
Submitted:
06 October 2024
Posted:
07 October 2024
You are already at the latest version
Abstract
In biology and medicine, synthetic polymers that mimic intrinsically disordered proteins (IDPs) exhibiting remarkable structural and conformational flexibility due to their lack of stable three-dimensional structures are highly desirable. This conformational flexibility facilitates self-organization, making such polymers highly useful for various biomedical applications. Specifically, intrinsically disordered synthetic polymers show great potential in drug delivery systems, organ transplantation, artificial organ design, and ensuring immune compatibility. There is a pressing need for new synthesis and characterization techniques to develop these polymers which are modeled after intrinsically disordered proteins and can be used for various biomedical purposes. In this paper, we outline our strategies for designing intrinsically disordered synthetic polymers for biomedical applications. These design approaches are inspired by the properties of intrinsically disordered proteins and use of disorder promoting oligopeptides.
Keywords:
IDPols
; intrinsically disordered proteins
; biomedical applications
; synthesis
; characterization
; disorder-promoting peptides
1. Introduction
Intrinsically disordered proteins (IDPs) are a unique class of proteins that unlike traditional proteins, do not have a stable or fixed three-dimensional structure in solution. Instead, they exist in a flexible, dynamic ensemble of conformations [1]. IDPs play critical roles in various biological processes, such as cell signaling, interactions with other proteins, DNA, or small molecules, and regulation [2]. Their flexibility allows them to bind multiple partners and participate in diverse cellular processes [3]. The lack of a rigid structure provides IDPs with the ability to adapt their shape to different binding partners and conditions, facilitating multiple interactions and functions [4]. Because they can change conformations easily, IDPs are often involved in regulatory processes that require rapid and reversible interactions. This makes them integral to signal transduction pathways [5]. IDPs are often subject to various post-translational modifications, such as phosphorylation, glycosylation, acetylation, and methylation, which can further modulate their function and interactions [6]. Mutations or dysregulations of IDPs are implicated in various diseases, such as cancer, neurodegenerative disorders, and cardiovascular diseases [7,8]. Their dynamic nature sometimes makes IDPs difficult to target with traditional drugs, presenting challenges and opportunities for therapeutic development [9].
Disorder-promoting amino acid residues refer to specific amino acids that are more likely to be found in intrinsically disordered regions of proteins [10]. Unlike structured regions of proteins, which have a defined three-dimensional shape, intrinsically disordered regions are flexible and lack a stable structure. Amino acid residues that are commonly considered disorder-promoting include proline, glycine, serine, glutamine, lysine, arginine, aspartic acid, and glutamic acid [11,12,13]. Proline causes kinks in the peptide chain due to its rigid structure and can disrupt regular secondary structures like α-helix and β-sheet. Glycine is highly flexible and lacks a side chain, giving it a high degree of conformational freedom which contributes to disorder. Serine can be engaged in hydrogen bonding, contributing to flexibility. Glutamine can form hydrogen bonds through the amide group, contributing to flexibility and disorder. The positive charge of lysine can interact with various partners, which can lead to a lack of a stable structure. Similar to lysine, arginine’s positive charge and ability to form multiple hydrogen bonds contribute to disorder. The negative charges of aspartic acid and glutamic acid can lead to repulsive interactions and destabilize ordered structures. These amino acids, when present in high concentrations within a protein sequence, increase the likelihood that the region will be intrinsically disordered. This intrinsic disorder can be essential for the protein’s function, allowing it to interact with multiple partners or change conformation easily in response to different cellular signals.
We introduced recently intrinsically disordered polymers (IDPols) bio-mimicking intrinsically disordered proteins (IDPs) by using structure-breaking amino acid residues [14,15]. In general, IDPols refer to polymers that lack a fixed or ordered three-dimensional structure in at least part of their sequence under physiological conditions [14]. Due to their lack of stable secondary and tertiary structures, IDPols are highly flexible and can easily change shape. IDPols can adopt multiple conformations depending on the environment, interactions, and presence of binding partners. Synthetic IDPols can be engineered for applications in materials science, including the design of flexible, biocompatible materials for medical applications like drug delivery systems, tissue engineering scaffolds, and responsive hydrogels [14]. The intrinsic disorder in these polymers provides them with unique properties that are not achievable by their well-ordered counterparts. This makes IDPs exceptionally versatile and useful in biomedical applications.
IDPols can be designed to respond to physiological conditions, such as pH or temperature, making them ideal for controlled drug delivery [14,16]. For instance, elastin-like polypeptides (ELPs) are a type of IDPols that was succesfully engineered to undergo phase transitions in response to temperature changes [17]. ELP-based drug delivery systems have been developed to release therapeutic agents at specific target sites, improving drug efficacy and reducing side effects [18]. Furthermore, IDPols mimic the natural extracellular matrix (ECM), providing a supportive environment for cell growth and tissue formation [19]. Silk-elastin-like polypeptides (SELPs) are used in the creation of hydrogels for tissue scaffolding [20]. These biomaterials can be tailored to support the growth and differentiation of cells, aiding in the regeneration of tissues, such as cartilage, bone, and skin. Additionally, IDPols’ ability to undergo conformational changes in response to specific stimuli can be harnessed for the development of sensitive biosensors [21]. IDPols can be in fact used to create biosensors that detect biomarkers associated with diseases [22]. For instance, IDPols have been incorporated into sensor platforms to identify cancer biomarkers or detect changes in glucose levels for diabetes management [23,24]. Furthermore, IDPols can be engineered to form bioadhesive materials that promote healing and provide structural support [25]. For example, mussel-inspired adhesives often utilize IDPols to replicate the adhesive properties of mussel foot proteins [26]. These adhesives can be used for wound closure applications, providing a biocompatible and strong bonding material that supports tissue repair. In addition, IDPol-based hydrogels can exhibit unique mechanical cues, making them suitable for various biomedical applications. In fact, hydrogels formed from IDPols such as resilin-like polypeptides (RLPs) show exceptional resilience and elasticity [27]. Such properties are advantageous in applications requiring dynamic mechanical performance, such as in soft tissue replacements or as injectable fillers for minimally invasive surgeries. In general, IDPols can be designed to change their properties in response to specific biological signals, enabling the creation of smart biomaterials. Specifically, IDPols that undergo conformational changes upon binding to specific ions or small molecules can be used to create smart implants or sensors that monitor and respond to physiological changes in real-time, providing feedback or releasing therapeutics in response to specific conditions [14]. In addition, IDPols can be functionalized for use in nanoparticle-based diagnostics and therapeutics [28]. IDPols attached to nanoparticles can be engineered to target specific tissues or cellular environments. This targeting mechanism is useful in molecular imaging, where IDPol-functionalized nanoparticles can help in the precise visualization of cancerous tissues or other pathological conditions [29].
