Submitted:
08 October 2024
Posted:
09 October 2024
You are already at the latest version
Abstract
During their life cycle, plants constantly respond to environmental changes. Abiotic stressors affect the photosynthetic and respiratory processes of plants. Reactive oxygen species (ROS) are produced during aerobic metabolism and play an important role as regulatory mediators in signaling processes, activating the plant's protective response to abiotic stress and restoring "oxidation-reduction homeostasis". Cells develop normally if the rates of ROS production and the ability to neutralize them are balanced. To implement oxidation-reduction signaling, this balance must be disrupted either by an increase in ROS concentration or a decrease in the activity of one or more antioxidant systems. Under abiotic stress, plants accumulate excessive amounts of ROS and if the ROS content exceeds the threshold amount dangerous for living organisms, it can lead to damage to all major cellular components. Adaptive resistance of plants to abiotic stressors depends on a set of mechanisms of adaptation to them. The accumulation of ROS in the cell depends on the type of abiotic stress, the strength of its impact on the plant, the duration of its impact and the recovery period. The aim of this review is to provide a general understanding of the processes occurring during ROS homeostasis in plants, oxidation-reduction processes in cellular compartments in response to abiotic stress and the participation of ROS in signaling processes activating adaptation processes to abiotic stress.
Keywords:
reactive oxygen species
; oxidative stress
; abiotic stress
; signaling wave
; acclimatization
1. Introduction
Plants are constantly exposed to stress as a result of environmental changes, so plants have developed a complex acclimation system. To activate this system, at least two types of signals are needed: the so-called reactive oxygen species (ROS) wave, which spreads at a tremendous speed from the initial site of stressor impact throughout the plant, and a specific signal from the type of abiotic stressor. The ROS wave, caused by abiotic stress, triggers a cascade of intercellular communication interactions that spreads through various plant tissues and is capable of transmitting a signal over long distances [1]. However, it has been noted that various abiotic stresses can also lead to specific fluctuations in the ROS wave, causing certain specific rearrangements of transcriptomes and metabolomes [1]. ROS wave is used for signaling and triggering plant adaptation mechanisms to abiotic stress. As a result of the ROS wave signaling, transcriptomic and metabolomic reprogramming of cells occurs. It is proposed that the ROS wave activates a general acclimation response, independent of specific abiotic stressors, which may induce broad transcriptome and metabolome reprogramming and render plants resilient to multiple different stresses.
The formation of free radicals in biological materials was discovered in the 1950s and it was hypothesized that oxygen radicals were generated by enzymatic reactions in vivo. Oxygen radicals were thought to be cell damaging and to cause oxidative damage to DNA, proteins, lipids, and other cellular components [2]. However, more recent studies have revealed positive functions of free radicals, including their involvement in metabolic processes and signaling pathways [3].
ROS have been shown to be produced to initiate redox signals to regulate a wide range of cellular reactions essential for life [4]. ROS, through redox signaling in plants, initiate a program of gene expression that helps maintain cellular redox homeostasis and are also involved in biological processes related to cell proliferation and differentiation [5]. While cell death was previously thought to be a direct consequence of ROS-induced oxidative stress, ROS are now believed to activate programmed cell death (PCD) pathways [6]. Thus, ROS-induced redox signaling acts as an important regulator of various physiological responses. The role of ROS as oxidants and/or components of redox signaling largely depends on the delicate balance between ROS production and detoxification in different subcellular organelles [7]. This is tightly controlled by the entire redox sensing and signaling networks that regulate cellular ROS levels in space and time by modifying the ROS production and detoxification mechanisms [8]. In case of low ROS in the cell, they act as positive signaling molecules actively participating in many biologically important processes. With increasing ROS concentration, they can act as highly toxic molecules. The accumulation of ROS in the cell depends on many parameters. The complex process of plant adaptation to abiotic stress involves a whole range of oxidant-antioxidant interaction mechanisms capable of flexibly altering redox signals [9].
2. Reactive Oxygen Species (ROS) and Their Formation in Plant Cells
Oxygen is an essential element required for normal plant development. In the ground state, oxygen (triplet oxygen, 3O2) has two unpaired electrons, which is not reactive [10]. In the process of oxidation-reduction reactions, molecular oxygen is an electron acceptor, it is able to attach from one to three electrons, which leads to the formation of reactive oxygen species (ROS). In biological systems, transition metal ions (Fe2+, Cu+) and semiquinones can act as electron donors, resulting in the activation of molecular oxygen [2]. The term ROS includes not only free radicals (superoxide radical, O2•–, and hydroxyl radical, OH•, perhydroxy radical (HO2•), peroxyl (RO2•), carbonate (CO3•−), semiquinone (SQ•-), alkoxy radical (RO•), and peroxy radical (ROO•)), but also molecules such as hydrogen peroxide (H2O2), singlet oxygen (1O2), ozone (O3), organic hydroperoxide (ROOH), hypoiodous acid (HOI), hypobromous acid (HOBr), and hypochlorous acid (HOCl) (Figure 1 ) [11,12,13,14]
As a result of oxidation-reduction reactions occurring in various compartments of plant cells, ROS are formed due to incomplete or partial reduction of oxygen molecules [15]. Several reactions associated with ROS formation are shown in Figure 2.
The mechanisms of ROS formation in biological systems are carried out in the course of non-enzymatic and enzymatic reactions. The mechanisms that trigger a certain type of reaction are regulated by the concentration of oxygen in the system. It is believed that at high concentrations of O2, ROS formation occurs via the non-enzymatic pathway, and at very low concentrations of O2, via the enzymatic pathway [16].
The most common ROS with high oxidizing capacity is the superoxide anion radical (O2•-). The superoxide anion is formed as a result of the monovalent reduction of the triplet state of molecular oxygen (Figure 2) [11]. Among the sources of ROS formed via the enzymatic pathway, mention should be made of xanthine oxidase (XO), the enzyme responsible for the initial activation of oxygen. XO can use xanthine, hypoxanthine, or acetaldehyde as electron donors (Figure 3) [17]. The latter has been shown to accumulate under oxygen deficiency, especially during hypoxia [18].
The next important step in ROS conversion is the dismutation of the superoxide anion (O2•–) by superoxide dismutase (SOD) to form H2O2. The level of H2O2 is regulated by an enzymatic set of catalases (CAT) and peroxidases (PX), localized in almost all compartments of the plant cell. The main function of PX is the neutralization of H2O2, however, this family of enzymes can also catalyze the formation of O2•– and H2O2 as a result of NADH oxidation. The oxidation of NADH with H2O2 produces the radical NAD•, which reduces O2 to O2•–, some of which is dismutated to H2O2 and O2 [19]. Thus, peroxidases and catalases play an important role in the fine regulation of ROS concentration in the cell through the activation and deactivation of H2O2 [11]. Alternatively, hydrogen peroxide can be converted to water by the enzymes catalase or glutathione peroxidase. In the glutathione peroxidase reaction, glutathione is oxidized to glutathione disulfide, which can be converted back to glutathione by glutathione reductase (GR) in a process that consumes NADPH [20].
Figure 4.
Oxidation and reduction reactions of GSH.
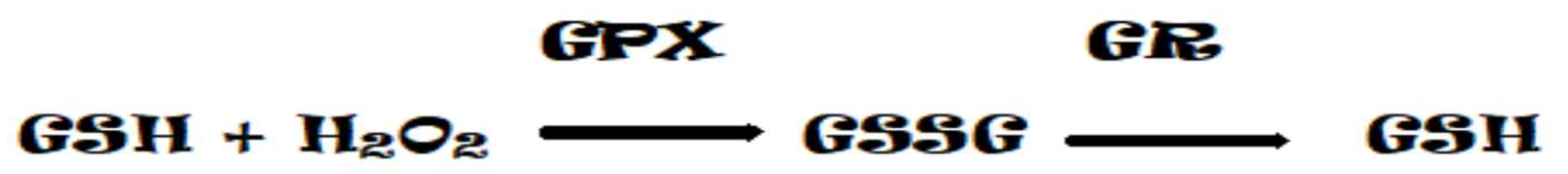
By scavenging O2•−, SODs reduce the risk of OH• formation via a metal-catalyzed Haber-Weiss-type reaction, as this reaction occurs 10,000 times faster than spontaneous dismutation [21]. The reaction results in the formation of H2O2, which is also a ROS with a dual role (toxicity and signaling). H2O2 can be effectively detoxified if the cellular antioxidant defense mechanism is sufficient. In some ways, this enzyme is unique in that its activity determines the concentrations of O2•− and H2O2, the two substrates of the Haber-Weiss reaction, and so it is likely to play a central role in the defense mechanism (Figure 5} [22].
In biological tissues, superoxide can also be non-enzymatically converted to the non-radical species hydrogen peroxide and singlet oxygen (1O2) [23]. In the presence of reduced transition metals (e.g., iron or copper ions), hydrogen peroxide can be converted to the highly reactive hydroxyl radical (•OH).
