
Submitted:
07 October 2024
Posted:
08 October 2024
You are already at the latest version
Abstract
Swine influenza A viruses (IAVsw) are important causes of disease in pigs but also constitute a public health risk. IAVsw strains show remarkable differences in pathogenicity. We aimed to generate airway organoids from the porcine lower respiratory tract and use these to establish well-differentiated airway epithelial cell (WD-AEC) cultures grown at air-liquid interface (ALI) for in vitro screening of IAVsw strain virulence. Epithelial cells were isolated from bronchus tissue of juvenile pigs, and airway organoids were cultured in extracellular matrix in medium with human growth factors. Single cell suspensions of these 3D organoids were seeded on transwell filters and differentiated at ALI to form a pseudostratified epithelium containing ciliated cells, mucus-producing cells and tight junctions. Inoculation with a low dose of IAVsw in a low volume inoculum resulted in virus replication without requiring addition of trypsin, and was quantified by detection of viral genome loads in apical washes. Interestingly, inoculation of an H3N2 strain known to cause severe disease in pigs induced greater reduction in trans-epithelial resistance and more damage to tight junctions than H1N2 or H1N1 strains associated with mild disease in pigs. We conclude that the porcine WD-AEC model is useful to assess the virulence of IAVsw strains.
Keywords:
air-liquid interface
; swine influenza virus
; airway epithelial cells
; transwell cultures
; pig
; organoids
1. Introduction
Influenza A virus (IAV) infections cause disease in multiple hosts, including humans, poultry and wild birds, but are also an important cause of respiratory tract infections in pigs [1,2]. Most swine IAV (IAVsw) infections are associated with relatively mild clinical signs, but infections can also result in severe disease and increased susceptibility to secondary infections within the porcine respiratory disease complex. Importantly, IAVsw strains pose a public health risk due to their ability to reassort which potentially leads to variants that are more transmissible and pathogenic in humans. [3]. Pigs are considered a potential ‘mixing vessel’ for exchange of IAV genetic segments, as they are susceptible to human, avian and porcine IAV strains. If a single host cell is infected by two different IAV strains, exchange of gene segments can lead to a virus with a novel combination of gene segments (‘reassortment’), potentially resulting in viruses with new infection properties in swine, birds or humans [1,4,5]. A prominent example is the 2009 Swine flu pandemic, caused by swine H1N1 virus after multiple reassortment events with other influenza A viruses [6] that led to sustained human-to-human transmission. IAVsw is enzootic in pigs in Europe [7], and occasional transmission events to humans have been reported.
Influenza A viruses have a single stranded negative sense RNA genome consisting of eight segments and belong to the family Orthomyxoviridae. The two major transmembrane glycoproteins hemagglutinin (HA) and neuraminidase (NA) are used to classify IAVs into subtypes. IAVsw strains require proteolytic cleavage of a monobasic arginine of the HA precursor protein by trypsin-like serine proteases such as TMPRRS2 or HAT to facilitate fusion of HA2 with the endosomal membrane [8,9]. Importantly, the porcine respiratory epithelium possess N-acetylneuraminic acids, the receptor used by IAV, linked to galactose both via alpha2,3 as well as via alpha2,6 linkage and can thus, in addition to IAVsw, also be infected by avian and human IAVs [9].
Traditional immortalized cell lines have low trypsin-like serine protease activity, and propagation of IAVsw in these cells requires the addition of an exogenous protease. IAVsw strains are often propagated in Madin-Darby canine kidney cells, either or not modified to express additional sialic acids [10,11]. However, replication in immortalized cells and addition of trypsin-like proteases to the culture media may result in culture adaptations that confound viral kinetics. Therefore, primary tissue culture, primary cell culture and organoid culture have been considered as alternatives that more closely mimic the respiratory tract epithelium of the host [12]. Primary-cell-based well-differentiated airway epithelial cell (WD-AEC) cultures differentiated at air-liquid interface (ALI) provide a reproducible and readily available in vitro model with endogenous protease activity and a heterogenous cell population mimicking the in vivo airway epithelium. However, these cells have limited in vitro expansion potential. Airway organoids (AOs) provide another powerful tool to model airway epithelium in vitro. AOs can be derived from pluripotent or tissue-residing stem cells and possess self-renewing capacity. Therefore, AOs can be expanded for many passages and can be cryostored and retrieved, which allows more experiments per tissue donor and increasing experimental reproducibility. Moreover, it was recently shown that human AOs can be differentiated at ALI into WD-AECs that can be used for in vitro viral infection studies [13,14,15].
Different porcine tissue or cell culture models have been developed to model respiratory virus infections. These include respiratory nasal explants [16,17], primary airway epithelial cells cultured submerged in medium [18,19], precision cut lung slices (PCLS) [20], primary airway epithelial cells grown at ALI [17,21,22,23] and airway organoids [24,25,26]. Here, we describe the development of porcine lower respiratory tract AO-derived WD-AEC cultures which were employed to characterize three different endemic IAVsw strains of subtypes H1N1, H1N2 and H3N1 in vitro. We aimed to investigate whether differences in viral kinetics and virulence can be recapitulated in vitro using AO-derived WD-AEC cultures.
2. Materials and Methods
Isolation of Porcine Airway Organoids (AO)
Porcine lungs were collected from pigs that were euthanized as healthy control animals in unrelated animal experiments (license AVD40100202010304, experiment number 2020.D-0024.002), or for sampling of primary cells for diagnostics (license AVD4010020174311, experiment number 2017.D-0063.004). Primary bronchial epithelial cells were isolated as described previously [27] and cultured in basement membrane extract (Geltrex BME, Gibco) submerged in airway organoid medium [28] containing 500ng/mL Rspondin 1, 25ng/mL FGF7, 100ng/mL FGF10, 100ng/mL Noggin (all Peprotech), 500nM A83-01 (Tocris), 5µM Y-27632 (Bio-Connect), 500µM SB202190 (Sigma), 1x B27 (Gibco), 1.25mM N-Acetylcysteine (Sigma), 5mM Nicotinamide (Sigma) in Advanced Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12 (Advanced DMEM/F12, Gibco) with 1x GlutaMax (Gibco), 10mM HEPES (Lonza), 1x antibiotic-antimycotic (Gibco) and 0.1 mg/mL primocin (InVivogen). Briefly, the main bronchi were dissected and digested overnight at 4°C on a platform rocker in DMEM/F12 (Gibco) with 10 µg/mL DNAse (Sigma-Aldrich), 1 mg/mL protease XIV (Sigma-Aldrich), 1 x antibiotic-antimycotic and 0.1 mg/mL primocin (InVivogen). Subsequently, epithelial cells were harvested by mechanical scraping and strained through a 70 µm filter (Corning). Red blood cells were lysed using Red Blood Cell Lysis buffer (Roche). After washing in PBS, a small fraction of the pellet was resuspended in BME, and seeded as 5x15µL droplets per well onto a 24 well plate. 500µL of airway organoid medium was added per well, and refreshed every 5 days. AOs were split every 2 weeks using TrypLE Express (Gibco), and AO cultures were maintained at 37°C, 5%CO2.
