
Submitted:
08 October 2024
Posted:
10 October 2024
You are already at the latest version
Abstract
Bacterial vectors for biomolecule delivery to targeted organelles, facilitating temporary or continuous protein production, have emerged as a promising approach for treating acquired and inherited diseases. This method offers a selective cancer eradication and targeting strategy with minimal side effects. Bacterial vectors provide an alternative to viral gene delivery, given their capacity to deliver large genetic materials while inducing minimal immunogenicity and cytotoxicity. Bacteria such as Bifidobacterium, Salmonella, and Clostridium have demonstrated potential for tumor-targeted biomolecule delivery. These vectors have also been used to transfer and amplify genes encoding biomolecules such as prodrug-converting enzymes, toxins, angiogenesis inhibitors, and cytokines. The microenvironment of necrotic tumors offers a unique opportunity for targeted therapy with the non-pathogenic anaerobic bacterium. For example, Clostridium sporogenes can germinate selectively in the necrotic regions upon injection as endospores, which helps to enhance the specificity of Clostridium sporogenes, resulting in tumor-specific colonization. Further, some invasive bacteria vectors engineered with the invasin gene from Yersinia Pseudotuberculosis, which enables specific binding to integrin receptors abundantly expressed in cancer cells and escape the endosome with incorporation of the Listeriolysin O gene from Listeria monocytogenes, have shown potential for application in cancer therapy. This review provides an overview of the use of the bacteria vector in cancer therapy, highlighting strategies to maximize delivery efficiency and address potential challenges.
Keywords:
Bacteria vector
; Cancer therapy
; Invasive E. coli
; Gene therapy
; Protein
; Cargo
; Vaccination
1. Introduction
Bacterial vectors for biomolecule delivery to targeted organelles, facilitating temporary or continuous protein production, have emerged as a promising approach for treating acquired and inherited diseases [1]. These vectors can be used to deliver controlled and sustained anti-cancer agents into tumor microenvironments singly or combined with other forms of therapy [2]. Bacteria vectors offer an attractive alternative to viral vectors due to their ability to deliver sizeable genetic material, proteins, and other forms of nucleic acids [3]. Bacteria vectors induce minimal immunogenicity and cytotoxicity, potentially high gene transfer efficiency, broad host range, rare re-arrangements in the delivered DNA, and safety since for gene transfer to occur, the donor bacteria have to die [4,5]. The tumor microenvironment provides an enabling environment for the survival and thriving of these bacteria vectors for selective targeting [6].
Although the efficiency and selectivity of these bacteria vectors for cargo delivery need to be optimized, bacteria such as Bifidobacterium [7], Salmonella [8], and Clostridium [9] have demonstrated the potential for tumor-targeting by preferentially replicating within solid tumors when injected from a distal site [10,11]. These vectors have also been used to transfer and amplify genes encoding biomolecules such as prodrug-converting enzymes, toxins, angiogenesis inhibitors, and cytokines [12,13,14]. Attenuated S. Typhimurium vectors represent the most widely applied bacterial vectors at the clinical trial level due to their ability to selectively colonize tumors to inhibit their growth and prolong survival after systemic infection [15].
Cancer is a many-sided disease, and combining other forms of therapies with bacteria therapy can help achieve synergistic effects by improving penetration limitations and maximizing the activities of other therapies while reducing systemic toxicity to the host [16]. Using clostridial species (Clostridium histolyticum), spore for targeted tumor killing by direct injection into the transplanted sarcomas of mice and subsequent vegetative growth of the organism resulted in significant tumor lysis and those mice that were co-treated with penicillin and antitoxin, an extended survival rate was observed compared to tumor-bearing mice that were not injected with clostridial spores [17].
Furthermore, the microenvironment of necrotic tumors offers a unique opportunity for targeted therapy with the non-pathogenic anaerobic bacterium Clostridium sporogenes, enabling it to germinate selectively in necrotic regions upon injection as endospores, which helps to enhance the specificity of Clostridium sporogenes resulting in tumor-specific colonization [18]. Here, we discuss the potential of an invasive bacteria vector engineered to invade mammalian cells, with the ability to bind specifically with β1 integrin receptors, which are abundantly expressed in cancer cells [19,20]. For example, E. coli DH10B is incorporated with genes from invasin gene from Yersinia Pseudotuberculosis and Listeriolysin O gene from Listeria monocytogenes, which facilitate the cellular entry and endosomal escape, respectively, before the cytosolic release of cargoes [21]. This bacteria vector is a diaminopimelic acid auxotroph, resulting in their suicidal tendency, which helps reduce the risk of systemic infection or organ colonization after the delivery of the therapeutic cargo [22]. The invasive bacteria vector will offer an exciting alternative to eliminating cancer cells, especially the cells in the core region of the tumor that are resistant to other forms of therapy [23,24]. The bacteria vector can deliver therapeutic genes, convert non-toxic pro-drugs to highly toxic metabolites, or perform bacteria immunotherapy. The bacteria can either be used as monotherapy or complementarily with other forms of therapy. This review highlights the potential of the bacteria vector for cancer therapy, mentioning the strategies to maximize delivery efficiency and potential challenges.
2. Engineered Bacteria Experimental Studies
Our laboratory and others have established the quantification of internalized E. coli vectors, optimized the time the vector can be wholly internalized into eukaryotic cells, and described its trafficking through the lysosomal and autophagy pathways [5,21,22]. E. coli invasion into eukaryotic cells is a pre-requisite to determining the molecular mechanisms of how it functions to obtain insights for improving its efficiency [25].
Our previous experiments showed high cellular infection of up to 70.47%, 27.4%, and 26.2% in MCF-7, A549, and HEK-293 cells, respectively. The transfection efficiency of E. coli vectors in vitro needs to be improved to harness it for cancer therapy. The efficiency of E. coli is considerably low compared to its viral counterpart due to several cellular barriers, including cellular uptake of the vector, endosomal escape, and nuclear entry [26]. To drastically improve E. coli payload delivery efficiency, we have combined it with other commercially available chemical vectors. For example, lipofectamine increases E. coli vector gene delivery efficiency up to 2.8 folds in HeLa cells [27]. Further, since endosomal escape is one of the crucial barriers that need to be overcome by integrin-mediated vector, we demonstrated in one of our works that the Inhibition of lysosomal V-ATPase enhances E. coli bactofection by 6.9, 3.2, 5.0, 2.8, and 4.5-fold in HeLa, HEK-293, A549, HT1080, and MCF-7 respectively, compared to untreated cells [28].
