
Submitted:
09 October 2024
Posted:
10 October 2024
Read the latest preprint version here
Abstract
Industrial hemp (Cannabis sativa L.) is primarily a dioecious plant, and monoecious varieties developed for high yield. Production practices vary by varieties prompting the need for the present study aimed to describe the floral characteristics and evaluate pollen quantity and viability of a monoecious (Henola) and three dioecious (CFX-2, Canda, and Joey) varieties. All varieties have five lanceolate anthers, basifixed to a threadlike filament. Anther length was significantly different but not anther width among varieties. The longest length (0.38 ± 0.046 cm) was recorded in Henola, and shortest (0.34 ± 0.043 cm) in CFX-2. Anther width ranged from 0.088 ± 0.0024 to 0.095 ± 0.0021cm. Pollen grains were triporate and spheroidal in shape and size differed significantly with the largest in Joey (27.83 ± 0.78 μm) and Henola (27.489 ± 0.99 μm), and smallest in Canda (22.04 ± 0.56 μm). The number of pollen grains differed significantly among varieties ranging from 29,183 in Henola to 104,548 in Joey. Even though Henola recorded the lowest pollen number it had the highest percentage (69.3%) of viable pollen and Canda recorded the lowest. There was a moderate, positive and significant relationship (r = 0.496) between anther length and the number of pollen grains in Joey. The relationship in Henola was moderate and non-significant (r = 0.356), weak and non-significant in Canda (r = 0.188), and in CFX-2 (r = 0.037). The findings from our study provide information for growers and researchers on hemp breeding and cultivation practices that may contribute to prevent cross pollination.
Keywords:
industrial hemp
; hemp
; hemp flower
; pollen
; pollen viability
; pollen number
1. Introduction
Industrial hemp (Cannabis sativa L.) is a plant of the Cannabaceae family cultivated globally for its multipurpose applications. North America and Europe are currently leading in the market largely due to favorable regulatory environments and high consumer awareness [1,2,3,4]. In the United States, the passage of the 2018 Farm Bill [5,6] propelled the crop’s reintroduction in the agroecosystems with the 2022 production valued at $238 million [7]. The fast-increasing popularity has been attributed to several factors including the recognition by growers of the crop’s diverse uses such as an alternative cover crop, in addition to its harmony with the environment, and the many economic opportunities including its potential as a value-added enterprise for entrepreneurs and businesses [8,9].
Hemp can be dioecious or monoecious and most varieties are photoperiod sensitive triggering the transition from the vegetative to the flowering growth stage in response to shortening daylight hours [10]. When growing the crop for medicinal purposes female flowers are highly sought after since they produce higher quantities of cannabinoids particularly cannabidiol (CBD). Cannabidiol continues to gain popularity globally because of its antioxidative, anxiolytic, and anti-inflammatory effects and its pharmacological potential in managing and treating neurological disorders including epilepsy and Alzheimer’s [11,12,13,14]. Hemp is also experiencing a resurgence in natural fiber production with applications in plastics, rubber, paper, and bioenergy among others [15,16,17,18,19,20]. The seeds are a valuable source of minerals, macronutrients, and phytonutrients [21,22], and have been reported to contain about 250–350 g/kg lipids, 20–25 g/kg protein, and 20–30 g/kg carbohydrates [23,24,25]. The lipid portion is rich in essential fatty acids and contains large amounts of omega-3 and omega-6 [26,27]. Several studies have reported extracts from leaves, inflorescences, and seeds exhibited antioxidant, antibacterial, and cytotoxic activity against cancer cells [28,29,30,31].
Despite the versatility of hemp, its cultivation is currently facing numerous challenges including the potential for cross-pollination. The plant is anemophilous, and the smooth exine layer of the pollen in conjunction with the lightweight facilitates genetic transfer for seed formation [32,33,34]. Dispersal of pollen during cultivation for CBD could result in pollination thereby altering the quality and quantity of CBD content [35]. Several tactics have been proposed and the most practiced include indoor cultivation with air filtration systems, planting feminized seeds or vegetatively propagated female clones [36,37], and manual removal of male plants before anthesis which is laborious and time-consuming. Unfortunately, these tactics have not yielded satisfactory outcomes since pollination by drifting pollen from neighboring grain, fiber, or CBD farms with male plants remains a serious limitation with reports of disputes and lawsuits between farmers [38,39].
Although the morphology of the hemp plant has been described [40,41,42,43], there is limited knowledge of floral variation among varieties that could influence and impact successful hemp cultivation and breeding. Most studied varieties in the United States are for CBD production. This study aims to examine and describe the flower and pollen of hemp and evaluate pollen quality (pollen viability) and quantity (number of pollen grains produced) of four hemp grain varieties that are either dioecious or monoecious.
2. Results and Discussion
2.1. Floral Characteristics of the Hemp Plant
Hemp is primarily a dioecious plant with male and female flowers on separate plants, a unique characteristic as only about 6% of plants exhibit this trait [44]. In this study, we used CFX-2, Canda, and Joey all dioecious varieties (Figure 1A), and Henola a monoecious variety with male and female flowers coexisting on the same plant (Figure 1B), bred in Europe primarily for higher grain yield [45]. Generally, the female and male plants are morphologically indistinguishable before flowering. However, their flowers have distinct morphologies with the female flower featuring dense leafy inflorescences compared to the male flower (Figure 1A) [41]. At flowering, the male plants become slender and taller with fewer leaves than the female plants which are shorter with a bunch of leaves associated with the terminal inflorescence (Figure 1A). In both the male and female plants, the flowers develop sequentially, with immature flowers located at the upper end of the inflorescence (Figure 1A and B).
The male plants and its flowers in dioecious varieties and male flowers in monoecious varieties eventually die after shedding pollen, and female flowers survive to produce seeds (Figure 2).
2.2. Anther Shape and Size
Anther size is vital since it directly impacts the amount of pollen produced, which can significantly influence pollination efficiency, reproductive success, and eventually crop yield [46]. Our findings reveal that anther size varied among varieties with a significant difference (F3, 232 =9.6 p < 0.0001) in length (Figure 4). The longest length was recorded in Henola (0.38 ± 0.046 cm), Joey (0.37 ± 0.054 cm), and Canda (0.37 ± 0.046 cm) and the shortest in CFX-2 (0.34 ± 0.043 cm).
Anther width was wider for Canda (0.095 ± 0.0021cm), and CFX-2 (0.094 ± 0.0022 cm) however, this was not significantly different (F3, 232 =1.9 p < 0.1503) from Henola and Joey with a much slender width of 0.088 ± 0.0024 cm and 0.089 ±0.0029) respectively. Based on anther width and length, it would appeared that these differences bear little or no ecological significance driven by the varieties. All hemp varieties have five lanceolate anthers, basifixed to a threadlike filament, dehiscence was observed to be latrorse with the anthers splitting open toward the sides to release pollen (Figure 3).
2.3. Pollen Shape, Size, Number, and Viability
Pollen shape and size influence the dispersal capability of pollen. Generally, anemophilous plants produce copious amounts of pollen, for example, a single hemp flower can generate about 350,000 pollen grains [47]. The abundant production is to offset the low efficiency of wind pollination [46] and to ensure successful pollination since some pollen are dispersed to the surrounding environment and apparently play no role in fertilization. All four hemp varieties were predominantly spheroidal in shape with three circular apertures (triporate) (Figure 5). Pollen size was significantly different (F3, 39= 13.4, p < 0.0001) with the largest size recorded in Joey (27.83 ± 0.78 μm) and Henola (27.489 ± 0.99 μm), followed by CFX-2 (26.75 ± 0.53 μm). The smallest pollen size was in Canda (22.04 ± 0.56 μm) and we can assume that pollen from this variety may be disperse further and more efficiently by wind.
Although there were no differences in the number of anthers, there was a significant difference in anther size, and in the number of pollen grains produced (F3, 70 = 20.9 p < 0.0001) among the four hemp varieties. The lowest number of pollen was recorded in Henola (Figure 6) the variety with the longest anther length (Figure 4).
Some studies [46,48,49] have documented a positive correlation between anther size and the quantity of pollen produced. In this study, data from all hemp varieties combined showed a weak and non-significant correlation between anther length (r = 0.19, p = 0.098) and the number of pollen grains produced (Figure 7a). Similarly, there was a non-significant correlation between anther width (r = 0.16, p = 0.175) and the number of pollen produced (Figure 7b). However, analysis of individual varieties showed a moderate, albeit positive and significant correlation (r = 0.496; p = 0.043) between anther length and the number of pollen grains in Joey (Figure 8d). All the other three varieties showed a non-significant relationship (r = 0.356; p = 0.124) in Henola, (r = 0.188; p = 0.47) in Canda and (r = 0.037; p = 0.877) in CFX-2 (Figure 8a, b and c). Henola being monoecious does not need a high number of pollen grains to ensure fertilization; it can be presumed that long anthers in Henola make it easier for the fewer pollen it produced to be retained within the flower. Seemingly, shorter anthers with more pollen would be an enabling mechanism for more efficient dispersal even though the relationship between these two variables did not appear significant for Canda and CFX-2.