Differentiation of AO on Air-Liquid-Interface (ALI)
Nunc Cell Culture inserts in Nunc carrier plates (both Thermo Fisher Scientific), 0.4 µm pore size, pore density <0.85×108 pores cm−2, 0.47 cm2 culture area, were coated with 33µg/mL collagen IV and 70µg/mL fibronectin (both Sigma-Aldrich). AOs were dissociated using TrypLE Express to obtain single cell suspensions and subsequently seeded at a density of 0.5-1×105 cells per transwell filter starting as liquid-liquid interface. Growth medium consisted of Airway Epithelial Cell Basal Medium (AECBM) prepared according to manufacturer’s instructions (Promocell), supplemented with 1 x antibiotic-antimycotic and 0.1 mg/mL primocin. Medium was refreshed every five days. A tight monolayer formed over the course of approximately one week, after which the growth medium was removed from the apical side, creating an air-liquid interface (ALI). The following day, cultures were transferred to basolateral differentiation medium containing bovine pituitary extract, l-epidermal growth factor, insulin (recombinant human), hydrocortisone, epinephrine, triiodo-l-thyronine, transferrin (recombinant human) and retinoic acid, as described previously [27]. Cells were maintained in differentiation medium at 37°C, 5%CO2, with medium exchanges every five days. Once per week, the apical side was washed with prewarmed PBS containing magnesium and calcium, to remove excess mucus.
Isolation and Propagation of IAVsw Strains
H1N1 Neth/Rhezerveen/CVI9121A/2012 (EPI_ISL_195178), H3N2 Neth/Ysselsteyn/CVI8866A/2012 (EPI_ISL_195222) and H1N2 Neth/Barger-Compascuum/CVI6324A/2012 (EPI_ISL_195176) were derived from naturally infected farmed pigs. Virus was isolated at Wageningen Bioveterinary Research, Lelystad, the Netherlands, from lung tissue samples as described previously [29]. Early passage virus stocks were produced in Madin-Darby canine kidney (MDCK) cells: 100uL virus stock was added to 90% confluent MDCK monolayer on T150 tissue culture flasks using 0.5ug/mL trypsin TPCK (Thermo Fisher). Upon development of CPE, infection was terminated at -80°C before collection and subsequent centrifugation of supernatant at 1000g for 10 minutes at 4°C.
TEER Measurements
To measure epithelial integrity, 400µL prewarmed PBS was added to the apical side of WD-AECs. Next, electrodes connected to an EVOM3 volt/Ohm meter (World Precision Instruments) were placed into the apical PBS and basolateral medium. The obtained value was multiplied by the surface area of the transwell filter and expressed in ohm*cm2.
Infection of WD-AECs with IAVsw
For IAVsw infection experiments, porcine WD-AECs of three different donors were used, and experiments were repeated three times. In each experiment, four transwell filters were infected with each IAVsw strain, of which one was fixed 24 hours post infection (hpi), while three were followed until 48hpi. Two uninfected filters were fixed at 0h for staining of the WD-AED cultures. Before inoculation, an apical wash with PBS was performed to remove mucus. Subsequently, virus stocks were rapidly thawed in a 37°C waterbath and diluted in PBS to a virus concentration of 104 or 105 TCID50/ml. 10µL of diluted inoculum was added apically onto the WD-AECs. After 1h at 37°C, four apical washes were performed to remove unbound inoculum, and a fifth wash was performed to collect and freeze a first sample for assessment of virus loads. After 16h, 24h and 48h, 400µL PBS was added to the apical side, briefly incubated at room temperature (RT), harvested and stored at -70°C until analysis. At 24h and 48h, TEER measurements were performed after the collection of apical washes, using a separate electrode per virus and thorough cleaning between measuring individual WD-AEC samples. At 48hpi, filters were fixed in 4% paraformaldehyde and stored in PBS at 4°C.
RNA Isolation and IAV qRT-PCR
RNA of apical washes stored in Molgen lysis buffer was isolated by an automated robot system (PurePrep 96) using the Molgen RNA isolation kit (OE00290096). A one-step quantitative reverse-transcriptase PCR (qRT-PCR) was performed using the TaqMan® Fast Virus 1-Step Master Mix (Applied Biosystems; Foster City, CA, USA), 500 mM of the forward (5’-CTTCTAACCGAGGTCGAAACGTA-3’) and reverse primers( 5’-CACTGGGCACGGTGAGC-3’), 250 nM probe (5’-6FAM-TCAGGCCCCCTCAAAGCCGA-QSY-3’) and 2 µL RNA template. The RT-qPCR was performed on a LightCycler480 platform with the following cycling conditions: 50 °C for 5 min, 95 °C for 20 s, 95 °C for 5 s and 58 °C for 15 s and 72 °C for 20 s for 45 cycles. To calculate viral copy numbers, gBlocks were ordered at IDT (Netherlands) covering the N-terminal 220bp gene fragment of segment 7 matrix genes of Influenza A virus (A/swine/Indiana/A02751461/2023 (H1N2)).
Histology and Immunohistochemistry
For transverse histology sections, formalin-fixed filters were embedded in paraffin blocks according to general pathology principles and cut as 7µm formalin-fixed paraffin-embedded (FFPE) sections for staining by hematoxylin and eosin, or for immunohistochemistry (IHC).
For IHC labeling of p63 and Muc5AC, antigen retrieval was performed for 15 min at 100 °C in 10 mM citate buffer at pH6 (DAKO) in a pressure cooker. For acetylated antigen, no antigen retrieval was performed. After unspecific antigen binding blocking for 30 min at RT with 1% normal goat serum, the primary antibodies were incubated in PBS with 1 % BSA for 45 min at RT. The following antibodies were used: anti-p63 (clone 4A4, Abcam), anti-Muc5AC (clone 45M1, Abcam) and anti-acetylated tubulin (clone 6-11B-1, Thermo Fisher Scientific). Subsequently, the EnVision+System HRP (DAKO) was applied at RT for 30 min, followed by incubation with 3,3’-diaminobenzidine (DAB) for 5 min before counterstaining with Mayer’s hematoxylin and mounting with Eukitt mounting medium (Sigma Aldrich).
Immunofluorescence
Paraformaldehyde-fixed filters were permeabilized using 0.25% Triton X-100 in PBS for 10 minutes at RT, followed by blocking in 5% BSA in PBS for 1h at RT. Anti-ZO-1 Alexa Fluor 594 (clone ZO1-1A12, Thermo Fisher Scientific) was diluted in 1 % BSA in PBS and incubated for 3h at RT, while anti-Muc5AC Alexa Fluor 488 (clone 45M1, Thermo Fisher Scientific) and anti-acTub-555 (clone 7E5H8, Thermo Fisher Scientific) were added after 2h to incubate for 1h at RT. For staining of IAV NP, the HB65 anti-NP antibody was labeled to Alexa Fluor 488 using ReadyLabelTM Antibody labeling kit according to the manufacturer’s instructions (Thermo Fisher Scientific), and added to the primary antibody incubation while omitting anti-Muc5AC-488. DAPI counterstaining for 10 minutes at RT was performed before mounting in ProLong Diamond Antifade mountant (Thermo Fisher scientific). Samples were imaged using the Leica Stellaris 5 WLL confocal laser scanning microscope.