2. Biological Functions of Disordered Peptides
Disorder-promoting residues play significant roles in various biological processes due to their flexible and dynamic nature [30]. Disorder-promoting residues often participate in transient interactions and can bind multiple targets via short linear motifs (SLiMs) or modular proteins, allowing them to act as versatile binding partners [30]. For instance, the p53 tumor suppressor protein contains disorder-promoting residues that facilitate its interaction with numerous protein partners involved in DNA repair, cell cycle regulation, and apoptosis [31]. The flexible nature of disordered peptides allows for the rapid modulation of interactions and activities, which is crucial for signal transduction and regulatory mechanisms. For instance, the transcription factor CREB (cAMP Response Element-Binding protein) has disorder-promoting residues that mediate interactions with other proteins and enable the regulation of gene expression in response to cellular signals [32]. Disordered peptides can function as flexible linkers connecting structured domains, facilitating the proper positioning and dynamics regulation of enzymatic and structural activities. The protein tau, which is associated with microtubule assembly in neurons, contains disordered regions that allow it to serve as a flexible spacer and linker [33]. However, its dysregulation and aggregation are linked to neurodegenerative disorders such as Alzheimer’s disease [34]. Furthermore, disordered peptides can act as molecular chaperones, assisting in the proper folding of other proteins and preventing aggregation [35]. For instance, the small heat shock protein Hsp27 has disorder-promoting residues that are crucial for its function as a molecular chaperone, helping to prevent the aggregation of other proteins under stress conditions [36]. Disordered peptides can serve as dynamic scaffolds, assembling and disassembling protein complexes in response to cellular conditions. The protein BRCA1, involved in DNA damage repair, has disorder-promoting residues that act as a scaffold to recruit various repair proteins to sites of DNA damage [37]. Disorder-promoting residues are often involved in the formation of biomolecular condensates through liquid-liquid phase separation, contributing to the organization of cellular compartments without membranes [38]. For instance, the RNA-binding protein FUS contains a sequence motif responsible for phase separation, enabling it to participate in the assembly of stress granules and nuclear bodies – a process crucial for cellular stress responses and the regulation of RNA metabolism [39]. Furthermore, the conformational versatility of disordered peptides enables them to participate in various dynamic processes, facilitating complex signaling networks and cellular responses. For example, eukaryotic translation initiation factor 4E-binding proteins (4E-BPs) use disorder-promoting residues to inhibit translation by binding to eIF4E, and their function is regulated through phosphorylation-induced conformational changes [40].
The therapeutic effects of disordered peptides containing disorder-promoting residues can be linked to their unique ability to engage in interactions that structured proteins cannot, providing advantages in specific biological contexts. As mentioned above, for example, the tumor suppressor protein p53 has intrinsically disordered regions that are crucial for its function in regulating cell cycle and apoptosis. Modulating the function of p53 with synthetic peptides can restore its normal function in cancer cells where it is mutated or inactivated. As we have shown previously, α-synuclein is a protein associated with Parkinson’s disease and has disordered regions that are crucial for its aggregation [41,42,43]. Peptides designed for interfering with α-synuclein aggregation can potentially serve as therapeutic agents to prevent or treat Parkinson’s disease. Furthermore, many antimicrobial peptides (AMPs) are intrinsically disordered and can adopt multiple conformations upon interaction with microbial membranes [44], enhancing their ability to disrupt these membranes and kill pathogens. Furthermore, for instance, apelin is an intrinsically disordered peptide that interacts with its receptor (APJ) and it has been shown to have beneficial effects on cardiovascular function [45]. Therapeutic peptides mimicking apelin can be used to treat heart failure and other cardiovascular diseases. Peptides derived from the Bcl-2 family of proteins can influence apoptotic pathways [46]. These peptides interact with intrinsically disordered regions of pro-apoptotic proteins, modulating cell death processes, which is crucial in cancer therapy. All in all, disorder promoting peptides hold significant therapeutic potential due to their flexibility and their ability to engage in dynamic interactions that structured proteins may not facilitate. Their roles in various diseases, from cancer to neurodegenerative and cardiovascular conditions, underline their importance in developing novel therapeutic strategies.
Recently, we have shown that the artificial intelligence (AI)-based structure prediction tool C-I-TASSER outperforms AlphaFold2, AlphaFold3, RoseTTAfold, and ESMFold in generating the structures of small-size intrinsically disordered peptides [47]. Specifically, C-I-TASSER (Contact-guided Iterative Threading ASSEmbly Refinement) is a computational method used for protein structure prediction [48]. It builds on the principles of the original I-TASSER but incorporates additional information from predicted contact maps to improve the accuracy of the predicted structures. Table 1 presents a list of disordered oligopeptides along with their predicted structures and biological functions generated using C-I-TASSER.
3. Synthesis of IDPols for Biomedical Applications using Disordered Peptides Containing Disorder-Promoting Residues
We recently designed the synthesis, characterization, and production protocols for the IDPols bio-mimicking the intrinsically disordered proteins using structure-breaking residues for soft robotics and biomedical applications [14,15]. Here, we develop an approach for synthesizing intrinsically disordered synthetic polymers using disorder promoting peptides for biomedical applications. In these approaches, biocompatibility and self-healing are considered. A sequence determines the function of a protein and we used ten disorder-promoting amino acid residues (Table 1). Here, we provide as an example the oligopeptide sequence GPSQKRDEEE.
