
Submitted:
09 October 2024
Posted:
10 October 2024
You are already at the latest version
Abstract
Alzheimer’s disease (AD) is the most widespread neurodegenerative disorder. Recently, it was found that mucus extract from Cornu aspersum have beneficial effects on memory and cognitive processes in a rat scopolamine model of AD. The present study elucidated mechanisms of action of standardized mucus snail extract (SE) enriched with fraction above 20 kDa on Alzheimer-type dementia in rats. Using proteomic analysis on two-dimensional polyacrylamide gel electrophoresis (2D–PAGE) on rat cortex extracts we compared protein expression in both groups: the first group treated intraperitoneally with scopolamine (Sco, 2 mg/kg, 11 days) and the second (Sco + SE) group treated intraperitoneally with Sco (Sco, 2 mg/kg) and protected by SE (0.5 ml/100 g bw) applied daily orally for 11 days. Brain cortex was separated and the expression of various proteins related to memory and cognitive functions were identified. We found that the expression of Ubiquitin carboxyl-terminal hydrolase isozyme L1, Calbindin, Vacuolar ATP synthase catalytic subunit A, Tropomyosin beta chain, 14-3-3 zeta/delta, Kinesin-1 heavy chain and Stathmin-4 significantly differs in SE-protected rats as compared to non-treated dement animals, and these brain proteins might be potential therapeutic targets for Alzheimer’s type dementia treatment.
Keywords:
Alzheimer’s disease
; scopolamine
; snail extract
; neuroprotection
; brain proteins
; dementia
1. Introduction
Alzheimer’s disease (AD), a progressive neurodegenerative disorder, is the most common cause of dementia — a gradual deterioration of memory, thinking, behaviour and social skills. These changes affect a person’s ability to function and in an advanced stage lead to death [1]. The causes of AD, as well as the exact mechanisms leading to neuronal death, have not yet been elucidated. It is generally accepted that selective oxidative stress in combination with a neuroinflammatory process leads to neurodegenerative changes in AD. Currently the most important alterations identified can be explained through: the cholinergic dysfunction, the amyloid cascade, the role of tau protein, APOE, calcium, and the involvement of the mitochondrial cascade and oxidative stress [2,3]. Other factors that affect the clinical development of AD include vascular pathology of blood–brain barrier that result in leakage and cause dementia [4]. Various epigenetic changes, including DNA methylation and hydroxymethylation, mtDNA methylation, noncoding RNA translation, and histone post-translational modifications also have been implicated in AD development [5]. AD is expressed by loss of neurons and synapses in the cerebral cortex and some subcortical areas. This loss leads to large-scale atrophy of the affected regions, including degeneration in the temporal and parietal lobes, as well as parts of the frontal cortex [6]. It is known also that Alzheimer’s disease (AD) is a polygenic and multifactorial disease characterized by the deposition of amyloid – β (Aβ) fibrils in the brain, leading to formation of plaques (Aβ plaques) and neurofibrillary tangles (NFTs), which are the two main pathological hallmarks of AD, and ultimately resulting in dendritic dysfunction, neuronal cell death, memory loss, behavioural changes, and organ shutdown [3].
But the changes in expression of some important brain proteins also can be a new important mechanism in AD pathology, for example as Ubiquitin carboxyl-terminal hydrolase isozyme L1, Calbindin, Vacuolar ATP synthase catalytic subunit A, Tropomyosin beta chain, 14-3-3 zeta/delta, Kinesin-1 heavy chain and Stathmin-4.
Animal models offer valuable tools for evaluating new therapeutic strategies for treatment of human diseases, as well as for studying the pathological mechanisms involved in these processes. In neuroscience research rat models are preferred over mouse models because of the more complex behavioural responses in rats and much more flexibility in dealing with novel situations [7,8].
In AD research, the rats have been used for decades as important model, for instance in studies on cholinergic dysfunction and memory impairment which played a crucial role in the development of the cholinesterase inhibitor drugs that are currently in use. In recent years, a number of transgenic rats as models for AD have been reported and new models are under development [8]. However transgenic model has its limitations because it does not show the complete model of AD, especially in sporadic form of AD, which is prevalent. The researchers developed alternative approaches to induce AD type of dementia using various chemical drugs: colchicine, scopolamine, okadaic acid, streptozotocin, trimethyltin, aluminium trichloride etc. but these chemicals cause damage to certain regions of the brain – hippocampus, cortex [9].
Scopolamine is used to induce cognitive dysfunction in experimental animals because it readily permeates the blood-brain barrier [10]. Sco is a muscarinic cholinergic receptor antagonist leading to memory impairment in rodents [11]. Intraperitoneal injection of Sco caused cholinergic dysfunction and impaired cognition in rats [12] and also causes an increasing amyloid-β deposition, which along with cholinergic dysfunction are hallmarks of the AD molecular pathology [10]. Prolonged use of Sco is known to cause damage to the hippocampal brain cholinergic nerves and to reduce memory of rodents in dose dependent manner. Sco caused decreased activity of choline acetyltransferase (the enzyme responsible for acetylcholine synthesis) in the cortex of AD patients [13]. Recent studies have pointed out that Sco increases the accumulation of reactive oxygen species inducing oxidative stress leading to memory impairment [14].
At present, the search for new natural compounds for multi-target treatment of AD is particularly relevant. In our previous research, we identified and characterized more than 40 novel peptides from C. aspersum mucus with potential antibacterial activity, which could be an alternative to conventional antibiotics due to low levels of resistance [15,16,17]. We identified a variety of metabolites by Nuclear Magnetic Resonance (NMR) and Mass Spectrometry (MS) profiling of C. aspersum mucus in two low molecular weight fractions (<1 kDa and <3 kDa) including free primary amino acids; sugars, such as glucose and sucrose; intermediates, such as fumarate; osmolites, such as betaine and choline; and several other organic acids, including isovaleric acid, lactic, tartaric, and acetic acid; as well as alantoin, glutathione, and antimicrobial peptides [18]. Detected choline is a basic metabolite and plays an important role as a methyl group donor in various vital metabolic processes, and also as a precursor for the synthesis of the neurotransmitter acetylcholine; participates in lipid transport and metabolism as well. Moreover, some Krebs cycle intermediates, such as pyruvate, oxaloacetate, and α-ketoglutarate, detected in the snail mucus could act as energy substrates in mitochondria and they manifest antioxidant properties in the brain [19]. In the same study, it was found that these Krebs cycle intermediates are effective protective compounds against oxidative stress in neuronal cells as they preserve HT22 cells from hydrogen peroxide-mediated toxicity. Since these intermediates did not have any toxic effects, they can be used in the therapeutic treatment of chronic neurodegenerative diseases.
The present study was designed to build on our previous research [20] related to the beneficial effects of mucus extract from C. aspersum snail in an experimental rat model of Alzheimer’s type dementia. Our original results provided the first insight into the capacity of SE from C. aspersum to improve memory in experimental AD type dementia. However, the complex mechanisms of this disease deserve more profound studies.
Hence, the goal of present study was to shed additional light on the capacity of SE to change the expression of some important brain proteins summarized in Table 1, as potential therapeutic targets for AD treatment, using advance 2D-PAGE proteomic analysis, MALDI-MS and MALDI -MS/MS and bioinformatics.
2. Results
2.1. Preparation of Mucus Extract
The crude mucus was collected from snails grown in Bulgarian eco farms using patented technology. After filtration and purification, a concentrated protein fraction with components above 20 kDa was added to purified initial mucus with a ratio between the two fractions of 1:1. This mixture is designated as snail extract (SE) and was used for treatment of the laboratory animals.
2.2. Treatment of Laboratory Animals by Mucus Extract and Scopolamine
To induce Alzheimer’s type dementia-type AD, laboratory rats were injected intraperitoneally with Scopolamine hydrobromide, designated as Sco. Male adult Wistar rats were used as laboratory animals. They were divided into two groups - the first (Sco group) was treated for 11 days intraperitoneally (Sco, 2 mg/kg) and the second (Sco + SE) treated intraperitoneally with Sco (Sco, 2 mg/kg ) and SE (0.5 ml/100 g bw) applied orally.
At the end of the experiment, the laboratory animals were euthanized and decapitated, and the prefrontal cortex was isolated from their brains. After homogenization of the cortex samples and their centrifugation (at 10 000 rpm for 30 min at 4 0C), the protein concentrations of the supernatants were evaluated by the Bradford method and the probes were prepared for 2 D PAGE.
2.3. Identification of Proteins on 2D -Gel Electrophoresis, Using Mass Spectrometry and Bioinformatics
Using two-dimensional gel electrophoresis (2D-PAGE) coupled with software MelanieTM Coverage 9.2, MALDI-MS and MALDI-MS/MS, we have analysed the cortex proteome and identified various proteins which change their expression in cortex after treatment with Sco (Figure 1A) and after treatment with Sco + SE (Figure 1B).
The proteins in rat cerebral cortex were separated by 2D electrophoresis and stained by Coomassie brilliant blue, were detected using MelanieTM Coverage 9.2 software in Coomassie stained three polyacrylamide gels (Standart, Sco control and Sco and SE together) (Figure 1).
After scanning of the gels with high resolution the differences in protein expression between Sco and Sco + SE group, as well as molecular weight (MW) and isoelectric point (pI) of corresponding proteins were determined by computer-aided 2D image analysis, carried out using MelanieTM Coverage 9.2. The software calculated the intensities in pixel of all spots. Prediction of the proteins from the 2D-PAGE spots of cortex was performed using the obtained data on their molecular weight (MW) and isoelectric point (pI) and comparison with the database UniProt, taxonomy Rattus norvegicus, brain (https://www.uniprot.org/).
