
Submitted:
07 October 2024
Posted:
08 October 2024
You are already at the latest version
Abstract
The CXC chemokine receptor 5 (CXCR5) is a member of the G protein-coupled receptor family that is highly expressed in B cells and a subset of T cells such as T follicular helper cells. Various types of cancers, including non-small cell lung cancer, breast cancer, and prostate cancer also express CXCR5; therefore, antibodies that specifically bind to CXCR5 could be useful for clarification of the mechanisms of cancer progression. In this study, we aimed to develop high-affinity monoclonal antibodies (mAbs) for flow cytometry against mouse CXCR5 (mCXCR5). The established anti-mCXCR5 mAb, Cx5Mab-3 (rat IgG2b, kappa), reacted with mCXCR5-overexpressed Chinese hamster ovary (CHO)-K1 (CHO/mCXCR5) by flow cytometry. Kinetic analyses using flow cytometry indicated that the dissociation constants (KD) of Cx5Mab-3 for CHO/mCXCR5 is 7.2 × 10−10 M, indicating that Cx5Mab-3 is useful for detecting mCXCR5 by flow cytometry with high affinity.
Keywords:
mouse CXCR5
; monoclonal antibody
; peptide immunization
; flow cytometry
1. Introduction
Chemokines are classified into four different subfamilies: CC, CXC, CX3C, and XC, which depend on the position and number of cysteine residues in their N-terminus.[1] The chemokine receptors belong to seven-transmembrane G-protein-coupled receptor (GPCR) families, and trigger intracellular signal transductions through binding to the ligands [1]. They play fundamental roles in development, homeostasis, immune system, cell proliferation, angiogenesis, and lymphocyte differentiation [2,3]. The CXC chemokine receptor type 5 (CXCR5) is predominantly expressed on the surface of B cells and a subset of T cells, such as T follicular helper (Tfh) cells [4]. The CXCR5+ Tfh cells interact with germinal center B cells and promote the differentiation into plasma cells or memory formation through somatic hypermutation and class-switch [5]. Therefore, CXCR5+ Tfh cells play an important role in secondary lymphoid tissue orchestration and lymphoid neogenesis in the spleen, lymph nodes, and Peyer’s patches [1,4].
CXCL13 is one of the ligands of CXCR5 [6]. The CXCL13/CXCR5 axis activates downstream signaling, including PI3K/Akt, MEK/ERK, and Rac pathways, which modulates immune cells to promote lymphocyte infiltration, activation, and differentiation to enhance the antitumor immune response [6]. Furthermore, CXCR5 is also expressed in cancer cells, which makes pivotal contributions to the development and progression [1,7,8,9,10,11]. Therefore, monoclonal antibodies (mAbs), which specifically bind to CXCR5 would be useful for cancer therapy and elucidation of the disease progression.
In this study, we report a novel anti-mCXCR5 mAb, which was successfully developed by the N-terminal peptide immunization.
1.1. Plasmids, Peptides, and Cell lines
The synthesized DNA (Eurofins Genomics KK) encoding mCXCR5 (Accession No.: NM_007551.3) was subsequently subcloned into a pCAGzeo vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). The mCXCR5 plasmid was transfected into LN229 and CHO-K1 cells, using a Neon transfection system (Thermo Fisher Scientific Inc., Waltham, MA). Stable transfectants were established by staining with anti-mCXCR5 mAb (clone L138D7; BioLegend, San Diego, CA) and sorted using a cell sorter (SH800; Sony Corp., Tokyo, Japan).
Eurofins Genomics KK (Tokyo, Japan) synthesized a partial sequence of the N-terminal extracellular region of mCXCR5 (1-MNYPLTLDMGSITYNMDDL-19) plus C-terminal cysteine. Subsequently, the keyhole limpet hemocyanin (KLH) was conjugated at the C-terminus of the peptide.
1.2. Development of mCXCR5-Producing Hybridomas
The Animal Care and Use Committee of Tohoku University (Permit number: 2022MdA-001) approved animal experiments. We immunized one rat intraperitoneally with 100 μg of the KLH-conjugated mCXCR5 peptide (mCXCR5-KLH) with Alhydrogel adjuvant 2% (InvivoGen). The hybridoma supernatants were subsequently screened using enzyme-linked immunosorbent assay (ELISA) with the mCXCR5 peptide, followed by flow cytometry using CHO/mCXCR5 and CHO-K1.
1.3. ELISA
The mCXCR5 peptide was immobilized on Nunc Maxisorp 96 well immunoplates (Thermo Fisher Scientific Inc.). After blocking, plates were then incubated with supernatants of hybridomas, followed by peroxidase-conjugated anti-rat immunoglobulins (Sigma-Aldrich Corp., St. Louis, MO). Next, enzymatic reactions were conducted, using ELISA POD Substrate TMB Kit (Nacalai Tesque, Inc.).
1.4. Flow Cytometric Analysis
Cells were treated with 10, 1, 0.1, or 0.01 μg/mL of Cx5Mab-3 or L138D7 for 30 min at 4°C. For peptide inhibition assay, Cx5Mab-3 (0.1 μg/mL) or L138D7 (0.1 μg/mL) were pre-incubated with 1 μg/mL of mCXCR5 peptide or dimethyl sulfoxide (DMSO) for 30 min at 4°C, and further incubated with CHO/mCXCR5 for 30 min at 4°C. The cells were treated with Alexa Fluor 488-conjugated anti-rat IgG (1:2000; Cell Signaling Technology, Inc., Danvers, MA). The fluorescence data were collected using the SA3800 Cell Analyzer (Sony Corp.). The KD was calculated using one-site binding models in GraphPad PRISM 6 (GraphPad Software, Inc., La Jolla, CA).