3.1. Modification of the Chain Extender with a Disorder-Promoting Oligopeptide
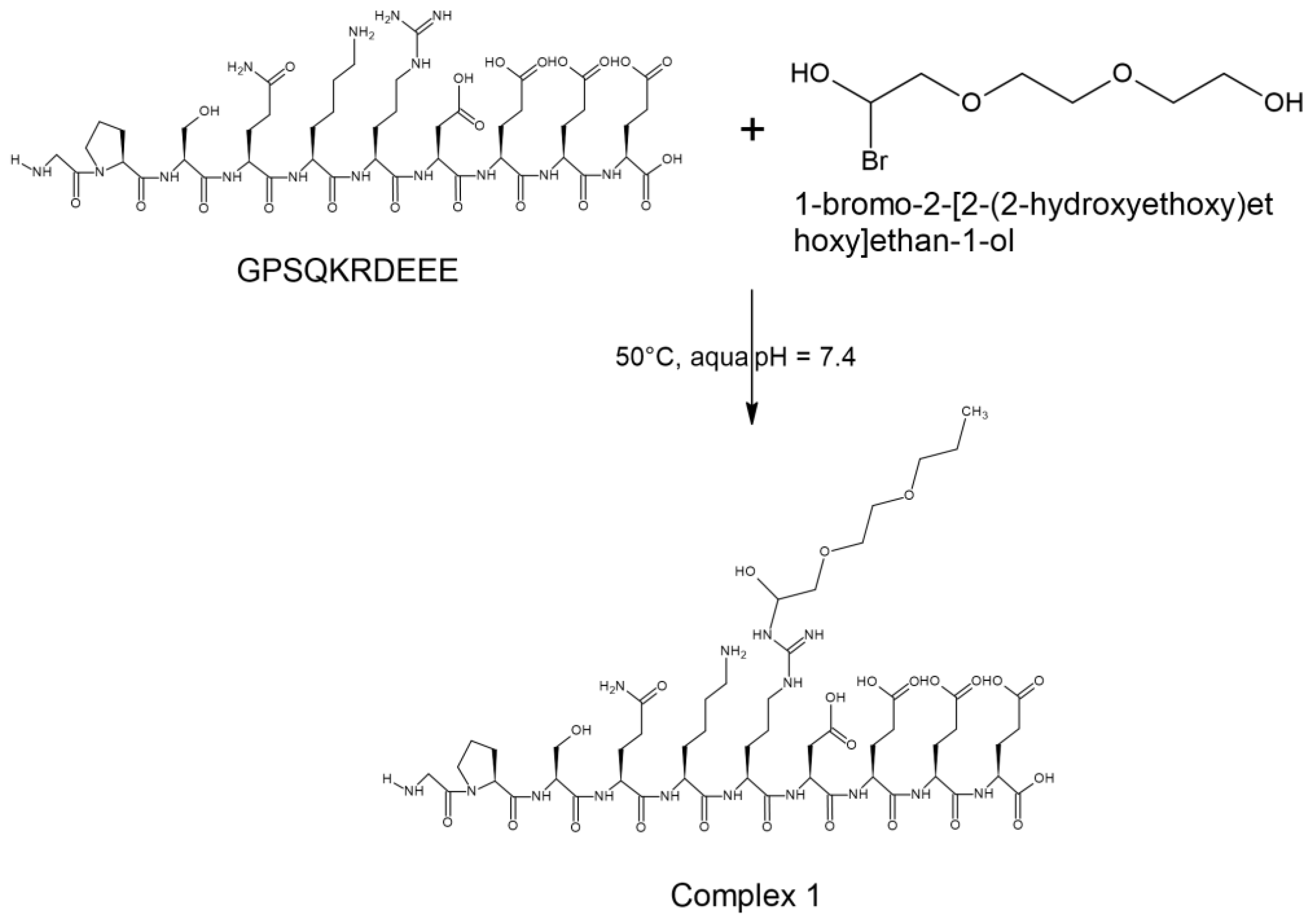
The approach involves the modification of the chain extender with a disorder-promoting oligopeptide. The reaction of the example GPSQKRDEEE oligopeptide with monobromo triethylene glycol yields a complex (Scheme 1).
This newly formed complex acts as a new type of chain extender and the modification takes only one step. The disordered oligopeptide is mixed with distilled water until it is completely dissolved (Scheme 1). After complete dissolution, monobrome triethylene glycol is added and the formed complex is dialyzed against ultrapure water for a few days using a dialysis membrane with appropriate pore size to remove the impurities from the medium. We carry out a protection reaction using tert-butyl hydrogen carbonate (BOC) for protecting the amine groups.
3.2. Synthesis of the Prepolymer and Polymer
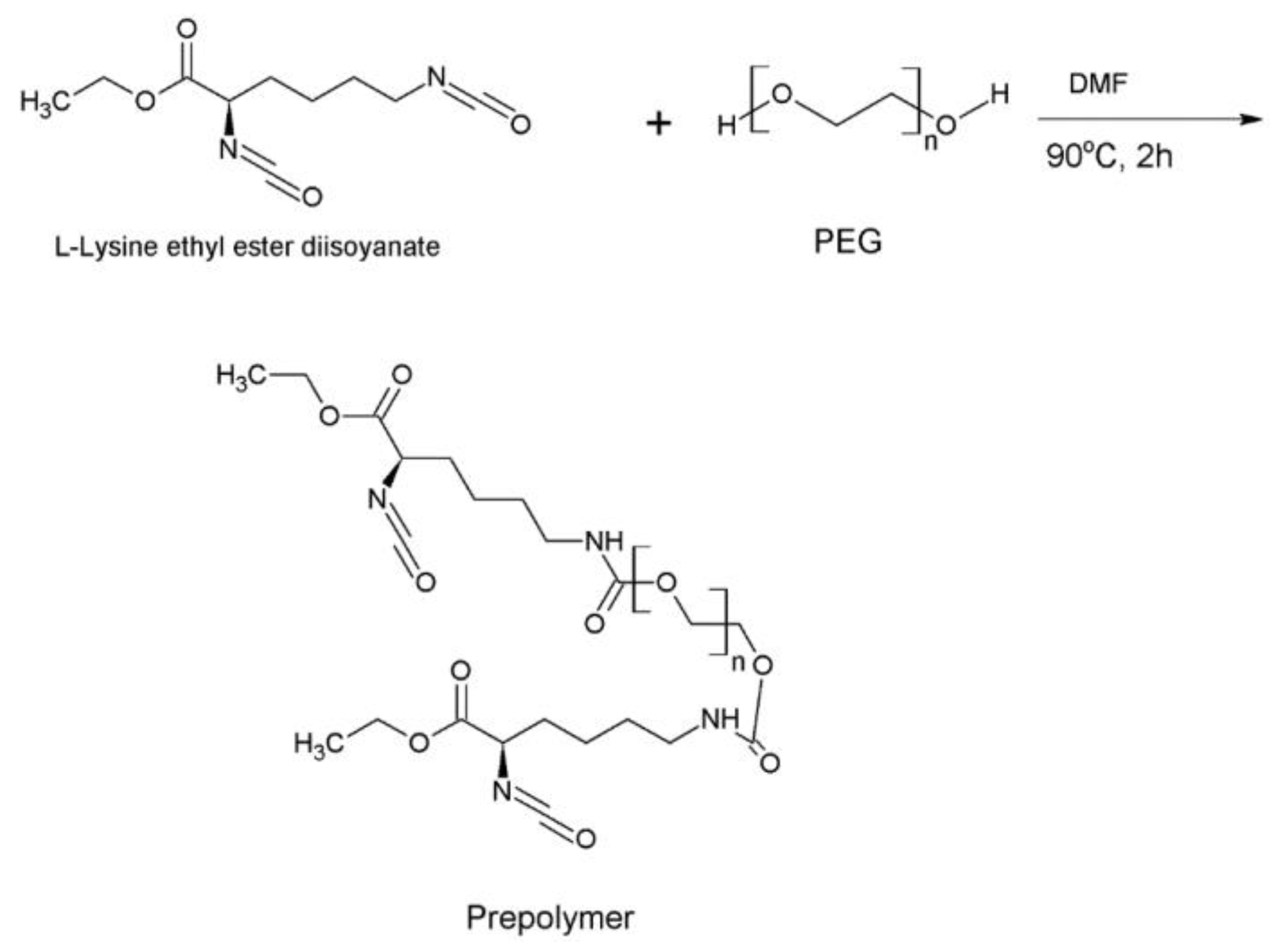
After we obtain the new chain extender, we start preparing the prepolymer. To produce the polymers, a controllable reaction system is important. Therefore, we prefer the two-step polyaddition reaction. However, we should mention here, that our goal is to obtain a biodegradable polymer, and we have to ensure that the polymer does not have a toxic effect after its degradation. Therefore, we choose the non-toxic L-lysine ethyl ester diisocyanate as the hard segment and poly(ethylene glycol) as our soft segment.
