
Submitted:
07 October 2024
Posted:
08 October 2024
You are already at the latest version
Abstract
The global biodiversity crisis has emphasized the unique contribution of functional diversity to ecosystem function, stability, and resilience. In this regard, the increasing availability of remotely sensed data together with the development of new sensors and approaches has the potential to improve our ability to quantify and monitor ecosystem traits and functions at unprecedent spatial, temporal, and spectral scales. In particular, air- and spaceborne hyperspectral data are making possible the measurement of plant-level functional traits to investigate ecosystem function and functional diversity in novel ways. In this review we pose that these developments together with similar ones focusing on soils and lithologies can help us understand relationships between functional diversity, ecosystem processes, and landsliding, and more broadly the disturbance dynamics of mountainscapes. Acknowledging the challenges associated with mountainscapes, this review 1) synthesizes broad established methods to retrieve functional traits from remotely sensed data, 2) summarizes approaches derived from remotely sensed data to characterize functional diversity and its relationships with ecosystem functioning, and 3) elaborates on how these methods and approaches can provide a needed holistic view of landslides. This holistic view recognizes the critical role that interactions between ecosystem and geomorphic processes play in the dynamics of mountainscapes mediated by landslides and the contribution of ecosystem diversity and processes to landslide susceptibility and recovery. In this “ecosystem-centered” view of landslides it might be necessary to scale from individual landslides and sites to entire landslide populations, assemblages, and landscapes.
Keywords:
functional diversity
; hyperspectral remote sensing
; landslides
; LiDAR
; plant functional traits
; soil-rock attributes
; spectral diversity
; tropical mountainscapes
1. Introduction
The global biodiversity crisis has emphasized the contribution of biodiversity to ecosystem stability and resilience, and highlighted monitoring needs at multiple scales [1,2]. In this context, functional diversity (FD) has emerged as an essential dimension of biodiversity due its potential to link ecosystem processes and function (EF) with taxonomic, phenotypic, and genomic diversity [3,4]. Remotely sensed multi-spectral data collected along the visible (VIS) through the long-waved infrared (LWIR) regions has already made possible the characterization of ecosystem traits and functions at several spatial and temporal scales [5,6,7,8,9,10,11], contributing in important ways to monitoring the Earth’s “pulse.” Simultaneously, important developments in imaging spectroscopy have made possible linking field-based leaf- and plant-level functional trait (FT) measurements with air- and spaceborne hyperspectral data to investigate ecosystem function and functional diversity at unprecedented spectral scales [12,13,14,15,16]. Here we pose that these developments together with similar ones focusing on soils and lithologies [17,18,19] can help us understand relationships between, functional diversity ecosystem processes, and landsliding, and more broadly the disturbance dynamics of mountainscapes.
The aforementioned sources of remotely sensed data to investigate functional diversity come with distinctive tradeoffs stemming from their spatial, temporal, and spectral resolutions [20,21,22]. A first set of trade-offs associated with multi-spectral data involves downscaling the data based on band-sharpening or bilinear interpolation methods to match the smallest field sampling unit used to characterize functional traits and functional diversity [e.g., 5,23,24]. Alternatively, downscaling may involve modeling fractional cover using spectral mixture analyses (SMA) to characterize pixel level plant functional types (PFT) and substrate composition, and ultimately habitat diversity [e.g., 25]. In both instances, subsequent upscaling using environmental data allows examination of diversity patterns at landscape and regional scales. A second set of trade-offs is associated with hyperspectral data and there is variation depending on whether the data is collected by aircrafts or satellites. The former may require upscaling to characterize functional traits and functional diversity from image scenes to regional scales [12,26,27]. The latter, like multi-spectral data, may require downscaling [28] or modeling fractional cover [29,30,31] to characterize functional traits and functional diversity at different scales. These tradeoffs may be amplified in studies investigating the relationships between ecosystems and landsliding given the challenges associated with the complex topography of mountainscapes [32,33], the large variation in landslide sizes [34,35], and the spectral characteristics of young landslides that contain a mixture of vegetated pixels and bare soil. In vegetation studies based on hyperspectral data it is recommended to remove bare soil pixels to eliminate background reflectance [36]; similarly in soil/rock studies the recommendation is to remove vegetated pixels [37].
A new generation of existing and planned spaceborne hyperspectral missions such as PRISMA [38], EnMAP [39], SBG [40,41], CHIME [42], and Planet’s Tanager satellites [43] open numerous possibilities for investigating taxonomic and functional diversity, and ecosystem function at a time of rapid global change. Although the challenges of spatial resolution may persist, the combination of this data with soil and lithological variables derived in a similar fashion [17,18,19] may enable the multiscale monitoring of tropical mountains, a mega diverse and climate-susceptible biome heavily influenced by numerous drivers of change, including landsliding.
Connecting functional diversity and ecosystem function hinges both on the selection of relevant functional traits at leaf- and plant-levels and linking these traits with their optical properties, and ultimately to air- and spaceborne sensors [Figure 1; 16,44]. This has involved the collection of relevant trait data in the field, development of models associating these traits with their optical properties, scaling functional traits, and assessing functional groups and functional diversity at multiple scales [15,26]. In this review we 1) synthesize broad established methods to retrieve functional traits from remotely sensed data, 2) summarize approaches derived from remotely sensed data to characterize functional diversity and its relationships with ecosystem functioning, and 3) elaborate on how these methods and approaches can provide a needed holistic view of landslides. This holistic view recognizes the critical role that interactions between ecosystem and geomorphic processes play in the dynamics of mountainscapes mediated by landslides and the contribution of ecosystem diversity and processes to landslide susceptibility and recovery. In this “ecosystem-centered” view of landslides it might be necessary to scale from individual landslides and sites to entire landslide populations, assemblages, and landscapes.
2. Functional Traits – From Field to Remotely Sensed Observations
In a broad sense, functional traits refer to attributes that play a role in the establishment, growth, survival, reproduction, and ultimately fitness, of an organism [45]. In a narrow sense, functional traits represent a subset of biochemical, morphological, physiological, and behavioral attributes that mediate organisms’ responses to environmental stressors, including disturbance, and have a marked effect on ecosystem functions and ecosystem stability at multiple scales [46,47,48,49,50]. Thus, upscaling functional trait measurements to regions [e.g., 51,52] can greatly help understand underlying causes of the observed variability, as well as consequences under global changes. In this regard, imaging spectroscopy is providing unique opportunities to observe spectral features [53,54] representative of leaf- and plant-level traits [44,55,56] that mediate ecosystem functions [12,15]. Moreover, imaging spectroscopy is providing opportunities to explore variation of spectral features in relation to geomorphic [57] and environmental [58,59] gradients, landscape heterogeneity [60], and spread of invasive species [61,62]. In addition to suggesting underlying causes of the observed variation, this work may also help understand the consequences of changes in the underlying drivers on trait distributions, and ultimately ecosystem functions which becomes important for monitoring and forecasting purposes [63,64,65] as well as for calibrating ecosystem and biosphere models [29,66,67,68].
Observing spectral features relies on the selection of leaf- and plant-level traits (Figure 1 Stage 1a) that collectively translate into critical ecosystem processes and functions, retrieval of this information from optical data using diverse approaches (Figure 1 Stage 1b and Stage 2), and scaling from leaf and plant to pixel, plot, landscape, and region levels (Figure 1 Stages 3 and 5e). Functional traits at the leaf level include pigments (Chlorophyll, Carotenoids, Anthocyanins), nutrients (Nitrogen, Phosphorus, Calcium, Potassium, Magnesium, Iron), structural compounds (Lignin, Cellulose, Polyphenols), water content (Equivalent Water Thickness, EWT), and mass per unit area (LMA) whereas at the plant level, tree height and canopy characteristics (e.g., size and shape, leaf area index, leaf angle distribution, fractional cover) [15,36,55,56,68,69,70,71]. At the individual level, both sets of traits are related to light capture and growth, photoprotection, stress resistance and defense, maintenance and metabolism. At the ecosystem level these traits inform about photosynthesis, primary production, carbon storage, nutrient cycling, decomposition, allocation and growth, stress resistance, and hydraulic regulation.
The retrieval of leaf- and plant-level functional traits from optical data is based on the development of relationships between trait measurements and individual bands, spectral indices, or the whole spectra via three broad sets of modeling approaches, namely statistical, physical-based, and hybrid [Figure 1 Stage 2; 16,65,72-76]. Statistical approaches involve models that establish relationships between a trait of interest and individual bands and spectral indices [66,77,78,79], uncorrelated, synthetic variables derived through principal component analysis (PCA) [53,80], or the entire spectrum using partial least squares regression (PSLR) [e.g., 68,81,82,83]. Physical-based approaches make use of coupled leaf-canopy radiative transfer models (RTM) to estimate canopy reflectance via forward mode or leaf traits by model inversion; these models require look-up tables as well as trait data sets calibrated both in the lab and field [84,85,86,87]. Finally, hybrid models combine both approaches [88,89,90].
Scaling and mapping plant functional traits (Figure 1 Stages 1-3 and 5e) are often part of large studies aimed at the development of new sensors, complex instruments, and applications [67,91,92,93], yet they may also take place in other contexts [94]. These studies consider retrieval methods, conduct simulations, collect in situ field and sensor calibration and validation measurements, and pre-process the imagery to different degrees. Central to scaling and mapping are in situ measurements of canopy reflectance at relatively homogenous sampling points using field spectroradiometers, or complex airborne instruments, and/or estimates derived from RTM models (Figure 1 Stage 1). Sampling points may represent elementary sampling units (ESU) within plots matching pixels of a given airborne or spaceborne sensor, tree crown pixels or full tree crowns, and field subplots within plots whose size and shape may represent pixels of a given sensor [23,68,70,93,95]. Extensive pre-processing of air- and spaceborne multispectral and hyperspectral data, may involve data fusion, estimating fractional vegetation cover, masking out clouds, bare ground, and tree-canopy shade, delineating tree canopies, and characterizing canopy geometry [Figure 1 Stage 1b; 15,91,96] [Figure 1 Stage 1b; 12,92,96]. At this stage, the models linking optical data with leaf- and plant-level traits are applied to the pre-processed remotely sensed data to scale up predictions and map these traits at pixel, plant, image scene, and region levels [Figure 1 Stage 3; 16,36,70]. Scaling may involve averaging pixel-level traits for individual tree crowns [70,97] or averaging the spectra of multiple pixels within sampling plots [12,36] or weighing vegetation indices or traits by the corresponding plant abundance [e.g., tree crown area; 68,98]. At scene and region scales, scaling up leaf- and plant-level traits may involve additional modeling using ancillary environmental data and resampling at resolutions lower than the original hyperspectral data [Figure 1 Stages 3 and 5e; 36,81,99].
3. Functional Diversity and Ecological Function
Functional diversity is broadly defined as the value and range of functional traits in a community [100,101]. In addition to facilitating comparisons at multiple scales, theoretical and empirical work suggests that functional diversity and species diversity are related [102,103,104], that intensity of disturbance influences functional diversity [105,106], that functional diversity greatly contributes to the stability of ecological function [101,107,108,109,110], and that the diversity of functions both within and across scales are central to ecosystem resilience [110,111]. This may explain efforts to examine variation of functional diversity in space [109,112,113,114] and time [115,116] in response to changing environmental conditions, characterize within (local; alpha diversity), between (landscape; beta diversity), and overall (regional; gamma diversity) functional diversity [117,118,119], and to develop qualitative and quantitative measures of functional diversity whether grouping species based on trait similarity or defining indices or metrics [120].
Using single or multiple functional traits it is possible to identify functional groups, i.e., groups of species sharing similar traits and most likely performing similar functions or exhibiting similar ecological strategies within ecosystems [Figure 1 Stage a; 110,121,122-126]. One large-scale functional grouping of plants uses categorical traits such as life form, leaf type, and leaf phenology to identify plant functional types (PFT); these traits are currently retrievable from remotely sensed data [127]. Another functional grouping of plants used leaf functional traits derived from airborne hyperspectral data and a cluster analysis to identify functional forest classes (FFC) and functional forest groups (FFG) that were used to characterize functional diversity at a country-scale [12]. These groupings can be incorporated into metrics that estimate functional richness (number of functional groups) or evenness (distribution of abundances of functional groups) at community scales [120,128]. In the context of this work, estimating functional diversity requires the identification of communities or pixel neighborhoods [54].
Functional diversity is more commonly expressed with metrics or indices based on single or multiple functional traits that can take discrete and/or continuous values [129,130,131]. A metric based on single functional traits is the community weighted mean of trait values (CWM) that expresses the importance of a trait at plot scales by weighing them by species or functional type abundances [Figure 1 Stage 5b; 5,114]. Concurrent CWM of trait values derived from field measurements and high resolution hyperspectral airborne data were used in a PLSR model to subsequently scale CWM of trait values to image scales [68]. Similarly, concurrent CWM of trait values, and high resolution multispectral spaceborne and environmental data were used as input in a Geographic Random Forest to map trait values in tropical regions [5].
