miR-21 as a prognostic biomarker in HCC treated by CT-guided high-dose rate brachytherapy. Radiat. Oncol. 2023, 18, 125. [CrossRef]
- Huang, X.; Xiong,Y.; Yang,J.; Yang,G.; Li,J. The Prognostic Significance of miR-21 Expression among Surgically Resected Hepatocellular Carcinoma Patients: Evidence from a Meta-Analysis and Retrospective Cohort Study. Biomed. Res. Int. 2020, 2020, 8848158. [CrossRef]
- Zhang, N.; Hu,Z.; Qiang,Y.; Zhu,X. Circulating miR-130b- and miR-21-based diagnostic markers and therapeutic targets for hepatocellular carcinoma. Mol. Genet. Genomic. Med. 2019, 7, e1012.
- Li, Z. Y.; Xi,Y.; Zhu,W.N.; Zeng,C.; Zhang,Z.Q.; Guo,Z.C.; Hao,D.L.; Liu,G.; Feng,L.; Chen,H.Z.; Chen,F.; Lv,X.; Liu,D.P.; Liang,C.C. Positive regulation of hepatic miR-122 expression by HNF4alpha. J Hepatol. 2011, 55, 602-611. [CrossRef]
- Bandiera, S.; Pfeffer,S.; Baumert,T.F.; Zeisel,M.B. miR-122--a key factor and therapeutic target in liver disease. J Hepatol. 2015, 62, 448-457. [CrossRef]
- Hao, P.; Li,Q.; Zhao,H. Mucin 1 expression is regulated by hsa_circ_0055054/microRNA‑122‑5p and promotes hepatocellular carcinoma development. Oncol. Lett. 2024, 28, 404.
- Zhao, X. F.; Li,N.; Lin,D.D.; Sun,L.B. Circulating MicroRNA-122 for the Diagnosis of Hepatocellular Carcinoma: A Meta-Analysis. Biomed. Res. Int. 2020, 2020, 5353695. [CrossRef]
- Fang, Y.; Yan,D.; Wang,L.; Zhang,J.; He,Q. Circulating microRNAs (miR-16, miR-22, miR-122) expression and early diagnosis of hepatocellular carcinoma. J Clin. Lab Anal. 2022, 36, e24541.
- Pelizzaro, F.; Cardin,R.; Sartori,A.; Imondi,A.; Penzo,B.; Aliberti,C.; Ponzoni,A.; Vitale,A.; Cillo,U.; Farinati,F. Circulating MicroRNA-21 and MicroRNA-122 as Prognostic Biomarkers in Hepatocellular Carcinoma Patients Treated with Transarterial Chemoembolization. Biomedicines. 2021, 9. [CrossRef]
- Ambade, A.; Satishchandran,A.; Szabo,G. Alcoholic hepatitis accelerates early hepatobiliary cancer by increasing stemness and miR-122-mediated HIF-1alpha activation. Sci Rep. 2016, 6, 21340. [CrossRef]
- Krishnan, K.; Steptoe,A.L.; Martin,H.C.; Pattabiraman,D.R.; Nones,K.; Waddell,N.; Mariasegaram,M.; Simpson,P.T.; Lakhani,S.R.; Vlassov,A.; Grimmond,S.M.; Cloonan,N. miR-139-5p is a regulator of metastatic pathways in breast cancer. RNA. 2013, 19, 1767-1780. [CrossRef]
- Wong, C. C.; Wong,C.M.; Tung,E.K.; Au,S.L.; Lee,J.M.; Poon,R.T.; Man,K.; Ng,I.O. The microRNA miR-139 suppresses metastasis and progression of hepatocellular carcinoma by down-regulating Rho-kinase 2. Gastroenterology. 2011, 140, 322-331. [CrossRef]
- Croft, D. R. and Olson,M.F. The Rho GTPase effector ROCK regulates cyclin A, cyclin D1, and p27Kip1 levels by distinct mechanisms. Mol. Cell Biol. 2006, 26, 4612-4627. [CrossRef]
- Huang, D.; Du,X.; Yuan,R.; Chen,L.; Liu,T.; Wen,C.; Huang,M.; Li,M.; Hao,L.; Shao,J. Rock2 promotes the invasion and metastasis of hepatocellular carcinoma by modifying MMP2 ubiquitination and degradation. Biochem. Biophys. Res. Commun. 2014, 453, 49-56. [CrossRef]
- Wang, Z.; Ding,Q.; Li,Y.; Liu,Q.; Wu,W.; Wu,L.; Yu,H. Reanalysis of microRNA expression profiles identifies novel biomarkers for hepatocellular carcinoma prognosis. Tumour. Biol. 2016, 37, 14779-14787. [CrossRef]
- Wang, X.; Gao,J.; Zhou,B.; Xie,J.; Zhou,G.; Chen,Y. Identification of prognostic markers for hepatocellular carcinoma based on miRNA expression profiles. Life Sci. 2019, 232, 116596. [CrossRef]
- Tonon, F.; Cemazar,M.; Kamensek,U.; Zennaro,C.; Pozzato,G.; Caserta,S.; Ascione,F.; Grassi,M.; Guido,S.; Ferrari,C.; Cansolino,L.; Trotta,F.; Kuzmanov,B.G.; Forte,G.; Martino,F.; Perrone,F.; Bomben,R.; Gattei,V.; Elvassore,N.; Murano,E.; Truong,N.H.; Olson,M.; Farra,R.; Grassi,G.; Dapas,B. 5-Azacytidine Downregulates the Proliferation and Migration of Hepatocellular Carcinoma Cells In Vitro and In Vivo by Targeting miR-139-5p/ROCK2 Pathway. Cancers. (Basel). 2022, 14.