We mix L-lysine ethyl ester diisocyanate (2 eq.) and PEG (1 eq.) in a DMF solution under constant stirring at 400 rpm (Scheme 2). Next, the new chain extender, which we call Complex 1, is added to the solution with the prepolymer at a ratio of 1 molar under continuous mixing in an inert atmosphere (Schem 3). After the addition takes place, the reaction continues for 3 h at constant temperature. The formed viscous solution is then dried using a vacuum oven for 36 h. All reactions are carried out under an inert atmosphere and the product is obtained by extracting the polymer in a chloroform solution.
4. Characterization and Production of IDPols
Rheological analyses are essential for determining the flow properties of polymers prior to their production [49]. These analyses are performed using a rotational rheometer equipped with parallel plate geometry, specifically a 25 mm diameter plate. To assess the polymers' flow behavior, a stress-strain sweep test is conducted at a constant strain of 1%, covering a frequency range of 0.1 to 100 rad/s, both at room temperature and elevated temperatures. Rheological testing also plays a key role in investigating self-healing properties, where recovery characteristics are measured [50]. For this analysis, an oscillation-time sweep test is conducted with strain variations ranging from 250% to 0.5%.
When the materials reach the appropriate condition for extrusion, the polymers are granulated to a diameter of 4 mm ± 1 mm at room temperature. Before loading them into the extruder, a pre-drying process is required, which is carried out for 2 to 4 hours in air circulation ovens operating at 90–100 °C. To produce the filaments, a twin-screw extruder with a cylindrical nozzle (ø 2.9 mm) is used, applying specific pressure at the polymers' designated Tm values.
It is also important to evaluate the mechanical properties post-healing. For this, the materials are cut using a razor and then rejoined, being kept in dark conditions for 24 hours. Following this period, the aforementioned mechanical tests are conducted on the specimens to assess their properties after healing. Additionally, dynamic thermomechanical analysis (DMA) tests are crucial for examining the thermomechanical behavior of 3D-printed polymers [51]. These tests will be performed using a DMA analyzer while gradually lowering the temperature. The glass transition temperature (Tg) of the specimens will be determined based on the peak of tan δ.
The drug release rate can be assessed by measuring the amount of drug released over time [52]. This can be achieved using various analytical techniques, such as UV-Vis spectroscopy, high-performance liquid chromatography (HPLC), or mass spectrometry.
To investigate the drug release behavior of self-healing polymers, a specific drug is chosen and encapsulated within the polymer. The total drug content is calculated based on the sample weight and the predicted weight ratio. These drug-encapsulated self-healing polymers are then immersed in a phosphate buffered saline (PBS) solution maintained at 37°C to monitor their drug-release behavior. At predetermined time intervals, a certain volume of solution is withdrawn from the PBS medium, and an equal volume of fresh PBS is added back. The withdrawn solution is analyzed using a UV-Vis spectrophotometer at the drug's characteristic absorption wavelength. Prior to these measurements, a calibration curve of absorbance versus concentration is established, ranging from 5 to 100 ppm. This calibration curve, represented by the equation y = mx + n (where y is the absorbance and x is the drug concentration in ppm), is used to determine cumulative drug release curves. The experiments are conducted in triplicate to ensure reliability. Drug release from a polymer matrix typically involves the processes of water penetration, hydration, swelling, diffusion of the dissolved drug, and erosion of the gelatinous layer. It's important to note that the mechanism of drug release is influenced by the drug dose, the pH of the solution, and the specific characteristics of both the polymer and the drug. The degree of swelling of the polymer matrix is directly related to the amount of drug released.
Cytotoxicity occurs when cells are negatively impacted by chemical substances or the physical characteristics of their environment [53]. To evaluate the biocompatibility of a biomaterial, it's crucial to determine its potential toxic effects on cells. A cytotoxicity test is used to assess whether self-healing polymers are harmful to cells. For this evaluation, the self-healing polymer is usually sterilized and prepared in an implantable form, such as disks or fibers. To evaluate the cytotoxicity of the self-healing polymer, an indirect MTT test is conducted. First, 2 mL of MEM Alpha medium is added to thoroughly dry samples, which are then incubated at 37°C with 5% CO2 for 24 hours. The medium containing the incubated polymer extracts is sterilized using a 0.22 µm syringe filter. Osteoblasts are seeded at 20,000 cells per well with a specific amount of MEM Alpha in a 48-well plate and incubated at 37°C with 5% CO2 for 24 hours. The osteoblast cells are then exposed to a culture medium containing the biopolymer extracts and incubated for another 24 hours. Subsequently, specific amounts of MEM Alpha and MTT solution prepared in PBS are added to each well, followed by an additional 4-hour incubation to allow formazan crystal formation. The solution is then removed, and a defined amount of DMSO is added to each well to dissolve the crystals. After a 20-minute incubation, the formazan solution is read at 570 nm using a microplate reader. Wells containing only MEM Alpha medium serve as the control samples.
The biodegradability of IDPols can be evaluated by measuring their degradation [54]. These bio-mimicked polymers are designed to imitate intrinsically disordered proteins that enzymes in the body can break down. Enzymatic degradation assays are used to determine the extent of biodegradation of these polymers. For a detailed degradation study, a solution containing collagenase type II in DPBS is prepared to assess the enzymatic breakdown of the self-healing polymers. In vitro degradation experiments involve weighing the dry polymers and incubating them at 37°C for 1, 3, 5, 7, 10, 24, and 48 hours in the collagenase solution. After incubation, the polymers are removed from the degradation medium, dried in a vacuum oven for 24 hours, and weighed again. For long-term degradation studies, DPBS serves as the degradation medium. Samples incubated at 37°C are taken out at the end of the 21st day, dried, and weighed.