The relationship between anther width and the number of pollen grains was negative and non-significant (r = -0.312, p = 0.18) in Henola (Figure 9c). The relationship was slightly, positive and non-significant in Canda (r = 0.1940, p = 0.456), and CFX-2 (r = 0.081, p = 0.734) (Figure 9a and b), moderate, positive and non-significant in Joey (r2 = 0.403, p = 0.109) (Figure 9d).
Generally, most insect-pollinated plants differ from wind pollinated plants in that insect pollinated plants produce nectar, have brightly colored flowers, and produced large sticky pollen grains with some possessing feeding and pollinating anthers (anther dimorphism). The pollinating anthers have been reported to be longer and produce more viable pollen grains than the feeding anthers [50,51]. It was argued that the longer anthers were to enable successful pollination while the shorter anthers were to fulfill the demand as a food resource for pollinators. Wind pollinated plants such as hemp do not produce nectar, but produce abundant small light weight pollen grains from their anthers. All hemp varieties in this study had five anthers and did not exhibit anther dimorphism. Henola, Joey and Canda had longest anther and the shortest was in CFX-2. According to [52], pollinators preferred pollen from plants with smaller pollen grains and rarely collected pollen from those with larger grains as bigger grains had relatively reduced nutritional content. Our findings do not support this because Joey variety with the same anther length and the same pollen size as Henola was reported by [53] to be the most preferred by bees, despite expressing lower protein, amino acid, and saturated and monosaturated fatty acid content compared to Henola. It is worth noting that although bees would collect pollen from hemp flowers, they are not attracted to the female flowers.
Figure 10 shows the Ampha Z32 output chart for pollen viability measure presented as a density scatter plot for each of the four hemp varieties showing percent viable pollen to the top right and non-viable pollen to the top left. Generally, the number of pollen grains produced is usually more compared to the number of viable pollen grains since only a fraction of the total pollen grains produced are capable of fertilizing an ovule. Knowledge of the viability of pollen is vital as it provides insight for species fitness, and survival of the next generation because of its direct connection to fertilization efficiency and the number of offspring.
Henola produced the lowest number of pollen grains (Figure 6), compared to the other varieties, however, the percentage of viable pollen was significantly (F3, 70 = 10.74, p < 0.0001) higher in Henola (Figure 11) compared to the others. Similar results of reduced pollen numbers for monoecious varieties have been reported [32]. The high pollen viability reported in this study for Henola could be associated with its high protein, amino acid, and saturated and monosaturated fatty acid contents reported by [53].
It is important to note that pollen viability and pollen size may have a direct impact on pollen dispersal as small size, lightweight pollen is easily dispersed [54] and spread over long distances by wind. In addition, since it may only require a single pollen grain to fertilize a female flower, the small pollen size in addition to more pollen produced by Canda and the high viability of Henola pollen would dramatically increase the chances of fertilization and cross-pollination of their pollen with other hemp plants; this is clearly undesirable in the cultivation of hemp for CBD production. In the United States, there is no national mandate for hemp growers to maintain buffer zones to mitigate cross pollination however, it is required in some jurisdictions [55]. In Europe and Canada, a standard buffer zone of 5 km is required [32]. However, according to [32] this was never validated experimentally and could have been the result of collective empirical observations. In addition, research has shown that hemp pollen can travel much further than 5 km. For instance, the occurrence of hemp pollen from North Africa in southwestern Europe a distance of several thousand kilometers [56]. Other crops such as maize with a relatively larger pollen size (between 80 and 125 μm) [57] compared to hemp pollen (22 to 28 μm) were transported over distances as far as 70 km. In Germany, regulations mandate 0.15 km buffer zones adjacent to conventional maize fields and 0.3 km adjacent to organic fields [58]. These recommended buffer zones may be effective because the amount of pollen transported decreases logarithmically with increasing distance from the source. Therefore, the risk of cross-pollination could be insignificant beyond the buffer zone from a pollen source. However, further research is needed to determine buffer zone distances for hemp as this might be different for each hemp variety or botanical classification of the variety.
Henola is a monoecious and dual-purpose variety bred for fiber and seed production compared to CFX-2, Canda, and Joey which are dioecious and grown predominantly for grain [45,59]. Ineffective fertilization and lower seed set and productivity have been linked to a lack of viable pollen [60]. Some studies have documented the seed yield of Henola to average around 3.25 t/ha [61,62] a value higher than the average grain yield of 2.21t/ha reported for CFX-2, Canda, and Joey [63], and also a higher Henola seed yield compared to other varieties [64]. This could be because pollination effectiveness and the resultant seed yield are due to increased pollen viability rather than increased pollen quantity [60,65].
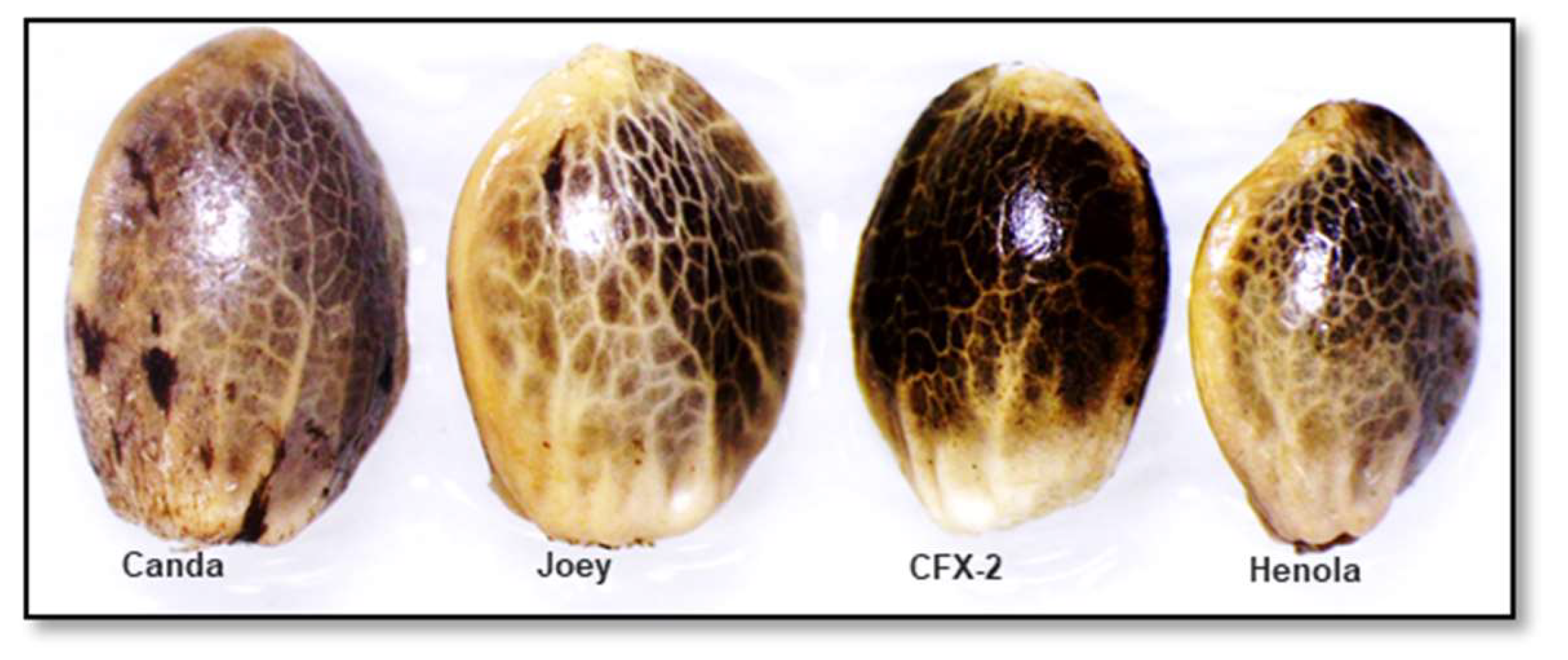
Figure 12 shows the spherical shape of the hemp seed. Among the four varieties, there was a significant difference (F3, 116 = 72.9 p < 0.0001) in seed length (Figure 12) with Canda having the longest length (0.44 ±0.003 cm) followed by Joey (0.41 ± 0.005 cm) and CFX-2 (0.40 ± 0.0028 cm) and Henola recorded the shortest length (0.35 ± 0.005 cm). Similarly, there was a significant difference (F3, 116 = 24.5, p < 0.000) in seed width with the shortest in Henola (0.24 ± 0.004 cm) and then Joey (0.27 ±0.006 cm) and similar widths in CFX-2 (0.28 ± 0.003) and Canda (0.29 ± 0.003). This was within the range reported for other hemp varieties [66]. Despite its small seed size, the lipid content, palmitic acid (C16:0), and the major fatty acid linolenic acid (C18:3 n3) were higher in Henola seed compared to the seeds of two monoecious (Futura 75 and Futura 83) hemp varieties [67]. Overall, seed metrics do not appear to be the primary target in variety selection by growers. Chemical composition and pollination efficiency may be most important traits.
