
Submitted:
10 October 2024
Posted:
11 October 2024
You are already at the latest version
Abstract
Sphingosine-1-phosphate (S1P) is a sphingolipid metabolic product produced via the phosphorylation of sphingosine by sphingosine kinases (SPHKs), serving as a powerful modulator of various cellular processes through its interaction with S1P receptors (S1PRs). Currently, the incompletely understood of mechanism in pancreatic diseases including pancreatitis and pancreatic cancer, largely limits therapeutic therapy options for these disorders. Recent evidence indicates that S1P significantly contributes to pancreatic diseases by modulating inflammation, promoting pyroptosis in pancreatic acinar cells, regulating the activation of pancreatic stellate cells, and affecting organelle functions in pancreatic cancer cells. Nevertheless, no review has encapsulated these advancements. Thus, this review compiles information about the involvement of S1P signaling in exocrine pancreatic disorders, including acute pancreatitis, chronic pancreatitis, and pancreatic cancer, as well as prospective treatment strategies to target S1P signaling for these conditions. The insights presented here possess the potential to offer valuable guidance for the implementation of therapies targeting S1P signaling in various pancreatic diseases.
Keywords:
S1P
; SPHKs
; S1PRs
; acute pancreatitis
; chronic pancreatitis
; pancreatic cancer
1. Introduction
The pancreas consists of two primary components: the exocrine pancreas, which constitutes the majority of pancreatic tissue responsible for secreting digestive enzymes, and the endocrine pancreas, composed of pancreatic islet cells that produce hormones [1,2]. Pancreatic diseases such as pancreatitis and pancreatic cancer (PC) can lead to primary loss of functional parenchyma and/or secondary impairment of exocrine pancreatic function [3]. These conditions may result from genetic factors, a high-fat diet, alcohol consumption, the presence of pancreatic duct stones, or other etiological factors [4,5,6].
Pancreatitis is a prevalent exocrine inflammatory condition of the pancreas, which includes both acute pancreatitis (AP) and chronic pancreatitis (CP) [7]. AP is characterized by edema, acinar cell necrosis, hemorrhage, and severe inflammation of the pancreas, typically manifesting as sudden onset and severe abdominal pain [8,9]. Meanwhile, CP is a clinical condition that develops as a result of ongoing inflammation and chronic fibrosis of the pancreatic acini, leading to irreversible structural damage [10]. PC, often referred to as the "King of Cancer," is a highly lethal malignancy, resulting in considerable economic burden on families and society [11]. Notably, the recurrent AP can give rise to CP, which stands as a prominent etiological factor in the development of PC [12,13]. Furthermore, the developmental defects of pancreas (DDP) are closely related to these pancreatic disorders [14]. However, the understanding of the pathogenesis underlying these pancreatic diseases remains incomplete, and there is currently a lack of specific treatments available. Recently, increasing evidence has suggested that sphingosine-1-phosphate (S1P)-associated cellular and biological mechanisms participated in the onset and progression of these pancreatic diseases.
Sphingolipids are essential components of all eukaryotic membranes [15]. S1P acts as a bioactive sphingolipid that is involved in various cellular processes including cell proliferation, survival, adhesion, and migration [16]. It is produced through the phosphorylation of sphingolipids by sphingosine kinases (SPHKs). S1P may transmit signals to cells via 5 specific G protein-coupled receptors (known as S1PR1–5) that are located on the cell membrane [17]. In recent decades, the advancement of molecular targeting of S1P signaling has become a significant focus of research in autoimmune diseases, inflammatory diseases, tumors, and associated areas [18,19,20,21,22,23,24]. In the case of pancreatic disease, S1P-mediated signaling has been identified as a crucial regulator in pancreatitis [25,26], PC [27,28], as well as the early development of the pancreas [29,30]. In this review, we provide a comprehensive overview of the regulatory mechanisms of the S1P signaling in AP, CP, PC, and DDP. This study emphasizes the importance of sphingolipid metabolism and elucidates the crucial link between S1P signaling and pathological mechanisms, which may provide valuable alternatives to traditional approaches for the management of pancreatic diseases.
2. Search Strategy
A comprehensive literature search was conducted using four electronic databases, namely PubMed, Web of Science, Chinese National Knowledge Infrastructure, and Wanfang data from the study's inception until September 27, 2024. We used search terms and keywords such as “sphingosine-1-phosphate”, “sphingosine kinase”, “S1P receptor”, “pancreas”, “pancreatitis”, “pancreatic cancer”, “pancreas development”. Moreover, some articles were found through the systematic tracking of citations in other scholarly publications or by accessing reputable journal websites.
We included studies elucidating the roles of SPHKs, S1P, S1PRs or related metabolites in pancreatic diseases including AP, CP, PC by integrating evidence from mechanistic studies, animal models, and in vitro observations. Studies were excluded if: (1) Articles in neither English nor Chinese languages; (2) Full texts were not available; (3) Duplicate articles and irrelevant studies. The data were sorted and summarized by different pancreatic diseases.
3. Basics of the S1P Signaling Pathway
3.1. Synthesis, Degeneration, and Transport of S1P
The de novo synthesis pathway (also known as anabolic pathway) of S1P occurs in the endoplasmic reticulum (ER), leading to the formation of ceramide, the central component of sphingolipids. The process initiates with serine and palmitoyl-CoA, which are converted to 3-ketodihydrosphingosine through the action of serine palmitoyltransferase. Subsequently, 3-ketodihydrosphingosine is metabolized to dihydrosphingosine by 3-ketodihydrosphingosine reductase. This is followed by the synthesis of dihydroceramide via ceramide synthase, and finally, dihydroceramide desaturase catalyzes the conversion of dihydroceramide to ceramide [31,32]. Ceramide can be metabolized into sphingomyelin and glucosylceramide in the Golgi apparatus [18]. Alternatively, the sphingomyelin pathway (also known as catabolic pathway) of S1P refers to the sphingomyelin metabolizing to ceramide by sphingomyelinase [22]. After ceramide generation, it is further metabolized to sphingosine by ceramidase, and then SPHKs phosphorylate sphingosine to S1P [31,33], which functions multiple cellular signals by binding with S1PRs located on the cell membrane. This process of S1P is called inside-out signaling. Notably, the generation of sphingosine occurs only through the degradation of ceramide.
The degradation of S1P occurs through the catalytic action of three distinct enzymes: S1P phosphatases (SPPs), S1P lyase (S1PL), and lipid phosphate phosphohydrolases (LPPs) [34]. In most cellular environments, S1PL irreversibly cleaves S1P into hexadecenal and ethanolamine phosphate. Additionally, 2 isoforms of SPP (SPP1 and SPP2) selectively dephosphorylate sphingoid base-1-phosphates, including S1P, leading to the generation of sphingosine [35,36]. The degradation of extracellular S1P is exclusively mediated by a specific group of enzymes known as the LPP isoforms [37].
Cells release S1P into the extracellular environment via various transporters. One of these transporters, known as spinster homolog 2 (SPNS2), is an organic ion transporter that operates independently of adenosine triphosphate (ATP) [38]. Recent studies have elucidated its involvement in developmental processes, maintenance of organ stability, regulation of circulating S1P levels, and modulation of inflammatory responses [39]. Members of the ATP-binding cassette (ABC) family, such as ABCA1 and ABCC1 (MRP1), are also implicated to transport S1P in the extracellular space [40].
3.2. Sphingosine Kinases
The regulation of SPHK1 and SPHK2 primarily occurs through phosphorylation, but also involves their interaction with other molecules in a spatial and temporal manner. In addition, although there is some overlap in their functions, they can be distinguished based on variances in subcellular localization and kinetic properties [33]. SPHK1 is primarily found in the cytosol but can be translocated to the plasma membrane upon stimulation by various cytokines and growth factors [41]. This translocation process is regulated through phosphorylation at Serine 225 by extracellular signal-regulated kinase 1/2 (ERK1/2), which enhances the enzyme's activity [42]. Following translocation, SPHK1 exhibits a propensity to associate with cholesterol and sphingolipid-enriched domains, thereby facilitating cellular proliferation and survival [43]. SPHK2, instead, is found in the plasma membrane, ER, mitochondria, and nucleus. Within the nucleus, SPHK2 has the ability to hinder DNA synthesis leading to cell cycle arrest [44]. Additionally, it can modify histone deacetylases which impacts epigenetic regulation of gene expression [45]. In stressful conditions, SPHK2 localizes in the ER where it promotes ceramide production facilitating apoptotic cell death induction [46].
3.3. S1P Receptors (S1PRs)
S1P exerts a paracrine or autocrine function through its binding to S1PRs [47]. In plasma, S1P are mostly produced by erythrocytes, and these S1P are binded with chaperones including apolipoprotein M (ApoM) and albumin in the circulation and presented to S1PRs [22,48]. S1PRs belong to the family of high-affinity G protein-coupled receptors, consisting of 5 distinct subtypes. S1PRs are selectively coupled to distinct G protein subtypes, with S1PR1 exclusively coupled to heterotrimeric Gαi/o, while S1PR2 and S1PR3 couple to Gαi/o, Gαq, and Gα12/13, and S1PR4 and S1PR5 to Gαi/o and Gα12/13 [18,49]. Additionally, S1PRs are widely expressed on numerous cell types and within tissues, yet they exhibit differential expression content in different parts [16]. S1PR1, S1PR2, and S1PR3 are widely present in many tissues, of which S1PR1 is dominant and mainly involved in the regulation of immune trafficking and immune activation [49,50]. While S1PR2 and S1PR3 are mainly involved in the regulation of endothelial barrier, macrophages and dendritic cell functions [51]. S1PR4 is mainly expressed in lymphatic and lung tissue, and S1PR5 is mainly expressed in the brain, spleen, and skin [16,52]. Collectively, the S1P signaling participates in diverse diseases and drug development targeting this system shows promising potential. The schematic diagram of S1P signaling pathway is shown in Figure 1. The downstream signaling pathways activated multiple cellular signaling such as the Rho family of small GTPases, phosphatidylinositide 3-kinase (PI3K), ERK, signal transducer and activator of transcription 3 (STAT3) [18,53]. These pathways subsequently facilitate a multitude of diverse effects in cells, thereby exerting either positive or negative influences on cellular proliferation, survival, and migration [52,54,55]. The dysregulation of the S1P signaling pathway also plays a pivotal role in pancreatic disorders, which is intricately linked to the expression of S1PRs [56,57,58,59].
3.4. Current Therapeutic Strategies Targeting S1P Signaling: Regulating SPHKs and S1PRs
Currently, a variety of selective and non-selective modulators targeting SPHKs and S1PRs have been extensively investigated, with some progressing to clinical trials and commercialization as drugs [53,54,60]. Some modulators have been reported to regulate various damage indicators and the proliferation of tumor cells in studies related to pancreatitis and PC [61,62,63], of which FTY720 and JTE-013 were the most studied. Fingolimod (FTY720) is an S1P analog approved by the U.S. FDA as an immunomodulatory drug for treating multiple sclerosis [17,58]. FTY720 can be phosphorylated by SPHK2 to become a biologically active form FTY720-P, binding to S1PR1-4 and S1PR5 with a particularly high affinity for S1PR1 [64]. Although FTY720-P is an agonist of the S1PRs, binding of FTY720-P to S1PR1 results in down-modulation and degradation of this receptor, thereby acting as a “functional antagonist” [65]. Moreover, FTY720 can also inhibit the activity of SPHK1 [53]. The representative modulators of S1P signaling are listed in Table 1.
The pancreatic protective effects of FTY720 have been demonstrated in several studies, as evidenced by the attenuation of inflammatory cytokines in AP models [66,67], inhibition of pancreatic fibrosis in CP rats [68], and reduction in tumor volume within the pancreas [69]. Similarly, the compound JTE-013 serves as a specific antagonist targeting S1PR2 [70], and is frequently employed to investigate the functional roles of S1PR2 in diverse pancreatic disorders. The administration of JTE-013 significantly ameliorated severe AP-induced intestinal injury [57], while also inhibiting the activation of pancreatic stellate cells and suppressing the proliferation and migration of PC cells [58,71]. In addition, SKI 5c and PF-543 emerging as potent and specific inhibitors of SPHK1 [59,72,73], have also been reported to ameliorate severe AP-induced pulmonary injury [61], and to mitigate pancreatic pathological damage in CP [59], respectively. Several other modulators, such as the dual inhibitors of SPHK1 and SPHK2, N,N-dimethylsphingosine (DMS) and SKI-II, the SPHK2 antagonist opaganib (ABC294640), the S1PR1 and S1PR3 antagonist VPC23019, and the S1PR2 agonist CYM5520, have been extensively utilized in cancer treatment and research [58,74,75,76,77]. Their potential in PC treatment has also been unveiled in recent years. ABC294640, SKI-II, and VPC23019 have been proved to exhibit anti-pancreatic cancer effects [62,78,79]. However, CYM5520 is demonstrated to promote the development of PC [58]. Overall, the modulation of SPHKs and S1PRs with these compounds may provide novel strategies for the management of pancreatic diseases.
4. Role of S1P Signaling Pathway in Pancreatic Diseases
4.1. AP
4.1.1. Functioning as Potential Biomarkers for Severity of AP
Numerous studies have demonstrated that S1P, together with SPHK and S1PRs, functions as significant biomarkers for predicting the severity and prognosis of AP. Clinical evidence suggested that the expression of SPHK1 and S1PR3 in peripheral immune cells such as leukocytes, neutrophils, monocytes, and lymphocytes were elevated in the early stage of AP patients, and subsequently recovered to the normal levels at the restoration stage [80,81,82]. Later, a positive correlation between the expression level of SPHK1 and the APACHE score, a key disease severity evaluating system, was found in AP patients [82]. In accordance with the clinical findings, S1P concentration and the expression of SPHK1 and S1PR2 in animal models of AP were elevated, indicating that the activation of S1P signaling may provide a risk factor for the onset of AP [25,26,83]. Our previous study using lipidomic analysis found that in L-ornithine-induced severe AP model rats, in contrast with the decrease of glycerophospholipids, pancreatic ceramide increased significantly, and its metabolic products also increased rapidly at 24 hours and peaked at 48 hours after modeling, such as glucosylceramides [84]. However, the research regarding the change of S1P level on AP is inconsistent. As revealed by 2 clinical studies encompassing 36 severe AP patients and 39 AP patients respectively, the S1P concentration in plasma or serum significantly decreased compared with the control volunteers [85,86],. The different results may be related to the severity and time course of AP, which remains under further investigation. A study revealed plasma S1P concentration significantly increased in mild AP patients on days 1 and 3, accompanied by the inhibition of ceramide synthesis. However, in severe AP patients, S1P decreased and the level of ceramide increased significantly on days 1 and 3, and they both subsequently returned to normal levels on day 7 [87]. These results suggest that S1P may be a reliable prognostic marker for AP due to the early sensitivity, and shows different changing patterns in mild and severe AP patients.