Metrics using multiple functional traits in a multidimensional framework describe the trait space occupied by species in a community using a convex hull (functional richness, FRic), the regularity of the spacing of the species (functional evenness, FEve), the mean distance of the species to the centroid of the trait space weighted by the relative abundance of each species (functional dispersion; FDis), and the degree to which species abundances are distributed towards the boundaries of the trait space (functional divergence; FDiv) [Figure 1 Stage 5b; 53,120,129,131] [Figure 1 Stage 5b; 53,120,129,130]. Alternatively, these indices may be estimated using a trait probability density (TPD) approach [117]. Using airborne LiDAR and hyperspectral data, as well as spaceborne multi-spectral data, Schneider et al. [15] and Helfenstein et al. [20] derived morphological and functional traits based on band ratios at pixel-scales, and used pixels and pixel neighborhoods as a surrogate for species and communities, respectively, to estimate FRic, FEve, and FDiv while exploring the influence of spatial (changes in neighborhood size) and spectral (convolving airborne hyperspectral data) scale; to visualize pixel-scale trait composition the authors used an RGB composite. In contrast, Zheng et al. [98] used LiDAR data to delineate tree crowns, characterized them based on morphological traits and the CWM of three band ratios, and subsequently estimated the same functional diversity metrics. Using a similar approach and hyperspectral data collected by an unoccupied aerial vehicle (UAV), Cimoli et al. [132] estimated FRic based on trait probability densities as the percentage of trait space occupied by a minimum density of pixels [TPD; 117]. Finally, using a radiative transfer model Pacheco-Labrador et al. [89] generated synthetic communities and found that the “observed” and “estimated” functional diversity metrics were in most instances well correlated, yet spatial but not spectral resolution and noise affected these correlations; similar results were found for spectral diversity metrics. More importantly, some functional diversity metrics could be predicted by spectral diversity metrics, yet the best correlations were observed at small or local scales. Here spectral diversity refers to the variability in spectral reflectance which has been increasingly recognized as an important component of biodiversity because it integrates information on plant taxonomic, phylogenetic, and functional diversity [133,134,135,136] and is a predictor of functional diversity [137].
In the previous studies, changes in neighborhood or pixel size were used to examine the influence of space on functional diversity metrics. Another approach involves partitioning functional diversity into alpha, beta, and gamma components (Figure 1 Stage 5b). Using three structural variables derived from spaceborne lidar, Schneider et al. [138] used a TPD approach to estimate alpha and beta functional diversity, the latter as the non-overlapping areas of two trait probability distributions representing two contiguous neighborhoods. Using high resolution hyperspectral data and principal component analyses at each pixel (species) in a given neighborhood (plot), Laliberté and collaborators [53,139] examined the within (alpha diversity), between (beta diversity), and overall (gamma diversity) plot variability of the principal components (Figure 1 Stage 5c). Similarly, Rossi et al. [54] used Rao’s quadratic entropy and spectral reflectance to derive alpha, beta, and gamma spectral diversity from Sentinel 2 multi-spectral data.
4. Diversity, Landsliding, and Mountainscapes
Early we posed that remotely sensed hyperspectral data may help understand the extent to which ecosystem and geomorphic processes interact to drive landsliding, and ultimately the large-scale dynamics of mountainscapes. Landsliding is a major disturbance transforming mountains worldwide and their ecological significance remains poorly understood [Figure 3; 140,141,142]. First, landslides create new substrates upon which communities and ecosystems reorganize and recover [141,142]. These substrates represent soil horizons exhumed all the way down to altered bedrock or even minimally weathered bedrock. Second, these substrates are heterogenous at local (individual landslides), regional (landslide populations), and landscape (landslide assemblages) scales [34,143,144]. Third, landslides leave long lasting legacies that may influence the process of landsliding itself [145,146]. Fourth, landsliding may contribute in important ways to the diversity [147,148,149,150] and functioning [151,152,153,154] of montane ecosystems. Whereas the creation of unique habitats and landscape configurations favors subsets of unique species, changes in abiotic conditions directly influence carbon and nutrient cycling, and rock weathering. Lastly, ecosystems may influence slope resistance and susceptibility to failure in complex ways given the dynamic nature of vegetation-soil-saprolite interactions during ecosystem development [141]. Advances in hyperspectral sensors, platforms, and data analytical approaches (Figure 1) can be used to characterize the vegetation based on functional traits, as well as the underlying soils and lithology based on physical and geochemical attributes. Together this has the potential to inform about the diversity and functioning of montane ecosystems at scales commensurate with landslide activity, and ultimately the long-term functional significance of landslides in mountainscapes.
4.1. Plant traits, Montane Ecosystems, and Landsliding
In mountainous environments, multi- and hyperspectral remote sensing has contributed to the characterization of plant communities [155,156,157,158], land cover/land use [159,160,161,162], tree line dynamics [33], and disturbance regimes including deforestation, forest die-off, and landsliding [Figure 2 and Figure 3; 163,164-167]. In particular, the efforts towards landslide mapping have been substantial [168]. In contrast, the use of multi- and hyperspectral remote sensing to characterize functional traits, functional diversity, and ecosystem function in mountainous environments, particularly in the context of landslide studies, has been limited.
Figure 2.
Functional and Spectral Diversity in Landslide-Impacted areas in the Sierra de las Minas of Guatemala (SLM) derived from PRISMA data. True-color RGB images (TCI) for A. a large portion of the SLM and B. True-color RGB images (TCI) of two focal regions used in C-D. C. RGB composite of three vegetation indices [normalized difference water index (NDWI), Anthocyanin Content Index (ACI), and Vogelman Index 2 (VOG)] and derived spectral alpha diversity (FT-SDα). D. RGB composite of three principal components (PCs) and derived spectral alpha diversity (PC-SDα).
Figure 2.
Functional and Spectral Diversity in Landslide-Impacted areas in the Sierra de las Minas of Guatemala (SLM) derived from PRISMA data. True-color RGB images (TCI) for A. a large portion of the SLM and B. True-color RGB images (TCI) of two focal regions used in C-D. C. RGB composite of three vegetation indices [normalized difference water index (NDWI), Anthocyanin Content Index (ACI), and Vogelman Index 2 (VOG)] and derived spectral alpha diversity (FT-SDα). D. RGB composite of three principal components (PCs) and derived spectral alpha diversity (PC-SDα).

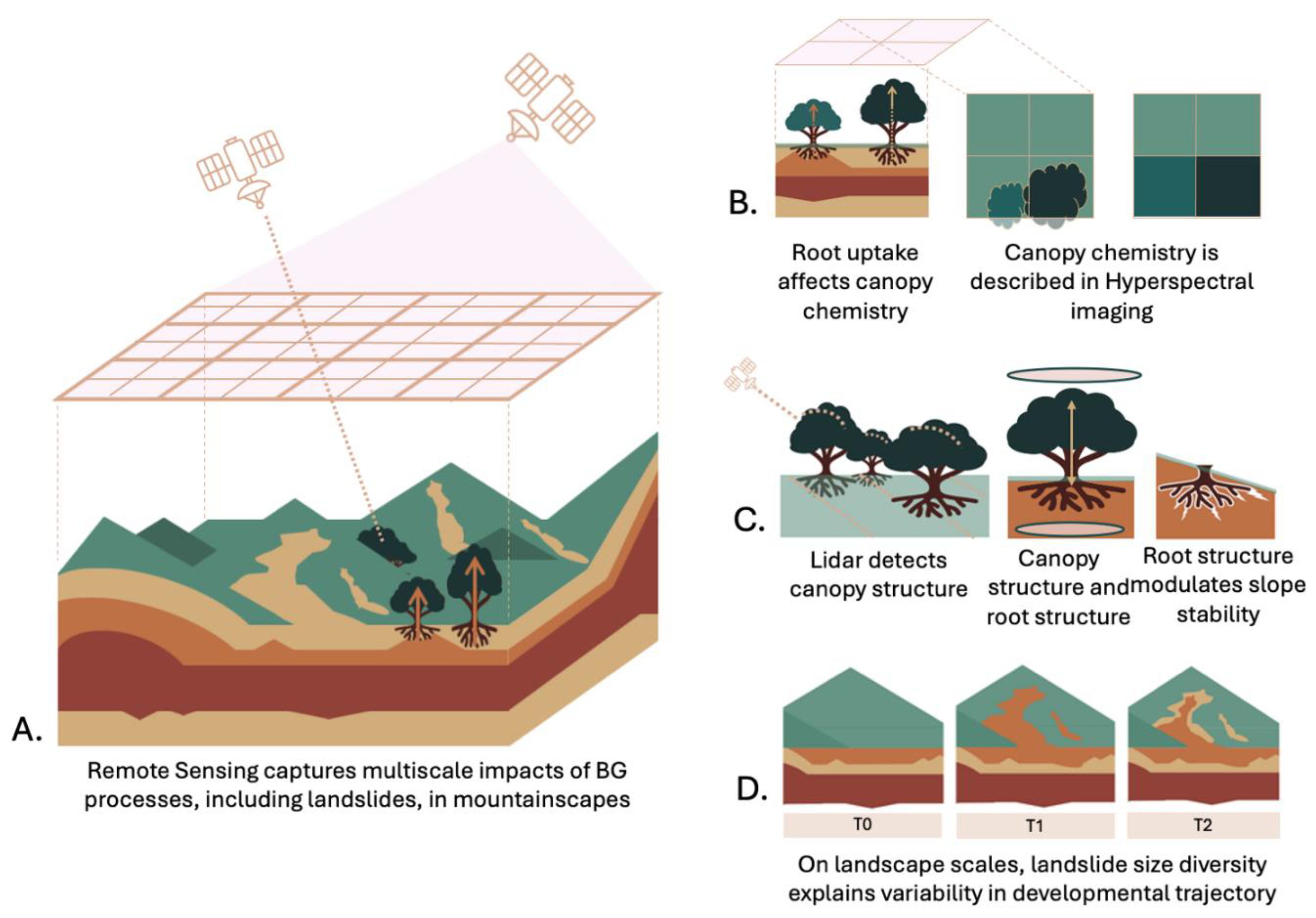
Figure 3.
Ecosystem-centered view of landslides can greatly benefit from new remote sensing data and missions A. Remotely sensed hyperspectral and LiDAR data captures landslides, vegetation, soil, and rock attributes at multiple scales. Integrated over space and time this data can inform ecosystem, biogeochemical, and geochemical processes. B. Parent materials influence soil and ecosystem properties. Root uptake of rock-derived nutrients influences leaf chemistry, which can be detected with hyperspectral data. C. LiDAR can be used to describe canopy structural variables which in turn may correlate with root structure [234]. Root structure in turn influences slope stability, affecting landsliding, which sets in motion various geochemical, biochemical and biological processes. D. At landscape scales, landslide age and size diversity explain variability in developmental trajectory in landscapes..
Figure 3.
Ecosystem-centered view of landslides can greatly benefit from new remote sensing data and missions A. Remotely sensed hyperspectral and LiDAR data captures landslides, vegetation, soil, and rock attributes at multiple scales. Integrated over space and time this data can inform ecosystem, biogeochemical, and geochemical processes. B. Parent materials influence soil and ecosystem properties. Root uptake of rock-derived nutrients influences leaf chemistry, which can be detected with hyperspectral data. C. LiDAR can be used to describe canopy structural variables which in turn may correlate with root structure [234]. Root structure in turn influences slope stability, affecting landsliding, which sets in motion various geochemical, biochemical and biological processes. D. At landscape scales, landslide age and size diversity explain variability in developmental trajectory in landscapes..
Studies of montane ecosystem function based on remote sensing have relied on vegetation indices (Normalized Vegetation Index-NDVI, Enhanced Vegetation Index-EVI, Normalized Difference Water Index-NDWI) and whole spectra to characterize canopy chemistry. Variation in time and space of phenological metrics derived from NDVI have been used to infer temperature sensitivity of mountain vegetation [169]. Similarly, NDVI and EVI have been used to estimate above-ground biomass and leaf area index of non-forest and forest montane ecosystems [9,170,171,172,173]. Air- and spaceborne hyperspectral data collected over temperate and tropical hilly and mountainous ecosystems has allowed landscape- to regional-level quantification of canopy nitrogen and lignin, while improving our understanding of nutrient dynamics [Figure 2; 58,64,174,175]. Extensive work with airborne hyperspectral data in Peru (eastern Andes) and Malaysia (Mount Kinabalu) has contributed unique insights into remotely sensed leaf chemistry associated with rock-derived nutrients [Figure 3b; 26,70,97,176]. Altogether, these studies have shed light into the contribution of abiotic (temperature, geomorphic and topographic settings) and biotic (species composition) factors driving trait variability and ecosystem, soil, geomorphic, and hydrologic processes.
The few studies using remotely sensed traits to characterize functional diversity in mountainous regions have used different traits, metrics and approaches, yet their underlying motivation has been to derive large-scale relationships between diversity and ecosystem function (BEF studies). Two studies conducted in Switzerland (Laegern Mountain) showed that functional richness (FRic) responded stronger to environmental conditions than functional evenness (FEve) and functional divergence (FDiv) [15,20]. More specifically, FRic was highest on the south aspect and lowest at the mountain ridge. At this same site, Schneider and collaborators [177] simulated gross primary productivity (GPP) and found that it was positively related with FRic; yet, there were large differences among sites differing with elevation. A fourth study conducted in China (southern aspect of Mt. Shennongjia) showed that FRic was highest at mid and high elevations [24]. Perhaps more importantly, this work revealed a humped-back relationship between above ground biomass and FRic derived from morphological traits, and a more complex, non-lineal relationship between productivity (NDVI) and FRic derived from physiological traits. Using a different approach in the eastern Andes of Peru, Duran and collaborators [178] found that the CWM for three leaf traits (LMA, non-structural carbohydrates, and percent water) were positively related to elevation, whereas a fourth one (Chlorophyll) and FRic negatively related. This work further showed that net primary (NPP) and gross primary (GPP) productivity were positively related with FRic. Interestingly, in the aforementioned work there is no reference to landslides.