3. Materials and Methods
3.1. Study Site and Plant Varieties
This study was conducted at the North Carolina Agricultural and Technical State University in Greensboro, North Carolina, USA. We used four hemp varieties (Canda, CFX-2, Henola, and Joey), which are among the most popular varieties grown in the United States for grain and fiber. Seeds were purchased from King’s AgiSeeds Inc. (1828 Freedom Road, Lancaster, PA, USA). The seeds of each variety were sown in the green house in 11.35 L plastic plant pot (28 cm wide and 24 cm high) with drainage filled with soil mix (Sunshine® Mix #1, Triangle Chemicals, Kinston, NC, USA). Seedlings were fertilized weekly with fish fertilizer (Alaska fish fertilizer 5-1-1, Lily Miller Brands, Atlanta, GA). After about two weeks, seedlings were thinned to about 4 plants per pot and a total of five pots per variety. Plants were fertilized bi-weekly with nitrogen 34-0-0 and watering was achieved through an automatic watering system regulated to water once daily for 10 minutes.
3.2. Pollen Viability
For each hemp variety, unopened mature flower buds were harvested, and one transferred into an Eppendorf tube, and stored in a refrigerator at 4°C. After 24 hours, Eppendorf tubes with opened anthers were selected and 2 mL of Ampha buffer #6 added to each tube following the manufacturer’s instructions. The suspension was hand-shaken to release the pollen grains into the solution. The pollen suspension was filtered using a 50µm Ampha filter and the pollen grains in the filtrate were counted using the Ampha Z32 Neutec pollen counter (NEUTEC GROUP Inc Farmingdale, NY). The Ampha Z32 uses impedance flow cytometry (IFC) to measure the electrical properties of cells. The IFC system uses microfluidic chip which permits measurements in the radio frequency range from 0.1 to 30 MHz with alternating current (AC). At a chosen frequency, data of cell size, membrane capacitance, cell concentration, and cytoplasmic conductivity of single cells are simultaneously obtained and related to biological key parameters such as cell viability and membrane permeability [68,69]. The Ampha Z32 data acquisition and processing algorithms display results on a scatterplot as counts of non-viable and viable pollen (Figure 10).
3.3. Pollen Staining and Measurement of Pollen Size
Mature unopened flower buds of each hemp variety were harvested and placed into separate Petri dishes. Petri dishes were manually shaken to release pollen grains which were transferred into 1.5 mL centrifuge tubes using a soft camel brush. Tubes were left unsealed to air dry for one hour before sealing and storing in a refrigerator at 4°C. Using a fresh culture swab, about 1000 pollen grains were deposited on a glass microscope slide (75 mm x 25 mm). Following this, 20 μL of modified Alexander stain [10 mL of 95% alcohol, 1 mL of diluted malachite green, 54.5 mL of distilled water, 25 mL of glycerol, 5 mL of diluted acid fuchsin, 0.5 mL of diluted orange G, and 4 mL of glacial acetic acid [24,25] was pipetted directly onto the pollen sample. Each slide was slowly heated on an electric burner to near boiling for about 10 seconds to allow the stain to set. Slides were allowed to cool for 2 minutes, and a coverslip was placed on each slide. Each slide was appropriately labeled, dated, and left at room temperature for 24 hours before being placed in a microscope slide storage folder. Pollen size was measured from the equatorial plane using an Olympus CX43 compound and SZX7 stereo microscopes and recorded. Photos were taken using an Olympus CX43 compound fitted with Olympus EP50 digital camera.
3.4. Flower and Seed Size
One unopened mature flower of each variety was randomly collected and placed in separate Petri dishes. We also randomly selected seeds from each variety and placed in separate Petri dishes. Measurements and photos were taken of length and width of anthers and seeds using an Olympus SZX7 stereo microscopes fitted with Olympus EP50 digital camera.
3.5. Statistical Analysis
Data wre analyzed using JMP Statistical Discovery software (JMP v.13.0.0 SAS Institute). Pollen size, pollen number and viability, length and width of anther and seeds of all four hemp varieties were subjected to one-way analysis of variance (ANOVA) with alpha at 5%. The Tukey-Kramer HSD test was used to separate means with p <0.05. Graphs were plotted using Microsoft Excel. The relationship between anther width and pollen production for all varieties combined and for each treatment was determined suing Pearson’s correlation and regression analysis.
Author Contributions
Conceptualization, B.N.D. and L.E.J.; methodology, B.N.D. and L.E.J.; formal analysis, B.N.D.; investigation, B.N.D. and L.E.J.; resources, B.N.D.; data curation, B.N.D.; writing—original draft preparation, B.N.D.; writing—review and editing, B.N.D. and L.E.J.; visualization, B.N.D. and L.E.J., project administration, B.N.D.; funding acquisition, B.N.D. and L.E.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Southern Sustainable Agriculture Research and Education (SSARE), Project No. LS20-333.
Data Availability Statement
Data sharing is not applicable.
Acknowledgments
We hereby acknowledge the assistance of the staff in the IPM Laboratory at NCA&T for their help in setting up experimental plots and data collection.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Amaducci, S.; Gusovious, H.J. Hemp - Cultivation, Extraction and Processing. In Industrial Applications of Natural Fibres: Structure, Properties and Technical Applications, Müssig, J., Ed.; John Wiley and Sons Ltd.: The Atrium, Southern Gate, Chichester, West Sussex, PO19 8SQ, United Kingdom, 2010, pp. 109- 134.
- Salentijn, E.M..; Zhang, Q.; Amaducci, S.; Yang, M.; Trindade, L.M. New developments in fiber hemp (Cannabis sativa L.) breeding. Ind. Crop. Prod. 2015, 68, 32-41.
- Schluttenhofer, C.; Yuan, L. (2017). Challenges towards revitalizing hemp: A multifaceted crop. Trends Plant Sci. 2017, 22, 917-929.
- Market Data Forecast. Global Industrial Hemp Market Size, Share, Trends, COVID-19 Impact & Growth Analysis Report—Segmented by Type, Application and Region (North America, Europe, Asia-Pacific, Latin America, Middle East, and Africa)—Industry Forecast (2022 to 2027). Available online: https://www.marketdataforecast.com/market-reports/industrial-hemp-market (accessed on 13 August 2024).
- Johnson, R. Defining Hemp: A Fact Sheet; Congressional Research Service: Washington, DC, USA, 2019.
- Malone, T.; Gomez, K. Hemp in the United States: A Case Study of Regulatory Path Dependence. Appl. Econ. Perspect. Policy 2019, 1–16.
- USDA-NASS. Agricultural Statistic 2023; National Hemp Report; USDA-NASS: Washington, DC, USA, 2023.
- Dingha, B.; Sandler, L.; Bhowmik, A.; Akotsen-Mensah, C.; Jackai, L.; Gibson, K.; Turco, R. Industrial Hemp Knowledge and Interest among North Carolina Organic Farmers in the United States. Sustainability 2019, 11, 2691.
- Kaur, G.; Kander, R. The Sustainability of Industrial Hemp: A Literature Review of Its Economic, Environmental, and Social Sustainability. Sustainability 2023, 15, 6457.
- Farinon, B.; Molinari, R.; Costantini, L.; Merendino, N. The Seed of Industrial Hemp (Cannabis sativa L.): Nutritional Quality and Potential Functionality for Human Health and Nutrition. Nutrients 2020, 12, 1935.
- Fiani, B.; Sarhadi, K.J.; Soula, M.; Zafar, A.; Quadri, S.A. Current application of cannabidiol (CBD) in the management and treatment of neurological disorders. Neurol. Sci. Off. J. Ital. Neurol. Soc. Ital. Soc. Clin. Neurophysiol. 2020, 41, 3085–3098.
- Britch, S.C.; Babalonis, S.; Walsh, S.L. Cannabidiol: Pharmacology and therapeutic targets. Psychopharmacology 2021, 238, 9–28.
- Castillo-Arellano, J.; Canseco-Alba, A.; Cutler, S.J.; León, F. The Polypharmacological Effects of Cannabidiol. Molecules 2023, 28, 3271.
- Wang, X.; Zhang, H.; Liu, Y.; Xu, Y.; Yang, B.; Li, H.; Chen, L. An overview on synthetic and biological activities of cannabidiol (CBD) and its derivatives. Bioorg. Chem. 2023, 140, 106810.