4.1.2. Mediating Local and Systematic Inflammation
As a severe inflammatory disease, the primary therapeutic measure and objective of AP is to manage inflammation in a targeted manner [88]. S1P signaling is closely associated with inflammatory signal transduction factors, such as IL-6, tumor necrosis factor (TNF)-α and nuclear factor-kappa B (NF-κB) [89]. It’s indicated that the levels of serum proinflammatory cytokines including IL-6, IL-1β and TNF-α showed similar shifts with SPHK1 expression in peripheral immune cells of severe AP patients, suggesting that the activation of SPHK1 may be closely related to the inflammation of severe AP [80,82]. Meanwhile, inhibition of S1PR2 by JTE-013 or genetic knockdown mitigated the severity of pancreatic injury and inflammation levels, as evidenced by a significant reduction in acinar cell death and inflammatory cytokine release, by regulating Rho-associated kinase (ROCK)/NF-κB signaling activation in acinar cells, and macrophage recruitment as well as polarization towards M1 phenotype [26]. FTY720, a non-selective S1PR modulator [64,90,91], significantly reduced pancreatic pathological injury, monocyte chemotactic protein-1 (MCP-1) level of hypertriglyceridemic AP mice models [92]. In addition, FTY720 decreased circulative IL-6, IL-10 and TNF-α levels, as well as the number of CD4+/CD8+ cells in necrotizing AP rats [66]. Although most literature indicated that inhibition of S1P signaling exerted a protective role in AP, similar protective effects have been observed with some S1P signaling agonists [26,61,83,93]. SEW2871, an S1PR1 agonist, significantly ameliorated parameters of cerulein-induced AP mice including pancreatic pathological injury and systematic inflammation, which was probably attributed to the inhibition of STAT3 phosphorylation in the pancreas [93].
4.1.3. Inducing Pyroptosis of Pancreatic Acinar Cells (PACs)
PACs are the predominant exocrine cell type undergoing dysfunction in AP [94]. In recent years there has been a growing interest in mitigating the severity of AP through the intervention targeting cell pyroptosis in PACs [95,96,97]. Pyroptosis was observed in acinar cells of cerulein-induced AP mice through detecting the activation of the pyroptosis characteristic protein, Caspase-1 and GSDMD. Genetic knockout of Sphk1 significantly attenuated cell damage and inhibited pyroptosis of PACs in AP mice [83]. Further in-depth investigation demonstrated that the upregulation of SPHK1 expression in acinar cells was implicated in the regulation of ER stress. Initiation of ER stress triggered protein kinase RNA-like endoplasmic reticulum kinase (PERK) activation, leading to enhanced thioredoxin-interacting protein (TXNIP) expression, which subsequently mediated Nod-like receptor family, pyrin domain containing 3 (NLRP3) inflammasome activation and facilitated the generation of active Caspase-1, thereby further promoting cellular pyroptosis [83].
4.1.4. Involving in AP-induced Organ Injury
The rapid production and release of a large number of inflammatory cytokines in sever AP can easily lead to systemic inflammatory response syndrome and multiple organ dysfunction syndromes [98]. AP-associated acute lung injury represents the most common and earliest organ dysfunction in disease development, which is characterized by significant pulmonary edema, hyperemia, and inflammatory infiltration in the alveoli [99]. The inhibition of SPHK1 by SKI 5c resulted in an increased survival rate of AP-associated acute lung injury rats, while concurrently reducing the MPO level in the lung and protein content of bronchoalveolar lavage fluid [61]. By contrast, others found that S1P signaling showed anti-inflammatory effects and protection against pulmonary injury in AP-associated acute lung injury [67,100]. Intraperitoneal administration of S1P mitigated pulmonary pathological injury, leading to a reduction in inflammatory mediators including IL-1β, IL-6, and TNF-α in bronchoalveolar lavage fluid. Additionally, it inhibited lung immune cell infiltration and suppressed NF-κB activity in alveolar macrophages [67,100]. These conflicting results reflected both the pro-inflammatory and anti-inflammatory effects of S1P signaling during AP-associated acute lung injury. As a powerful angiogenic factor that strengthens lung endothelial cell integrity and suppresses vascular permeability and alveolar flooding in preclinical animal models of acute lung injury, S1P holds therapeutic potential [101]. It’s supposed that the role of S1P signaling in lung inflammation and injury might depend on the type of insult and the degree of oxidative stress, which should be further investigated in long-term studies.
Intestinal damage is another severe complication associated with severe AP, which includes gut mucosal barrier functional changes, intestinal immune barrier dysfunction, and gastrointestinal microbial imbalance, exaggerates the severity of disease by increasing intestinal permeability, microbial infections, bacterial translocation, and release of proinflammatory substances [98,102]. The inhibition of S1PR2 by JTE-013 significantly attenuated pathological injury and pyroptosis in intestinal tissues of severe AP mice [57]. Additionally, S1PR2 was reported to exert a positive regulatory effect on macrophage pyroptosis, and this process might be attributed to the upregulation of the Ras homolog family member A (RhoA)/ROCK signaling pathway [57].
4.2. CP
Irreversible fibrosis of the pancreas is the main characteristic of CP [103]. Pancreatic fibrosis is essentially an increase in synthesis and decrease in degradation of collagen-based extracellular matrix, of which pancreatic stellate cells (PSCs) are key cells involved in pancreatic injury [104]. It has been demonstrated that S1P signaling was closely associated with the development of diverse tissue fibrosis by interacting with excessive extracellular matrix synthesis and high profibrotic marker gene expression [105,106,107,108]. The concentration of S1P was found to be significantly elevated in in vivo and in vitro CP models [59,71]. It was also found that administration of S1P resulted in severe impairment of pancreatic function and histomorphology in CP rats by S1P-S1PR2 signaling-mediated autophagy and the NLRP3 inflammasome activation in PSCs [71]. Through further crosstalk experiments, it was revealed that the S1P derived from PACs contributed to the fibrosis of CP by inducing autophagy and activating PSCs through the adenosine monophosphate-activated protein kinase (AMPK)/mammalian target of rapamycin (mTOR) signaling pathway [59]. Moreover, the S1PR modulator FTY720 has been shown to exhibit an anti-fibrotic role by significantly reducing the expression of fibrosis-related regulatory factors, including IFN-γ and transforming growth factor (TGF)-β1 [68]. Due to the non-selectivity of FTY720 in regulating S1PR, how FTY720 regulates S1PR in specific disease models remains elusive, and there is still a lack of in-depth explanation of FTY720 in regulatory mechanisms of pancreatitis. Moreover, since there are a limited number of studies investigating the correlation between S1P signaling and CP, further elucidation is required regarding the corresponding downstream molecular regulatory mechanisms and signaling pathways.
4.3. PC
4.3.1. Targeting S1P Signaling for PC Therapy
S1P plays a pivotal role in multiple features of cancer, including sustained activation of cell proliferation signaling pathways, evasion of growth inhibitory signals, resistance to programmed cell death, promotion of uncontrolled replication, induction of angiogenesis and initiation of invasion and metastasis processes [109,110,111]. Targeting SPHK2 has proved to successfully improve tumor outcomes via PhotoImmunoNanoTherapy, which regulated cellular stress and inflammatory genetic programs through epigenetic mechanisms [112]. Moreover, the SPHK2 selective inhibitor ABC294640 has been reported to have antitumor properties alone or in combination with the multikinase inhibitor sorafenib, an effective treatment in renal cancer, hepatic cancer, and pancreatic adenocarcinoma [63,78,113]. Surprisingly, the sensitivity of PC cells to gemcitabine was enhanced when the ceramide/S1P ratio was increased through the utilization of pharmacologic methods (such as SPHK1 inhibitor or ceramide analogue) or small interfering RNA-based approaches that either elevated intracellular ceramide levels or reduced SPHK1 activity [114]. To summarize, exploring the modulation of S1P signaling could offer a promising and innovative approach to treating PC.
4.3.2. Impacting Mitochondria-Mediated Apoptosis of PC Cells
Mitochondria, derived from aerobic bacteria, are pivotal organelles in eukaryotes, and functions importantly in tumorigenesis of PC [115]. Recent studies have indicated that mitochondria may play indispensable roles in the chemoresistance of PC by influencing apoptosis via mitochondrial functions [116,117,118,119,120,121]. It has been found that treatment with ABC294640, an antagonist of SPHKs, significantly delayed tumor growth in mice injected with pancreatic ductal adenocarcinoma (PDAC) cells, which was related to the inhibition of apoptosis by the mitochondrial Kv1.3 ion channel blocker PAPTP [122,123,124]. On the other hand, regulation of S1PRs also modulates mitochondrial-mediated apoptosis in PC cells. SPHK1 antagonist mebendazole was reported to induce apoptosis in PC cells through the intrinsic mitochondrial pathway [125]. Similar conclusions were drawn from experiments conducted on PSN1 cells treated with SKI-II [62]. The administration of FTY720 effectively suppressed the growth of PC cells in both humans and mice, resulting in enhanced apoptosis and cell cycle arrest [69]. Furthermore, treatment with either FTY720 or gemcitabine, a drug of choice for PC therapy, whether used alone or in combination, induced mitochondrial depolarization [69].
4.3.3. Inhibiting S1P Signaling Improves PC by Inducing ER Stress of PC Cells
The intersection between ER stress and the ceramide/S1P axis occurs at multiple crucial points in PC. Notably, both anabolic pathway-mediated generation and S1P lyase-mediated degradation of S1P are processed in ER [126]. Previous research has demonstrated that ER stress activates NF-κB through the activation of receptors located on the plasma membrane: S1PR1, S1PR2, and S1PR3 [127,128]. Conversely, ER stress can stimulate the production of S1P through an increase in SPHK2 expression and utilize it as part of downstream signaling pathways [129]. Evidence from preclinical models of cancer indicates that cancer cells can enhance SPHK2 expression as a survival mechanism in response to ER stress [126]. It has been proven that the dysregulated metabolism of sphingolipids is influenced by both PC and inhibiting S1P can enhance ER stress and immunogenic cell death (ICD), leading to an improved therapeutic response to oxaliplatin in PC. The provided experimental data demonstrates that the induction of ER stress is significantly enhanced by downregulating S1P expression through the knockdown of Sphk2 or employing the SPHK2 inhibitor ABC294640, at various doses and in different human and murine PC cell lines, resulting in increased phosphorylation of the PERK/eukaryotic initiation factor 2alpha (eIF2α) pathway and even immunogenic cell death [130]. Moreover, the in vivo findings reveal a notable increase in HMGB1 (a key mediator of ICD) staining intensity, accompanied by elevated expression of CD3 and CD8 markers in tumors treated with ABC294640 and oxaliplatin, as well as reduced tumor weight [130]. These findings provide compelling evidence for the significance of investigating sphingolipid metabolism in PC and exploring the potential therapeutic targeting of SPHK2/S1P in PC.
4.3.4. Mediating PSCs Activation That Transferring to PC
Tumor-stromal interactions are widely acknowledged to play a pivotal role in the progression of PC cancer [131]. PSCs are stellate-shaped stromal cells situated at the basolateral aspect of acinar cells or in the adjacent perivascular and periductal areas within the normal pancreas [132]. Growth factors, chemokines, cytokines, miRNAs, and exosomes secreted by PSCs have been observed to exert their effects within the tumor microenvironment. They can either activate PSCs through autocrine signaling or elicit paracrine signals on epithelial tumor cells, thereby facilitating enhanced proliferation, migration, and invasion of these cells [133,134]. Studies of S1P and PSCs are well-documented, it is widely acknowledged that S1P promotes PSC activation in pancreatic-related diseases and primary PSCs [28,59,71]. For instance, the following studies were conducted to provide evidence supporting the role of S1P in triggering PSC activation, leading to the subsequent release of paracrine factors that promote PC cell invasion and growth [28]. They utilized conditioned media obtained from PSCs treated with S1P and applied it to the human pancreatic cancer cell lines, PANC1 and L3.6. The results showed that the conditioned media derived from S1P-stimulated PSCs significantly enhanced both migration and invasion of PANC1 cells as determined [28]. On the contrary, inhibition of S1PR2 by JTE-013 or knockdown of S1pr2 by S1pr2-shRNA resulted in reduced tumor cell migration and invasion compared to control PSCs. Accordingly, in both orthotopic and subcutaneous models of PC, the growth kinetics of tumors formed by co-implantation of S1pr2sh PSCs and L3.6 cells was notably slower compared to those formed by co-implantation of regular PSCs and L3.6 cells, resulting in a significant reduction in the final tumor weight [28].
4.3.5. Being Activated by Bile Acids (BAs)
BAs are widely acknowledged for their capacity to enhance the breakdown and absorption of fats, while also playing a pivotal role in lipid metabolism [135]. Patients diagnosed with PC, particularly those harboring tumors located in the head of the pancreas, frequently encounter biliary obstruction and elevated levels of BAs [136]. In PC cells, BAs diminish susceptibility to programmed cell death, facilitate progression through the cell cycle, and augment expression of inflammatory mediators and cellular motility [137]. Furthermore, BAs may disrupt biomembranes at high concentrations [138,139]. Conjugated BAs, such as taurocholic acid (TCA), have been shown to activate S1PR2, and the ERK1/2 and protein kinase B (AKT) signaling pathways [140,141,142]. The growth and migration of S1PR2-expressing cell lines PANC-2-luc and AsPC-1 were enhanced in a dose-dependent manner by the activation of TCA or the S1PR2 agonist CYM5520, while they did not have any impact on the growth of MIA PaCa-2 and BxPC-3 cells, which primarily exhibit S1PR5 expression. Based on the aforementioned results, an in vivo study was conducted using a murine model of cholestasis associated with metastatic PC, yielding consistent findings with those obtained from in vitro experiments [58]. The findings suggest that TCA functions as an agonist, activating S1PR2 in pancreatic cells and thereby promoting the development of PC.