Recent studies improved the E. coli vector gene delivery capabilities by non-covalent coupling with cell-penetrating peptides and elucidating the interaction to form a hybrid vector using atomic force microscopy (Unpublished) and combining the sophistication and real-time possibility of imaging flow cytometry for E. coli tracking and gene delivery in cancer cells. The imaging flow cytometry combines the features of fluorescent microscopy and laser scanning cytometry, enabling the acquisition and identification of a heterogeneous population of cells [29,30]. The red fluorescing E. coli vector DH10B asd− (pGB2Ωinv-hly, pUltraRFPKM) is a useful molecular tool for tracking studies to understand better and unveil the molecular mechanism of E. coli vector and navigation through the endosomal and autophagy pathways of cancer cells [25,31].
However, extensive studies are required on the E. coli therapeutics cargo delivery into cancer cells both in vitro and in vivo for E. coli to make it to clinical trials like its counterparts, such as Salmonella Typhimurium [32,33,34], Listeria monocytogenes and Clostridium sp. Understanding how the gut microbiota changes upon anti-cancer therapeutics delivery will be pertinent, as the possibility that the host’s resident flora affects the host tumor’s susceptibility to cancer therapies.
3. Gut Microbiome and Anti-Cancer Payload Delivery
Over the last decade, evidence has linked microbiota with the physiology and function of the host. Human microbiomes consist of complex communities of bacteria, yeast, fungi, protozoa, archaea, and viruses that inhabit the surface barriers of the human body [35,36]. These microorganisms comprise the human microbiota while collecting these microorganism’s genomes makes up the human microbiome [37]. The gut microbiome dysfunction can lead to many diseases, such as inflammation, cancer, and neurodegenerative disease [38,39,40]. Overwhelming evidence has suggested the link between commensal microbiota and cancer. Gastrointestinal microbiota plays a pivotal role in modulating responses to cancer immunotherapy, and some data demonstrates that the microbial community within the tumor microenvironment can contribute to therapeutic efficacy [37,41,42]. Therefore, it is pertinent to understand how the gut microbiome would respond to therapeutic cargo delivery into cancer cells and how both can be used synergically to alleviate cancer. Previous reports have shown that gut microbiota may shape responses to forms of cancer therapy through effect on multiple metabolic pathways [43].
The emergence of metagenomics and transcriptomics sequencing of diverse cell populations to quantify the microbial community has helped shed more light on the gut microbiome composition, function, and dynamics [44]. However, the basic mechanistic understanding of the individual genetic factor that drives the overall function of the gut microbiota needs to be adequately understood. These will give us better insights into harnessing the gut microbiome for anti-cancer benefits. There are still questions about whether targeting the gut and tumor microbiotas will be a better option by delivering therapeutic cargo to remove or populate specific beneficial or harmful bacteria within host-associated microbial communities (Figure. 1).
Figure 1.
A proposed therapeutic approach targeting the gut microbiome. This approach delivers an efficacious therapeutic payload against bacteria that produce toxins, genotoxin, metabolites, and reactive oxygen species in the gut. This inflammation promotes tumor growth and development. By reducing the populations of these bacteria, healthy bacteria will be promoted, resulting in the shrinking of the tumor. For example, some bacteria species stimulate an inflammatory response that supports carcinogenesis by producing genotoxic metabolites by Bacteroides fragilis, E. coli, and Campylobacter jejuni, respectively. Bacteria, especially part of the residence flora, can be engineered to produce anti-genotoxic substances that reduce the bacteria responsible for this substance and promote gut-friendly bacteria.
Figure 1.
A proposed therapeutic approach targeting the gut microbiome. This approach delivers an efficacious therapeutic payload against bacteria that produce toxins, genotoxin, metabolites, and reactive oxygen species in the gut. This inflammation promotes tumor growth and development. By reducing the populations of these bacteria, healthy bacteria will be promoted, resulting in the shrinking of the tumor. For example, some bacteria species stimulate an inflammatory response that supports carcinogenesis by producing genotoxic metabolites by Bacteroides fragilis, E. coli, and Campylobacter jejuni, respectively. Bacteria, especially part of the residence flora, can be engineered to produce anti-genotoxic substances that reduce the bacteria responsible for this substance and promote gut-friendly bacteria.