Another aggressive oxidant is the hydroxyl radical (•OH), which is involved in oxidative damage to DNA bases [14]. •OH is produced by iron-catalyzed H2O2 and O2•− [24,25,26].•OH can be formed by the ionization of H2O and also by photolytic decomposition of alkyl hydroperoxides. It is generally assumed that HO• in biological systems is formed by the redox cycle of the Fenton reaction, where free iron (Fe2+) reacts with hydrogen peroxide (H2O2), and the Haber-Weiss reaction, which results in the formation of Fe2+ when superoxide reacts with ferric iron (Fe3+) (Figure 5). In addition to the iron redox cycle described above, a number of other transition metals, including Cu, Ni, Co, and V, may be responsible for the formation of HO• in living cells.
3. Oxidative Stress
Redox reactions are common in living things and are responsible for the majority of ROS generation [27]. Excessive ROS production, if not scavenged, can lead to so-called “oxidative stress”. Physiological levels and types of ROS in different compartments are highly dependent on the energetic load to which the cellular response is subjected. Even within a single cell, there are at least eight different organelle compartments (mitochondrial matrix, lysosomes, smooth ER/SR, rough ER, Golgi, peroxisomes, nucleus, cytosol), each with its own redox balance [28]. Therefore, the term oxidative stress should be used when the levels and types of oxidants in a cell or organelle, on average, significantly exceed the level associated with normal homeostatic function for each compartment. In plant cells, redox homeostasis is a normal state in which the antioxidant defense system aims to maintain a balance between ROS production and antioxidant activity [29]. Under normal conditions, ROS production and levels are safe for normal cellular function, including proliferation, differentiation, signaling, and intercellular communication [12,30,31]. The relationship between ROS and redox changes/regulation in cells is commonly referred to as redox biology and is thought to play a key role in ROS-mediated signaling and/or cellular metabolic regulation [1,32,33,34]. Because membranes can function as barriers to redox levels, each subcellular compartment can contain its own redox state that will correspond to its own steady-state ROS level, contributing to the formation of a cellular-specific ROS signature during abiotic stress [35].
Reactive oxygen species are a by-product of aerobic metabolism in various cellular organelles such as chloroplasts, mitochondria, peroxisomes, plasma membranes and cell wall [14,36]. The specific generation of ROS in the cell is highly localized and different pathways are extensively involved in this process [37,38]. To better understand the ROS scavenging tactics in different subcellular compartments, it is imperative to first study the subcellular compartment-specific ROS generation in cells.
ROS, as signaling molecules, play an important role in biological processes, the so-called “redox biology” [6,39,40,41]. For participation in the redox signaling process, the basal amount of ROS is crucial [6,41]. In case the levels of reduced glutathione are too high, the so-called “reductive stress” can occur, which also has detrimental consequences for the cell [42,43]. However, under abiotic stress, when ROS levels in plants are above or below normal levels, a redox imbalance occurs that can cause impairment and/or even loss of certain cellular functions. Low ROS levels trigger signaling that alters normal plant metabolism, while excess ROS initiates oxidative damage to cells [6,26]. Therefore, the mechanisms that ensure a stable equilibrium between ROS production and its neutralization must function together and be harmonized [44,45].
Thus, redox homeostasis is a prerequisite for the normal functioning of cells and organisms [46]. It has been previously argued that the cell normally maintains cytosolic thiols in a highly reduced redox state, thus not supporting the existence of reductive stress [42]. Under stress conditions, ROS accumulation in plant cells disrupts the “redox state” of various proteins, enzymes, receptors, altering or participating in various signal transduction pathways in response to an abiotic factor [41,44]. There is a growing understanding that the role of redox metabolism enzymes and metabolites goes far beyond a simple ROS scavenging function. First, as specialized processors of ROS signaling, these enzymes act as integral parts of a complex signaling system [47].
4. Regulation of ROS in Cellular Compartments
Reactive oxygen species are a by-product of aerobic metabolism in various cellular organelles such as chloroplasts, mitochondria, peroxisomes, plasma membranes, and cell walls (Figure 6) [14,36]. ROS generation is specific in different cellular compartments [37,48]. Abiotic stresses that limit CO2 availability through stomatal closure enhance the production of ROS such as O2•- and 1O2 in chloroplasts, which in turn can initiate retrograde and anterograde signaling [49]. ROS signaling molecules are mainly produced in the apoplast by NADPH oxidases, some oxidases and peroxidases, and in other cellular compartments through various pathways [50,51,52,53]. The signaling as well as metabolic processes are represented as a continuous process of ROS production and scavenging, which occurs in all cellular compartments [44]. Each cellular compartment has its own ROS level, which determines its ROS signature and depends on the cell type, the type and strength of the stressor. With the help of a system of sensors that decipher the cellular signature, the mechanisms of adaptation to various abiotic stresses are activated.
4.1. Regulation of ROS in Сhloroplasts
Chloroplasts are considered to be the major site of ROS production, which depends on the interaction of chlorophyll (Chl) and light [14,54]. Under stress conditions, stomatal conductance and CO2 assimilation rate are significantly reduced and thus generate excited triplet Chl (3Chl.), which inhibits photosynthesis etc. and promotes excessive ROS generation [55,56]. Photosystems I and II (PS I and PS II) are the major sites in chloroplasts where 1O2 and O2•− are largely produced [2]. However, the amount of O2•− generated in PS I by the Mehler reaction is converted to H2O2 by SOD. ROS is generated in chloroplasts in the reaction centers via PS I and II by trapping excess photons in PS II and scavenging electrons by molecular oxygen via PS [49,57,58]. Excess energy in PS II leads to the formation of the triplet state of 3Chl. in the antenna complex of PS II. In PS II, 1O2 is formed due to energy transfer, as well as the formation of O2•−, H2O2, and OH• [59,60,61,62,63]. In the absence of stress, electrons leak from the excited PS, reducing NADP to NADPH, which then enters the Calvin cycle where CO2 is reduced as an electron acceptor. Conversely, the electron transport chain (ETC) is overloaded, resulting in electron leakage from ferredoxin to O2, forming O2•− [64]. Under high light intensity with low CO2 demand, direct electron transfer to molecular oxygen occurs due to stomatal closure via the Mehler reaction in PS I [65]. In PS I, the conversion of O2•− to H2O2 depends on inefficient photochemical and non-photochemical quenching [65]. These superoxide radicals are converted to H2O2, which is further converted to water [66]. While SODs catalyze the dismutation of O2•− to O2 and H2O2, thylakoid membrane-associated ascorbate peroxidases (APXs) are involved in the detoxification of H2O2 via the ascorbate-glutathione cycle [57]. This electron sink (also called the water–water cycle) is critical for balancing the oxidation and reduction of the electron transport chain to maximize photosynthetic efficiency [67].
Inefficient photochemical quenching under various stress conditions results in excessive shortening of the electron transport chain. Excess energy trapped in PS II then promotes the formation of 1O2, thereby also promoting the oxidation of PS II-bound electron acceptors. OH•, the most reactive free radical, is formed from H2O2 via the Fenton reaction in the presence of reduced transition metal ions such as Cu+ or Fe2+, which are formed by the reduction of Cu2+ and Fe3+ (Figure 5). Although chloroplasts have evolved antioxidants and scavenging systems to maintain ROS homeostasis, severe or prolonged stress conditions can rapidly deplete detoxifiers and inactivate scavenging systems. Consequently, unfavourable conditions lead to overproduction of ROS and increased damage to chloroplasts due to the reactivity of ROS towards macromolecules in their vicinity.
4.2. Regulation of ROS in Mitochondria
In non-green parts of plants, mitochondria are the main site of ROS generation [68]. ROS produced in mitochondria reduce both mitochondrial energy transport and other subcellular functions. Respiratory complexes I and III are the main sources of mitochondrial ROS, especially O2•−. Next, O2•−formed in both complexes is converted into H2O2 as a result of catalysis by MnSOD and Cu/ZnSOD dismutases [69,70].
ROS production in mitochondria is lower than in chloroplasts. The mitochondrial respiratory chain was originally described as flavin and cytochrome c-containing proteins in the inner matrix of mitochondria [71,72]. This model proposed that the four major complexes, i.e., NADH-coenzyme Q reductase (complex I), succinate coenzyme Q reductase or succinate dehydrogenase (complex II or SDH), ubiquinol cytochrome c reductase (complex III), and cytochrome c oxidase (complex IV) of the respiratory chain, are randomly distributed in the matrix and linked by the redox active enzymes coenzyme Q and cytochrome c [73], This model was refined with complexes I, III, and IV forming supercomplexes that allow efficient electron transfer with minimal O2•− formation. However, ROS from complexes I, II, and III are not only involved in the random release of electrons from the ETC and their transfer to molecular oxygen, but are now considered important mediators in physiological cell signaling. ROS production must be strictly controlled to avoid its overproduction, which can cause damage to mitochondrial and extramitochondrial macromolecules and cause cell death [74].
Under abiotic stresses, such as high temperatures and/or drought, respiration rate is high and transpiration and photosynthesis are low, increasing the need for mitochondrial ATP to compensate for ATP production by chloroplasts [47]. In the mitochondria, O2 is reduced to O2•− by NADPH dehydrogenase [complex I] [73]. Although mitochondria are the source of ROS production, this organelle also has its own detoxification system consisting of an alternative oxidase (AOX) and MnSOD. O2•− produced in this organelle is rapidly converted to stable, membrane-permeable H2O2 by MnSOD or ascorbate peroxidase (APX). AOX works in concert with MnSOD to maintain the reducing O2•− to O2 [68,74]. Giraud et al. [75] reported that AOX mutants of Arabidopsis were sensitive to drought and light stress, suggesting a role for this enzyme in ROS detoxification. The greatest amount of ROS was found in mitochondria as O2•− molecules, which are converted during detoxification by MnSOD and APX to H2O2 and eventually O2 [76]. As in chloroplasts, mitochondria produce ROS even in an unstressed state at basal levels, and any form of stress causes a decrease in ROS levels due to ATP synthesis [68,69,77,78].