- Chundawat, S.P.S.; Beckham, G.T.; Himmel, M.E.; Dale, B.E. Deconstruction of lignocellulosic biomass to fuels and chemicals. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 121-145.
- Hu, R.; Lim, J.K. Fabrication and mechanical properties of completely biodegradable hemp fiber reinforced polylactic acid composites. J. Compos. Mater. 2016, 41, 1655-1669.
- Angelini, L.G.; Tavarini S.; Candilo, M.D. Performance of new and traditional fiber hemp (Cannabis sativa L.) cultivars for novel application: Stem bark, and core yield and chemical composition. J. Nat. Fibers 2016,13, 238-252.
- Cherney, J.H.; Small, E. Industrial Hemp in North America: Production, Politics and Potential. Agronomy 2016, 6, 58.
- Zhao, J.; Xu, Y.; Wang, W.; Griffin, J.; Roozeboom, K.; Wang, D. Bioconversion of industrial hemp biomass for bioethanol production: A review. Fuel 2020, 281, 118725.
- Placido, D.F.; Lee, C.C. Potential of Industrial Hemp for Phytoremediation of Heavy Metals. Plants 2022, 11, 595.
- Pollastro, F.; Minassi, A.; Fresu, L.G. (2018). Cannabis phenolics and their bioactivities. Curr. Med. Chem. 2018, 25, 1160-1185.
- Leonard, W.; Zhang, P.; Ying, D.; Fang, Z. Hempseed in food industry: Nutritional value, health benefits, and industrial applications. Compr. Rev. Food Sci. Food Saf. 2020, 19, 282-308.
- Chen, T.; He, J.; Zhang, J.; Zhang, H.; Qian, P.; Hao, J.; Li, L. Analytical Characterization of Hempseed (Seed of Cannabis sativa L.) Oil from Eight Regions in China. J. Diet. Suppl. 2010, 7, 117–129.
- House, J.D.; Neufeld, J.; Leson, G. Evaluating the Quality of Protein from Hemp Seed (Cannabis sativa L.) Products Through the Use of the Protein Digestibility-Corrected Amino Acid Score Method. J. Agric. Food Chem. 2010, 58, 11801–11807.
- Vonapartis, E.; Aubin, M.P.; Seguin, P.; Mustafa, A.F.; Charron, J.B. Seed composition of ten industrial hemp cultivars approved for production in Canada. J. Food Compos. Anal. 2015, 39, 8–12.
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance-A review. Life Sci. 2018, 203, 255–267.
- Alonso-Esteban, J.I.; González-Fernández, M.J.; Fabrikov, D.; Sánchez-Mata, M.; Torija-Isasa, E.; Guil-Guerrero, J.L. Fatty acids and minor functional compounds of hemp (Cannabis sativa L.) seeds and other Cannabaceae species. J. Food Compos. Anal. 2023, 115, 104962.
- Orlando, G.; Recinella, L.; Chiavaroli, A.; Brunetti, L.; Leone, S.; Carradori, S.; Di Simone, S.; Ciferri, M.C.; Zengin, G.; Ak, G.; et al. Water Extract from Inflorescences of Industrial Hemp Futura 75 Variety as a Source of Anti-Inflammatory, Anti-Proliferative and Antimycotic Agents: Results from In Silico, In Vitro and Ex Vivo Studies. Antioxidants 2020, 9, 437.
- di Giacomo, V.; Recinella, L.; Chiavaroli, A.; Orlando, G.; Cataldi, A.; Rapino, M.; Di Valerio, V.; Politi, M.; Antolini, M.D.; Acquaviva, A.; et al. Metabolomic Profile and Antioxidant/Anti-Inflammatory Effects of Industrial Hemp Water Extract in Fibroblasts, Keratinocytes and Isolated Mouse Skin Specimens. Antioxidants 2021, 10, 44.
- Alonso-Esteban, J.I.; Pinela, J.; Ćirić, A.; Calhelha, R.C.; Soković, M.; Ferreira, I.C.F.R.; Barros, L.; Torija-Isasa, E.; Sánchez-Mata, M.d.C. Chemical composition and biological activities of whole and dehulled hemp (Cannabis sativa L.) seeds. Food Chem. 2022, 374, 131754.
- Serventi, L.; Flores, G.A.; Cusumano, G.; Barbaro, D.; Tirillini, B.; Venanzoni, R.; Angelini, P.; Acquaviva, A.; Di Simone, S.C.; Orlando, G.; et al. Comparative Investigation of Antimicrobial and Antioxidant Effects of the Extracts from the Inflorescences and Leaves of the Cannabis sativa L. cv. strawberry. Antioxidants 2023, 12, 219.
- Small, E.; Antle, T. A Preliminary Study of Pollen Dispersal in Cannabis sativa in Relation to Wind Direction. J. Ind. Hemp 2003, 8, 37-50.
- Kurtz, L.E.; Brand, M.H.; Lubell-Brand, J.D. Production of tetraploid and triploid hemp. HortSci. 2020, 55, 1703-1707.
- Gómez-Mena, C.; Honys, D.; Datla, R.; Testillano, P.S. Advances in pollen research: biology, biotechnology, and plant breeding applications. Front. Plant Sci. 2022, 13, 876502.
- Todd, J.; Song, H.; Van Acker, R. Does pollination alter the cannabinoid composition and yield of extracts from hemp (Cannabis sativa L. cv. Finola) flowers?. Ind. Crop. and Prod. 2022, 183, 114989.
- Ushiyama, T.; Du, M.; Inoue, S.; Shibaike, H.; Yonemura, S.; Kawashima, S.; Amano, K. Three-dimensional prediction of maize pollen dispersal and cross-pollination, and the effects of windbreaks. Environ. Biosafety Res. 2009, 8, 183-202.
- Meier, C.; Mediavilla ,V. Factors influencing the yield and the quality of hemp essential oil. J. Int. Hemp Assoc. 1998, 5, 16–20.
- Capital Press. Hemp boom spurs cross-pollination disputes. https://www.capitalpress.com/state/oregon/hemp-boom-spurs-cross-pollination-disputes/article_efd1e99c-c903-11e9-8bdd-73e58f5946b5.html Available online (accessed on 06 July 2024).
- National Cannabis Industry Association (NCIA). Cross-pollination poised to prompt litigation in light of new USDA hemp rules. Available online https://thecannabisindustry.org/member-blog-cross-pollination-poised-to-prompt-litigation-in-light-of-new-usda-hemp-rules/ (accessed on 06 July 2024).
- Chabbert, B.; Kurek, B.; Beherec, O. Physiology and botany of industrial hemp. In Hemp Industrial Production and Uses; Bouloc, P., Ed.; Cabi Publication: Wallingford, UK, 2013; pp. 27–47.
- Amaducci, S.; Scordia, D.; Liu, F.H.; Zhang, Q.; Guo, H.; Testa, G.; Cosentino, S.L. Key cultivation techniques for hemp in Europe and China. Ind. Crop. Prod. 2015, 68, 2-16.
- Leme, F.M.; Schönenberger, J.; Staedler, Y.M.; Teixeira, S.P. Comparative floral development reveals novel aspects of structure and diversity of flowers in Cannabaceae. Bot. J. Linn. Soc. 2020,193, 64–83.
- Hesami, M.; Pepe, M.; Jones, A.M.P. Morphological Characterization of Cannabis sativa L. Throughout Its Complete Life Cycle. Plants 2023, 12, 3646.
- Renner, S.S.; Ricklefs, R.E. “Dioecy and its correlates in the flowering plants.” Am. J. Bot. 1995, 82, 596-606.
- Frankowski, J.; Wawro, A.; Batog, J.; Burczyk, H. New Polish Oilseed Hemp Cultivar Henola - Cultivation, Properties and Utilization for Bioethanol Production. J. Nat. Fibers. 2021, 19, 7283–7295.
- Fernández-Illescas, F.; Nieva, J.; Márquez-García, B.; Muñoz-Rodríguez, A. Pollen production in halophytic species of the Chenopodiaceae in a Mediterranean marsh. Grana. 2010, 49, 300–307.
- Faegri, K.; Iverson, J.; Kaland, P.E.; Krzywinski, K. Textbook of pollen analysis, 4th ed.; John Wiley and Sons: Chichester, UK, 1989; 328.
- Bhowmik, S.; Datta, B.K. Pollen production in relation to ecological class of some hydrophytes and marsh plants. Am. J. Plant Sci. 2013, 4, 324-332.
- Milatović, D.; Nikolić, D.; Janković, S.; Janković, D.; Stanković, J. Morphological characteristics of male reproductive organs in some walnut (Juglans regia L.) genotypes. Sci. Hortic. 2020, 272, 109587.
- Pinheiro-Costa, B.K.; Mesquita-Neto, J.N.; Rego, J.O.; Schlindwein, C. Trade off between quantity and size of pollen grains in the heterandrous flowers of Senna pendula (Fabaceae). Acta Bot. Bras. 2018, 32, 446-453.