2. Results
2.1. Development of Anti-mCXCR5 mAbs Using N-Terminal Peptide Immunization
To develop anti-mCXCR5 mAbs, one rat was immunized with the KLH-conjugated mCXCR5 peptide (Figure 1A). Spleen was then excised from the rat, after which splenocytes were fused with myeloma P3U1 cells (Figure 1B). Developed hybridomas were subsequently seeded into fourteen 96-well plates and cultivated for six days. Then, positive wells for the naked mCXCR5 peptide were selected using ELISA, followed by the selection of mCXCR5-expressing cell-reactive and CHO-K1-non-reactive supernatants using flow cytometry (Figure 1C). The ELISA screening identified 74 out of 1342 wells (5.5%), which strongly reacted with the naked mCXCR5 peptide. The flow cytometric screening identified 18 out of the 74 wells (24.3%), which exhibited strong signals to CHO/mCXCR5 cells but not to CHO-K1 cells. After the limiting dilution and several additional screenings, anti-mCXCR5 mAb, Cx5Mab-3 (rat IgG2b, kappa) was finally established (Figure 1D).
2.2. Flow Cytometric Analysis Using Cx5Mab-3
To check the reactivity of Cx5Mab-3 and L138D7, we conducted flow cytometry against CHO/mCXCR5 and CHO-K1. Cx5Mab-3 recognized CHO/mCXCR5 cells dose-dependently at 10, 1, 0.1, and 0.01 μg/mL (Figure 2A). Parental CHO-K1 cells were not recognized even at 10 μg/mL of all mAbs (Figure 2B). The reactivity of Cx5Mab-3 was also observed in LN229/mCXCR5 cells (Figure 3). The sensitivity against CHO/mCXCR5 or LN229/mCXCR5 cells was similar between Cx5Mab-3 and L138D7 (Figs. 2 and 3).
We next performed a peptide-blocking assay. As shown in Figure 4, Cx5Mab-3 and L138D7 reacted with CHO/mCXCR5 cells. The reaction of Cx5Mab-3 was completely neutralized by mCXCR5 peptide, indicating that the reaction by Cx5Mab-3 was mediated by the recognition of N-terminus of mCXCR5. Because the reaction of L138D7 was not neutralized, the epitope of L138D7 is different from that of Cx5Mab-3.
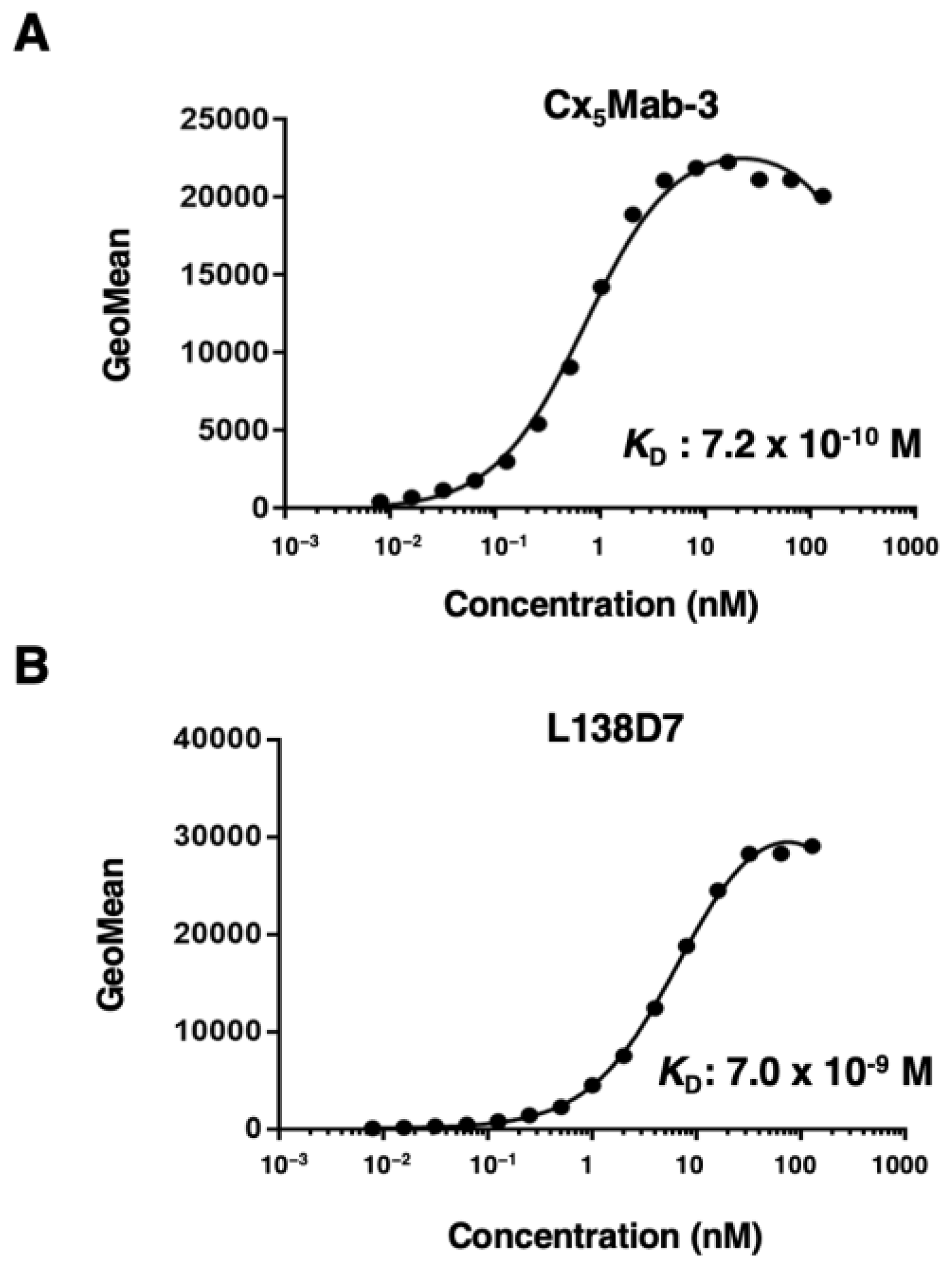
2.3. Determination of Dissociation Constant of Anti-mCXCR5 mAbs against CHO/mCXCR5 Cells
We determined the apparent dissociation constant (KD) of Cx5Mab-3 and L138D7 against mCXCR5 by flow cytometry. The geometric mean of the fluorescence intensity of CHO/mCXCR5 at each concentration of Cx5Mab-3 and L138D7 was plotted. By fitting one-site binding models, the KD values of Cx5Mab-3 and L138D7 for CHO/mCXCR5 were determined as 7.2 × 10−10 M and 7.0 × 10−9 M (Figure 5), indicating that Cx5Mab-3 possess higher affinity than L138D7 for CHO/mCXCR5 cells.
4. Discussion
In this study, we developed a novel anti-mCXCR5 mAb (clone Cx5Mab-3) using N-terminal peptide immunization and investigated for flow cytometry to detect mCXCR5. We checked the reactivity of Cx5Mab-3 in western blotting; however, mCXCR5 protein was not detected by Cx5Mab-3 (data not shown), although Cx5Mab-3 detects the N-terminal peptide of mCXCR5 in ELISA (Figure 1). A commercially available anti-mCXCR5 mAb (clone L138D7) was developed by immunizing rats with mCXCR5-transfected cells and is valid only for flow cytometry [12]. Because the reaction of L138D7 was not neutralized by N-terminal peptide (Figure 4), the epitope of L138D7 might be the conformational one, which is located in the other extracellular domains. In the future study, we will determine the binding epitope of L138D7.