4.3. Regulation of ROS in Peroxisomes
Another important site for ROS production is the peroxisomes, where a number of oxidases catalyze various reactions and generate H2O2 and O2•− as by-products. Glycolate oxidase (GOX) is thought to be the major source of ROS production in peroxisomes [79]. This GOX in peroxisomes induces stomatal closure; as a result, the rate of stomatal gas exchange is significantly reduced and thus reduces CO2 for RubisCO formation, inducing photorespiration and H2O2 production [80]. In addition, xanthine oxidase (XOD) activity can also generate O2•− and uric acid in the peroxisomal matrix, which are further dismutated to H2O2 by the metalloenzymes SOD and urate oxidase, respectively [81,82]. As in mitochondria and chloroplasts, the peroxisome produces O2•− even during normal metabolism. Due to the delicate balance between uptake and production, ROS levels in peroxisomes are kept under control [83]. There are two O2•− production sites in the peroxisomal matrix, where xanthine oxidases convert xanthine and hypoxanthine to uric acid and O2•−, and at the proximal membrane, O2 is used as an electron acceptor by NADH and cytochrome b to generate O2•− [84]. Metabolic processes that produce H2O2 in the peroxisome include β-oxidation, radical disproportionation, and the flavin oxidase pathway [38,85,86]. CATs detoxify the H2O2 system, while APX and ascorbic acid (AsA)-GSH scavenge H2O2 in the peroxisome [44,87,88]. Reduced AsA-GSH levels result in lipid peroxidation in peroxisomes. In addition to these enzymes, POX, a polyamine catabolizing enzyme, has been shown to regulate stress-responsive genes by facilitating ROS production and removal in the peroxisome [24,37,89].
4.4. Regulation of ROS in the Apoplast
NADPH oxidases produced in stomatal guard cells in Arabidopsis produce ROS in the apoplast as a result of ABA-induced stomatal closure [90,91]. AtRbohD and AtRbohF are two genes that regulate NADPH oxidases in Arabidopsis [92,93]. In addition to NADPH oxidases, peroxidases, cell wall-associated oxidases, polyamine oxidases, and oxalate oxidases play a role in H2O2 generation in the apoplast [75,94]. One of the important sources of apoplastic ROS production are cell wall-associated oxidases [75]. Oxalate oxidase, cell wall protein, is known to release H2O2 and CO2 from oxalic acid [95]. This enzyme is mainly involved in plant defense against biotic and abiotic stress. Amine oxidases are found in the apoplast and contribute to plant defense by producing H2O2. These oxidases induce oxidative deamination of polyamines with the help of cofactors. As observed by Heyno et al., the generation of hydroxyl ions in the apoplastic region of the cell completely or partially promotes the formation of cell wall-associated peroxidases [96]. Increased H2O2 production leads to increased levels of polyamines and Ca2+. This leads to increased production of H2O2, which in turn activates the antioxidant mechanism along with increased synthesis of higher polyamines and second messengers such as Ca2+. ABA levels are also elevated, leading to activation of polyamine-activated signaling pathways in response to abiotic stress [89].
Various enzymes promote ROS formation in the apoplast, among which the most important are quinine reductase, NADPH oxidase, SOD, and PX [13,14]. Some apoplastic enzymes can also lead to ROS production under normal and stress conditions. Other oxidases responsible for the transfer of two electrons to oxygen (amino acid oxidases and glucose oxidase) can promote the accumulation of H2O2. Also, extracellular germline-like oxalate oxidase catalyzes the formation of H2O2 and CO2 from oxalate in the presence of oxygen [97]. Amine oxidases catalyze the oxidation of biogenic amines to the corresponding aldehyde with the release of NH3 and H2O2. Data on the accumulation of polyamine (putrescine) during oxygen starvation in rice and wheat shoots [98] and the predominant localization of amine oxidase in the apoplast suggest the participation of amine oxidase in the production of H2O2 during oxygen starvation.
4.5. Regulation of ROS in Cell Walls and Plasma Membranes
Plant cell walls accumulate oxidative radicals OH•, O2•−, H2O2 and 1O2 under stress. Peroxidases, lipoxygenases, oxidases and polyamines localized in the cell walls are responsible for ROS formation. These oxygen radicals are also responsible for the lipid peroxidation of polyunsaturated fatty acids found in the plant cell wall, which produce ROS [99]. Peroxidases found in cell walls catalyze the formation of H2O2 via NADH, through malate dehydrogenase. ROS generation by cell wall-associated peroxidases triggers biotic responses and changes in K+ levels in drought-stressed plants [38,100]. Liu et al. in their review described the role of polyamine in plant stress response as mediated by antioxidant systems or ROS suppression [101]. Therefore, exogenous application of polyamines or activation of polyamine-related genes would result in activation of antioxidant processes in situ in plants exposed to drought, salinity, nutrient deficiency, temperature and other stresses [37,77,102].
As for the plasma membrane, NADPH oxidases localized therein are responsible for the generation of O2•− by transferring an electron from NADPH to O2, which is then converted by SOD to H2O2. NADPH oxidases in plasma membranes play a crucial role in the stress response of plant cells to environmental factors such as abiotic stresses [77,100],. In certain cases, several enzymes have been found to catalyze the conversion of O2 to O2•−. For example, in soybean, in addition to NADPH oxidase, quinone reductase also functions in the presence of menadione, facilitating the conversion of O2 to O2•− [77,100,102].
In the endoplasmic reticulum (ER), both O2•−and H2O2 are produced by fatty acid oxidation by GOX and urate oxidase activities [103]. In addition, a small amount of O2•−is produced in the ER as a by-product of oxidation and hydroxylation processes involving cytochrome P450 and cytochrome P540 reductase in the presence of reduced NADPH [38]. Compared with other cellular compartments, the rate of ROS production is comparatively lower in the cytosol, where the redox balance is largely maintained by cytoplasmic NADPH as a central component. However, in addition to ROS production, the cytosol plays a key role in the redox signaling process in plant cells. In general, ROS signaling from various cellular organelles passes through the cytoplasm to modulate gene expression in the cell nucleus [104].
5. ROS under Abiotic Stress
Abiotic stresses are closely related to climate change and hinder plant growth and development; hence, it also negatively affects crop yield and quality [13,105]. Environmental impacts on plants can be both long-term and short-term and vary in strength. Depending on the set of mechanisms launched by plants in response to stress, which can vary significantly for each plant species and even for different varieties of the same species, their tolerance to stress and rapid acclimatization can be determined [106]. During abiotic stress, ROS production is impaired, resulting in disruption of ROS metabolic and signaling pathways. Metabolic ROS can directly alter the redox status of enzymes involved in the metabolism process, which may lead to changes in the metabolic rate. Alteration of metabolic reactions in the cell may negatively affect defense mechanisms to abiotic stresses (Figure 7) [107].
The steady-state ROS level and redox state of the plant cell differ depending on the type of abiotic stress the plant faces. It follows that different environmental conditions lead to the production of a specific set of subcellular ROS and redox signature, which in turn leads to the activation of an acclimation response adapted to them.