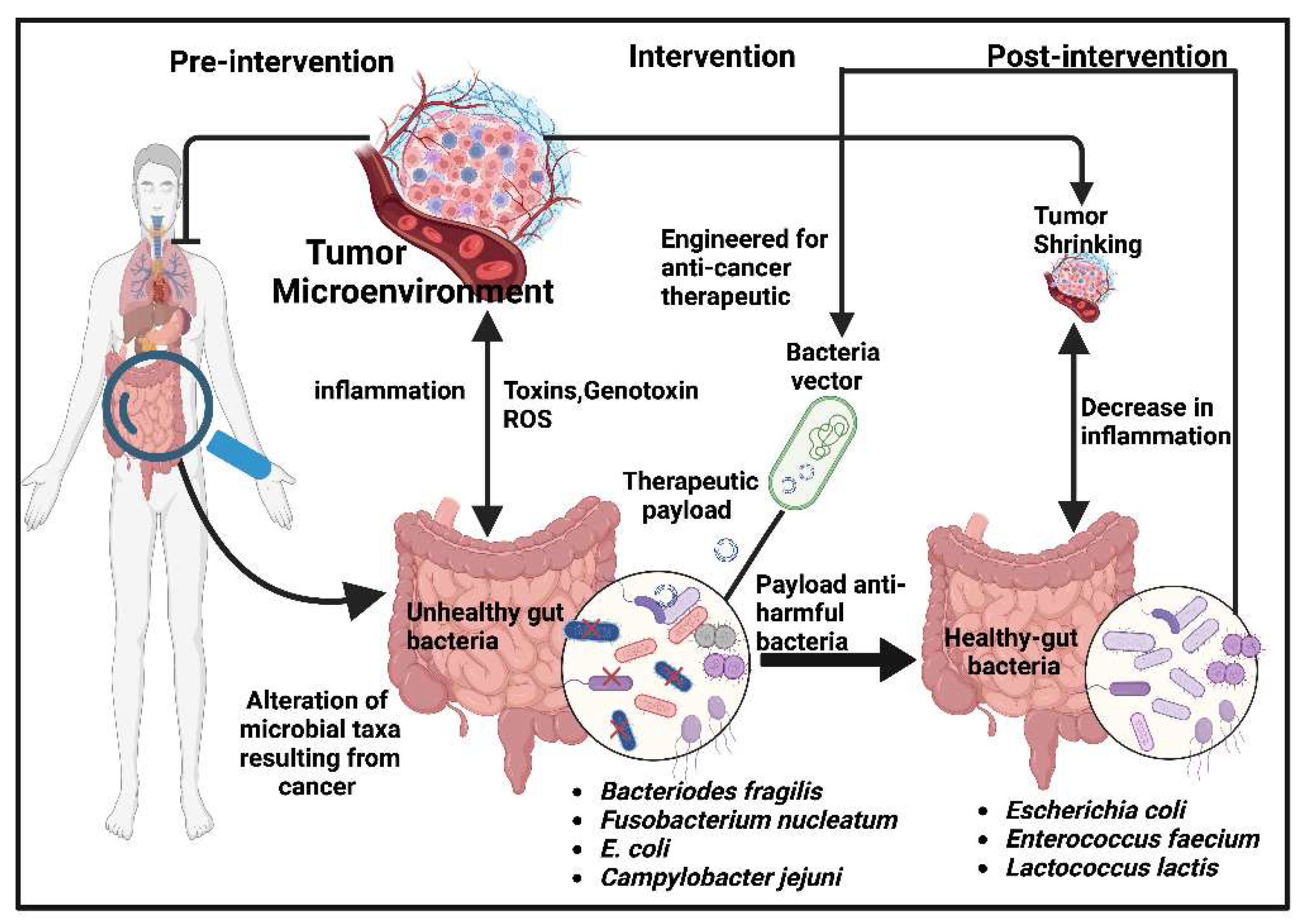
Bacteria species stimulate an inflammatory response that promotes carcinogenesis via toxins produced by Bacteroides fragilis, increasing the production of reactive oxygen species and leading to aberrant signaling pathways in human and mouse tumors, which may prevent antitumor immune function [45]. Further, some bacteria produce genotoxic metabolites, such as colibactin from E. coli, and Campylobacter jejuni produces cytolethal toxin, which induces carcinogenicity in mice [38,46,47]. There is striking evidence that F. nucleatum plays a pivotal role in the development and progression of colon adenomas and colon cancer and has been found in nodal and distal metastasis. On the contrary, some bacteria taxa have been associated with good gut health, including Firmicutes, Bacteroides, Lactobacillus, and Enterococcus [48]. Genetically engineered bacteria have the potential to modulate gut microbiota (Figure. 1) or host physiology by secreting proteins, such as human interleukin-10, to reduce inflammation.
4. Strategies for the Application of Invasive Bacteria in Cancer Therapy
The conventional therapeutic approach for human malignancy, such as radiotherapies, chemotherapies, and surgery, presents significant health limitations, including but not limited to poor tumor-specific targeting, significant adverse effects, insufficient tumor permeability, rapid tumor relapse, and metastasis [49]. However, cancer immunotherapy has appeared as a promising option to promote the recognition and elimination of tumor cells by stimulating the immune system through the induction of innate and adaptive responses with different treatment strategies [50].
These responses capacitate the immune system to eliminate or protect against several tumors by releasing pro-inflammatory cytokines. Immune therapies such as checkpoint inhibitors, monoclonal anti-cytotoxic T-Lymphocyte associated protein 4 (CTL-4), and programmed death protein 1 (PD1) are now being used clinically, with inspiring results [51,52,53,54,55]. However, immune therapy may decrease immunotolerance, resulting in immune-related adverse events and kidney-related toxicity, which limits their use.
To address these shortcomings, bacteria-mediated anti-tumor therapy provides an alternative option. Bacteria’s unique characteristics include the ability to destroy tumor cells from the inside and subsequently induce innate and adaptive antitumor immune responses, which help to eliminate tumor cells effectively [56]. Interestingly, compared to most other therapeutics, the efficacy of tumor-targeting bacteria is independent of the tumor’s genetic makeup.
Invasive bacteria vectors can be used in different ways in cancer therapy. One approach is to use bacteria to deliver therapeutic genes into cancer cells. These have been used in in vivo studies in phagocytotic and non-phagocytic cells with modified E. coli BM2710/pGB2Ωinv-hly carrying pC1ΩTGF-β1. The engineered E. coli vector was used to deliver a therapeutic gene (pC1ΩTGF-β1) into the intestinal mucosa through oral administration, significantly reducing the severity of experimental colitis in mice [57]. Further, the re-introduction of a deficient gene sensitizes tumor cells to other chemotherapeutic agents. Hepatocellular Carcinoma (HCCs) are generally highly resistant to chemotherapeutic agents and radiotherapy. It is believed that molecular changes during carcinogenesis, such as overexpression of the multidrug resistance gene and the loss of tumor suppressor gene p53, may allow tumor cell populations to become resistant to most therapeutic approaches. Using bacteria as a vector, the wild-type p53 gene can be reintroduced into HCCs to sensitize it to a chemotherapeutic agent [58].
Current research has focused on identifying new pro-drug activating enzymes that produce highly toxic and freely diffusible metabolites. The expression of bacterial enzymes in tumor cells to convert non-toxic pro-drugs to highly toxic metabolites with anti-cancer effects. E. coli DH5a-lux/bG can hydrolyze glucuronide substrates and emit luminescence. The bG and the luxCDABE gene cluster for prodrug activation, luminance emission, and specific conversion of the glucuronide prodrug 9ACG to the topoisomerase I poison 9-amino camptothecin (9AC). The bacteria localize and replicate in human tumor xenografts and produce substantial antitumor activity in combination with systemic 9ACG prodrug therapy [59]. Cytosine deaminase converts non-toxic 5-fluorocytosine (5-FU) into the chemotherapeutic agent 5-fluorouracil (5-FU). 5-FU is highly toxic because it is further metabolized into product that infers with DNA and RNA synthesis. Upon administration of the S. Typhimurium (VNP20009) strain, cytosine deaminase and 5-FU were expressed in patients, and the conversion of 5-FC to 5-FU indicated the bacterial production of functional CD in the tumor [60].