After the conducted 2D electrophoresis, mass spectrometric analysis and online database searches we found proteins that perform different functions - structural, antioxidant, metabolic, signaling, involved in the folding of other proteins, etc. The key proteins with their change in expression in Sco and Sco + SE rats are presented in the Table 2. Selected spots from the polyacrylamide gels were excised and subjected to trypsin digestion and prepared for subsequent mass spectrometric analysis. After extraction of peptides from each protein spot they were subjected to MALDI-MS and some of them to MS/MS spectrometric analysis (Figure 2 and Figure 3). Protein identification was performed by MASCOT software (http://www.matrixscience.com) on the bases MALDI-MS of peptides exracted from gel after triptic digestion of protein spots. The monoisotopic m/z values of the peptide mass fingerprint were compared to known proteins in the Swiss-Prot database, as used the following search parameters: Peptide Mass Fingerprint; Database: SwissProt; Taxonomy: Rattus; Enzyme: Trypsin; Allow up to: 1 - 3 missed cleavages; Monoisotopic; Mass Values MH+; Peptide tolerance: ±1.2 - 2.5 Da and a probabilistic score at p<0.05. For example presented MS spectrum on figure shows 42% protein sequence coverage and e =2.1e-12. The theoretical and observed pI and MW values were in good agreement. Proteins detected in Mascot were checked against the UniProtKB online database (http://www.uniprot.org) in order to verify their localization and characterize their functions.
Some proteins presented in Table 3 were confirmed by MS/MS analysis of tryptic peptides obtained from the respective spots. The MS spectra of the obtained peptides, after tryptic digestion of protein from spot A14 of 2D-PAGE, are presented on Figure 2A.
Analysis of the MS/MS spectrum of the peptide [M+H]+ at m/z 1548.37, obtained after hydrolysis of the protein of spot A14, represents the sequence SVTEQGAELSNEER (Figure 2B), which corresponds to 14-3-3 protein zeta/delta (1433Z_RAT).
Another protein that significantly changes its expression is Stathmin-4 (STMN4_RAT) determined with MW 22 073 Da at pI 5.7 was presented at site A16 by mass spectrometric analysis (Figure 3A,B). It was confirmed by MS following Mascot Peptide Fingerprint and MS/MS analysis.
Peptide sequences generated from the MS/MS spectra were put in BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) to confirm the corresponding proteins. Table 2 presents amino acid sequences of tryptic peptides of several spots from 2D-PAGE images, determined by MS/MS-analyses. Using the same method eight proteins showed in Table 2 were successfully confirmed.
3. Discussion
Because of the multifactorial nature of AD, it is difficult to reveal exact mechanisms underlying this disorder. It is likely that oxidative stress triggers the disease by increasing the production and aggregation of Aβ amyloid peptides facilitating τ-hyperphosphorylation and its aggregation into NFTs [39]. Abnormal accumulation of β-amyloid promotes ROS formation through activation of NMDA receptors is also of importance [40]. Studies have shown that amyloid β interacts with cholinergic receptors, affecting their function [41].
Recently, our research showed that mucus extract from C. aspersum demonstrated complex therapeutic potential in conditions of scopolamine altered behaviour, biochemistry, and gene expression in rat’s brain in Alzheimer’s type dementia. The obtained results showed that the used snail extract possessed moderate antioxidant properties, acetylcholinesterase inhibitory activity and modulates monoamine content in memory-related brain structures. The confirmation of these results by principal components analysis demonstrated a very close position between the Sco + snail and control groups, supporting the restorative effect of SE in demented animals. Moreover, we reported also a very pronounced effect of the snail extract on gene expression [20].
The obtained results by proteomic analysis on 2D-PAGE of the cortex extract of rats treated with scopolamine and treated with Sco+SE, showed a change in the expression of a number of proteins with diverse functions – structural, regulatory, metabolic, signaling, antioxidant, etc., which are involved in various aspects of AD progression.
The functions and the biological role of identified key proteins from rat cortex are summarized in Table 1 based on protein function in rat brain published in many studies.
Cytoskeletal proteins are important for the construction and functioning of the cytoskeleton. They included actin, tubulin, GFAP (Glial Fibrillary Acidic Protein). A number of studies have shown that the actin cytoskeleton plays a key role in synaptic function and plasticity, since it takes part in different aspects of the coordinated machinery that modulates synaptic transmission [27].
Our results showed an increase in the expression of β-actin in the Sco-treated animals and a decrease in the rats treated with scopolamine and SE together. The increased levels of β-actin in the demented rats likely reflect increased production of the plaque-forming Aβ peptide that is deposited outside neurons. Thus, actin may be positioned at the crossroads between the amyloid cascade and synaptic dysfunction and is likely a key link between them in the pathogenesis of AD.
Tubulin is the major constituent of microtubules, which are composed of α–β-tubulin heterodimers forming linear protofilaments that form a hollow polar cylinder. Microtubules (MT) are essential components of the cytoskeleton of the cell, which has locomotory functions. In the cell, MTs are among the most complex structures in terms of both chemical and functional properties [30]. Our studies showed a decrease in the expression of tubulin in rats with Alzheimer’s type dementia and an increase in the subsequent treatment of the demented animals with SE.
GFAP is a key protein responsible for the cytoskeleton structure of glial cells and for maintaining their mechanical strength, regulating astrocyte morphology and function, as well as maintaining neighboring neurons and the blood-brain barrier, and is not found outside the CNS [29]. Our studies showed no significant differences in GFAP expression in the two groups of rats. This may be due to an earlier stage of development of dementia of the Alzheimer’s type in the Sco group.
Proteins involved in signal pathways mediate significant events in cellular life cycle, and they are closely associated with AD development. Cytoskeletal protein regulators, which are often part of signaling cascades, also play a key role in AD progression. Such proteins are 14-3-3, stathmin, kinesin, cofilin, tropomyosin. 14-3-3 is an adapter protein involved in the regulation of a wide range of general and specialized signaling pathways [35].
Our studies showed a twofold increased expression of 14-3-3 in the Sco group of rats compared to the group treated with Sco and SE together. These results correlate with literature data suggesting an important role of 14-3-3 in regulating MT dynamics by phosphorylating the MT-associated protein tau. Moreover, the 14-3-3 protein is likely to be one of the key components in elucidating the mechanism of action of snail extract on improving memory in Sco-affected demented animals.
Stathmin is a ubiquitous cytosolic phosphoprotein primarily expressed in the nervous system and a member of a family of phosphoproteins that bind to tubulin and destabilize MTs. Microtubules undergo continuous assembly and disassembly in the cytoskeleton of the cell. Stathmin-4 (STM-4) regulates the dynamics of MTs. STM-4 in the unphosphorylated or hypophosphorylated state binds to tubulin and prevents the polymerization of the latter, thereby preventing microtubule assembly. After phosphorylation, stathmin is released from tubulin and allows the formation of MTs. Dysregulation of stathmin and MTs dynamics has been observed in aged animals and in patients with Alzheimer’s disease and depression [37].
Our results showed a multifold increased expression of stathmin in Sco-treated animals and very low levels in the group of rats treated with Sco and SE together. This in turn suggests a positive effect of SE on the cytoskeleton of neurons in general. In our studies, a moderate increase in the expression levels of kinesin-1 heavy chain was observed in the Sco group of rats, which confirms the literature data [36,42].
Cofilin is a ubiquitously expressed protein. Results obtained from multiple studies suggest its critical role in the pathogenesis of AD [23]. Our studies showed a slightly increased expression of cofilin in the animals treated with Sco compared to those co-treated with Sco and SE.
Tropomyosin (Tm) has been shown to be an integral component of the neurofibrillary pathology (NFP) of AD. Studies have shown that a protein other than tau, neurofilaments, MAP2, or ubiquitin and antigenically related to tropomyosin is involved in the abnormal filaments characteristic of NFP of AD. However, the exact role of Tm in AD pathology is still not well understood [33].
Our studies showed a fold reduction in Tm expression levels in rats co-treated with Sco and SE compared to the scopolamine group. This suggests the key role of Tm in AD. The mechanisms of this action remain to be elucidated.
Obligate metabolic enzymes participate in biochemical pathways and cycles in all eukaryotic cells They could be used to determine the metabolic state of brain cells or brain tissue in different pathologic and physiologic conditions. An important represantive is malate dehydrogenase (MDH). MDH catalyses the reduction of aromatic alpha-keto acids in the presence of NADH and plays an essential role in the malate-aspartate shuttle and the citric acid cycle, important for the mitochondrial supply of NADH for oxidative phosphorylation. But the functional significance of changes in MDH levels in AD is unknown [26].
Our data on decreased MDH expression in the Sco group of rats suggest disturbances in neuronal metabolism in AD-type dementia. There is a positive effect of snail mucus on the metabolism.
Heat-shock proteins have many functions – protection of newly synthesized proteins in the ribosomes, correction of the wrong folding of protein in physiological and pathological conditions, regulation of the transduction pathway, replication/transcription and anti-oxidant function. Heat-shock protein 60 (Hsp60) is a chaperone that is localized in the mitochondria and is involved in the correct folding of proteins. Many studies have shown that Hsp60 is localized outside of mitochondria such as in the cytosol, in extracellular vesicles, or on the cell surface. The role of Hsp60 in AD is still unclear. Furthermore, extracellular release of Hsp60 increases the production of other pro-inflammatory factors and promotes neuronal cell death. Hyperactivation of microglia in response to certain negative factors contributes to the progression of several neurodegenerative diseases, including AD [31].
Our results showed an increase in the expression of Hsp60 in the animals treated with Sco and SE together, which is evidence of its protective role.
Ubiquitin carboxyl-terminal hydrolase isozyme L1 (UCH L1) protein belongs to the Ubiquitin system which is a non-lysosomed protein degradation pathway [44]. It is an important cell regulation system and it may be regulated at protein degradation level but not at transcriptional level [35].
Our research showed a decrease in the expression of UCH L1 in the Sco group of rats and an increase in the group of animals treated with Sco and SE together. Restoring Uch-L1 activity may represent a new therapeutic strategy for AD.
Proteins involved in Ca2+ homeostasis and cell bioenergetics that we determined are calbindin, creatine kinase, ATP-synthase. The regulation of intracellular calcium homeostasis is a very complex mechanism that is vital for several cellular pathways and is thus involved in cell survival and death. Cellular bioenergetics is the basis of life processes in all cells under physiological and pathological conditions. Calbindin (CB) buffers cytosolic calcium. It can stimulate membrane Ca2+-ATPase and 3′,5′-cyclic nucleotide phosphodiesterase. CB has a critical role in preventing neuronal death as well as maintaining calcium homeostasis. Although a significant reduction in CB expression was observed in the brains of AD mice and humans, it is not known whether these changes contribute to AD-related dysfunction. Experiments have shown that removal of CB from the amyloid precursor protein presenilin in transgenic mice worsens the pathogenesis of AD, suggesting a critical role of CB in the pathogenesis of AD [25]. Our results confirm the literature data showing a decrease in CB expression in demented animals compared to the rats treated with Sco and SE together. Mitochondrial bioenergetics is not only essential with ATP synthesis, but is also key to the regulation of several other mitochondrial and cellular processes that are affected in AD. Dysregulation of these processes can further increase the extent of mitochondrial dysfunction and cellular damage induced by dysfunctional bioenergetics, creating a vicious cycle that will ultimately lead the cell to apoptosis [32]. Our studies showed a two-fold reduction in the expression of ATP synthase in the rats treated with Sco compared to the group of animals treated with Sco and snail extract together. These results direct attention to this enzyme as a potential therapeutic target for pharmacological preparations aimed at preventing mitochondrial dysfunction in AD and, accordingly, inhibiting apoptosis in neurons.