- Panzitt, K.; Tschernatsch,M.M.; Guelly,C.; Moustafa,T.; Stradner,M.; Strohmaier,H.M.; Buck,C.R.; Denk,H.; Schroeder,R.; Trauner,M.; Zatloukal,K. Characterization of HULC, a novel gene with striking up-regulation in hepatocellular carcinoma, as noncoding RNA. Gastroenterology. 2007, 132, 330-342. [CrossRef]
- Shah, M. and Sarkar,D. HCC-Related lncRNAs: Roles and Mechanisms. Int. J Mol. Sci. 2024, 25. [CrossRef]
- Lumkul, L.; Jantaree,P.; Jaisamak,K.; Wongkummool,W.; Lapisatepun,W.; Orrapin,S.; Udomruk,S.; Lo,P.L.; Chaiyawat,P. Combinatorial Gene Expression Profiling of Serum HULC, HOTAIR, and UCA1 lncRNAs to Differentiate Hepatocellular Carcinoma from Liver Diseases: A Systematic Review and Meta-Analysis. Int. J Mol. Sci. 2024, 25.
- Wang, B. G.; Lv,Z.; Ding,H.X.; Fang,X.X.; Wen,J.; Xu,Q.; Yuan,Y. The association of lncRNA-HULC polymorphisms with hepatocellular cancer risk and prognosis. Gene. 2018, 670, 148-154.
- Sonohara, F.; Inokawa,Y.; Hayashi,M.; Yamada,S.; Sugimoto,H.; Fujii,T.; Kodera,Y.; Nomoto,S. Prognostic Value of Long Non-Coding RNA HULC and MALAT1 Following the Curative Resection of Hepatocellular Carcinoma. Sci Rep. 2017, 7, 16142. [CrossRef]
- Yang, Z.; Lu,Y.; Xu,Q.; Tang,B.; Park,C.K.; Chen,X. HULC and H19 Played Different Roles in Overall and Disease-Free Survival from Hepatocellular Carcinoma after Curative Hepatectomy: A Preliminary Analysis from Gene Expression Omnibus. Dis. Markers. 2015, 2015, 191029. [CrossRef]
- Ji, P.; Diederichs,S.; Wang,W.; Boing,S.; Metzger,R.; Schneider,P.M.; Tidow,N.; Brandt,B.; Buerger,H.; Bulk,E.; Thomas,M.; Berdel,W.E.; Serve,H.; Muller-Tidow,C. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene. 2003, 22, 8031-8041.
- Golam, R. M.; Khalil,M.A.F.; Shaker,O.G.; Ahmed,T.I.; Elguaad,M.K.A.; Hassan,E.A.; El-Ansary,M.R.M.; Ismail,A.; Kandil,Y.I.; Mohammed,O.A.; Doghish,A.S. The clinical significance of long non-coding RNAs MALAT1 and CASC2 in the diagnosis of HCV-related hepatocellular carcinoma. PLoS. One. 2024, 19, e0303314. [CrossRef]
- Liao, X.; Chen,J.; Luo,D.; Luo,B.; Huang,W.; Xie,W. Prognostic value of long non-coding RNA MALAT1 in hepatocellular carcinoma: A study based on multi-omics analysis and RT-PCR validation. Pathol. Oncol. Res. 2022, 28, 1610808. [CrossRef]
- Li, J.; Wang,X.; Tang,J.; Jiang,R.; Zhang,W.; Ji,J.; Sun,B. HULC and Linc00152 Act as Novel Biomarkers in Predicting Diagnosis of Hepatocellular Carcinoma. Cell Physiol Biochem. 2015, 37, 687-696. [CrossRef]
- Liu, G.; Yang,Z.F.; Sun,J.; Sun,B.Y.; Zhou,P.Y.; Zhou,C.; Guan,R.Y.; Wang,Z.T.; Yi,Y.; Qiu,S.J. The LINC00152/miR-205-5p/CXCL11 axis in hepatocellular carcinoma cancer-associated fibroblasts affects cancer cell phenotypes and tumor growth. Cell Oncol. (Dordr. ). 2022, 45, 1435-1449.