The use of bacteria in cancer immunotherapy has been demonstrated by several studies with bacteria preferentially growing within the tumor cores due to the immune-privileged nature and the often hypoxic and necrotic tumor microenvironment with its possibility of locally affecting tumor growth through the recruitment and activation of the host’s immune system [61,62,63]. The cells in this hypoxic region of the necrotic tumor microenvironment are shown to be resistant to chemotherapy and radiotherapy, which provides the opportunity for selective targeting with bacteria [64,65]. In a previous study, an engineered non-pathogenic Escherichia coli strain specifically lyses within the tumor microenvironment. It releases an encoded nanobody antagonist of CD47 (CD47nb)12, an anti-phagocytic receptor commonly overexpressed in several human cancer types. Increases activation of tumor-infiltrating T cells and stimulates rapid tumor regression, preventing metastasis and leading to long-term survival in a syngeneic tumor model in mice. Also, the local injection of CD47nb-expressing bacteria stimulates systemic tumor-antigen-specific immune responses that reduce the growth of untreated tumors [66].
The invasive bacteria vector will offer an exciting alternative to eliminating cancer cells, especially the cells in the core region of the tumor that are resistant to other forms of therapy. The bacteria vector can deliver therapeutic genes, convert non-toxic pro-drugs to highly toxic metabolites, or perform bacteria immunotherapy. The bacteria can either be used as monotherapy or complementarily with other forms of therapy.
Figure 2.
Strategies of tumor treatment, both monotherapy and complementary therapy. A. Bacteria vectors including E. coli, Salmonella Typhimurium, Clostridium sp, and Bifidobacterium sp. B) The tumor microenvironment is a hypoxic region of the necrotic tumor microenvironment resistant to chemotherapy and radiotherapy, allowing selective targeting with bacteria. Immunotherapy, on the other hand, results in decreased immunotolerance and other immune-related adverse events and kidney-related toxicity, which limits their use.
Figure 2.
Strategies of tumor treatment, both monotherapy and complementary therapy. A. Bacteria vectors including E. coli, Salmonella Typhimurium, Clostridium sp, and Bifidobacterium sp. B) The tumor microenvironment is a hypoxic region of the necrotic tumor microenvironment resistant to chemotherapy and radiotherapy, allowing selective targeting with bacteria. Immunotherapy, on the other hand, results in decreased immunotolerance and other immune-related adverse events and kidney-related toxicity, which limits their use.
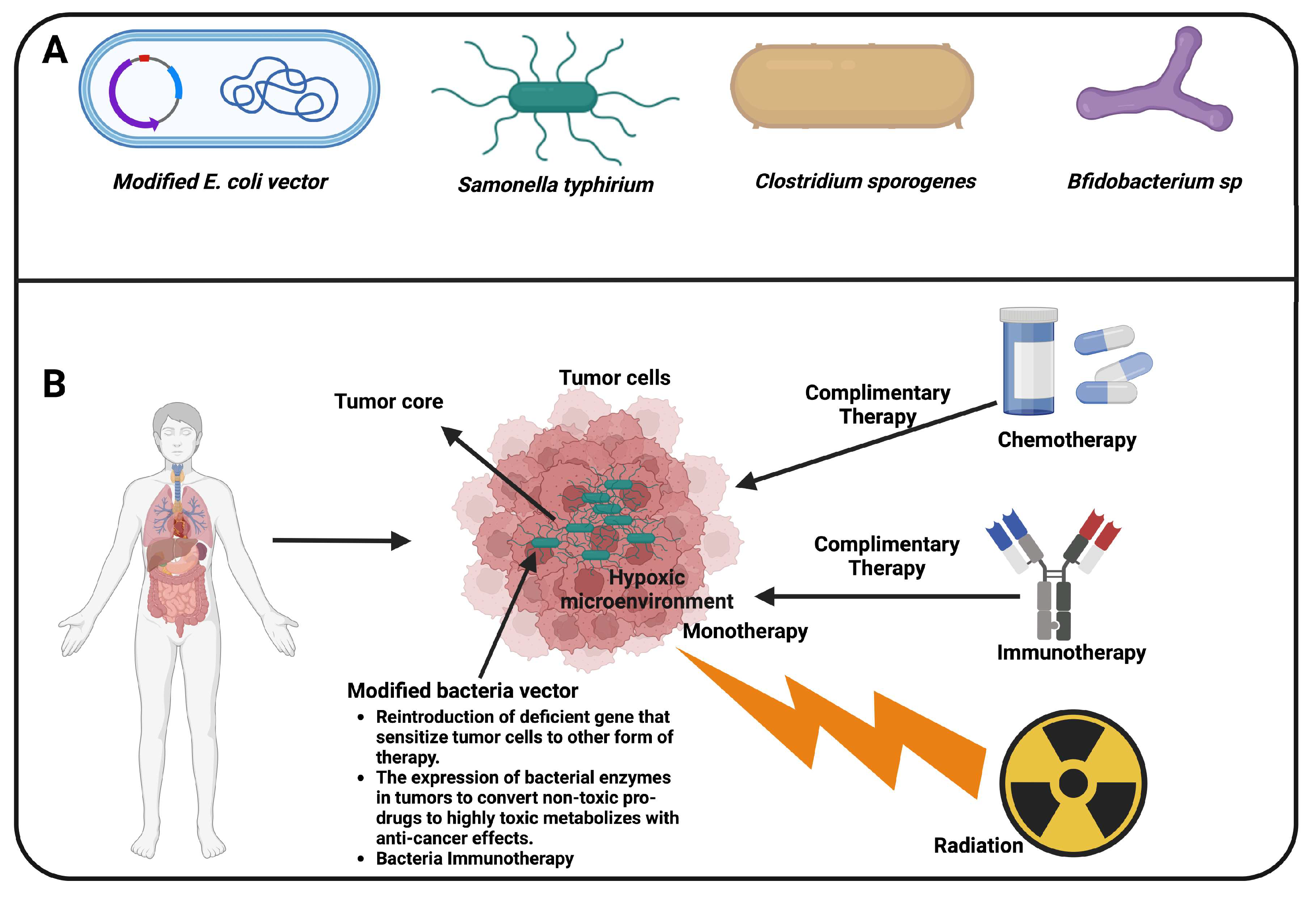
5. Perspective and Limitation, and Conclusion
Invasive bacteria used for payload delivery are primarily engineered to acquire improved anti-tumor activities, therapeutic index, and safety [67,68]. The deletion of significant virulence genes is often required to minimize their pathogenicity. The deletion of the lipopolysaccharide (LPS) in the outer membrane of the E. coli vector makes it diaminopimelic acid (DAP) autotroph, which helps to prevent the stimulation of tumor necrosis factor (TNF) and shock in gram-negative sepsis [25]. These benefits include an automatic self-targeting ability and the possibility of genetic manipulation to produce newly engineered attenuated strains [69]. Nevertheless, invasive bacteria for anti-cancer treatment have not yet been clinically established and require more research before their use in cancer treatment.