It has been reported that the development of therapeutic agents targeting the CXCL13/CXCR5 axis can be effective in treating cancers and inflammatory diseases [13]. CXCR5+ CD4+ Tfh cells mainly contribute to the functions to stimulate B cell proliferation, antibody/B cell receptor class-switch, and antibody production in infection, cancer, and autoimmunity [14]. Moreover, CXCR5+ CD8+ T cells possess not only abovementioned function but also maintain a cytolytic capacity in infection, autoimmunity, and tumor microenvironments resembling CD8 T+ effector cells [15,16]. In inflammatory diseases and autoimmunity, CXCR5+ CD8+ T cells are implicated in disease progression [17]. In contrast, CXCR5 expression on CD8+ T cells generally is indicative of progenitor memory stem-like exhausted cells, which are more responsive to immune checkpoint blockade therapy in tumors [18]. Furthermore, CD8+ lymphocyte populations with high PD-1 expression in tumors secrete high levels of CXCL13 [19]. Secretion of CXCL13 by the tumor-infiltrating lymphocytes with high PD-1 serves to attract other immune cell subsets to the tumor microenvironment, including Tfh cells and B cells [19]. Therefore, CXCL13 will predict response to anti-PD-1 therapy that correlates with increased overall survival and durable responses [19], and represent novel biomarkers for response to targeted PD1/PD-L1 therapy [20].
An inverse correlation between the expression of CXCR5 and p53 tumor suppressor was also reported in human breast cancer cell lines [21]. CXCR5 is increased by silencing p53 in MCF7 cells, which potentiates CXCL13-mediated chemotaxis [21]. A CXCR5 promoter analysis revealed that p53 suppresses the transcriptional activity of NF-κB, which is essential for the upregulation of CXCR5 [21]. The related tumor suppressors p63 and p73 also exhibit similar mechanisms for CXCR5 gene regulation [22]. Since CXCL13 is one of the overexpressed chemokines in breast cancer tissues compared with normal breast tissues, mAb therapies against CXCR5 could be an essential strategy to treat the tumor [22]. Therefore, Cx5Mab-3 may help obtain the proof of concept in preclinical studies.
In our previous studies, we changed the isotype of mAbs into mouse IgG2a to retain the antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC), which led to high antitumor activities in mouse xenograft models [23,24,25]. Since the subclass of Cx5Mab-3 was rat IgG2b, it might possess ADCC and CDC activities. In further studies, we will evaluate whether the depletion of CXCR5-expressing immunosuppressive cells can enhance antitumor activities in mouse syngeneic and xenograft models of human cancers.