5. Computational Approaches
Studying IDPols involves understanding their physical, chemical, and mechanical properties, which can be quite complex due to their dynamic, large and intricate molecular structures. Computational approaches can help in studying their physical, chemical, and mechanical properties. Molecular dynamics (MD) simulations involves simulating the physical movements of atoms and molecules in IDPols over time, allowing to study their dynamic behavior, structural properties, and interactions at a molecular level [55]. Furthermore, Monte Carlo (MC) simulations are utilized for studying polymers in thermodynamic equilibrium by using random sampling to compute the properties of polymer systems, which is particularly useful for examining phase transitions and thermodynamic properties [56]. Additionally, quantum mechanics (QM) calculations focus on understanding the electronic structure of IDPols [57]. Methods such as density functional theory (DFT) can be used to study the electronic properties and chemical reactivity of polymer chains [58]. Moreover, coarse-grained modeling simplifies the polymer model by reducing the number of degrees of freedom, allowing for the study of larger IDPols over longer timescales [59]. It is particularly useful for investigating the macroscopic properties of IDPols. Finite element analysis (FEA) is used to study the mechanical behavior of polymers under various conditions by dividing the polymer material into smaller, finite elements, and solving the behavior of these elements [60]. In addition, molecular docking and binding studies can help in understanding how IDPols interact with other molecules, which is crucial in drug delivery and designing polymers with specific binding properties. Finally, machine learning and data-driven approaches are increasingly being used to predict polymer properties and optimize polymer design by training models on experimental and simulation data [61].
6. Conclusions
Here, we present new chemical reaction mechanism and experimental designs for the studies of IDPols biomimicking intrinsically disordered proteins using disorder promoting peptides. These polymers can be manufactured by polymer and material industries for biomedical applications including tissue engineering, drug delivery, artificial organ design, immune compatibility and organ transplantation.
Author Contributions
Conceptualization, O. C.-W.; methodology, O. C.-W.; investigation, F. N. C.; writing—original draft preparation, O. C.-W., F. N. C., V.N.U.; writing—review and editing O. C.-W., F. N. C., V.N.U.; supervision, O. C.-W., V.N.U.
Funding
This study was supported by the Turkish – German University Scientific Research Projects Commission under the grant no: 2024BF05
References
- Dunker, A.K.; Lawson, J.D.; Brown, C.J.; Williams, R.M.; Romero, P.; Oh, J.S.; Oldfield, C.J.; Campen, A.M.; Ratliff, C.M.; Hipps, K.W.; Ausio, J.; Nissen, M.S.; Reeves, R.; Kang, C.; Kissinger, C.R.; Bailey, R.W.; Griswold, M.D.; Chiu, W.; Garner, E.C.; Obradovic, Z. ; Intrinsically disordered protein, J Mol Graph Model 2001, 19, 26–59. [CrossRef]
- Coskuner-Weber, O.; Mirzanli, O.; Uversky, V.N. , Intrinsically disordered proteins and proteins with intrinsically disordered regions in neurodegenerative diseases, Biophys Rev 2022, 14, 679–707. [CrossRef]
- Coskuner, O.; Uversky, V.N. , Intrinsically disordered proteins in various hypotheses on the pathogenesis of Alzheimer’s and Parkinson’s diseases, in: Progress in Molecular Biology and Translational Science, Elsevier, 2019: pp. 145–223. [CrossRef]
- Coskuner, O.; Murray, I.V.J. , Adenosine Triphosphate (ATP) Reduces Amyloid-β Protein Misfolding in vitro, JAD 2014, 41, 561–574. [CrossRef]
- Wright, P.E.; Dyson, H.J. , Intrinsically disordered proteins in cellular signalling and regulation, Nat Rev Mol Cell Biol 2015, 16, 18–29. [CrossRef]
- Darling, A.L.; Uversky, V.N. , Intrinsic Disorder and Posttranslational Modifications: The Darker Side of the Biological Dark Matter, Frontiers in Genetics 2018, 9, 158. 9. [CrossRef]
- Coskuner-Weber, O.; Uversky, V. , Insights into the Molecular Mechanisms of Alzheimer’s and Parkinson’s Diseases with Molecular Simulations: Understanding the Roles of Artificial and Pathological Missense Mutations in Intrinsically Disordered Proteins Related to Pathology, IJMS 2018, 19, 336. [CrossRef]
- Uversky, V.N. , Wrecked regulation of intrinsically disordered proteins in diseases: pathogenicity of deregulated regulators, Front Mol Biosci 2014, 1, 6. 1. [CrossRef]
- Ruan, H.; Sun, Q.; Zhang, W.; Liu, Y.; Lai, L. , Targeting intrinsically disordered proteins at the edge of chaos, Drug Discovery Today 2019, 24, 217–227. [CrossRef]
- He, B.; Wang, K.; Liu, Y.; Xue, B.; Uversky, V.N.; Dunker, A.K. , Predicting intrinsic disorder in proteins: an overview, Cell Res 2009, 19, 929–949. [CrossRef]
- Theillet, F.-X.; Kalmar, L.; Tompa, P.; Han, K.-H.; Selenko, P.; Dunker, A.K.; Daughdrill, G.W.; Uversky, V.N. , The alphabet of intrinsic disorder: I. Act like a Pro: On the abundance and roles of proline residues in intrinsically disordered proteins, Intrinsically Disord Proteins 1 (2013) e24360. [CrossRef]
- Uversky, V.N. , The alphabet of intrinsic disorder: II. Various roles of glutamic acid in ordered and intrinsically disordered proteins, Intrinsically Disord Proteins 1 (2013) e24684. [CrossRef]
- Coskuner, O.; Wise-Scira, O. , Arginine and Disordered Amyloid-β Peptide Structures: Molecular Level Insights into the Toxicity in Alzheimer’s Disease, ACS Chem. Neurosci. 2013, 4, 1549–1558. [Google Scholar] [CrossRef] [PubMed]
- Yuce-Erarslan, E.; Domb, J.; Kasem, H.; Uversky, V.N.; Coskuner-Weber, O. , Intrinsically Disordered Synthetic Polymers in Biomedical Applications, Polymers 2023, 15, 2406. [CrossRef]
- Coskuner-Weber, O.; Yuce-Erarslan, E.; Uversky, V.N. , Paving the Way for Synthetic Intrinsically Disordered Polymers for Soft Robotics, Polymers 2023, 15, 763. [CrossRef]
- Bhatia, S. , Nanotechnology in Drug Delivery, 0 ed., Apple Academic Press, 2017. [CrossRef]
- Juanes-Gusano, D.; Santos, M.; Reboto, V.; Alonso, M.; Rodríguez-Cabello, J.C. , Self-assembling systems comprising intrinsically disordered protein polymers like elastin-like recombinamers, J Pep Sci 28 (2022). [CrossRef]
- MacEwan, S.R.; Chilkoti, A. , Applications of elastin-like polypeptides in drug delivery, Journal of Controlled Release 2014, 190, 314–330. [CrossRef]
- Jose, G.; Shalumon, K.T.; Chen, J.-P. , Natural Polymers Based Hydrogels for Cell Culture Applications, CMC 2020, 27, 2734–2776. [CrossRef]
- Sarangthem, V.; Singh, T.D.; Dinda, A.K. , Emerging Role of Elastin-Like Polypeptides in Regenerative Medicine, Advances in Wound Care 2021, 10, 257–269. [CrossRef]
- Wang, B.; Patkar, S.S.; Kiick, K.L. , Application of Thermoresponsive Intrinsically Disordered Protein Polymers in Nanostructured and Microstructured Materials, Macromol. Biosci. 2021, 21, 2100129. [Google Scholar] [CrossRef]
- Pasinszki, T.; Krebsz, M.; Tung, T.T.; Losic, D. , Carbon Nanomaterial Based Biosensors for Non-Invasive Detection of Cancer and Disease Biomarkers for Clinical Diagnosis, Sensors 2017, 17, 1919. [CrossRef]
- Psoma, S.D.; Kanthou, C. , Wearable Insulin Biosensors for Diabetes Management: Advances and Challenges, Biosensors 2023, 13, 719. [CrossRef]
- Sempionatto, J.R.; Lasalde-Ramírez, J.A.; Mahato, K.; Wang, J.; Gao, W. , Wearable chemical sensors for biomarker discovery in the omics era, Nat Rev Chem 2022, 6, 899–915. [CrossRef]
- Samui, A.B. (Ed.) ; Smart polymers: basics; applications; First edition; Press, C.R., Boca Raton London New York, 2022.