Because some brain hemoglobin may be derived from the peripheral circulation due to a compromised blood-brain barrier often seen in the brains of elderly individuals and those with AD, research by Chuang and colleagues suggests that the genesis of some plaques may be a consequence from prolonged accumulation of amyloid at sites of vascular injury [45]. Our results showed a fold-increased expression of Hemoglobin (Hb) in the Sco group of rats compared to the insignificant expression in the group of animals treated with Sco and SE together. This is probably indirect evidence of the close relationship between Hb and Aβ in the progression of AD and the improvement of the disease after treatment with snail mucus.
Antioxidant proteins play an important role in nervous system diseases and physiological conditions as well. These are thioredoxin, Cu-Zn Superoxide dismutase (CuZn-SOD).
Thioredoxin (Trx) plays a role in the reversible S-nitrosylation of cysteine residues in target proteins and thereby contributes to the intracellular nitric oxide response. In the brain, Trx is activated upon ischemia/reperfusion and appears to play an important role not only in oxidative stress but also in signal transduction. In vitro and in vivo experiments have demonstrated that enhancement of endogenous Trx expression and administration of exogenous Trx inducers play neuroprotective roles in AD, including inhibition of oxidative stress, inflammation and apoptosis, and activation of survival signaling pathways. These findings suggest that increasing Trx expression and activity may provide a strategy to delay AD progression [21].
Our experiments showed small differences in the expression of Trx in the two groups of experimental animals. There was rather a qualitative difference in the corresponding spot from 2D electrophoresis, namely a distinct spot in the Sco group of rats and several spots of weaker intensity in the rats treated with Sco and SE together (Figure 1). We hypothesize that the snail extract probably activates protein isoenzymes that are involved in neuroprotective mechanisms under conditions of oxidative stress.
4. Materials and Methods
4.1. Snail Extract
The snail mucus was collected from C. aspersum snails, grown in Bulgarian eco-farms using patented technology without suffering for the snails [46]. After homogenization, the crude mucus extract was purified and enriched with the protein fraction over 20 kDa, concentrated by ultrafiltration on 20 kDa polyethersulfone membrane (Microdyn NadirTM from SterliTech Corporation, Goleta. USA). The snail extract used for the experiments in this study contained 50% crude mucus extract and 50% fraction containing compounds with molecular weight above 20 kDa. The snail extract protein concentration was determined by Bradford assay [47].
4.2. Laboratory Animals
Male adult Wistar rats six to eight weeks old, were purchased from a local vendor (Erboj, Slivnica, Sofia). Rats were housed three per cage under constant laboratory conditions (25 ± 30C, 12/12 h light/dark cycle, food and water available ad libitum). Habituation period was for 5 days before starting the experiment.
All experiments have been performed strictly according to the national regulations and European Communities Council Directive (86/609/EEC) also “Principles of laboratory animal care” (NIH publication No. 85-23) concerning the protection of animals used for scientific and experimental purposes. The research with laboratory animals was approved by the Committee of Bioethics at the Institute of Neurobiology, Bulgarian academy of Sciences.
4.3. Scopolamine Induced Dementia
The rats were injected with Scopolamine hydrobromide (2 mg/kg, intraperitoneally- i.p.) for 11 consecutive days. This was done in order to induce AD type dementia. The dose of scopolamine was determined in our previous studies [20].
4.4. Treatment of the Laboratory Animals
The animals were divided into 2 groups: 1) Sco treated control group and 2) Sco + SE treated group. Sco and Sco + SE groups were injected with scopolamine hydrobromide at a dose of 2 mg/kg. Sco + SE group received snail extract from C. aspersum orally (0.5 ml/100 g bw), and Sco group received dH2O orally (0.5 ml/100 g bw) instead SE. Sco was dissolved in distilled water ex tempore before each administration. SE was applied 1h before the Sco injection daily for 16 days (5 days before and 11 days simultaneously with Sco), as describe previously [20].
4.5. Extraction of Proteins from the Rat Brain
Rat brains were rapidly removed after the animals were euthanized via mild CO2 inhalation followed by decapitation. The samples from rat cortex (about 3-4 g) were washed with 10 ml of ice cold PBS buffer (0.2 g KCl, 8 g NaCl, 1.44 Na2HPO4, 0.24 g KH2PO4). Then the samples were homogenized in 0.1 M phosphate buffer, pH 7.8, containing protease inhibitors (Calbiochem Protease Inhibitor Cocktail Set 111). After complete homogenization, the samples were centrifuged at 10 000 rpm for 30 min at 40C. Of the pure supernatants obtained, protein concentrations were assessed using a Bradford Protein Assay Kit (Bio-Rad, Hercules, CA 94547 USA). The supernatants were used for examination by 2D - PAGE.
4.6. 2D-PAGE Analysis
2D-PAGE analysis was conducted to examine change of expression of the proteins as was described in [48]. After determination of the protein concentration of supernatants with the Protein Assay Kit (BioRad), approximately 250 μg of protein was mixed with IPG rehydration buffer, containing 8mol/L urea, 2% w/v CHAPS, 0.3% dithiothreitol (DTT), final volume of 360 μL. The strips were rehydrated for 12 h and focused (IEF) with PROTEAN® i12™ IEF System. At the end of the IEF program, the IPG strips were equilibrated in a 50 mmol/L Tris-HCl solution, pH 8.8, containing 6 mol/L urea, 30% glycerol, 2% SDS, and 1% DTT, for 10 min, after which the solution was replaced with the same solution, but the DTT was exchanged by 5% iodoacetamide. The strips were placed on home-casted vertical 12.0% SDS-PAGE gels. Staining was performed using Coomassie Brilliant Blue G-250 Dye (CBB G-250, Thermo Fisher Scientific).
4.7. Trypsin Digestion of Protein Spots
The trypsin digestion of proteins from 2D PAGE was conducted as was described by Rosenfeld et al. [49]. After two times washing with 150 μL of 200 mmol/L ammonium bicarbonate in 50% ACN/MQ (30 min at 30°C) and subsequently discolored gel pieces were dried in a SpeedVac (Thermo Savant, Holbrook, NY). A volume of 10 μl digestion buffer was added to the dried gel spots. It contained 50 mM ammonium bicarbonate, pH 7.8 and modified trypsin per microliter (Promega). The tubes were kept on ice for 45 min to allow the gel pieces to be completely soaked with the protease solution. Digestion reaction was performed overnight at 37oC. Then the supernatants were recovered and the resulting peptides were extracted twice with 35 μl of 60% ACN / 0.1% DIEA. The extracts were pooled and dried in the speed vac machine (Labconco® Centrivap Concentrator, Kansas City, MO, USA). The extracted peptides were redissolved in 10 μl of 0.1% formic acid and matrix solution (a saturated solution of alphacyano-4-hydroxycinnamic acid in acetonitrile/water 50:50 with 0.1% TFA, for peptide measurements of <10 kDa; and spotted on the MALDI plate.
4.8. Mass Spectrometric Analyses (MS and MS/MS)
After the tryptic digestion, the proteins were analyzed by an Autoflex™III, High-Performance MALDI-TOF & TOF/TOF System (Bruker Daltonics). It is a highly effective system on the base of 200 Hz frequency-tripled Nd–YAG laser, whose wavelengths operates at 355 nm. Samples were prepared by mixing 2.0 μl of the sample with a 2.0 μl matrix solution (7 mg / ml α-cyano-4-hydroxycinnamic acid (CHCA) in 50% acetonitrile (ACN) containing 0.1% trifluoroacetic acid (TFA) and was put on a stainless plate with 192 pits. MS mode and collision energy of 4200 after a total of 3500 shots were used after drying the samples at room temperature. The mass scale was calibrated by solution of protein standards. The external calibration of the apparatus was performed with a mixture of angiotensin I, glu-fibrinopeptide B, ACTH (1–17), and ACTH.
4.9. Identification of Proteins
Identification of proteins from MALDI/MS spectra was achieved using different protein databases by Peptide Mass Fingerprints search by MASCOT http://www.matrixscience.com (MASCOT server, Matrixscience, London, UK). The used parameters were: Type of search: Peptide Mass Fingerprint; Database: SwissProt; Taxonomy: Rattus; Enzyme: Trypsin; Allow up to: 1 - 3 missed cleavages; Monoisotopic; Mass Values MH+; Peptide tolerance: 1.2 - 2.5 and a probabilistic score at p<0.05. Proteins detected in Mascot were checked against the UniProtKB online database (http://www.uniprot.org). The amino acid sequences revealed by manual interpretation of the MS/MS spectra were alignment with Swiss Prot and NCBI BLAST databases (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Evaluation of gel maps was conducted by MelanieTM Coverage 9.2. Difference in protein expression in rat cortex between Sco Control and Sco + SE treated animals, as well as molecular weight (Mw) and isoelectric point (pI) of corresponding proteins were determined by computer-aided 2D image analysis, carried out using MelanieTM Coverage 9.2 software.
5. Conclusion
We found large differences in protein expression in the two groups of rats (Sco and Sco+SE) of Ubiquitin carboxyl-terminal hydrolase isozyme L1, Calbindin, Vacuolar ATP synthase catalytic subunit A, Tropomyosin beta chain, 14-3-3 zeta/delta, Kinesin-1 heavy chain, Stathmin-4 and Hemoglobin subunit alpha-1/2. While increased expression of Hb α subunit is controversial and could reflect the impaired blood-brain barrier, the remaining proteins might be potential drug targets in Alzheimer’s type dementia treatment.
Supplementary Materials
Not applicable.