Author Contributions
K.I. and M.H. performed the experiments. H.S., T.T., and M.K.K. analyzed the data. Y.K designed the experiments. K.I. and H.S. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest involving this article.
References
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. Febs j 2018;285(16): 2944-2971. [CrossRef]
- Luther, S.A.; Cyster, J.G. Chemokines as regulators of T cell differentiation. Nat Immunol 2001;2(2): 102-107. [CrossRef]
- Sokol, C.L.; Luster, A.D. The chemokine system in innate immunity. Cold Spring Harb Perspect Biol 2015;7(5). [CrossRef]
- Su, Z.; Chen, L.; Niu, Q.; Yang, B.; Huang, Z. Association of Gene Polymorphisms in CXC Chemokine Receptor 5 with Rheumatoid Arthritis Susceptibility. Iran J Allergy Asthma Immunol 2022;21(5): 537-548. [CrossRef]
- Shi, J.; Hou, S.; Fang, Q.; et al. PD-1 Controls Follicular T Helper Cell Positioning and Function. Immunity 2018;49(2): 264-274.e264. [CrossRef]
- Kazanietz, M.G.; Durando, M.; Cooke, M. CXCL13 and Its Receptor CXCR5 in Cancer: Inflammation, Immune Response, and Beyond. Front Endocrinol (Lausanne) 2019;10: 471. [CrossRef]
- Shiels, M.S.; Pfeiffer, R.M.; Hildesheim, A.; et al. Circulating inflammation markers and prospective risk for lung cancer. J Natl Cancer Inst 2013;105(24): 1871-1880. [CrossRef]
- Eide, H.A.; Halvorsen, A.R.; Sandhu, V.; et al. Non-small cell lung cancer is characterised by a distinct inflammatory signature in serum compared with chronic obstructive pulmonary disease. Clin Transl Immunology 2016;5(11): e109. [CrossRef]
- Hussain, M.; Adah, D.; Tariq, M.; et al. CXCL13/CXCR5 signaling axis in cancer. Life Sci 2019;227: 175-186. [CrossRef]
- Panse, J.; Friedrichs, K.; Marx, A.; et al. Chemokine CXCL13 is overexpressed in the tumour tissue and in the peripheral blood of breast cancer patients. Br J Cancer 2008;99(6): 930-938. [CrossRef]
- Singh, S.; Singh, R.; Sharma, P.K.; et al. Serum CXCL13 positively correlates with prostatic disease, prostate-specific antigen and mediates prostate cancer cell invasion, integrin clustering and cell adhesion. Cancer Lett 2009;283(1): 29-35. [CrossRef]
- https://www.biolegend.com/ja-jp/products/purified-anti-mouse-cd185-cxcr5-antibody-8454, accessed on 10 May 2024.
- Hsieh, C.H.; Jian, C.Z.; Lin, L.I.; et al. Potential Role of CXCL13/CXCR5 Signaling in Immune Checkpoint Inhibitor Treatment in Cancer. Cancers (Basel) 2022;14(2). [CrossRef]
- Crotty, S. Follicular helper CD4 T cells (TFH). Annu Rev Immunol 2011;29: 621-663. [CrossRef]
- Im, S.J.; Hashimoto, M.; Gerner, M.Y.; et al. Defining CD8+ T cells that provide the proliferative burst after PD-1 therapy. Nature 2016;537(7620): 417-421. [CrossRef]
- Brummelman, J.; Mazza, E.M.C.; Alvisi, G.; et al. High-dimensional single cell analysis identifies stem-like cytotoxic CD8(+) T cells infiltrating human tumors. J Exp Med 2018;215(10): 2520-2535. [CrossRef]
- He, R.; Hou, S.; Liu, C.; et al. Follicular CXCR5- expressing CD8(+) T cells curtail chronic viral infection. Nature 2016;537(7620): 412-428. [CrossRef]
- Kallies, A.; Zehn, D.; Utzschneider, D.T. Precursor exhausted T cells: key to successful immunotherapy? Nat Rev Immunol 2020;20(2): 128-136. [CrossRef]
- Thommen, D.S.; Koelzer, V.H.; Herzig, P.; et al. A transcriptionally and functionally distinct PD-1(+) CD8(+) T cell pool with predictive potential in non-small-cell lung cancer treated with PD-1 blockade. Nat Med 2018;24(7): 994-1004. [CrossRef]
- de Chaisemartin, L.; Goc, J.; Damotte, D.; et al. Characterization of chemokines and adhesion molecules associated with T cell presence in tertiary lymphoid structures in human lung cancer. Cancer Res 2011;71(20): 6391-6399. [CrossRef]
- Mitkin, N.A.; Hook, C.D.; Schwartz, A.M.; et al. p53-dependent expression of CXCR5 chemokine receptor in MCF-7 breast cancer cells. Sci Rep 2015;5: 9330. [CrossRef]
- Mitkin, N.A.; Muratova, A.M.; Sharonov, G.V.; et al. p63 and p73 repress CXCR5 chemokine receptor gene expression in p53-deficient MCF-7 breast cancer cells during genotoxic stress. Biochim Biophys Acta Gene Regul Mech 2017;1860(12): 1169-1178. [CrossRef]
- Takei, J.; Kaneko, M.K.; Ohishi, T.; et al. A defucosylated anti-CD44 monoclonal antibody 5-mG2a-f exerts antitumor effects in mouse xenograft models of oral squamous cell carcinoma. Oncol Rep 2020;44(5): 1949-1960. [CrossRef]
- Hosono, H.; Takei, J.; Ohishi, T.; et al. Anti-EGFR monoclonal antibody 134-mG2a exerts antitumor effects in mouse xenograft models of oral squamous cell carcinoma. Int J Mol Med 2020;46(4): 1443-1452. [CrossRef]
- Itai, S.; Ohishi, T.; Kaneko, M.K.; et al. Anti-podocalyxin antibody exerts antitumor effects via antibody-dependent cellular cytotoxicity in mouse xenograft models of oral squamous cell carcinoma. Oncotarget 2018;9(32): 22480-22497. [CrossRef]
Figure 1.