Collectively, these studies show that SPHK/S1P signaling promotes cell proliferation, migration, and survival in vitro, as well as enhances tumor growth and activity in vivo. Furthermore, numerous other scholarly articles corroborate this assertion [79,143,144,145,146,147,148]. Yet, the role of S1P in the development and treatment of PC remains a subject of controversy, with some literature suggesting its potential beneficial effects while lacking specific mechanistic studies [149,150,151]. Given the above, the present section emphasizes the pivotal role of S1P-dependent mechanisms in tumor growth, proliferation, and metastasis, providing an overview of current and emerging therapeutic strategies targeting enzymes involved in sphingolipid metabolism and/or signaling for precision PC therapy.
5. S1P Signaling Participates in the Developmental Defects of Pancreas (DDP)
DDP, such as annular pancreas, pancreas divisum, and dorsal pancreatic agenesis, frequently involve congenital genetic abnormalities, which have been identified as a significant etiology for pancreatitis and other gastrointestinal disorders [14,152,153]. The pancreas originates from a small ventral bud and a larger dorsal bud. During the rotation of the foregut, the ventral pancreas undergoes rotational movement towards the dorsal pancreas, ultimately fusing together to form a fully developed pancreas with interconnected ducts [154]. Among the various developmental congenital anomalies, dorsal pancreatic agenesis is one of the rare entities [154]. DDP was reported to be rescued by S1P supplementation [29]. The data suggested that the mechanism of S1P may involve a direct effect on the mesenchyme, which was reported to benefit dorsal pancreatic endoderm [155], instead of a direct effect on the endoderm. The researchers further speculated that blood vessel-derived S1P escaped from the circulation, then bound S1PRs on dorsal pancreatic mesenchymal cells or endothelial cells, stimulating mesenchymal cell proliferation in a cell-autonomous manner or indirectly via intercellular signaling, which rescued dorsal pancreatic growth [29]. Subsequently, the research team conducted further investigations into the impact of S1PR defects on early pancreas development [30]. S1PRs ablation resulted in a reduced volume of the dorsal and ventral pancreata, but it had no effect on endocrine or exocrine differentiation [30]. The defects of the dorsal pancreas in S1pr–/– mice were found to be attributed to reduced proliferation of Pdx1+ progenitors, and induced hypervascularization may underlie the adverse impact of S1PR ablation on endodermal growth [30]. Nevertheless, the specific S1PR types and downstream regulatory mechanisms were not clearly clarified in the studies [29,30]. A more recent study confirmed that S1P played a key role in pancreas development linking specification and lineage allocation through S1PR2 [56]. The findings revealed a delay in pancreas development and severe impairments in both the endocrine and acinar lineages of S1pr2-deficient mice embryos. S1P/S1PR2 signaling performed a function through Gαi-mediated yes-associated protein (YAP) stabilization and Notch attenuation, in turn partially activating connective tissue growth factor (CTGF) which participated in regulating the survival of endocrine and acinar progenitors [56]. Taken together, these studies highlight the role of S1P signaling in diseases associated with aberrant pancreas development and may propose novel therapeutic strategies.

Table 1.
Representative modulators of S1P signaling.
| Compound | Structure | Primarily used as | Ref. |
|---|---|---|---|
| N,N-dimethylsphingosine (DMS) | 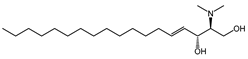 |
Antagonist of SPHK1 and SPHK2 | [74,156] |
| SKI-II | 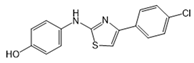 |
Antagonist of SPHK1 and SPHK2 | [75,157] |
| SKI 5c | 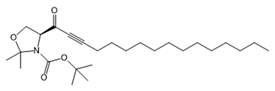 |
Antagonist of SPHK1 | [72] |
| PF-543 | 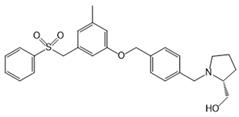 |
Antagonist of SPHK1 | [59,73] |
| Opaganib (ABC294640) | 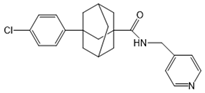 |
Antagonist of SPHK2 | [76] |
| K145 | 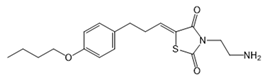 |
Antagonist of SPHK2 | [158] |
| Fingolimod (FTY720) | 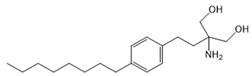 |
Modulator of S1PR1, S1PR3, S1PR4 and S1PR5, antagonist of SPHK1 | [17,53,58] |
| Etrasimod (APD334) | 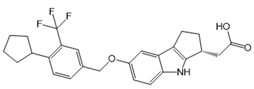 |
Modulator of S1PR1, S1PR4 and S1PR5 | [159] |
| Siponimod (BAF312) | 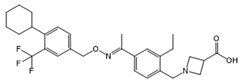 |
Modulator of S1PR1 and S1PR5 | [160] |
| Ozanimod (RPC1063) | 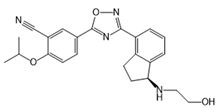 |
Agonist of S1PR1 and S1PR5 | [161] |
| VPC23019 | 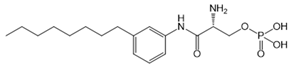 |
Antagonist of S1PR1 and S1PR3 | [77] |
| SEW2871 | 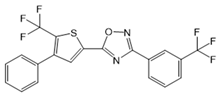 |
Agonist of S1PR1 | [162] |
| Ponesimod (ACT-128800) | 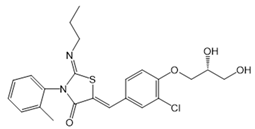 |
Agonist of S1PR1 | [163] |
| NIBR-0213 | 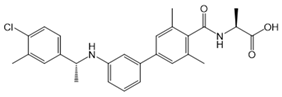 |
Antagonist of S1PR1 | [164] |
| CYM5520 | 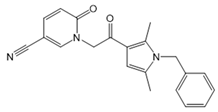 |
Agonist of S1PR2 | [58] |
| JTE-013 | 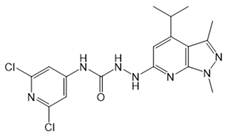 |
Antagonist of S1PR2 | [70] |
| CYM5541 | 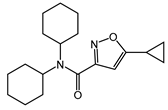 |
Agonist of S1PR3 | [165] |
| TY52156 | 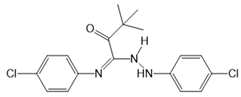 |
Antagonist of S1PR3 | [164] |
| CYM50358 | 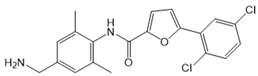 |
Antagonist of S1PR4 | [166] |
| A-971432 | 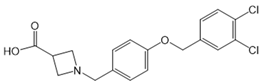 |
Agonist of S1PR5 | [167] |
6. Conclusions and Future Prospectives
The fluctuations in levels of the sphingolipid metabolite S1P, as well as the altered expression of its synthesizing enzymes SPHKs and receptors S1PRs, exert significant impacts on both physiological and pathological functions of the pancreas. Details are presented in Table 2 and Figure 2. In pancreatitis, S1P signaling exerts local protective effects on the pancreas by modulating inflammatory signals and acinar cell pyroptosis. Moreover, it regulates the extent of pathological injury in AP models through other organs, especially on the lung-intestinal axis. In addition, the underlying mechanism of S1P signaling in CP primarily revolves around the regulation of PSC activation and fibrosis. The regulatory effects of S1P signaling on pancreatitis predominantly involve NF-κB, STAT3 phosphorylation, PERK/TXNIP/NLRP3, RhoA/ROCK, AMPK/mTOR pathways as well as IFN-γ and TGF-β1. In PC, inhibiting SPHKs/S1PRs can modulate the intrinsic mitochondrial pathway or enhance endoplasmic reticulum stress to impede cancer progression primarily encompasses pathways including AKT, PERK/eIF2α, JAK2/STAT3 and FAK/Vimentin, etc. Additionally, TCA-induced activation of S1PR2 also contributes to PC development. Importantly, targeting the S1P signaling pathway may improve drug sensitivity in PC treatment. Nevertheless, the correlation between S1P signaling and pancreatitis or PC remains controversial. On one hand, it may be crucial for S1P to remain within a certain range, as deviations towards elevated or diminished concentrations can significantly contribute to the progression of these diseases. On the other hand, the role of S1P signaling in these diseases may vary depending on sample types, specific disease models, injury severity levels and time course, as well as variations in the specific receptors it targets. In addition to acquired factors, it is noteworthy that the impact of S1P on pancreatic diseases may also be attributed to congenital developmental abnormalities. Defects in S1P signaling that occur during the embryonic period can lead to aberrant development of pancreas, deficient proliferation of pancreatic progenitors, loss of endocrine and acinar differentiation, etc. These abnormalities could potentially serve as significant etiological factors underlying pancreatic diseases, such as pancreatitis.
In the future, the potential breakthroughs of S1P signaling in pancreatic diseases warrant further research. The directions can include but are not limited to the following: (1) Considering the "two-way" effect of S1P signaling in pancreatic diseases, it’s necessary to conduct more basic investigations into the role and functional levels of S1P and its specific receptor across diverse cell types and disease contexts; (2) Currently, the majority of studies investigating the mechanism of S1P receptor in pancreatic diseases primarily focus on S1PR2, with limited research conducted on other receptors. Thus, additional research is warranted to investigate the role of other receptors and explore their in-depth mechanisms. (3) It is of great significance to elucidate whether there are crosstalk or direct correlations between other cell types of neighboring microenvironments, such as nerve cells and immune cells, with pancreatic cells through S1P secretion and associated signaling pathways. (4) There is currently a scarcity of clinical studies exploring S1P signaling as a therapeutic target for pancreatic diseases. Further clinical practice needs to be performed to determine whether S1P signaling can serve as an effective therapeutic intervention. With a more profound comprehension of S1P signaling, future research endeavors focusing on the modulation of S1P signaling, along with the development of SPHK or S1PR agonists/antagonists may emerge as a prominent area of interest, potentially leading to innovative therapeutic interventions for pancreatic diseases.
Author Contributions
F.F. and W.L. drafted the manuscript. X.Z. and Y.W. helped in searching for literature. C.H. and D.D. designed the review and revised the manuscript.
Acknowledgments
The study was supported by the Key Research Project of Sichuan Province (No. 2024YFFK0153 to C.H.) and Natural Science Foundation of Sichuan Province (No.2023NSFSC1755 to D.D.).
Conflicts of Interest
The authors declare that they have no competing interests.
Abbreviations
ABC, ATP-binding cassette; AKT, protein kinase B; AMPK, adenosine monophosphate-activated protein kinase; AP, acute pancreatitis; ATP, adenosine triphosphate; BAs, bile acids; CP, chronic pancreatitis; CTGF, connective tissue growth factor; DDP, developmental defects of pancreas; eIF2α, eukaryotic initiation factor 2alpha; ER, endoplasmic reticulum; ERK1/2, extracellular signal-regulated kinase 1/2; FAK, focal adhesion kinase; JAK2, Janus kinases2; ICD, immunogenic cell death; LPPs, lipid phosphate phosphohydrolases; MCP-1, monocyte chemotactic protein-1; mTOR, mammalian target of rapamycin; NF-κB, nuclear factor-kappa B; NLRP3, Nod-like receptor family, pyrin domain containing 3; PACs, pancreatic acinar cells; PC, pancreatic cancer; PDAC, pancreatic ductal adenocarcinoma; PERK, protein kinase RNA-like endoplasmic reticulum kinase; PI3K, phosphatidylinositide 3-kinase; PSCs, pancreatic stellate cells; RhoA, Ras homolog family member A; ROCK, Rho-associated kinase; S1P, sphingosine-1-phosphate; S1PL, S1P lyase; S1PRs, S1P receptors; SPHKs, sphingosine kinases; SPNS2, spinster homolog 2; SPPs, S1P phosphatases; STAT3, signal transducer and activator of transcription 3; TCA, taurocholic acid; TGF, transforming growth factor; TNF, tumor necrosis factor; TXNIP, thioredoxin-interacting protein; YAP, yes-associated protein
References
- Kimita, W.; Petrov, M.S. Iron metabolism and the exocrine pancreas. Clin. Chim. Acta. 2020, 511, 167-176. [CrossRef]
- Zhou, Q.; Melton, D.A. Pancreas regeneration. Nature 2018, 557, 351-358. [CrossRef]
- Struyvenberg, M.R.; Martin, C.R.; Freedman, S.D. Practical guide to exocrine pancreatic insufficiency - Breaking the myths. BMC Med. 2017, 15, 29. [CrossRef]
- Singh, V.K.; Yadav, D.; Garg, P.K. Diagnosis and management of chronic pancreatitis: A review. JAMA 2019, 322, 2422-2434.
- Wang, G.J.; Gao, C.F.; Wei, D.; Wang, C.; Ding, S.Q. Acute pancreatitis: etiology and common pathogenesis. World J. Gastroenterol. 2009, 15, 1427-1430. [CrossRef]
- Zhang, R.; Peng, X.; Du, J.X.; Boohaker, R.; Estevao, I.L.; Grajeda, B.I.; Cox, M.B.; Almeida, I.C.; Lu, W. Oncogenic KRASG12D reprograms lipid metabolism by upregulating SLC25A1 to drive pancreatic tumorigenesis. Cancer Res. 2023, 83, 3739-3752. [CrossRef]
- Mayerle, J.; Sendler, M.; Hegyi, E.; Beyer, G.; Lerch, M.M.; Sahin-Tóth, M. Genetics, cell biology, and pathophysiology of pancreatitis. Gastroenterology 2019, 156, 1951-1968. [CrossRef]
- Chan, Y.C.; Leung, P.S. Acute pancreatitis: Animal models and recent advances in basic research. Pancreas 2007, 34, 1-14.