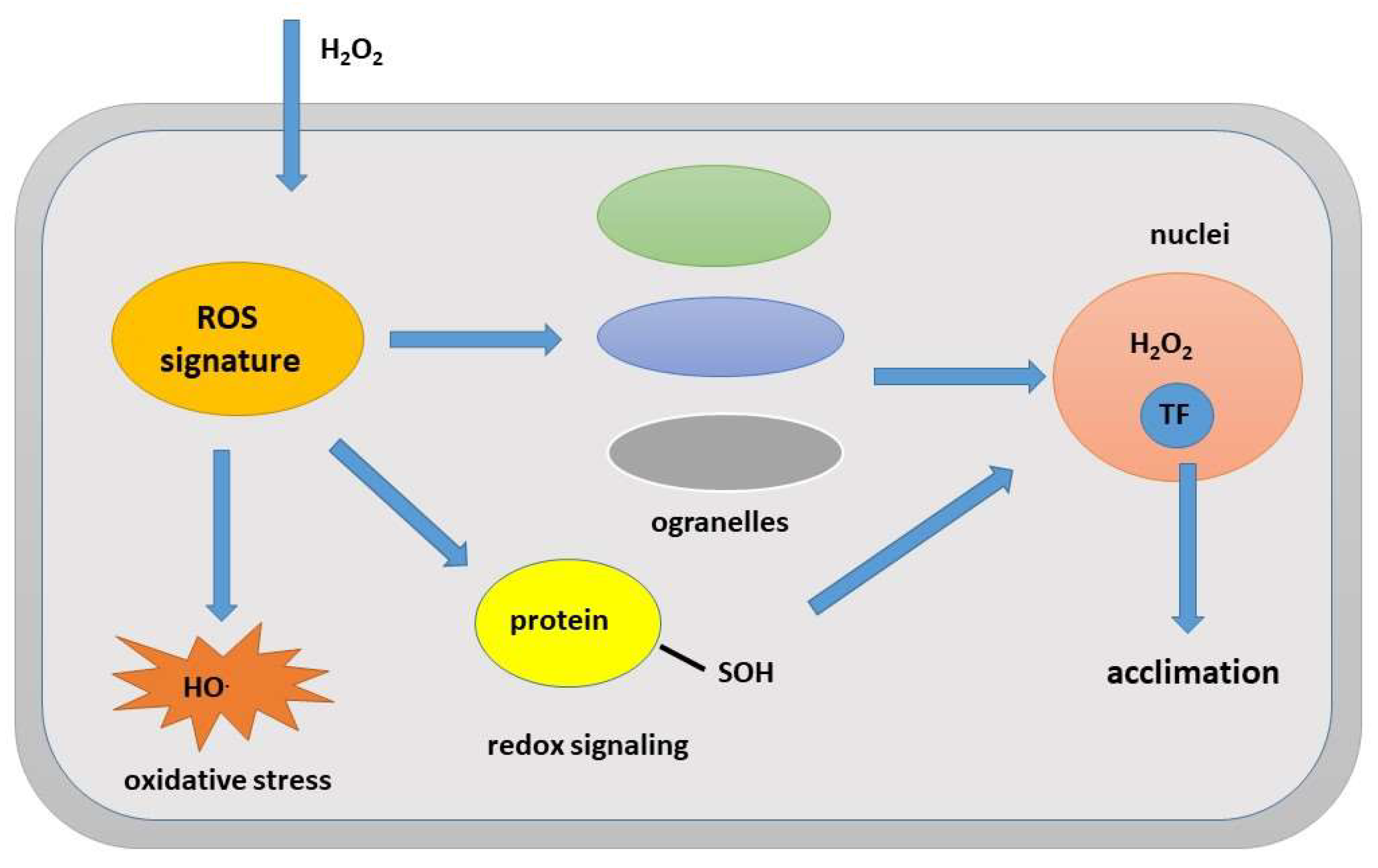
Excess ROS and ROS modification lead to changes in the redox balance in the cell, which may alter the function of key regulatory proteins regulating transcription and/or translation [3,32]. In contrast, signaling ROS directly alter the redox state of regulatory proteins and alter transcription and translation, resulting in activation of acclimation responses to abiotic stresses, counteracting their negative impact on metabolism and reducing the level of metabolic ROS. Signaling ROS are produced in the apoplast [108]. These signaling ROS are then translocated into the cytoplasm via regulated aquaporins [109] where they alter the redox status of key regulatory proteins such as TFs that influence gene expression (Figure 8)[34,79,110].
As signaling molecules, ROS are distributed in all metabolically active plant tissues and are controlled by the ROS gene network [111,112,113]. ROS, together with Ca2+ ions, participate in long-range systemic signaling, participating in the activation of acclimation to abiotic stresses [52,108]. Plant hormones such as abscisic acid (ABA) and jasmonic acid (JA) trigger ROS production, initiating a systemic signal (ROS wave) [114]. Hormonally activated ROS move in a loop and activate acclimation mechanisms [115,116]. This mechanism is realized in a feedback loop that activates ROS and calcium, thus causing a response of the whole plant to acclimate to abiotic stresses .
Under stress conditions, compartments such as the chloroplast, peroxisomes, and mitochondria have been shown to exhibit expansion of membrane structures that contact the nuclear envelope and can directly alter the ROS state of nuclei [32]. Metabolic and signaling ROS can be produced in different subcellular compartments (e.g., metabolic ROS in the chloroplast and signaling ROS in the apoplast). However, they can influence each other’s levels and even shuttle between compartments [109]. Although ROS production is a common phenomenon and part of plant cellular metabolism, environmental stresses lead to excessive ROS formation, which are not only highly reactive but also toxic in nature and damage lipids, proteins, carbohydrates, and DNA [2,15]. Higher concentrations of ROS have damaging effects on cellular organelles and tissues of shoots and roots. However, more and more studies have shown that ROS play a dual role in plants [1,6]. An important positive role of ROS is to scavenge excess electrons in the chloroplast from the photosynthetic apparatus, thereby preventing antenna overload and subsequent damage. A similar scavenging function is also performed by ROS in the mitochondria. In plant cells, there are several levels of ROS detoxification pathways and mechanisms, which allows ROS to redirect electron flow and prevent overload of various cellular systems during stress [49]. ROS, as well as their ability to convert into other forms of ROS, may mediate the regulation of metabolic fluxes during stress, preventing damage or excessive accumulation of toxic products. The most beneficial role of ROS during abiotic stress is shown in signaling processes that activate acclimation processes (Figure 8) [3,34,51,53,117]. Plants with impaired ROS production or ROS scavenging were found to be more sensitive to abiotic stresses and also failed to mediate systemic signaling during abiotic stress [50,118,119]. However, these useful functions of ROS can be demonstrated while the cell detoxifies excess ROS. In case of disruption of the detoxification system, as well as in case of significant and prolonged abiotic stress, excess ROS is extremely toxic for the cell, leading to the destruction of biomolecules and cell death. Thus, ROS molecules are important molecules regulating metabolism and triggering signals for acclimation processes in response to abiotic stress.
6. Conclusion
Abiotic stresses, both climate change and intense, impede plant growth and negatively affect crop yield and quality. To cope with the constant environmental impact, plants have developed a complex acclimation system. The acclimation system is triggered by a cascade of signaling reactions. ROS formation in plants is a natural metabolic process. ROS play both positive and negative roles. At low concentrations, ROS are signaling molecules that activate proliferation-differentiation processes. During abiotic stress, ROS production is impaired, which leads to disruption of ROS metabolic and signaling pathways. The steady-state level of ROS and the redox state of the plant cell differ depending on the type of abiotic stress the plant faces. It follows that different environmental conditions lead to the formation of a specific set of subcellular ROS and redox signature, which in turn leads to the activation of an acclimation response adapted to them. Thus, ROS is an essential molecule. signaling about abiotic stress and triggering the acclimation process. At the site of exposure to the stressor, ROS are produced, which trigger a wave of ROS that spreads at a tremendous speed throughout the plant. The ROS wave is used for signaling and triggering the mechanisms of plant adaptation to abiotic stress. As a result of ROS wave signaling, transcriptomic and metabolomic reprogramming of cells occurs.
However, the processes of ROS interaction with other molecules, signal transduction mechanisms, detection of signaling cascades, participation of ROS in metabolic processes, regulation of genes involved in the processes of plant acclimation to various types of abiotic stress are complex and require further research.
Funding
The reported study was supported by FGUM-2022-0003 of the Ministry of Science and Higher Education of the Russian Federation.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mittler, R.: Vanderauwera, S.: Suzuki, N.: Miller, G.: Tognett,i V.B.: Vandepoele, K.: Gollery, M.: Shulaev, V.: Van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011. 16, 300–309. [CrossRef]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, Oxidative Damage and Oxygen Deprivation Stress. Ann Bot. 2003, 91, 179–194. [CrossRef]
- Foyer C.H., Noctor G. Redox signaling in plants. Antioxid. Redox Signal. 2013,18, 2087–2090. [CrossRef]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [CrossRef]
- Zeng, J.; Dong, Z.; Wu, H.; Tian, Z.; Zhao, Z. Redox regulation of plant stem cell fate. EMBO J. 2017, 36, 2844-2855. [CrossRef]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [CrossRef]
- Dangl, J.L; Jones, J.D. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826-833. [CrossRef]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [CrossRef]
- Chan, K.X.;, Mabbitt, P.D.; Phua, S.Y.; Mueller, J.W.; Nisar, N,; Gigolashvili, T.; Stroeher, E.; Grassl, J.; Arlt, W.; Estavillo, G.M.; Jackson, C.J.; Pogson, B.J. Sensing and signaling of oxidative stress in chloroplasts by inactivation of the SAL1 phosphoadenosine phosphatase. Proc Natl Acad Sci U S A 2016, 113, E4567-4576. [CrossRef]
- Mailloux, J.R. Application of mitochondria-targeted pharmaceuticals for the treatment of heart disease. Curr. Pharm. Des. 2016, 22, 4763–4779. [CrossRef]
- Elstner, E.F. Metabolism of activated oxygen species. In: Davies DD, ed. Biochemistry of plants, Vol. 11. London: Academic Press, 1987, 253–315. [CrossRef]
- Droge, W. Free radicals in the physiological control of cell function. Physiol Rev 2002, 82, 47–95. [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Parvin, K.; Bhuiyan, T.F.; Anee, T.I.; Nahar, K.; Hossen, M.; Zulfiqar, F.; Alam, M.; Fujita, M. Regulation of ROS metabolism in plants under environmental stress: A review of recent experimental evidence. Int. J. Mol. Sci. 2020, 21, 8695. [CrossRef]
- Bhattacharjee, S. . Reactive Oxygen Species in Plant Biology. Springer; New Delhi, India: ROS and oxidative stress: Origin and implication. 2019, 1–31. [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [CrossRef]
- Bolwell, G.P.; Wojtaszek, P. Mechanisms for generation of reactive oxygen species in plant defence--a broad perspective. Physiological and Molecular Plant Pathology 1997, 51, 347–366. [CrossRef]
- Pfister-Sieber, M.; Braendle, R. Aspects of plant behavior under anoxia and post-anoxia. Proceedings of the Royal Society of Edinburgh 1994, 102B, 313–324. [CrossRef]
- Lamb, C.; Dixon, R.A. The oxidative burst in plant disease resistance. Annual Review of Plant Physiology and Plant Molecular Biology 1997, 48, 251–275. [CrossRef]
- Dorion, S.; Ouellet, J.C.; Rivoal, J. Glutathione metabolism in plants under stress: beyond reactive oxygen species detoxification. Metabolites 2021, 11, 641. [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem 2010, 48, 909–930. [CrossRef]
- Kandhari, P. Generic differences in antioxidant concentration in the fruit tissues of four major cultivars of apples. Master Thesis, University of Maryland, College Park 2004.