3. Results
3.1. Establishment of Porcine AO-Derived WD-AECs
Porcine airway epithelial cells were isolated from primary bronchi, expanded in extracellular matrix as 3D airway organoids (AOs) and subsequently seeded as single cells on transwell filters (Figure 1A). Once the cell layers reached confluency, the apical medium was removed to generate an air-liquid interface (ALI). If sufficient tight junctions had formed, no more cell culture medium leaked through the cell layer, facilitating the apical exposure to air. Subsequently, the cell layer differentiated over the course of 3-5 weeks into a pseudostratified muco-ciliated respiratory epithelium (Figure 1B) containing basal and goblet cells (Figure 1C).
Epithelial integrity was furthermore evaluated by measuring transepithelial electrical resistance (TEER) and remained stable from week 2 of differentiation onwards (Figure 1D). Using laser scanning confocal microscopy, tight junctions were detected early after differentiation by staining with an antibody to ZO-1. Goblet cells, visualized by staining of MUC5AC, appeared after one week of differentiation and were most numerous at 2-4 weeks of differentiation. Cilia, stained by an anti-acetylated tubulin antibody, were visible starting from week 3 and were fully developed by week 5 (Figure 2).
3.2. Swine Influenza Virus Strains Replicate in Porcine WD-AECs
To functionally evaluate porcine WD-AECs, we employed inoculations with IAVsw as an infection model (Figure 3A and Supplemental figure S1). WD-AECs of three different porcine donors were washed with PBS to remove mucus and subsequently inoculated with 10uL of 102 or 103 TCID50 of H1N1, H1N2 or H3N2 strains for one hour. Apical washes collected at 16, 24 or 48hpi were analyzed by qRT-PCR. For all three virus strains, an increase in viral RNA genome loads were observed over time. For H1N2 a dose-response was maintained throughout 48h, while for H1N1 and H3N2 a dose-dependent difference in viral RNA copies was visible only at earlier time points (Figure 3B). TEER values remained stable for H1N2-inoculated WD-AECs, while for H3N2, TEER values reached almost zero after 48h. TEER values for H1N1-inoculated WD-AECs declined over time (Figure 3C).
3.3. IAVsw Strains Cause Epithelial Damage on WD-AECs
Transverse FFPE sections of WD-AECs fixed after 48hpi demonstrated substantial loss of cells after H3N2 inoculation, while the epithelial cell layer of H1N2-inoculated WD-AECs remained comparable to PBS-inoculated WD-AECs (Figure 3D and Supplemental figure S1). For H1N1, only the dose of 103 appeared to cause damage to the cell layer, although the effect was much less compared to H3N2-infected cultures.
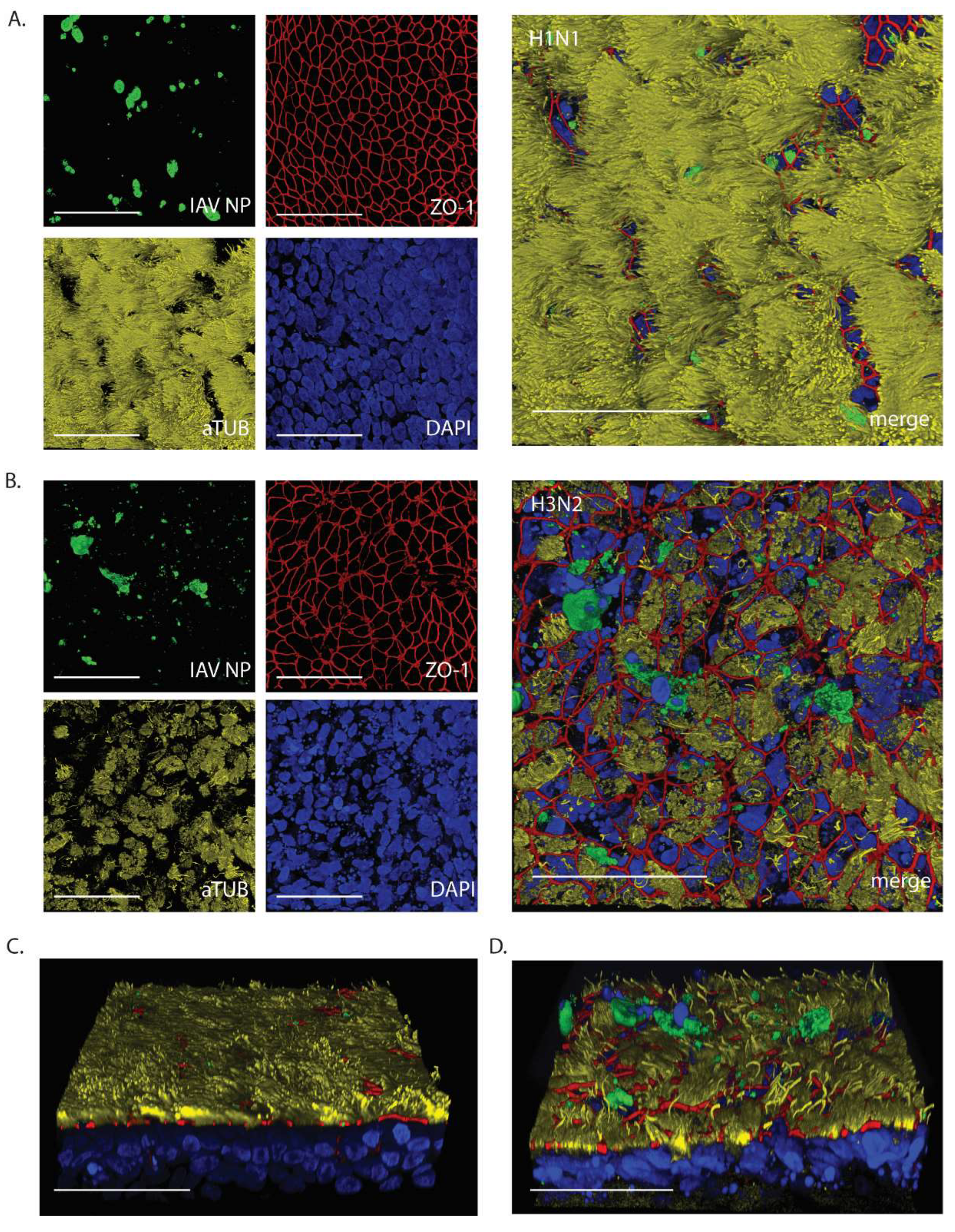
At 24hpi, WD-AECs inoculated with 103 of H1N1 or H3N2 were evaluated by confocal laser scanning microscopy (Figure 4). While the epithelium composition remained comparable to intact WD-AECs with H1N1 (see Figure 2), striking cytopathogenic effects were observed for H3N2-inoculated WD-AECs. Tight junctions were discontinuous, cilia were shortened and less abundant, and there was an increase of karyorrhectic debris. At the same time, IAV-infected cells were visible in both cell layers, as shown by staining against viral NP. Interestingly, IAV NP-positive cells were often located apically within or slightly above the WD-AEC layer (Figure 4C and D).
4. Discussion
Here, we describe the generation of porcine airway organoids from lung tissue of adult pigs, and subsequent seeding on transwells leading to the generation of pseudostratified well-differentiated airway epithelial cells (WD-AEC) cultured at air-liquid interface (ALI) to obtain an in vitro stem cell-derived model of the mucociliary respiratory epithelium of pigs. Subsequently, we used these WD-AED cultures for characterization of IAVsw strains of subtypes H1N1, H1N2 and H3N2.