- Trevizan, R.; Caetano,A.P.S.; Brito, V.L.G.; Oliveira, P.E.; Telles, F.J. Stamen and pollen heteromorphism linked to the division of labour in Melastomataceae species, Flora. 2023, 305, 152315.
- Hao, K.; Tian, Z.X.; Wang, Z.C.; Huang, S.Q. Pollen grain size associated with pollinator feeding strategy. Proc. Biol. Sci. 2020, 287, 20201191.
- Dingha, B.N.; Jackai, L.E. Chemical Composition of Four Industrial Hemp (Cannabis sativa L.) Pollen and Bee Preference. Insects 2023, 14, 668.
- Ackerman, J.D. Abiotic pollen and pollination: ecological, functional, and evolutionary perspectives. Plant Syst. Evol. 2000, 222, 167–185.
- Floraflex. Hemp Farming Regulations and Licensing: Navigating Legal Requirements. Available online: https://floraflex.com/default/blog/post/hemp-farming-regulations-and-licensing-navigating-legal-requirements (accessed August 16 2024).
- Cabezudo, B.; Recio, M.; Sánchez-Laulhé, J.M.; Trigo, M.D.M.; Toro, F.J.; Polvorinos, F. Atmospheric transportation of marihuana pollen from North Africa to the southwest of Europe. Atmos. Environ. 1997, 31, 3323-3328.
- Brunet, Y.; Foueillassar, X.; Audran, A.; Garrigou, D.; Dayau, S.; Tardieu, L. Evidence for long-range transport of viable maize pollen. In 1st European Conference on the Coexistence of Genetically Modified Crops with Conventional and Organic Crops, Boelt, B., Slagelse, Eds.; Danish Institute of Agricultural Sciences: Helsingor, Denmark, 2003; pp. 74–76.
- Hofmann, F.; Epp, R.; Kruse, L. et al. Monitoring of Bt-Maize pollen exposure in the vicinity of the nature reserve Ruhlsdorfer Bruch in northeast Germany 2007 to 2008. Environ. Sci. Eur. 2010, 22, 229–251.
- Sieracka, D.; Zaborowicz, M.; Frankowski, J. Identification of Characteristic Parameters in Seed Yielding of Selected Varieties of Industrial Hemp (Cannabis sativa L.) Using Artificial Intelligence Methods. Agriculture 2023, 13, 1097.
- Impe, D.; Reitz, J.; Köpnick, C.; Rolletschek, H.; Börner, A.; Senula, A.; Nagel, M. Assessment of pollen viability for wheat. Front. Plant Sci. 2020, 10, 1588.
- Teleszko, M.; Zając, A.; Rusak, T. Hemp Seeds of the Polish ‘Bialobrzeskie’ and ‘Henola’ Varieties (Cannabis sativa L. var. sativa) as Prospective Plant Sources for Food Production. Molecules 2022, 27, 1448.
- Stramkale, V.; Morozova, I.; Černova, L.; Stramkalis, A. Industrial Hemp Varieties Productivity Potential in the Latvian Climatic Conditions. In Proceedings of the 14th International Scientific and Practical Conference. 15-16 June 2023.
- Lan, Y.; Zha, F.; Peckrul, A.; Hanson, B.; Johnson, B.; Rao, J.; Chen, B. Genotype x Environmental Effects on Yielding Ability and Seed Chemical Composition of Industrial Hemp (Cannabis sativa L.) Varieties Grown in North Dakota, USA. J. Am. Oil. Chem. Soc. 2019, 96, 1417-1425.
- Bajwa, P.; Singh, S.; Singh, M.; Kafle, A.; Parkash, V.; et al. Assessing the production potential of industrial hemp in the semi-arid west Texas. Technol. Agron. 2023, 3,17.
- Matthews, F.R.; Bramlett, D.L. Pollen Quantity and Viability Affect Seed Yields from Controlled Pollinations of Loblolly Pine. South. J. Appl. For. 1986, 10, 78-80.
- Moon, Y.; Cha, Y.; Lee, J.; Kim, K.; Kwon, D.; Kang, Y. Investigation of Suitable Seed Sizes, Segregation of Ripe Seeds, and Improved Germination Rate for the Commercial Production of Hemp Sprouts (Cannabis sativa L.). J. Sci. Food Agric. 2020, 100, 2819-2827.
- Gimeno-Martínez, D.; Igual, M.; García-Segovia, P.; Martínez-Monzó, J.; Navarro-Rocha, J. Characterisation of the Fat Profile of Different Varieties of Hemp Seeds (Cannabis sativa L.) for Food Use. Biol. Life Sci. Forum 2023, 26, 89.
- Cheung, K.C.; Di Berardino, M.; Schade-Kampmann, G.; Hebeisen, M.; Pierzchalski, A.; Bocsi, J.; Mittag, A.; Tárnok, A. Microfluidic impedance-based flow cytometry. Cytometry A 2010, 77, 648-66.
- Xu Y, Xie X, Duan Y, Wang L, Cheng Z, Cheng J. A review of impedance measurements of whole cells. Biosens. Bioelectron. 2016, 77, 824-36.
Figure 1.
(A) Dioecious and (B) monoecious hemp plants (a) unopened flowers and (b) opened sepal enclosing tightly packed stamens on a panicle of a male plant.
Figure 1.
(A) Dioecious and (B) monoecious hemp plants (a) unopened flowers and (b) opened sepal enclosing tightly packed stamens on a panicle of a male plant.

Figure 2.
(A) Dioecious female and (B) monoecious hemp plants both show immature seeds.

Figure 3.
The inflorescence of the male hemp plant shows (a) an immature flower bud, (b) a partially opened flower (c) a flower with exposed anthers and sepals (d) a flower with visible filaments and dehisced anthers.
Figure 3.
The inflorescence of the male hemp plant shows (a) an immature flower bud, (b) a partially opened flower (c) a flower with exposed anthers and sepals (d) a flower with visible filaments and dehisced anthers.

Figure 4.
Single isolated anthers of the four hemp varieties showing differences in size.

Figure 5.
Stained pollen grains (a) Joey (b) Henola (c) CFX-2 and (d) Canda observed under light microscopy at magnification 40x.
Figure 5.
Stained pollen grains (a) Joey (b) Henola (c) CFX-2 and (d) Canda observed under light microscopy at magnification 40x.
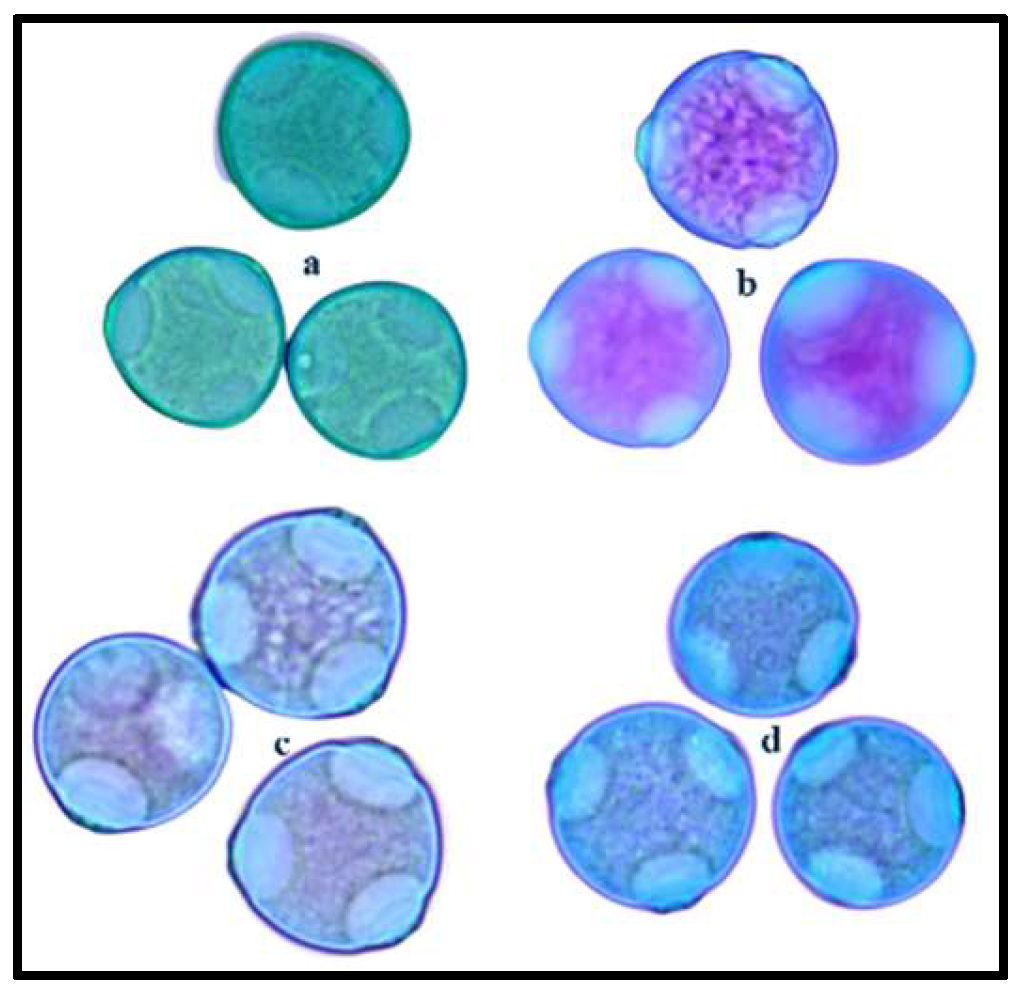
Figure 6.