- Ayala, A.; Muñoz, M.F.; Argüelles, S Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid Med Cell Longev. 2012, 2014, 360438. [CrossRef]
- Gupta, D.K.; Palma, J.M.; Corpas, F.J. Reactive Oxygen Species and Oxidative Damage in Plants under Stress; Springer: New York, NY, USA 2015. [CrossRef]
- Joshi, R.; Wani, S.H.; Singh, B.; Bohra, A.; Dar, Z.A.; Lone, A.A.; Pareek, A.; Singla-Pareek, S.L. Transcription factors and plants response to drought stress: Current understanding and future directions. Front. Plant Sci. 2016, 7, 1029. [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Selenium pretreatment upregulates the antioxidant defense and methylglyoxal detoxification system and confers enhanced tolerance to drought stress in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 143, 1758–1776. [CrossRef]
- Decros, G.; Baldet, P.; Beauvoit, B.; Stevens, R.; Flandin, A.; Colombié, S.; et al. Get the balance right: ROS homeostasis and redox signalling in fruit. Front. Plant Sci. 2019, 10, 1091. [CrossRef]
- Naviaux, R.K. Oxidative shielding or oxidative stress? J Pharmacol Exp Ther 2012, 342, 608–618. [CrossRef]
- Paciolla, C.; Paradiso, A.; de Pinto, M. “Cellular redox homeostasis as central modulator in plant stress response,” in Redox State as a Central Regulator of Plant-Cell Stress Responses, eds Gupta D., Palma J., Corpas F. (Berlin: Springer;) 2016, 1–23. [CrossRef]
- Bae, Y.S.; Oh, H.; Rhee, S.G.; Yoo, Y.D. Regulation of reactive oxygen species generation in cell signaling. Mol Cells 2011, 32, 491–509. [CrossRef]
- Hernandez-Garcia, D.; Wood, C.D.; Castro-Obregon, S.; Covarrubias, L. Reactive oxygen species: a radical role in development? Free Radic Biol Med. 2010, 49, 130–143. [CrossRef]
- Foyer, C.H.; Noctor, G. Stress-triggered redox signalling: what’s in pROSpect? Plant, Cell Environ. 2016, 39, 951–964. [CrossRef]
- Dietz, K.J. Efficient high light acclimation involves rapid processes at multiple mechanistic levels. J. Exp. Bot. 2015, 66, 2401–2414. [CrossRef]
- Dietz, K.J. Thiol-based peroxidases and ascorbate peroxidases: why plants rely on multiple peroxidase systems in the photosynthesizing chloroplast? Mol. Cells 2016, 39, 20–25. [CrossRef]
- Noctor, G.; Foyer, C.H. Intracellular redox compartmentation and ROS-related communication in regulation and signaling. Plant Physiol. 2016, 171, 1581–1592. [CrossRef]
- Kohli, S.K.; Khanna, K.; Bhardwaj, R.; Abd Allah, E.F.; Ahmad, P.; Corpas, F.J. Assessment of subcellular ROS and NO metabolism in higher plants: Multifunctional signaling molecules. Antioxidants. 2019, 8, 641. [CrossRef]
- Podgorska, A.; Burian, M.; Szal, B. Extra-cellular but extra-ordinarily important for cells: Apoplastic reactive oxygen species metabolism. Front. Plant Sci. 2017, 8, 1353. [CrossRef]
- Janků, M.; Luhová, L.; Petřivalský, M. On the origin and fate of reactive oxygen species in plant cell compartments. Antioxidants. 2019, 8, 105. [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [CrossRef]
- Kumar, V.; Irfan, M.; Ghosh, S.; Chakraborty, N.; Chakraborty, S.; Datta, A. Fruit ripening mutants reveal cell metabolism and redox state during ripening. Protoplasma 2016, 253, 581–594. [CrossRef]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Breusegem, F.V. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663-679. [CrossRef]
- Gilbert, H.F. Molecular and cellular aspects of thiol-disulfide exchange. Adv Enzymol Relat Areas Mol Biol 1990, 63, 69–172. [CrossRef]
- Rajasekaran, N.S.; Connell, P.; Christians, E.S.; Yan, L.J.; Taylor, R.P.; Orosz, A.; Zhang, X.Q.; Stevenson, T.J.; Peshock, R.M.; Leopold, J.A.; Barry, W.H.; Loscalzo, J.; Odelberg, S.J.; Benjamin, I.J. Human alpha B-crystallin mutation causes oxido-reductive stress and protein aggregation cardiomyopathy in mice. Cell 2007, 130, 427–439. [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [CrossRef]
- Panieri, E.; Santoro, M.M. ROS signaling and redox biology in endothelial cells. Cell Mol. Life Sci. 2015, 72, 3281–3303. [CrossRef]
- Hansen, J.M.; Go, Y.M.; Jones, D.P. Nuclear and mitochondrial compartmentation of oxidative stress and redox signaling. Annu Rev Pharmacol Toxicol 2006, 46, 215–234. [CrossRef]
- Leister, D. Piecing the puzzle together: the central role of reactive oxygen species and redox hubs in chloroplast retrograde signaling. Antioxid. Redox Signal 2019, 30, 1206–1219. [CrossRef]
- Chan, Z.; Yokawa, K.; Kim, W.Y.; Song, C.P. ROS regulation during plant abiotic stress responses. Front. Plant Sci. 2016, 7, 1536. [CrossRef]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141,391–396. [CrossRef]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory burst oxidases: the engines of ROS signaling. Curr. Opin. Plant Biol. 2011, 14, 691–699. [CrossRef]
- Vaahtera, L.; Brosche, M.; Wrzaczek, M.; Kangasjarvi, J. Specificity in ROS signaling and transcript signatures. Antioxid. Redox Signal. 2014, 21, 1422–1441. [CrossRef]
- Gilroy, S.; Białasek, M.; Suzuki, N.; Górecka, M.; Devireddy, A.R.l Karpiński, S.; Mittler, R. ROS, Calcium, and Electric Signals: Key Mediators of Rapid Systemic Signaling in Plants. Plant Physiol. 2016, 171, 1606–1615. [CrossRef]
- Mignolet-Spruyt, L., Xu, E., Idanheimo, N., Hoeberichts, F.A., Muhlenbock, P., Brosche, M., Van Breusegem, F. and Kangasjarvi, J. Spreading the news: subcellular and organellar reactive oxygen species production and signalling. J. Exp. Bot. 2016, 67, 3831–3844. [CrossRef]
- Raja,V.; Majeed, U.; Kang, H.; Andrabi, K.I.; John, R. Abiotic stress: Interplay between ROS, hormones and MAPKs. Environ. Exp. Bot. 2017, 137, 142–157. [CrossRef]
- Shakirova, F.M.; Allagulova, C.R.; Maslennikova, D.R.; Klyuchnikova, E.O.; Avalbaev, A.M.; Bezrukova, M.V. Salicylic acid-induced protection against cadmium toxicity in wheat plants. Environ. Exp. Bot. 2016, 122, 19–28. [CrossRef]
- Singh, A.; Kumar, A.; Yadav, S.; Singh, I K. Reactive oxygen species-mediated signaling during abiotic stress. Plant Gene 2019, 18, 100–173. [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [CrossRef]
- Triantaphylides, C.; Havaux, M. Singlet oxygen in plants: production, detoxification and signaling. Trends Plant Sci. 2009, 14, 219–228. [CrossRef]
- Pospisil, P.; Arato, A.; Krieger-Liszkay, A.; Rutherford, A.W. Hydroxyl radical generation by photosystem II. Biochemistry 2004, 43, 6783–6792. [CrossRef]
- Pospisil, P. Production of reactive oxygen species by photosystem II. Biochim. Biophys. Acta. 2009, 1787. 1151–1160. [CrossRef]
- Pospisil, P. The role of metals in production and scavenging of reactive oxygen species in photosystem II. Plant Cell Physiol. 2014, 55, 1224–1232. [CrossRef]
- Pospisil, P. Production of reactive oxygen species by photosystem II as a response to light and temperature stress. Front Plant Sci. 2016, 7, 1950. [CrossRef]
- Pospisil, P.; Yamamoto, Y. Damage to photosystem II by lipid peroxidation products. BBA Gen. Subj. 2017, 1861, 457–466. [CrossRef]
- Shumbe, L.; Chevalier, A.; Legeret, B.; Taconnat, L.; Monnet, F.; Havaux, M. Singlet oxygen-induced cell death in Arabidopsis under high-light stress is controlled by OXI1 kinase. Plant Physiol. 2016,. 170, 1757–1771. [CrossRef]
- Havaux, M.; Strasser, R.J.; Greppin, H. A theoretical and experimental analysis of the qP and q N coefficients of chlorophyll fluorescence quenching and their relation to photochemical and nonphotochemical events. Photosynth Res. 1991, 27, 41–55. [CrossRef]
- Susek, R.E.; Ausubel, F.M.; Chory, J. Signal transduction mutants of Arabidopsis uncouple nuclear CAB and RBCS gene expression from chloroplast development. Cell. 1993, 74, 787–799. [CrossRef]
- Asada, K. The water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Annual Review of Plant Physiology and Plant Molecular Biology 1999, 50, 601–639. [CrossRef]
- Rhoads, D.M.; Umbach, A.L.; Subbaiah, C.C.; Siedow, J.N. Mitochondrial reactive oxygen species. Contribution to oxidative stress and interorganellar signaling. Plant Physiol. 2006, 141, 357–366. [CrossRef]
- Mattos, L.; Moretti, C. Oxidative stress in plants under drought conditions and the role of different enzymes. Enzym. Eng. 2015, 5, 1–6.