Interestingly, one of the uniquely safe attributes of bacterial vectors is their sensitivity to clinically available antibiotic treatments, enabling their control post-administration, an invaluable property for safe gene therapy. However, despite the potential for bacterial delivery systems, it is crucial to highlight their potential adverse side effects and the need to optimize their delivery into the targeted organ. Recombinant bacteria in humans must be carefully controlled and ascertained to prevent lateral gene transfer to the resident bacteria and limit the vector’s environmental spread [70].
The specificity of bacteria vectors, such as E. coli [19,22,25], Salmonella typhimurium [71,72], and Shigella flexneri [73], to bind specifically to integrin receptors for delivery of therapeutic cargo into cancer cells is one unique characteristic that makes them candidates for cancer gene therapy [74]. The significant expression of integrin on cancer cells provides a focal point for therapeutic cargo delivery into cancer cells. Integrin is a cell adhesion receptor that plays a crucial role in cell proliferation, migration, and survival [75,76]. For example, evidence has shown that β1 integrin receptors were significantly expressed in melanoma [77], Ovarian [78,79], and non-small-cell lung carcinoma [80], which could be explored to deliver plasmid DNA, pro-drug, mRNA, or protein for cancer therapy.
Furthermore, cancer therapy with engineered bacteria provides an alternative option to meet the challenges of late-stage cancers initially insensitive to the therapies. Manipulating the gut microbiome by combining bacteria with cancer drugs may increase their effective delivery to the cancer sites. Using this strategy, the gut microbiome breaks down and metabolizes oral drugs, helping dispense and distribute them into lymphatic and blood circulation and the gut-brain axis [81]. Also, the bacteria S. Typhimurium can sense hypoxia displayed deep inside and help penetrate the tumors. However, the bacteria used as drug carriers face challenges, such as biocompatibility, motility deterioration after drug loading, and lack of in vivo verification [82].
Industrially, bacterial vectors are cheap to manufacture and practical compared to viral vectors, which are particularly cumbersome, time-consuming, and more expensive for gene therapy [83]. The necessary infrastructure and expertise already exist for bacterial vector manufacturing at low cost on an industrial scale, as bacterial culture systems have long been in operation in the biotechnology industries [84].
Overall, developing bacterial vectors with the potential to deliver therapeutic agents is an exciting area of research, but extensive work is still needed. Cargo delivery or the use of invasive bacteria in anticancer is gaining acceptance for its potential to provide positive clinical outcomes. However, more work needs to be done to improve some systems’ safety and efficacy so that this approach can yield dividends in the coming years.
Author Contributions
Conceptualization, RA., and KN.; Writing- original draft preparation., RA.; Writing-review and editing, RA and KN. The authors contributed equally.
Funding
None
Conflicts of Interest
Authors declare no conflict of interest.
References
- R. Gardlik, J.H. R. Gardlik, J.H. Fruehauf, Bacterial vectors and delivery systems in cancer therapy, IDrugs, 13 (2010) 701-706.
- B.F.-L. Sieow, K.S. B.F.-L. Sieow, K.S. Wun, W.P. Yong, I.Y. Hwang, M.W. Chang, Tweak to treat: reprograming bacteria for cancer treatment, Trends in cancer, 7 (2021) 447-464.
- A. Heggie, T.L. A. Heggie, T.L. Thurston, T. Ellis, Microbial messengers: nucleic acid delivery by bacteria, Trends in Biotechnology, (2024).
- C.K. Baban, M. C.K. Baban, M. Cronin, D. O’Hanlon, G.C. O’Sullivan, M. Tangney, Bacteria as vectors for gene therapy of cancer, Bioengineered bugs, 1 (2010) 385-394.
- I. Fajac, S. I. Fajac, S. Grosse, J.-M. Collombet, G. Thevenot, S. Goussard, C. Danel, C. Grillot-Courvalin, Recombinant Escherichia coli as a gene delivery vector into airway epithelial cells, Journal of controlled release, 97 (2004) 371-381.
- Z. Wang, W. Z. Wang, W. Sun, R. Hua, Y. Wang, Y. Li, H. Zhang, Promising dawn in tumor microenvironment therapy: engineering oral bacteria, International Journal of Oral Science, 16 (2024) 24.
- K. Yazawa, M. K. Yazawa, M. Fujimori, J. Amano, Y. Kano, S.i. Taniguchi, Bifidobacterium longum as a delivery system for cancer gene therapy: selective localization and growth in hypoxic tumors, Cancer gene therapy, 7 (2000) 269-274.
- J.H. Zheng, J.-J. J.H. Zheng, J.-J. Min, Targeted cancer therapy using engineered Salmonella typhimurium, Chonnam medical journal, 52 (2016) 173-184.
- J. Pahle, L. J. Pahle, L. Menzel, N. Niesler, D. Kobelt, J. Aumann, M. Rivera, W. Walther, Rapid eradication of colon carcinoma by Clostridium perfringens Enterotoxin suicidal gene therapy, BMC cancer, 17 (2017) 1-14.
- M. Yang, F. M. Yang, F. Yang, W. Chen, S. Liu, L. Qiu, J. Chen, Bacteria-mediated cancer therapies: opportunities and challenges, Biomaterials Science, 9 (2021) 5732-5744.
- C.-H. Lee, Engineering bacteria toward tumor targeting for cancer treatment: current state and perspectives, Applied microbiology and biotechnology, 93 (2012) 517-523.