- Habtezion, A.; Gukovskaya, A.S.; Pandol, S.J. Acute pancreatitis: A multifaceted set of organelle and cellular interactions. Gastroenterology 2019, 156, 1941-1950. [CrossRef]
- Hammad, A.Y.; Ditillo, M.; Castanon, L. Pancreatitis. Surg. Clin. North Am. 2018, 98, 895-913.
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2021, 71, 209-249. [CrossRef]
- Lee, B.; Jones, E.K.; Manohar, M.; Li, L.; Yadav, D.; Conwell, D.L.; Hart, P.A.; Vege, S.S.; Fogel, E.L.; Serrano, J., et al. Distinct serum immune profiles define the spectrum of acute and chronic pancreatitis from the multicenter prospective evaluation of chronic pancreatitis for epidemiologic and translational studies (PROCEED) study. Gastroenterology 2023, 165, 173-186. [CrossRef]
- Zheng, L.; Xue, J.; Jaffee, E.M.; Habtezion, A. Role of immune cells and immune-based therapies in pancreatitis and pancreatic ductal adenocarcinoma. Gastroenterology 2013, 144, 1230-40. [CrossRef]
- Schnedl, W.J.; Piswanger-Soelkner, C.; Wallner, S.J.; Reittner, P.; Krause, R.; Lipp, R.W.; Hohmeier, H.E. Agenesis of the dorsal pancreas and associated diseases. Dig. Dis. Sci. 2009, 54, 481-7. [CrossRef]
- Green, C.D.; Maceyka, M.; Cowart, L.A.; Spiegel, S. Sphingolipids in metabolic disease: The good, the bad, and the unknown. Cell Metab. 2021, 33, 1293-1306. [CrossRef]
- Bravo, G.; Cedeño, R.R.; Casadevall, M.P.; Ramió-Torrentà, L. Sphingosine-1-Phosphate (S1P) and S1P signaling pathway modulators, from current insights to future perspectives. Cells 2022, 11. [CrossRef]
- Colombo, E.; Farina, C. Lessons from S1P receptor targeting in multiple sclerosis. Pharmacol. Ther. 2022, 230, 107971. [CrossRef]
- Verstockt, B.; Vetrano, S.; Salas, A.; Nayeri, S.; Duijvestein, M.; Vande Casteele, N. Sphingosine 1-phosphate modulation and immune cell trafficking in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 351-366. [CrossRef]
- Baeyens, A.; Bracero, S.; Chaluvadi, V.S.; Khodadadi-Jamayran, A.; Cammer, M.; Schwab, S.R. Monocyte-derived S1P in the lymph node regulates immune responses. Nature 2021, 592, 290-295. [CrossRef]
- Schneider, G. S1P signaling in the tumor microenvironment. Adv. Exp. Med. Biol. 2020, 1223, 129-153.
- Stepanovska, B.; Huwiler, A. Targeting the S1P receptor signaling pathways as a promising approach for treatment of autoimmune and inflammatory diseases. Pharmacol. Res. 2020, 154, 104170. [CrossRef]
- Cartier, A.; Hla, T. Sphingosine 1-phosphate: Lipid signaling in pathology and therapy. Science 2019, 366. [CrossRef]
- Mendelson, K.; Evans, T.; Hla, T. Sphingosine 1-phosphate signalling. Development. 2014, 141, 5-9.
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: an enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol. 2003, 4, 397-407. [CrossRef]
- Konończuk, T.; Łukaszuk, B.; Mikłosz, A.; Chabowski, A.; Żendzian-Piotrowska, M.; Kurek, K. Cerulein-induced acute pancreatitis affects sphingomyelin signaling pathway in rats. Pancreas 2018, 47, 898-903. [CrossRef]
- Yang, J.; Tang, X.; Li, B.; Shi, J. Sphingosine 1-phosphate receptor 2 mediated early stages of pancreatic and systemic inflammatory responses via NF-kappa B activation in acute pancreatitis. Cell Commun. Signal. 2022, 20, 157. [CrossRef]
- Yuza, K.; Nakajima, M.; Nagahashi, M.; Tsuchida, J.; Hirose, Y.; Miura, K.; Tajima, Y.; Abe, M.; Sakimura, K.; Takabe, K., et al. Different roles of sphingosine kinase 1 and 2 in pancreatic cancer progression. J. Surg. Res. 2018, 232, 186-194. [CrossRef]
- Bi, Y.; Li, J.; Ji, B.; Kang, N.; Yang, L.; Simonetto, D.A.; Kwon, J.H.; Kamath, M.; Cao, S.; Shah, V. Sphingosine-1-phosphate mediates a reciprocal signaling pathway between stellate cells and cancer cells that promotes pancreatic cancer growth. Am. J. Pathol. 2014, 184, 2791-2802. [CrossRef]
- Edsbagge, J.; Johansson, J.K.; Esni, F.; Luo, Y.; Radice, G.L.; Semb, H. Vascular function and sphingosine-1-phosphate regulate development of the dorsal pancreatic mesenchyme. Development. 2005, 132, 1085-92. [CrossRef]
- Sand, F.W.; Hörnblad, A.; Johansson, J.K.; Lorén, C.; Edsbagge, J.; Ståhlberg, A.; Magenheim, J.; Ilovich, O.; Mishani, E.; Dor, Y., et al. Growth-limiting role of endothelial cells in endoderm development. Dev. Biol. 2011, 352, 267-277. [CrossRef]
- Fyrst, H.; Saba, J.D. An update on sphingosine-1-phosphate and other sphingolipid mediators. Nat. Chem. Biol. 2010, 6, 489-497. [CrossRef]
- Harrison, P.J.; Dunn, T.M.; Campopiano, D.J. Sphingolipid biosynthesis in man and microbes. Nat. Prod. Rep. 2018, 35, 921-954. [CrossRef]
- Jozefczuk, E.; Guzik, T.J.; Siedlinski, M. Significance of sphingosine-1-phosphate in cardiovascular physiology and pathology. Pharmacol. Res. 2020, 156, 104793. [CrossRef]
- Liu, X.; Zhang, Q.H.; Yi, G.H. Regulation of metabolism and transport of sphingosine-1-phosphate in mammalian cells. Mol. Cell Biochem. 2012, 363, 21-33. [CrossRef]
- Bandhuvula, P.; Saba, J.D. Sphingosine-1-phosphate lyase in immunity and cancer: Silencing the siren. Trends Mol. Med. 2007, 13, 210-217. [CrossRef]
- Mandala, S.M. Sphingosine-1-phosphate phosphatases. Prostaglandins. Other. Lipid. Mediat. 2001, 64, 143-56. [CrossRef]
- Książek, M.; Chacińska, M.; Chabowski, A.; Baranowski, M. Sources, metabolism, and regulation of circulating sphingosine-1-phosphate. J. Lipid Res. 2015, 56, 1271-1281. [CrossRef]
- Kawahara, A.; Nishi, T.; Hisano, Y.; Fukui, H.; Yamaguchi, A.; Mochizuki, N. The sphingolipid transporter spns2 functions in migration of zebrafish myocardial precursors. Science 2009, 323, 524-527. 10.1126/science.1167449.
- Spiegel, S.; Maczis, M.A.; Maceyka, M.; Milstien, S. New insights into functions of the sphingosine-1-phosphate transporter SPNS2. J. Lipid Res. 2019, 60, 484-489. [CrossRef]
- Aye, I.L.; Singh, A.T.; Keelan, J.A. Transport of lipids by ABC proteins: interactions and implications for cellular toxicity, viability and function. Chem. Biol. Interact. 2009, 180, 327-339. [CrossRef]
- Leclercq, T.M.; Pitson, S.M. Cellular signalling by sphingosine kinase and sphingosine 1-phosphate. IUBMB. Life 2006, 58, 467-472. [CrossRef]
- Pitson, S.M.; Moretti, P.A.; Zebol, J.R.; Lynn, H.E.; Xia, P.; Vadas, M.A.; Wattenberg, B.W. Activation of sphingosine kinase 1 by ERK1/2-mediated phosphorylation. EMBO J. 2003, 22, 5491-5500.
- Pitson, S.M.; Xia, P.; Leclercq, T.M.; Moretti, P.A.; Zebol, J.R.; Lynn, H.E.; Wattenberg, B.W.; Vadas, M.A. Phosphorylation-dependent translocation of sphingosine kinase to the plasma membrane drives its oncogenic signalling. J. Exp. Med. 2005, 201, 49-54. [CrossRef]
- Igarashi, N.; Okada, T.; Hayashi, S.; Fujita, T.; Jahangeer, S.; Nakamura, S. Sphingosine kinase 2 is a nuclear protein and inhibits DNA synthesis. J. Biol. Chem. 2003, 278, 46832-9. [CrossRef]
- Hait, N.C.; Allegood, J.; Maceyka, M.; Strub, G.M.; Harikumar, K.B.; Singh, S.K.; Luo, C.; Marmorstein, R.; Kordula, T.; Milstien, S., et al. Regulation of histone acetylation in the nucleus by sphingosine-1-phosphate. Science 2009, 325, 1254-1257. [CrossRef]
- Maceyka, M.; Sankala, H.; Hait, N.C.; Le Stunff, H.; Liu, H.; Toman, R.; Collier, C.; Zhang, M.; Satin, L.S.; Merrill, A.H., Jr., et al. SphK1 and SphK2, sphingosine kinase isoenzymes with opposing functions in sphingolipid metabolism. J. Biol. Chem. 2005, 280, 37118-37129. [CrossRef]
- Strub, G.M.; Maceyka, M.; Hait, N.C.; Milstien, S.; Spiegel, S. Extracellular and intracellular actions of sphingosine-1-phosphate. Adv. Exp. Med. Biol. 2010, 688, 141-155.
- Spiegel, S.; Milstien, S. The outs and the ins of sphingosine-1-phosphate in immunity. Nat. Rev. Immunol. 2011, 11, 403-415. [CrossRef]
- Yanagida, K.; Hla, T. Vascular and immunobiology of the circulatory sphingosine 1-phosphate gradient. Annu. Rev. Physiol. 2017, 79, 67-91. [CrossRef]
- Wu, J.; Shi, Y. Progress in regulation of vascular function by sphingosine-1-phosphate in atherosclerosis. Chinese Journal of Pathophysiology 2023, 39, 2288-2295.
- Tsai, H.C.; Han, M.H. Sphingosine-1-phosphate (S1P) and S1P signaling pathway: Therapeutic targets in autoimmunity and inflammation. Drugs 2016, 76, 1067-1079. [CrossRef]
- Aarthi, J.J.; Darendeliler, M.A.; Pushparaj, P.N. Dissecting the role of the S1P/S1PR axis in health and disease. J. Dent. Res. 2011, 90, 841-854. [CrossRef]
- Xie, Z.; Liu, H.; Geng, M. Targeting sphingosine-1-phosphate signaling for cancer therapy. Sci. China Life Sci. 2017, 60, 585-600. [CrossRef]
- McGinley, M.P.; Cohen, J.A. Sphingosine 1-phosphate receptor modulators in multiple sclerosis and other conditions. Lancet 2021, 398, 1184-1194. [CrossRef]
- Burg, N.; Salmon, J.E.; Hla, T. Sphingosine 1-phosphate receptor-targeted therapeutics in rheumatic diseases. Nat. Rev. Rheumatol. 2022, 18, 335-351. [CrossRef]
- Serafimidis, I.; Rodriguez-Aznar, E.; Lesche, M.; Yoshioka, K.; Takuwa, Y.; Dahl, A.; Pan, D.; Gavalas, A. Pancreas lineage allocation and specification are regulated by sphingosine-1-phosphate signalling. PLoS Biol. 2017, 15, e2000949. [CrossRef]
- Lin, T.; Peng, M.; Zhu, Q.; Pan, X. S1PR2 participates in intestinal injury in severe acute pancreatitis by regulating macrophage pyroptosis. Front. Immunol. 2024, 15, 1405622. [CrossRef]
- Sarkar, J.; Aoki, H.; Wu, R.; Aoki, M.; Hylemon, P.; Zhou, H.; Takabe, K. Conjugated bile acids accelerate progression of pancreatic cancer metastasis via S1PR2 signaling in cholestasis. Ann. Surg. Oncol. 2023, 30, 1630-1641. [CrossRef]
- Wang, D.; Han, S.; Lv, G.; Hu, Y.; Zhuo, W.; Zeng, Z.; Tang, J.; Huang, Y.; Wang, F.; Wang, J., et al. Pancreatic acinar cells-derived sphingosine-1-phosphate contributes to fibrosis of chronic pancreatitis via inducing autophagy and activation of pancreatic stellate cells. Gastroenterology 2023, 165, 1488-1504. [CrossRef]
- Sukocheva, O.A.; Furuya, H.; Ng, M.L.; Friedemann, M.; Menschikowski, M.; Tarasov, V.V.; Chubarev, V.N.; Klochkov, S.G.; Neganova, M.E.; Mangoni, A.A., et al. Sphingosine kinase and sphingosine-1-phosphate receptor signaling pathway in inflammatory gastrointestinal disease and cancers: A novel therapeutic target. Pharmacol. Ther. 2020, 207, 107464. [CrossRef]
- Mo, C. The role of sphingosine kinase 1 in acute lung injury with severe acute pancreatitis. 2015, Southwest Medical University.
- Speirs, M.M.P.; Swensen, A.C.; Chan, T.Y.; Jones, P.M.; Holman, J.C.; Harris, M.B.; Maschek, J.A.; Cox, J.E.; Carson, R.H.; Hill, J.T., et al. Imbalanced sphingolipid signaling is maintained as a core proponent of a cancerous phenotype in spite of metabolic pressure and epigenetic drift. Oncotarget 2019, 10, 449-479. [CrossRef]
- Beljanski, V.; Lewis, C.S.; Smith, C.D. Antitumor activity of sphingosine kinase 2 inhibitor ABC294640 and sorafenib in hepatocellular carcinoma xenografts. Cancer Biol. Ther. 2011, 11, 524-534. [CrossRef]
- Naseh, M.; Vatanparast, J.; Rafati, A.; Bayat, M.; Haghani, M. The emerging role of FTY720 as a sphingosine 1-phosphate analog for the treatment of ischemic stroke: The cellular and molecular mechanisms. Brain Behav. 2021, 11, e02179.