Generation of porcine AOs was largely based on a protocol previously described for human airway AOs [28]. Indeed, human growth factors are sufficiently conserved to allow their use for expansion of porcine airway epithelial stem cells [24]. However, airway organoids grown in 3D culture are not conveniently used as model for respiratory virus infection, as their cilia are usually directed inwards [28]. This is a disadvantage when mimicking the natural infection route via inhaled air, because the apical side of the respiratory epithelium is not easily accessible. This challenge can be resolved by applying protocols to culture apical-out airway organoids although these cultures will remain at a liquid-liquid interface [30].
We chose to prepare single cell preparations of AOs grown in 3D, and seed these in transwell filters to generate WD-AEC cultures grown at ALI [13,31]. By doing so, ciliated cells are facing upwards and are exposed to air, making them easily accessible for inoculation and apical washes, and the cultures resemble the bronchial epithelium found in vivo. A major advantage of using AO-derived WD-AECs instead of primary-cell derived WD-AECs is the stem-cell-based renewal of organoids. Thus, cells obtained from a single donor can be expanded to high numbers, allowing long-term use of a single donor. This increases the inter-experimental comparability, allows the comparison between donors and can be used to reflect animal-to-animal variation which is observed in vivo.
Interestingly, during the differentiation process, we observed an increase of goblet cells at weeks 2/3 which subsequently decreased. This coincided with an observed increased production of mucus during this time. We observed that the frequencies of goblet cells declined in subsequent weeks, when ciliated cells appeared. By staining the individual cell populations, we determined that, in our hands, porcine WD-AECs are suitable for virus infection studies after 5 weeks of differentiation.
Conventional cell lines require addition of an exogenous protease for initial infection with influenza viruses with hemagglutinins containing a monobasic cleavage site. In contrast, AO-derived WD-AECs can be readily infected without the requirement of exogenous protease activity. Moreover, the high susceptibility and permissiveness of the cells was shown using of a low inoculum dose of 102 or 103 TCID50, allowing room for exponential virus replication and dissemination. We chose to use two parallel doses with a tenfold difference to demonstrate that differences in in vitro virulence were not caused by minor dose calculation errors.
Using porcine WD-AECs, it was feasible to detect differences in viral kinetics and cytopathogenicity between the three different IAVsw strains. The strains were isolated from pigs during a field investigation in 2012, during which H3N2 viruses were the most virulent circulating strains (Norbert Stockhofe, personal communication). In our experiments, H1N2 reached lower viral RNA copy numbers than H1N1 and H3N2, while no substantial differences in genome loads were observed between H3N2 and H1N1. However, when evaluating TEER values and histology, H3N2 clearly inflicted the most pronounced cytopathic effect. The high cytopathogenicity of the H3N2 strain was reproduced in three independent experiments, using tissues from three different porcine donors. Moreover, these results are in line with previously reported data using virus isolates of the same IAVsw subtypes on porcine precision-cut lung slices [20]. In vivo, similar cytokine responses and viral lung titers were previously observed for H3N2 and H1N1 isolates [32], although there are reports describing more severe clinical disease upon infection with H3N2 [20]. H1N2 isolates seem to cause slightly delayed immune responses compared to H1N1 and H3N2 isolates [32].
Several other studies have reported on IAVsw infections in porcine ALI models generated from primary cells. Wu et al. used a relatively high inoculum dose (approximately 105 infectious units) of a H1N1 or H3N2 strain and detected viral shedding over a period of more than a week, associated with loss of ciliated cells but retained TEER values [21]. Krunkosky et al. used a low inoculum (250 plaque forming units) and showed efficient virus replication in the absence of substantial drops in TEER values [17]. We consistently observed a reduction in TEER values after infection with our IAVsw H3N2 strain, coinciding with loss of tight junction integrity and clear loss of cells detected by histochemistry. Whether this is related to differences in the culture properties or differences between the virus strains used will need to be determined. We intend to use our AO-derived ALI model to isolate currently circulating IAVsw strains, and compare their in vitro and in vivo virulence.
This study has several limitations. We used airway organoids of three porcine donors, but in addition to donor differences we also observed batch-to-batch differences between cultures, with substantial differences in the TEER values detected at the start of the infection. Moreover, we used virus strains that were isolated more than ten years ago. Although these had been well characterized in vitro, no detailed in vivo virulence data were available. Finally, the virus strains were isolated in MDCK cells, potentially resulting in tissue culture adaptations[19]. Future studies will be performed with primary isolates obtained in ALI cultures, thus retaining the in vivo properties of the virus as much as possible.
In conclusion, porcine airway organoid-derived WD-AECs offer a potential bridge between in vivo animal models or clinical studies and in vitro experiments in conventional immortalized cell lines, because of their similarity with in vivo respiratory epithelium. WD-AECs therefore offer a valuable tool to characterize new virus variants, as well as to isolate virus from field samples. Moreover, in vitro studies to evaluate therapeutics, e.g., antivirals, can be performed, as well as studies to evaluate the host range of viruses when comparing WD-AECs from several animal species. The next step is to combine these WD-AEC cultures with immune cells, to recapitulate the complex interactions between respiratory epithelium and the immune system.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Replicates of IAVsw infection experiments on porcine WD-AECs of two different donors.
Author Contributions
Conceptualization, N.M.G, M.V. and R.d.S.; methodology, N.M.G., R.d.S., M.V., S.v.O. and R.D.; software, L.B. and R.d.S.; validation, R.D. and S.v.O.; formal analysis, R.D., M.V. and N.M.G.; investigation, R.D., S.v.O. and N.M.G.; resources, J.W. and R.d.S.; data curation, N.M.G. and M.V.; writing—original draft preparation, N.M.G.; writing—review and editing, M.V., L.B., J.W., R.d.S.; visualization, R.D., L.B., N.M.G., R.d.S.; supervision, R.d.S.; project administration, N.M.G., M.V. and R.d.S.; funding acquisition, J.W., R.d.S., M.V. and N.M.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Next Level Animal Sciences program of Wageningen University and Research.
Data Availability Statement
All data will be made available upon reasonable request.