- Rasmusson, A.G.; Geisler, D.A.; Møller, I.M. The multiplicity of dehydrogenases in the electron transport chain of plant mitochondria. Mitochondrion 2008, 8, 47–60. [CrossRef]
- Braidot, E.; Petrussa, E.; Vianello, A.; Macri, F. Hydrogen peroxide generation by higher plant mitochondria oxidizing complex I of complex II substrates. FEBS Letters 1999, 451, 347–350. [CrossRef]
- Gille, L.; Nohl, H. The ubiquinol/bc1 redox couple regulates mitochondrial oxygen radical formation. Archives of Biochemistry and Biophysics 2001, 388, 34–38. [CrossRef]
- Möller, I.M. Plant mitochondria and oxidative stress: electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annual Review of Plant Physiology and Plant Molecular Biology 2001, 52, 561–91. [CrossRef]
- Blokhina, O.; Fagerstedt, K.V. Reactive oxygen species and nitric oxide in plant mitochondria: Origin and redundant regulatory systems. Physiol. Plant. 2010, 138, 447–462. [CrossRef]
- Giraud, E.; Ho, L.H.; Clifton, R.; Carroll, A.; Estavillo, G.; Tan, Y.F.; Howell, K.A.; Ivanova, A.; Pogson, B.J.; Millar, A.H. The absence of ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress. Plant Physiol. 2008, 147, 595–610. [CrossRef]
- Scialò, F.; Fernández-Ayala, D.J.; Sanz, A. Role of mitochondrial reverse electron transport in ROS signaling: Potential roles in health and disease. Front. Physiol. 2017, 8, 428. [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [CrossRef]
- Kerchev, P.; Waszczak, C.; Lewandowska, A.; Willems, P.; Shapiguzov, A.; Li, Z. Lack of GLYCOLATE OXIDASE1, but not GLYCOLATE OXIDASE2, attenuates the photorespiratory phenotype of CATALASE2- deficient Arabidopsis. Plant Physiol. 2016, 171, 1704–1719. [CrossRef]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Signal. 2009, 11, 861–905. [CrossRef]
- Corpas, F.J.; del Río, L.A.; Palma, J.M. Impact of nitric oxide (NO) on the ROS metabolism of peroxisomes. Plants 2019, 8,37. [CrossRef]
- Reumann, S.; Chowdhary, G.; Lingner, T. Characterization, prediction and evolution of plant peroxisomal targeting signals type 1 (PTS1s). Biochim. Biophys. Acta 2016, 1863, 790–803. [CrossRef]
- Palma, J.M., Corpas, F.J., del Río, L.A. Proteome of plant peroxisomes: New perspectives on the role of these organelles in cell biology. Proteomics 2009, 9, 2301–2312. [CrossRef]
- Kostić, A.; Dimitrijević, D.S.; Stojanović, G.S.; Palić, I.R.; Đorđević, A.S.; Ickovski, J.D. Xanthine oxidase: Isolation, assays of activity, and inhibition. J. Chem. 2015, 2015, 1–8. [CrossRef]
- Schrader, M.; Fahimi, H.D. Peroxisomes and oxidative stress. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2006, 1763, 1755–1766. [CrossRef]
- Corpas, F.J.; Barroso, J.B.; Palma, J.M.; Rodriguez-Ruiz, M. Plant peroxisomes: A nitro-oxidative cocktail. Redox Biol. 2017, 11, 535–542. [CrossRef]
- Jimenez, A.; Hernandez, J.A.; del Río, L.A.; Sevilla, F. Evidence for the presence of the ascorbate-glutathione cycle in mitochondria and peroxisomes of pea leaves. Plant Physiol. 1997, 114, 275–284. [CrossRef]
- Vandenabeele, S.; Vanderauwera, S.; Vuylsteke, M.; Rombauts, S.; Langebartels, C.; Seidlitz, H.K.; Zabeau, M.; Van Montagu, M.; Inzé, D.; Van Breusegem, F. Catalase deficiency drastically affects gene expression induced by high light in Arabidopsis thaliana. Plant J. 2004, 39, 45–58. [CrossRef]
- Gupta, K.; Sengupta, A.; Chakraborty, M.; Gupta, B. Hydrogen peroxide and polyamines act as double edged swords in plant abiotic stress responses. Front. Plant Sci. 2016, 7, 1343. [CrossRef]
- Rajab, H.; Khan, M S.; Malagoli, M.; Hell, R.; Wirtz, M. Sulfate-induced stomata closure requires the canonical ABA signal transduction machinery. Plants 2019, 8, 21. [CrossRef]
- Máthé, C.; Garda, T.; Freytag, C. The Role of Serine-threonine protein phosphatase PP2A in plant oxidative stress signaling—Facts and hypotheses. Int. J. Mol. Sci. 2019, 20, 3028. [CrossRef]
- Kwak, J.M.; Mori, I.C.; Pei, Z.M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones, J.D.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. [CrossRef]
- Torres, M.A.; Dangl, J.L. Functions of the respiratory burst oxidase in biotic interactions, abiotic stress and development. Curr. Opin. Plant Biol. 2005, 8, 397–403. [CrossRef]
- Li, Y.; Xiang, J.; Wang, Y.; Zheng, L.; Fan, Y.; Li, Y.; Zhao, F. Analysis of antioxidant characteristics and related gene expression profiles of rice drought-tolerance lines derived from embryo-soaking with alternanthera philoxeroides DNA solution. J. Bot. Sci. 2015, 4, 30–36.
- Rietz, S.; Bernsdorff, F.E.; Cai, D. Members of the germin-like protein family in Brassica napus are candidates for the initiation of an oxidative burst that impedes pathogenesis of Sclerotinia sclerotiorum. J. Exp. Bot. 2012, 63, 5507–5519. [CrossRef]