- Heinritz, C.; Ng, X.J.; Scheibel, T. , Bio-inspired Protein-Based and Activatable Adhesion Systems, Adv Funct Materials 2024, 34, 2303609. [CrossRef]
- Balu, R.; Whittaker, J.; Dutta, N.K.; Elvin, C.M.; Choudhury, N.R. , Multi-responsive biomaterials and nanobioconjugates from resilin-like protein polymers, J. Mater. Chem. B 2014, 2, 5936–5947. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.T.; Zhao, Y. , Engineered Hybrid Nanoparticles for On-Demand Diagnostics and Therapeutics, Acc. Chem. Res. 2015, 48, 3016–3025. [Google Scholar] [CrossRef] [PubMed]
- Kröger, A.P.P.; Paulusse, J.M.J. , Single-chain polymer nanoparticles in controlled drug delivery and targeted imaging, Journal of Controlled Release 2018, 286, 326–347. [CrossRef]
- Chakrabarti, P.; Chakravarty, D. , Intrinsically disordered proteins/regions and insight into their biomolecular interactions, Biophysical Chemistry 2022, 283, 106769. [CrossRef]
- Uversky, V.N. , p53 Proteoforms and Intrinsic Disorder: An Illustration of the Protein Structure-Function Continuum Concept, Int J Mol Sci 2016, 17, 1874. [CrossRef]
- Liu, J.; Perumal, N.B.; Oldfield, C.J.; Su, E.W.; Uversky, V.N.; Dunker, A.K. , Intrinsic Disorder in Transcription Factors, Biochemistry 2006, 45, 6873–6888. [CrossRef]
- Melková, K.; Zapletal, V.; Narasimhan, S.; Jansen, S.; Hritz, J.; Škrabana, R.; Zweckstetter, M.; Jensen, M.R.; Blackledge, M.; Žídek, L. , Structure and Functions of Microtubule Associated Proteins Tau and MAP2c: Similarities and Differences, Biomolecules 2019, 9, 105. 9. [CrossRef]
- Iqbal, K.; Liu, F.; Gong, C.-X.; Grundke-Iqbal, I. , Tau in Alzheimer disease and related tauopathies, Curr Alzheimer Res 2010, 7, 656–664. [CrossRef]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. , Molecular chaperones in protein folding and proteostasis, Nature 2011, 475, 324–332. [CrossRef]
- Westerheide, S.D.; Raynes, R.; Powell, C.; Xue, B.; Uversky, V.N. , HSF Transcription Factor Family, Heat Shock Response, and Protein Intrinsic Disorder, CPPS 2012, 13, 86–103. [CrossRef]
- Cortese, M.S.; Uversky, V.N.; Dunker, A.K. , Intrinsic disorder in scaffold proteins: Getting more from less, Progress in Biophysics and Molecular Biology 2008, 98, 85–106. [CrossRef]
- Coskuner-Weber, O.; Uversky, V.N. , Liquid-Liquid Phase Separation Associated with Intrinsically Disordered Proteins: Experimental and Computational Tools, Curr Protein Pept Sci (2024). [CrossRef]
- Carey, J.L.; Guo, L. , Liquid-Liquid Phase Separation of TDP-43 and FUS in Physiology and Pathology of Neurodegenerative Diseases, Front. Mol. Biosci. 2022, 9, 826719. [Google Scholar] [CrossRef]
- Dawson, J.E.; Bah, A.; Zhang, Z.; Vernon, R.M.; Lin, H.; Chong, P.A.; Vanama, M.; Sonenberg, N.; Gradinaru, C.C.; Forman-Kay, J.D. , Non-cooperative 4E-BP2 folding with exchange between eIF4E-binding and binding-incompatible states tunes cap-dependent translation inhibition, Nat Commun 2020, 11, 3146. [CrossRef]
- Coskuner, O.; Wise-Scira, O. , Structures and Free Energy Landscapes of the A53T Mutant-Type α-Synuclein Protein and Impact of A53T Mutation on the Structures of the Wild-Type α-Synuclein Protein with Dynamics, ACS Chem. Neurosci. 2013, 4, 1101–1113. [Google Scholar] [CrossRef] [PubMed]
- Wise-Scira, O.; Dunn, A.; Aloglu, A.K.; Sakallioglu, I.T.; Coskuner, O. , Structures of the E46K Mutant-Type α-Synuclein Protein and Impact of E46K Mutation on the Structures of the Wild-Type α-Synuclein Protein, ACS Chem. Neurosci. 2013, 4, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Wise-Scira, O.; Aloglu, A.K.; Dunn, A.; Sakallioglu, I.T.; Coskuner, O. , Structures and Free Energy Landscapes of the Wild-Type and A30P Mutant-Type α-Synuclein Proteins with Dynamics, ACS Chem. Neurosci. 2013, 4, 486–497. [Google Scholar] [CrossRef]
- Bello-Madruga, R.; Burgas, M.T. , The limits of prediction: Why intrinsically disordered regions challenge our understanding of antimicrobial peptides, Computational and Structural Biotechnology Journal 2024, 23, 972–981. [CrossRef]