Number of pollen grains of the four hemp varieties. Means with the same letters are not significantly different (p > 0.05).
Figure 6.
Number of pollen grains of the four hemp varieties. Means with the same letters are not significantly different (p > 0.05).
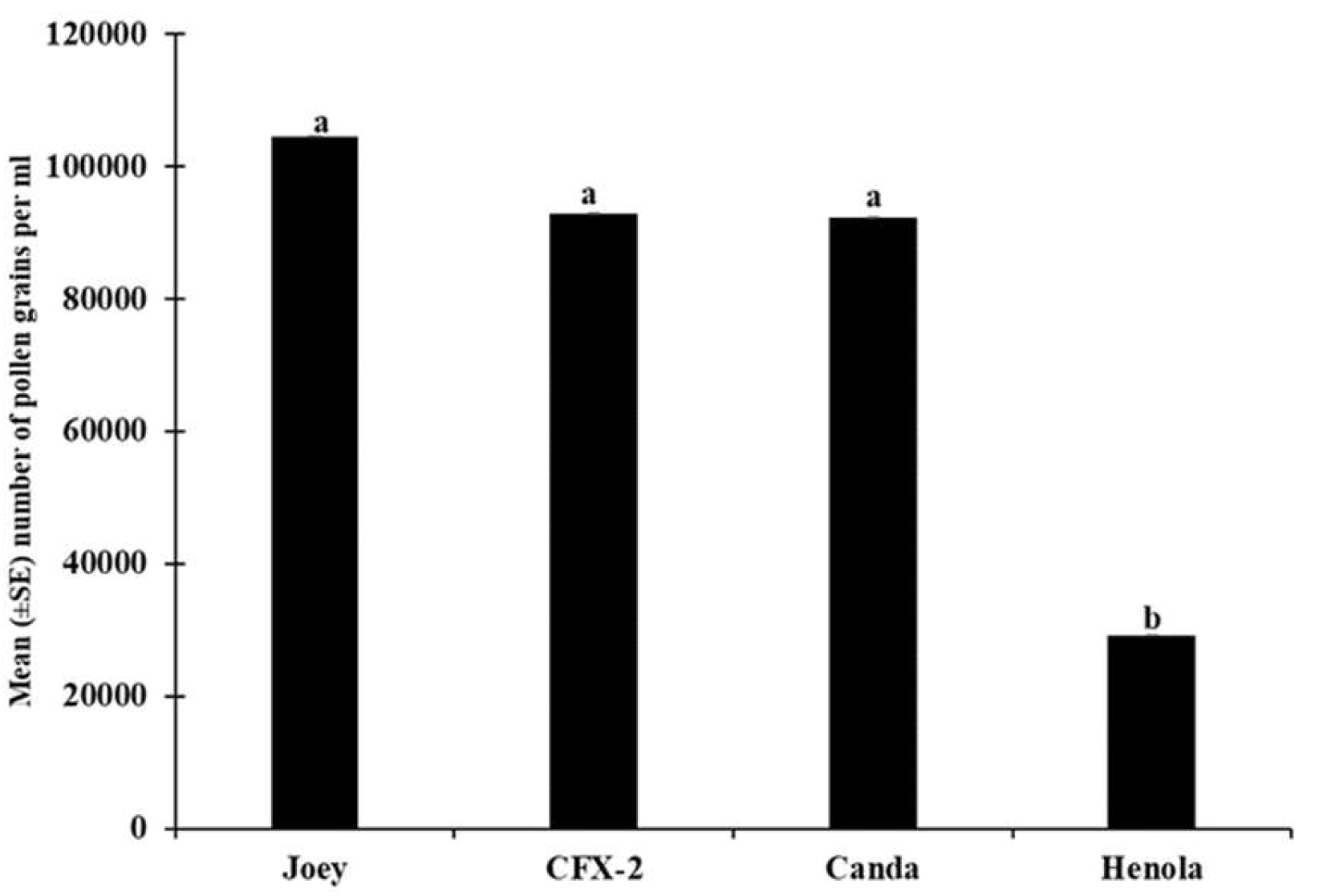
Figure 7.
Linear regression from all four hemp varieties showing the relationship between anther length and pollen production (a) anther length (y = 24652.906 + 146776.29*x; n = 74; r2 = 0.039; p = 0.0977), (b) anther width (y = 37238.184 + 371801.39x; n = 74; r2 = 0.025; p = 0.175).
Figure 7.
Linear regression from all four hemp varieties showing the relationship between anther length and pollen production (a) anther length (y = 24652.906 + 146776.29*x; n = 74; r2 = 0.039; p = 0.0977), (b) anther width (y = 37238.184 + 371801.39x; n = 74; r2 = 0.025; p = 0.175).
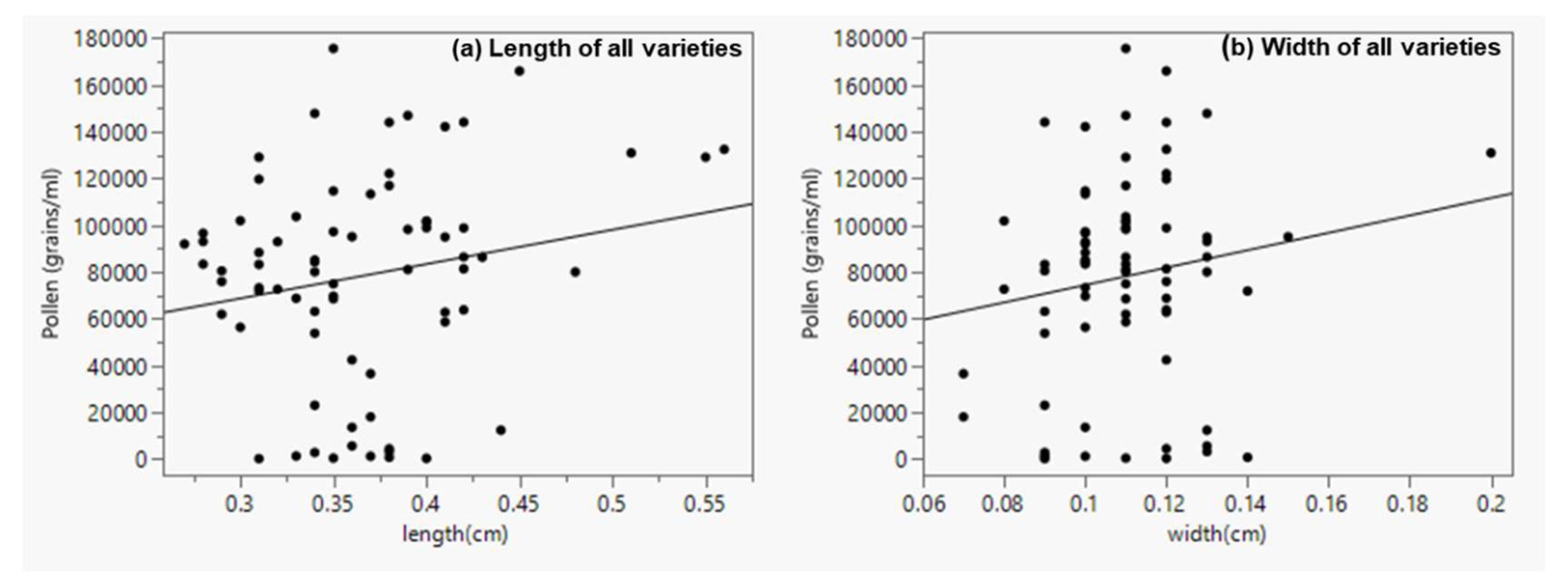
Figure 8.
Linear regression showing the relationship between anther length and pollen production (a) Canda (y = 42228.932 + 142950.78x; n = 17; r2 = 0.035; p = 0.47), (b) CFX-2 (y = 88729.339 + 12468.785x; n = 20; r2 = 0.0014; p = 0.877), (c) Henola (y = -124002.9 + 411238.3x; n = 20; r2 = 0.127; p = 0.124), (d) Joey (y = -6658.993 + 268920.47x; n = 17; r2 = 0.247; p = 0.043).