- Heyno, E.; Mary, V.; Schopfer, P.; Krieger-Liszkay, A. Oxygen activation at the plasma membrane: Relation between superoxide and hydroxyl radical production by isolated membranes. Planta 2011, 234, 35–45. [CrossRef]
- Bolwell, G.P.; Wojtaszek, P. Mechanisms for generation of reactive oxygen species in plant defence—a broad perspective. Physiological and Molecular Plant Pathology 1997, 51, 347–366. [CrossRef]
- Reggiani, R.; Bertani, A. Effect of decreasing oxygen concentration on polyamine metabolism in rice and wheat shoots. Journal of Plant Physiology 1989, 135, 375–377. [CrossRef]
- Sewelam, N.; Kazan, K.; Schenk, P.M. Global plant stress signaling: Reactive oxygen species at the cross-road. Front. Plant Sci. 2016, 7, 187. [CrossRef]
- Sharma, V.; Anderson, D.; Dhawan, A. Zinc oxide nanoparticles induce oxidative DNA damage and ROS-triggered mitochondria mediated apoptosis in human liver cells (HepG2). Apoptosis 2012, 17, 852–870. [CrossRef]
- Liu, J.H.; Wang, W.; Wu, H.; Gong, X.; Moriguchi, T. Polyamines function in stress tolerance: From synthesis to regulation. Front. Plant Sci. 2015, 6, 827. [CrossRef]
- Cuypers, A.; Hendrix, S.; Amaral dos Reis, R.; De Smet, S.; Deckers, J.; Gielen, H.; Jozefczak, M.; Loix, C.; Vercampt, H.; Vangronsveld, J. Hydrogen peroxide, signaling in disguise during metal phytotoxicity. Front. Plant Sci. 2016, 7, 470. [CrossRef]
- Kumar, S.P.J.; Prasad, R.S., Banerjee, R.; Thammineni, C. Seed birth to death: Dual functions of reactive oxygen species in seed physiology. Ann. Bot. 2015, 116, 663–668. [CrossRef]
- Van Breusegem, F.; Bailey-Serres, J.; Mittler, R. Unraveling the tapestry of networks involving reactive oxygen species in plants. Plant Physiol. 2008, 147, 978–984. [CrossRef]
- Yadav, S.; Modi, P.; Dave, A.; Vijapura, A.; Patel, D.; Patel, M. Effect of abiotic stress on crops. In: Hasanuzzaman M., Filho M.C.M.T., Fujita M., Nogueira T.A.R., editors. Sustainable Crop Production. IntechOpen; London, UK 2020, 3–24. [CrossRef]
- Kononenko, N.V.; Lazareva, E.M.; Fedoreyeva L.I.. Mechanisms of Antioxidant Resistance in Different Wheat Genotypes under Salt Stress and Hypoxia. Int J Mol Sci. 2023, 24,16878. [CrossRef]
- .Miller, G.A.D.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R.O.N. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [CrossRef]
- Gilroy, S.; Suzuki, N.; Miller, G.; Choi, W.G.; Toyota, M.; Devireddy, A.R.; Mittler, R. A tidal wave of signals: Calcium and ROS at the forefront of rapid systemic signaling. Trends Plant Sci. 2014, 19, 623–630. [CrossRef]
- Tian, S.; Wang, X.; Li, P.; Wang, H.; Ji, H.; Xie, J.; Qiu, Q.; Shen, D.; Dong, H. Plant aquaporin AtPIP1;4 links apoplastic H2O2 induction to disease immunity pathways. Plant Physiol. 2016, 171, 1635–1650. [CrossRef]
- Huang, S.; VanAken, O.; Schwarzländer, M.; Belt, K.; Millar, A.H. The roles of mitochondrial reactive oxygen species in cellular signaling and stress responses in plants. Plant Physiol 2016, 171, 1551–1559. [CrossRef]
- Mittler, R.; Finka, A.; Goloubinoff, P. How do plants feel the heat? Trends Biochem. Sci. 2012, 37, 118–125. [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [CrossRef]
- Fichman, Y.; Mittler, R. Rapid systemic signaling during abiotic and biotic stresses: Is the ROS wave master of all trades? Plant J. 2020, 102, 887–896. [CrossRef]
- Suzuki, N.; Miller, G.; Salazar, C.; Mondal, H.A.; Shulaev, E.; Cortes, D.F.; Shuman, J.L.; Luo, X.; Shah, J.; Schlauch, K.; Shulaev, V.; Muttler, R. Temporal-Spatial Interaction between Reactive Oxygen Species and Abscisic Acid Regulates Rapid Systemic Acclimation in Plants. Plant Cell. 2013, 25, 3553–3569. [CrossRef]
- Devireddy, A.R.; Zandalinas, S.I.; Fichman, Y.; Mittler, R. Integration of reactive oxygen species and hormone signaling during abiotic stress. Plant J. 2021, 105, 459–476. [CrossRef]
- Zandalinas, S.I.; Fichman, Y.; Devireddy, A.R.; Sengupta, S.; Azad, R.K.; Mittler, R. Systemic signaling during abiotic stress combination in plants. Proc. Natl. Acad. Sci. USA. 2020, 117, 13810–13820. [CrossRef]
- Considine, M.; Sandalio, L.M.; Foyer, C.H. Unravelling how plants benefit from ROS and NO reactions, while resisting oxidative stress. Ann. Bot. 2015, 116, 469–473. [CrossRef]
- Davletova, S.; Rizhsky, L.; Liang, H.; Shengqiang, Z.; Oliver, D.J.; Coutu, J.; Shulaev, V.; Schlauch, K.; Mittler, R. Cytosolic Ascorbate Peroxidase 1 Is a Central Component of the Reactive Oxygen Gene Network of Arabidopsis. Plant Cell. 2005, 17, 268–281. [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [CrossRef]
Figure 1.
ROS species: free radicals and free molecules.
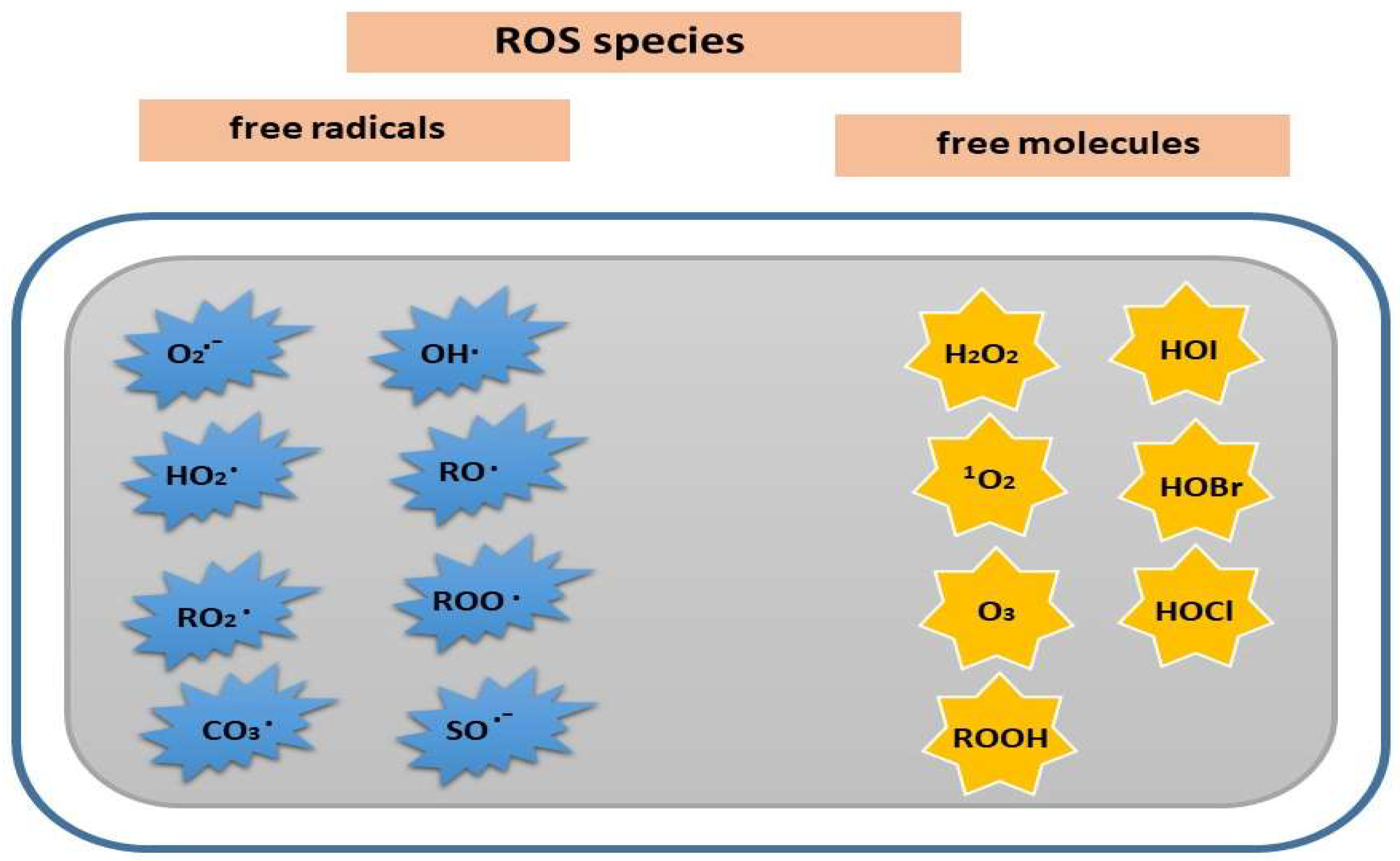
Figure 2.
ROS formation reactions.
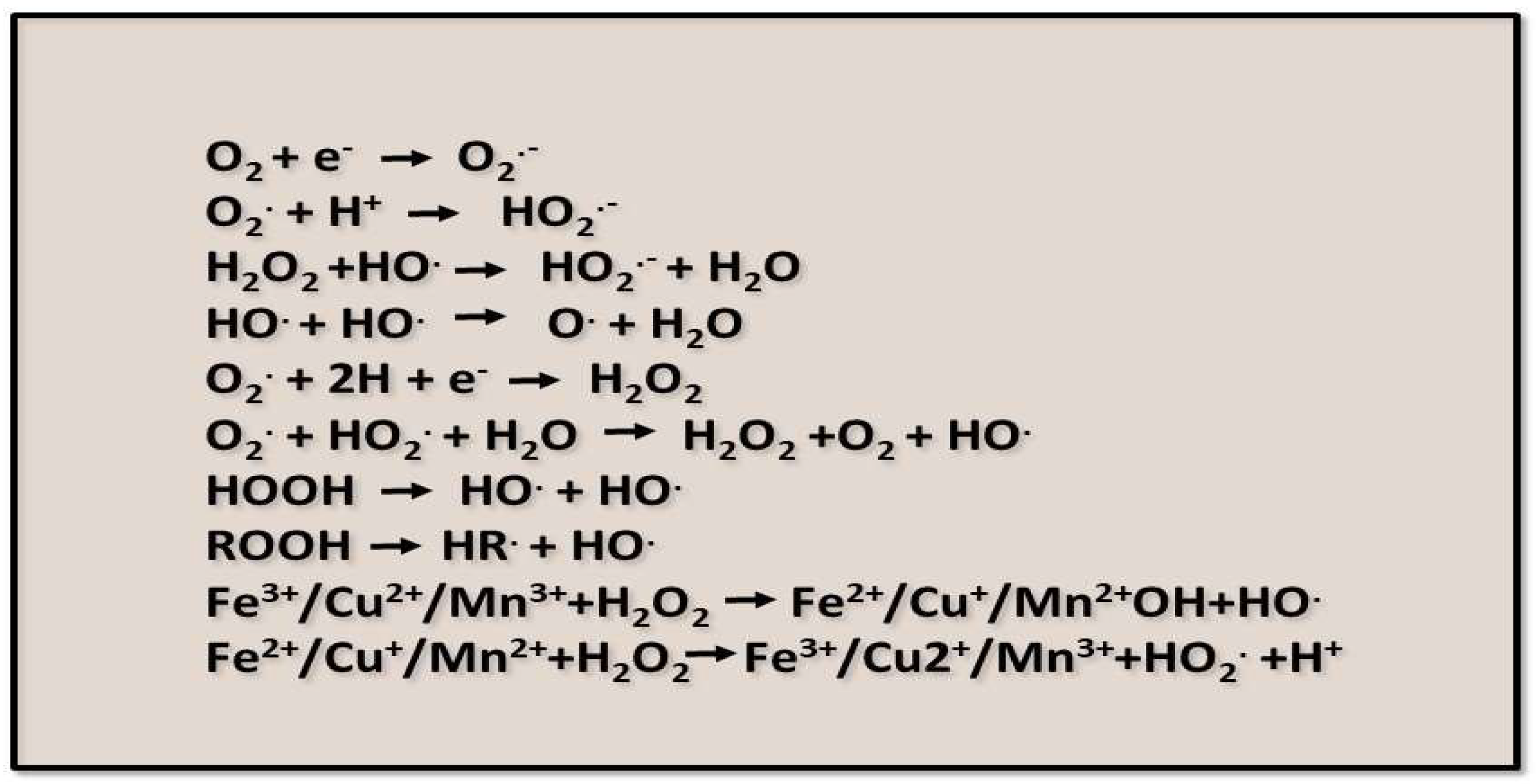
Figure 3.