Author Contributions
V.A., P.D., and L.V. were responsible for the study concept and design. V.A., L.V., and A.D. contributed to the investigation and data acquisition. V.A., L.V., and A.D. drafted the manuscript. R.K., P.D., L.T., and L.V. provided critical revision of the manuscript for important intellectual content. All authors have read and approved the final version for publication.
Funding
This work was supported by Grant KП-06-H73/5-05.12.2023 from the Bulgarian National Science Fund.
Institutional Review Board Statement
The experiments have been performed strictly according to the national regulations and European Communities Council Directive (86/609/EEC) also “Principles of laboratory animal care” (NIH publication No. 85-23) concerning the protection of animals used for scientific and experimental purposes. The research with laboratory animals was approved by the Committee of Bioethics at the Institute of Neurobiology, Bulgarian Academy of Sciences.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Acknowledgments
Project BG05M2OP001-1.002-0019: “Clean Technologies for Sustainable Environment—Waters, Waste, Energy for a Circular Economy”.
Conflicts of Interest
All the authors declare that the research was conducted in the absence of any commercial or financial relationships that could be considered as a potential conflict of interest.
References
- Knopman, D.S.; Amieva, H.; Petersen, R.C.; Chételat, G.; Holtzman, D.M.; Hyman, B.T.; Nixon, R.A; Jones, D.T. Alzheimer disease. Nat Rev Dis Primers 2021, 7(1), 33. [CrossRef]
- Sanabria-Castro, A.; Alvarado-Echeverría, I.; Monge-Bonilla, C. Molecular Pathogenesis of Alzheimer’s Disease: An Update. Ann Neurosci 2017, 24(1), 46-54. [CrossRef]
- Hajjo, R.; Sabbah, D.A.; Abusara, O.H.; Al Bawab, A.Q. A Review of the Recent Advances in Alzheimer’s Disease Research and the Utilization of Network Biology Approaches for Prioritizing Diagnostics and Therapeutics. Diagnostics 2022; 12(12): 2975, 1-40. [CrossRef]
- Nation, D.A.; Sweeney, M.D.; Montagne, A.; Sagare, A.P.; D’Orazio, L.M.; Pachicano, M.; Sepehrband, F.; Nelson, A.R.; Buennagel, D.P.; Harrington, M.G.; Benzinger, T.L.S.; Fagan, A.M.; Ringman, J.M.; Schneider,L.S.; Morris, J.C.; Chui, H.C.; Law, M.; Toga, A.W.; Zlokovic, B.V. Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat. Med 2019, 25, 270–276. [CrossRef]
- Perkovic, M.N.; Paska, A.V.; Konjevod, M.; Kouter, K.; Strac, D.S.; Erjavec, G.N.; Pivac, N. Epigenetics of Alzheimer’s Disease. Biomolecules 2021, 11(2): 195, 1-38. [CrossRef]
- Wenk, G.L. Neuropathologic changes in Alzheimer’s disease. JCP 2003, 64 (9), 7-10. https://pubmed.ncbi.nlm.nih.gov/12934968/.
- Deacon, R.M. Housing, husbandry andbhandling of rodents for behavioral experiments. Nat Protoc 2006, 1(2), 936–946. [CrossRef]
- Benedikz, E.; Kloskowska, E.; Winblad, B. The rat as an animal model of Alzheimer’s disease. J. Cell. Mol. Med 2009, 13(6), 1034-1042. [CrossRef]
- Malekzadeh, S.; Edalatmanesh, M.A.; Mehrabani, D.; Shariati, M. Drugs Induced Alzheimer’s Disease in Animal Model. GMJ 2017, 6(3), 1-12. [CrossRef]
- Chen, W.N.; Yeong, K.Y. Scopolamine, a Toxin-Induced Experimental Model, Used for Research in Alzheimer’s Disease. CNS Neurol Disord Drug Targets 2020, 19(2), 85-93. [CrossRef]
- Yadang, F.S.A.; Nguezeye, Y.; Kom, C.W.; Betote, P.H.D.; Mamat, A.; Tchokouaha, L.R.Y.; Taiwé, G.S.; Agbor, G.A.; Bum, E.N. Scopolamine-Induced Memory Impairment in Mice: Neuroprotective Effects of Carissa edulis (Forssk.) Valh (Apocynaceae) Aqueous Extract. Int J Alzheimers Dis 2020, 6372059, 1-10. [CrossRef]
- Oh, J.H.; Choi, B.J.; Chang, M.S.; Park, S.K. Nelumbo nucifera semen extract improves memory in rats with scopolamine-induced amnesia through the induction of choline acetyltransferase expression. Neurosci Lett 2009, 461(1), 41-44. [CrossRef]
- Perry, E.K.; Tomlinson, B.E.; Blessed, G.; Bergmann, K.; Gibson, P.H.; Perry, R.H. Correlation of cholinergic abnormalities with senile plaques and mental test scores in senile dementia. Br Med J 1978, 2(6150), 1457-1459. [CrossRef]
- Bunadri, P.; Neerati, V.; Merugu, S.; Akondi, B.R. Neuroprotective effect of resveratrol against Scopolamine-induced cognitive impairment and oxidative stress in rats. Arch Biol Sci 2013, 65(4), 1381–1386. [CrossRef]
- Dolashki, A.; Velkova, L.; Daskalova, E.; Zheleva N.; Topalova, Y.; Atanasov, V.; Voelter, W.; Dolashka, P. Antimicrobial Activities of Different Fractions from Mucus of the Garden Snail Cornu aspersum. Biomedicines 2020, 8(9), 315, 1-17. [CrossRef]
- Topalova, Y.; Belouhova, M.; Velkova, L.; Dolashki, A.; Zheleva, N.; Daskalova, E.; Kaynarov, D.; Voelter, W.; Dolashka, P. Effect and Mechanisms of Antibacterial Peptide Fraction from Mucus of C. aspersum against Escherichia coli NBIMCC 8785. Biomedicines 2022, 10(3), 672, 1-19. [CrossRef]
- Velkova, L.; Dolashki, A.; Petrova, V.; Pisareva, E.; Kaynarov, D.; Kermedchiev, M.; Todorova, M.; Dolashka, P. Antibacterial Properties of Peptide and Protein Fractions from Cornu aspersum Mucus. Molecules 2024, 29, 2886. [CrossRef]
- Vassilev, N.G.; Simova, S.D.; Dangalov, M.; Velkova, L.; Atanasov, V.; Dolashki, A.; Dolashka, P. An 1H NMR- and MS-Based Study of Metabolites Profiling of Garden Snail Helix aspersa Mucus. Metabolites 2020, 10(9), 360, 1-15. [CrossRef]
- Sawa, K.; Uematsu, T.; Korenaga, Y.; Hirasawa, R.; Kikuchi, M., Murata K.; Zhang, J.; Gai, X.; Sakamoto, K.; Koyama, T.; Satoh, T. Krebs cycle intermediates protective against oxidative stress by modulating the level of reactive oxygen species in neuronal HT22 cells. Antioxidants 2017, 6(1), 21, 1-9. [CrossRef]
- Tancheva, L.; Lazarova, M.; Velkova, L.; Dolashki, A.; Uzunova, D.; Minchev, B.; Petkova-Kirova, P.; Hassanova, Y.; Gavrilova, P.; Tasheva, K.; Taseva, T.; Hodzhev, Y.; Atanasov, A.G.; Stefanova, M.; Alexandrova, A.; Tzvetanova, E.; Atanasov, V.; Kalfin, R.; Dolashka, P. Beneficial Effects of Snail Helix aspersa Extract in an Experimental Model of Alzheimer’s Type Dementia. JAD 2022, 88(1), 155-175. [CrossRef]
- Jia, J.; Zeng, X.; Xu, G.; Wang, Z. The Potential Roles of Redox Enzymes in Alzheimer’s Disease: Focus on Thioredoxin. ASN neuro 2021, 13, 1759091421994351, 1-13. [CrossRef]
- Tsvetanova, E.; Alexandrova, A.; Georgieva, A.; Tancheva, L.; Lazarova, M.; Dolashka, P.; Velkova, L.; Dolashki, A.; Atanasov, V.; Kalfin, R. Effect of mucus extract of Helix aspersa on scopolamine-induced cognitive impairment and oxidative stress in rat’s brain. BCC 2020, 52(D), 107-111.