The development of anti-mCXCR5 mAbs. (A) CHO/mCXCR5 cells were immunized into a Sprague–Dawley rat. (B) The spleen cells were fused with P3U1 cells. (C) To select anti-mCXCR5 mAb-producing hybridomas, the supernatants were screened by ELISA as 1st screening and by flow cytometry using CHO-K1 and CHO/mCXCR5 cells as 2nd screening. (D) After limiting dilution, an anti-mCXCR5 mAb, Cx5Mab-3 was finally established.
Figure 1.
The development of anti-mCXCR5 mAbs. (A) CHO/mCXCR5 cells were immunized into a Sprague–Dawley rat. (B) The spleen cells were fused with P3U1 cells. (C) To select anti-mCXCR5 mAb-producing hybridomas, the supernatants were screened by ELISA as 1st screening and by flow cytometry using CHO-K1 and CHO/mCXCR5 cells as 2nd screening. (D) After limiting dilution, an anti-mCXCR5 mAb, Cx5Mab-3 was finally established.
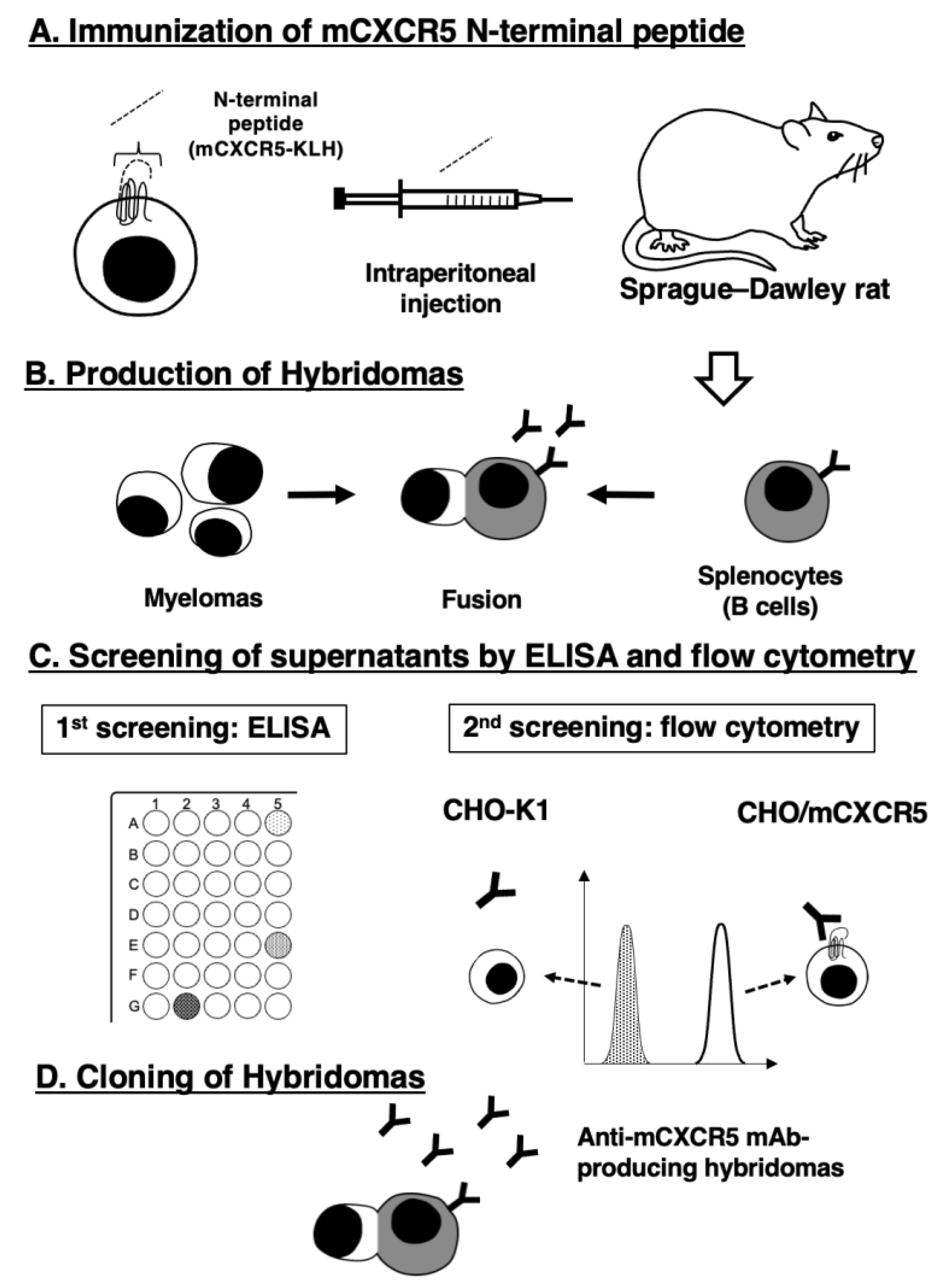
Figure 2.