- Wang, B.; Yang,S.; Zhao,W. Long Non-Coding RNA NRAD1 and LINC00152 are Highly Expressed and Associated with Prognosis in Patients with Hepatocellular Carcinoma. Onco. Targets. Ther. 2020, 13, 10409-10416. [CrossRef]
- Shehab-Eldeen, S.; Essa,A.; Arafat,E.S.; Sleem,A.S.; Alhosary,A.A.; Darwish,E.; Essa,A.; Al-Omair,O.A.; Al-Khoufi,E.A.; Al Abdulqader,A.K.; Nada,A. Serum LINC00152 and UCA1 in HCV-Induced Hepatocellular Carcinoma: Clinical Significance and Prognostic Value. Biologics. 2023, 17, 137-149. [CrossRef]
- Hajjari, M. and Salavaty,A. HOTAIR: an oncogenic long non-coding RNA in different cancers. Cancer Biol. Med. 2015, 12, 1-9.
- Ishibashi, M.; Kogo,R.; Shibata,K.; Sawada,G.; Takahashi,Y.; Kurashige,J.; Akiyoshi,S.; Sasaki,S.; Iwaya,T.; Sudo,T.; Sugimachi,K.; Mimori,K.; Wakabayashi,G.; Mori,M. Clinical significance of the expression of long non-coding RNA HOTAIR in primary hepatocellular carcinoma. Oncol. Rep. 2013, 29, 946-950. [CrossRef]
- Han, C.; Yang,Y.; Guo,L.; Guan,Q.; Ruan,S. The expression of long non-coding RNA HOTAIR in advanced hepatocellular carcinoma and its prognostic correlation with sunitinib therapy. Arch. Med. Sci. 2022, 18, 71-78. [CrossRef]
- Lou, Z. H.; Xu,K.Y.; Qiao,L.; Su,X.Q.; Ou-Yang,Y.; Miao,L.B.; Liu,F.; Wang,Y.; Fu,A.; Ren,X.H.; Huang,J.S.; Li,Z.Y.; Bao,J.F. Diagnostic Potential of the Serum lncRNAs HOTAIR, BRM and ICR for Hepatocellular Carcinoma. Front Biosci. (Landmark. Ed). 2022, 27, 264. [CrossRef]
- Wen, P.; Qi,X.; Zheng,R. Value of the HOTAIR expression assay in predicting therapy target in hepatocellular carcinoma: A meta-analysis and bioinformatics analysis. Int. J Biol. Markers. 2024, 39, 239-247. [CrossRef]
- Yu, T.; Wang,Y.; Fan,Y.; Fang,N.; Wang,T.; Xu,T.; Shu,Y. CircRNAs in cancer metabolism: a review. J Hematol. Oncol. 2019, 12, 90. [CrossRef]
- Zhang, X.; Zhou,H.; Jing,W.; Luo,P.; Qiu,S.; Liu,X.; Zhu,M.; Liang,C.; Yu,M.; Tu,J. The Circular RNA hsa_circ_0001445 Regulates the Proliferation and Migration of Hepatocellular Carcinoma and May Serve as a Diagnostic Biomarker. Dis. Markers. 2018, 2018, 3073467. [CrossRef]
- Xu, Q.; Zhou,L.; Yang,G.; Meng,F.; Wan,Y.; Wang,L.; Zhang,L. Overexpression of circ_0001445 decelerates hepatocellular carcinoma progression by regulating miR-942-5p/ALX4 axis. Biotechnol. Lett. 2020, 42, 2735-2747. [CrossRef]
- Shi, Y.; Sun,X.; He,X. Overexpression of Aristaless-Like Homeobox-4 Inhibits Proliferation, Invasion, and EMT in Hepatocellular Carcinoma Cells. Oncol. Res. 2017, 25, 11-18. [CrossRef]
- Wang, P.; Xu,L.L.; Zheng,X.B.; Hu,Y.T.; Zhang,J.F.; Ren,S.S.; Hao,X.Y.; Li,L.; Zhang,M.; Xu,M.Q. Correlation between the expressions of circular RNAs in peripheral venous blood and clinicopathological features in hepatocellular carcinoma. Ann. Transl. Med. 2020, 8, 338. [CrossRef]
- Chang, Z.; Song,Y.; Luo,F.; Yang,X.; Cai,Y.; Guo,H. Circular RNA SMARCA5 Promotes a Poor Prognosis and Radiotherapy Resistance for Patients with Hepatocellular Carcinoma. Ann. Clin. Lab Sci. 2023, 53, 573-577.