- Kang, D.E.; Woo, J.A. Cofilin, a Master Node Regulating Cytoskeletal Pathogenesis in Alzheimer’s Disease. JAD 2019, 72(1), 131-144. [CrossRef]
- Bishop, P.; Rocca, D.; Henley, J.M. Ubiquitin C-terminal hydrolase L1 (UCH-L1): structure, distribution and roles in brain function and dysfunction. Biochem J 2016, 473(16), 2453-2462. [CrossRef]
- Kook, S.Y.; Jeong, H.; Kang, M.J.; Park, R.; Shin, H.J.; Han, S.H.; Son, S.M.; Song, H.; Baik. S.H.; Moon, M.; Yi, E.C.; Hwang, D.; Mook-Jung, I. Crucial role of calbindin-D28k in the pathogenesis of Alzheimer’s disease mouse model. Cell Death Differ 2014, 21(10), 1575-1587. [CrossRef]
- Shi, Q.; Gibson, G.E. Up-regulation of the mitochondrial malate dehydrogenase by oxidative stress is mediated by miR-743a. J Neurochem 2011, 118(3), 440-448. [CrossRef]
- Pelucchi, S.; Stringhi, R.; Marcello, E. Dendritic Spines in Alzheimer’s Disease: How the Actin Cytoskeleton Contributes to Synaptic Failure. Int J Mol Sci 2020, 21(3), 908, 1-23. [CrossRef]
- Bürklen, T.S.; Schlattner, U.; Homayouni, R.; Gough, K.; Rak, M.; Szeghalmi, A.; Wallimann, T. The creatine kinase/creatine connection to Alzheimer’s disease: CK-inactivation, APP-CK complexes and focal creatine deposits. J Biomed Biotechnol 2006, 2006(3), 35936, 1-12. [CrossRef]
- Kamphuis, W.; Middeldorp, J.; Kooijman, L.; Sluijs, J.A.; Kooi, E.J.; Moeton, M.; Freriks, M.; Mizee, M.R.; Hol, E.M. Glial fibrillary acidic protein isoform expression in plaque related astrogliosis in Alzheimer’s disease. Neurobiol Aging 2014, 35(3), 492-510. [CrossRef]
- Sferra, A.; Nicita, F.; Bertini, E. Microtubule Dysfunction: A Common Feature of Neurodegenerative Diseases. Int J Mol Sci 2020, 21(19), 7354, 1-37. [CrossRef]
- Campanella, C.; Pace, A.; Bavisotto, C.C.; Marzullo, P.; Gammazza, A.M.; Buscemi, S.; Piccionello, A.P. Heat Shock Proteins in Alzheimer’s Disease: Role and Targeting. Int J Mol Sci 2018, 19(9), 2603, 1-22. [CrossRef]
- Patro, S.; Ratna, S.; Yamamoto, H.A.; Ebenezer, A.T.; Ferguson D.S.; Kaur, A.; McIntyre, B.C.; Snow, R.; Solesio, M.E. ATP Synthase and Mitochondrial Bioenergetics Dysfunction in Alzheimer’s Disease. Int J. Mol Sci 2021, 22(20), 11185, 1-17. [CrossRef]
- Gunning, P.; O’Neill, G.; Hardeman, E. Tropomyosin-based regulation of the actin cytoskeleton in time and space. Physiol Rev 2008, 88(1), 1-35. [CrossRef]
- Gu, Q.; Cuevas, E.; Raymick, J.; Kanungo, J.; Sarkar, S. Downregulation of 14-3-3 Proteins in Alzheimer’s Disease. Mol Neurobiol 2020, 57(1), 32-40. [CrossRef]
- Qureshi, H.Y.; Li, T.; MacDonald, R.; Cho, C.M.; Leclerc, N.; Paudel, H.K. Interaction of 14-3-3ζ with microtubule-associated protein tau within Alzheimer’s disease neurofibrillary tangles. Biochemistry 2013, 52(37), 6445–6455. [CrossRef]
- Selvarasu, K.; Singh, A.K.; Iyaswamy, A.; Sreenivasmurthy, S.G.; Krishnamoorthi, S.; Bera, A.K.; Huang, J.D.; Durairajan, S.S.K. Reduction of kinesin I heavy chain decreases tau hyperphosphorylation, aggregation, and memory impairment in Alzheimer’s disease and tauopathy models. Front Mol Biosci 2022, 9, 1050768, 1-16. [CrossRef]
- Uchida, S.; Shumyatsky, G.P. Deceivingly dynamic: Learning-dependent changes in stathmin and microtubules. Neurobiol Learn Mem 2015, 124, 52-61. [CrossRef]
- Walser, M.; Svensson, J.; Karlsson, L.; Motalleb, R.; Åberg, M.; Kuhn, H.G.; Isgaard, J.; Åberg, N.D. Growth Hormone and Neuronal Hemoglobin in the Brain-Roles in Neuroprotection and Neurodegenerative Diseases. Front. Endocrinol 2021, 11, 606089, 1-15. [CrossRef]
- Zhao, Y.; Zhao, B. Oxidative stress and the pathogenesis of Alzheimer’s disease. Oxid Med Cell Longev 2013, 316523, 1-11. [CrossRef]
- Makhaeva, G.F.; Lushchekina, S.V.; Boltneva, N.P.; Sokolov, V.B.; Grigoriev, V.V.; Serebryakova, O.G.; Vikhareva, E.A.; Aksinenko, A.Y.; Barreto, G.E.; Aliev, G.; Bachurin, S.O. Conjugates of γ-carbolines and phenothiazine as new selective inhibitors of butyrylcholinesterase and blockers of NMDA receptors for Alzheimer disease. Sci Rep 2015, 5, 13164, 1-11. [CrossRef]
- Ni, R.; Marutle, A.; Nordberg, A. Modulation of α7 nicotinic acetylcholine receptor and fibrillary amyloid-β interactions in Alzheimer’s disease brain. JAD 2013, 33(3), 841–851. [CrossRef]
- Sun T.; Yu N.; Zhai L.K.; Li N.; Zhang C.; Zhou L.; Huang Z.; Jiang X.Y.; Shen Y.; Chen Z.Y. (2013) c-Jun NH2-terminal kinase (JNK)-interacting protein-3 (JIP3) regulates neuronal axon elongation in a kinesin- and JNK-dependent manner. J Biol Chem 2013, 288(20), 14531-14543. [CrossRef]
- DeSalle, L.M.; Pagano, M. Regulation of the G1 to S transition by the ubiquitin pathway. FEBS Lett 2001, 490(3), 179-189. [CrossRef]
- Chait, B.T.; Kent, S.B. Weighing naked proteins: practical, high-accuracy mass measurement of peptides and proteins. Science 1992, 257(5078), 1885-1894. [CrossRef]
- Chuang, J.Y.; Lee, C.W.; Shih, Y.H.; Yang, T.; Yu, L.; Kuo, Y.M. Interactions between amyloid-beta and hemoglobin: implications for amyloid plaque formation in Alzheimer’s disease. PloS One 2012, 7(3):e33120, 1-10. [CrossRef]
- Dolashka, P.; Atanasov, D. Device for Collecting Extracts from Garden Snail. BG Utility Model Application Number 2656, 08.11.2013. Patent Number 2097, 31 August 2015. Available online: https://portal.bpo.bg/bpo_online/-/bpo/utility-model-detail (accessed on 19 May 2024).
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976, 72, 248-254. https://10.1006/abio.1976.9999.
- O’Farrell PH. High resolution two-dimensional electrophoresis of proteins. J Biol Chem. 1975, 250(10), 4007-21.
- Rosenfeld, J.; Capdevielle, J.; Guillemot, J.C.; Ferrara, P. In-gel digestion of proteins for internal sequence analysis after one- or two-dimensional gel electrophoresis. Anal. Biochem., 1992, 203, 173-179.
Figure 1.
2D-PAGE of cellular proteins from rat Cortex: (A) treated with Sco and (B) treated with Sco + SE (pI 3–10, 12.0% SDS-polyacrylamide gels - Coomassie Brilliant Blue G-250 staining). Note: Annotated numbers refer to the identification of the proteins in Table 1.
Figure 1.
2D-PAGE of cellular proteins from rat Cortex: (A) treated with Sco and (B) treated with Sco + SE (pI 3–10, 12.0% SDS-polyacrylamide gels - Coomassie Brilliant Blue G-250 staining). Note: Annotated numbers refer to the identification of the proteins in Table 1.
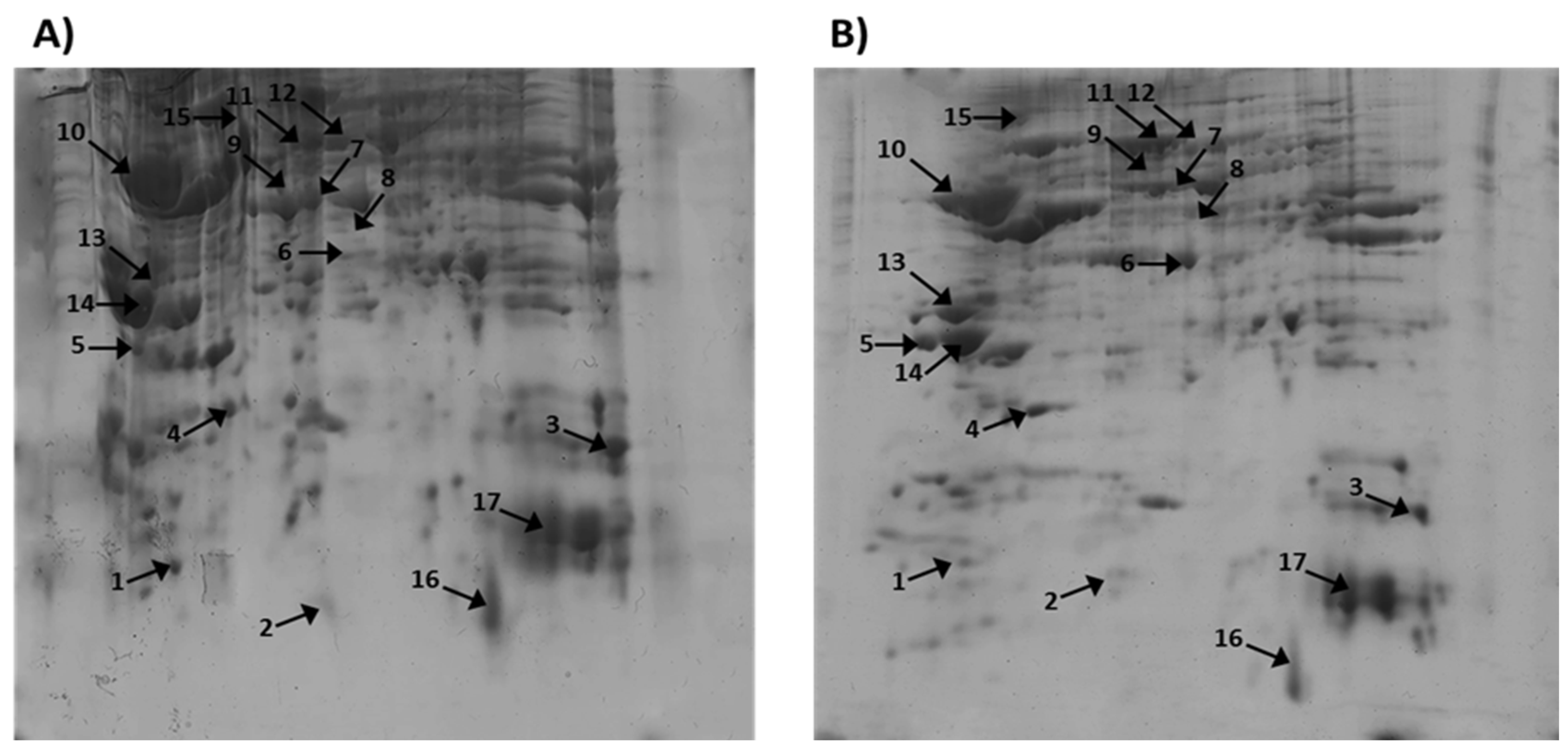
Figure 2.