Flow cytometry of mCXCR5-expressed CHO-K1 cells using Cx5Mab-3 and L138D7. CHO/mCXCR5 (A) and CHO-K1 (B) cells were treated with 0.01–10 µg/mL of Cx5Mab-3 or L138D7. Then, cells were treated with anti-rat IgG conjugated with Alexa Fluor 488 (black line). The filled gray represents the negative control (blocking buffer).
Figure 2.
Flow cytometry of mCXCR5-expressed CHO-K1 cells using Cx5Mab-3 and L138D7. CHO/mCXCR5 (A) and CHO-K1 (B) cells were treated with 0.01–10 µg/mL of Cx5Mab-3 or L138D7. Then, cells were treated with anti-rat IgG conjugated with Alexa Fluor 488 (black line). The filled gray represents the negative control (blocking buffer).
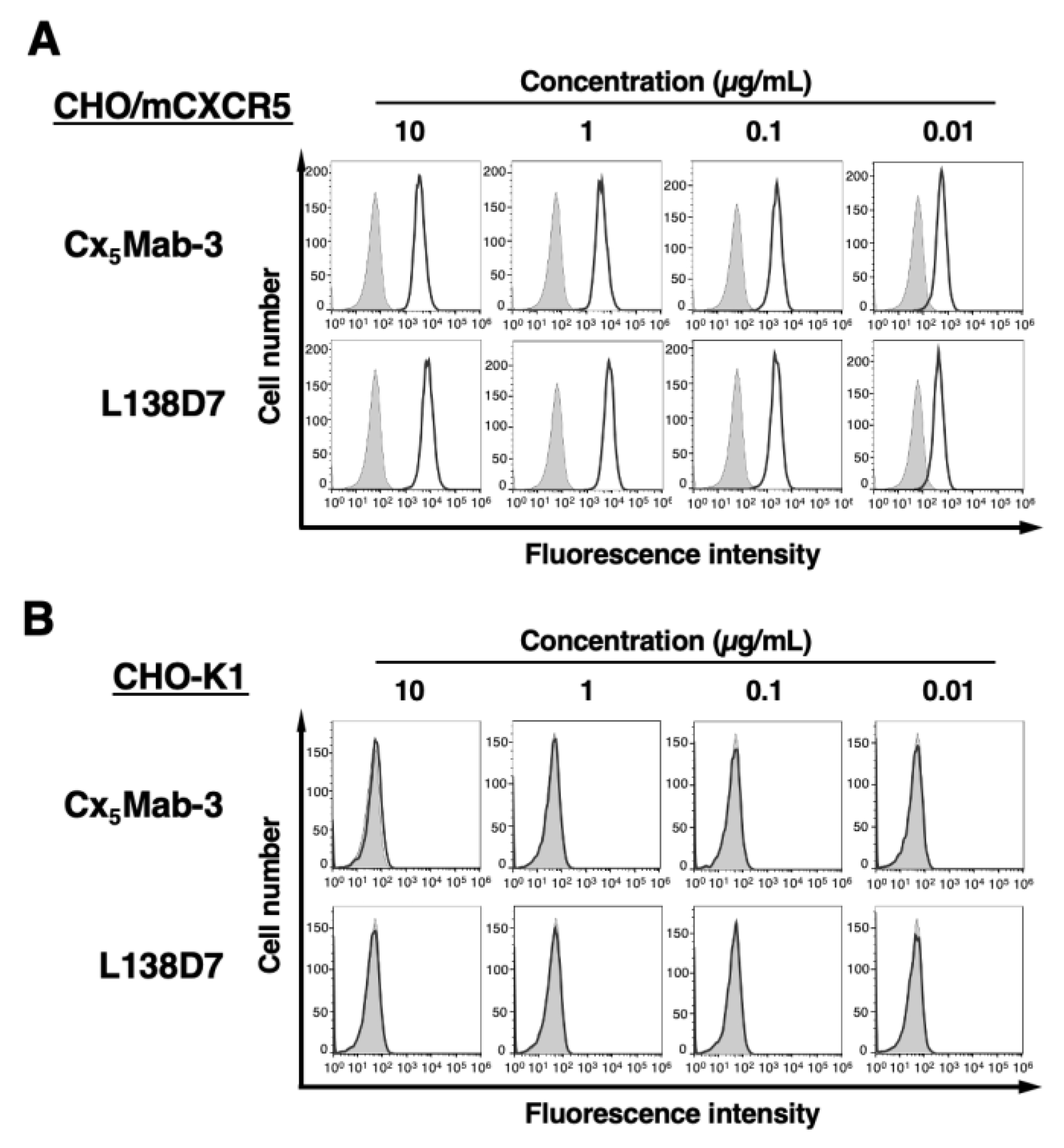
Figure 3.
