
Submitted:
08 October 2024
Posted:
09 October 2024
You are already at the latest version
Abstract
Plant‒fungal associations often drive plant invasion success by increasing host growth, disease resistance, and tolerance to environmental stress. In this study, to determine whether the numerically dominant foliar endophyte Colletotrichum can directly enhance the resistance of the invasive plant Ageratina adenophora to disease and environmental stressors, we planted sterile A. adenophora seedlings, inoculated them with three endophytic fungal strains of Colletotrichum with distinct genetic backgrounds and compared their role in the resistance of A. adenophora to the pathogenicity of three foliar pathogens from the Didymellaceae family, as well as to abiotic stress, including drought and nutrient stress. We observed that the foliar endophyte Colletotrichum reduced photosynthesis-related physiological indicators, increased resistance-related indicators, and decreased the biomass of A. adenophora in the absence of pathogen attack, drought and nutrient stress. However, some endophytic Colletotrichum strains exhibit positive effects on resistance to certain foliar pathogen attacks, and some endophytes increase the biomass of A. adenophora under drought or nutrient stress. Our results revealed that there are certain positive effects of foliar Colletotrichum endophytes on A. adenophora in response to biotic and abiotic stresses, which may be beneficial for its invasion.
Keywords:
Plant invasion
; Ageratina Adenophora
; Colletotrichum
; Plant pathogen
; Foliar endophytes
; Drought and nutrient stress
1. Introduction
Endophytes are a group of microorganisms (bacteria, actinobacteria, or fungi) that reside in the internal tissues of plants in symbiotic associations without causing any disease symptoms [1,2,3]. Among endophytes, endophytic fungi are among the important components of plant-microorganism interaction systems. Endophytic fungi include Chytridiomycota, Zygomycota, Ascomycota, and Basidiomycota, which occur in diverse environments [4]. On the basis of their lifestyle, functional diversity, biology and mode of transmission, endophytic fungi are classified as systemic/true endophytes (mutualistic for the entirety of their lifespan) or transient/nonsystemic endophytes (adapting transient modes of lifestyles, mutualistic and parasitic being the two most common modes, during different life cycle stages) [5]. The mutualistic relationships of these endophytic fungi with host plants are well known to have profound beneficial impacts on plant communities, such as improving plant fitness, increasing tolerance to biotic and abiotic stresses, and increasing plant biomass. In turn, the host plant provides fungal endophytes with refuge and nutrients and allows them to survive within the next generation of host plants.
The association of a plant with endophytic fungi is, therefore, a supreme beneficial relationship for both partners, especially the plant. Fungal endophytes can stimulate the growth of plants directly or indirectly by triggering the solubilization of phosphorus, potassium, and zinc [6,7]. Endophytic fungi also confer resistance against plant pathogens through enhanced production of plant secondary metabolites, endophyte-specific metabolite synthesis, induction of systemic or local immune responses or competitive elimination of pathogens [8]. Plants are subjected to various abiotic stresses, such as drought stress, temperature stress, salinity stress and heavy metal toxicity stress, during their life cycle [9]. In addition to their intrinsic mechanism of adaptation, endophytes also enhance the alleviation of the negative effects of stress conditions faced by host plants through two mechanisms: (a) activation of the stress response after host exposure and (b) synthesis of antistress biochemicals by endophytes [10].
Endophytic fungi have similar important ecological functions in invasive ecosystems, and the presence or absence of mutualist fungi may partially explain why some alien plants become invasive [11]. First, endophytic fungi can directly act on invasive plants to improve their growth, reproduction, defense against pathogens and herbivores and environmental tolerance. For example, Aschehoug, E. T.; Metlen, K. L.; Callaway, R. M. and Newcombe, G. [12] reported that inoculation with the endophyte Alternaria (CID120) could directly promote the growth of the invasive plant Centaurea stoebe. Endophytic fungi can improve the reproductive success of invasive plants by increasing germination [13] or facilitating the allocation of plant resources to adopt more efficient reproductive strategies [14]. For example, endophytic infection of Cyperus rotundus plants reduces investment in expensive sexual reproductive structures (inflorescence) but increases investment in the primary mode of reproduction (asexual tub production). This trade-off may increase population growth and thus establishment success [14]. The invasive annual Anthemis cotula leaves have endophytic microorganisms with plant growth-promoting traits, such as ammonia production, indole acetic acid (IAA) production, phosphate dissolution and biocontrol of active substances [15]. Currie et al. reported that foliar endophytic fungi (Colletotrichum acutatum, Alternaria alternata and Cladosporium oxysporum) can effectively reduce the pathogenicity of Puccinia komarovii, a rust pathogen, to the invasive plant Impatiens glandulifera [16]. The invasive grass Cirsium arvense was inoculated with the endophyte Chaetomium cochliodes to produce oxolipin and jasmonate metabolites, which induced the production of plant defense metabolites for systemic resistance [17]. Rudgers and Clay reported that Neotyphodium coenophialum significantly reduced the abundance and diversity of herbivorous insects in the invasive Lolium arundinaceum [18]. The survival rate of Phragmites australis seedlings under salt stress could be improved by inoculation with endophytes from the dark septum (DSE). When exposed to salt stress, P. australis can establish reciprocal relationships with DSEs [19].
Second, by interfering with native plants, endophytic fungi can also indirectly increase the competitive advantage of invasive plants; for example, they can change the soil nutrient content and soil through the cycle of endophytic fungal species to inhibit the growth of native plants and indirectly promote the development of their own species [20]. AMF can also reduce the dependence of host plants on symbiosis with AMF without changing their biomass and indirectly promote the establishment of host grass and subsequent invasion [21].
Ageratina adenophora (Sprengel) R.M. King & H. Robinson is a perennial herb of Asteraceae native to the Mexican area of America. This plant has invaded more than 40 tropical to temperate regions worldwide, causing severe ecological impacts and economic losses [20]. Since the invasion of Yunnan Province from the China–Myanmar border in the 1940s, A. adenophora has been widely distributed in Yunnan Province and neighboring Sichuan, Guizhou, Guangxi, Xizang and other provinces [22] and is one of the 18 most severely invasive plants in China [23]. The A. adenophora can accumulate beneficial microorganisms (such as nitrogen-fixing bacteria) around its roots to gain a sustained growth advantage [24,25]. Moreover, some promoting bacteria have a more significant growth-promoting effect on A. adenophora than on local plants do [26]. A. adenophora can also accumulate large amounts of native plant pathogenic fungi in its leaf tissue in the form of endophytic fungi, which may help its successful invasion [27]; these dominant genera of foliar endophytic fungi include Colletotrichum, Nemania, Phomopsis, and Xylaria [28].
Previously, some Colletotrichum endophytes were shown to have a weak negative impact on the growth of A. adenophora planted in agricultural soil and forest soil but can increase the pathogenicity of the foliar pathogen Diaporthe helianthi and slightly reduce the herbivory of A. adenophora seedlings in the wild [29]. Species of Colletotrichum are commonly found in many plant hosts as pathogens, endophytes and occasionally saprobes. It has been reported that Colletotrichum, an endophyte, exists in Dendrobium orchids [30], Taxus mairei [31], Citrus grandis cv. “Tomentosa” [32], tea plants [33], and wild banana plants (Musa acuminata) [34]. Colletotrichum plays both pathogenic and endophytic roles depending on the interaction between Colletotrichum and the specific host, as well as biotic and abiotic factors in the environment [35]. Because A. adenophora can be infected frequently by diverse foliar pathogens in its invaded range, particularly members of the family Didymellaceae [36], it is unclear whether the widely occurring foliar Colletotrichum endophytes also increase the pathogenicity of the dominant foliar pathogens from the family Didymellaceae. Moreover, although some Colletotrichum endophytes have a weak negative impact on the growth of A. adenophora [29], it is necessary to determine whether they can increase host plant resistance to environmental stressors such as drought stress or nutrient stress because A. adenophora occurs in the wild in harsh environments (roadsides and railway embankments), which are characterized by drought and low nutrient availability [20].
In this study, we planted sterile seedlings of A. adenophora, inoculated them with endophytic fungal strains of Colletotrichum with distinct genetic backgrounds and compared their role in the resistance of invasive A. adenophora to the pathogenicity of three foliar pathogen members from the Didymellaceae family, as well as to abiotic stress, including drought and nutrient stress. We expected that, compared with plants inoculated with A. adenophora without endophytes, those inoculated with Colletotrichum would (1) have greater biomass, (2) decrease the pathogenicity of the pathogen on their leaves, and (3) alleviate the effects of drought stress and nutrient stress on plant growth.
2. Materials and Methods
2.1. Description of Fungi
The three Colletotrichum strains AX39, AX198 and AX115 used in this study were previously isolated from healthy leaves of A. adenophora in Yunnan Province, China. The ITS sequences of the strains were searched for similar DNA sequences in GenBank via BLAST, and the results revealed that both AX39 (GenBank accession number: PP227195) and AX198 (GenBank accession number: PP227202) presented the highest ITS sequence similarity with the C. gloeosporioides complex, whereas AX115 (GenBank accession number: KC507204) presented the highest ITS sequence similarity with the C. acutatum complex. Three Didymellaceae strains, G56, Y122 and S188, used in this study were previously collected from the roots and healthy and diseased leaves of A. adenophora. Didymellaceae species are distributed throughout a broad range of environments, and most are economically important plant fungal pathogens with a wide host range [37]. Our previous study characterized their virulence and host range and reported that these three strains are A. adenophora host-specific pathogens with weak virulence to native plants but strong virulence to the host A. adenophora [36]. Among them, G56 and Y122 belong to Mesophoma ageratinae, and S188 belongs to Mesophoma speciosa [38].