The closest reference to functional traits retrieved from remotely sensed data in landslide studies include GRVI (green-red vegetation index), NDVI (Table S1), and tree height [179]. The former was used in combination with back estimations of lateral apparent cohesion contributed by roots to understand slope resistance/susceptibility to intense rainfall events in China [180]. In contrast, NDVI and tree height are used in studies characterizing vegetation recovery and its role in stabilizing sediment fluxes and sequestering carbon, respectively. A common metric used, or that can be derived, from studies using NDVI is the Vegetation Recovery Ratio (VRR), a ratio of the vegetation gained over time relative to the vegetation that was lost (Table S1). Some interesting observations, as well as limitations, emerge from this work. First, the variability of NDVI within [181] and among studies (Table S1) can be substantial. Second, the presence of active and inactive landslides, together with the variability of VRR in any given population, suggests that successional trajectories are highly variable which may explain the within-study NDVI variability [e.g., 181]. Third, NDVI0, VRR, and annual VRR varied among studies, in part reflecting differences among sites but also, the time covered by the studies. Fourth, VRR decreases with elevation and slope [182]. Fifth, rainfall-triggered landslides may recover faster than those triggered by earthquakes [183]. Finally, the limited use of ecological principles precludes a deeper understanding of ecosystem-geomorphic interactions. For example, NDVI is not modeled to characterize ecosystem development, nor is it related to variables such as above ground biomass or tree height that are informative of important ecosystem processes. Similarly, the variability of NDVI at the scale of individual landslides may be used to characterize functional diversity; integrated over time and across landslides within a population may help understand the contribution of landslides to the diversity of montane ecosystems. In contrast to the aforementioned studies, Freund and collaborators [179] focused on a landslide assemblage to examine changes in vegetation height and above ground biomass in landslides of different ages along a broad elevation gradient in Peru's Kosñipata Valley. These authors combined a 1 m-resolution top-of-canopy height (TCH) raster derived from high-resolution full waveform airborne LiDAR data [184] with mapped landslides. These were characterized by TCH’s mean (TCHmean) and standard deviation (TCHsd), and TCH was subsequently used to estimate biomass at a 30-m resolution. Over a period of 25 years, TCHmean increased linearly with time; residual vegetation on landslides translated on average into larger TCHmean compared to landslides without residual vegetation. In contrast, in landslides without residual vegetation the linear relationship between TCHsd and time exhibited a steeper slope than that of landslides with residual vegetation. Above ground biomass increased linearly with time, and the rate of increase was larger at low elevations followed by mid and high elevations. Using orthoimages and digital surface models generated from data collected by instruments mounted on UAVs, two independent studies in Japan found that residual vegetation on landsliding greatly contributed to their recovery [185,186].
4.2. Soil and Lithology Attributes, Montane Ecosystems, and Landsliding
In mountainous environments, parent material strongly influences soil [187,188,189,190] and ecosystem [191,192,193] properties with likely consequences on slope stability, and ultimately landsliding (Figure 3). Yet, it is also likely that the exposure of fresh rock resulting from landslide activity sets in motion geochemical, biogeochemical, and biological processes that have been poorly linked but that may have profound consequences on the biosphere. Addressing these possibilities, however, is difficult given the poor characterization of parent materials in many mountainous regions around the world. In this regard, multi- and hyperspectral remotely sensed data has been invaluable for lithological mapping of hilly and mountainous regions [17,37,194,195]. This data has also been used, albeit to a lesser extent, to quantify rock mineralogy, geochemistry, and weathering degree [196,197,198,199,200,201] and characterize soil properties [202,203,204,205,206,207,208]. One common thread among most of this work is its focus on bare rock and soil or ecosystems with sparse vegetation such as grasslands or croplands. In densely vegetated mountains, landslide activity exposes bare substrates, and this may open opportunities to study rock and soil processes borrowing work conducted beyond these environments [209,210,211,212,213].
In hilly and mountainous humid environments with dense canopies two complementary approaches may help characterize underlying parent material and soil properties, landslides undergoing succession, and/or ecosystem-geomorphic interactions. Although both focus on the vegetation, their emphasis varies. A first approach uses remote sensing to establish relationships between leaf/canopy traits retrieved from multi- and hyperspectral data (See Section 3) and soil fertility. In planted and natural forests, a reduction in leaf area and foliar concentration of Chlorophyll (Chl), Nitrogen (N), and Phosphorus (P) is associated with nutrient deficiencies, that in the case of rock-derived nutrients may be inherited from parent material [Figure 3b; 193]. In the Archipelago of Hawaii, three studies that used airborne LiDAR and hyperspectral data to obtain tree heights, including derived estimates of above ground biomass, and foliar N and P have contributed different insights into leaf traits[191–19-ecosystem function relationships. Across a substrate age gradient - surrogate for degree of weathering and nutrient limitation - foliar N was slightly larger at the two intermediate-age sites, whereas mean tree height decreased, but its variability increased, with substrate age [214]. In Kaua’i, the oldest island of the Archipelago, remotely sensed foliar P resembled spatial predictions of foliar P based on analytical measurements, elevation, and rainfall [215]. More specifically, concentration of foliar P was highest in lower slopes and depositional areas and lowest on ridges and the stable shield surfaces. A negative correlation between foliar P and strontium isotopes indicated that increased fertility was the result of inputs from rock weathering. Finaly, in Hawai’i, the youngest of the islands, remotely sensed foliar N did not change with substrate characteristics but the same was not true for above ground biomass [64]. In Costa Rica, low concentrations of remotely sensed foliar N were observed on slopes whereas high concentrations on ridges [58]; several soil N metrics followed this pattern [216]. In the Andean foothills of eastern Peru, remotely sensed foliar concentrations of N and rock-derived nutrients (P, Calcium-Ca, Magnesium-Mg, and Potassium-K) increased non-linearly with incision depth; the opposite was true for LMA [176,217]. Furthermore, remotely sensed foliar Ca and K were significantly and positively correlated with the corresponding available nutrient in soil whereas foliar and soil P were negatively correlated. In Mt. Kinabalu, Malaysian Borneo, a study centered on two lithologies (sedimentary and ultramafic lithologies) and 32 catchments (16 per lithology) found that remotely sensed foliar P and N were significantly and positively correlated with a measure of hillslope disequilibrium (HD; decaying to steepening) in areas underlain by both lithologies; for remotely sensed foliar Ca this was true but only in areas underlain by sedimentary lithologies [57]. Finally, LMA was negatively correlated with HD. In addition to informing about underlying rock and soil properties, this work also suggests a role of erosional processes in supplying rock-derived nutrients, otherwise referred to as soil rejuvenation.
The second approach uses remotely sensed data to identify vegetation anomalies and/or indicator species associated with mineralization and geo-environments [218,219,220,221,222,223,224,225,226,227,228,229]. Deeply rooted in Phyto/Bio geochemistry and Geobotany [230,231,232,233], Geobotanical Remote Sensing (GBRS) investigates associations among plant traits, soil properties, underlying mineral deposits, and spectral signatures in areas densely covered by vegetation for the purpose of mineral exploration, geologic delineation, and phytoremediation. At the leaf level, plant traits include concentration of metal elements, water content, and spectra whereas at the crown, individual and population levels tree height and leaf phenology, and at the community level species composition and structure; soil properties include concentration of mineral elements (Figure 3b). The principle behind GBRS is that plants growing at sites with anomalous concentrations of metal elements have altered physiologies and that these stress states are observable in the chemical and spectral signatures of leaves, and remotely sensed plant canopies. At any given site targeted for mineral exploration, the within species responses to different mineral elements as well as the among species responses to a single element can be highly variable and this has the potential to identify indicator species. In the context of ecosystem-geomorphic interactions studies this work can potentially inform about changes in the redistribution of biomass between ecosystem above and belowground compartments and their influence on slope stability and landslide regeneration (Figure 3c). In addition, it can inform about plant-microbial associations that may help plants survive under stressful conditions.
5. Hyperspectral Remote Sensing Can Integrate Plant Traits, Soil-Rock Attributes, and Landslide Studies to Understand the Diversity, Functioning, and Dynamics of Mountainscapes
An increasing availability of airborne and spaceborne hyperspectral data, together with planned missions, can help develop a needed holistic view of landslides. This holistic view recognizes the critical role that interactions between ecosystem and geomorphic processes play in the dynamics of mountainscapes mediated by landslides and the contribution of ecosystem diversity and processes to landslide susceptibility and recovery. In this “ecosystem-centered” view of landslides it might be necessary to scale from individual landslides and sites to entire landslide populations, assemblages, and landscapes. This is because theoretical and observational data show that the distribution of landslide sizes is very diverse, and this can partially explain the variability in landslide recovery or developmental trajectories (Figure 3d).
Landslides create new substrates and landscape configurations, and the reorganization and recovery of ecosystems have implications beyond stabilizing sediment fluxes and sequestering carbon. Landslides alter abiotic conditions, including carbon and nutrient cycling, rock weathering, and soil formation while offering unique habitats that favor subsets of unique species. Hyperspectral and LiDAR data may help characterize the vegetation based on functional traits, the underlying soils and lithology based on physical and chemical attributes, and ultimately the functional diversity of landslides and landscapes. At this point we could interrogate the landscapes to answer questions about the contribution of landslides to the diversity, functioning, and disturbance dynamics of mountainscapes, but also about the contribution of ecosystem development to landslide hazard. For example, understanding changes during ecosystem development in above and belowground attributes influencing slope stability may help constrain sets of attributes influencing recovery, understand variation in successional trajectories, and landscape memory.
Ecosystems may influence slope resistance and susceptibility to failure in complex ways due to the dynamic nature of vegetation-soil-saprolite interactions. Hyperspectral and LiDAR data can potentially help identify ecosystem attributes that interact in tandem with geomorphic ones to understand and even forecast sites likely to fail. A focus on tree heights and biomass, including its allocation to above and below ground parts, at individual to plot/pixel scales could be a starting point. A combination of leaf- and canopy-level functional traits and biomass derived from hyperspectral and LiDAR data together with field work may help characterize and define functional groups based on attributes influencing slope stability. Furthermore, understanding this variability in space and time and relationships with the formation of landslides, may help test a priori hypotheses as well as postulate new ones regarding ecosystem-geomorphic interactions. This variability may be related to biotic (e.g., species) and abiotic (climate, soil fertility, geologic substrate, degree of weathering) conditions.
In sum, an “ecosystem-centered” view of landslides greatly aided by hyperspectral and LiDAR data can reveal unknown relationships between ecosystem and geomorphic processes from local to regional scales, while informing about the diversity and functioning of montane ecosystems, and ultimately the long-term functional significance of landslides in mountainscapes.
6. Conclusion
This review aimed at synthesizing broad established methods to retrieve functional traits from remotely sensed data, summarizing approaches derived from remotely sensed data to characterize functional traits and functional diversity, and elaborating on how these methods and approaches can provide a needed holistic view of landslides. We also posed that it was important to bring together developments in hyperspectral remote sensing stemming from research in different disciplines. It became clear through our work that the extent to which hyperspectral and LiDAR data has been used to investigate montane ecosystems, and more specifically tropical ones, has been limited. This includes the use of this data to characterize functional traits and functional diversity, and using these, to understand ecosystem-geomorphic interactions. It also became clear that integrating different disciplines using similar data and approaches can provide a fresh view of mountainscapes, including their dynamics mediated by landsliding.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. File S1 includes Table S1 summarizing landslide studies characterizing Vegetation Recovery Rates (VRR) over time and Figure S1 with spectral features representing by functional traits and principal components used in the RGB composites in Figure 2.
Author Contributions
Study conception and design (AK and CR) and manuscript preparation (AK and CR).
Funding
Support for this work was provided by a NASA Puerto Rico Space Grant Fellowship (AK), NSF DEB 1556878 (CR) and REPS Supplemental Funding (CR, AK), and the Puerto Rico Science Trust (CR).
Data Availability Statement
Not applicable
Acknowledgements
We are thankful to Y. Ortiz and L. Ospina for contributing to lively discussions on functional traits, functional diversity, and remote sensing, to F. Schneider for clarifying questions regarding spectroscopy, and to B.E. Hubbard for providing a friendly review of an early version of this manuscript.