Flow cytometry of mCXCR5-expressed LN229 cells using Cx5Mab-3 and L138D7. LN229/mCXCR5 (A) and LN229 (B) cells were treated with 0.01–10 µg/mL of Cx5Mab-3 or L138D7. The cells were treated with anti-rat IgG conjugated with Alexa Fluor 488 (black line). The filled gray represents the negative control (blocking buffer).
Figure 3.
Flow cytometry of mCXCR5-expressed LN229 cells using Cx5Mab-3 and L138D7. LN229/mCXCR5 (A) and LN229 (B) cells were treated with 0.01–10 µg/mL of Cx5Mab-3 or L138D7. The cells were treated with anti-rat IgG conjugated with Alexa Fluor 488 (black line). The filled gray represents the negative control (blocking buffer).
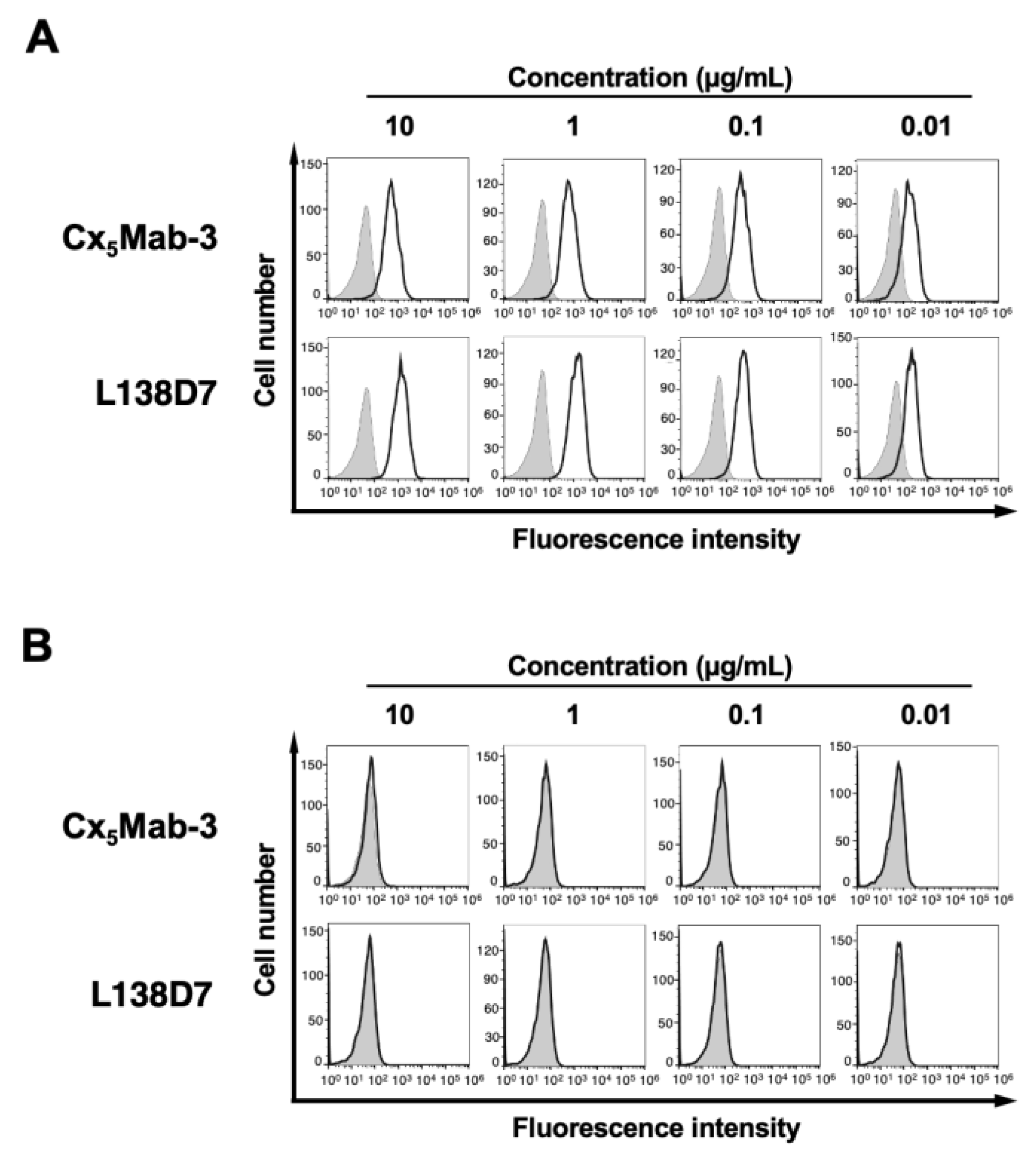
Figure 4.
A peptide-blocking assay using Cx5Mab-3 with mCXCR5 peptide. Cx5Mab-3 (0.1 µg/mL) plus control (1% DMSO in blocking buffer), Cx5Mab-3 plus mCCR5 peptide (1 μg/ml), L138D7 (0.1 µg/mL) plus control (1% DMSO in blocking buffer), or L138D7 plus mCCR5 peptide (1 μg/ml) were reacted with CHO/mCXCR5 for 30 min at 4°C, followed by treatment with Alexa Fluor 488-conjugated anti-rat IgG. The filled gray represents the negative control (blocking buffer). DMSO, dimethyl sulfoxide.
Figure 4.