- De Oliveira, A.A.; Vergara, A.; Wang, X.; Vederas, J.C.; Oudit, G.Y. , Apelin pathway in cardiovascular, kidney, and metabolic diseases: Therapeutic role of apelin analogs and apelin receptor agonists, Peptides 2022, 147, 170697. [CrossRef]
- Orzáez, M.; Gortat, A.; Mondragón, L.; Pérez-Payá, E. , Peptides and Peptide Mimics as Modulators of Apoptotic Pathways, ChemMedChem 2009, 4, 146–160. [CrossRef]
- Weber, O.C. , Intrinsically Disordered Proteins by Homology Modeling and Replica Exchange Molecular Dynamics Simulations: A Case Study of Amyloid-β42, JOTCSA (2024) 1151–1164. [CrossRef]
- Zheng, W.; Zhang, C.; Li, Y.; Pearce, R.; Bell, E.W.; Zhang, Y. , Folding non-homologous proteins by coupling deep-learning contact maps with I-TASSER assembly simulations, Cell Reports Methods 2021, 1, 100014. [CrossRef]
- Xie, F.; Halley, P.J.; Avérous, L. , Rheology to understand and optimize processibility, structures and properties of starch polymeric materials, Progress in Polymer Science 2012, 37, 595–623. [CrossRef]
- Di, X.; Li, L.; Jin, Q.; Yang, R.; Li, Y.; Wang, X.; Wu, G.; Yuan, C.; Sensitive, H. ; Degradable, and Rapid Self-Healing Hydrogel Sensor with Semi-Interpenetrating Network for Recognition of Micro-Expressions, Small (2024) 2403955. [CrossRef]
- Baqasah, H.; He, F.; Zai, B.A.; Asif, M.; Khan, K.A.; Thakur, V.K.; Khan, M.A. , In-Situ Dynamic Response Measurement for Damage Quantification of 3D Printed ABS Cantilever Beam under Thermomechanical Load, Polymers 2019, 11, 2079. [CrossRef]
- Shah, V.P.; Elkins, J.S.; Williams, R.L. , Evaluation of the Test System Used for In Vitro Release of Drugs for Topical Dermatological Drug Products, Pharmaceutical Development and Technology 1999, 4, 377–385. [CrossRef]
- Akter, M.; Sikder, M.T.; Rahman, M.M.; Ullah, A.K.M.A.; Hossain, K.F.B.; Banik, S.; Hosokawa, T.; Saito, T.; Kurasaki, M. , A systematic review on silver nanoparticles-induced cytotoxicity: Physicochemical properties and perspectives, Journal of Advanced Research 2018, 9, 1–16. [CrossRef]
- Lucas, N.; Bienaime, C.; Belloy, C.; Queneudec, M.; Silvestre, F.; Nava-Saucedo, J.-E. , Polymer biodegradation: Mechanisms and estimation techniques – A review, Chemosphere 2008, 73, 429–442. [CrossRef]
- Barrat, J.-L.; Baschnagel, J.; Lyulin, A. , Molecular dynamics simulations of glassy polymers, Soft Matter 2010, 6, 3430. [CrossRef]
- Mavrantzas, V.G. , Using Monte Carlo to Simulate Complex Polymer Systems: Recent Progress and Outlook, Front. Phys. 2021, 9, 661367. [Google Scholar] [CrossRef]
- Alvarez-Estrada, R.F.; Calvo, G.F. , Models for polymers and biopolymers based on quantum mechanics, Molecular Physics 2002, 100, 2957–2970. [CrossRef]
- McCormick, T.M.; Bridges, C.R.; Carrera, E.I.; DiCarmine, P.M.; Gibson, G.L.; Hollinger, J.; Kozycz, L.M.; Seferos, D.S. , Conjugated Polymers: Evaluating DFT Methods for More Accurate Orbital Energy Modeling, Macromolecules 2013, 46, 3879–3886. [CrossRef]
- Kmiecik, S.; Gront, D.; Kolinski, M.; Wieteska, L.; Dawid, A.E.; Kolinski, A. , Coarse-Grained Protein Models and Their Applications, Chem. Rev. 2016, 116, 7898–7936. [Google Scholar] [CrossRef]
- Fisher, F.T.; Brinson, L.C. , Viscoelastic interphases in polymer–matrix composites: theoretical models and finite-element analysis, Composites Science and Technology 2001, 61, 731–748. [CrossRef]
- Cencer, M.M.; Moore, J.S.; Assary, R.S. , Machine learning for polymeric materials: an introduction, Polymer International 2022, 71, 537–542. [CrossRef]
Scheme 1.
The reaction of a disorder-promoting peptide sequence with monobromo triethylene glycol yields a complex.
Scheme 1.
The reaction of a disorder-promoting peptide sequence with monobromo triethylene glycol yields a complex.
Scheme 2.
Prepolymer synthesis mechanism using L-lysine ethyl ester diisocyanate.
Scheme 3.
Synthesis of an IDPol using a disorder promoting oligopeptide.