2.2. Growth Effects of Colletotrichum on Its Host A. adenophora
2.2.1. Planting Host A. adenophora and Inoculation with Colletotrichum
The effects of three Colletotrichum strains (AX39, AX115 and AX198) on the growth of A. adenophora were first tested. In addition to the control treatment without inoculation of Colletotrichum strains, there were 4 groups in the experiment, and 5 POTS plants were planted in each group as replicates, for a total of 20 POTS plants. Seeds of A. adenophora collected from wild plants in Xishan Forest Park, Kunming, were immersed in 75% ethanol for 10 minutes and then in 3% sodium hypochlorite for 10 minutes and then washed three times with sterile water. The surface-disinfected seeds were soaked in sterile water for 12 hours, and 4 seeds were cultured in Murashige and Skoog (MS) medium tissue culture bottles with tweezers in an artificial climate chamber (RXZ-380D, Ningbo Southeast Instrument Co., Ltd., Ningbo, China) for one month (25 °C ± 1, relative humidity 80% ± 5, photoperiod 12 h, light intensity 12,000 lux) (Figure S1a).
For preparation of the spore suspension, Colletotrichum strains were cultured on PDA media (200 g potato, 20 g glucose, 20 g agar, 1000 ml water, natural pH). The purified fungus was inoculated into 50 ml of sterilized PDB liquid media (200 g potato, 20 g glucose, 1000 ml water, and natural pH) in 200 ml conical flasks and cultured at 28 °C with shaking for 3–5 days (Figure S1b). After massive sporulation, the mycelium was filtered and removed with 4 layers of sterilized lens paper (Li et al. 2023), and the spores were collected by centrifugation of the filtered spore mixture. A spore suspension was prepared by adding 0.5% sterilized gelatin solution, and the concentration was adjusted to ~106 CFU/ml for each strain. The spore suspension was stored at 4 °C until use.
For inoculation, the seedling leaves were sprayed on both sides with a 0.5% gelatin suspension with 5×106 CFU/ml spores of Colletotrichum strains (1 ml per bottle). The plants in the control group were sprayed with 0.5% sterilized gelatin solution. After 24 h of culture in the chamber, the same inoculation procedure was performed. Fungal infection of leaves was detected via the [39] method within three days after inoculation. Three leaves were picked from each plant (at the bottom, middle and top of the plant), and the middle part of the leaves (avoiding the vein part) was cut into 2 mm2 pieces. The surface was disinfected (soaked in 0.5% sodium hypochlorite solution for 2 min, soaked in 75% ethanol for 2 min, washed with sterile water three times, and dried with sterilized filter paper). Nine small pieces (2 mm2) were placed on 2% MEA media (0.6 g of malt extract, 0.06 g of soybean peptone, 20 g of agar powder, and 1000 ml of water) at room temperature. Three plates of medium were used for each treatment. After 3–5 days, the number of small leaves with fungal growth was recorded, and the fungi were cultured on PDA media supplemented with streptomycin sulfate and ampicillin to verify that they were originally inoculated strains (Figure S1c).
The inoculated plants were transplanted to polypropylene (PP) cups filled with 100 g of sterilized Danish humus soil (Pindstrup sphagnum, Denmark) and 100 ml of water and placed in a plant growth chamber to grow for 1 month (Figure S1d). Afterward, the plants were watered with 50 ml of sterile water every two days and 50 ml of Hoagland nutrient solution once a week. The PP cups were sealed with a PTFE membrane to block air microorganisms. The plants were subsequently transplanted to a pot with an outer diameter of 23.5 cm and a height of 14 cm and cultured in a greenhouse for 2 months (Figure S3). A volume of 200 ml of sterile water was used to irrigate the plants every two days, and 200 ml of Hoagland nutrient solution was used to irrigate the plants once a week. Fungal infection of leaves was tested again in the 4th and 9th weeks during this period.
2.2.2. Measuring Physiological Traits and Growth Indices
The following plant physiological traits were measured: chlorophyll content (chlorophyll tester, Shandong Fangke Instrument Co., Ltd., model FK-YL01), total sugar content (Model A145-1-1 plant soluble sugar content test kit (colorimetric method), Nanjing Jiancheng Bioengineering Institute), peroxidase (POD) activity (A084-3-1 peroxidase (POD) assay kit (colorimetric method)) and total phenol (TP) content (A143-1-1 plant total phenol test box (colorimetric method)).
We measured the unit leaf area dry weight for the 2nd and 5th pairs of leaves from the top to the bottom of A. adenophora. We punched five leaf discs with an area of approximately 0.5 cm2 for each leaf via an 8 mm diameter stopper borer. These leaf discs were dried for 48 h at 65 °C in an oven to obtain dry weights, after which the unit leaf area of dry matter weight was calculated. All the plants were removed from the soil to clean the roots, and the attached water was blotted dry with filter paper. The stems and roots were cut, and the stem length, root length and branch number of the plants were recorded. After drying at 60 °C for 48 h, the aboveground dry weight, underground dry weight and root shoot ratio of the plants were obtained by combining the leaf dry weights harvested earlier.
2.3. The Resistance to Pathogens on A. adenophora Leaves by the Endophyte Colletotrichum
In the above 2.2 experiment, A. adenophora plants were inoculated with the endophytic fungi AX39, AX115, and AX198, whereas the control plants did not receive any endophytic fungi. After the physiological indices were measured in the 2.2 experiment, one week before the plant biomass was harvested, we selected 4 leaves from the fourth pair of equal areas for each pot of plants and inoculated them with the pathogenic fungi S188, G56, and Y122 from the Didymellaceae family and the control group (agar without fungal mycelia). Inoculation with the pathogen was performed as described previously [40]. Briefly, the pathogens were cultured on PDA media for 7 days, after which agar discs (6 mm2) containing fungal mycelia were inoculated into the fourth pair of leaves in each pot. There were 5 POTS in each group. Small wounds were made by lightly touching the underside of the leaf with a sterilized toothpick, resulting in a wound area of ~0.2 cm2. The inoculum agar was pressed against the wound on the underside of the leaf using Scotch tape and clipped in place with a bent hair clip. Control inoculation was performed with agar without fungal mycelia. After allowing for a period of 7 days for disease development, observations were made on the leaf surfaces, and the area of the diseased spot was measured. The size of the diseased spot was used as an indicator to evaluate the effect of endophytic fungal inoculation on the resistance of A. adenophora to pathogen infection.
2.4. The Resistance of A. adenophora Against to Drought Stress and Nutrient Stress by the Endophyte Colletotrichum
Sterile and endophyte-infected A. adenophora seedlings were planted and prepared as described in Section 2.2. These plants were divided into four groups: the control treatment without endophytes and the inoculation treatment with the AX39, AX115 and AX198 strains. At the same time, the soil was divided into three groups: normal treatment, drought stress treatment [the soil was watered with 100 mL/cup 20% PEG-6000 solution for the first time (drought stress, 20), with an osmotic potential of −4.906 bars to produce drought stress] [41], and nutrient stress treatment (the whole process without supplying nutrient solution), with 5 POTS in each treatment, for a total of 60 POTS of plants. After the plants were irrigated with sterile water according to the water requirements, Hoagland nutrient solution was added once a week for the normal and drought treatments, while only sterile water was added for the nutrient stress treatments. After 45 days of growth in an artificial climate chamber (Figure S4), the plants were harvested, and the chlorophyll content, branch number, root length, stem length, aboveground and underground dry weight and root shoot ratio of the plants were measured as described above.
2.5. Data Analysis
The fungal infection status of the leaves was expressed as the isolation rate (number of small leaves with fungal growth per plate/the total number of small leaves per plate × 100%). The response index (RI) was calculated to evaluate the direction (i.e., positive or negative) and intensity of the fungal effect by calculating physiological indices, biomass data and leaf spot area. The formula is as follows: (treatment-control)/control (Williamson and Richardson 1988). The Kolmogorov-Smirnov test was used to determine the normality of the data, and the nonparametric Mann-Whitney U test (for two groups) and Kruskal-Wallis test (for three or more groups) were used to test between-group differences for the data that were not normally distributed. For normally distributed data, analysis of variance (ANOVA) and post hoc comparisons were used to evaluate the differences between groups. Independent sample T tests (2 groups) and Duncan tests and Dunnett T3 tests were used for post hoc multiple comparisons (three or more groups) to test for differences between groups. Normality tests, homogeneity of variance tests, one-way analysis of variance, post hoc multiple comparisons, independent sample T tests and nonparametric analyses were performed via SPSS v.25.0 (IBM, Chicago, IL, USA). The data were plotted in GraphPad Prism 7 (GraphPad Software, Inc., La Jolla, CA, USA).
3. Results
3.1. Effects of Colletotrichum on Host A. adenophora Growth
To determine whether these endophytes can successfully colonize the host, we first detected the isolation rate of A. adenophora at 1, 4 and 9 weeks after Colletotrichum inoculation (Figure S2). The isolation rates of AX39, AX115 and AX198 decreased with increasing host growth time, with average isolation rates of 69.45%, 66.67% and 76.85%, respectively, in the first week and decreasing to 14.58%, 36.67% and 14.17%, respectively, in the ninth week. The isolation rates of strains AX39 and AX198 decreased significantly in the ninth week compared with those in the first week. There was no significant difference in colonization among AX39, AX198 and AX115 in each period.
Compared with those without inoculation, inoculation with AX39, AX115 and AX198 reduced plant photosynthesis-related physiological indices (i.e., chlorophyll content and soluble sugar content) (Figure 1ab) and increased plant resistance-related physiological indices [i.e., total phenolic (TP) and peroxidase (POD) activity] (Figure 1cd). The chlorophyll contents of the plants inoculated with the three strains were extremely significantly lower than those of the plants without inoculation, and the POD activities of the plants inoculated with AX39 and AX198 were significantly greater than those of the plants without inoculation. AX39 and AX115 had greater effects on reducing the chlorophyll content and total sugar content than AX198. Inoculation with fungal endophytes also reduced plant biomass (aboveground dry weight, underground dry weight, number of branches and the root shoot ratio) but increased stem length and root length (Figure 2a-f). Among them, both AX39 and AX115 commonly had greater response indices for aboveground biomass and stem length than did AX198; moreover, AX39 and AX115 also adversely affected the LMA (dry matter weight per unit leaf area) (g·m-2) of the second pair and the fifth pair of leaves, but AX198 positively affected the LMA of A. adenophora (Figure 3ab).