Acknowledgments
The authors thank Ruth Bossers-de Vries, Helmi Fijten, Evelien Germeraad, Bart van der Hee, Stan Peters, Bregtje Smid, Norbert Stockhofe, Androniki Voulgari-Kokota and the pathology department of WBVR for their contributions to the study and/or critical feedback to the manuscript. Schematic illustrations in Figure 1 and 3 were generated with BioRender.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Castrucci, M.R.; Donatelli, I.; Sidoli, L.; Barigazzi, G.; Kawaoka, Y.; Webster, R.G. Genetic reassortment between avian and human influenza A viruses in Italian pigs. Virology 1993, 193, 503–506. [Google Scholar] [CrossRef]
- Ma, W. Swine influenza virus: Current status and challenge. Virus Res 2020, 288, 198118. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.K.; Chang, J.; Arendsee, Z.W.; Venkatesh, D.; Souza, C.K.; Kimble, J.B.; Lewis, N.S.; Davis, C.T.; Vincent, A.L. Swine Influenza A Viruses and the Tangled Relationship with Humans. Cold Spring Harb Perspect Med 2021, 11, a038737. [Google Scholar] [CrossRef]
- Cogdale, J.; Kele, B.; Myers, R.; Harvey, R.; Lofts, A.; Mikaiel, T.; Hoschler, K.; Banyard, A.C.; James, J.; Mollett, B.C.; et al. A case of swine influenza A(H1N2)v in England, November 2023. Euro Surveill 2024, 29, 2400002. [Google Scholar] [CrossRef] [PubMed]
- Hennig, C.; Graaf, A.; Petric, P.P.; Graf, L.; Schwemmle, M.; Beer, M.; Harder, T. Are pigs overestimated as a source of zoonotic influenza viruses? Porcine Health Manag 2022, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Novel Swine-Origin Influenza, A.V.I.T.; Dawood, F.S.; Jain, S.; Finelli, L.; Shaw, M.W.; Lindstrom, S.; Garten, R.J.; Gubareva, L.V.; Xu, X.; Bridges, C.B.; et al. Emergence of a novel swine-origin influenza A (H1N1) virus in humans. N Engl J Med 2009, 360, 2605–2615. [Google Scholar] [CrossRef]
- Simon, G.; Larsen, L.E.; Durrwald, R.; Foni, E.; Harder, T.; Van Reeth, K.; Markowska-Daniel, I.; Reid, S.M.; Dan, A.; Maldonado, J.; et al. European surveillance network for influenza in pigs: surveillance programs, diagnostic tools and Swine influenza virus subtypes identified in 14 European countries from 2010 to 2013. PLoS One 2014, 9, e115815. [Google Scholar] [CrossRef]
- Bottcher, E.; Matrosovich, T.; Beyerle, M.; Klenk, H.D.; Garten, W.; Matrosovich, M. Proteolytic activation of influenza viruses by serine proteases TMPRSS2 and HAT from human airway epithelium. J Virol 2006, 80, 9896–9898. [Google Scholar] [CrossRef]
- Peitsch, C.; Klenk, H.D.; Garten, W.; Bottcher-Friebertshauser, E. Activation of influenza A viruses by host proteases from swine airway epithelium. J Virol 2014, 88, 282–291. [Google Scholar] [CrossRef]
- Harada, Y.; Takahashi, H.; Trusheim, H.; Roth, B.; Mizuta, K.; Hirata-Saito, A.; Ogane, T.; Odagiri, T.; Tashiro, M.; Yamamoto, N. Comparison of suspension MDCK cells, adherent MDCK cells, and LLC-MK2 cells for selective isolation of influenza viruses to be used as vaccine seeds. Influenza Other Respir Viruses 2020, 14, 204–209. [Google Scholar] [CrossRef]
- Suderman, M.; Moniwa, M.; Alkie, T.N.; Ojkic, D.; Broes, A.; Pople, N.; Berhane, Y. Comparative Susceptibility of Madin-Darby Canine Kidney (MDCK) Derived Cell Lines for Isolation of Swine Origin Influenza A Viruses from Different Clinical Specimens. Viruses 2021, 13, e13. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A. Cell culture keeps pace with influenza virus. Lancet Respir Med 2018, 6, 805–806. [Google Scholar] [CrossRef] [PubMed]
- Lamers, M.M.; Mykytyn, A.Z.; Breugem, T.I.; Wang, Y.; Wu, D.C.; Riesebosch, S.; van den Doel, P.B.; Schipper, D.; Bestebroer, T.; Wu, N.C.; et al. Human airway cells prevent SARS-CoV-2 multibasic cleavage site cell culture adaptation. Elife 2021, 10, e66815. [Google Scholar] [CrossRef]
- Rijsbergen, L.C.; van Dijk, L.L.A.; Engel, M.F.M.; de Vries, R.D.; de Swart, R.L. In Vitro Modelling of Respiratory Virus Infections in Human Airway Epithelial Cells - A Systematic Review. Front Immunol 2021, 12, 683002. [Google Scholar] [CrossRef]
- Bauer, L.; Rijsbergen, L.C.; Leijten, L.; Benavides, F.F.; Noack, D.; Lamers, M.M.; Haagmans, B.L.; de Vries, R.D.; de Swart, R.L.; van Riel, D. The pro-inflammatory response to influenza A virus infection is fueled by endothelial cells. Life Sci Alliance 2023, 6, e202201837. [Google Scholar] [CrossRef]
- Glorieux, S.; Van den Broeck, W.; van der Meulen, K.M.; Van Reeth, K.; Favoreel, H.W.; Nauwynck, H.J. In vitro culture of porcine respiratory nasal mucosa explants for studying the interaction of porcine viruses with the respiratory tract. J Virol Methods 2007, 142, 105–112. [Google Scholar] [CrossRef]
- Krunkosky, M.; Krunkosky, T.M.; Meliopoulos, V.; Kyriakis, C.S.; Schultz-Cherry, S.; Tompkins, S.M. Establishment of Swine Primary Nasal, Tracheal, and Bronchial Epithelial Cell Culture Models for the Study of Influenza Virus Infection. J Virol Methods 2024, 327, 114943. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasan, C.C.; Thomas, M.; Antony, L.; Wormstadt, T.; Hildreth, M.B.; Wang, D.; Hause, B.; Francis, D.H.; Li, F.; Kaushik, R.S. Development and characterization of swine primary respiratory epithelial cells and their susceptibility to infection by four influenza virus types. Virology 2019, 528, 152–163. [Google Scholar] [CrossRef]
- Meliopoulos, V.; Cherry, S.; Wohlgemuth, N.; Honce, R.; Barnard, K.; Gauger, P.; Davis, T.; Shult, P.; Parrish, C.; Schultz-Cherry, S. Primary Swine Respiratory Epithelial Cell Lines for the Efficient Isolation and Propagation of Influenza A Viruses. J Virol 2020, 94, e01091–01020. [Google Scholar] [CrossRef]
- Meng, F.; Punyadarsaniya, D.; Uhlenbruck, S.; Hennig-Pauka, I.; Schwegmann-Wessels, C.; Ren, X.; Durrwald, R.; Herrler, G. Replication characteristics of swine influenza viruses in precision-cut lung slices reflect the virulence properties of the viruses. Vet Res 2013, 44, 110. [Google Scholar] [CrossRef]