Figure 8.
Linear regression showing the relationship between anther length and pollen production (a) Canda (y = 42228.932 + 142950.78x; n = 17; r2 = 0.035; p = 0.47), (b) CFX-2 (y = 88729.339 + 12468.785x; n = 20; r2 = 0.0014; p = 0.877), (c) Henola (y = -124002.9 + 411238.3x; n = 20; r2 = 0.127; p = 0.124), (d) Joey (y = -6658.993 + 268920.47x; n = 17; r2 = 0.247; p = 0.043).
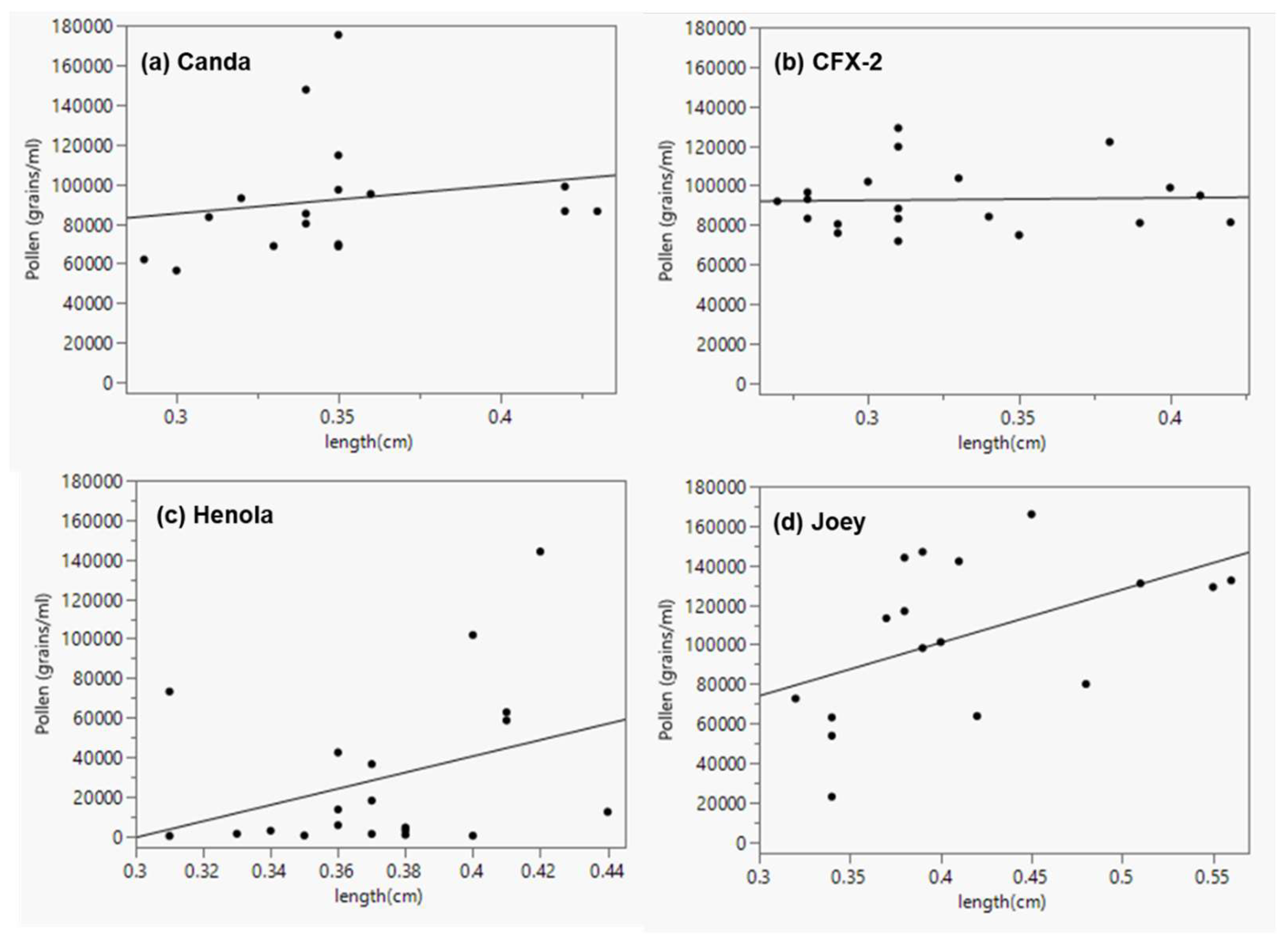
Figure 9.
Linear regression showing the relationship between anther width and pollen production (a) Canda (y = 44789.822 + 422524.62x; n = 17; r2 = 0.038; p = 0.46), (b) CFX-2 (y = 81976.305 + 97682.84x; n = 20; r2 = 0.007; p = 0.73), (c) Henola (y = 93212.894 - 606914.64x; n = 20; r2 = 0.097; p = 0.181), (d) Joey (y = 36483.659 + 605804.08x; n = 17; r2 = 0.162; p = 0.109).
Figure 9.
Linear regression showing the relationship between anther width and pollen production (a) Canda (y = 44789.822 + 422524.62x; n = 17; r2 = 0.038; p = 0.46), (b) CFX-2 (y = 81976.305 + 97682.84x; n = 20; r2 = 0.007; p = 0.73), (c) Henola (y = 93212.894 - 606914.64x; n = 20; r2 = 0.097; p = 0.181), (d) Joey (y = 36483.659 + 605804.08x; n = 17; r2 = 0.162; p = 0.109).
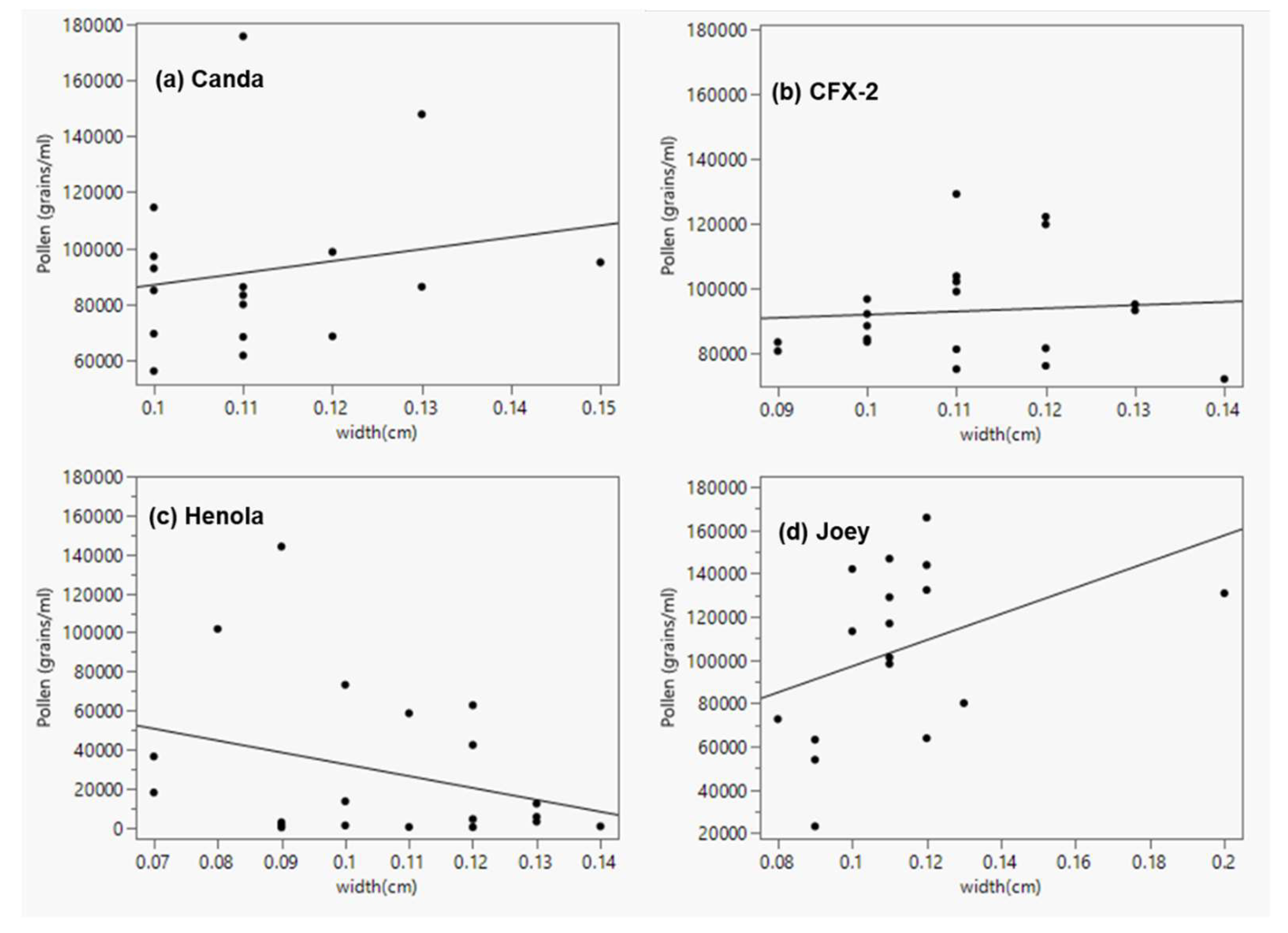
Figure 10.