3.2. Resistance of Foliar Pathogens to A. adenophora Inoculated with the Endophyte Colletotrichum
The host A. adenophora showed no leaf spot when inoculated with any of the three Colletotrichum strains by either spraying the spore suspension (Figure 4a) or wounding and inoculating the plants with agar discs of Colletotrichum (Figure 4b). We subsequently tested the resistance of A. adenophora inoculated with the endophyte Colletotrichum to three foliar pathogens from the Didymellaceae family. With respect to infection by the foliar pathogens G56 and Y122 (members of Mesophoma ageratinae (Didymellaceae family)), inoculation with the AX39 and AX115 endophytes resulted in a larger leaf disease spot area than did inoculation without endophytes, suggesting that inoculation with G56 and Y122 promoted infection by AX39 and AX115. In contrast, the leaf disease spot area in the group inoculated with the AX198 endophyte was smaller than that in the uninoculated group, indicating that AX198 attenuated the pathogenic effects of the G56 and Y122 pathogens (Figure 5abd). With respect to infection by the pathogen Mesophoma speciosa (Didymellaceae family) S188, the areas of leaf disease spots inoculated with AX39 and AX115 were smaller than those inoculated with the uninoculated endophyte, indicating that AX39 and AX115 weakened the pathogenic effect of the pathogen S188; however, AX198 had no effect on S188 infection (Figure 5cd).
3.3. The Resistance of A. adenophora Inoculated with the Endophyte Colletotrichum against Drought Stress and Nutrient Stress
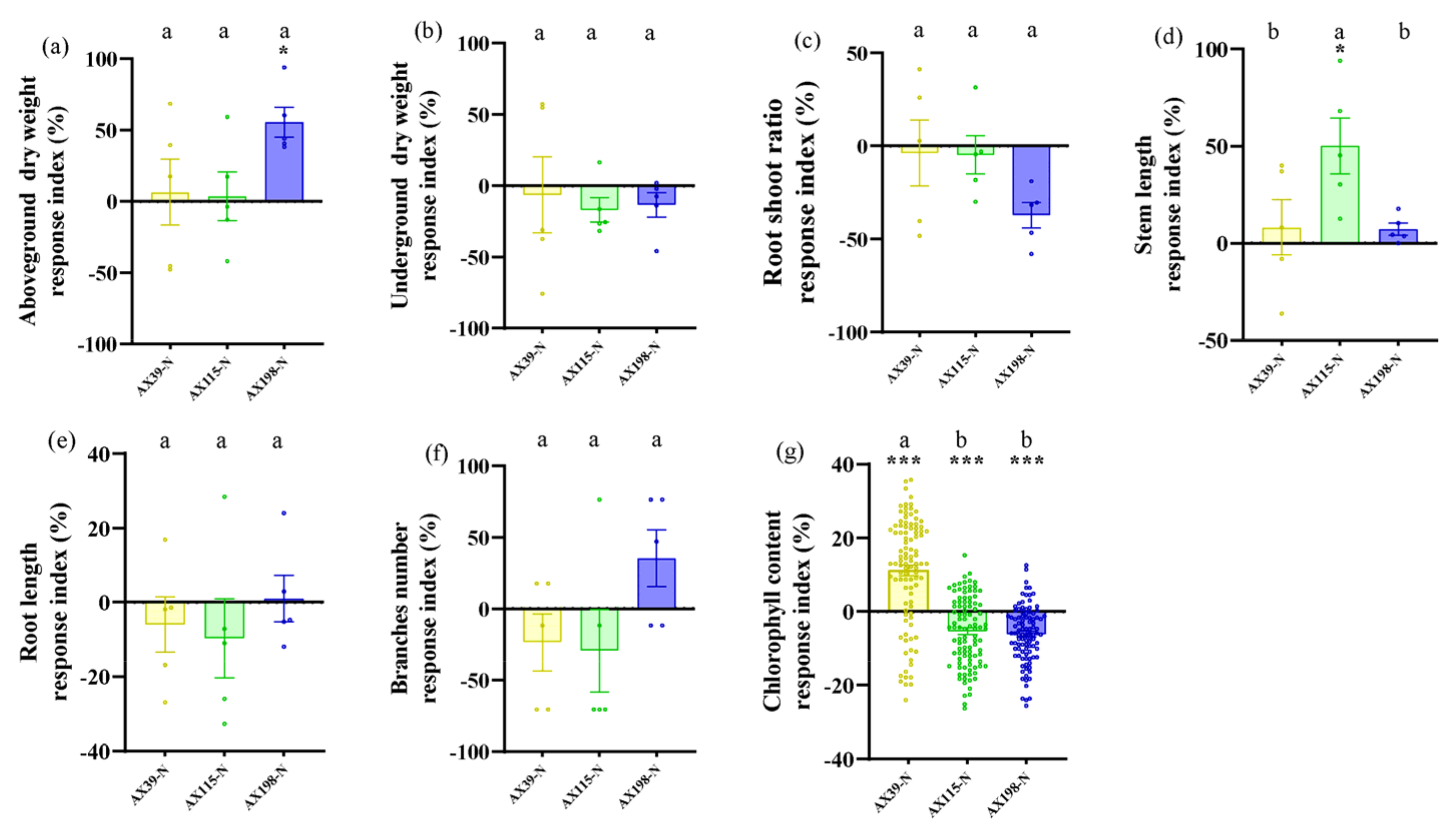
Compared with A. adenophora without endophyte inoculation, inoculation with the AX198 strain significantly increased stem length (Figure 6d) and chlorophyll content (Figure 6g) under drought stress; inoculation with AX198 significantly increased aboveground dry weight (Figure 7a), and inoculation with AX115 increased stem length (Figure 7d) under nutrient stress. In contrast to those under drought stress, inoculation with AX39 significantly increased the chlorophyll content, whereas inoculation with both AX115 and AX198 drastically decreased the chlorophyll content under nutrient stress (Figure 7g).
4. Discussion
Understanding the function of fungal endophytes associated with invasive plants has become an increasing concern of invasion biologists. In general, recently introduced fungal endophytes can directly help invasive plants improve growth, reproduction, defense against pathogens and herbivores and environmental tolerance [12,13,14,16]. Previously, a high abundance of the endophytic fungus Colletotrichum was found to be enriched in the leaves of A. adenophora [28]. In this study, we further tested the effects of these endophytic fungi on the growth of A. adenophora under pathogen and environmental stress conditions. (1) The host A. adenophora did not grow well after asymptomatic infection with the endophytic fungus Colletotrichum, as indicated by reduced plant photosynthesis-related physiological indices (chlorophyll content and soluble sugar content), increased plant resistance-related physiological indices [total phenolic (TP) and peroxidase (POD) activity], and decreased biomass. (2) Endophytic Colletotrichum fungi had some positive effects on the leaves of A. adenophora, but genetically distinct endophytic fungi presented different responses to different pathogens. (3) Under drought or nutrient stress, the endophyte Colletotrichum promoted the growth of A. adenophora, especially in terms of increasing stem length and aboveground biomass and regulating the leaf chlorophyll content.
In contrast to expectation 1, although the leaf of the host A. adenophora was not affected by inoculation with any of the strains of Colletotrichum inoculated via spore suspension (Figure 4a) or by wounding and inoculation via agar discs (Figure 4b), inoculation with Colletotrichum had adverse effects on A. adenophora growth because all the strains reduced the A. adenophora biomass (Figure 2). This adverse effect also reduced plant photosynthetic efficiency by reducing the contents of chlorophyll and soluble sugars (Figure 1ab). Photosynthesis is considered one of the main driving forces of plant growth, and leaf pathogens affect one of the most important physiological processes. Many studies have shown that pathogen infection results in a decrease in the photosynthetic rate and photosynthetic unit changes, and these changes may be due to cutting or damage to the photosynthetic apparatus [42]. Photosynthesis decreases in bean plants infected with Colletotrichum lindemuthianum [43], and photosynthesis decreases in acai leaves infected with Colletotrichum [42]. Moreover, it was also demonstrated that Monographella albescens was able to impair the photosynthetic process in both symptomatic and asymptomatic leaf areas [44]. In fact, many pathogens may impair photosynthesis in asymptomatic, although colonized, tissues, i.e., often called virtual lesions [45,46,47]. Therefore, inoculation with the endophytic fungus Colletotrichum inhibited photosynthesis in A. adenophora to some extent, leading to a further reduction in biomass, which also indicated that these endophytic Colletotrichum fungi in A. adenophora behave physiologically like pathogens. Previous studies have shown that beneficial microbes, like pathogens, can elicit a plant immune response (Wippel 2023), e.g., the induction of the plant hypersensitive response, including the formation of reactive oxygen species (ROS), the induction of salicylic acid (SA), jasmonic acid (JA) signalling, the upregulation of pathogenesis-related (PR) genes and plant defensin 1.2 (PDF1.2) [48,49]. It is valuable to determine whether the expression of immune-related genes such as PR1/2/5 and PDF1.2 is elicited in A. adenophora by the colonization of these endophytes in the future.