- Wu, N.H.; Yang, W.; Beineke, A.; Dijkman, R.; Matrosovich, M.; Baumgartner, W.; Thiel, V.; Valentin-Weigand, P.; Meng, F.; Herrler, G. The differentiated airway epithelium infected by influenza viruses maintains the barrier function despite a dramatic loss of ciliated cells. Sci Rep 2016, 6, 39668. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, L.; Liu, B.; Feng, Y.; Zhou, H.; Zhang, Z.; Wu, Y.; Wang, J.; Gan, Y.; Yuan, T.; et al. Establishment and comparison of air-liquid interface culture systems for primary and immortalized swine tracheal epithelial cells. BMC Cell Biol 2018, 19, 10. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.Y.; Shin, D.L.; Li, G.; Wu, N.H.; Herrler, G. Time-dependent viral interference between influenza virus and coronavirus in the infection of differentiated porcine airway epithelial cells. Virulence 2021, 12, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Li, L.; Xue, M.; Zhao, L.; Liu, X.; Wang, W.; Feng, L.; Liu, P. Long-Term Expanding Porcine Airway Organoids Provide Insights into the Pathogenesis and Innate Immunity of Porcine Respiratory Coronavirus Infection. J Virol 2022, 96, e0073822. [Google Scholar] [CrossRef] [PubMed]
- Bonillo-Lopez, L.; Carmona-Vicente, N.; Tarrés-Freixas, F.; Kochanowski, K.; Martínez, J.; Perez, M.; Sibila, M.; Correa-Fiz, F.; Aragon, V. Porcine Nasal Organoids as a model to study the interactions between the swine nasal microbiota and the host. bioRxiv 2024, (preprint). [CrossRef]
- Davila, K.M.S.; Nelli, R.K.; Mora-Diaz, J.C.; Sang, Y.; Miller, L.C.; Gimenez-Lirola, L.G. Transcriptome Analysis in Air-Liquid Interface Porcine Respiratory Epithelial Cell Cultures Reveals That the Betacoronavirus Porcine Encephalomyelitis Hemagglutinating Virus Induces a Robust Interferon Response to Infection. Viruses 2024, 16, e939. [Google Scholar] [CrossRef]
- Bordes, L.; Gerhards, N.M.; Peters, S.; van Oort, S.; Roose, M.; Dresken, R.; Venema, S.; Vrieling, M.; Engelsma, M.; van der Poel, W.H.M.; et al. H5N1 clade 2.3.4.4b avian influenza viruses replicate in differentiated bovine airway epithelial cells cultured at air-liquid interface. J Gen Virol 2024, 105, e002007. [Google Scholar] [CrossRef]
- Sachs, N.; Papaspyropoulos, A.; Zomer-van Ommen, D.D.; Heo, I.; Bottinger, L.; Klay, D.; Weeber, F.; Huelsz-Prince, G.; Iakobachvili, N.; Amatngalim, G.D.; et al. Long-term expanding human airway organoids for disease modeling. EMBO J 2019, 38, e100300. [Google Scholar] [CrossRef]
- Heinen, P.P.; de Boer-Luijtze, E.A.; Bianchi, A.T.J. Respiratory and systemic humoral and cellular immune responses of pigs to a heterosubtypic influenza A virus infection. J Gen Virol 2001, 82, 2697–2707. [Google Scholar] [CrossRef]
- Liu, C.; Diong, X.; Liu, P.; Lin, X. Advances in porcine respiratory and intestinal organoids: status and potential application for virus infections. One Health Adv 2024, 2, 22. [Google Scholar]
- Rijsbergen, L.C.; Lamers, M.M.; Comvalius, A.D.; Koutstaal, R.W.; Schipper, D.; Duprex, W.P.; Haagmans, B.L.; de Vries, R.D.; de Swart, R.L. Human Respiratory Syncytial Virus Subgroup A and B Infections in Nasal, Bronchial, Small-Airway, and Organoid-Derived Respiratory Cultures. mSphere 2021, 6, e00237–00221. [Google Scholar] [CrossRef]
- Crisci, E.; Mussa, T.; Fraile, L.; Montoya, M. Review: influenza virus in pigs. Mol Immunol 2013, 55, 200–211. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Establishment of AO-derived WD-AECs. A) Schematic outline of the isolation of porcine bronchial epithelial cells from the primary bronchi which were subsequently grown as 3D organoids in extracellular matrix before seeding in 2D on transwell filters which were cultured at air-liquid interface upon confluency. B) Transverse histology sections over the course of 7 weeks showing the development of a pseudostratified ciliated respiratory epithelium. Hematoxylin and eosin stain, 40x objective. C) Immunohistochemistry (IHC) staining of WD-AECs 7 weeks post airlift and porcine bronchus epithelial cells. P63 staining to visualize basal cells, Muc5AC staining to visualize mucus (goblet cells) and acetylated tubulin staining to visualize cilia. WD-AEC resembles in vivo bronchial epithelium in cellular composition and morphology, despite a reduced thickness. (D) Development of transepithelial electrical resistance (TEER) over the course of differentiation of WD-AECs. After an initial increase post airlift, TEER values remained consistent. For B, C and D: Differentiation was followed for three separate donors. representative data from one experiment are shown.
Figure 1.
Establishment of AO-derived WD-AECs. A) Schematic outline of the isolation of porcine bronchial epithelial cells from the primary bronchi which were subsequently grown as 3D organoids in extracellular matrix before seeding in 2D on transwell filters which were cultured at air-liquid interface upon confluency. B) Transverse histology sections over the course of 7 weeks showing the development of a pseudostratified ciliated respiratory epithelium. Hematoxylin and eosin stain, 40x objective. C) Immunohistochemistry (IHC) staining of WD-AECs 7 weeks post airlift and porcine bronchus epithelial cells. P63 staining to visualize basal cells, Muc5AC staining to visualize mucus (goblet cells) and acetylated tubulin staining to visualize cilia. WD-AEC resembles in vivo bronchial epithelium in cellular composition and morphology, despite a reduced thickness. (D) Development of transepithelial electrical resistance (TEER) over the course of differentiation of WD-AECs. After an initial increase post airlift, TEER values remained consistent. For B, C and D: Differentiation was followed for three separate donors. representative data from one experiment are shown.
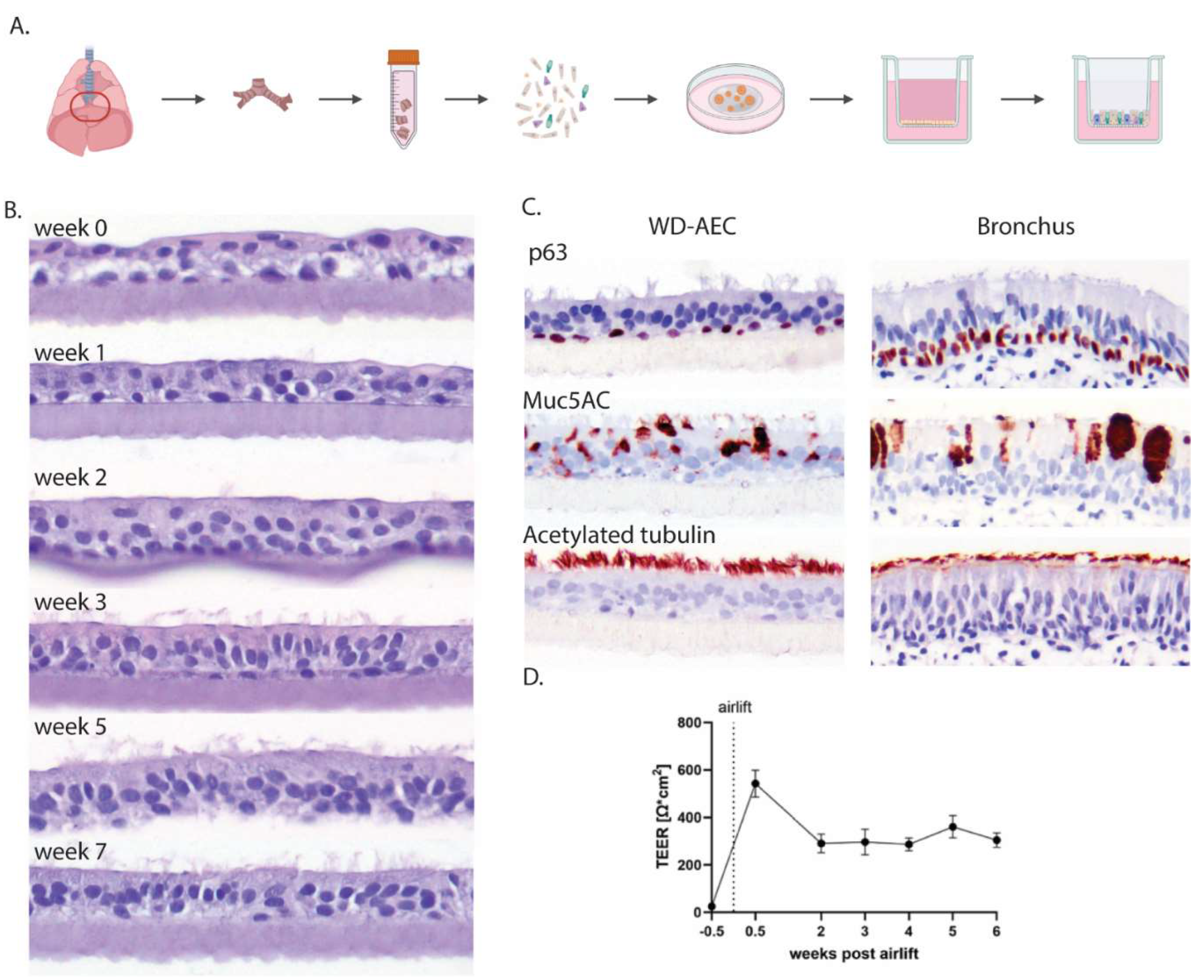
Figure 2.