- Blaho, V.A.; Hla, T. Regulation of mammalian physiology, development, and disease by the sphingosine 1-phosphate and lysophosphatidic acid receptors. Chem. Rev. 2011, 111, 6299-320. [CrossRef]
- Müller, C.A.; Belyaev, O.; Burr, W.; Munding, J.; McArthur, N.; Bergmann, U.; Werner, J.; Tannapfel, A.; Uhl, W. Effects of FTY720 and rapamycin on inflammation in taurocholate-induced acute pancreatitis in the rat. Pancreas 2012, 41, 1086-1091. [CrossRef]
- Liu, H.B.; Cui, N.Q.; Wang, Q.; Li, D.H.; Xue, X.P. Sphingosine-1-phosphate and its analogue FTY720 diminish acute pulmonary injury in rats with acute necrotizing pancreatitis. Pancreas 2008, 36, 10-15. [CrossRef]
- Okamoto, T.; Yamada, T.; Kuno, A.; Ogawa, K.; Tang, M.; Sano, H.; Ohara, H.; Nakao, H.; Kataoka, H.; Shirai, T., et al. FTY720, an immunosuppressant, attenuates chronic pancreatitis in rats by suppressing T-cell infiltration. Pancreas. 2005, 30, 64-70. [CrossRef]
- Lankadasari, M.B.; Aparna, J.S.; Mohammed, S.; James, S.; Aoki, K.; Binu, V.S.; Nair, S.; Harikumar, K.B. Targeting S1PR1/STAT3 loop abrogates desmoplasia and chemosensitizes pancreatic cancer to gemcitabine. Theranostics 2018, 8, 3824-3840.
- Yang, S.; Li, H.W.; Tian, J.Y.; Wang, Z.K.; Chen, Y.; Zhan, T.T.; Ma, C.Y.; Feng, M.; Cao, S.F.; Zhao, Y., et al. Myeloid-derived growth factor suppresses VSMC dedifferentiation and attenuates postinjury neointimal formation in rats by activating S1PR2 and its downstream signaling. Acta Pharmacol. Sin. 2024, 45, 98-111. [CrossRef]
- Cui, L.; Li, C.; Zhang, G.; Zhang, L.; Yao, G.; Zhuo, Y.; Cui, N.; Zhang, S. S1P/S1PR2 promote pancreatic stellate cell activation and pancreatic fibrosis in chronic pancreatitis by regulating autophagy and the NLRP3 inflammasome. Chem. Biol. Interact. 2023, 380, 110541. [CrossRef]
- Sun, D.; Wang, S. Sphingosine kinases are involved in the regulation of all-trans retinoic acid sensitivity of K562 chronic myeloid leukemia cells. Oncol. Lett. 2021, 22, 581. [CrossRef]
- Yi, X.; Tang, X.; Li, T.; Chen, L.; He, H.; Wu, X.; Xiang, C.; Cao, M.; Wang, Z.; Wang, Y., et al. Therapeutic potential of the sphingosine kinase 1 inhibitor, PF-543. Biomed Pharmacother. 2023, 163, 114401. [CrossRef]
- Uzunova, V.; Tzoneva, R.; Stoyanova, T.; Pankov, R.; Skrobanska, R.; Georgiev, G.; Maslenkova, L.; Tsonchev, Z.; Momchilova, A. Dimethylsphingosine and miltefosine induce apoptosis in lung adenocarcinoma A549 cells in a synergistic manner. Chem. Biol. Interact. 2019, 310, 108731. [CrossRef]
- Grbčić, P.; Tomljanović, I.; Klobučar, M.; Kraljević Pavelić, S.; Lučin, K.; Sedić, M. Dual sphingosine kinase inhibitor SKI-II enhances sensitivity to 5-fluorouracil in hepatocellular carcinoma cells via suppression of osteopontin and FAK/IGF-1R signalling. Biochem. Biophys. Res. Commun. 2017, 487, 782-788. [CrossRef]
- Dai, L.; Bai, A.; Smith, C.D.; Rodriguez, P.C.; Yu, F.; Qin, Z. ABC294640, A novel sphingosine kinase 2 inhibitor, induces oncogenic virus-infected cell autophagic death and represses tumor growth. Mol. Cancer Ther. 2017, 16, 2724-2734.
- Sekine, Y.; Suzuki, K.; Remaley, A.T. HDL and sphingosine-1-phosphate activate stat3 in prostate cancer DU145 cells via ERK1/2 and S1P receptors, and promote cell migration and invasion. Prostate 2011, 71, 690-699.
- Beljanski, V.; Knaak, C.; Zhuang, Y.; Smith, C.D. Combined anticancer effects of sphingosine kinase inhibitors and sorafenib. Invest. New Drugs 2011, 29, 1132-1142. [CrossRef]
- Tian, B.; Sui, S.; Meng, Q.; Liu, P.; Lin, H.; Xin, L.; Li, Z.; Wang, L. The influence of sphlngosine 1-phosphate on migration and invasion of pancreatic cancer PANC1 cells. Chin. J. Pancreatol. 2016, 16, 73-76.
- Deng, R.; Tang, L.; Li, D.; Li, K. Sphingosine kinase 1 change in early stage for severe acute pancreatitis. Journal of Frontiers of Medicine. 2015, 2.
- Yang, L.; Zhong, M.; Li, Y.; Zheng, X.; Yan, H.; Tang, L.; Li, D. Value of sphingosine kinase 1 and C-reactive protein in prediction of severity degree of acute pancreatitis. Medical Journal of Chinese People's Liberation Army. 2014, 26, 5.
- Li, Q.; Wang, C.; Zhang, Q.; Tang, C.; Li, N.; Li, J. The role of sphingosine kinase 1 in patients with severe acute pancreatitis. Ann. Surg. 2012, 255, 954-962. [CrossRef]
- Tang, J. Mechanism of sphingosine kinase 1 mediating acinar cell pyroptosis through PERK/TXNIP/NLRP3 signaling axis in acute pancreatitis. 2022, Huazhong University of Science and Technology.
- Yang, J.X.; Wang, M.J.C.; Qiu, Q.; Huang, Y.; Wang, Y.Q.; Pu, Q.L.; Jiang, N.; Wang, R.; Wen, L.; Zhang, X.Y., et al. Time-course lipidomics of ornithine-induced severe acute pancreatitis model reveals the free fatty acids centered lipids dysregulation characteristics. Metabolites 2023, 13. [CrossRef]
- Wollny, T.; Wątek, M.; Wnorowska, U.; Piktel, E.; Góźdź, S.; Kurek, K.; Wolak, P.; Król, G.; Żendzian-Piotrowska, M.; Bucki, R. Hypogelsolinemia and decrease in blood plasma sphingosine-1-phosphate in patients diagnosed with severe acute pancreatitis. Dig. Dis. Sci. 2022, 67, 536-545. [CrossRef]
- Xiao, J.; Lin, H.; Liu, B.; Xia, Z.; Zhang, J.; Jin, J. Decreased S1P and SPHK2 are involved in pancreatic acinar cell injury. Biomark. Med. 2019, 13, 627-637. [CrossRef]
- Konończuk, T.; Łukaszuk, B.; Żendzian-Piotrowska, M.; Dąbrowski, A.; Krzyżak, M.; Ostrowska, L.; Kurek, K. Plasma sphingolipids in acute pancreatitis. Int. J. Mol. Sci. 2017, 18. [CrossRef]
- Habtezion, A. Inflammation in acute and chronic pancreatitis. Curr. Opin. Gastroenterol. 2015, 31, 395-9. [CrossRef]
- Sun, G.; Wang, B.; Wu, X.; Cheng, J.; Ye, J.; Wang, C.; Zhu, H.; Liu, X. How do sphingosine-1-phosphate affect immune cells to resolve inflammation? Front. Immunol. 2024, 15, 1362459.
- Corbett, B.; Luz, S.; Sotuyo, N.; Pearson-Leary, J.; Moorthy, G.S.; Zuppa, A.F.; Bhatnagar, S. FTY720 (Fingolimod), a modulator of sphingosine-1-phosphate receptors, increases baseline hypothalamic-pituitary adrenal axis activity and alters behaviors relevant to affect and anxiety. Physiol. Behav. 2021, 240, 113556. [CrossRef]
- Sykes, D.A.; Riddy, D.M.; Stamp, C.; Bradley, M.E.; McGuiness, N.; Sattikar, A.; Guerini, D.; Rodrigues, I.; Glaenzel, A.; Dowling, M.R., et al. Investigating the molecular mechanisms through which FTY720-P causes persistent S1P1 receptor internalization. Br. J. Pharmacol. 2014, 171, 4797-4807. [CrossRef]
- Liu, J.; Xu, P.; Zhang, L.; Kayoumu, A.; Wang, Y.; Wang, M.; Gao, M.; Zhang, X.; Wang, Y.; Liu, G. FTY720 attenuates acute pancreatitis in hypertriglyceridemic apolipoprotein CIII transgenic mice. Shock 2015, 44, 280-286. [CrossRef]
- Zou, L.; Ke, L.; Wu, C.; Tong, Z.; Li, W.; Li, N.; Li, J. SEW2871 Alleviates the severity of caerulein-induced acute pancreatitis in mice. Biol. Pharm. Bull. 2015, 38, 1012-1019. [CrossRef]
- Leung, P.S.; Ip, S.P. Pancreatic acinar cell: its role in acute pancreatitis. Int. J. Biochem. Cell Biol. 2006, 38, 1024-1030. [CrossRef]
- Sun, B.; Chen, Z.; Chi, Q.; Zhang, Y.; Gao, B. Endogenous tRNA-derived small RNA (tRF3-Thr-AGT) inhibits ZBP1/NLRP3 pathway-mediated cell pyroptosis to attenuate acute pancreatitis (AP). J. Cell. Mol. Med. 2021, 25, 10441-10453.
- Lin, T.; Song, J.; Pan, X.; Wan, Y.; Wu, Z.; Lv, S.; Mi, L.; Wang, Y.; Tian, F. Downregulating gasdermin D reduces severe acute pancreatitis associated with pyroptosis. Med. Sci. Monit. 2021, 27, e927968. [CrossRef]
- Gao, L.; Dong, X.; Gong, W.; Huang, W.; Xue, J.; Zhu, Q.; Ma, N.; Chen, W.; Fu, X.; Gao, X., et al. Acinar cell NLRP3 inflammasome and gasdermin D (GSDMD) activation mediates pyroptosis and systemic inflammation in acute pancreatitis. Br. J. Pharmacol. 2021, 178, 3533-3552. [CrossRef]
- Ge, P.; Luo, Y.; Okoye, C.S.; Chen, H.; Liu, J.; Zhang, G.; Xu, C.; Chen, H. Intestinal barrier damage, systemic inflammatory response syndrome, and acute lung injury: A troublesome trio for acute pancreatitis. Biomed. Pharmacother. 2020, 132, 110770. [CrossRef]
- Zhou, J.; Zhou, P.; Zhang, Y.; Wang, G.; Fan, Z. Signal pathways and markers involved in acute lung injury induced by acute pancreatitis. Dis. Markers 2021, 2021, 9947047. [CrossRef]
- Yang, X; Liu, H; Cui, N; Li, D; Li, J; Wang, Q. The protective effects of sphingosine-1-phosphate on lung injury in acute necrotizing pancreatitis in rats. The Chinese Journal of General Surgery. 2008, 17, 7-232.
- Natarajan, V.; Dudek, S.M.; Jacobson, J.R.; Moreno-Vinasco, L.; Huang, L.S.; Abassi, T.; Mathew, B.; Zhao, Y.; Wang, L.; Bittman, R., et al. Sphingosine-1-phosphate, FTY720, and sphingosine-1-phosphate receptors in the pathobiology of acute lung injury. Am. J. Respir. Cell Mol. Biol. 2013, 49, 6-17.
- Wang, Z.; Liu, J.; Li, F.; Luo, Y.; Ge, P.; Zhang, Y.; Wen, H.; Yang, Q.; Ma, S.; Chen, H. The gut-lung axis in severe acute pancreatitis-associated lung injury: The protection by the gut microbiota through short-chain fatty acids. Pharmacol. Res. 2022, 182, 106321. [CrossRef]
- Kichler, A.; Jang, S. Chronic pancreatitis: Epidemiology, diagnosis, and management updates. Drugs 2020, 80, 1155-1168. [CrossRef]
- Apte, M.; Pirola, R.; Wilson, J. The fibrosis of chronic pancreatitis: new insights into the role of pancreatic stellate cells. Antioxid. Redox Signal. 2011, 15, 2711-2722. [CrossRef]
- Sobel, K.; Menyhart, K.; Killer, N.; Renault, B.; Bauer, Y.; Studer, R.; Steiner, B.; Bolli, M.H.; Nayler, O.; Gatfield, J. Sphingosine 1-phosphate (S1P) receptor agonists mediate pro-fibrotic responses in normal human lung fibroblasts via S1P2 and S1P3 receptors and Smad-independent signaling. J. Biol. Chem. 2013, 288, 14839-14851. [CrossRef]
- Yang, L.; Yue, S.; Yang, L.; Liu, X.; Han, Z.; Zhang, Y.; Li, L. Sphingosine kinase/sphingosine 1-phosphate (S1P)/S1P receptor axis is involved in liver fibrosis-associated angiogenesis. J. Hepatol. 2013, 59, 114-123.