As expected 2, endophytic Colletotrichum fungi had some positive effects on the dominant pathogen Didymellaceae when inoculated on the leaves of A. adenophora (Figure 5cd). Didymellaceae are the dominant foliar pathogens. These pathogens occur widely on A. adenophora in the invaded range, and some members exhibit high virulence and a narrow host range [36]. In particular, their virulence evolves with invasion history [50]. Therefore, these pathogens may have important impacts on the expansion of A. adenophora along with a delayed invasion history. Our findings indicated that the Colletotrichum-Didymellaceae interaction may be important for understanding the role of foliar fungi in plant invasions. Similarly, many studies have verified that foliar fungi can help invasive host plants resist pathogens; for example, foliar endophytic fungi of the invasive plant Impatiens glandulifera appear to be antagonistic to rust fungus (Puccinia komarovii) [16]. Cryptosporiopsis quercina isolated from Tripterygium wilfordii, an endophytic fungus, produces a special active peptide, cyclohexenone, which inhibits the growth of the plant pathogens Sclerotinia sclerotiorum and Botrytis cinerea [51].
Nonetheless, our findings suggest that the mode of action of endophytic fungi against pathogens is much more complex and depends on the different genetic backgrounds of both endophytic fungi and the specific pathogen being investigated. For example, endophytes of different genotypes presented different resistances to pathogens, with the endophyte AX198 being more active against the pathogens G56 and Y122 than AX39 and AX115 (Figure 5abc). The resistance of endophytic fungi to different pathogen genotypes also differs, with the endogenous fungi AX39 and AX115 promoting infection by the pathogens M. ageratinae G56 and Y122 but weakening infection by the pathogen M. speciosa S188 (Figure 5). These results suggested that the resistance of the AX198 strain differed from that of the AX39 and AX115 strains to pathogens, which reflected the differences among strains from different complexes. Second, the results also indicated that AX198 resulted in different LMA responses (Figure 3) and different physiological responses (Figure 1) in A. adenophora. Because most Colletotrichum strains inhibit the growth of Didymellaceae strains both in vitro and in vivo (unpublished data), the resistance of these endophytic Colletotrichum strains may occur through competitive exclusion of the Didymellaceae pathogen. On the other hand, our results also suggest a synergistic interaction between Colletotrichum and other pathogens, which results in greater virulence against A. adenophora. Similar to our previous study, the foliar fungus Colletotrichum sp. promoted the pathogenicity of Diaporthe helianthi on A. adenophora leaves [29]. Coinfections are common in nature and usually increase the pathogenicity of pathogens to the host [52]. For example, compared with inoculation with either of the two pathogens separately, coinfection with Verticillium dahliae or Colletotrichum coccodes causes more severe foliar disease symptoms and crown rot in potato (Nicola) [53].
The interactions between endophytic microorganisms and plants are influenced by environmental factors, including nutrient conditions [54] and drought conditions [55]. As expected 3, inoculation with the endophyte Colletotrichum could alleviate the effects of drought stress and nutrient stress on A. adenophora by increasing several growth indicators of A. adenophora, including the aboveground biomass, stem length and chlorophyll content. The response intensity of A. adenophora under drought or nutrient stress also varied with the genetic background of these endophytes. Relatively, the strain AX198 promoted the growth of A. adenophora more strongly than the other two strains did (Figure 6 and Figure 7). This result is consistent with that of strain AX198 being more active against the pathogens G56 and Y122 than AX39 and AX115 (Figure 5abc). Moreover, the greater LMA after AX198 inoculation was consistent with these results (Figure 3). Interestingly, compared with those under normal hydrothermal and nutrient conditions, both AX39 and AX115 resulted in longer stem lengths (see Figure 2d), which reflects a complex response to abiotic stress and depends on the different genetic backgrounds of endophytic fungi. More physiological indicators for stress resistance, including superoxide dismutase (SOD) and catalase (CAT) activity, membrane relative permeability, and malondialdehyde (MDA) content [56,57,58,59,60], are valuable for understanding the detailed physiological changes induced by genetically distinct endophytes in A. adenophora under stress conditions.
Furthermore, the effects of the endophyte Colletotrichum on plant A. adenophora may be dose-dependent of environmental factor. For example, Zhang et al. studied the effects of endophytic fungal infection on the germination and seedling growth of perennial ryegrass under different salt stress levels [61]. They reported that at low concentrations of NaCl, infection with endophytic fungi had little effect on seedling growth, but at high concentrations, the fungus had a significant promoting effect on seedling growth. Cheplick et al. reported that inoculation with endophytic fungi reduced the number of tillers in perennial ryegrass under drought stress, but the number of tillers increased after the recovery period [62]. The endophytic fungus Colletotrichum tofieldiae Ct0861, which is a symbiotic fungus of Arabidopsis thaliana, promotes plant growth and reproduction only under phosphate starvation conditions [63,64]. In soils with different water-holding capacities, the effects of fungal endophytes on the growth of perennial ryegrass differ because of their different effects under drought conditions [65]. Therefore, it is necessary to explore the interactions between endophytic microorganisms and plants under different drought and nutrient stresses (including different nutrient element stresses).
In general, our findings suggested the high abundance of the leaf endophytic fungus Colletotrichum did not significantly promote the growth of A. adenophora under normal hydrothermal and nutrient conditions; however, these endophytic Colletotrichum strains could help A. adenophora resist foliar fungal pathogens and improve the response to drought or nutrient stress, depending on the genetic background of the endophytic fungi. Our results provide novel clues to explain why A. adenophora contains a high abundance of Colletotrichum endophytes [27], but some Colletotrichum endophytes have a weak negative impact on the growth of A. adenophora [29]. Nonetheless, as plant mutualists, microbiomes are not a single organism but a community of species with complex interactions among microbial taxa and between microbes and their shared host. The presence of an individual strain is detrimental to the host, but multiple strains can be beneficial [66]. Therefore, the effects of multiple Colletotrichum inoculations on the stress resistance of Colletotrichum are also worth studying in the future.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Han-Bo Zhang: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Validation, Writing – review & editing. Ai-Ling Yang: Investigation, Methodology, Software, Writing – original draft, Writing – review & editing. Yu-Xuan Li: Investigation, Methodology. Zhao-Ying Zeng: Investigation, Methodology.
Funding
This work was funded by the National Key R&D Program of China (No. 2022YFC2601100).
Acknowledgments
We thank Jin-Peng Li and Lu Cheng at Yunnan University for their help with the experiments and Lin Chen and Kai Fang at Yunnan University for their help with the experiments and data analysis.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
References
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. Endophytes: a treasure house of bioactive compounds of medicinal importance. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef] [PubMed]
- Specian, V.; Sarragiotto, M.H.; Pamphile, J.A.; Clemente, E. Chemical characterization of bioactive compounds from the endophytic fungus Diaporthe helianthi isolated from Luehea divaricata. Braz. J. Microbiol. 2012, 43, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.; Vieira, M.; Cota, B.; Johann, S.; Alves, T.; Zani, C.; Rosa, C. Endophytic fungi of tropical forests: a promising source of bioactive prototype molecules for the treatment of neglected diseases. In: Ekinci D (ed) Drug development—a case study based insight into modern strategies. Intech, Croatia, 2011; pp. 1–18. [CrossRef]
- Wani, Z.A.; Ashraf, N.; Mohiuddin, T.; Riyaz-Ul-Hassan, S. Plant-endophyte symbiosis, an ecological perspective. Appl. Microbiol. Biot. 2015, 99, 2955–2965. [Google Scholar] [CrossRef]
- Ji, S.H.; Gururani, M.A.; Chun, S.C. Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiol. Res. 2014, 169, 83–98. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Ahemad, M.; Oves, M.; Wani, P.A. Plant growth promotion by phosphate solubilizing fungi–current perspective. Arch. Agron. Soil Sci. 2010, 56, 73–98. [Google Scholar] [CrossRef]
- Arnold, A.E.; Mejía, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. U. S. A. 2003, 100, 15649–15654. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: a key player in plant abiotic stress tolerance. Biol. Plant 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Lata, R.; Chowdhury, S.; Gond, S.K.; White, J.F., Jr. Induction of abiotic stress tolerance in plants by endophytic microbes. Lett. Appl. Microbiol. 2018, 66, 268–276. [Google Scholar] [CrossRef]
- Evans, H.C. The endophyte-enemy release hypothesis: implications for classical biological control and plant invasions. CABI 2008, 20–25. [Google Scholar] [CrossRef]
- Aschehoug, E.T.; Metlen, K.L.; Callaway, R.M.; Newcombe, G. Fungal endophytes directly increase the competitive effects of an invasive forb. Ecology 2012, 93, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Uchitel, A.; Omacini, M.; Chaneton, E.J. Inherited fungal symbionts enhance establishment of an invasive annual grass across successional habitats. Oecologia 2011, 165, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Bastias, D.A.; Gianoli, E.; Gundel, P.E. Fungal endophytes can eliminate the plant growth-defence trade-off. New Phytol. 2021, 230, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Bashir, I.; War, A.F.; Rafiq, I.; Reshi, Z.A.; Rashid, I.; Shouche, Y.S. Uncovering the secret weapons of an invasive plant: The endophytic microbes of Anthemis cotula. HELIYON 2024, 10. [Google Scholar] [CrossRef] [PubMed]
- Currie, A.F.; Gange, A.C.; Abrazak, N.; Ellison, C.A.; Maczey, N.; Wood, S.V. Endophytic fungi in the invasive weed Impatiens glandulifera: a barrier to classical biological control? Weed Res. 2020, 60, 50–59. [Google Scholar] [CrossRef]