Reactions of dismutation of O2 into H2O.
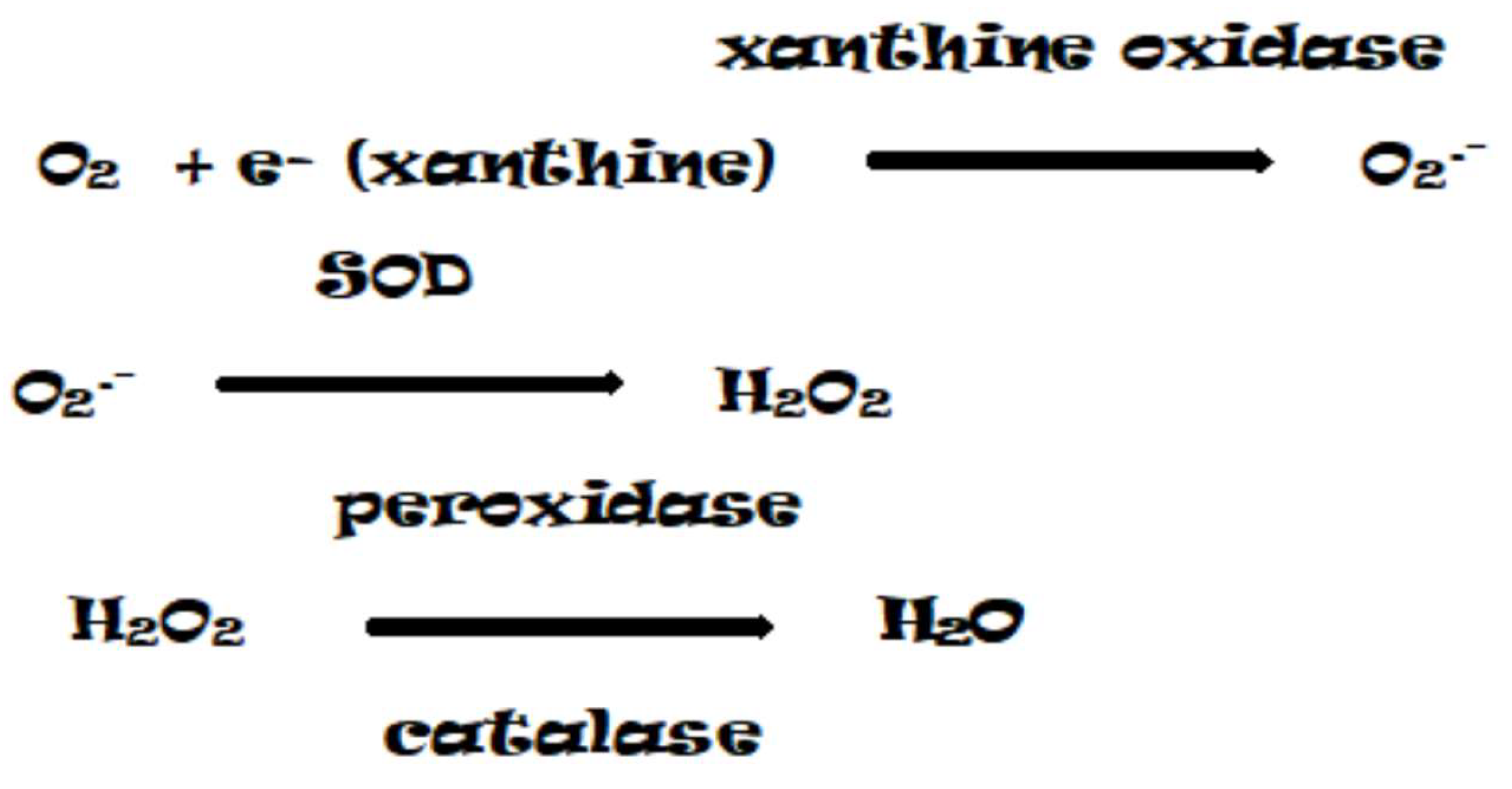
Figure 5.
Haber-Weiss and Fenton reactions.
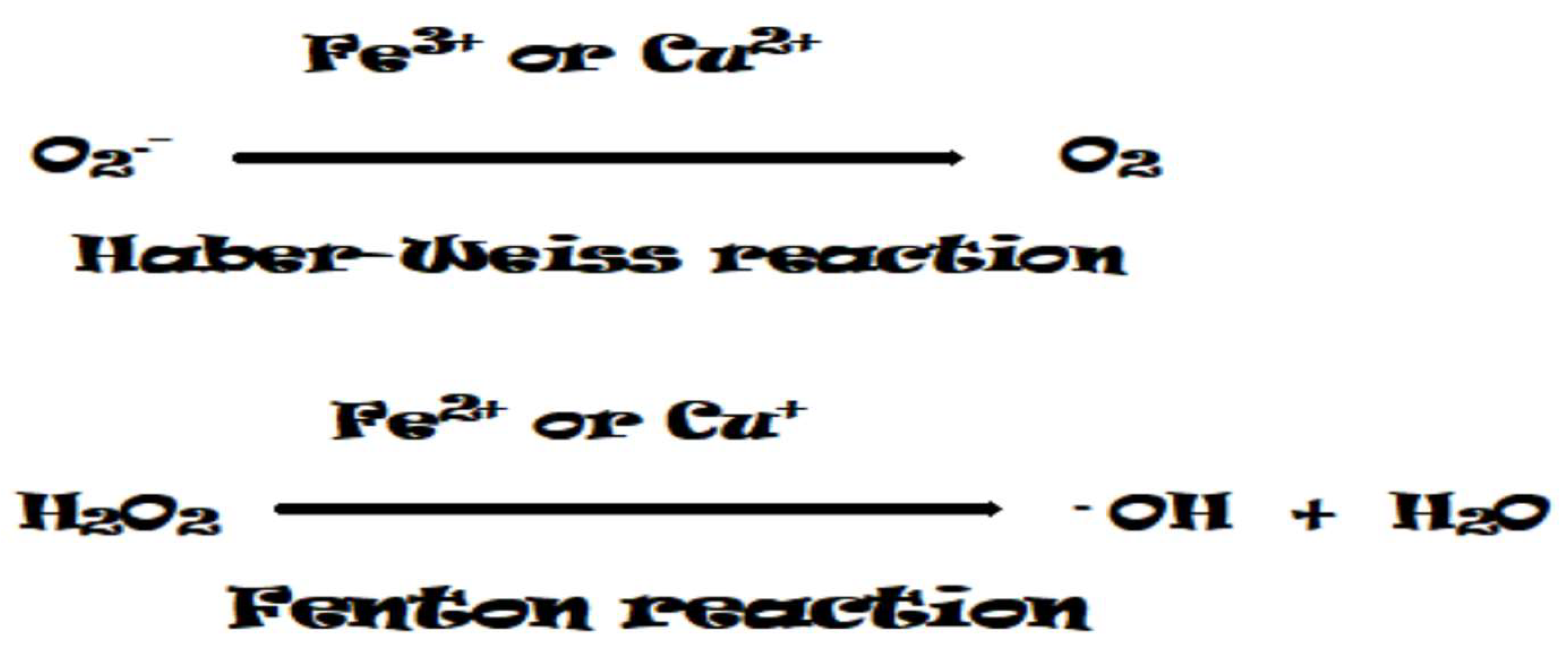
Figure 6.
Regulation of ROS in cellular compartments.
Figure 7.
Negative effects caused in plant cells under abiotic stress.
Figure 8.
Participation of ROS in the mechanisms of plant acclimatization to abiotic stress.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