- Zhang, X.; Ritter, J.K.; Li, N. Sphingosine-1-phosphate pathway in renal fibrosis. Am. J. Physiol. Renal. Physiol. 2018, 315, 752-756. [CrossRef]
- Wang, E.; He, X.; Zeng, M. The role of S1P and the related signaling pathway in the development of tissue fibrosis. Front. Pharmacol. 2018, 9, 1504. [CrossRef]
- Furuya, H.; Shimizu, Y.; Kawamori, T. Sphingolipids in cancer. Cancer Metastasis Rev. 2011, 30, 567-576. [CrossRef]
- Bao, Y.; Guo, Y.; Zhang, C.; Fan, F.; Yang, W. Sphingosine kinase 1 and sphingosine-1-phosphate signaling in colorectal cancer. Int. J. Mol. Sci. 2017, 18. [CrossRef]
- Spiegel, S.; Milstien, S. Functions of the multifaceted family of sphingosine kinases and some close relatives. J. Biol. Chem. 2007, 282, 2125-2129. [CrossRef]
- Barth, B.M.; Shanmugavelandy, S.S.; Kaiser, J.M.; McGovern, C.; Altınoğlu, E.; Haakenson, J.K.; Hengst, J.A.; Gilius, E.L.; Knupp, S.A.; Fox, T.E., et al. PhotoImmunoNanoTherapy reveals an anticancer role for sphingosine kinase 2 and dihydrosphingosine-1-phosphate. ACS Nano 2013, 7, 2132-2144. [CrossRef]
- Wilhelm, S.M.; Adnane, L.; Newell, P.; Villanueva, A.; Llovet, J.M.; Lynch, M. Preclinical overview of sorafenib, a multikinase inhibitor that targets both Raf and VEGF and PDGF receptor tyrosine kinase signaling. Mol. Cancer Ther. 2008, 7, 3129-3140. [CrossRef]
- Guillermet-Guibert, J.; Davenne, L.; Pchejetski, D.; Saint-Laurent, N.; Brizuela, L.; Guilbeau-Frugier, C.; Delisle, M.B.; Cuvillier, O.; Susini, C.; Bousquet, C. Targeting the sphingolipid metabolism to defeat pancreatic cancer cell resistance to the chemotherapeutic gemcitabine drug. Mol. Cancer Ther. 2009, 8, 809-820. [CrossRef]
- Fu, Y.; Ricciardiello, F.; Yang, G.; Qiu, J.; Huang, H.; Xiao, J.; Cao, Z.; Zhao, F.; Liu, Y.; Luo, W., et al. The role of mitochondria in the chemoresistance of pancreatic cancer cells. Cells 2021, 10. [CrossRef]
- Gerhard, M.C.; Schmid, R.M.; Häcker, G. Analysis of the cytochrome c-dependent apoptosis apparatus in cells from human pancreatic carcinoma. Br. J. Cancer 2002, 86, 893-898. [CrossRef]
- Yang, L.; Cao, Z.; Yan, H.; Wood, W.C. Coexistence of high levels of apoptotic signaling and inhibitor of apoptosis proteins in human tumor cells: implication for cancer specific therapy. Cancer Res. 2003, 63, 6815-6824.
- Anderson, S.; Bankier, A.T.; Barrell, B.G.; de Bruijn, M.H.; Coulson, A.R.; Drouin, J.; Eperon, I.C.; Nierlich, D.P.; Roe, B.A.; Sanger, F., et al. Sequence and organization of the human mitochondrial genome. Nature 1981, 290, 457-465. [CrossRef]
- Xue, R.; Meng, Q.; Lu, D.; Liu, X.; Wang, Y.; Hao, J. Mitofusin2 induces cell autophagy of pancreatic cancer through inhibiting the PI3K/Akt/mTOR signaling pathway. Oxid. Med. Cell. Longev. 2018, 2018, 2798070. [CrossRef]
- Jamil, M.; Cowart, L.A. Sphingolipids in mitochondria-from function to disease. Front. Cell Dev. Biol. 2023, 11, 1302472. [CrossRef]
- Duan, M.; Gao, P.; Chen, S.X.; Novák, P.; Yin, K.; Zhu, X. Sphingosine-1-phosphate in mitochondrial function and metabolic diseases. Obes. Rev. 2022, 23, e13426.
- Leanza, L.; Henry, B.; Sassi, N.; Zoratti, M.; Chandy, K.G.; Gulbins, E.; Szabò, I. Inhibitors of mitochondrial Kv1.3 channels induce Bax/Bak-independent death of cancer cells. EMBO Mol. Med. 2012, 4, 577-593.
- Leanza, L.; Romio, M.; Becker, K.A.; Azzolini, M.; Trentin, L.; Managò, A.; Venturini, E.; Zaccagnino, A.; Mattarei, A.; Carraretto, L., et al. Direct pharmacological targeting of a mitochondrial ion channel selectively kills tumor cells in vivo. Cancer Cell 2017, 31, 516-531. [CrossRef]
- Patel, S.H.; Wilson, G.C.; Wu, Y.; Keitsch, S.; Wilker, B.; Mattarei, A.; Ahmad, S.A.; Szabo, I.; Gulbins, E. Sphingosine is involved in PAPTP-induced death of pancreas cancer cells by interfering with mitochondrial functions. J. Mol. Med. (Berl). 2024, 102, 947-959. [CrossRef]
- Limbu, K.R.; Chhetri, R.B.; Oh, Y.S.; Baek, D.J.; Park, E.Y. Mebendazole impedes the proliferation and migration of pancreatic cancer cells through SK1 inhibition dependent pathway. Molecules 2022, 27. [CrossRef]
- Lépine, S.; Allegood, J.C.; Park, M.; Dent, P.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate phosphohydrolase-1 regulates ER stress-induced autophagy. Cell Death Differ. 2011, 18, 350-61. [CrossRef]
- Park, K.; Ikushiro, H.; Seo, H.S.; Shin, K.O.; Kim, Y.I.; Kim, J.Y.; Lee, Y.M.; Yano, T.; Holleran, W.M.; Elias, P., et al. ER stress stimulates production of the key antimicrobial peptide, cathelicidin, by forming a previously unidentified intracellular S1P signaling complex. Proc. Natl. Acad. Sci. U. S. A. 2016, 113, 1334-1342. [CrossRef]
- Blom, T.; Bergelin, N.; Meinander, A.; Löf, C.; Slotte, J.P.; Eriksson, J.E.; Törnquist, K. An autocrine sphingosine-1-phosphate signaling loop enhances NF-kappaB-activation and survival. BMC Cell Biol. 2010, 11, 45.
- Lee, S.Y.; Hong, I.K.; Kim, B.R.; Shim, S.M.; Sung Lee, J.; Lee, H.Y.; Soo Choi, C.; Kim, B.K.; Park, T.S. Activation of sphingosine kinase 2 by endoplasmic reticulum stress ameliorates hepatic steatosis and insulin resistance in mice. Hepatology 2015, 62, 135-146. [CrossRef]
- Gao, Z.; Janakiraman, H.; Xiao, Y.; Kang, S.W.; Dong, J.; Choi, J.; Ogretmen, B.; Lee, H.S.; Camp, E.R. Sphingosine-1-phosphate inhibition increases endoplasmic reticulum stress to enhance oxaliplatin sensitivity in pancreatic cancer. World J. Oncol. 2024, 15, 169-180. [CrossRef]
- Wang, Z.; Dong, S.; Zhou, W. Pancreatic stellate cells: Key players in pancreatic health and diseases (Review). Mol. Med. Rep. 2024, 30. [CrossRef]
- Apte, M.V.; Wilson, J.S.; Lugea, A.; Pandol, S.J. A starring role for stellate cells in the pancreatic cancer microenvironment. Gastroenterology 2013, 144, 1210-1219. [CrossRef]
- Yoshida, N.; Masamune, A.; Hamada, S.; Kikuta, K.; Takikawa, T.; Motoi, F.; Unno, M.; Shimosegawa, T. Kindlin-2 in pancreatic stellate cells promotes the progression of pancreatic cancer. Cancer. Lett. 2017, 390, 103-114. [CrossRef]
- Pothula, S.P.; Pirola, R.C.; Wilson, J.S.; Apte, M.V. Pancreatic stellate cells: Aiding and abetting pancreatic cancer progression. Pancreatology 2020, 20, 409-418. [CrossRef]
- Wang, Y.; Xu, H.; Zhang, X.; Ma, J.; Xue, S.; Shentu, D.; Mao, T.; Li, S.; Yue, M.; Cui, J., et al. The role of bile acids in pancreatic cancer. Curr. Cancer. Drug. Targets. 2024, 24, 1005-1014. [CrossRef]
- Gál, E.; Veréb, Z.; Kemény, L.; Rakk, D.; Szekeres, A.; Becskeházi, E.; Tiszlavicz, L.; Takács, T.; Czakó, L.; Hegyi, P., et al. Bile accelerates carcinogenic processes in pancreatic ductal adenocarcinoma cells through the overexpression of MUC4. Sci. Rep. 2020, 10, 22088. [CrossRef]
- Režen, T.; Rozman, D.; Kovács, T.; Kovács, P.; Sipos, A.; Bai, P.; Mikó, E. The role of bile acids in carcinogenesis. Cell. Mol. Life Sci. 2022, 79, 243. [CrossRef]
- Feng, H.Y.; Chen, Y.C. Role of bile acids in carcinogenesis of pancreatic cancer: An old topic with new perspective. World J. Gastroenterol. 2016, 22, 7463-7477. [CrossRef]
- Nagathihalli, N.S.; Beesetty, Y.; Lee, W.; Washington, M.K.; Chen, X.; Lockhart, A.C.; Merchant, N.B. Novel mechanistic insights into ectodomain shedding of EGFR Ligands Amphiregulin and TGF-α: Impact on gastrointestinal cancers driven by secondary bile acids. Cancer Res. 2014, 74, 2062-2072. [CrossRef]
- Liu, R.; Zhao, R.; Zhou, X.; Liang, X.; Campbell, D.J.; Zhang, X.; Zhang, L.; Shi, R.; Wang, G.; Pandak, W.M., et al. Conjugated bile acids promote cholangiocarcinoma cell invasive growth through activation of sphingosine 1-phosphate receptor 2. Hepatology 2014, 60, 908-918. [CrossRef]
- Nagahashi, M.; Takabe, K.; Liu, R.; Peng, K.; Wang, X.; Wang, Y.; Hait, N.C.; Wang, X.; Allegood, J.C.; Yamada, A., et al. Conjugated bile acid-activated S1P receptor 2 is a key regulator of sphingosine kinase 2 and hepatic gene expression. Hepatology 2015, 61, 1216-1226. [CrossRef]
- Yang, C.; Yuan, H.; Gu, J.; Xu, D.; Wang, M.; Qiao, J.; Yang, X.; Zhang, J.; Yao, M.; Gu, J., et al. ABCA8-mediated efflux of taurocholic acid contributes to gemcitabine insensitivity in human pancreatic cancer via the S1PR2-ERK pathway. Cell Death Discov. 2021, 7, 6. [CrossRef]
- Xu, J.; Zhou, L.; Du, X.; Qi, Z.; Chen, S.; Zhang, J.; Cao, X.; Xia, J. Transcriptome and lipidomic analysis suggests lipid metabolism reprogramming and upregulating SPHK1 promotes stemness in pancreatic ductal adenocarcinoma stem-like cells. Metabolites 2023, 13. [CrossRef]
- Maines, L.W.; Schrecengost, R.S.; Zhuang, Y.; Keller, S.N.; Smith, R.A.; Green, C.L.; Smith, C.D. Opaganib protects against radiation toxicity: Implications for homeland security and antitumor radiotherapy. Int. J. Mol. Sci. 2022, 23. [CrossRef]
- Guo, Y.X.; Ma, Y.J.; Han, L.; Wang, Y.J.; Han, J.A.; Zhu, Y. Role of sphingosine 1-phosphate in human pancreatic cancer cells proliferation and migration. Int. J. Clin. Exp. Med. 2015, 8, 20349-20354.
- Yang, M.; Wang, L.; Li, S. Clinical significanceof SPHK1 genein pancreatic adenocarcinoma and its effect on proliferation and migration of pancreatic cancer cells. Acta Medicinae Universitatis Scientiae et Technologiae Huazhong. 2021, 50, 8.
- Cai, D.; Su, Y.; Tang, J. Department effect of sphingosine kinase 1 on the proliferation and apoptosis in human pancreatic cancer cell line SW1990. Hainan Medical Journal. 2017, 28, 4.
- Aoki, H.; Aoki, M.; Katsuta, E.; Ramanathan, R.; Idowu, M.O.; Spiegel, S.; Takabe, K. Host sphingosine kinase 1 worsens pancreatic cancer peritoneal carcinomatosis. J. Surg. Res. 2016, 205, 510-517. [CrossRef]
- Karam, M.; Ives, A.; Auclair, C. Is Sphingosine-1-phosphate a regulator of tumor vascular functionality? Cancers (Basel) 2022, 14.
- Gu, Y.; Shea, J.; Slattum, G.; Firpo, M.A.; Alexander, M.; Mulvihill, S.J.; Golubovskaya, V.M.; Rosenblatt, J. Defective apical extrusion signaling contributes to aggressive tumor hallmarks. eLife 2015, 4, e04069. [CrossRef]
- Yamada, T.; Okajima, F.; Ohwada, S.; Kondo, Y. Growth inhibition of human pancreatic cancer cells by sphingosylphosphorylcholine and influence of culture conditions. Cell. Mol. Life Sci. 1997, 53, 435-441. [CrossRef]
- Ohno, Y.; Kanematsu, T. Annular pancreas causing localized recurrent pancreatitis in a child: Report of a case. Surg. Today 2008, 38, 1052-1055. [CrossRef]
- Gutta, A.; Fogel, E.; Sherman, S. Identification and management of pancreas divisum. Expert Rev Gastroenterol Hepatol. 2019, 13, 1089-1105. [CrossRef]
- Dhakal, B.; Pant, S.; Choudhary, S.; Basnet, B.; Neupane, S. Dorsal pancreatic agenesis: A case report. Ann. Med. Surg. (Lond). 2023, 85, 2949-2952. [CrossRef]
- Scharfmann, R. Control of early development of the pancreas in rodents and humans: implications of signals from the mesenchyme. Diabetologia 2000, 43, 1083-1092. [CrossRef]
- Kim, J.W.; Kim, Y.W.; Inagaki, Y.; Hwang, Y.A.; Mitsutake, S.; Ryu, Y.W.; Lee, W.K.; Ha, H.J.; Park, C.S.; Igarashi, Y. Synthesis and evaluation of sphingoid analogs as inhibitors of sphingosine kinases. Bioorg. Med. Chem. 2005, 13, 3475-85. [CrossRef]
- Antoon, J.W.; White, M.D.; Burow, M.E.; Beckman, B.S. Dual inhibition of sphingosine kinase isoforms ablates TNF-induced drug resistance. Oncol. Rep. 2012, 27, 1779-86. [CrossRef]
- Shi, Y.; Qiao, J.; Mu, B.; Zuo, B.; Yuan, J. 3-(2-amino-ethyl)-5-[3-(4-butoxyl-phenyl)-propylidene]-thiazolidine-2,4-dione (K145) ameliorated dexamethasone induced hepatic gluconeogenesis through activation of Akt/FoxO1 pathway. Biochem. Biophys. Res. Commun. 2017, 493, 286-290. [CrossRef]
- Sandborn, W.J.; Vermeire, S.; Peyrin-Biroulet, L.; Dubinsky, M.C.; Panes, J.; Yarur, A.; Ritter, T.; Baert, F.; Schreiber, S.; Sloan, S., et al. Etrasimod as induction and maintenance therapy for ulcerative colitis (ELEVATE): two randomised, double-blind, placebo-controlled, phase 3 studies. Lancet 2023, 401, 1159-1171.