Conflicts of Interest
None
References
- Chapin, F.S., 3rd; Carpenter, S.R.; Kofinas, G.P.; Folke, C.; Abel, N.; Clark, W.C.; Olsson, P.; Smith, D.M.; Walker, B.; Young, O.R. , et al. Ecosystem stewardship: Sustainability strategies for a rapidly changing planet. Trends Ecol. Evol. 2010, 25, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.C. The Post-2020 Global Biodiversity Framework: How did we get here, and where do we go next? Integr. Conserv. 2023, 2, 1–9. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Schneider, F.D.; Santos, M.J.; Armstrong, A.; Carnaval, A.; Dahlin, K.M.; Fatoyinbo, L.; Hurtt, G.C.; Schimel, D.; Townsend, P.A. , et al. Integrating remote sensing with ecology and evolution to advance biodiversity conservation. Nat. Ecol. Evol. 2022, 6, 506–519. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Functional diversity: back to basics and looking forward. Ecol. Lett. 2006, 9, 741–758. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Gutiérrez, J.; Rifai, S.; Shenkin, A.; Oliveras, I.; Bentley, L.P.; Svátek, M.; Girardin, C.A.J.; Both, S.; Riutta, T.; Berenguer, E. , et al. Pantropical modelling of canopy functional traits using Sentinel-2 remote sensing data. Remote Sens. Environ. 2021, 252, 112122. [Google Scholar] [CrossRef]
- Maxwell, A.E.; Wilson, B.T.; Holgerson, J.J.; Bester, M.S. Comparing harmonic regression and GLAD Phenology metrics for estimation of forest community types and aboveground live biomass within forest inventory and analysis plots. Int. J. Appl. Earth Obs. 2023, 122, 103435. [Google Scholar] [CrossRef]
- Neinavaz, E.; Schlerf, M.; Darvishzadeh, R.; Gerhards, M.; Skidmore, A.K. Thermal infrared remote sensing of vegetation: Current status and perspectives. Int. J. Appl. Earth Obs. 2021, 102, 102415. [Google Scholar] [CrossRef]
- Skidmore, A.K.; Pettorelli, N. Agree on biodiversity metrics to track from space. Nature 2015, 523, 504–512. [Google Scholar] [CrossRef]
- Wallis, C.I.B.; Homeier, J.; Peña, J.; Brandl, R.; Farwig, N.; Bendix, J. Modeling tropical montane forest biomass, productivity and canopy traits with multispectral remote sensing data. Remote Sens. Environ. 2019, 225, 77–92. [Google Scholar] [CrossRef]
- Xiong, X.; Butler, J.J. MODIS and VIIRS calibration history and future outlook. Remote Sens. 2020, 12, 2523. [Google Scholar] [CrossRef]
- Helmer, E.H.; Goodwin, N.R.; Gond, V.; Souza, Jr.; C.M., *!!! REPLACE !!!*; Asner, G.P. Characterizing tropical forests with multispectral imagery. In Land Resources: Monitoring, Modeling and Mapping; Thenkabail, P.S., Ed.; Taylor & Francis Group: Boca Raton, Florida, USA, 2015; pp. 367–396. [Google Scholar]
- Asner, G.P.; Martin, R.E.; Knapp, D.E.; Tupayachi, R.; Anderson, C.B.; Sinca, F.; Vaughn, N.R.; Llactayo, W. Airborne laser-guided imaging spectroscopy to map forest trait diversity and guide conservation. Science 2017, 355, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Kokaly, R.F.; Asner, G.P.; Ollinger, S.V.; Martin, M.E.; Wessman, C.A. Characterizing canopy biochemistry from imaging spectroscopy and its application to ecosystem studies. Remote Sens. Environ. 2009, 113, S78–S91. [Google Scholar] [CrossRef]
- Pascual-Venteo, A.B.; Portalés, E.; Berger, K.; Tagliabue, G.; Garcia, J.L.; Pérez-Suay, A.; Rivera-Caicedo, J.P.; Verrelst, J. Prototyping crop traits retrieval models for CHIME: Dimensionality reduction strategies applied to PRISMA data. Remote Sens. 2022, 14, 2448. [Google Scholar] [CrossRef] [PubMed]
- Schneider, F.D.; Morsdorf, F.; Schmid, B.; Petchey, O.L.; Hueni, A.; Schimel, D.S.; Schaepman, M.E. Mapping functional diversity from remotely sensed morphological and physiological forest traits. Nat. Commun. 2017, 8, 1441. [Google Scholar] [CrossRef] [PubMed]
- Ustin, S.L.; Roberts, D.A.; Gamon, J.A.; Asner, G.P.; Green, R.O. Using imaging spectroscopy to study ecosystem processes and properties. BioScience 2004, 54, 523–534. [Google Scholar] [CrossRef]
- Abrams, M.; Yamaguchi, Y. Twenty Years of ASTER Contributions to Lithologic Mapping and Mineral Exploration. Remote Sens. 2019, 11, 1394. [Google Scholar] [CrossRef]
- Misbah, K.; Laamrani, A.; Khechba, K.; Dhiba, D.; Chehbouni, A. Multi-sensors remote sensing applications for assessing, monitoring, and mapping NPK content in soil and crops in African agricultural land. Remote Sens. 2022, 14, 81. [Google Scholar] [CrossRef]
- Novais, J.J.; Poppiel, R.R.; Lacerda, M.P.C.; Oliveira, M.P.; Demattê, J.A.M. Spectral Mixture Modeling of an ASTER Bare Soil Synthetic Image Using a Representative Spectral Library to Map Soils in Central-Brazil. AgriEngineering 2023, 5, 156–172. [Google Scholar] [CrossRef]
- Helfenstein, I.S.; Schneider, F.D.; Schaepman, M.E.; Morsdorf, F. Assessing biodiversity from space: Impact of spatial and spectral resolution on trait-based functional diversity. Remote Sens. Environ. 2022, 275, 113024. [Google Scholar] [CrossRef]
- Lechner, A.M.; Foody, G.M.; Boyd, D.S. Applications in remote sensing to forest ecology and management. One Earth 2020, 2, 405–412. [Google Scholar] [CrossRef]
- Pettorelli, N.; Schulte to Bühne, H.; Tulloch, A.; Dubois, G.; Macinnis-Ng, C.; Queirós, A.M.; Keith, D.A.; Wegmann, M.; Schrodt, F.; Stellmes, M. , et al. Satellite remote sensing of ecosystem functions: opportunities, challenges and way forward. Remote Sens. Ecol. Conserv. 2018, 4, 71–93. [Google Scholar] [CrossRef]
- Ma, X.; Mahecha, M.D.; Migliavacca, M.; van der Plas, F.; Benavides, R.; Ratcliffe, S.; Kattge, J.; Richter, R.; Musavi, T.; Baeten, L. , et al. Inferring plant functional diversity from space: the potential of Sentinel-2. Remote Sens. Environ. 2019, 233, 111368. [Google Scholar] [CrossRef]
- Zheng, Z.; Schmid, B.; Zeng, Y.; Schuman, M.C.; Zhao, D.; Schaepman, M.E.; Morsdorf, F. Remotely sensed functional diversity and its association with productivity in a subtropical forest. Remote Sens. Environ. 2023, 290, 113530. [Google Scholar]
- Fernández-García, V.; Marcos, E.; Fernández-Guisuraga, J.M.; Fernández-Manso, A.; Quintano, C.; Suárez-Seoane, S.; Calvo, L. Multiple endmember spectral mixture analysis (MESMA) applied to the study of habitat diversity in the fine-grained landscapes of the Cantabrian Mountains. Remote Sens. 2021, 13. [Google Scholar] [CrossRef]
- Asner, G.P.; Anderson, C.B.; Martin, R.E.; Tupayachi, R.; Knapp, D.E.; Sinca, F. Landscape biogeochemistry reflected in shifting distributions of chemical traits in the Amazon forest canopy. Nat. Geosci. 2015, 8, 567–573. [Google Scholar] [CrossRef]
- Asner, G.P.; Knapp, D.E.; Anderson, C.B.; Martin, R.E.; Vaughn, N. Large-scale climatic and geophysical controls on the leaf economics spectrum. Proc. Natl. Acad. Sci. U S A 2016, 113, E4043–E4051. [Google Scholar] [CrossRef] [PubMed]
- Zini, S.; Barbato, M.P.; Piccoli, F.; Napoletano, P. Deep learning hyperspectral pansharpening on large scale PRISMA dataset. Remote Sens. 2024, 16, 2079. [Google Scholar] [CrossRef]
- Bogan, S.A.; Antonarakis, A.S.; Moorcroft, P.R. Imaging spectrometry-derived estimates of regional ecosystem composition for the Sierra Nevada, California. Remote Sens. Environ. 2019, 228, 14–30. [Google Scholar] [CrossRef]
- Hacker, P.W.; Coops, N.C.; Laliberté, E.; Michaletz, S.T. Variations in accuracy of leaf functional trait prediction due to spectral mixing. Ecol. Indic. 2022, 136, 108687. [Google Scholar]
- Quintano, C.; Fernández-Manso, A.; Fernández-Guisuraga, J.M.; Calvo, L. Unmixing PRISMA hyperspectral images by multiple endmember spectral mixture analysis (MESMA) to assess fire severity in Mediterranean forest ecosystems. Imaging Spectrometry XXVI: Applications; Sensors, and Processing, 2023; p. 126880I. [Google Scholar]
- Waser, L.T.; Rüetschi, M.; Psomas, A.; Small, D.; Rehush, N. Mapping dominant leaf type based on combined Sentinel-1/-2 data – Challenges for mountainous countries. ISPRS J. Photogramm. Rem. Sens. 2021, 180, 209–226. [Google Scholar]
- Weiss, D.J.; Walsh, S.J. Remote Sensing of Mountain Environments. Geogr. Comp. 2009, 3, 1–21. [Google Scholar] [CrossRef]
- Brown, J.H.; Gupta, V.K.; Li, B.-L.; Milne, B.T.; Restrepo, C.; West, G.B. The fractal nature of nature: power laws, ecological complexity and biodiversity. Philosophical Transactions of the Royal Society of London Series B Biological Sciences 2002, 357, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Medwedeff, W.G.; Clark, M.K.; Zekkos, D.; West, A.J. Characteristic landslide distributions: An investigation of landscape controls on landslide size. Earth Planet. Sc. Lett. 2020, 539, 116203. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Anderson, C.B.; Knapp, D.E. Quantifying forest canopy traits: Imaging spectroscopy versus field survey. Remote Sens. Environ. 2015, 158, 15–27. [Google Scholar]
- Bertoldi, L.; Massironi, M.; Visonà, D.; Carosi, R.; Montomoli, C.; Gubert, F.; Naletto, G.; Pelizzo, M.G. Mapping the Buraburi granite in the Himalaya of Western Nepal: Remote sensing analysis in a collisional belt with vegetation cover and extreme variation of topography. Remote Sens. Environ. 2011, 115, 1129–1144. [Google Scholar] [CrossRef]
- Loizzo, R.; Ananasso, C.; Guarini, R.; Lopinto, E.; Candela, L.; Pisani, A.R. The PRISMA Hyperspectral Mission. In Living Planet Symposium 2016, 9-13 May 2016, Prague, Czech Republic; 13 May 2016. [Google Scholar]
- Storch, T.; Honold, H.-P.; Chabrillat, S.; Habermeyer, M.; Tucker, P.; Brell, M.; Ohndorf, A.; Wirth, K.; Betz, M.; Kuchler, M. , et al. The EnMAP imaging spectroscopy mission towards operations. Remote Sens. Environ. 2023, 294, 113632. [Google Scholar]
- Schneider, F.D.; Ferraz, A.; Schimel, D. Watching Earth’s interconnected systems at work. Eos 2019, 100. [Google Scholar] [CrossRef]
- Stavros, E.N.; Chrone, J.; Cawse-Nicholson, K.; Freeman, A.; Glenn, N.F.; Guild, L.; Kokaly, R.; Lee, C.; Luvall, J.; Pavlick, R. , et al. Designing an Observing System to Study the Surface Biology and Geology (SBG) of the Earth in the 2020s. Journal of Geophysical Research: Biogeosciences 2023, 128, e2021JG006471. [Google Scholar]
- Nieke, J.; Rast, M. Status: Copernicus Hyperspectral Imaging Mission for the Environment (CHIME). IGARSS 2019, 4609–4611. [Google Scholar]
- Planets Lab PBC. Tanager-1 Is Ready For Launch: Planet’s First Hyperspectral Satellite. Available online: https://www.planet.com/pulse/tanager-1-is-ready-for-launch-planets-first-hyperspectral-satellite/.