Ampha cytometer chart at 2.00 MHz for four hemp varieties.
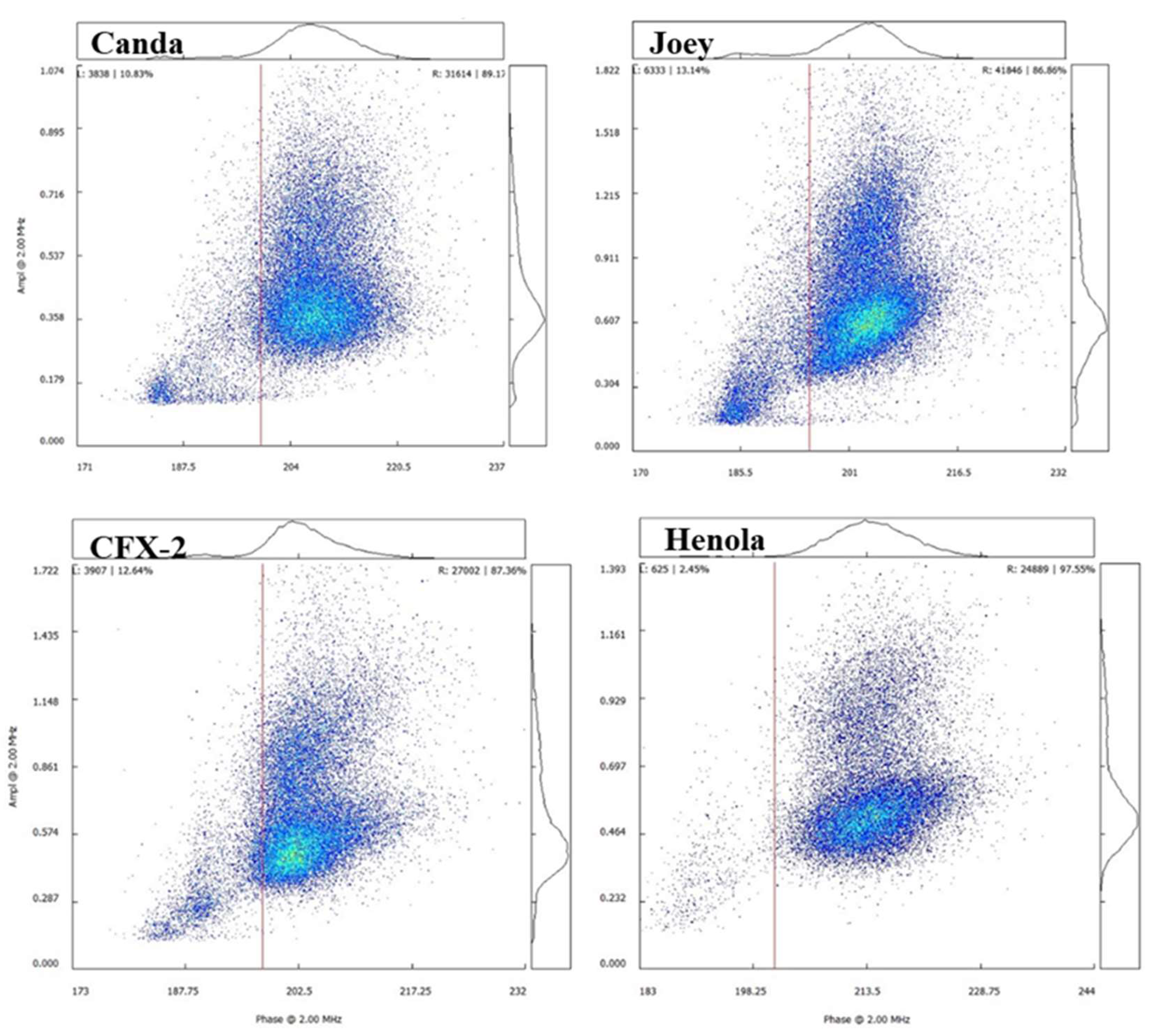
Figure 11.
Percentage of viable pollen of four hemp varieties after storage for 24 hours. Means with the same letters are not significantly different (p > 0.05).
Figure 11.
Percentage of viable pollen of four hemp varieties after storage for 24 hours. Means with the same letters are not significantly different (p > 0.05).
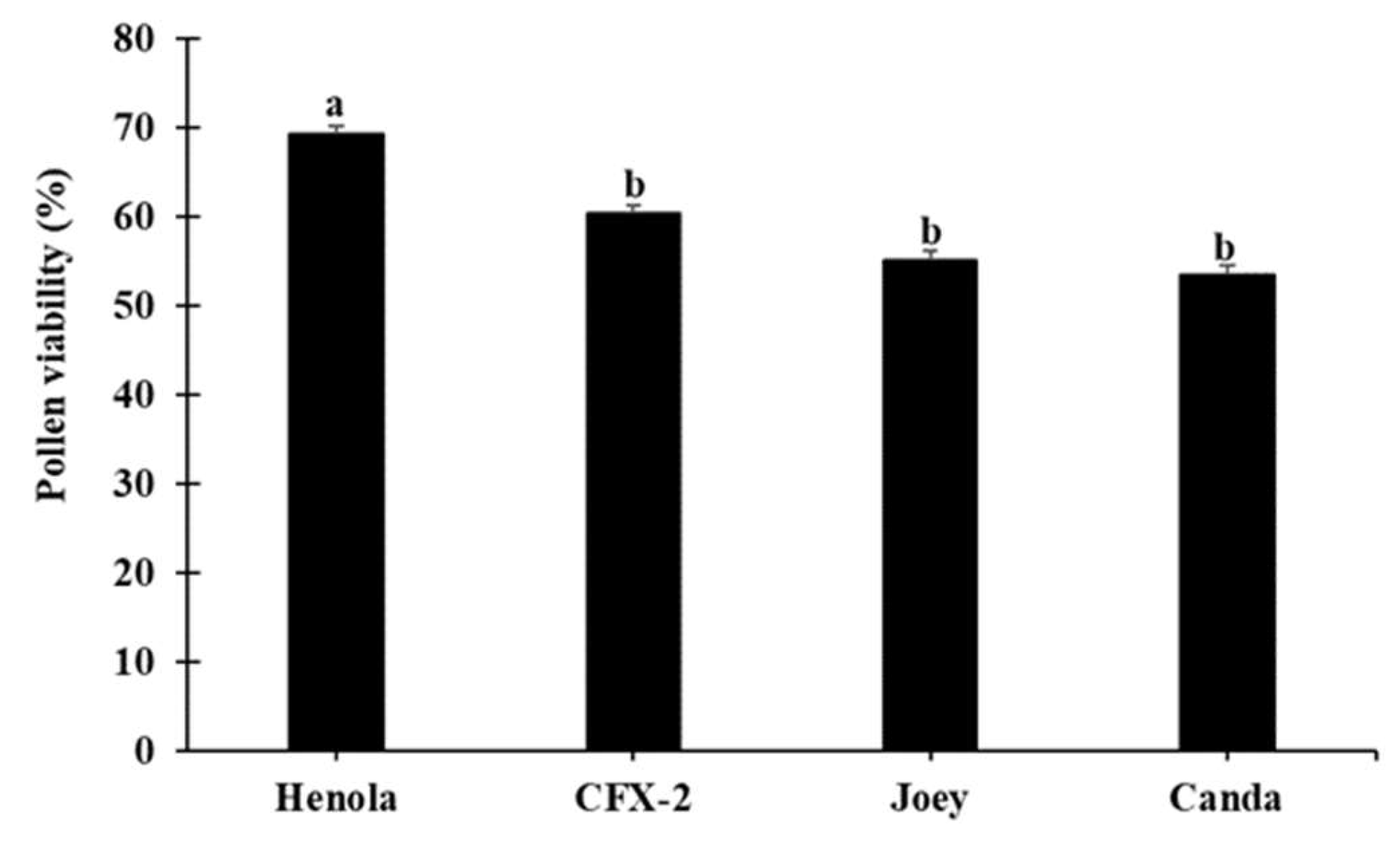
Figure 12.
Spherical shape and variation of seed size among four hemp varieties.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