- Liu, T.; Song,Z.; Gai,Y. Circular RNA circ_0001649 acts as a prognostic biomarker and inhibits NSCLC progression via sponging miR-331-3p and miR-338-5p. Biochem. Biophys. Res. Commun. 2018, 503, 1503-1509. [CrossRef]
- Qin, M.; Liu,G.; Huo,X.; Tao,X.; Sun,X.; Ge,Z.; Yang,J.; Fan,J.; Liu,L.; Qin,W. Hsa_circ_0001649: A circular RNA and potential novel biomarker for hepatocellular carcinoma. Cancer Biomark. 2016, 16, 161-169. [CrossRef]
- Zhang, X.; Qiu,S.; Luo,P.; Zhou,H.; Jing,W.; Liang,C.; Tu,J. Down-regulation of hsa_circ_0001649 in hepatocellular carcinoma predicts a poor prognosis. Cancer Biomark. 2018, 22, 135-142. [CrossRef]
- Fu, L.; Wu,S.; Yao,T.; Chen,Q.; Xie,Y.; Ying,S.; Chen,Z.; Xiao,B.; Hu,Y. Decreased expression of hsa_circ_0003570 in hepatocellular carcinoma and its clinical significance. J Clin. Lab Anal. 2018, 32. [CrossRef]
- Zhang, X.; Chen,W.; Guo,D.; Li,Y.; Zhao,Y.; Ren,M.; Lu,G.; Lu,X.; He,S. Circ_0003570 Suppresses the progression of hepatocellular carcinoma through miR-182-5p/STARD13 regulatory axis. Biol. Proced. Online. 2022, 24, 14. [CrossRef]
- Jang, S. Y.; Kim,G.; Tak,W.Y.; Kweon,Y.O.; Lee,Y.R.; Han,Y.S.; Han,J.R.; Park,J.G.; Kang,M.K.; Lee,H.W.; Lee,W.K.; Park,S.Y.; Hur,K. Circular Noncoding RNA hsa_circ_0003570 as a Prognostic Biomarker for Hepatocellular Carcinoma. Genes (Basel). 2022, 13. [CrossRef]
- Kang, M. K.; Kim,G.; Park,J.G.; Jang,S.Y.; Lee,H.W.; Tak,W.Y.; Kweon,Y.O.; Park,S.Y.; Lee,Y.R.; Hur,K. Tissue Circular RNA_0004018 and 0003570 as Novel Prognostic Biomarkers for Hepatitis B-Related Hepatocellular Carcinoma. Genes (Basel). 2023, 14. [CrossRef]
- Jiang, Z.; Shen,L.; Wang,S.; Wu,S.; Hu,Y.; Guo,J.; Fu,L. Hsa_circ_0028502 and hsa_circ_0076251 are potential novel biomarkers for hepatocellular carcinoma. Cancer Med. 2019, 8, 7278-7287. [CrossRef]
- Liu, H.; Lan,T.; Li,H.; Xu,L.; Chen,X.; Liao,H.; Chen,X.; Du,J.; Cai,Y.; Wang,J.; Li,X.; Huang,J.; Yuan,K.; Zeng,Y. Circular RNA circDLC1 inhibits MMP1-mediated liver cancer progression via interaction with HuR. Theranostics. 2021, 11, 1396-1411. [CrossRef]
- Zhang, T.; Jing,B.; Bai,Y.; Zhang,Y.; Yu,H. Circular RNA circTMEM45A Acts as the Sponge of MicroRNA-665 to Promote Hepatocellular Carcinoma Progression. Mol. Ther. Nucleic Acids. 2020, 22, 285-297. [CrossRef]
- Martinez-Quetglas, I.; Pinyol,R.; Dauch,D.; Torrecilla,S.; Tovar,V.; Moeini,A.; Alsinet,C.; Portela,A.; Rodriguez-Carunchio,L.; Sole,M.; Lujambio,A.; Villanueva,A.; Thung,S.; Esteller,M.; Zender,L.; Llovet,J.M. IGF2 Is Up-regulated by Epigenetic Mechanisms in Hepatocellular Carcinomas and Is an Actionable Oncogene Product in Experimental Models. Gastroenterology. 2016, 151, 1192-1205. [CrossRef]
- Chen, D.; Zhang,C.; Lin,J.; Song,X.; Wang,H. Screening differential circular RNA expression profiles reveal that hsa_circ_0128298 is a biomarker in the diagnosis and prognosis of hepatocellular carcinoma. Cancer Manag. Res. 2018, 10, 1275-1283. [CrossRef]
Figure 1.
miRNA biogenesis and functions. In the cell nucleus, a long precursor named pri-miRNA is processed by the enzyme Drosha to pre-miRNA; this in turn is exported to the cytoplasm by the Exportin 5 enzyme; here, the DICER enzyme produces the double stranded mature RNA (miRNA). The miRNA antisense strand is then loaded onto RISC allowing the recognition of the target mRNA resulting in the translation inhibition (via an imperfect base pairing) or in mRNA degradation (via perfect base pairing). Recent findings indicate that via direct/indirect mechanisms, miRNA can also promote gene expression. This figure has been created with BioRender.com.
Figure 1.
miRNA biogenesis and functions. In the cell nucleus, a long precursor named pri-miRNA is processed by the enzyme Drosha to pre-miRNA; this in turn is exported to the cytoplasm by the Exportin 5 enzyme; here, the DICER enzyme produces the double stranded mature RNA (miRNA). The miRNA antisense strand is then loaded onto RISC allowing the recognition of the target mRNA resulting in the translation inhibition (via an imperfect base pairing) or in mRNA degradation (via perfect base pairing). Recent findings indicate that via direct/indirect mechanisms, miRNA can also promote gene expression. This figure has been created with BioRender.com.