- Asner, G.P.; Martin, R.E.; Knapp, D.E.; Tupayachi, R.; Anderson, C.; Carranza, L.; Martinez, P.; Houcheime, M.; Sinca, F.; Weiss, P. Spectroscopy of canopy chemicals in humid tropical forests. Remote Sens. Environ. 2011, 115, 3587–3598. [Google Scholar] [CrossRef]
- Reich, P.B.; Wright, I.J.; Cavender-Bares, J.; Craine, J.M.; Oleksyn, J.; Westoby, M.; Walters, M.B. The evolution of plant functional variation: Traits, spectra, and strategies. Int. J. Plant Sci. 2003, 164, S143–S164. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive la différence: plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Diaz, S.; Purvis, A.; Cornelissen, J.H.; Mace, G.M.; Donoghue, M.J.; Ewers, R.M.; Jordano, P.; Pearse, W.D. Functional traits, the phylogeny of function, and ecosystem service vulnerability. Ecol. Evol. 2013, 3, 2958–2975. [Google Scholar] [CrossRef]
- Lavorel, S.; Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Naeem, S.; Wright, J.P. Disentangling biodiversity effects on ecosystem functioning: deriving solutions to a seemingly insurmountable problem. Ecol. Lett. 2003, 6, 567–579. [Google Scholar]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Bruelheide, H.; Dengler, J.; Purschke, O.; Lenoir, J.; Jiménez-Alfaro, B.; Hennekens, S.M.; Botta-Dukát, Z.; Chytrý, M.; Field, R.; Jansen, F. , et al. Global trait–environment relationships of plant communities. Nat. Ecol. Evol. 2018, 2, 1906–1917. [Google Scholar] [CrossRef] [PubMed]
- Šímová, I.; Violle, C.; Svenning, J.-C.; Kattge, J.; Engemann, K.; Sandel, B.; Peet, R.K.; Wiser, S.K.; Blonder, B.; McGill, B.J. , et al. Spatial patterns and climate relationships of major plant traits in the New World differ between woody and herbaceous species. J. Biogeogr. 2018, 45, 895–916. [Google Scholar] [CrossRef]
- Laliberte, E.; Schweiger, A.K.; Legendre, P. Partitioning plant spectral diversity into alpha and beta components. Ecol Lett 2020, 23, 370–380. [Google Scholar]
- Rossi, C.; Kneubühler, M.; Schütz, M.; Schaepman, M.E.; Haller, R.M.; Risch, A.C. Remote sensing of spectral diversity: A new methodological approach to account for spatio-temporal dissimilarities between plant communities. Ecol. Indic. 2021, 130, 108106. [Google Scholar] [CrossRef]
- Curran, P.J. Remote sensing of foliar chemistry. Remote Sens. Environ. 1989, 30, 271–278. [Google Scholar] [CrossRef]
- Gholizadeh, H.; Friedman, M.S.; McMillan, N.A.; Hammond, W.M.; Hassani, K.; Sams, A.V.; Charles, M.D.; Garrett, D.R.; Joshi, O.; Hamilton, R.G. , et al. Mapping invasive alien species in grassland ecosystems using airborne imaging spectroscopy and remotely observable vegetation functional traits. Remote Sens. Environ. 2022, 271, 112887. [Google Scholar] [CrossRef]
- Chadwick, K.D.; Asner, G.P. Geomorphic transience moderates topographic controls on tropical canopy foliar traits. Ecol. Lett. 2020, 23, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Balzotti, C.S.; Asner, G.P.; Taylor, P.G.; Cleveland, C.C.; Cole, R.; Martin, R.E.; Nasto, M.; Osborne, B.B.; Porder, S.; Townsend, A.R. Environmental controls on canopy foliar nitrogen distributions in a Neotropical lowland forest. Ecol. Appl. 2016, 26, 2449–2462. [Google Scholar] [PubMed]
- Osborne, B.B.; Nasto, M.K.; Asner, G.P.; Balzotti, C.S.; Cleveland, C.C.; Sullivan, B.W.; Taylor, P.G.; Townsend, A.R.; Porder, S. Climate, topography, and canopy chemistry exert hierarchical control over soil N cycling in a Neotropical lowland forest. Ecosystems 2017, 20, 1089–1103. [Google Scholar]
- Miraglio, T.; Coops, N.C.; Wallis, C.I.B.; Crofts, A.L.; Kalacska, M.; Vellend, M.; Serbin, S.P.; Arroyo-Mora, J.P.; Laliberté, E. Mapping canopy traits over Québec using airborne and spaceborne imaging spectroscopy. Sci. Rep. 2023, 13, 17179. [Google Scholar]
- Asner, G.P.; Vitousek, P.M. Remote analysis of biological invasion and biogeochemical change. Proc. Natl. Acad. Sci. U S A 2005, 102, 4383–4386. [Google Scholar]
- Hall, S.J.; Asner, G.P. Biological invasion alters regional nitrogen-oxide emissions from tropical rainforests. Glob. Change Biol. 2007, 13, 2143–2160. [Google Scholar] [CrossRef]
- Asner, G.P.; Nepstad, D.; Cardinot, G.; Ray, D. Drought stress and carbon uptake in an Amazon forest measured with spaceborne imaging spectroscopy. Proc. Natl. Acad. Sci. U S A 2004, 101, 6039–6044. [Google Scholar] [CrossRef]
- Balzotti, C.S.; Asner, G.P. Biotic and abiotic controls over canopy function and structure in humid Hawaiian forests. Ecosystems 2018, 21, 331–348. [Google Scholar]
- Asner, G.P. Hyperspectral remote sensing of canopy chemistry, physiology and diversity in tropical rainforests - Chapter 12. In Hyperspectral Remote Sensing of tTopical and Subtropical Forests; Kalacska, M., Sanchez-Azofeifa, G.A., Eds.; Taylor and Francis Group: Boca Raton, Florida, US, 2008; p. 352. [Google Scholar]
- Gamon, J.A.; Rahman, A.F.; Dungan, J.L.; Schildhauer, M.; Huemmrich, K.F. Spectral Network (SpecNet)—What is it and why do we need it? Remote Sens. Environ. 2006, 103, 227–235. [Google Scholar] [CrossRef]
- Kattenborn, T. Linking canopy reflectance and plant functioning through radiative transfer models. Dr. rer. nat. Thesis, Karlsruher Institut für Technologie, Karlsruhe, Germany, 2018. [Google Scholar]
- Wang, Z.; Chlus, A.; Geygan, R.; Ye, Z.; Zheng, T.; Singh, A.; Couture, J.J.; Cavender-Bares, J.; Kruger, E.L.; Townsend, P.A. Foliar functional traits from imaging spectroscopy across biomes in eastern North America. New Phytol. 2020, 228, 494–511. [Google Scholar] [CrossRef] [PubMed]
- Asner, G.P.; Martin, R.E.; Ford, A.J.; Metcalfe, D.J.; Liddell, M.J. Leaf chemical and spectral diversity in Australian tropical forests. Ecol. Appl. 2009, 19, 236–253. [Google Scholar] [CrossRef]
- Chadwick, K.; Asner, G. Organismic-scale remote sensing of canopy foliar traits in lowland tropical forests. Remote Sens. 2016, 8, 87. [Google Scholar] [CrossRef]
- Matheny, A.M.; Mirfenderesgi, G.; Bohrer, G. Trait-based representation of hydrological functional properties of plants in weather and ecosystem models. Plant Divers. 2017, 39, 1–12. [Google Scholar] [CrossRef]
- Asner, G.P.; Haxo, B.; Knapp, D.E. Computing for analysis and modeling of hyperspectral imagery. In High Performance Computing for Remote Sensing; Plaza, S., Chang, C.I., Eds.; Chapman and Hall Press: New York, New York, US, 2007; pp. 110–130. [Google Scholar]
- Doughty, C.E.; Asner, G.P.; Martin, R.E. Predicting tropical plant physiology from leaf and canopy spectroscopy. Oecologia 2011, 165, 289–299. [Google Scholar]
- Féret, J.-B.; François, C.; Gitelson, A.; Asner, G.P.; Barry, K.M.; Panigada, C.; Richardson, A.D.; Jacquemoud, S. Optimizing spectral indices and chemometric analysis of leaf chemical properties using radiative transfer modeling. Remote Sens. Environ. 2011, 115, 2742–2750. [Google Scholar] [CrossRef]
- Serbin, S.P.; Townsend, P.A. Scaling functional traits from leaves to canopies. Chapter 3. In Remote Sensing of Plant Biodiversity; Cavender-Bares, J., Gamon, J.A., Townsend, P.A., Eds.; Springer Open, 2020; pp. 43–82. [Google Scholar]
- Verrelst, J.; Camps-Valls, G.; Muñoz-Marí, J.; Rivera, J.P.; Veroustraete, F.; Clevers, J.G.P.W.; Moreno, J. Optical remote sensing and the retrieval of terrestrial vegetation bio-geophysical properties – A review. ISPRS J. Photogramm. Rem. Sens. 2015, 108, 273–290. [Google Scholar] [CrossRef]
- Bannari, A.; Morin, D.; Bonn, F.; Huete, A.R. A review of vegetation indices. Remote Sens. Rev. 1995, 13, 95–120. [Google Scholar] [CrossRef]
- Beauchemin, M.; Fung, K.B. On statistical band selection for image visualization. Phogrammetric Engineering and Remote Sensing 2001, 67, 571–574. [Google Scholar]
- Ollinger, S.V. Sources of variability in canopy reflectance and the convergent properties of plants. New Phytol. 2011, 189, 375–394. [Google Scholar] [CrossRef] [PubMed]
- Higgins, M.A.; Asner, G.P.; Martin, R.E.; Knapp, D.E.; Anderson, C.; Kennedy-Bowdoin, T.; Saenz, R.; Aguilar, A.; Joseph Wright, S. Linking imaging spectroscopy and LiDAR with floristic composition and forest structure in Panama. Remote Sens. Environ. 2014, 154, 358–367. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Carranza-Jimenez, L.; Sinca, F.; Tupayachi, R.; Anderson, C.B.; Martinez, P. Functional and biological diversity of foliar spectra in tree canopies throughout the Andes to Amazon region. New Phytol. 2014, 204, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Burnett, A.C.; Anderson, J.; Davidson, K.J.; Ely, K.S.; Lamour, J.; Li, Q.; Morrison, B.D.; Yang, D.; Rogers, A.; Serbin, S.P. A best-practice guide to predicting plant traits from leaf-level hyperspectral data using partial least squares regression. J. Exp. Biol. 2021, 72, 6175–6189. [Google Scholar] [CrossRef]
- Feilhauer, H.; Asner, G.P.; Martin, R.E.; Schmidtlein, S. Brightness-normalized Partial Least Squares Regression for hyperspectral data. J. Quant. Spectrosc. Ra. 2010, 111, 1947–1957. [Google Scholar] [CrossRef]
- Féret, J.-B.; François, C.; Asner, G.P.; Gitelson, A.A.; Martin, R.E.; Bidel, L.P.R.; Ustin, S.L.; le Maire, G.; Jacquemoud, S. PROSPECT-4 and 5: Advances in the leaf optical properties model separating photosynthetic pigments. Remote Sens. Environ. 2008, 112, 3030–3043. [Google Scholar] [CrossRef]
- Kötz, B.; Schaepman, M.; Morsdorf, F.; Bowyer, P.; Itten, K.; Allgöwer, B. Radiative transfer modeling within a heterogeneous canopy for estimation of forest fire fuel properties. Remote Sens. Environ. 2004, 92, 332–344. [Google Scholar] [CrossRef]
- Svendsen, D.H.; Hernández-Lobato, D.; Martino, L.; Laparra, V.; Moreno-Martínez, Á.; Camps-Valls, G. Inference over radiative transfer models using variational and expectation maximization methods. Machine Learning 2023, 112, 921–937. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Noland, T.L.; Mohammed, G.H.; Sampson, P.H. Scaling-up and model inversion methods with narrowband optical indices for chlorophyll content estimation in closed forest canopies with hyperspectral data. IEEE Trans. Geosci. Remote Sens. 2001, 39, 1491–1507. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E. Spectral and chemical analysis of tropical forests: Scaling from leaf to canopy levels. Remote Sens. Environ. 2008, 112, 3958–3970. [Google Scholar] [CrossRef]
- Pacheco-Labrador, J.; de Bello, F.; Migliavacca, M.; Ma, X.; Carvalhais, N.; Wirth, C. A generalizable normalization for assessing plant functional diversity metrics across scales from remote sensing. Methods Ecol. Evol. 2023, 14, 2123–2136. [Google Scholar] [CrossRef]
- Verrelst, J.; Rivera-Caicedo, J.P.; Reyes-Muñoz, P.; Morata, M.; Amin, E.; Tagliabue, G.; Panigada, C.; Hank, T.; Berger, K. Mapping landscape canopy nitrogen content from space using PRISMA data. ISPRS J. Photogramm. Rem. Sens. 2021, 178, 382–395. [Google Scholar]
- Asner, G.P.; Knapp, D.E.; Boardman, J.; Green, R.O.; Kennedy-Bowdoin, T.; Eastwood, M.; Martin, R.E.; Anderson, C.; Field, C.B. Carnegie Airborne Observatory-2: Increasing science data dimensionality via high-fidelity multi-sensor fusion. Remote Sens. Environ. 2012, 124, 454–465. [Google Scholar] [CrossRef]
- Lee, C.M.; Cable, M.L.; Hook, S.J.; Green, R.O.; Ustin, S.L.; Mandl, D.J.; Middleton, E.M. An introduction to the NASA Hyperspectral InfraRed Imager (HyspIRI) mission and preparatory activities. Remote Sens. Environ. 2015, 167, 6–19. [Google Scholar]
- McCorkel, J.; Kuester, M.; Johnson, B.R.; Kampe, T.U. NEON ground validation capabilities for airborne and space-based imagers. Proceedings SPIE, 2011; p. 81530Z. [Google Scholar]
- Dechant, B.; Kattge, J.; Pavlick, R.; Schneider, F.D.; Sabatini, F.; Moreno, A.; Butler, E.; Bodegom, P.; Vallicrosa, H.; Kattenborn, T. , et al. Intercomparison of global foliar trait maps reveals fundamental differences and limitations of upscaling approaches. Remote Sens. Environ. 2024, 311, 114276. [Google Scholar] [CrossRef]