- Hartley, S.E.; Eschen, R.; Horwood, J.M.; Gange, A.C.; Hill, E.M. Infection by a foliar endophyte elicits novel arabidopside-based plant defence reactions in its host, Cirsium arvense. New Phytol. 2015, 205, 816–827. [Google Scholar] [CrossRef]
- Rudgers, J.A.; Clay, K. An invasive plant-fungal mutualism reduces arthropod diversity. Ecol. Lett. 2008, 11, 831–840. [Google Scholar] [CrossRef]
- Mateu, M.G.; Baldwin, A.H.; Maul, J.E.; Yarwood, S.A. Dark septate endophyte improves salt tolerance of native and invasive lineages of Phragmites australis. ISME J. 2020, 14, 1943–1954. [Google Scholar] [CrossRef]
- Poudel, A.S.; Jha, P.K.; Shrestha, B.B.; Muniappan, R. Biology and management of the invasive weed Ageratina adenophora (Asteraceae): current state of knowledge and future research needs. Weed Res. 2019, 59, 79–92. [Google Scholar] [CrossRef]
- Perez, L.I.; Gundel, P.E.; Parisi, P.A.G.; Moyano, J.; Fiorenza, J.E.; Omacini, M.; Nunez, M.A. Can seed-borne endophytes promote grass invasion by reducing host dependence on mycorrhizas? Fungal Ecol. 2021, 52. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Y.Z. Invasion dynamics and potential spread of the invasive alien plant species Ageratina adenophora (Asteraceae) in China. Divers. Distrib. 2006, 12, 397–408. [Google Scholar] [CrossRef]
- Feng, J.; Zhu, Y. Alien invasive plants in China: risk assessment and spatial patterns. BIODIVERS. CONSERV. 2010, 19, 3489–3497. [Google Scholar] [CrossRef]
- Xu, C.W.; Yang, M.Z.; Chen, Y.J.; Chen, L.M.; Zhang, D.Z.; Mei, L.; Shi, Y.T.; Zhang, H.B. Changes in non-symbiotic nitrogen-fixing bacteria inhabiting rhizosphere soils of an invasive plant Ageratina adenophora. Appl. Soil. Ecol. 2012, 54, 32–38. [Google Scholar] [CrossRef]
- Fang, K.; Bao, Z.S.N.; Chen, L.; Zhou, J.; Yang, Z.P.; Dong, X.F.; Zhang, H.B. Growth-promoting characteristics of potential nitrogen-fixing bacteria in the root of an invasive plant Ageratina adenophora. PeerJ 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Fang, K.; Wang, Y.Z.; Zhang, H.B. Differential effects of plant growth-promoting bacteria on invasive and native plants. S. Afr. J. Bot. 2019, 124, 94–101. [Google Scholar] [CrossRef]
- Chen, L.; Zhou, J.; Zeng, T.; Miao, Y.F.; Mei, L.; Yao, G.B.; Fang, K.; Dong, X.F.; Sha, T.; Yang, M.Z.; et al. Quantifying the sharing of foliar fungal pathogens by the invasive plant Ageratina adenophora and its neighbours. New Phytol. 2020, 227, 1493–1504. [Google Scholar] [CrossRef]
- Mei, L.; Zhu, M.; Zhang, D.Z.; Wang, Y.Z.; Guo, J.; Zhang, H.B. Geographical and temporal changes of foliar fungal endophytes associated with the invasive plant Ageratina adenophora. Microb. Ecol. 2014, 67, 402–409. [Google Scholar] [CrossRef]
- Fang, K.; Chen, L.M.; Zhang, H.B. Evaluation of foliar fungus-mediated interactions with below and aboveground enemies of the invasive plant Ageratina adenophora. Ecol. Evol. 2021, 11, 526–535. [Google Scholar] [CrossRef]
- Ma, X.; Nontachaiyapoom, S.; Jayawardena, R.S.; Hyde, K.D.; Gentekaki, E.; Zhou, S.; Qian, Y.; Wen, T.; Kang, J. Endophytic Colletotrichum species from Dendrobium spp. in China and Northern Thailand. MycoKeys 2018, 23–57. [Google Scholar] [CrossRef]
- Wang, Y.T.; Lo, H.S.; Wang, P.H. Endophytic fungi from Taxus mairei in Taiwan: first report of Colletotrichum gloeosporioides as an endophyte of Taxus mairei. BOT. STUD. 2008, 49, 39–43. [Google Scholar]
- Liu, J.W.; Manawasinghe, I.S.; Liao, X.N.; Mao, J.; Dong, Z.Y.; Jayawardena, R.S.; Wanasinghe, D.N.; Shu, Y.X.; Luo, M. Endophytic Colletotrichum (Sordariomycetes, Glomerellaceae) species associated with Citrus grandis cv. "Tomentosa" in China. MycoKeys 2022, 163-188. [CrossRef]
- Win, P.M.; Matsumura, E.; Fukuda, K. Effects of pesticides on the diversity of endophytic fungi in tea plants. Microb. Ecol. 2021, 82, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Photita, W.; Lumyong, S.; Lumyong, P.; McKenzie, E.H.C.; Hyde, K.D. Are some endophytes of Musa acuminata latent pathogens? FUNGAL DIVERS. 2004, 16, 131–140. [Google Scholar]
- Crawford, K.M.; Knight, T.M. Competition overwhelms the positive plant-soil feedback generated by an invasive plant. Oecologia 2017, 183, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, A.L.; Li, Y.X.; Zhang, H.B. Virulence and host range of fungi associated with the invasive plant Ageratina adenophora. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Hou, L.W.; Duan, W.J.; Crous, P.W.; Cai, L. Didymellaceae revisited. Stud. Mycol. 2017, 105–159. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.L.; Chen, L.; Cheng, L.; Li, J.P.; Zeng, Z.Y.; Zhang, H.B. Two Novel Species of Mesophoma gen. nov. from China. Curr. Microbiol. 2023, 80. [Google Scholar] [CrossRef]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Gilbert, G.S.; Webb, C.O. Phylogenetic signal in plant pathogen-host range. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 4979–4983. [Google Scholar] [CrossRef]
- Michel, E.; Kaufmann, M.R. The osmotic potential of polyethylene glycol 60001. Plant Physiol. 1973, 5, 914–916. [Google Scholar] [CrossRef]
- Castro, G.L.S.; Silva Junior, D.D.; Bueno, A.C.S.O.; Silva, G.B. Anthracnose in acai palm leaves reduces leaf gas exchange and chlorophyll a fluorescence. TROP. PLANT PATHOL. 2017, 42, 13–20. [Google Scholar] [CrossRef]
- Alves, A.A.; da Silva Guimaraes, L.M.; de Melo Chaves, A.R.; DaMatta, F.M.; Alfenas, A.C. Leaf gas exchange and chlorophyll a fluorescence of Eucalyptus urophylla in response to Puccinia psidii infection. ACTA PHYSIOL. PLANT. 2011, 33, 1831–1839. [Google Scholar] [CrossRef]
- Tatagiba, S.D.; DaMatta, F.M.; Rodrigues, F.A. Leaf gas exchange and Chlorophyll a fluorescence imaging of rice leaves infected with Monographella albescens. Phytopathology 2015, 105, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Bastiaans, L. Ratio between virtual and visual lesion size as a measure to describe reduction in leaf photosynthesis of rice due to leaf blast. Phytopathology 1991, 81, 611–615. [Google Scholar] [CrossRef]
- Berger, S.; Sinha, A.K.; Roitsch, T. Plant physiology meets phytopathology: plant primary metabolism and plant-pathogen interactions. J. Exp. Bot. 2007, 58, 4019–4026. [Google Scholar] [CrossRef]
- Domiciano, G.P.; Resende, R.; Rodrigues, F.; DaMatta, F. Alteração na fotossíntese de plantas infectadas por patógenos. Revisão Anual De Patologia De Plantas 2009, 17, 305–339. [Google Scholar]
- Zarei, A.; Körbes, A.P.; Younessi, P.; Montiel, G.; Champion, A.; Memelink, J. Two GCC boxes and AP2/ERF-domain transcription factor ORA59 in jasmonate/ethylene-mediated activation of the PDF1.2 promoter in Arabidopsis. PLANT MOL. BIOL. 2011, 75, 321–331. [Google Scholar] [CrossRef]
- Anisimova, O.K.; Shchennikova, A.V.; Kochieva, E.Z.; Filyushin, M.A. Pathogenesis-related genes of PR1, PR2, PR4, and PR5 families are involved in the response to Fusarium infection in garlic (Allium sativum L.). Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Fang, K.; Zhou, J.; Chen, L.; Li, Y.X.; Yang, A.L.; Dong, X.F.; Zhang, H.B. Virulence and community dynamics of fungal species with vertical and horizontal transmission on a plant with multiple infections. Plos Pathog. 2021, 17. [Google Scholar] [CrossRef]
- Li, J.Y.; Strobel, G.A. Jesterone and hydroxy-jesterone antioomycete cyclohexenone epoxides from the endophytic fungus Pestalotiopsis jesteri. Phytochemistry 2001, 57, 261–265. [Google Scholar] [CrossRef]
- Laine, A.L. Context-dependent effects of induced resistance under co-infection in a plant-pathogen interaction. EVOL. APPL. 2011, 4, 696–707. [Google Scholar] [CrossRef]
- Tsror, L.; Hazanovsky, M. Effect of coinoculation by Verticillium dahliae and Colletotrichum coccodes on disease symptoms and fungal colonization in four potato cultivars. Plant Pathol. 2001, 50, 483–488. [Google Scholar] [CrossRef]
- Dai, Z.; Su, W.; Chen, H.; Barberan, A.; Zhao, H.; Yu, M.; Yu, L.; Brookes, P.C.; Schadt, C.W.; Chang, S.X.; et al. Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of Actinobacteria and Proteobacteria in agro-ecosystems across the globe. GLOBAL CHANGE BIOL. 2018, 24, 3452–3461. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.; Styer, A.B.; Pierroz, G.; Goncalves, A.P.; Pasricha, R.; Hazra, A.B.; Bubner, P.; Coleman-Derr, D. Drought drives spatial variation in the millet root microbiome. FRONT. PLANT SCI. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alamri, S.A.; Ali, H.M.; Alayafi, A.A. Bacillus firmus (SW5) augments salt tolerance in soybean (Glycine max L.) by modulating root system architecture, antioxidant defense systems and stress-responsive genes expression. PLANT PHYSIOLO. BIOCH. 2018, 132, 375–384. [Google Scholar] [CrossRef]
- Li, J.; Meng, B.; Chai, H.; Yang, X.; Song, W.; Li, S.; Lu, A.; Zhang, T.; Sun, W. Arbuscular mycorrhizal fungi alleviate drought stress in C3 (Leymus chinensis) and C4 (Hemarthria altissima) grasses via altering antioxidant enzyme activities and photosynthesis. FRONT. PLANT SCI. 2019, 10. [Google Scholar] [CrossRef]
- Batool, T.; Ali, S.; Seleiman, M.F.; Naveed, N.H.; Ali, A.; Ahmed, K.; Abid, M.; Rizwan, M.; Shahid, M.R.; Alotaibi, M.; et al. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Mengistu, A.A. Endophytes: colonization, behaviour, and their role in defense mechanism. Int. J. Microbiol. 2020. [Google Scholar] [CrossRef]
- AlKahtani, M.D.F.; Hafez, Y.M.; Attia, K.; Rashwan, E.; Al Husnain, L.; AlGwaiz, H.I.M.; Abdelaal, K.A.A. Evaluation of silicon and proline application on the oxidative machinery in drought-stressed sugar beet. ANTIOXIDANTS 2021, 10. [Google Scholar] [CrossRef]
- Zhang, P.; Hu, L.; Fu, J. Effects of endophytic fungi on seed germination of perennial ryegrass under salt stress. Pratacultural Science 2012, 29, 1094–1099. [Google Scholar]
- Cheplick, G.P.; Perera, A.; Koulouris, K.F. Effect of drought on the growth of Lolium perenne genotypes with and without fungal endophytes. Funct. Ecol. 2000, 14, 657–667. [Google Scholar] [CrossRef]
- Garcia, E.; Alonso, A.; Platas, G.; Sacristan, S. The endophytic mycobiota of Arabidopsis thaliana. FUNGAL DIVERS. 2013, 60, 71–89. [Google Scholar] [CrossRef]
- Hiruma, K.; Gerlach, N.; Sacristan, S.; Nakano, R.T.; Hacquard, S.; Kracher, B.; Neumann, U.; Ramirez, D.; Bucher, M.; O'Connell, R.J.; et al. Root endophyte Colletotrichum tofieldiae confers plant fitness benefits that are phosphate status dependent. Cell 2016, 165, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, J.; Zhang, G.; Lin, W.; Tian, P. Effects of fungal endophytes on the growth of perennial ryegrass under drought condition. Pratacultural Science 2016, 33, 599–607. [Google Scholar]
- Laurich Jason, R.; Lash, E.; O'Brien Anna, M.; Pogoutse, O.; Frederickson Megan, E. Community interactions among microbes give rise to host-microbiome mutualisms in an aquatic plant. mBio. 2024, 15, e00972-00924. [Google Scholar] [CrossRef]
Figure 1.