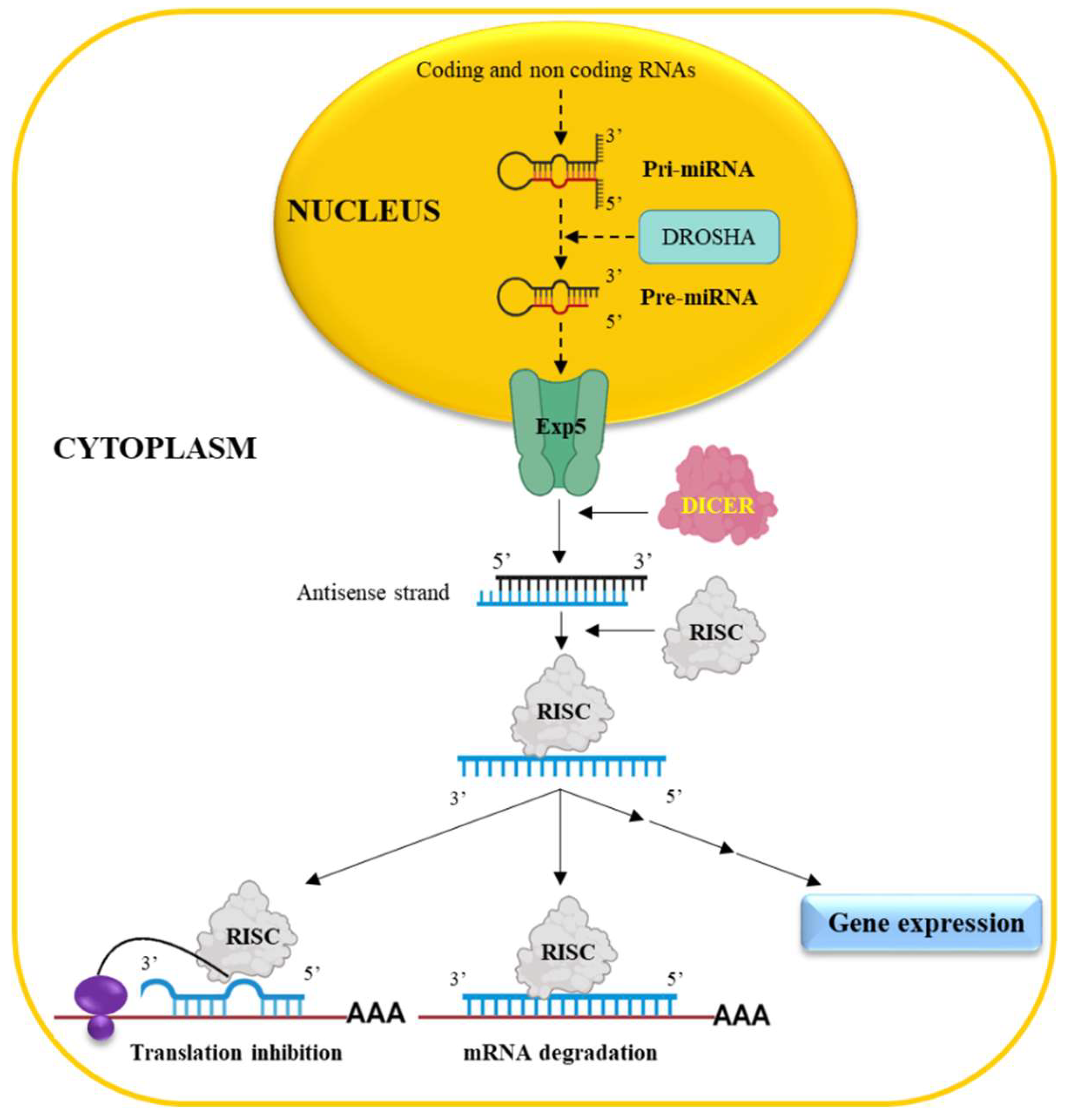
Figure 2.
LncRNAs biogenesis and functions. LncRNAs can be spliced, capped and poly-adenylated. Following transcription, they assume a three-dimensional (3D) structure responsible for the biological effects such as the recruitment of transcription activators/repressors to the promoters of their target genes thus regulating gene expression, the inhibition of miRNA activity via the sponging effect, the ability to act as scaffolds for protein for the formation of protein complexes. This figure has been created with BioRender.com.
Figure 2.
LncRNAs biogenesis and functions. LncRNAs can be spliced, capped and poly-adenylated. Following transcription, they assume a three-dimensional (3D) structure responsible for the biological effects such as the recruitment of transcription activators/repressors to the promoters of their target genes thus regulating gene expression, the inhibition of miRNA activity via the sponging effect, the ability to act as scaffolds for protein for the formation of protein complexes. This figure has been created with BioRender.com.