Mass spectrometric analysis of protein on spot A14 from 2D-PAGE: A) MS spectrum of peptides obtained after trypsin digestion of spot 14 with experimental MW 27.754 kDa and pI 4.73; B) MS/MS spectrum from MALDI-TOF-TOF analysis of peptide at m/z 1548.37 from spot 14: 1433Z_RAT.
Figure 2.
Mass spectrometric analysis of protein on spot A14 from 2D-PAGE: A) MS spectrum of peptides obtained after trypsin digestion of spot 14 with experimental MW 27.754 kDa and pI 4.73; B) MS/MS spectrum from MALDI-TOF-TOF analysis of peptide at m/z 1548.37 from spot 14: 1433Z_RAT.
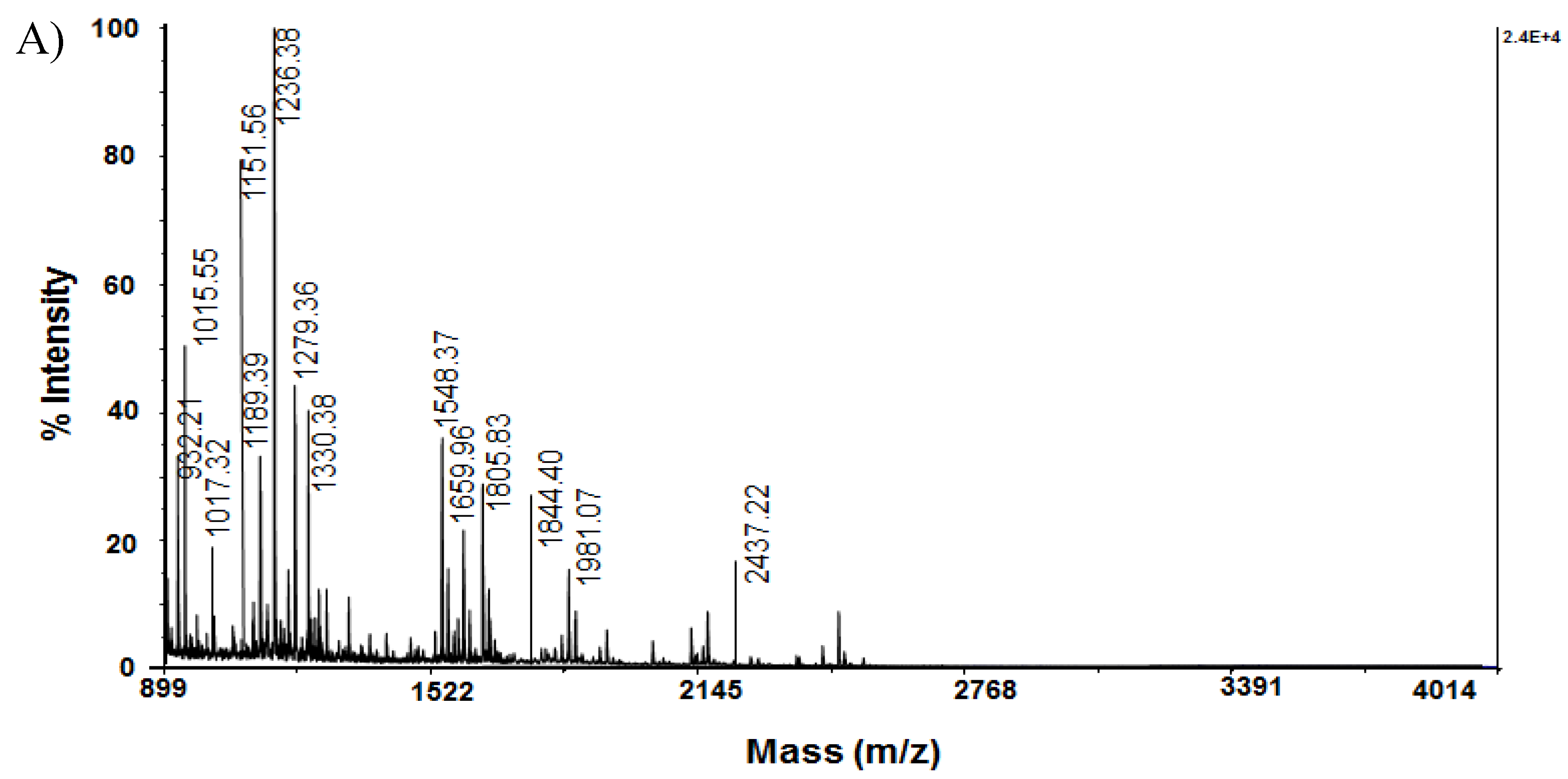
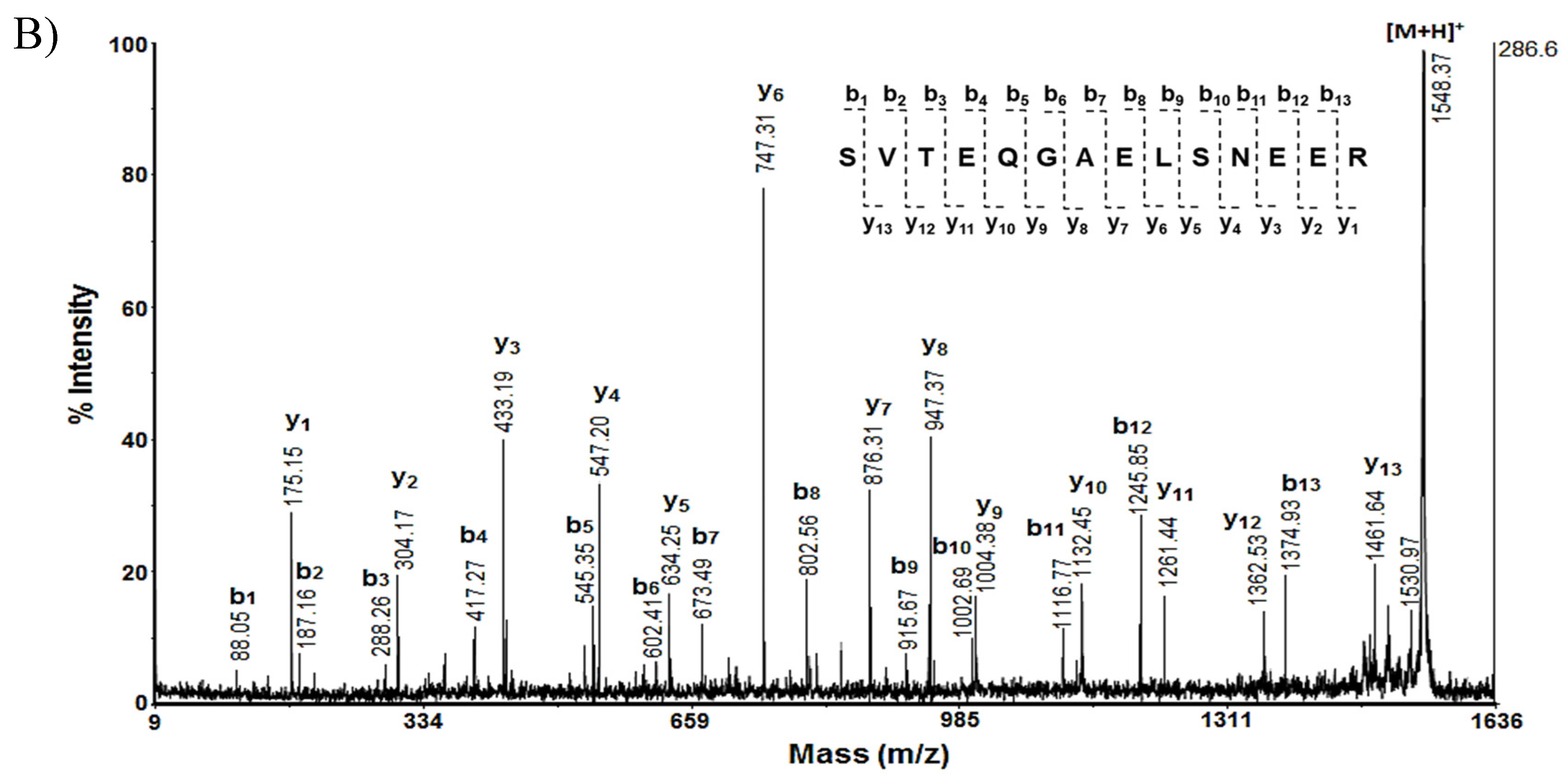
Figure 3.
Mass spectrometric analysis of protein on spot A 16 from 2D-PAGE: A) MS spectrum of peptides obtained after trypsin digestion of spot 16 with experimental MW 22.073 kDa and pI 5.76; B) MS/MS spectrum from MALDI-TOF-TOF analysis of peptide at m/z 1572.46 from spot 16: STMN4_RAT.
Figure 3.
Mass spectrometric analysis of protein on spot A 16 from 2D-PAGE: A) MS spectrum of peptides obtained after trypsin digestion of spot 16 with experimental MW 22.073 kDa and pI 5.76; B) MS/MS spectrum from MALDI-TOF-TOF analysis of peptide at m/z 1572.46 from spot 16: STMN4_RAT.