- J.M. Pawelek, K.B. J.M. Pawelek, K.B. Low, D. Bermudes, Bacteria as tumour-targeting vectors, The lancet oncology, 4 (2003) 548-556.
- S. Zhou, C. S. Zhou, C. Gravekamp, D. Bermudes, K. Liu, Tumour-targeting bacteria engineered to fight cancer, Nature Reviews Cancer, 18 (2018) 727-743.
- D. Lin, X. D. Lin, X. Feng, B. Mai, X. Li, F. Wang, J. Liu, X. Liu, K. Zhang, X. Wang, Bacterial-based cancer therapy: An emerging toolbox for targeted drug/gene delivery, Biomaterials, 277 (2021) 121124.
- I.Y. Lin, T.T.H. I.Y. Lin, T.T.H. Van, P.M. Smooker, Live-attenuated bacterial vectors: tools for vaccine and therapeutic agent delivery, Vaccines, 3 (2015) 940-972.
- R.B. Mokhtari, T.S. R.B. Mokhtari, T.S. Homayouni, N. Baluch, E. Morgatskaya, S. Kumar, B. Das, H. Yeger, Combination therapy in combating cancer, Oncotarget, 8 (2017) 38022.
- N.P. Minton, Clostridia in cancer therapy, Nature Reviews Microbiology, 1 (2003) 237-242.
- J.R. Möse, G. J.R. Möse, G. Möse, Oncolysis by clostridia. I. Activity of Clostridium butyricum (M-55) and other nonpathogenic clostridia against the Ehrlich carcinoma, Cancer research, 24 (1964) 212-216.
- A. Osahor, K. A. Osahor, K. Deekonda, C.-W. Lee, E.U.-H. Sim, A. Radu, K. Narayanan, Rapid preparation of adherent mammalian cells for basic scanning electron microscopy (SEM) analysis, Analytical biochemistry, 534 (2017) 46-48.
- M. Larsen, U. M. Larsen, U. Griesenbach, S. Goussard, D. Gruenert, D. Geddes, R.a. Scheule, S. Cheng, P. Courvalin, C. Grillot-Courvalin, E. Alton, Bactofection of lung epithelial cells in vitro and in vivo using a genetically modified Escherichia coli, Gene therapy, 15 (2008) 434-442.
- C. Grillot-Courvalin, S. C. Grillot-Courvalin, S. Goussard, P. Courvalin, Bacteria as gene delivery vectors for mammalian cells, Horizontal Gene Transfer, (2002) 261-265.
- K. Narayanan, P.E. K. Narayanan, P.E. Warburton, DNA modification and functional delivery into human cells using Escherichia coli DH10B, Nucleic acids research, 31 (2003) e51-e51.
- D. Morrissey, G.C. D. Morrissey, G.C. O’Sullivan, M. Tangney, Tumour targeting with systemically administered bacteria, Current gene therapy, 10 (2010) 3-14.
- K.S. Allemailem, Innovative approaches of engineering tumor-targeting bacteria with different therapeutic payloads to fight cancer: A smart strategy of disease management, International Journal of Nanomedicine, (2021) 8159-8184.
- R.O. Akinsola, M. R.O. Akinsola, M. Adewoyin, C.-W. Lee, E.U.-H. Sim, K. Narayanan, RFP-based method for real-time tracking of invasive bacteria in a heterogeneous population of cells, Analytical biochemistry, 634 (2021) 114432.
- I.A. Khalil, K. I.A. Khalil, K. Kogure, H. Akita, H. Harashima, Uptake pathways and subsequent intracellular trafficking in nonviral gene delivery, Pharmacological reviews, 58 (2006) 32-45.
- K. Narayanan, C.W. K. Narayanan, C.W. Lee, A. Radu, E.U.H. Sim, Escherichia coli bactofection using Lipofectamine, Analytical biochemistry, 439 (2013) 142-144.
- R.O. Akinsola, C.W. R.O. Akinsola, C.W. Lee, E.U.H. Sim, K. Narayanan, Inhibition of lysosomal vacuolar proton pump down-regulates cellular acidification and enhances E. coli bactofection efficiency, Analytical biochemistry, 616 (2021) 114088.
- S. Dimitriadis, L. S. Dimitriadis, L. Dova, I. Kotsianidis, E. Hatzimichael, E. Kapsali, G.S. Markopoulos, Imaging Flow Cytometry: Development, Present Applications, and Future Challenges, Methods and Protocols, 7 (2024) 28.
- J.P. Robinson, R. J.P. Robinson, R. Ostafe, S.N. Iyengar, B. Rajwa, R. Fischer, Flow cytometry: the next revolution, Cells, 12 (2023) 1875.
- R.O. Akinsola, Investigating the E. coli vector trafficking through the endo-lysosomal and autophagy pathways to improve its efficiency, Monash University.
- C.R. Pangilinan, C.-H. C.R. Pangilinan, C.-H. Lee, Salmonella-based targeted cancer therapy: updates on a promising and innovative tumor immunotherapeutic strategy, Biomedicines, 7 (2019) 36.
- K.M. Broadway, B.E. K.M. Broadway, B.E. Scharf, Salmonella typhimurium as an anticancer therapy: recent advances and perspectives, Current Clinical Microbiology Reports, 6 (2019) 225-239.
- K. Liang, Q. K. Liang, Q. Liu, P. Li, H. Luo, H. Wang, Q. Kong, Genetically engineered Salmonella Typhimurium: Recent advances in cancer therapy, Cancer letters, 448 (2019) 168-181.
- E. Dekaboruah, M.V. E. Dekaboruah, M.V. Suryavanshi, D. Chettri, A.K. Verma, Human microbiome: an academic update on human body site specific surveillance and its possible role, Archives of microbiology, 202 (2020) 2147-2167.
- S. Runge, S.P. S. Runge, S.P. Rosshart, The mammalian metaorganism: a holistic view on how microbes of all kingdoms and niches shape local and systemic immunity, Frontiers in Immunology, 12 (2021) 702378.
- V. Matson, C.S. V. Matson, C.S. Chervin, T.F. Gajewski, Cancer and the microbiome—influence of the commensal microbiota on cancer, immune responses, and immunotherapy, Gastroenterology, 160 (2021) 600-613.