Physiological indices of A. adenophora inoculated with endophytic Colletotrichum strains. (a) Chlorophyll content, (b) soluble sugar content, (c) total phenol content, and (d) peroxidase activity. Dots with different colours represent the raw data of each sample inoculated with the Colletotrichum AX39, AX115 and AX198 strains. The RI represents the response index, where the negative RI in panels (a) and (b) indicates reduced chlorophyll content and soluble sugar content in the treatment with Colletotrichum spp. infection compared with the control without Colletotrichum spp. infection. The positive RIs in panels (c) and (d) indicate increased total phenol content and peroxidase POD activity in the treatment with Colletotrichum spp. infection compared with the control without Colletotrichum spp. infection. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each treatment group and the control group (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to determine whether the difference in the RI was significant among the treatments inoculated with AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
Figure 1.
Physiological indices of A. adenophora inoculated with endophytic Colletotrichum strains. (a) Chlorophyll content, (b) soluble sugar content, (c) total phenol content, and (d) peroxidase activity. Dots with different colours represent the raw data of each sample inoculated with the Colletotrichum AX39, AX115 and AX198 strains. The RI represents the response index, where the negative RI in panels (a) and (b) indicates reduced chlorophyll content and soluble sugar content in the treatment with Colletotrichum spp. infection compared with the control without Colletotrichum spp. infection. The positive RIs in panels (c) and (d) indicate increased total phenol content and peroxidase POD activity in the treatment with Colletotrichum spp. infection compared with the control without Colletotrichum spp. infection. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each treatment group and the control group (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to determine whether the difference in the RI was significant among the treatments inoculated with AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
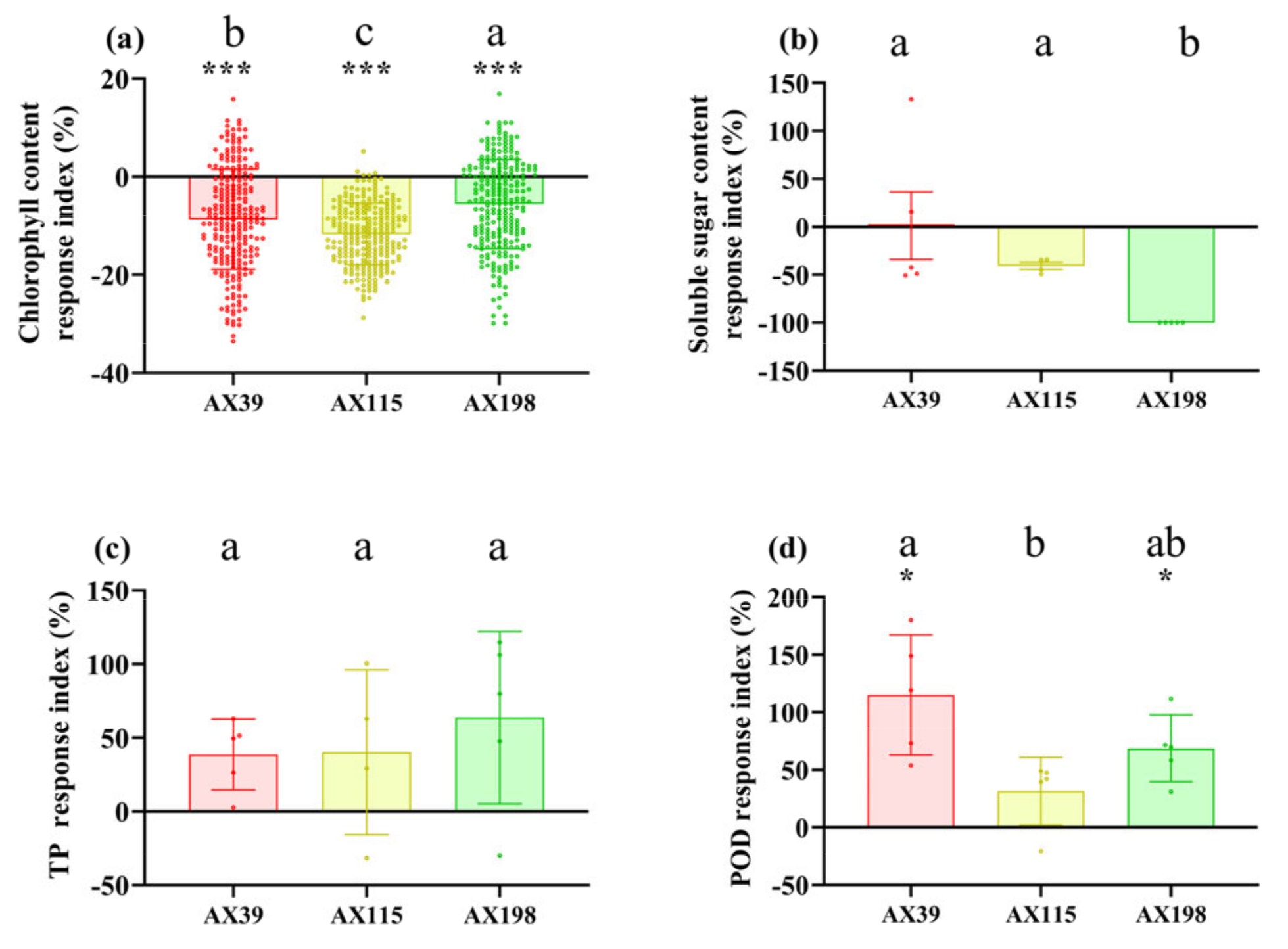
Figure 2.
Biomass of A. adenophora inoculated with endophytic Colletotrichum strains. (a) Aboveground parts, (b) underground parts, (c) branch number, (d) stem length, (e) root length, and (f) root shoot ratio. Dots with different colours represent the raw data of each sample inoculated with the Colletotrichum AX39, AX115 and AX198 strains. A negative RI indicates a reduced biomass of A. adenophora in the experimental treatment with Colletotrichum spp. infection compared with that in the control without Colletotrichum spp. infection. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each treatment group and the control group (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to determine whether the difference in the RI was significant among the treatments inoculated with AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
Figure 2.