Development of WD-AECs over time. Confocal laser scanning microscopy images showing the development of tight junctions (ZO-1), goblet cells (Muc5ac) and cilia (acetylated tubulin) over the course of 6 weeks after airlifting transwell filters. Goblet cells were most abundant at 2-4 weeks, while cilia appeared after 3 weeks. Tight junctions were detected from week 0 onwards, but only started to show a regular pattern from week 2 onwards. Merged images show nuclei in blue, tight junctions in red, goblet cells in green and cilia in yellow. Scale bar represents 100µm. 40x objective. Representative data from one experiment are shown.
Figure 2.
Development of WD-AECs over time. Confocal laser scanning microscopy images showing the development of tight junctions (ZO-1), goblet cells (Muc5ac) and cilia (acetylated tubulin) over the course of 6 weeks after airlifting transwell filters. Goblet cells were most abundant at 2-4 weeks, while cilia appeared after 3 weeks. Tight junctions were detected from week 0 onwards, but only started to show a regular pattern from week 2 onwards. Merged images show nuclei in blue, tight junctions in red, goblet cells in green and cilia in yellow. Scale bar represents 100µm. 40x objective. Representative data from one experiment are shown.
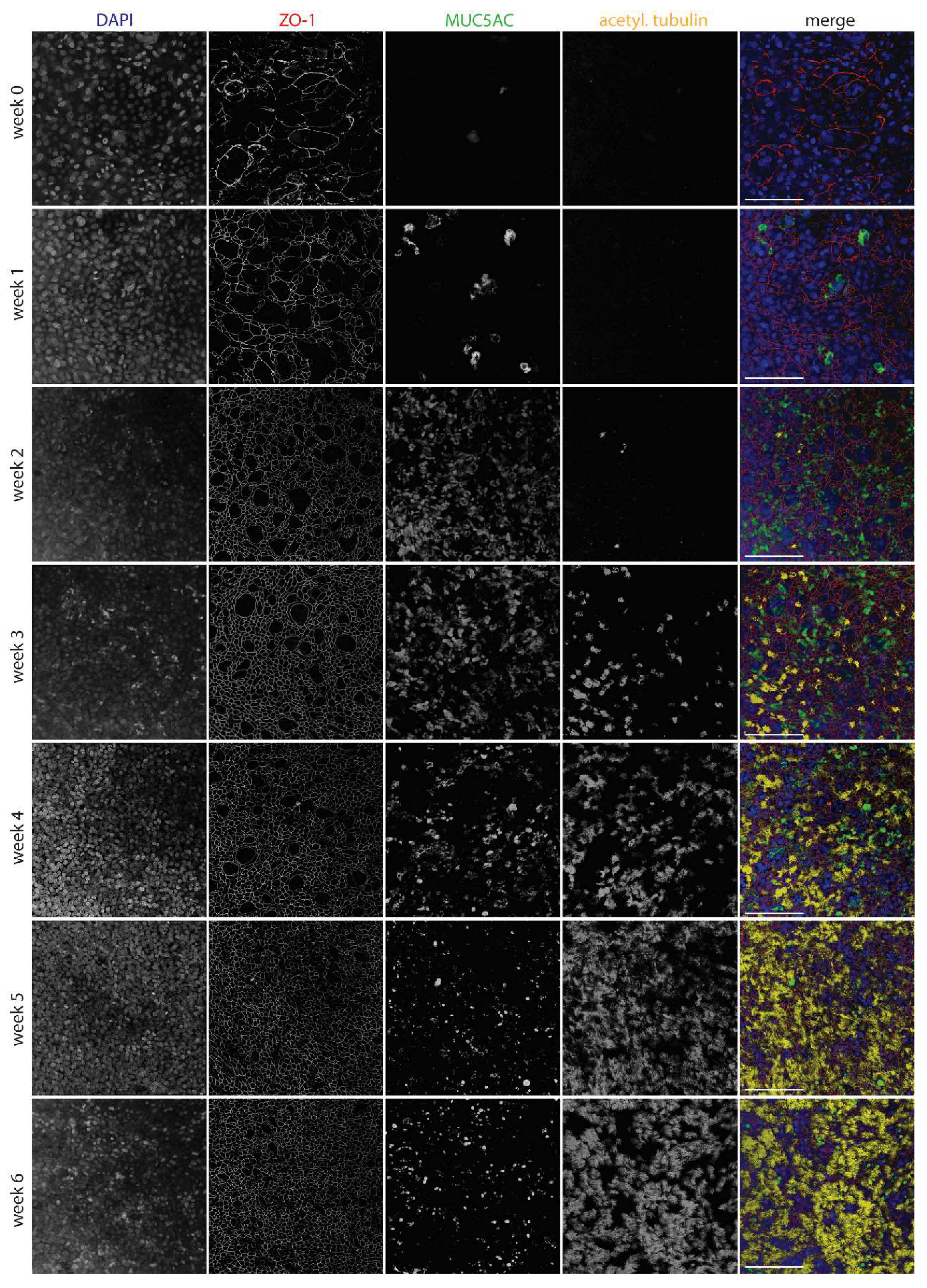
Figure 3.
Infection of porcine WD-AECs with IAVsw. (A) Experimental design of IAVsw inoculation of porcine WD-AEC cultures. Three different IAVsw strains were used (H1N1, H1N2, H3N2) and applied apically at a dose of 102 or 103 TCID50 in a volume of 10µl. After 1h incubation, transwell filters were washed 4 times. An apical wash was collected at 16h, 24h and 48h post inoculation, and transepithelial electrical resistance (TEER) was measured at 0h, 24h and 48h. At 48h, filters were fixed and stained by hematoxylin and eosin (HE) and against IAV nucleoprotein (NP). (B) Viral RNA loads in apical washes. All viruses replicated in porcine WD-AECs. For H1N2, a dose response curve was observed over all evaluated time points, which was only observed for the earlier time points for the other two viruses. (C) TEER values as a measure of epithelial integrity. TEER values declined for H3N2- and H1N1-inoculated WD-AECs over 48h, but remained constant in H1N2-inoculated WD-AECs. (D) Transverse histology sections stained by HE at 48h post infection. The epithelial cell layer remained intact for H1N2-inoculated WD-AECs, while there was substantial thinning of the cell layer in H3N2-inoculated WD-AECs. For H1N1, the higher dose seemed to cause more cell loss compared to the lower dose. 40x objective. B, C and D show representative results from one out of three experiments (pig04).