- Y. Chen, J. Y. Chen, J. Zhou, L. Wang, Role and mechanism of gut microbiota in human disease, Frontiers in cellular and infection microbiology, 11 (2021) 625913.
- D. Zheng, T. D. Zheng, T. Liwinski, E. Elinav, Interaction between microbiota and immunity in health and disease, Cell research, 30 (2020) 492-506.
- L.J. Spielman, D.L. L.J. Spielman, D.L. Gibson, A. Klegeris, Unhealthy gut, unhealthy brain: The role of the intestinal microbiota in neurodegenerative diseases, Neurochemistry international, 120 (2018) 149-163.
- A.M. Barbosa, A. A.M. Barbosa, A. Gomes-Gonçalves, A.G. Castro, E. Torrado, Immune system efficiency in cancer and the microbiota influence, Pathobiology, 88 (2021) 170-186.
- P. Raoul, V. P. Raoul, V. De Gaetano, G. Sciaraffia, G. Ormea, M. Cintoni, C. Pozzo, A. Strippoli, A. Gasbarrini, M.C. Mele, E. Rinninella, Gastric Cancer, Immunotherapy, and Nutrition: The Role of Microbiota, Pathogens, 13 (2024) 357.
- E. Montassier, T. E. Montassier, T. Gastinne, P. Vangay, G. Al-Ghalith, S. Bruley des Varannes, S. Massart, P. Moreau, G. Potel, M. de La Cochetière, E. Batard, Chemotherapy-driven dysbiosis in the intestinal microbiome, Alimentary pharmacology & therapeutics, 42 (2015) 515-528.
- G. Goel, T. G. Goel, T. Requena, S. Bansal, Human-Gut Microbiome: Establishment and Interactions, Academic Press2022.
- C. Xing, Y. C. Xing, Y. Du, T. Duan, K. Nim, J. Chu, H.Y. Wang, R.-F. Wang, Interaction between microbiota and immunity and its implication in colorectal cancer, Frontiers in Immunology, 13 (2022) 963819.
- F. Taieb, C. F. Taieb, C. Petit, J.-P. Nougayrède, E. Oswald, The enterobacterial genotoxins: cytolethal distending toxin and colibactin, EcoSal plus, 7 (2016) 10.1128/ecosalplus. ESP-0008-2016.
- K. Hartl, M. K. Hartl, M. Sigal, Microbe-driven genotoxicity in gastrointestinal carcinogenesis, International Journal of Molecular Sciences, 21 (2020) 7439.
- S. Vivarelli, R. S. Vivarelli, R. Salemi, S. Candido, L. Falzone, M. Santagati, S. Stefani, F. Torino, G.L. Banna, G. Tonini, M. Libra, Gut microbiota and cancer: from pathogenesis to therapy, Cancers, 11 (2019) 38.
- H.H. Chen, M.T. H.H. Chen, M.T. Kuo, Improving radiotherapy in cancer treatment: Promises and challenges, Oncotarget, 8 (2017) 62742.
- A.R. Kumar, A.R. A.R. Kumar, A.R. Devan, B. Nair, B.S. Vinod, L.R. Nath, Harnessing the immune system against cancer: current immunotherapy approaches and therapeutic targets, Molecular biology reports, (2021) 1-21.
- W. Torres, V. W. Torres, V. Lameda, L.C. Olivar, C. Navarro, J. Fuenmayor, A. Pérez, A. Mindiola, M. Rojas, M.S. Martínez, M. Velasco, Bacteria in cancer therapy: beyond immunostimulation, J. Cancer Metastasis Treat, 4 (2018).
- P.S. Hegde, D.S. P.S. Hegde, D.S. Chen, Top 10 challenges in cancer immunotherapy, Immunity, 52 (2020) 17-35.
- R. Franzin, G.S. R. Franzin, G.S. Netti, F. Spadaccino, C. Porta, L. Gesualdo, G. Stallone, G. Castellano, E. Ranieri, The use of immune checkpoint inhibitors in oncology and the occurrence of AKI: where do we stand?, Frontiers in immunology, 11 (2020) 574271.
- S. Tan, D. S. Tan, D. Li, X. Zhu, Cancer immunotherapy: Pros, cons and beyond, Biomedicine & Pharmacotherapy, 124 (2020) 109821.
- M.A. Perazella, A.C. M.A. Perazella, A.C. Shirali, Immune checkpoint inhibitor nephrotoxicity: what do we know and what should we do?, Kidney International, 97 (2020) 62-74.
- Y. Liu, L. Y. Liu, L. Niu, N. Li, Y. Wang, M. Liu, X. Su, X. Bao, B. Yin, S. Shen, Bacterial-Mediated Tumor Therapy: Old Treatment in a New Context, Advanced Science, 10 (2023) 2205641.
- I. Castagliuolo, E. I. Castagliuolo, E. Beggiao, P. Brun, L. Barzon, S. Goussard, R. Manganelli, C. Grillot-Courvalin, G. Palu, Engineered E. coli delivers therapeutic genes to the colonic mucosa, Gene Therapy, 12 (2005) 1070-1078.
- L. Mohr, S. L. Mohr, S. Shankara, S.K. Yoon, T.U. Krohne, M. Geissler, B. Roberts, H.E. Blum, J.R. Wands, Gene therapy of hepatocellular carcinoma in vitro and in vivo in nude mice by adenoviral transfer of the Escherichia coli purine nucleoside phosphorylase gene, Hepatology, 31 (2000) 606-614.
- C. Cheng, Y. C. Cheng, Y. Lu, K. Chuang, W. Hung, J. Shiea, Y. Su, C. Kao, B. Chen, S. Roffler, T. Cheng, Tumor-targeting prodrug-activating bacteria for cancer therapy, Cancer gene therapy, 15 (2008) 393-401.
- J. Nemunaitis, C. J. Nemunaitis, C. Cunningham, N. Senzer, J. Kuhn, J. Cramm, C. Litz, R. Cavagnolo, A. Cahill, C. Clairmont, M. Sznol, Pilot trial of genetically modified, attenuated Salmonella expressing the E. coli cytosine deaminase gene in refractory cancer patients, Cancer gene therapy, 10 (2003) 737-744.