- Ogasawara, A.; Takeuchi, H.; Komiya, H.; Ogawa, Y.; Nishimura, K.; Kubota, S.; Hashiguchi, S.; Takahashi, K.; Kunii, M.; Tanaka, K., et al. Anti-inflammatory effects of siponimod on astrocytes. Neurosci. Res. 2022, 184, 38-46. [CrossRef]
- Sandborn, W.J.; Feagan, B.G.; Wolf, D.C.; D'Haens, G.; Vermeire, S.; Hanauer, S.B.; Ghosh, S.; Smith, H.; Cravets, M.; Frohna, P.A., et al. Ozanimod induction and maintenance treatment for ulcerative colitis. N. Engl. J. Med. 2016, 374, 1754-1762. [CrossRef]
- Mork, B.E.; Lamerand, S.R.; Zhou, S.; Taylor, B.K.; Sheets, P.L. Sphingosine-1-phosphate receptor 1 agonist SEW2871 alters membrane properties of late-firing somatostatin expressing neurons in the central lateral amygdala. Neuropharmacology 2022, 203, 108885. [CrossRef]
- Markham, A. Ponesimod: First approval. Drugs 2021, 81, 957-962. [CrossRef]
- Hafizi, R.; Imeri, F.; Stepanovska Tanturovska, B.; Manaila, R.; Schwalm, S.; Trautmann, S.; Wenger, R.H.; Pfeilschifter, J.; Huwiler, A. Sphk1 and Sphk2 differentially regulate erythropoietin synthesis in mouse renal interstitial fibroblast-like cells. Int. J. Mol. Sci. 2022, 23. [CrossRef]
- Paradiso, E.; Lazzaretti, C.; Sperduti, S.; Antoniani, F.; Fornari, G.; Brigante, G.; Di Rocco, G.; Tagliavini, S.; Trenti, T.; Morini, D., et al. Sphingosine-1 phosphate induces cAMP/PKA-independent phosphorylation of the cAMP response element-binding protein (CREB) in granulosa cells. Mol. Cell. Endocrinol. 2021, 520, 111082. [CrossRef]
- Cencetti, F.; Bernacchioni, C.; Tonelli, F.; Roberts, E.; Donati, C.; Bruni, P. TGFβ1 evokes myoblast apoptotic response via a novel signaling pathway involving S1P4 transactivation upstream of Rho-kinase-2 activation. Faseb. J. 2013, 27, 4532-4546.
- Di Pardo, A.; Castaldo, S.; Amico, E.; Pepe, G.; Marracino, F.; Capocci, L.; Giovannelli, A.; Madonna, M.; van Bergeijk, J.; Buttari, F., et al. Stimulation of S1PR5 with A-971432, a selective agonist, preserves blood-brain barrier integrity and exerts therapeutic effect in an animal model of Huntington's disease. Hum. Mol. Genet. 2018, 27, 2490-2501.
Figure 1.
Schematic diagram of S1P signaling pathway.
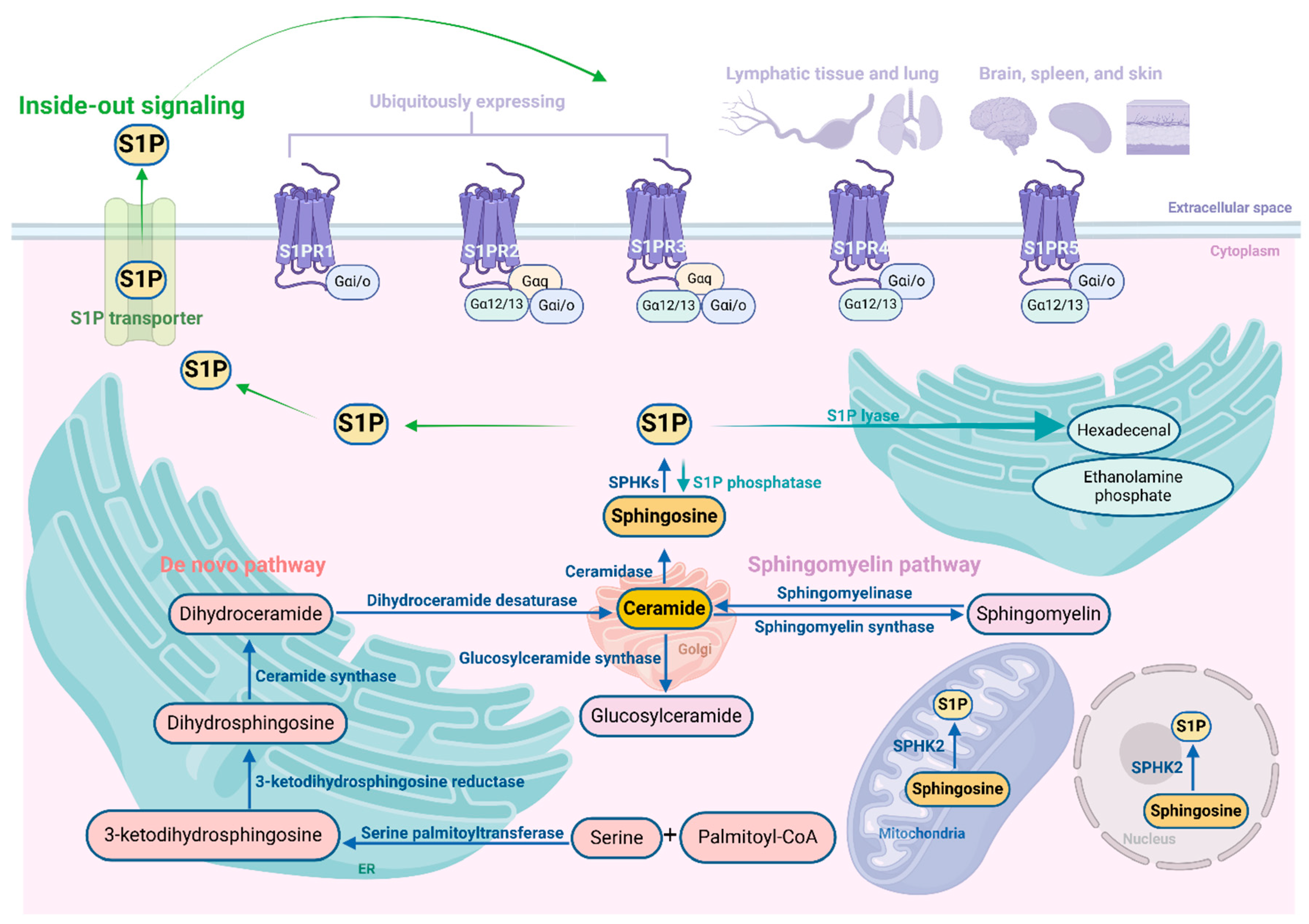
Figure 2.
Role and relative mechanisms of S1P signaling in pancreatic diseases.
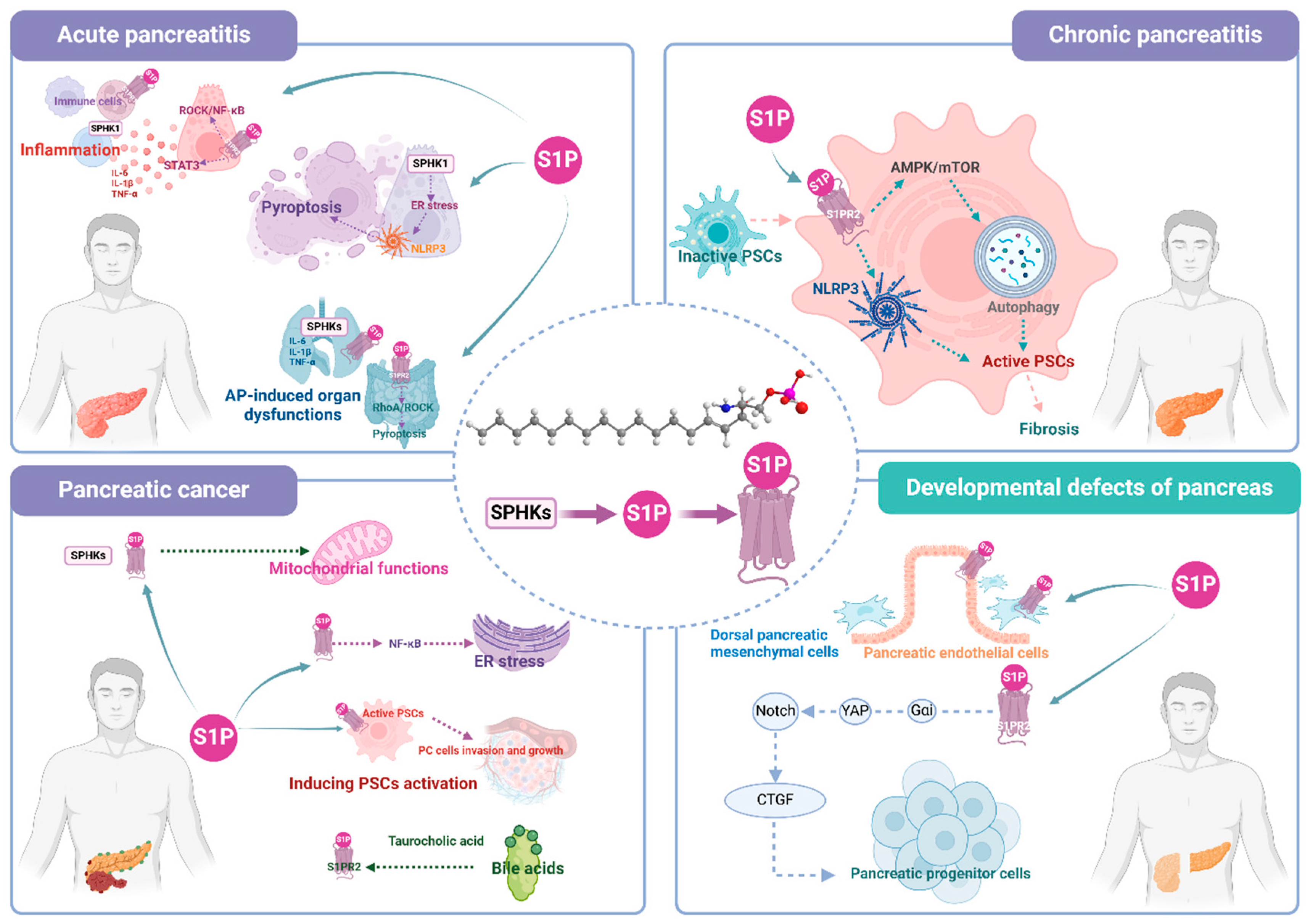
Table 2.
Functions of SPHKs/S1P/S1PRs signaling in pancreatic diseases.