Biomass of A. adenophora inoculated with endophytic Colletotrichum strains. (a) Aboveground parts, (b) underground parts, (c) branch number, (d) stem length, (e) root length, and (f) root shoot ratio. Dots with different colours represent the raw data of each sample inoculated with the Colletotrichum AX39, AX115 and AX198 strains. A negative RI indicates a reduced biomass of A. adenophora in the experimental treatment with Colletotrichum spp. infection compared with that in the control without Colletotrichum spp. infection. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each treatment group and the control group (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to determine whether the difference in the RI was significant among the treatments inoculated with AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
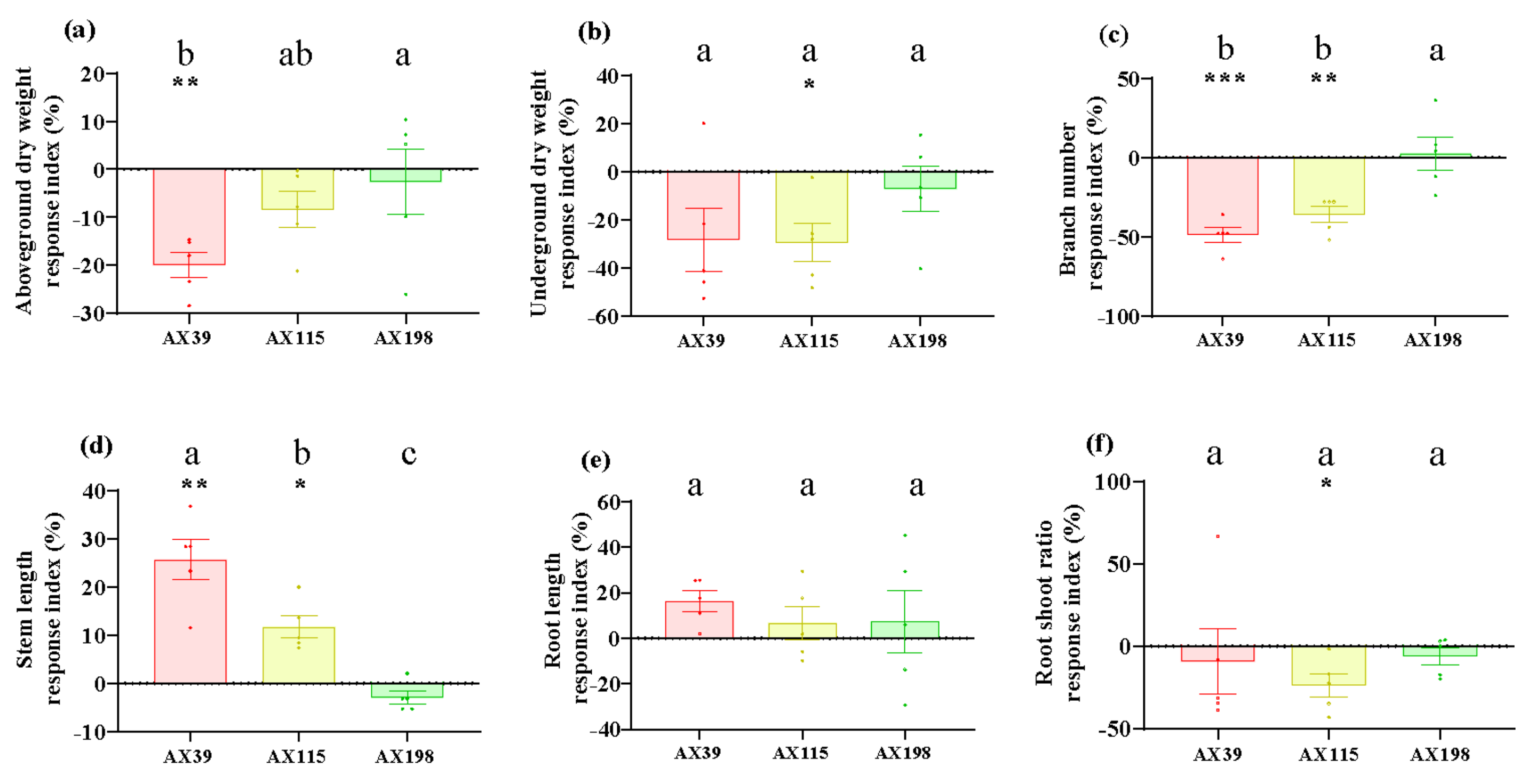
Figure 3.
LMA (dry weight per unit area) of A. adenophora inoculated with endophytic Colletotrichum strains. (a) The second pair of leaves, (b) the fifth pair of leaves. Dots with different colours represent the raw data of each sample inoculated with the Colletotrichum AX39, AX115 and AX198 strains. A negative RI indicates a reduced LMA of A. adenophora in the experimental treatment with Colletotrichum spp. infection compared with that in the control without Colletotrichum spp. infection. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each treatment group and the control group (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to determine whether the difference in the RI was significant among the different treatments inoculated with AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
Figure 3.
LMA (dry weight per unit area) of A. adenophora inoculated with endophytic Colletotrichum strains. (a) The second pair of leaves, (b) the fifth pair of leaves. Dots with different colours represent the raw data of each sample inoculated with the Colletotrichum AX39, AX115 and AX198 strains. A negative RI indicates a reduced LMA of A. adenophora in the experimental treatment with Colletotrichum spp. infection compared with that in the control without Colletotrichum spp. infection. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each treatment group and the control group (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to determine whether the difference in the RI was significant among the different treatments inoculated with AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
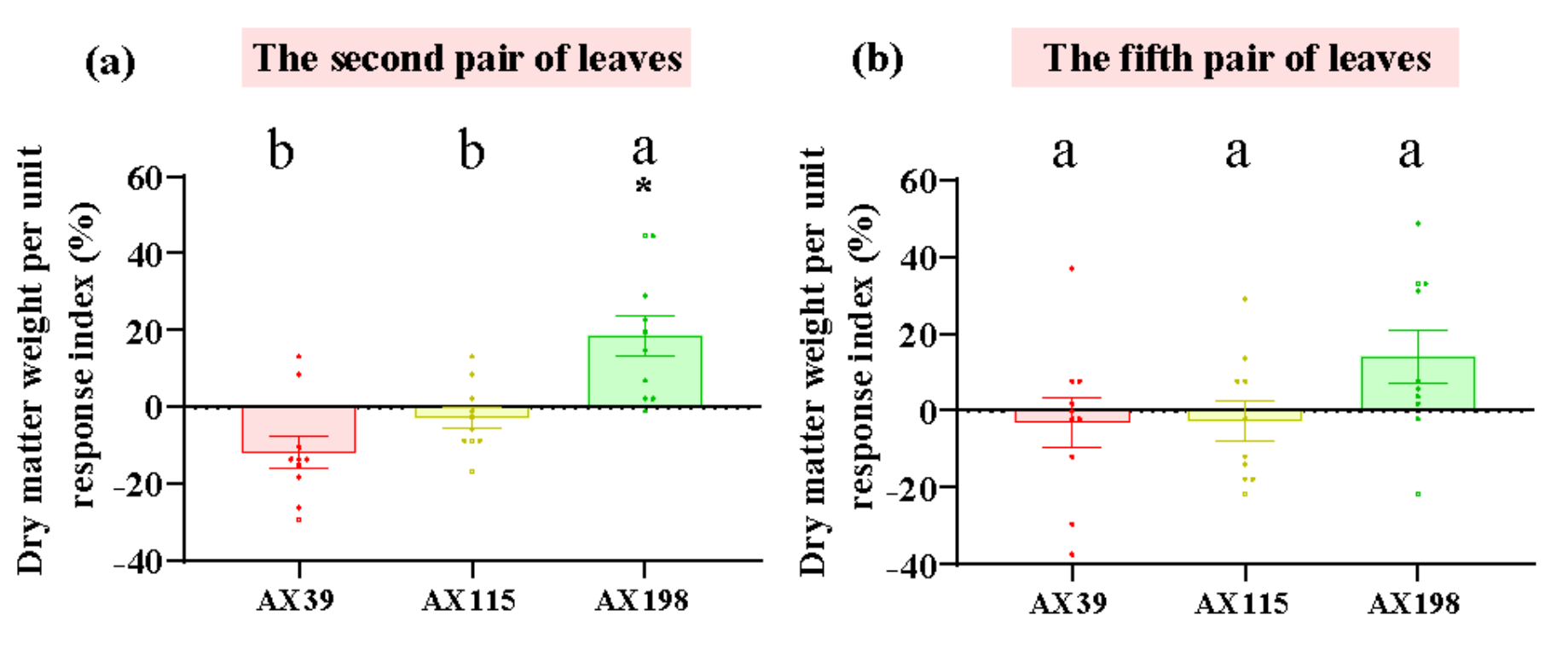
Figure 4.
The asymptomatic leaves of A. adenophora plants inoculated with Colletotrichum spore mixture (a) and wounded and inoculated with agar discs of Colletotrichum (b). "CK" represents the control group without Colletotrichum inoculation.
Figure 4.
The asymptomatic leaves of A. adenophora plants inoculated with Colletotrichum spore mixture (a) and wounded and inoculated with agar discs of Colletotrichum (b). "CK" represents the control group without Colletotrichum inoculation.

Figure 5.
Pathogenicity effects of inoculating endophyte Colletotrichum strains on A. adenophora after challenge with the (a) pathogen G56, (b) pathogen Y122 and (c) pathogen S188. Dots with different colours represent the raw data of each sample inoculated with the Colletotrichum AX39, AX115 and AX198 strains. The specific leaf spot area and morphology are shown in (d); scale bar =10 mm, and "CK" represents the control group without Colletotrichum inoculation. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each treatment group and the control group (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to determine whether the difference in the RI was significant among the different treatments inoculated with AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
Figure 5.