- J. Jiang, Y. J. Jiang, Y. Huang, Z. Zeng, C. Zhao, Harnessing engineered immune cells and bacteria as drug carriers for cancer immunotherapy, ACS nano, 17 (2023) 843-884.
- J.M. Brown, W.R. J.M. Brown, W.R. Wilson, Exploiting tumour hypoxia in cancer treatment, Nature Reviews Cancer, 4 (2004) 437-447.
- D.-H. Nguyen, A. D.-H. Nguyen, A. Chong, Y. Hong, J.-J. Min, Bioengineering of bacteria for cancer immunotherapy, nature communications, 14 (2023) 3553.
- D.C. Singleton, A. D.C. Singleton, A. Macann, W.R. Wilson, Therapeutic targeting of the hypoxic tumour microenvironment, Nature reviews Clinical oncology, 18 (2021) 751-772.
- Z. Chen, F. Z. Chen, F. Han, Y. Du, H. Shi, W. Zhou, Hypoxic microenvironment in cancer: molecular mechanisms and therapeutic interventions, Signal transduction and targeted therapy, 8 (2023) 70.
- S. Chowdhury, S. S. Chowdhury, S. Castro, C. Coker, T.E. Hinchliffe, N. Arpaia, T. Danino, Programmable bacteria induce durable tumor regression and systemic antitumor immunity, Nature medicine, 25 (2019) 1057-1063.
- A. Elsherbeny, H. A. Elsherbeny, H. Bayraktutan, U.C. Oz, C. Moloney, J.C. Ashworth, A.M. Grabowska, C. Alexander, Responsive Nanomaterial Delivery Systems for Pancreatic Cancer Management, Advanced Therapeutics, 7 (2024) 2300330.
- X. Xie, Y. X. Xie, Y. Zhang, F. Li, T. Lv, Z. Li, H. Chen, L. Jia, Y. Gao, Challenges and opportunities from basic cancer biology for nanomedicine for targeted drug delivery, Current cancer drug targets, 19 (2019) 257-276.
- K. Mugwanda, S. K. Mugwanda, S. Hamese, W.F. Van Zyl, E. Prinsloo, M. Du Plessis, L.M. Dicks, D.B. Thimiri Govinda Raj, Recent advances in genetic tools for engineering probiotic lactic acid bacteria, Bioscience Reports, 43 (2023) BSR20211299.
- E. Skippington, M.A. E. Skippington, M.A. Ragan, Lateral genetic transfer and the construction of genetic exchange communities, FEMS microbiology reviews, 35 (2011) 707-735.
- A. Chabloz, J.V. A. Chabloz, J.V. Schaefer, I. Kozieradzki, S.J. Cronin, D. Strebinger, F. Macaluso, J. Wald, T.H. Rabbitts, A. Plückthun, T.C. Marlovits, Salmonella-based platform for efficient delivery of functional binding proteins to the cytosol, Communications biology, 3 (2020) 342.
- M.N. Dharmasena, C.M. M.N. Dharmasena, C.M. Feuille, C.E.C. Starke, A.A. Bhagwat, S. Stibitz, D.J. Kopecko, Development of an acid-resistant Salmonella Typhi Ty21a attenuated vector for improved oral vaccine delivery, PloS one, 11 (2016) e0163511.
- A. Phalipon, P. A. Phalipon, P. Sansonetti, Live AttenuatedShigella flexneriMutants as Vaccine Candidates Against Shigellosis and Vectors for Antigen Delivery, Biologicals, 23 (1995) 125-134.
- S. Bashiardes, T. S. Bashiardes, T. Tuganbaev, S. Federici, E. Elinav, The microbiome in anti-cancer therapy, Seminars in immunology, Elsevier, 2017, pp. 74-81.
- J.S. Desgrosellier, D.A. J.S. Desgrosellier, D.A. Cheresh, Integrins in cancer: biological implications and therapeutic opportunities, Nature Reviews Cancer, 10 (2010) 9.
- P. Majumder, Integrin-mediated delivery of drugs and nucleic acids for anti-angiogenic cancer therapy: Current landscape and remaining challenges, Bioengineering, 5 (2018) 76.
- J. Nip, H. J. Nip, H. Shibata, D.J. Loskutoff, D.A. Cheresh, P. Brodt, Human melanoma cells derived from lymphatic metastases use integrin alpha v beta 3 to adhere to lymph node vitronectin, The Journal of clinical investigation, 90 (1992) 1406-1413.
- J.K. Slack-Davis, K.A. J.K. Slack-Davis, K.A. Atkins, C. Harrer, E.D. Hershey, M. Conaway, Vascular cell adhesion molecule-1 is a regulator of ovarian cancer peritoneal metastasis, Cancer research, 69 (2009) 1469-1476.
- C.N. Landen, T.-J. C.N. Landen, T.-J. Kim, Y.G. Lin, W.M. Merritt, A.A. Kamat, L.Y. Han, W.A. Spannuth, A.M. Nick, N.B. Jennnings, M.S. Kinch, Tumor-selective response to antibody-mediated targeting of αvβ3 integrin in ovarian cancer, Neoplasia, 10 (2008) 1259-1267.
- M. Adachi, T. M. Adachi, T. Taki, M. Higashiyama, N. Kohno, H. Inufusa, M. Miyake, Significance of integrin α5 gene expression as a prognostic factor in node-negative non-small cell lung cancer, Clinical cancer research, 6 (2000) 96-101.
- T. Hillman, Bacteriobot Drug-Liposome Carriers: An Optimization of Cancer-Drug Delivery to the Colon by Manipulating the Gut Microbiome, Nanoparticle, 1 (2019) 1-10.
- A. Sousa, A.N. A. Sousa, A.N. Phung, N. Škalko-Basnet, S. Obuobi, Smart delivery systems for microbial biofilm therapy: dissecting design, drug release and toxicological features, Journal of Controlled Release, 354 (2023) 394-416.
- J. El Andari, D. J. El Andari, D. Grimm, Production, processing, and characterization of synthetic AAV gene therapy vectors, Biotechnology journal, 16 (2021) 2000025.
- T.A. Beacham, J.B. T.A. Beacham, J.B. Sweet, M.J. Allen, Large scale cultivation of genetically modified microalgae: a new era for environmental risk assessment, Algal Research, 25 (2017) 90-100.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