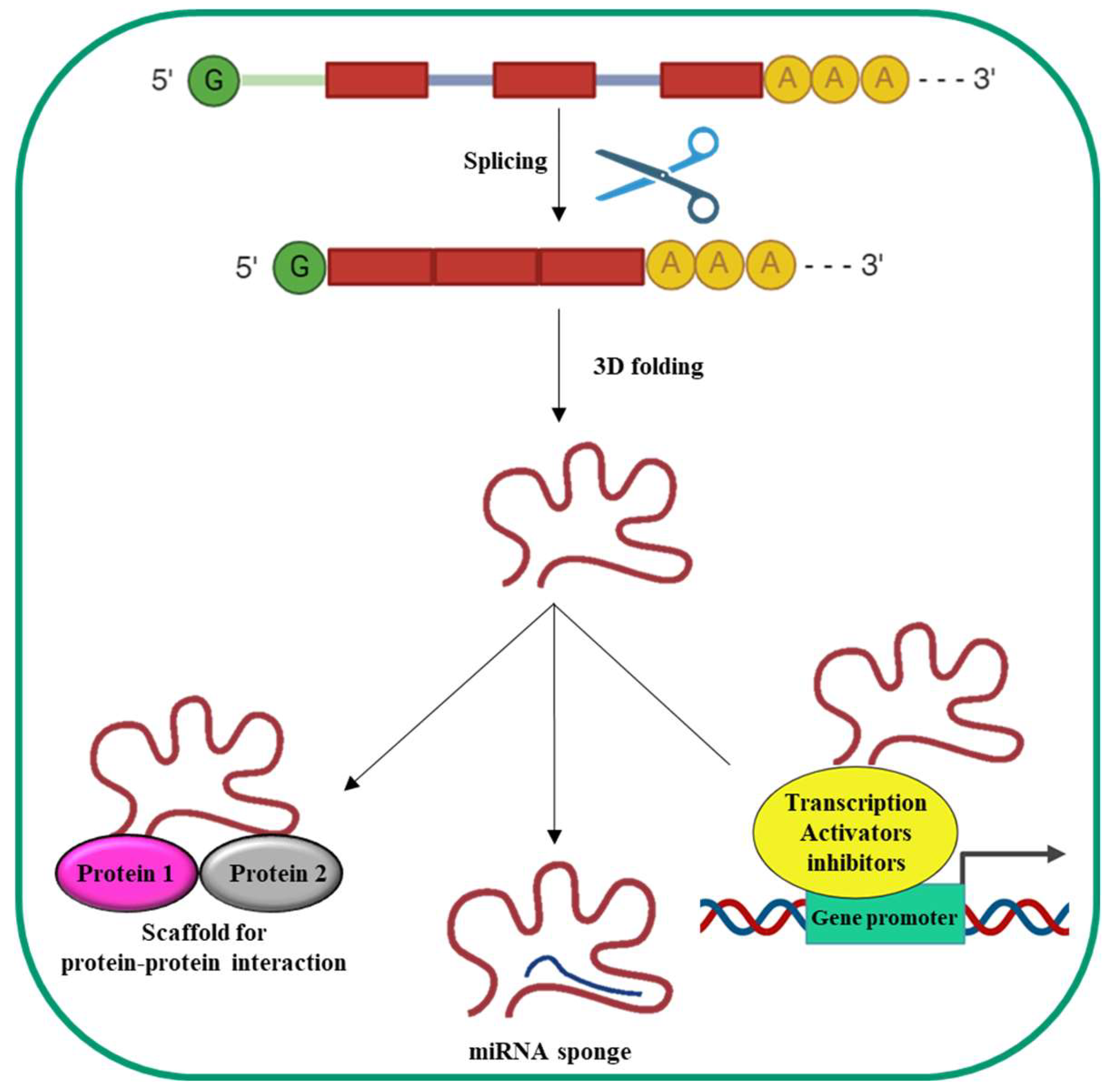
Figure 3.
CircRNAs biogenesis and functions. Although circRNAs have multiple biogenesis mechanisms, a common one is represented by back-splicing. Back-splicing can be induced by protein dimerization, sequence complementarity of flanking introns and exon skipping mechanisms. Following the formation of a circular RNA, circRNA is exported into the cytoplasm, where it can bind miRNAs via complementary regions, interact with specific mRNAs, undergo translation to generate small peptides, interact with proteins and interact with gene promoters. This figure has been created with BioRender.com.
Figure 3.
CircRNAs biogenesis and functions. Although circRNAs have multiple biogenesis mechanisms, a common one is represented by back-splicing. Back-splicing can be induced by protein dimerization, sequence complementarity of flanking introns and exon skipping mechanisms. Following the formation of a circular RNA, circRNA is exported into the cytoplasm, where it can bind miRNAs via complementary regions, interact with specific mRNAs, undergo translation to generate small peptides, interact with proteins and interact with gene promoters. This figure has been created with BioRender.com.