- Wocher, M. Unlocking the benefits of spaceborne imaging spectroscopy for sustainabile agriculture. Dr. rer. nat. Thesis, Universität at München, München, 2022. [Google Scholar]
- Baret, F.; Guyot, G. Potentials and limits of vegetation indices for LAI and APAR assessment. Remote Sens. Environ. 1991, 35, 161–173. [Google Scholar] [CrossRef]
- Martin, R.E.; Chadwick, K.D.; Brodrick, P.G.; Carranza-Jimenez, L.; Vaughn, N.R.; Asner, G.P. An approach for foliar trait retrieval from airborne imaging spectroscopy of tropical forests. Remote Sens. 2018, 10, 199. [Google Scholar] [CrossRef]
- Zheng, Z.; Zeng, Y.; Schneider, F.D.; Zhao, Y.; Zhao, D.; Schmid, B.; Schaepman, M.E.; Morsdorf, F. Mapping functional diversity using individual tree-based morphological and physiological traits in a subtropical forest. Remote Sens. Environ. 2021, 252, 112170. [Google Scholar]
- Knox, N.M.; Skidmore, A.K.; Prins, H.H.T.; Asner, G.P.; van der Werff, H.M.A.; de Boer, W.F.; van der Waal, C.; de Knegt, H.J.; Kohi, E.M.; Slotow, R. , et al. Dry season mapping of savanna forage quality, using the hyperspectral Carnegie Airborne Observatory sensor. Remote Sens. Environ. 2011, 115, 1478–1488. [Google Scholar] [CrossRef]
- Díaz, S.; Lavorel, S.; Chapin, F.S.; Tecco, P.A.; Gurvich, D.E.; Grigulis, K. Functional Diversity — at the Crossroads between Ecosystem Functioning and Environmental Filters. In Terrestrial Ecosystems in a Changing World; Canadell, J.G., Pataki, D.E., Pitelka, L.F., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2007; pp. 91–91. [Google Scholar]
- Tilman, D. Functional Diversity. In Encyclopedia of Biodiversity; 2001; Volume 3, pp. 109–120. [Google Scholar]
- Ali, A. Biodiversity–ecosystem functioning research: Brief history, major trends and perspectives. Biol. Conserv. 2023, 285, 110210. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin Iii, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S. , et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and Ecosystem Functioning. Annu. Rev. Ecol. Syst 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Biswas, S.R.; Mallik, A.U. Disturbance effects on species diversity and functional diversity in riparian and upland plant communities. Ecology 2010, 91, 28–35. [Google Scholar] [CrossRef]
- Cursach, J.; Rita, J.; Gómez-Martínez, C.; Cardona, C.; Capó, M.; Lázaro, A. The role of landscape composition and heterogeneity on the taxonomical and functional diversity of Mediterranean plant communities in agricultural landscapes. PLOS ONE 2020, 15, e0238222. [Google Scholar] [CrossRef]
- de Bello, F.; Lavorel, S.; Hallett, L.M.; Valencia, E.; Garnier, E.; Roscher, C.; Conti, L.; Galland, T.; Goberna, M.; Májeková, M. , et al. Functional trait effects on ecosystem stability: assembling the jigsaw puzzle. Trends Ecol. Evol. 2021, 36, 822–836. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M.; Barbier, M.; Filotas, E.; Gravel, D.; Isbell, F.; Miller, S.J.; Montoya, J.M.; Wang, S.; Aussenac, R.; Germain, R. , et al. Biodiversity as insurance: From concept to measurement and application. Biol. Rev. 2021, 96, 2333–2354. [Google Scholar] [CrossRef]
- McWilliam, M.; Hoogenboom, M.O.; Baird, A.H.; Hughes, T.P. Biogeographical disparity in the functional diversity and redundancy of corals. Proc. Natl. Acad. Sci. U S A 2018, 115, 3084–3089. [Google Scholar] [CrossRef]
- Peterson, G.; Allen, C.R.; Holling, C.S. Ecological Resilience, Biodiversity, and Scale. Ecosystems 1998, 1, 6–18. [Google Scholar] [CrossRef]
- Restrepo, C.; Renjifo, L.M.; Marples, P. Frugivorous birds in fragmented neotropical montane forests: Landscape pattern and body mass distribution. In Tropical Forest Remnants: Ecology, Management and Conservation of Fragmented Ecosystems; Laurance, W.F., Bierregaard, R.O., Eds.; University of Chicago Press: Chicago, 1997; pp. 171–189. [Google Scholar]
- Castro Sánchez-Bermejo, P.; deCastro-Arrazola, I.; Cuesta, E.; Davis, A.L.V.; Moreno, C.E.; Sánchez-Piñero, F.; Hortal, J. Aridity drives the loss of dung beetle taxonomic and functional diversity in three contrasting deserts. J. Biogeogr. 2022, 49, 2243–2255. [Google Scholar] [CrossRef]
- Frainer, A.; Primicerio, R.; Dolgov, A.; Fossheim, M.; Johannesen, E.; Lind, S.; Aschan, M. Increased functional diversity warns of ecological transition in the Arctic. Proceedings of the Royal Society B Biological Sciences 2021, 288, 202110054. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.G.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing functional diversity in the field – methodology matters! Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- Spasojevic, M.J.; Suding, K.N. Inferring community assembly mechanisms from functional diversity patterns: the importance of multiple assembly processes. J. Ecol. 2012, 100, 652–661. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y.; Liu, S.; Chai, Y.; Dang, H.; Yue, M.; Liu, X.; Guo, Y. Patterns of diversity and community assembly change across local to regional scales: An evidence of deterministic assembly processes along resource availability gradient at temperate forest. Ecol. Indic. 2021, 132, 108261. [Google Scholar] [CrossRef]
- Carmona, C.P.; de Bello, F.; Mason, N.W.H.; Lepš, J. Traits without borders: Integrating functional diversity across scales. Trends Ecol. Evol. 2016, 31, 382–394. [Google Scholar] [CrossRef] [PubMed]
- de Bello, F.; Thuiller, W.; Lepš, J.; Choler, P.; Clément, J.-C.; Macek, P.; Sebastià, M.-T.; Lavorel, S. Partitioning of functional diversity reveals the scale and extent of trait convergence and divergence. J. Veg. Sci. 2009, 20, 475–486. [Google Scholar] [CrossRef]
- Villéger, S.; Grenouillet, G.; Brosse, S. Decomposing functional β-diversity reveals that low functional β-diversity is driven by low functional turnover in European fish assemblages. Global Ecol. Biogeogr. 2013, 22, 671–681. [Google Scholar] [CrossRef]
- Petchey, O.L.; O’Gorman, E.J.; Flynn, D.F.B. A functional guide to functional diversity measures - Chapter 4. In Biodiversity, Ecosystem Functioning, and Human Wellbeing: An Ecological and Economic Perspective; Naeem, S., Bunker, D.E., Hector, A., Loreau, M., Perrings, C., Eds.; Oxford University Press: UK, 2009; pp. 49–59. [Google Scholar]
- Cardinale, B.J.; Srivastava, D.S.; Duffy, J.E.; Wright, J.P.; Downing, A.L.; Sankaran, M.; Jouseau, C. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 2006, 443, 989–992. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and Its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Holling, C.S. Cross-Scale Morphology, Geometry, and Dynamics of Ecosystems. Ecol. Monogr. 1992, 62, 447–502. [Google Scholar]
- MacArthur, R.; Wilson, E.O. The Theory of Island Biogeography. Princeton University Press: Princeton, New Jersey, US, 1967; p. 203. [Google Scholar]
- Reich, P.B. The world-wide ‘fast–slow’ plant economics spectrum: a traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. Leaf Life-Span in Relation to Leaf, Plant, and Stand Characteristics among Diverse Ecosystems. Ecol. Monogr. 1992, 62, 365–392. [Google Scholar] [CrossRef]
- Ustin, S.L.; Gamon, J.A. Remote sensing of plant functional types. New Phytol. 2010, 186, 795–816. [Google Scholar] [CrossRef]
- Mason, N., W. H. ; Mouillot, D.; Lee, W.G.; Wilson, J.B.; Setälä, H. Functional Richness, Functional Evenness and Functional Divergence: The Primary Components of Functional Diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.H.; Mouillot, D. Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Schleuter, D.; Daufresne, M.; Massol, F.; Argillier, C. A user's guide to functional diversity indices. Ecol. Monogr. 2010, 80, 469–484. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a mutlifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef]
- Cimoli, E.; Lucieer, A.; Malenovský, Z.; Woodgate, W.; Janoutová, R.; Turner, D.; Haynes, R.S.; Phinn, S. Mapping functional diversity of canopy physiological traits using UAS imaging spectroscopy. Remote Sens. Environ. 2024, 302, 113958. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E. Spectranomics: Emerging science and conservation opportunities at the interface of biodiversity and remote sensing. GECCO 2016, 8, 212–219. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Gamon, J.A.; Hobbie, S.E.; Madritch, M.D.; Meireles, J.E.; Schweiger, A.K.; Townsend, P.A. Harnessing plant spectra to integrate the biodiversity sciences across biological and spatial scales. Am. J. Bot. 2017, 104, 966–969. [Google Scholar] [CrossRef]
- Schweiger, A.K.; Cavender-Bares, J.; Townsend, P.A.; Hobbie, S.E.; Madritch, M.D.; Wang, R.; Tilman, D.; Gamon, J.A. Plant spectral diversity integrates functional and phylogenetic components of biodiversity and predicts ecosystem function. Nat. Ecol. Evol. 2018, 2, 976–982. [Google Scholar] [CrossRef]
- Wang, R.; Gamon, J.A. Remote sensing of terrestrial plant biodiversity. Remote Sens. Environ. 2019, 231, 111218. [Google Scholar] [CrossRef]
- Rocchini, D.; Santos, M.J.; Ustin, S.L.; Feret, J.B.; Asner, G.P.; Beierkuhnlein, C.; Dalponte, M.; Feilhauer, H.; Foody, G.M.; Geller, G.N. , et al. The spectral species concept in living color. J Geophys Res Biogeosci 2022, 127, e2022JG007026. [Google Scholar] [CrossRef] [PubMed]
- Schneider, F.D.; Ferraz, A.; Hancock, S.; Duncanson, L.I.; Dubayah, R.O.; Pavlick, R.P.; Schimel, D.S. Towards mapping the diversity of canopy structure from space with GEDI. Env. Res. Lett. 2020, 15, 115006. [Google Scholar] [CrossRef]
- Schweiger, A.K.; Laliberte, E. Plant beta-diversity across biomes captured by imaging spectroscopy. Nat. Commun. 2022, 13, 2767. [Google Scholar] [CrossRef]
- Eichel, J.; Stoffel, M.; Wipf, S. Go or grow? Feedbacks between moving slopes and shifting plants in high mountain environments. Progress in Physical Geography: Earth and Environment 2023, 47, 967–985. [Google Scholar] [CrossRef]
- Restrepo, C.; Walker, L.R.; Shiels, A.B.; Bussmann, R.; Claessens, L.; Fisch, S.; Lozano, P.; Negi, G.; Paolini, L.; Poveda, G. , et al. Landsliding and Its Multiscale Influence on Mountainscapes. BioScience 2009, 59, 685–698. [Google Scholar] [CrossRef]
- Walker, L.R.; Shiels, A.B. Landslide Ecology. Cambridge University Press: Cambridge, 2013. [Google Scholar]
- Myster, R.W.; Fernandez, D.S. Spatial gradients and patch structure on two Puerto Rican landslides. Biotropica 1995, 27, 149–159. [Google Scholar] [CrossRef]
- Tanyaş, H.; Allstadt, K.E.; van Westen, C.J. An updated method for estimating landslide-event magnitude. Earth Surf. Processes 2018, 43, 1836–1847. [Google Scholar] [CrossRef]
- Samia, J.; Temme, A.; Bregt, A.; Wallinga, J.; Guzzetti, F.; Ardizzone, F.; Rossi, M. Do landslides follow landslides? Insights in path dependency from a multi-temporal landslide inventory. Landslides 2017, 14, 547–558. [Google Scholar] [CrossRef]
- Shimokawa, E. A natural recovery process of vegetation on landslide scars and landslide periodicity in forested drainage basings. In Symposium on Effects of Forest Land use on Erosion and Slope Stabililty; O’Loughlin, C.L., Pearce, A.J., Eds.; Environment and Policy Institute East-West Center, University of Hawaii: Honolulu, Hawaii, United States, 1984; pp. 99–107. [Google Scholar]
- Alexandrowicz, Z.; Margielewski, W. Impact of mass movements on geo- and biodiversity in the Polish Outer (Flysch) Carpathians. Geomorphology 2010, 123, 290–304. [Google Scholar] [CrossRef]
- Freund, C.A.; Silman, M.R. Developing a more complete understanding of tropical montane forest disturbance ecology through landslide research. Front. For. Glob. Change 2023, 6, 1091387. [Google Scholar]