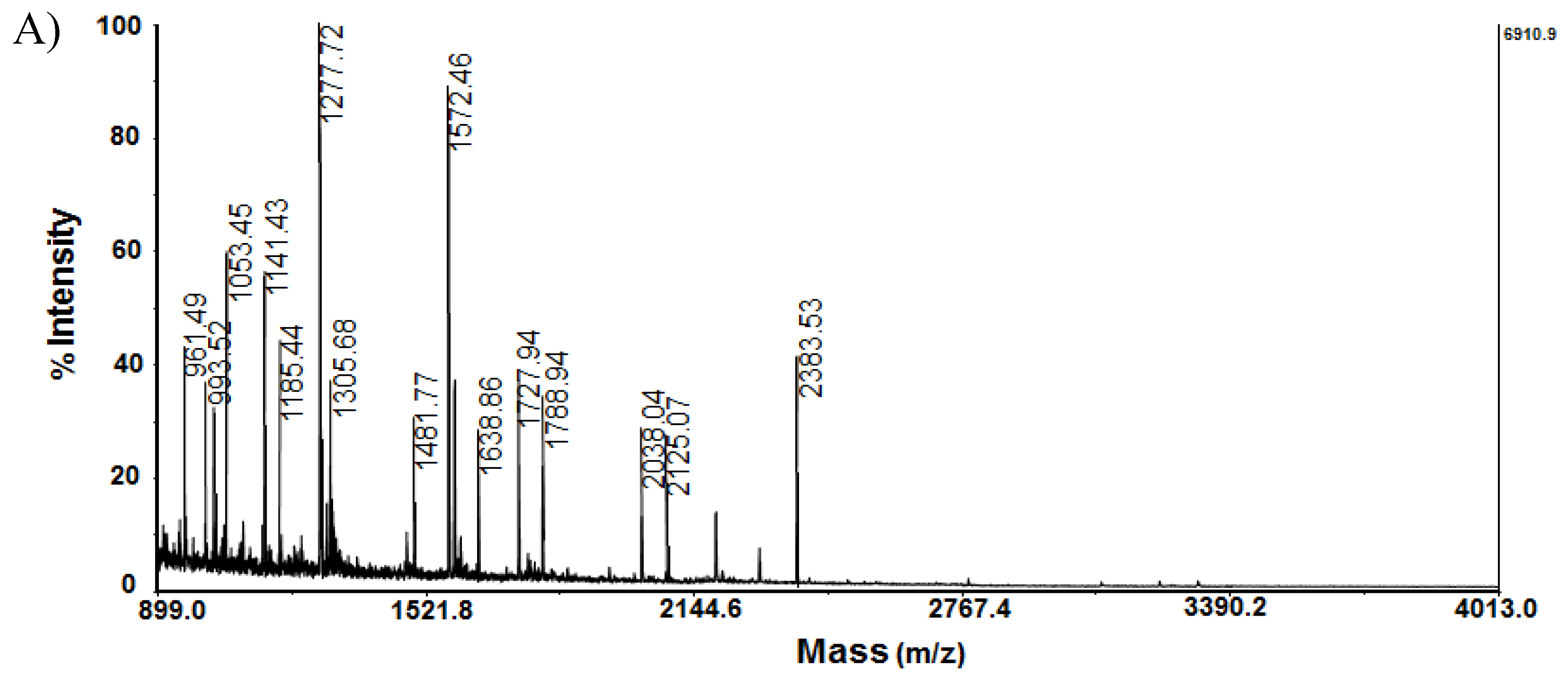
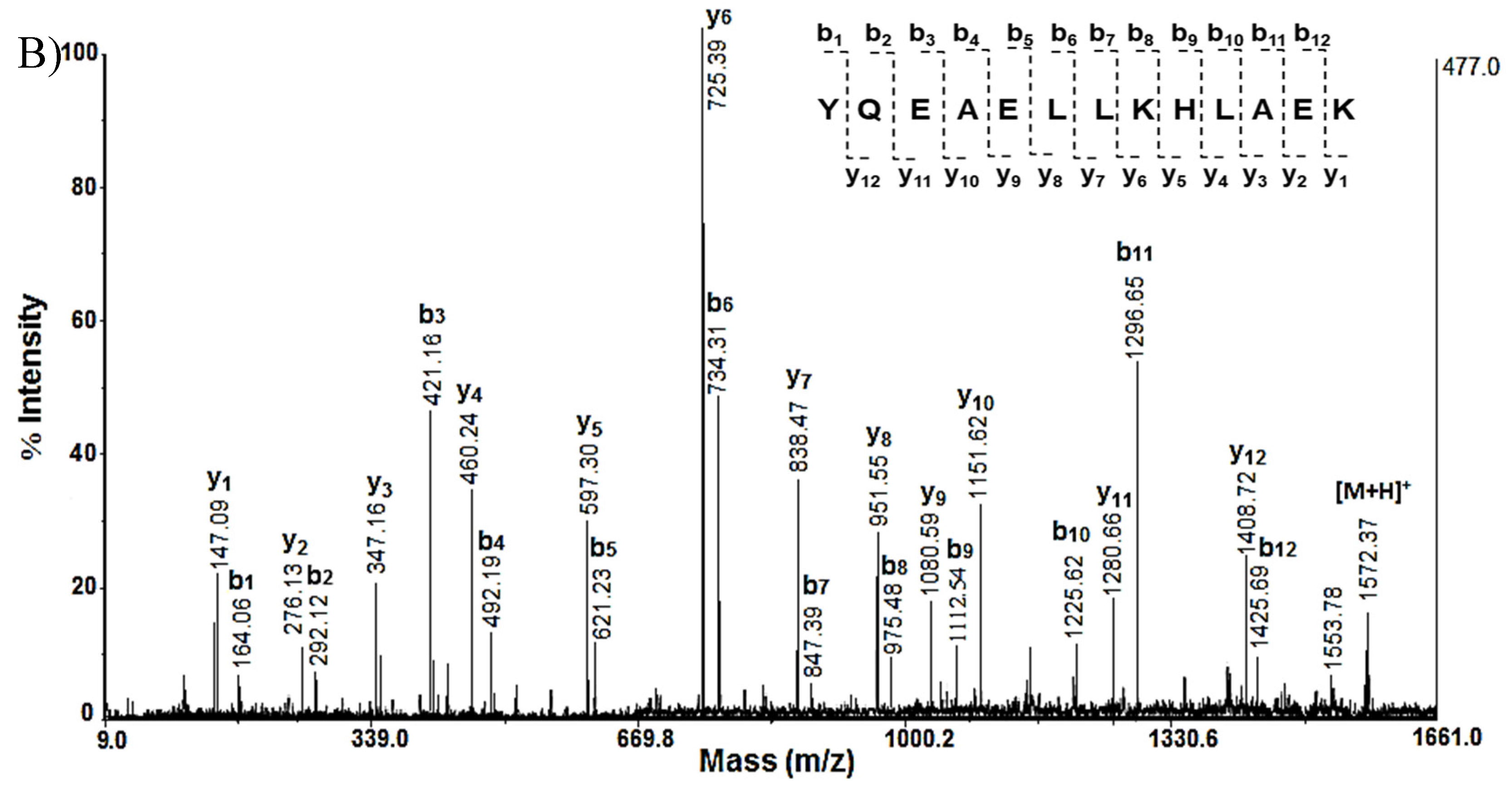

Table 1.
Functions and biological role of key proteins from rat cortex.
| Spot № | Protein Name | Molecular Function | Biological Process |
|---|---|---|---|
| 1 | Thioredoxin P11232 (THIO_RAT) | Thioredoxin (Trh) nitrosylates the active Cys site of caspase 3 in response to nitric oxide and thereby inhibits caspase-3 activity. | Trh has multiple biological functions, including protective cellular mechanisms against oxidative stress and cytokine-induced damage. Enhancement of endogenous Trx expression and administration of exogenous Trx inducers play neuroprotective roles in BA and activate pro-survival signaling pathways [21]. |
| 2 | Superoxide dismutase [Cu-Zn] P07632, SODC_RAT) | CuZnSOD catalyzes the dismutation of the highly reactive superoxide anion to O2 and to the less reactive species H2O2. | It is suggested that CuZnSOD plays a major role in the antioxidant defense system of nervous tissue. In aging and in AD disease, a decrease in the expression levels of the enzyme is usually observed [22]. |
| 3 | Cofilin-1; P45592, (COF1_RAT) | Cofilin binds to F-actin and exhibits pH-sensitive F-actin depolymeri- zing activity. | Cofilin is required for neural tube morphogenesis and neural crest cell migration. Activated cofilin acts as a bridge between actin and microtubule dynamics by displacing tau from microtubules and leading to tauopathy [23]. |
| 4 | Ubiquitin carboxyl-terminal hydrolase isozyme L1 Q00981 (UCHL1_RAT) | This enzyme is a thiol protease that recognizes and hydrolyzes a peptide bond at the C-terminal glycine of ubiquitin. | There is evidence that UCH L1 binds and co-localizes with monoubiquitin and prolongs the half-life of ubiquitin. and Uch-L1 reduction is part of a cycle that favors Aβ accumulation in vascular injury [24]. |
| 5 | Calbindin P07171 (CALB1_RAT) | Calbindin (CB) buffers cytosolic calcium. It can stimulate membrane Ca2+-ATPase and 3′,5′-cyclic nucleotide phosphodiesterase. | CB has a critical role in maintaining calcium homeostasis and preventing neuronal death. Experiments have shown that removal of CB from the amyloid precursor protein presenilin in transgenic mice worsens the pathogenesis of AD, suggesting a critical role of CB in the pathogenesis of AD [25]. |
| 6 | Malate dehydrogenase, cytoplasmic O88989 (MDHC_RAT) | Catalyzes the reduction of aromatic alpha-keto acids in the presence of NADH. | It plays an essential role in the malate-aspartate shuttle and the citric acid cycle, important for the mitochondrial supply of NADH for oxidative phosphorylation. The functional significance of MDH elevation in AD is unknown [26]. |
| 7 | Actin, cytoplasmic 1 P60711 (ACTB_RAT) | Actins are highly conserved proteins that are involved in various types of cell motility and are ubiquitously expressed in all eukaryotic cells. | A number of studies have shown that the actin cytoskeleton plays a key role in synaptic function and plasticity. It may be positioned at the crossroads of pathways contributing to AD pathogenesis, i.e., between the amyloid cascade and synaptic dysfunction [27]. |
| 8 | Creatine kinase B-type P07335 (KCRB_RAT) | Creatine kinase (CK ) reversibly catalyzes the transfer of phosphate between ATP and various phosphogens such as creatine phosphate. | CK plays a major role in the cellular energetics of the brain, so any disruption of this enzyme can worsen AD pathology. Cytosolic brain-type creatine kinase (BB-CK) is inactivated by oxidation in patients with AD [28]. |
| 9 | Glial fibrillary acidic protein P47819 (GFAP_RAT) | GFAP is a class III intermediate filament. It is a cell-specific marker that distinguishes astrocytes from other glial cells during development of the central nervous system. | GFAP is a key protein and is a marker of astroglial damage. It is responsible for the structure of the cytoskeleton of glial cells, regulation of the morphology and function of astrocytes, as well as for maintaining the blood-brain barrier. In AD, amyloid plaques are surrounded by reactive astrocytes with increased expression of GFAP filaments [29]. |
| 10 | Tubulin alpha-1B chain Q6P9V9 (TBA1B_RAT) | Tubulin is the main constituent of microtubules, which are composed of α–β-tubulin heterodimers forming linear protofilaments that form a hollow polar cylinder. | Microtubules (MT) are essential components of the cytoskeleton of the cell, which has locomotory functions. A number of studies show that MT-dysfunction can contribute to or be the cause of neurodegenerative processes and AD in particular. Disruption of the neuronal cytoskeleton is a feature of the neurodegenerative brain, and tubulin levels are decreased in the AD brain [30]. |
| 11 | 60 kDa heat shock protein, mitochondrial P63039 (CH60_RAT) | Hsp60 is a chaperone that is localized in the mitochondria and is involved in the correct folding of proteins. | Hsp60 is a protein that, together with Hsp10, is considered essential for mitochondrial protein folding. The role of Hsp60 in AD is still unclear [31,33]. |
| 12 | V-type proton ATPase catalytic subunit A P50516 (VATA_MOUSE) | ATP-synthase is the last enzyme of the mitochondrial electron transport chain, where oxidative phosphorylation takes place in order to synthesize ATP, which is a universal energy carrier in the cell. | In mammalian cells, ATP synthase, in addition to ATP synthesis, can also perform its degradation (ATPase), which indicates the important role of this enzyme in the regulation of cellular bioenergetics and metabolism. The mechanisms of bioenergetic dysfunction incl. dysregulation of ATP synthase in AD remains unclear [32]. |
| 13 | Tropomyosin beta chain P58775 (TPM2_RAT) | Tropomyosin (Tm) binds to actin filaments in muscle and non-muscle cells. In non-muscle cells, it is involved in the stabilization of cytoskeletal actin filaments. | Tm has been shown to be an integral component of the neurofibrillary pathology of Alzheimer’s disease. However, the exact role of Tm in AD pathology is still not well understood [33]. |