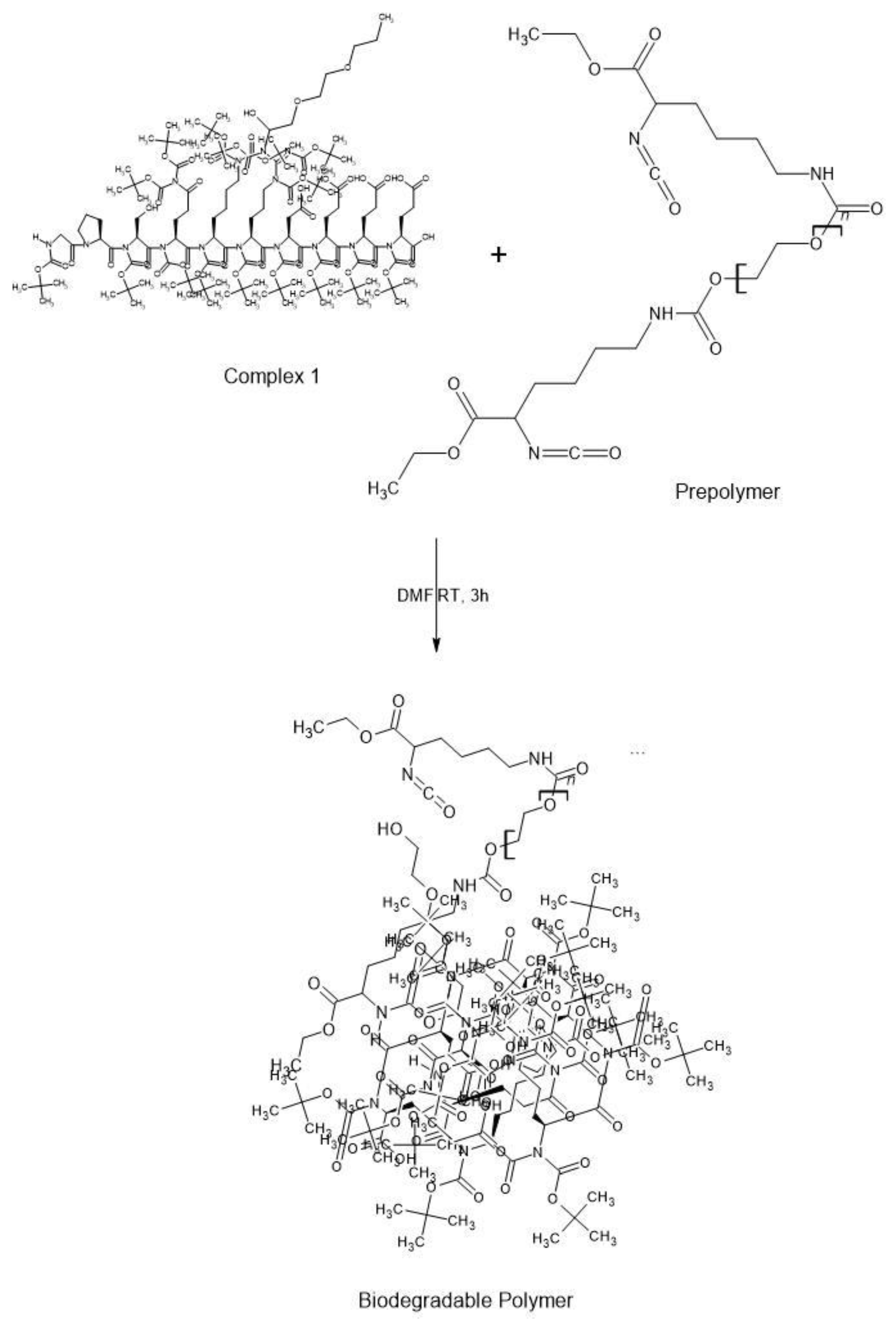

Table 1.
Representative disordered oligopeptides along with their sequence, predicted structure, and biological function.
Table 1.
Representative disordered oligopeptides along with their sequence, predicted structure, and biological function.
| Sequence | Structure | Degree of Accuracy (C-scoreb) | Function | |
|---|---|---|---|---|
| Peptide 1 | GPSQKRDEEE |  |
-1.38 | hydrolase activity |
| Peptide 2 | PSGREQKDKK |  |
-1.46 | cation binding 3',5'-cyclic-nucleotide phosphodiesterase activity |
| Peptide 3 | DQKRESPGDP |  |
-1.33 | peptide transporter activity |
| Peptide 4 | KERQDSPGKK |  |
-1.16 | nucleic acid binding |
| Peptide 5 | SDPKQGRDEP |  |
-1.18 | transporter activity |
| Peptide 6 | RKGEQDPSKG |  |
-1.30 | -tRNA binding -Sep-tRNA:Cys-tRNA -synthase activity -heparan sulfate proteoglycan binding -heparin binding -asparaginase activity -complement component C3b binding |
| Peptide 7 | EGPSKQRDGE |  |
-1.15 | glucosidase activity |
| Peptide 8 | DQGRKPSQED |  |
-1.03 | -fructokinase activity -glucose binding -ATP binding -glucokinase activity |
| Peptide 9 | PRGKDEQSGP |  |
-1.56 | cation binding |
| Peptide 10 | SKGEQPRDGP |  |
-1.26 | cation binding |
| Peptide 11 | KDSGPRQEGK |  |
-1.28 | metal ion binding |
| Peptide 12 | RQDEGPSKPR |  |
-1.18 | RNA glycosylase activity |
| Peptide 13 | GPRQSKEDGQ |  |
-1.34 | -oxidoreductase activity -antioxidant activity -ATP binding -GMP synthase (glutamine-hydrolyzing) activity -protein disulfide isomerase activity |
| Peptide 14 | KGRPQDEGSK |  |
-1.03 | -DNA binding -RNA polymerase III activity -RNA polymerase II activity -RNA polymerase I activity |
| Peptide 15 | DPSQGKREPD |  |
-0.84 | cation binding |
| Peptide 16 | QEGPRKSDGP |  |
-1.30 | translation initiation factor activity |
| Peptide 17 | SKPGQERDKS |  |
-0.84 | cation binding |
| Peptide 18 | RKDPSGEQRP |  |
-1.18 | DNA binding |
| Peptide 19 | EPQGRSDPKR |  |
-1.18 | 4-hydroxy-tetrahydrodipicolinate synthase |
| Peptide 20 | DGEQKSPRQD |  |
-1.31 | -transition metal ion binding -endopeptidase activity |
| Peptide 21 | QKDRPGESKP |  |
-1.36 | cellulase activity |
| Peptide 22 | GRKDESPQGR |  |
-1.15 | xylanase activity |
| Peptide 23 | SPRKEQGDPS |  |
-1.25 | -cofactor binding -aryl-acylamidase activity -protein homodimerization activity -(+)-trans-carveol dehydrogenase activity -amino acid binding -ATP binding -iron ion binding -phenylalanine 4-monooxygenase activity |
| Peptide 24 | PKRGSQDPEK |  |
-1.33 | -oxidoreductase activity, acting on CH-OH group of donors |
| Peptide 25 | EGRDPQSKGR |  |
-1.28 | -purine ribonucleoside triphosphate binding -adenyl ribonucleotide binding -purine ribonucleoside binding -nucleic acid binding |
| Peptide 26 | QPGSRKDEPG |  |
-1.37 | -transferase activity, transferring acyl groups other than amino-acyl groups |
| Peptide 27 | SGKRQEDPGR |  |
-1.26 | catalytic activity |
| Peptide 28 | DEGPRSKGQK |  |
-1.48 | -3-oxo-pimeloyl-[acp] methyl ester reductase activity -trans-2-enoyl-CoA reductase (NADPH) activity -2,4-dienoyl-CoA reductase (NADPH) activity -receptor binding -3-oxoacyl-[acyl-carrier-protein] reductase (NADPH) activity -nucleotide binding |
| Peptide 29 | RQPKGESDPQ |  |
-1.26 | -purine ribonucleoside triphosphate binding -RNA polymerase activity -adenyl ribonucleotide binding -purine ribonucleoside binding -substrate-specific channel activity -ion transmembrane transporter activity -cysteine-type peptidase activity -helicase activity -endopeptidase activity -nucleic acid binding |
| Peptide 30 | KSDPGRQEKS |  |
-1.05 | -adenine phosphoribosyltransferase activity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