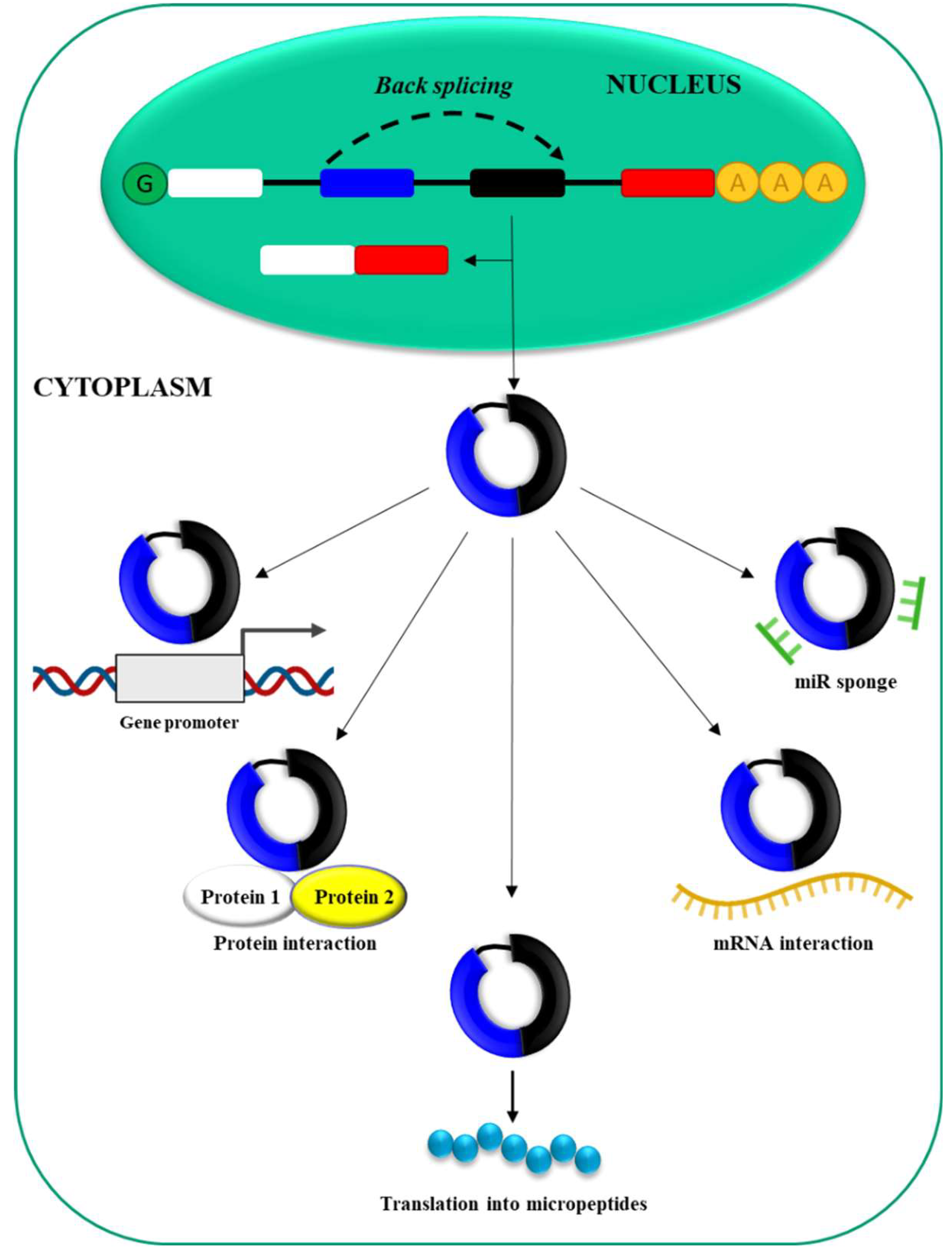
Figure 4.
Extracellular vesicles. Extracellular vesicles (EVs) are a heterogeneous group of lipid bilayer particles, synthesized and secreted by different cell types into the extracellular environment. EVs encapsulate a variety of bioactive molecules such as proteins, lipids and nucleic acids. Nowadays it is known that EVs can contain ncRNAs and can deliver these molecules to distant cells both under physiological and pathological conditions. This figure has been created with BioRender.com.
Figure 4.
Extracellular vesicles. Extracellular vesicles (EVs) are a heterogeneous group of lipid bilayer particles, synthesized and secreted by different cell types into the extracellular environment. EVs encapsulate a variety of bioactive molecules such as proteins, lipids and nucleic acids. Nowadays it is known that EVs can contain ncRNAs and can deliver these molecules to distant cells both under physiological and pathological conditions. This figure has been created with BioRender.com.