- Furusawa, J.; Makoto, K.; Utsumi, S. A large-scale field experiment of artificially caused landslides with replications revealed the response of the ground-dwelling beetle community to landslides. Ecol. Evol. 2023, 13, e9939. [Google Scholar] [CrossRef] [PubMed]
- Neto, C.; Cardigos, P.; Oliveira, S.C.; Zêzere, J.L. Floristic and vegetation successional processes within landslides in a Mediterranean environment. Sci. Total Environ. 2017, 574, 969–981. [Google Scholar] [PubMed]
- He, J.-b.; Wu, Y.-h.; Bing, H.-j.; Zhu, H.; Zhou, J. Soil chronosequence derived from landslides on the upper reach of Minjiang River, western China. J. Mt. Sci. 2023, 20, 1282–1292. [Google Scholar] [CrossRef]
- Lasota, J.; Kraj, W.; Honkowicz, B.; Staszel, K.; Błońska, E. Nutrient status of tree seedlings in a site recovering from a landslide. Forests 2020, 11, 709. [Google Scholar] [CrossRef]
- Ramos Scharrón, C.E.; Castellanos, E.J.; Restrepo, C. The transfer of modern organic carbon by landslide activity in tropical montane ecosystems. Journal of Geophysical Research: Biogeosciences 2012, 117, G03016. [Google Scholar]
- Restrepo, C.; Vitousek, P.; Neville, P. Landslides significantly alter land cover and the distribution of biomass: an example from the Ninole ridges of Hawai'i. Plant Ecology 2003, 166, 131–143. [Google Scholar] [CrossRef]
- Dennison, P.E.; Roberts, D.A. Endmember selection for multiple endmember spectral mixture analysis using endmember average RMSE. Remote Sens. Environ. 2003, 87, 123–135. [Google Scholar] [CrossRef]
- Kluczek, M.; Zagajewski, B.; Kycko, M. Airborne HySpex hyperspectral versus multitemporal Sentinel-2 images for mountain plant communities mapping. Remote Sens. 2022, 14. [Google Scholar] [CrossRef]
- Kokaly, R.F.; Despain, D.G.; Clark, R.N.; Livo, K.E. Mapping vegetation in Yellowstone National Park using spectral feature analysis of AVIRIS data. Remote Sens. Environ. 2003, 84, 437–456. [Google Scholar]
- Marcinkowska-Ochtyra, A.; Zagajewski, B.; Raczko, E.; Ochtyra, A.; Jarocińska, A. Classification of high-mountain vegetation communities within a diverse giant mountains ecosystem using airborne APEX hyperspectral imagery. Remote Sens. 2018, 10, 570. [Google Scholar] [CrossRef]
- Basnet, B.; Vodacek, A. Tracking land use/land cover dynamics in cloud prone areas using moderate resolution satellite data: A case study in Central Africa. Remote Sens. 2015, 7, 6683–6709. [Google Scholar] [CrossRef]
- Bicudo da Silva, R.F.; Millington, J.D.A.; Moran, E.F.; Batistella, M.; Liu, J. Three decades of land-use and land-cover change in mountain regions of the Brazilian Atlantic Forest. Landscape Urban Plan. 2020, 204, 103948. [Google Scholar] [CrossRef]
- Colby, J.D.; Keating, P.L. Land cover classification using Landsat TM imagery in the tropical highlands: The influence of anisotropic reflectance. Int. J. Remote Sens. 1998, 19, 1479–1500. [Google Scholar] [CrossRef]
- Peyre, G.; Osorio, D.; François, R.; Anthelme, F. Mapping the páramo land-cover in the Northern Andes. Int. J. Remote Sens. 2021, 42, 7777–7797. [Google Scholar] [CrossRef]
- Brandt, J.S.; Townsend, P.A. Land use – land cover conversion, regeneration and degradation in the high elevation Bolivian Andes. Landscape Ecol. 2006, 21, 607–623. [Google Scholar] [CrossRef]
- Chen, X.; Chen, W.; Xu, M. Remote-sensing monitoring of postfire vegetation dynamics in the Greater Hinggan Mountain Range Based on long time-series data: Analysis of the effects of six topographic and climatic factors. Remote Sens. 2022, 14. [Google Scholar] [CrossRef]
- Hamunyela, E.; Brandt, P.; Shirima, D.; Do, H.T.T.; Herold, M.; Roman-Cuesta, R.M. Space-time detection of deforestation, forest degradation and regeneration in montane forests of Eastern Tanzania. Int. J. Appl. Earth Obs. 2020, 88, 102063. [Google Scholar]
- Miletić, B.R.; Matović, B.; Orlović, S.; Gutalj, M.; Đorem, T.; Marinković, G.; Simović, S.; Dugalić, M.; Stojanović, D.B. Quantifying forest cover loss as a response to drought and dieback of Norway Spruce and evaluating sensitivity of various vegetation indices using remote sensing. Forests 2024, 15, 662. [Google Scholar] [CrossRef]
- Restrepo, C.; Alvarez, N. Landslides and Their Contribution to Land-cover Change in the Mountains of Mexico and Central America1. Biotropica 2006, 38, 446–457. [Google Scholar] [CrossRef]
- Novellino, A.; Pennington, C.; Leeming, K.; Taylor, S.; Alvarez, I.G.; McAllister, E.; Arnhardt, C.; Winson, A. Mapping landslides from space: A review. Landslides 2024, 21, 1041–1052. [Google Scholar]
- Chapman, D.S. Greater phenological sensitivity to temperature on higher Scottish mountains: new insights from remote sensing. Glob. Change Biol. 2013, 19, 3463–3471. [Google Scholar] [CrossRef]
- Cho, M.A.; Skidmore, A.K. Hyperspectral predictors for monitoring biomass production in Mediterranean mountain grasslands: Majella National Park, Italy. Int. J. Remote Sens. 2009, 30, 499–515. [Google Scholar] [CrossRef]
- Schino, G.; Borfecchia, F.; De Cecco, L.; Dibari, C.; Iannetta, M.; Martini, S.; Pedrotti, F. Satellite estimate of grass biomass in a mountainous range in central Italy. Agrofor. Syst. 2003, 59, 157–162. [Google Scholar] [CrossRef]
- Soenen, S.A.; Peddle, D.R.; Hall, R.J.; Coburn, C.A.; Hall, F.G. Estimating aboveground forest biomass from canopy reflectance model inversion in mountainous terrain. Remote Sens. Environ. 2010, 114, 1325–1337. [Google Scholar] [CrossRef]
- Uscanga, A.; Shuler, S.; Silva, L.C.R. Incorporating small-scale disturbances in models of forest structure and aboveground biomass of tropical mountains. Ecosphere 2024, 15, e4744. [Google Scholar] [CrossRef]
- Ollinger, S.V.; Smith, M.L.; Martin, M.E.; Hallett, R.A.; Goodale, C.L.; Aber, J.D. Regional variation in foliar chemistry and N cycling among forests of diverse history and composition. Ecology 2002, 83, 339–355. [Google Scholar]
- Townsend, P.A. Application of imaging spectroscopy to mapping canopy nitrogen in the forests of the Central Appalachian mountains using Hyperion and AVIRIS. IEEE Trans. Geosci. Remote Sens. 2003, 41, 1347–1354. [Google Scholar] [CrossRef]
- Chadwick, K.D.; Asner, G.P. Landscape evolution and nutrient rejuvenation reflected in Amazon forest canopy chemistry. Ecol. Lett. 2018, 21, 978–988. [Google Scholar] [CrossRef]
- Schneider, F.D.; Longo, M.; Paul-Limoges, E.; Scholl, V.M.; Schmid, B.; Morsdorf, F.; Pavlick, R.P.; Schimel, D.S.; Schaepman, M.E.; Moorcroft, P.R. Remote sensing-based forest modeling reveals positive effects of functional diversity on productivity at local spatial scale. Journal of Geophysical Research: Biogeosciences 2023, 128, e2023JG007421. [Google Scholar] [CrossRef]
- Durán, S.M.; Martin, R.E.; Díaz, S.; Maitner, B.S.; Malhi, Y.; Salinas, N.; Shenkin, A.; Silman, M.R.; Wieczynski, D.J.; Asner, G.P. , et al. Informing trait-based ecology by assessing remotely sensed functional diversity across a broad tropical temperature gradient. Sci. Adv. 2019, 5, eaaw8114. [Google Scholar] [CrossRef] [PubMed]
- Freund, C.A.; Clark, K.E.; Curran, J.F.; Asner, G.P.; Silman, M.R. Landslide age, elevation and residual vegetation determine tropical montane forest canopy recovery and biomass accumulation after landslide disturbances in the Peruvian Andes. J. Ecol. 2021, 109, 3555–3571. [Google Scholar] [CrossRef]
- Li, M.; Ma, C.; Du, C.; Yang, W.; Lyu, L.; Wang, X. Landslide response to vegetation by example of July 25–26, 2013, extreme rainstorm, Tianshui, Gansu Province, China. B. Eng. Geol. Environ. 2021, 80, 751–764. [Google Scholar]
- Yunus, A.P.; Fan, X.; Tang, X.; Jie, D.; Xu, Q.; Huang, R. Decadal vegetation succession from MODIS reveals the spatio-temporal evolution of post-seismic landsliding after the 2008 Wenchuan earthquake. Remote Sens. Environ. 2020, 236, 111476. [Google Scholar]
- Jiao, Q.; Zhang, B.; Liu, L.; Li, Z.; Yue, Y.; Hu, Y. Assessment of spatio-temporal variations in vegetation recovery after the Wenchuan earthquake using Landsat data. Nat. Hazards 2014, 70, 1309–1326. [Google Scholar] [CrossRef]
- Saito, H.; Uchiyama, S.; Teshirogi, K. Rapid vegetation recovery at landslide scars detected by multitemporal high-resolution satellite imagery at Aso volcano, Japan. Geomorphology 2022, 398. [Google Scholar] [CrossRef]
- Asner, G.P.; Mascaro, J. Mapping tropical forest carbon: Calibrating plot estimates to a simple LiDAR metric. Remote Sens. Environ. 2014, 140, 614–624. [Google Scholar] [CrossRef]
- Nakata, Y.; Hayamizu, M.; Ishiyama, N. Assessing primary vegetation recovery from earthquake-induced landslide scars: A real-time kinematic unmanned aerial vehicle approach. Ecol. Eng. 2023, 193, 107019. [Google Scholar]
- Thapa, P.S.; Daimaru, H.; Yanai, S. Analyzing vegetation recovery and erosion status after a large Landslide at Mt. Hakusan, Central Japan. Ecol. Eng. 2024, 198, 107144. [Google Scholar] [CrossRef]
- Badía, D.; Martí, C.; Aznar, J.M.; León, J. Influence of slope and parent rock on soil genesis and classification in semiarid mountainous environments. Geoderma 2013, 193-194, 13–21. [Google Scholar] [CrossRef]
- Feldman, S.B.; Zelazny, L.W.; Baker, J.C. High-elevation forest soils of the Southern Appalachians: I. Distribution of parent materials and soil-landscape relationships. Soil Sci. Soc. Am. J. 1991, 55, 1629–1637. [Google Scholar] [CrossRef]
- Mage, S.M.; Porder, S. Parent material and topography determine soil phosphorus status in the Luquillo Mountains of Puerto Rico. Ecosystems 2013, 16, 284–294. [Google Scholar] [CrossRef]
- Wilson, S.G.; Dahlgren, R.A.; Margenot, A.J.; Rasmussen, C.; O'Geen, A.T. Expanding the Paradigm: The influence of climate and lithology on soil phosphorus. Geoderma 2022, 421, 115809. [Google Scholar] [CrossRef]
- Hahm, W.J.; Rempe, D.M.; Dralle, D.N.; Dawson, T.E.; Lovill, S.M.; Bryk, A.B.; Bish, D.L.; Schieber, J.; Dietrich, W.E. Lithologically controlled subsurface critical zone thickness and water storage capacity determine regional plant community composition. Water Resour. Res. 2019, 55, 3028–3055. [Google Scholar]
- Hahm, W.J.; Riebe, C.S.; Lukens, C.E.; Araki, S. Bedrock composition regulates mountain ecosystems and landcape evolution. Proc. Natl. Acad. Sci. U S A 2014, 111, 3338–3343. [Google Scholar] [CrossRef]
- Watt, M.S.; Pearse, G.D.; Dash, J.P.; Melia, N.; Leonardo, E.M.C. Application of remote sensing technologies to identify impacts of nutritional deficiencies on forests. ISPRS J. Photogramm. Rem. Sens. 2019, 149, 226–241. [Google Scholar]
- Chen, Y.; Wang, Y.; Zhang, F.; Dong, Y.; Song, Z.; Liu, G. Remote sensing for lithology mapping in vegetation-covered regions: Methods, challenges, and opportunities. Minerals 2023, 13, 1153. [Google Scholar] [CrossRef]
- Peyghambari, S.; Zhang, Y. Hyperspectral remote sensing in lithological mapping, mineral exploration, and environmental geology: An updated review. J. Appl. Remote Sens. 2021, 15, 031501. [Google Scholar] [CrossRef]
- Bai, S.; Zhao, J. A new strategy to fuse remote sensing data and geochemical data with different machine learning methods. Remote Sens. 2023, 15, 930. [Google Scholar] [CrossRef]
- Fan, Y.-H.; Wang, H. Application of remote sensing to identify Copper–Lead–Zinc deposits in the Heiqia area of the West Kunlun Mountains, Chinas. Sci. Rep. 2020, 10, 12309. [Google Scholar] [CrossRef]
- He, J.; Chen, D.; Zhan, Y.; Ren, X.; Li, Q. A retrieval model of surface geochemistry composition based on remotely sensed data. Open Geosci. 2023, 15, 20220514. [Google Scholar] [CrossRef]
- Riaza, A.; Strobl, P.; Beisl, U.; Hausold, A.; Müller, A. Spectral mapping of rock weathering degrees on granite using hyperspectral DAIS 7915 spectrometer data. Int. J. Appl. Earth Obs. 2001, 3, 345–354. [Google Scholar]