| 14 | 14-3-3 protein zeta/delta P63102 (1433Z_RAT) | 14-3-3 is an adapter protein involved in the regulation of a wide range of general and specialized signaling pathways. Binding usually results in modulation of the activity of the protein to which 14-3-3 binds. | 14-3-3 isoforms regulate a wide range of cellular processes such as the cell cycle, transcription, intracellular trafficking, apoptosis, and autophagy [34]. In AD, they are thought to contribute to NFT formation through τ-hyperphosphorylation [35]. |
| 15 | Kinesin-1 heavy chain Q2PQA9 (KINH_RAT) | Kinesin is a microtubule-dependent motor protein required for the normal distribution of mitochondria and lysosomes. Required for anterograde axonal transport of MAPK8IP3/JIP3, which is essential for MAPK8IP3/JIP3 function in axon elongation [43] | Studies have shown that Kinesin-1 heavy chain, which is part of a key molecular motor protein, is involved in tau homeostasis in AD cells and animal models. It has been proposed that reduction of Kinesin-1 heavy chain levels is sufficient to prevent and/or delay abnormal tau behavior in AD and other tauopathies [36]. |
| 16 | Stathmin-4 P63043 (STMN4_RAT) | Stathmin (STM) is a ubiquitous cytosolic phosphoprotein primarily expressed in the nervous system and a member of a family of phosphoproteins that bind to tubulin and destabilize MTs. | STM-4 in the unphosphorylated or hypophosphorylated state binds to tubulin and prevents its polymerization, thereby preventing MTs assembly. After phosphorylation, STM-4 is released from tubulin and allows the formation of MTs. Dysregulation of STM and MTs dynamics has been observed in aged animals and in patients with AD and depression [37]. |
| 17 | Hemoglobin subunit alpha-1/2P01946 (HBA_RAT) | It participates in the transfer of oxygen from the lung to the various peripheral tissues. | Studies have shown that hemoglobin (Hb) binds to Aβ and co-localizes with plaques and vascular amyloid deposits in the brains of AD patients after death. Research by Chuang et al., 2012 suggests that the genesis of some plaques may be a consequence of prolonged amyloid accumulation at sites of vascular injury [38]. |
Table 2.
Key proteins from rat cortex of Sco-treated and Sco + SE treated animals with their MW, pI and expression calculated by Melanie Coverage 9.2 software and confirmed by the MASCOT software based on the performed MALDI-MS analyses.
Table 2.
Key proteins from rat cortex of Sco-treated and Sco + SE treated animals with their MW, pI and expression calculated by Melanie Coverage 9.2 software and confirmed by the MASCOT software based on the performed MALDI-MS analyses.
| № Spot Cortex |
Protein Name (UniProtKB) |
Volume (pixels) Sco |
Volume (pixels) Sco + SE |
MW Exp. |
pI Exp. |
MW Theor |
pI Theor |
Expression |
|---|---|---|---|---|---|---|---|---|
| 1 | Thioredoxin; P11232 (THIO_RAT) | 6050118 | 5441182 | 15.081 | 4.48 | 11.673 | 4.80 |  |
| 2 | Superoxide dismutase [Cu-Zn]; P07632 ( SODC_RAT) | 6261736 | 7118268 | 18.158 | 5.51 | 15.912 | 5.88 |  |
| 3) | Cofilin-1; P45592, (COF1_RAT) | 1157164 | 1101119 | 20 | 7.46 | 18.533 | 8.22 |  |
| 4 | Ubiquitin carboxyl-terminal hydrolase isozyme L1; Q00981 (UCHL1_RAT) | 2204242 | 3605218 | 22 | 4.9 | 24.838 | 5.14 |  |
| 5 | Calbindin; P07171 (CALB1_RAT) | 8396846 | 11756730 | 28 | 3.94 | 29.994 | 4.71 |  |
| 6 | Malate dehydrogenase, cytoplasmic; O88989 (MDHC_RAT) | 12166374 | 15772607 | 36 | 5.63 | 36.483. | 6.16 |  |
| 7 | β –Actin; P60711 (ACTB_RAT) | 18729148 | 13744467 | 48 | 5.35 | 41.737 | 5.2 |  |
| 8 | Creatine Kinase B-type; P07335 (KCRB_RAT) |
375048 | 448176 | 40 | 5.52 | 42.725 | 5.4 |  |
| 9 | Glial Fibrillary Acidic Protein; P47819 (GFAP_RAT) |
582304 | 583050 | 53 | 5.17 | 49.957 | 5.02 |  |
| 10 | Tubulin α-1 chain; Q6P9V9 (TBA1B_RAT) | 22419986 | 7427465 | 53 | 4.35 | 50.152 | 4.94 |  |
| 11 | 60 kDa heat shock protein, mitochondrial; P63039 (CH60_RAT) | 10059479 | 13101991 | 71 | 5.44 | 60.955 | 5.91 |  |
| 12 | Vacuolar ATP synthase catalytic subunit A; P50516 (VATA_RAT) | 2211577 | 3933414 | 73 | 5.28 | 68.326 | 5.41 |  |
| 13 | Tropomyosin beta chain P58775 (TPM2_RAT) |
2502851 | 451024 | 32.817 | 4.66 | 41.253 | 4.45 |  |
| 14 | 14-3-3 protein zeta/delta P63102 (1433Z_RAT) |
15894376 | 7086121 | 27.754 | 4.73 | 33.326 | 4.46 |  |
| 15 | Kinesin-1 heavy chain Q2PQA9 (KINH_RAT) |
4906008 | 3993752 | 109.463 | 6.06 | 126.318 | 5.51 |  |
| 16 | Stathmin-4; P63043 (STMN4_RAT) | 39377068 | 10224790 | 22.073 | 5.76 | 17.650 | 6.42 |  |
| 17 | Hemoglobin subunit alpha-1/2; P01946 (HBA_RAT) | 27582716 | 2551968 | 15.319 | 7.82 | 21.167 | 7.7 |  |
Table 3.
Amino acid sequences of tryptic peptides of several spots from 2D-PAGE images, determined by MS/MS-analyses.
Table 3.
Amino acid sequences of tryptic peptides of several spots from 2D-PAGE images, determined by MS/MS-analyses.
| Spot No | AAS of peptide |
Mass [M+H]+ | Spot No | AAS of peptide |
Mass [M+H]+ | |
|---|---|---|---|---|---|---|
|
6 |
DLDVAVLVGSMPR | 1371.71 |
14 |
MKGDYYR | 932.21 | |
| FVEGLLPNDFSR | 1393.68 | DSTLIMQLLR | 1189.39 | |||
| SQIALKLGVTADDVK | 1558.84 | KEMQPTHPIR | 1236.38 | |||
| VIVVGNPANTNCLTASK | 1700.89 | SVTEQGAELSNEER | 1548.37 | |||
| GEFITTVQQRGAAVIK | 1719.94 | LAEQAERYDDMAACMK | 1844.40 | |||
|
7 |
EITALAPSTMK | 1161.61 |
15 |
KMEENEK | 908.29 | |
| IWHHTFYNELR | 1515.74 | YQQEVDRIK | 1177.35 | |||
| SYELDPGQVITIGNER | 1790.87 | EYELLSDELNQK | 1479.48 | |||
| YPIEHGIVTNWDDMEK | 1946.89 | TQMLDQEELLASTRR | 1791.35 | |||
| VAPEEHPVLLTEAPLNPK | 1954.06 | GLEETVAKELQTLHNLR | 1949.51 | |||
| 10 | YMACCLLYR | 1135.50 | 16 | MTLAAYKEK | 1053.45 | |
| QLFHPEQLITGK | 1410.77 | EAHLAAMLER | 1141.43 | |||
| SIQFVDWCPTGFK | 1527.73 | RKYQEAELLK | 1277.72 | |||
| IHFPLATYAPVISAEK | 1756.67 | YQEAELLKHLAEK | 1572.46 | |||
| EDAANNYARGFYTIGK | 1790.87 | MKELPLVSLFCSCFLSDPLNK | 2383.53 | |||
| KKMQMLK | 907.24 | MFAAFPTTK | 1013.31 | |||
| HIAEDSDR | 941.24 | LRVDPVNFK | 1087.42 | |||
| 13 | AEFAERSVAK | 1107.91 | 17 | IGGHGGEYGEEALQR | 1572.47 | |
| LEEAEKAADESER | 1475.70 | TYFSHIDVSPGSAQVK | 1735.55 | |||
| LEEAEKAADESERGMK | 1791.71 | AADHVEDLPGALSTLSDLHAHK | 2296.75 | |||
| TIDDLEDEVYAQKMKYK | 2088.89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