Figure 3.
Infection of porcine WD-AECs with IAVsw. (A) Experimental design of IAVsw inoculation of porcine WD-AEC cultures. Three different IAVsw strains were used (H1N1, H1N2, H3N2) and applied apically at a dose of 102 or 103 TCID50 in a volume of 10µl. After 1h incubation, transwell filters were washed 4 times. An apical wash was collected at 16h, 24h and 48h post inoculation, and transepithelial electrical resistance (TEER) was measured at 0h, 24h and 48h. At 48h, filters were fixed and stained by hematoxylin and eosin (HE) and against IAV nucleoprotein (NP). (B) Viral RNA loads in apical washes. All viruses replicated in porcine WD-AECs. For H1N2, a dose response curve was observed over all evaluated time points, which was only observed for the earlier time points for the other two viruses. (C) TEER values as a measure of epithelial integrity. TEER values declined for H3N2- and H1N1-inoculated WD-AECs over 48h, but remained constant in H1N2-inoculated WD-AECs. (D) Transverse histology sections stained by HE at 48h post infection. The epithelial cell layer remained intact for H1N2-inoculated WD-AECs, while there was substantial thinning of the cell layer in H3N2-inoculated WD-AECs. For H1N1, the higher dose seemed to cause more cell loss compared to the lower dose. 40x objective. B, C and D show representative results from one out of three experiments (pig04).
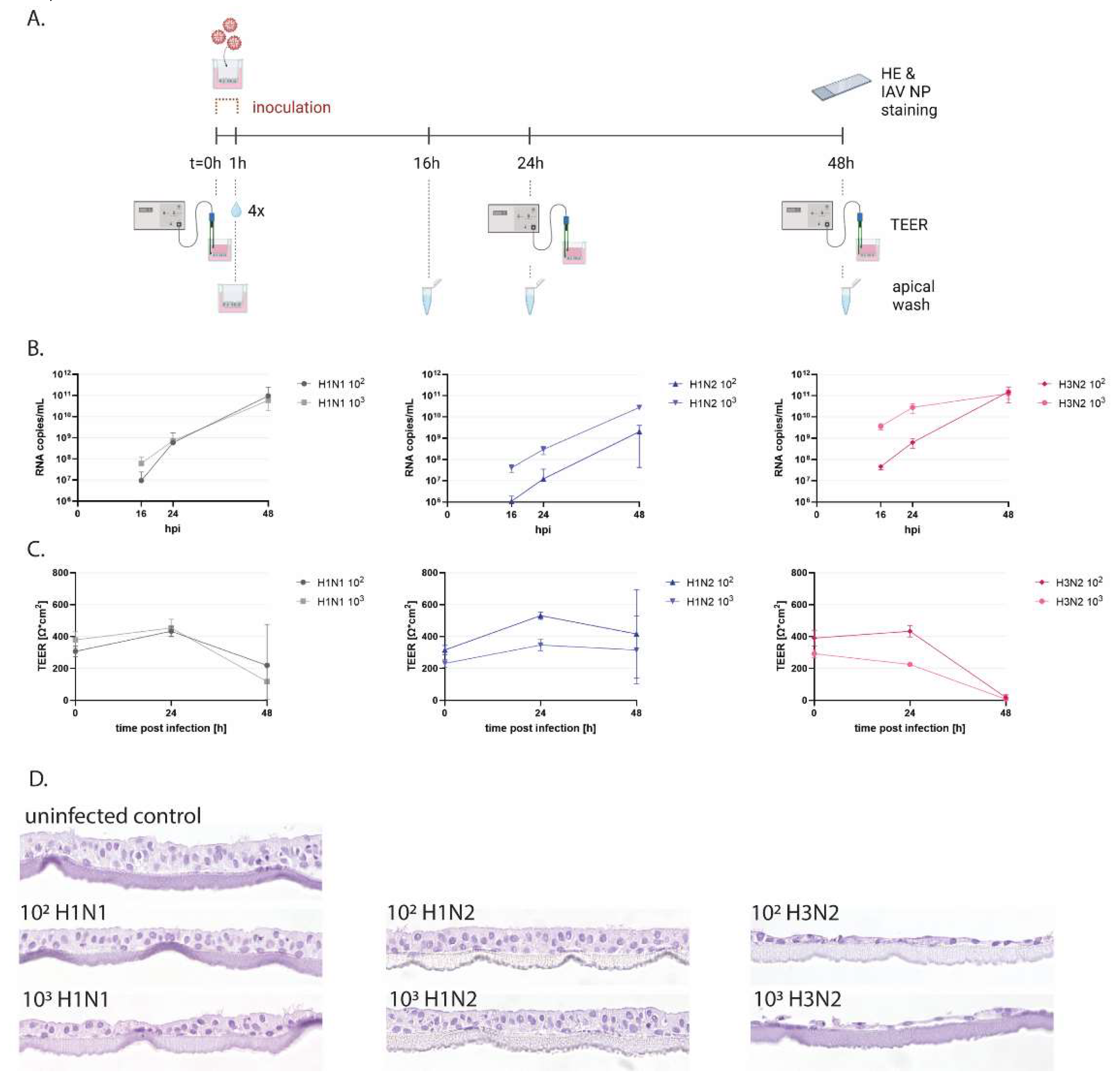
Figure 4.
IAV NP expression after 24h post infection with 103 TCID50. Confocal microscopy images to visualize expression of IAV NP and epithelial composition. (A) H1N1-inoculated WD-AECs showed NP expression (green), integer tight junctions (red), a thick layer of cilia (yellow) and predominantly intact nuclei (blue). (B) H3N2-inoculated filters showed NP expression, while at the same time tight junctions were compromised, cilia expression was reduced and more fragmented nuclei were visible. (C) Angled side-view of WD-AEC shown in (A). (D) Angled side-view of WD-AEC shown in (B). Note that IAV NP staining (green) is visible within or above the apical side of WD-AEC. All images: Scale bar represents 50µm. 100x objective. Representative data from one experiment are shown.
Figure 4.
IAV NP expression after 24h post infection with 103 TCID50. Confocal microscopy images to visualize expression of IAV NP and epithelial composition. (A) H1N1-inoculated WD-AECs showed NP expression (green), integer tight junctions (red), a thick layer of cilia (yellow) and predominantly intact nuclei (blue). (B) H3N2-inoculated filters showed NP expression, while at the same time tight junctions were compromised, cilia expression was reduced and more fragmented nuclei were visible. (C) Angled side-view of WD-AEC shown in (A). (D) Angled side-view of WD-AEC shown in (B). Note that IAV NP staining (green) is visible within or above the apical side of WD-AEC. All images: Scale bar represents 50µm. 100x objective. Representative data from one experiment are shown.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