- Sousa, F.J.; Sousa, D.J. Spatial patterns of chemical weathering at the basal tertiary nonconformity in California from multispectral and hyperspectral optical remote sensing. Remote Sens. 2019, 11, 2528. [Google Scholar] [CrossRef]
- Thomas, M.; Walter, M.R. Application of hyperspectral infrared analysis of hydrothermal alteration on Earth and Mars. Astrobiology 2002, 2, 335–351. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, T.; Chabrillat, S.; Pignatti, S.; Milewski, R.; Karyotis, K.; Brell, M.; Ruhtz, T.; Bochtis, D.; Zalidis, G. Evaluation of airborne HySpex and spaceborne PRISMA hyperspectral remote sensing data for soil organic matter and carbonates estimation. Remote Sens. 2023, 15, 1106. [Google Scholar] [CrossRef]
- Baptista, G.M.M.; Corrêa, R.S.; dos Santos, P.F.; Madeira Netto, J.S.; Meneses, P.R. Use of imaging spectroscopy for mapping and quantifying the weathering degree of tropical soils in Central Brazil. Appl. Environ. Soil Sci. (Online) 2011, 2011, 641328. [Google Scholar]
- Gomez, C.; Lagacherie, P.; Coulouma, G. Regional predictions of eight common soil properties and their spatial structures from hyperspectral Vis–NIR data. Geoderma 2012, 189-190, 176–185. [Google Scholar] [CrossRef]
- Leone, A.P.; Wright, G.G.; Corves, C. The application of satellite remote sensing for soil studies in upland areas of Southern Italy. Int. J. Remote Sens. 1995, 16, 1087–1105. [Google Scholar] [CrossRef]
- Palacios-Orueta, A.; Pinzón, J.E.; Ustin, S.L.; Roberts, D.A. Remote sensing of soils in the Santa Monica Mountains: II. Hierarchical foreground and background analysis. Remote Sens. Environ. 1999, 68, 138–151. [Google Scholar]
- Yu, H.; Kong, B.; Wang, G.; Du, R.; Qie, G. Prediction of soil properties using a hyperspectral remote sensing method. Arch. Acker Pfl. Boden. 2018, 64, 546–559. [Google Scholar]
- Zou, J.; Wei, Y.; Zhang, Y.; Liu, Z.; Gai, Y.; Chen, H.; Liu, P.; Song, Q. Remote sensing inversion of soil organic matter in cropland combining topographic factors with spectral parameters. Front. Environ. Sci. 2024, 12, 1420557. [Google Scholar] [CrossRef]
- Ge, Y.; Thomasson, J.A.; Sui, R. Remote sensing of soil properties in precision agriculture: A review. Front. Earth Sci. 2011, 5, 229–238. [Google Scholar] [CrossRef]
- Mulder, V.L.; de Bruin, S.; Schaepman, M.E.; Mayr, T.R. The use of remote sensing in soil and terrain mapping — A review. Geoderma 2011, 162, 1–19. [Google Scholar] [CrossRef]
- Park, J.; Kim, K. Quantification of rock mass weathering using spectral imaging. J. South. Afr. Inst. Min. Metall. 2019, 119, 1039–1046. [Google Scholar]
- van der Meer, F.D.; van der Werff, H.M.A.; van Ruitenbeek, F.J.A.; Hecker, C.A.; Bakker, W.H.; Noomen, M.F.; van der Meijde, M.; Carranza, E.J.M.; Smeth, J.B.d.; Woldai, T. Multi- and hyperspectral geologic remote sensing: A review. Int. J. Appl. Earth Obs. 2012, 14, 112–128. [Google Scholar] [CrossRef]
- Yu, H.; Kong, B.; Wang, Q.; Liu, X.; Liu, X. 14 - Hyperspectral remote sensing applications in soil: a review. In Hyperspectral Remote Sensing; Pandey, P.C., Srivastava, P.K., Balzter, H., Bhattacharya, B., Petropoulos, G.P., Eds.; Elsevier, 2020; pp. 269–291. [Google Scholar]
- Vitousek, P.; Asner, G.P.; Chadwick, O.A.; Hotchkiss, S. Landscape-level variation in forest structure and biogeochemistry across a substrate age gradient in Hawaii. Ecology 2009, 90, 3074–3086. [Google Scholar] [CrossRef] [PubMed]
- Porder, S.; Asner, G.P.; Vitousek, P.M. Ground-based and remotely sensed nutrient availability across a tropical landscape. Proc. Natl. Acad. Sci. U S A 2005, 102, 10909–10912. [Google Scholar] [CrossRef]
- Weintraub, S.R.; Taylor, P.G.; Porder, S.; Cleveland, C.C.; Asner, G.P.; Townsend, A.R. Topographic controls on soil nitrogen availability in a lowland tropical forest. Ecology 2015, 96, 1561–1574. [Google Scholar] [CrossRef]
- Chadwick, K.D.; Asner, G.P. Tropical soil nutrient distributions determined by biotic and hillslope processes. Biogeochemistry 2016, 127, 273–289. [Google Scholar] [CrossRef]
- Amaral, C.H.d.; Almeida, T.I.R.d.; Souza Filho, C.R.d.; Roberts, D.A.; Fraser, S.J.; Alves, M.N.; Botelho, M. Characterization of indicator tree species in neotropical environments and implications for geological mapping. Remote Sens. Environ. 2018, 216, 385–400. [Google Scholar] [CrossRef]
- Chakraborty, R.; Kereszturi, G.; Pullanagari, R.; Durance, P.; Ashraf, S.; Anderson, C. Mineral prospecting from biogeochemical and geological information using hyperspectral remote sensing - Feasibility and challenges. J. Geochem. Explor. 2022, 232, 106900. [Google Scholar]
- Collins, W.; Chang, S.-H.; Raines, G.L.; Canney, F.; Ashley, R. Airborne biogeophysical mapping of hidden mineral deposits. Econ. Geol. 1983, 78, 737–749. [Google Scholar] [CrossRef]
- Goetz, A.F.H.; Rock, B.N.; Rowan, L.C. Remote sensing for exploration: An overview. Econ. Geol. 1983, 78, 573–590. [Google Scholar] [CrossRef]
- Hede, A.N.H.; Kashiwaya, K.; Koike, K.; Sakurai, S. A new vegetation index for detecting vegetation anomalies due to mineral deposits with application to a tropical forest area. Remote Sens. Environ. 2015, 171, 83–97. [Google Scholar] [CrossRef]
- Hede, A.N.H.; Koike, K.; Kashiwaya, K.; Sakurai, S.; Yamada, R.; Singer, D.A. How can satellite imagery be used for mineral exploration in thick vegetation areas? Geochem. Geophy. Geosy. 2017, 18, 584–596. [Google Scholar] [CrossRef]
- Milton, N.M.; Collins, W.; Chang, S.-H.; Schmidt, R.G. Remote detection of metal anomalies on Pilot Mountain, Randolph County, North Carolina. Econ. Geol. 1983, 78, 605–617. [Google Scholar] [CrossRef]
- Rodriguez-Gomez, C.; Kereszturi, G.; Jeyakumar, P.; Pullanagari, R.; Reeves, R.; Rae, A.; Procter, J.N. Remote exploration and monitoring of geothermal sources: A novel method for foliar element mapping using hyperspectral (VNIR-SWIR) remote sensing. Geothermics 2023, 111, 102716. [Google Scholar]
- Rowan, L.C.; Crowley, J.K.; Schmidt, R.G.; Ager, C.M.; Mars, J.C. Mapping hydrothermally altered rocks by analyzing hyperspectral image (AVIRIS) data of forested areas in the Southeastern United States. J. Geochem. Explor. 2000, 68, 145–166. [Google Scholar] [CrossRef]
- Sabins, F.F. Remote sensing for mineral exploration. Ore Geol. Rev. 1999, 14, 157–183. [Google Scholar] [CrossRef]
- Schellekens, J., H.; Gilbes, F.; Rivera, G.D.; Yuri, C.Y.; Chardón, S.; Fong, Y. Reflectance spectra of tropical vegetation as a response to metal enrichment in the substrate of west-central Puerto rico. Caribb. J. Earth Sci. 2005, 39, 9–12. [Google Scholar]
- Shin, J.H.; Yu, J.; Wang, L.; Kim, J.; Koh, S.-M. Investigation of spectral variation of pine needles as an indicator of arsenic content in soils. Minerals 2019, 9, 498. [Google Scholar] [CrossRef]
- Alekseenko, V.A.; Shvydkaya, N.V.; Alekseenko, A.V.; Machevariani, M.M.; Bech, J.; Pashkevich, M.A.; Puzanov, A.V.; Nastavkin, A.V.; Roca, N. Element accumulation patterns of native plant species under the natural geochemical stress. Plants 2021, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Brooks, R.R.; Dunn, C.E.; Hall, G.E.M. Biological Systems in Mineral Exploration and Processing; Ellis Horwood: New York, United States, 1995. [Google Scholar]
- Dunn, C.E. Biogeochemistry in Mineral Exploration; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Ufimtseva, M.D. The patterns in accumulation of chemical elements by higher plants and their responses in biogeochemical provinces. Geochem. Int. 2015, 53, 441–455. [Google Scholar] [CrossRef]
- Hardiman, B.S.; Gough, C.M.; Butnor, J.R.; Bohrer, G.; Detto, M.; Curtis, P.S. Coupling Fine-Scale Root and Canopy Structure Using Ground-Based Remote Sensing. Remote Sens. 2017, 9. [Google Scholar] [CrossRef]
Figure 1.
Workflows to derive functional traits from remotely sensed data and estimate functional diversity metrics. Stage 1a: Field Data. Leaf samples and spectra are collected in the field from field plots and plant canopies; leaf samples undergo analytical work. Stage 1b: RS Data. Optical and LiDAR data are collected at sampling sites to provide spectral and structural information of vegetation that may require various levels of preprocessing, such as masking of shade and bare ground. Stage 2: Modeling leaf trait- spectra relationships. Leaf and canopy traits are modelled as a function of light reflectance using different approaches. Stage 3: Scaling. Models developed as part of Stage 2 are used with RS data to assign trait values to pixels. Stage 4: Spectral Features. Traits, principal components, spectral species derived from remotely sensed data represent features used to estimate diversity metrics. Stage 5a: Functional Groups. Clustering of pixels from the functional trait maps at known sites (plots) can be used to define forest functional groups. Stage 5b: Functional Diversity Metrics. Pixels from the functional trait maps can be used to estimate functional diversity metrics. Single traits are used to estimate Community Weighted Means (CWM) whereas multiple traits to estimate Functional Richness (FRic), Functional Dispersion (FDis), Functional Divergence (FDiv), and Functional Evenness (FEv = Functional Evenness). Stage 5c: Diversity-Scale Relationships. Spectral features and functional diversity metrics can be used to understand the extent of their variability with scale [alpha (SDα), beta (LCSDb), and gamma (SDγ) diversity]. Additionally, metrics including Feature Contribution to Alpha Diversity highlight the contributions of bands to spectral diversity of the region (FCAD) [53]. Stage 5d: Scaling up to Regions. Depending upon the extent of the region, scaling may involve additional steps to model average spectra as a function of environmental variables.
Figure 1.
Workflows to derive functional traits from remotely sensed data and estimate functional diversity metrics. Stage 1a: Field Data. Leaf samples and spectra are collected in the field from field plots and plant canopies; leaf samples undergo analytical work. Stage 1b: RS Data. Optical and LiDAR data are collected at sampling sites to provide spectral and structural information of vegetation that may require various levels of preprocessing, such as masking of shade and bare ground. Stage 2: Modeling leaf trait- spectra relationships. Leaf and canopy traits are modelled as a function of light reflectance using different approaches. Stage 3: Scaling. Models developed as part of Stage 2 are used with RS data to assign trait values to pixels. Stage 4: Spectral Features. Traits, principal components, spectral species derived from remotely sensed data represent features used to estimate diversity metrics. Stage 5a: Functional Groups. Clustering of pixels from the functional trait maps at known sites (plots) can be used to define forest functional groups. Stage 5b: Functional Diversity Metrics. Pixels from the functional trait maps can be used to estimate functional diversity metrics. Single traits are used to estimate Community Weighted Means (CWM) whereas multiple traits to estimate Functional Richness (FRic), Functional Dispersion (FDis), Functional Divergence (FDiv), and Functional Evenness (FEv = Functional Evenness). Stage 5c: Diversity-Scale Relationships. Spectral features and functional diversity metrics can be used to understand the extent of their variability with scale [alpha (SDα), beta (LCSDb), and gamma (SDγ) diversity]. Additionally, metrics including Feature Contribution to Alpha Diversity highlight the contributions of bands to spectral diversity of the region (FCAD) [53]. Stage 5d: Scaling up to Regions. Depending upon the extent of the region, scaling may involve additional steps to model average spectra as a function of environmental variables.
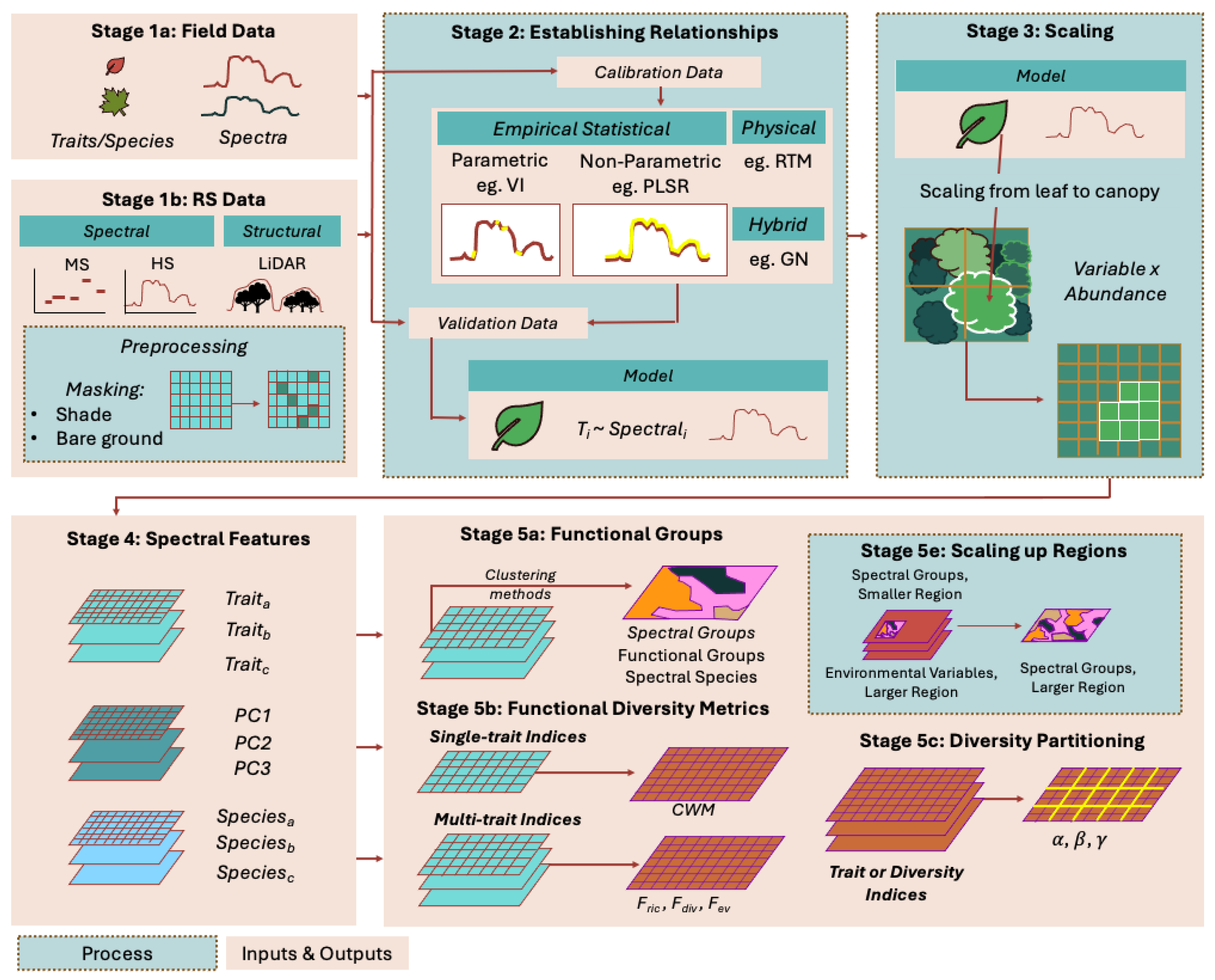
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