Pathogenicity effects of inoculating endophyte Colletotrichum strains on A. adenophora after challenge with the (a) pathogen G56, (b) pathogen Y122 and (c) pathogen S188. Dots with different colours represent the raw data of each sample inoculated with the Colletotrichum AX39, AX115 and AX198 strains. The specific leaf spot area and morphology are shown in (d); scale bar =10 mm, and "CK" represents the control group without Colletotrichum inoculation. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each treatment group and the control group (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to determine whether the difference in the RI was significant among the different treatments inoculated with AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
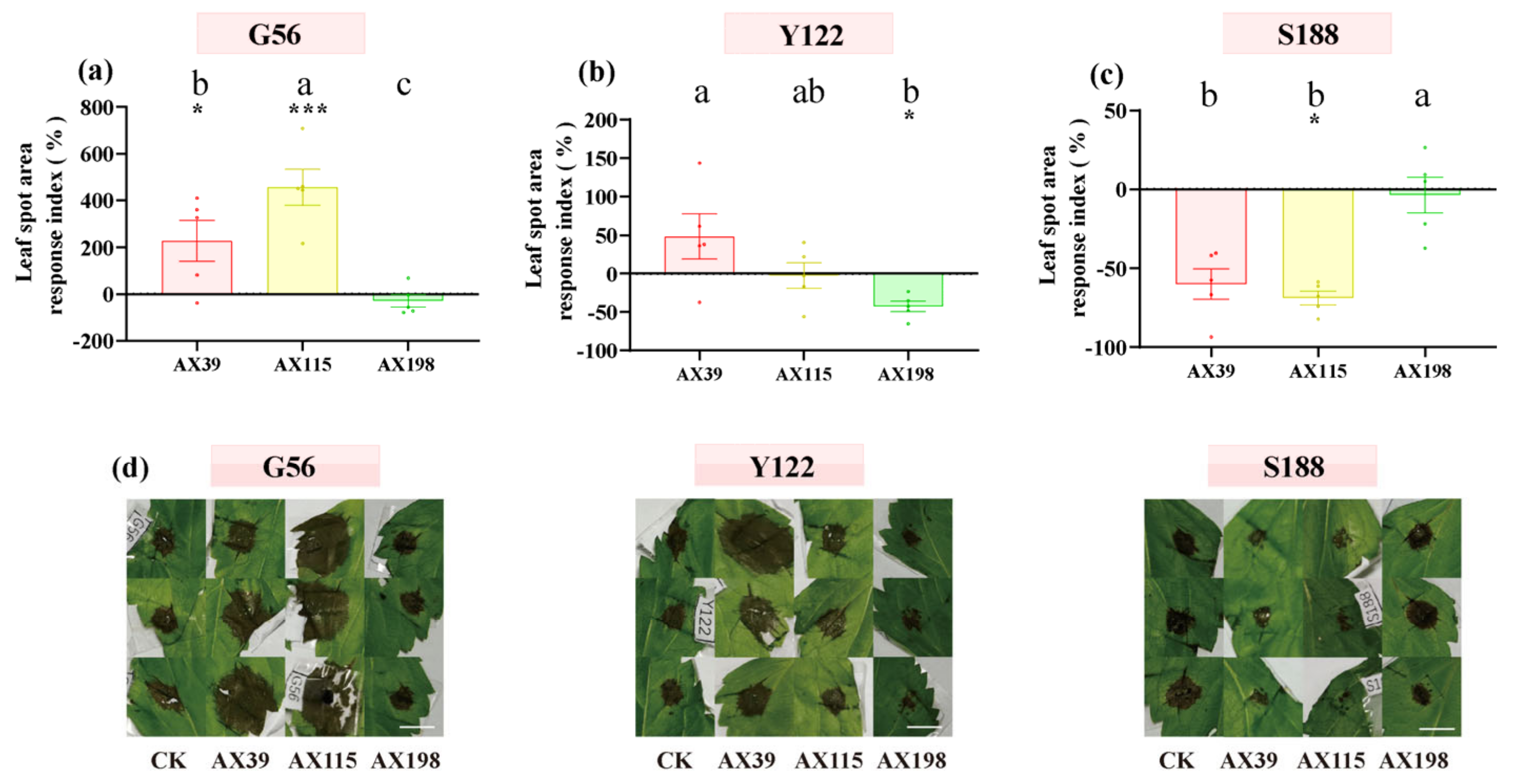
Figure 6.
Biomass and chlorophyll content of A. adenophora inoculated with endophyte Colletotrichum strains under normal treatment and drought stress. (a) Aboveground parts, (b) underground parts, (c) root shoot ratio, (d) stem length, (e) root length, (f) branch number, and (g) chlorophyll content. A positive RI indicates an increased biomass of A. adenophora in the drought stress (-W) treatment with Colletotrichum strain (AX39, AX115, or AX198) inoculation compared with that without Colletotrichum inoculation. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each drought stress treatment group and the normal treatment group (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to show that the differences in the RIs were significant among the treatments inoculated with strains AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
Figure 6.
Biomass and chlorophyll content of A. adenophora inoculated with endophyte Colletotrichum strains under normal treatment and drought stress. (a) Aboveground parts, (b) underground parts, (c) root shoot ratio, (d) stem length, (e) root length, (f) branch number, and (g) chlorophyll content. A positive RI indicates an increased biomass of A. adenophora in the drought stress (-W) treatment with Colletotrichum strain (AX39, AX115, or AX198) inoculation compared with that without Colletotrichum inoculation. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each drought stress treatment group and the normal treatment group (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to show that the differences in the RIs were significant among the treatments inoculated with strains AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
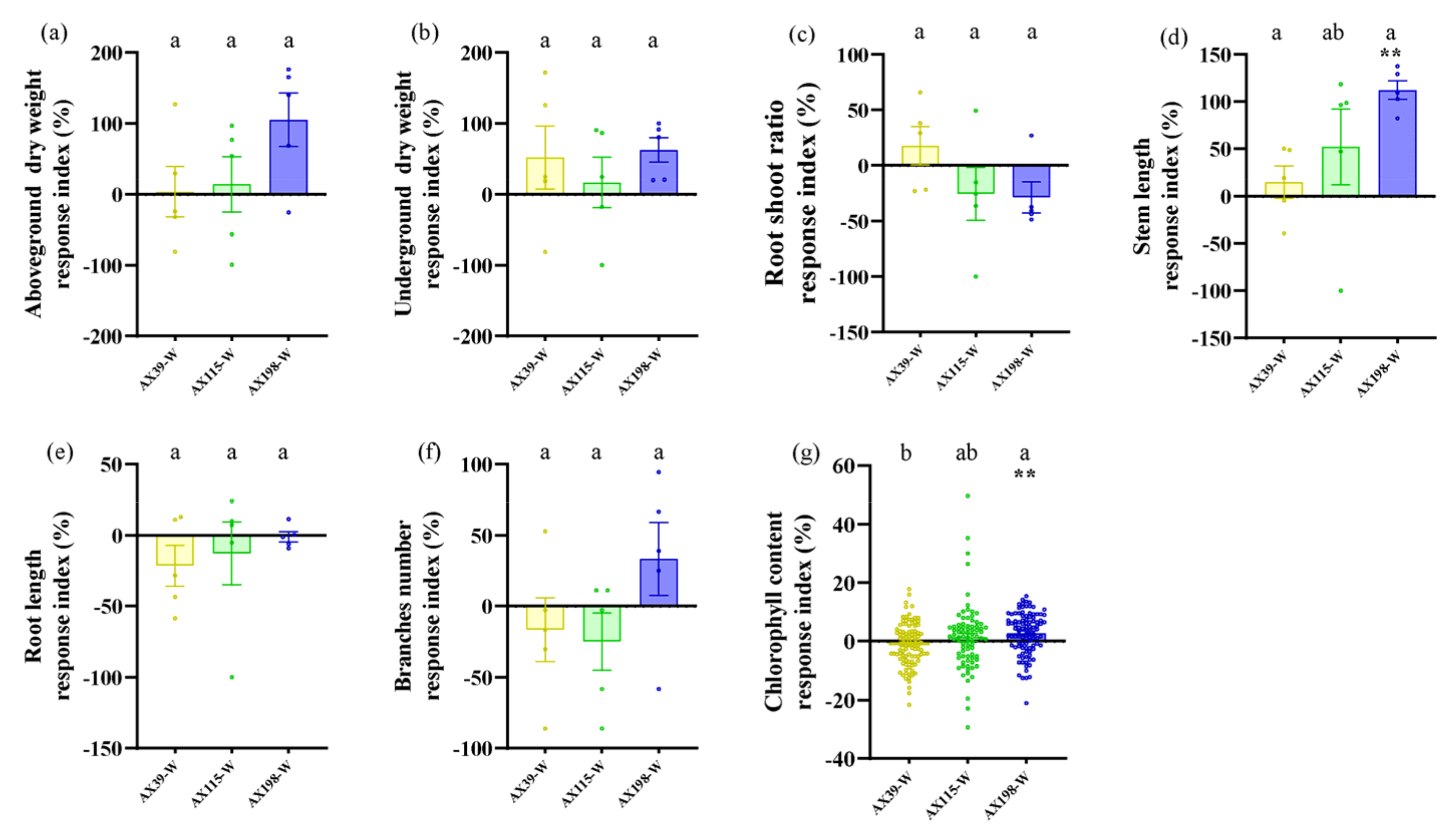
Figure 7.
Biomass and chlorophyll content of A. adenophora plants inoculated with endophytic Colletotrichum strains under normal conditions and nutrient stress conditions. (a) Aboveground parts, (b) underground parts, (c) root shoot ratio, (d) stem length, (e) root length, (f) branch number, and (g) chlorophyll content. A positive RI indicates an increased biomass of A. adenophora in the nutrient stress (-N) treatment with Colletotrichum strain (AX39, AX115, or AX198) inoculation compared with that without Colletotrichum inoculation. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each nutrient stress treatment and the normal treatment (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to show that the differences in the RIs were significant among the treatments inoculated with strains AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
Figure 7.
Biomass and chlorophyll content of A. adenophora plants inoculated with endophytic Colletotrichum strains under normal conditions and nutrient stress conditions. (a) Aboveground parts, (b) underground parts, (c) root shoot ratio, (d) stem length, (e) root length, (f) branch number, and (g) chlorophyll content. A positive RI indicates an increased biomass of A. adenophora in the nutrient stress (-N) treatment with Colletotrichum strain (AX39, AX115, or AX198) inoculation compared with that without Colletotrichum inoculation. Nonparametric Mann-Whitney U tests or independent sample T tests were used to identify the differences between each nutrient stress treatment and the normal treatment (* < 0.05, ** < 0.01, *** <0.001). Post hoc comparisons were performed via Duncan’s test for equal variance and Dunnett’s test for unequal variance (T3 test) to show that the differences in the RIs were significant among the treatments inoculated with strains AX39, AX115 and AX198, with different letters indicating significant differences. The error bars are the standard errors.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