A peptide-blocking assay using Cx5Mab-3 with mCXCR5 peptide. Cx5Mab-3 (0.1 µg/mL) plus control (1% DMSO in blocking buffer), Cx5Mab-3 plus mCCR5 peptide (1 μg/ml), L138D7 (0.1 µg/mL) plus control (1% DMSO in blocking buffer), or L138D7 plus mCCR5 peptide (1 μg/ml) were reacted with CHO/mCXCR5 for 30 min at 4°C, followed by treatment with Alexa Fluor 488-conjugated anti-rat IgG. The filled gray represents the negative control (blocking buffer). DMSO, dimethyl sulfoxide.
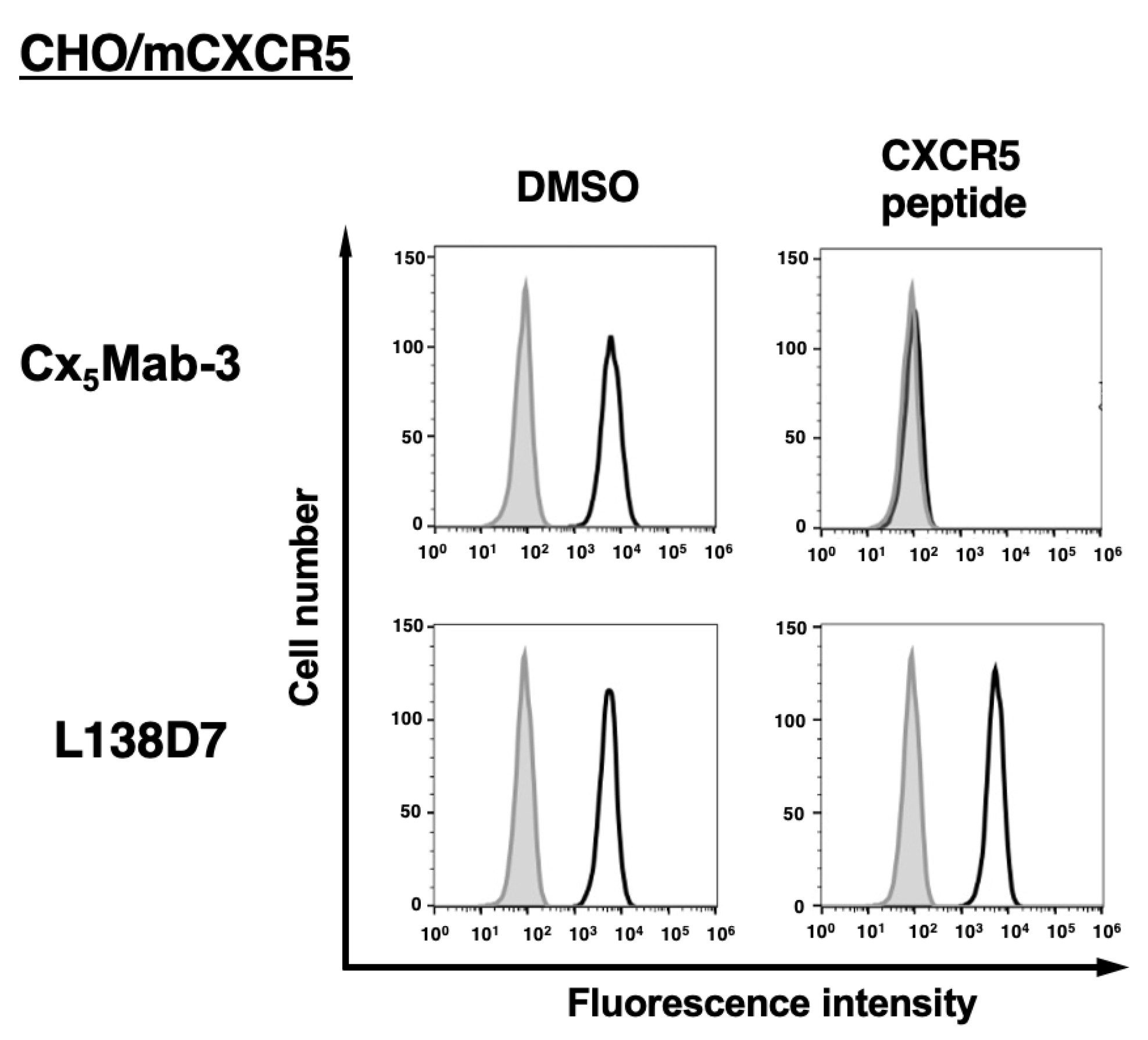
Figure 5.
The binding affinity of Cx5Mab-3. CHO/mCXCR5 cells were suspended in serially diluted Cx5Mab-3 (A) or L138D7 (B). The cells were treated with anti-rat IgG conjugated with Alexa Fluor 488. The fluorescence data were subsequently collected using the SA3800 Cell Analyzer, followed by the calculation of the KD using GraphPad PRISM 6.
Figure 5.
The binding affinity of Cx5Mab-3. CHO/mCXCR5 cells were suspended in serially diluted Cx5Mab-3 (A) or L138D7 (B). The cells were treated with anti-rat IgG conjugated with Alexa Fluor 488. The fluorescence data were subsequently collected using the SA3800 Cell Analyzer, followed by the calculation of the KD using GraphPad PRISM 6.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