| Object | Disease | Subject | Model | Treatment | Effects | Mechanism | Ref. |
|---|---|---|---|---|---|---|---|
| SPHK1 | AP | Severe AP patients | / | / | ↑ Peripheral blood leukocytes SPHK1 in the early stage of severe AP patients | [80] | |
| SPHK1 | AP | Mild and severe AP patients | / | / | ↑ Peripheral blood leukocytes SPHK1 in the early stage of mild and severe AP patients. | / | [81] |
| SPHK1/S1PR3 | AP | Severe AP patients | / | / | ↑ SPHK1 and S1PR3 in the early stage of severe AP patients, but recovered to normal level at the restoration stage. SPHK1 expression of peripheral neutrophils, monocytes, and CD4+ T lymphocytes was positively correlated with the APACHE score of severe AP patients | [82] | |
| S1P | AP | Wistar rats | Cerulein | / | ↑Pancreatic S1P | / | [25] |
| S1P | AP | Severe AP patients | / | / | ↓Plasma S1P in severe AP patients | / | [85] |
| SPHK2/S1P | AP | AP patients | / | / | ↓Serum S1P in AP patients | / | [86] |
| Female C57BL/6 mice | Cerulein | / | ↓Serum S1P, pancreatic S1P and SPHK2 | ||||
| AR42J cells | Cerulein | / | ↓S1P, SPHK2 | ||||
| S1P | AP | AP patients | / | / | ↑ Plasma S1P in the early stage (days 1 and 3) of mild AP patients, then returned to normal level at day 7 | / | [87] |
| ↓ Plasma S1P in the early stage (days 1 and 3) of severe AP patients, then returned to normal level at day 7 | |||||||
| S1PR2 | AP | Male ICR mice | Cerulein, injection of TCA into the pancreatic duct | / | ↑Pancreatic S1PR2 | S1PR2 regulated ROCK/NF-κB signaling | [26] |
| Inhibition of S1PR2 by JTE-013, knockdown of S1pr2 | ↓Pancreatic damage, NF-κB | ||||||
| PACs, primary peritoneal macrophages, RAW264.7 cells | TCA | / | ↑S1PR2 | ||||
| Inhibition of S1PR2 by JTE-013, knockdown of S1pr2 | ↓NF-κB, macrophage recruitment and macrophage polarization toward the M1 phenotype | ||||||
| S1PR | AP | Female apolipoprotein CIII transgenic C57BL/6J mice | Cerulein | Modulation of S1PR by FTY720 | ↓Pancreatic pathological injury, MCP-1 | / | [92] |
| S1PR | AP | Wistar rats | Injection of 5% sodium taurocholate into the biliopancreatic duct | Modulation of S1PR by FTY720 | ↓IL-6, IL-10 and TNF-α in plasma/serum, necrosis, inflammation and number of CD4+/CD8+ cells in pancreas | / | [66] |
| S1PR1 | AP | Male ICR mice | Cerulein | Activation of S1PR1 by SEW2871 | ↓Pathological injury of pancreas, serum amylase, lipase, IL-6 and TNF-α, pancreatic MPO, number of CD45+CD4+ T lymphocytes in the peripheral blood, infiltration of CD4+ T cells in pancreas, inflammation | S1PR1 regulated the phosphorylation of STAT3 | [93] |
| SPHK1 | AP | Male C57BL/6J mice | Cerulein | / | ↑Pancreatic SPHK1 | SPHK1 regulated PERK/TXNIP/NLRP3 signaling | [83] |
| Knockout of Sphk1 | ↓Pancreatic damage, pyroptosis, endoplasmic reticulum stress | ||||||
| 266-6 cells | CCK8 | / | ↑ SPHK1 | ||||
| Knockdown of Sphk1 | ↓LDH, pyroptosis, endoplasmic reticulum stress | ||||||
| SPHK1 | AP | SD rats | Injection of 5% sodium taurocholate into the biliopancreatic duct | / | ↑SPHK1 in pancreas and peripheral blood neutrophils | / | [61] |
| Inhibition of SPHK1 by SKI 5c | ↑Survival rate | ||||||
| ↓Serum amylase, lipase, TNF-α and IL-1β; MPO in the lung, protein content of bronchoalveolar lavage fluid, pathological injury of the lung | |||||||
| S1P/S1PR | AP | Male Wistar rats | Injection of 5% sodium taurocholate into the biliopancreatic duct | 100 μg/kg S1P, i.p., once, modulation of S1PR by FTY720 | ↓IL-1β, IL-6, TNF-α, protein concentration, total cell count, PMN percentage in bronchoalveolar lavage fluid, NF-κB activity of alveolar macrophages, capillary leakage and MPO in the lung. Pathological injury of pancreas and lung | / | [67] |
| S1P | AP | Wistar rats | Injection of 5% sodium taurocholate into the biliopancreatic duct | 50 μg/kg S1P, i.p., once | ↓Serum amylase and lipase, protein concentration, leucocyte and neutrophil count of bronchoalveolar lavage fluid, MPO of the lung tissue, pathological injury of pancreas and lung | / | [100] |
| S1PR2 | AP | Male C57BL/6 mice | Cerulein+ lipopolysaccharide | Inhibition of S1PR2 by JTE-013 | ↓Pathological injury of pancreas, inflammation, intestinal tissue injury and pyroptosis | S1PR2 regulated RhoA/ROCK signaling | [57] |
| THP-1 cells | Lipopolysaccharide +ATP | Knockdown or overexpression of S1pr2 | S1PR2 positively regulated macrophage pyroptosis, and negatively regulated cohesin expression in FHC cells after co-culture of FHC and THP-1 cells | ||||
| S1P/S1PR2 | CP | Male Wistar rats | Dibutyltin dichloride | / | ↑Plasma and pancreatic S1P | S1P binding to S1PR2 promoted PSC activation and pancreatic fibrosis in CP by regulating autophagy and the NLRP3 inflammasome sequentially | [71] |
| 200 μg/kg/day S1P, i.p., 4 weeks | ↑Pancreatic damage, fibrosis, autophagy, S1PR2, NLRP3 | ||||||
| PSCs | / | 5 μM S1P, 24h | ↑PSC activation, autophagy, S1PR2, NLRP3 | ||||
| Inhibition of S1PR2 by JTE-013, knockdown of S1pr2 | ↓PSC activation, autophagy, NLRP3 | ||||||
| SPHK1/S1P/S1PR2 | CP | C57BL/6 mice | Cerulein, pancreatic duct ligation | / | ↑Serum S1P, pancreatic SPHK1, S1PR2 | PACs-derived S1P contributed to fibrosis of CP via inducing autophagy and activation of PSCs through the AMPK/mTOR signaling | [59] |
| Knockout of Sphk1, inhibition of SPHK1 or S1PR2 by PF-543 and JTE-013, respectively | ↓Pathological injury of pancreas, fibrosis, inflammation, atrophy of the pancreas | ||||||
| PACs | CCK, hypoxia | / | ↑SPHK1, S1P in PACs | ||||
| Knockout of Sphk1 in PACs, PSCs treated with S1P, knockdown of S1pr2 or inhibition of S1PR2 in PSCs | SPHK1/S1P/S1PR2 signaling positively regulated activation and autophagy of PSCs | ||||||
| S1PR | CP | Male Wistar rats | Male WBN/Kob rats | Modulation of S1PR by FTY720 | ↑Pancreas weights | S1PR regulated IFN-γ and TGF-β1 expression | [68] |
| ↓Pancreatic MPO activity, hydroxyproline content, pathological injury of pancreas, inflammation, fibrosis, necrosis, infiltration of CD4 and CD8-positive T cells in the pancreas | |||||||
| S1P | PC | C57BL/6 mice | PANC-2 cells, subcutaneous | PhotoImmunoNanoTherapy | ↓Tumor volume | / | [112] |
| Athymic nude mice | Human BxPC-3-GFP cells, orthotopic | PhotoImmunoNanoTherapy | ↑Surem S1P | ||||
| SPHK2 | PC | BxPC-3 cells | / | Inhibition of SPHK2 by ABC294640, sorafenib | ↓Cell viability, ↑Cell apoptosis | / | [78] |
| SCID mice | Human BxPC-3 cells, subcutaneous | Inhibition of SPHK2 by ABC294640, sorafenib | ↓Tumor growth, ↑Tumor cell apoptosis | ||||
| SPHK2 | PC | SCID mice | Human BxPC-3 cells, subcutaneous | Inhibition of SPHK2 by ABC294640 | ↓Tumor growth | / | [63] |
| SPHK1 | PC | BxPC-3 or PANC-1 cells | Gemcitabine | Inhibition of SPHK1 by SKI or knockdown of Sphk1 | ↓Cell viability | / | [114] |
| Overexpression of Sphk1 | ↑Cell viability | ||||||
| SPHKs/S1P | PC | C57BL/6 mice | PDAC cells, metastatic | PAPTP + ABC294640 | ↓Tumor growth | SPHKs regulated the mitochondrial Kv1.3 ion | [124] |
| MIA PaCa-2 cells | Inhibition of Kv1.3 by PAPTP | / | ↑Sphingosine, S1P-phosphatase | ||||
| Inhibition of S1P-phosphatase by XY-14 | ↓Sphingosine, death of pancreas cancer cells | ||||||
| Inhibition of SPHK2 by ABC294640 | ↑Sphingosine, death of pancreas cancer cells | ||||||
| SPHK1/S1P | PC | MIA PaCa-2, PANC-1 or Capan-1 cells | / | Inhibition of SPHK1 by mebendazole | ↓Cell migration, proliferation and viability, ↑Cell mitochondrial apoptosis | SPHK1 regulated the intrinsic mitochondrial pathway, JAK2/STAT3 and FAK/Vimentin signaling pathway | [125] |
| SPHKs | PC | PSN1 cells | / | Inhibition of SPHKs by SKI-II | ↓Cell proliferation | SPHKs regulated the ratio of S1P/C16 Cer | [62] |
| S1PR1 | PC | MIA PaCa-2 or PAN02 cells | / | Inhibition of S1PR1 by FTY720 | ↓Cell migration, proliferation | S1PR1 regulated the mitochondrial membrane potential, S1PR1-STAT3 loop, and epithelial to mesenchymal transition | [69] |
| MIA PaCa-2 or PAN02 cells | Gemcitabine | Inhibition of S1PR1 by FTY720 | ↑Cell death, ↓Cell proliferation | ||||
| NOD.CB17-Prkdcscid/J mice or C57BL/6 mice | luciferase-tagged MIA PaCa-2 cells | Treatment of FTY720 and gemcitabine | ↓Tumor volume, tumor cell metastasis, proliferation, ↑Cell apoptosis or necrosis | ||||
| SPHK2 | PC | MIA PaCa-2 or PANC-1 cells | Oxaliplatin | Inhibition of SPHK2 by ABC294640 or knockdown of Sphk2 | ↓Cell viability, ↑ER stress | SPHK2 modulated ER stress, thereby regulating PERK/eIF2α phosphorylation and ICD | [130] |
| Male C57BL/6 mice | KPC cells, orthotopic | Oxaliplatin +ABC294640 | ↓Tumor weight, ↑ICD | ||||
| S1P/S1PR2 | PC | PANC-1 or L3.6 cells | Conditioned media collected from S1P-treated PSCs | / | ↑Cell proliferation, migration | S1P regulated tumor microenvironment and the interactions of PSCs with cancer cells | [28] |
| PANC-1 cells | Conditioned media collected from S1P-treated PSCs | Inhibition of S1PR2 by JTE-013, Knockdown of S1pr2 | ↓Cell migration, invasion | ||||
| Male nude mice | L3.6 cells + PSCs or ASPC-1 cells + PSCs, orthotopic | / | ↑Tumor volume, weight, metastasis | ||||
| Knockdown of S1pr2 | ↓Tumor volume, weight, metastasis | ||||||
| S1PR2 | PC | PANC-1 or CFPAC-1 cells | Gemcitabine | Inhibition of S1PR2 by JTE-013 | ↑Gemcitabine -induced apoptosis, ↓Cell migration, invasion | TCA contributes to gemcitabine ineffectiveness by activating S1PR2/ERK signaling | [142] |
| S1P/S1PR2 | PC | Male C57BL/6 mice | PANC-2-luc cells + bile duct ligation, orthotopic,metastatic | Activation of S1PR2 by CYM5520 | ↑Tumor growth | / | [58] |
| Treated with anti-S1P-antibody, sphingomab | ↓Survival, ↑tumor burden | ||||||
| PANC-2-luc or ASPC-1 cells | TCA, CYM5520 | / | ↑Cell growth | ||||
| Inhibition of S1PR2 by JTE-013 | ↓Cell growth, migration, viability | ||||||
| Inhibition of all S1PRs except S1PR2 by FTY720 | ↑Cell viability | ||||||
| SPHKs | PC | PAN02 cells | / | Knockdown of Sphk1 | ↑Cell proliferation, migration | / | [27] |
| PAN02 cells | / | Knockdown of Sphk2 | ↓Cell proliferation, migration | ||||
| Male C57BL/6 mice | Sphk1 KO PAN02 cells | / | ↑Survival | ||||
| Male C57BL/6 mice | Sphk2 KO PAN02 cells | / | ↓Survival | ||||
| Sphingosine | PC | Male nude mice | PANC-1 or PANC-1 TRCs, orthotopic | / | Sphingosine significantly decreased in TRCs | / | [143] |
| SPHK2 | PC | Male C57BL/6 mice | PAN02 cells | Inhibition of SPHK2 by ABC294640 | ↓Tumor growth | / | [144] |
| S1P | PC | Capan-1 or PANC-1 cells | / | S1P, 0.5 and 1 μΜ; Inhibition Src by PP2 | ↓Cell proliferation, migration | / | [145] |
| SPHK1 | PC | BxPC-3 cells | / | Overexpression of Sphk1 | ↑Cell proliferation, migration | / | [146] |
| SPHK1 | PC | SWl990 cells | / | Inhibition of SPHK1 by DMS | ↓Cell proliferation, ↑Cell apoptosis | / | [147] |
| Activation of SPHK1 by phorbol 12-myristate13-acetate | ↑Cell proliferation, ↓Cell apoptosis | ||||||
| S1P/S1PRs | PC | PANC-1 cells | / | S1P, 20-200 nΜ; Inhibition of S1PRs by VPC23019 | ↓Cell migration, invasion | / | [79] |
| S1P | PC | Female athymic nude mice | PANC-2-SAL, TPAN1-IFA, metastatic | Inhibition of S1P Lyase by LX2931 | ↓Tumor volume, Hypoxia marker | S1P regulated tumor hypoxia and therapy efficacy in solid tumors | [149] |
| S1PR2 | PC | Nude mice | HPAF II tumor cells expressing S1PR2-GFP | / | ↓Tumor size and metastatic frequency | / | [150] |
| S1P | PC | PANC-1 or MIA PaCa-2 cells | / | S1P, 0-10 μΜ | ↓DNA synthesis | / | [151] |
| SPHK1 | PC | C57BL/6 mice | PANC-2-luc cells | Knockout of Sphk1 | ↓Tumor burden, cell proliferation | / | [148] |
| S1P | DDP | In vitro culture of pancreatic explants of mice embryos | Cdh2–/– (N-cadherin knockout) | S1P, 0.1 µM | ↑ Early morphogenesis of the dorsal pancreas, formation of the dorsal pancreatic bud, dorsal pancreatic mesenchymal cell proliferation, development of dorsal pancreatic endoderm, mesenchyme and endothelium | S1P stimulated mesenchymal cell proliferation | [29] |
| S1PR | DDP | C57BL6 mice embryos | / | Knockout of S1pr | ↓ Volume of the dorsal and ventral pancreata, proliferation of Pdx1+ progenitors | S1PR regulated proliferation rate of Pdx1+ progenitors, hypervascularization | [30] |
| ↑ Aberrant development of the pancreatic endoderm, vascular density of dorsal pancreas | |||||||
| S1PR2 | DDP | C57BL/6J mice embryos | / | Knockout of S1pr2 | ↓ Development of pancreas, survival and commitment of pancreas progenitors, endocrine and acinar differentiation | S1PR2 regulated Gαi-mediated YAP stabilization and Notch attenuation, then partially activated CTGF | [56] |
| ↑ Lpar1, S1PR3 in the epithelium |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