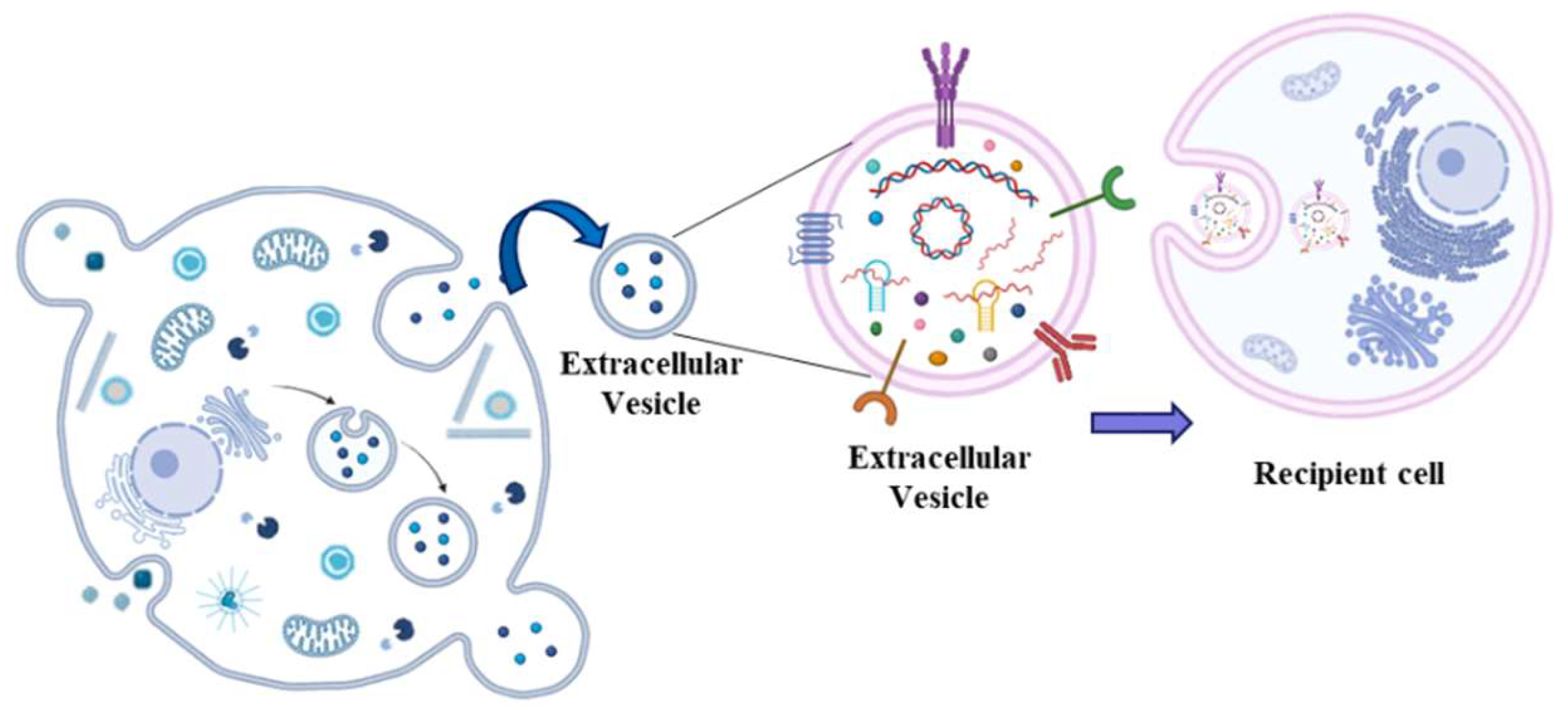

Table 1.
miRNAs considered as potential biomarkers for hepatocellular carcinoma.
| miRNA | Prognostic significance | Levels in HCC | Ref |
|---|---|---|---|
| miR-221 | Poor | Increased (tissue) Decreased (serum) |
[74,75,76,77] |
| miR-18-a | Poor | Increased (tissue) | [74] |
| miR-487a | Poor | Increased (tissue) | [80] |
| miR-33a | Poor/Good | Reduced (tissue) | [87,88] |
| miR-105-1 | Poor | Reduced (tissue) | [93,94] |
| miR-138-5p | Poor | Reduced (tissue) | [96,97,98] |
| miR-21 | Poor | Increased (Tissue/serum) | [101, 102,103, 104] |
| miR-122 | Poor | Decreased (tissue, serum) | [108,109] |
| miR-139-5p | Poor | Decreased (tissue) | [116,117,118] |
Table 2.
lncRNAs considered as potential biomarkers for hepatocellular carcinoma.
| lncRNA | Prognostic significance | Levels in HCC | References |
|---|---|---|---|
| HULC | Poor | Increased (serum, tissue) |
[119, 121, 122, 123, 128] |
| MALAT1 | Poor | Increased (serum/tissue) | [126,127] |
| Linc00152 | Poor | Increased (plasma, tissue) |
[128,130,131] |
| HOTAIR | Poor | Increased (tissue, serum) |
[133, 134, 135, 136] |
Table 3.
circRNAs considered as potential biomarkers for hepatocellular carcinoma.
| circRNA | Prognostic significance | Levels in HCC | References |
|---|---|---|---|
| 0001445 | Poor | Decreased (tissue, plasma) Increased (tissue) |
[138,139,141] [142] |
| 0001649 | Poor | Decreased (tissue) | [144,145] |
| 0003570 | Poor | Decreased (tissue) | [146,147,149] |
| 0028502/0076251 | Poor | Decreased (tissue) |
[150] |
| CircDLC1 | Poor | Decreased (tissue) |
[151] |
| 0066659 | Poor | Increased (tissue) |
[152] |
| 0128298 | Poor | Increased (tissue) |
[154] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



