
Submitted:
10 October 2024
Posted:
10 October 2024
You are already at the latest version
Abstract
The aim of the study was to analyze the association between single nucleotide polymorphism (SNP) rs80860411A>C located in the intergenic region of chromosome 13 (near the GALNT15 gene encoding Polypeptide N-Acetylgalactosaminyltransferase15, GALNTL2) and fat-tening and slaughter traits of pigs. The study was conducted on 426 individuals belonging to three breeds: Polish Landrace (n = 192), Polish Large White (n = 187) and Pulawska (n = 48). The rs80860411 genotypes were determined using the PCR-RFLP method. Association analysis was performed for each breed separately, as well as for the entire study group. The study showed that rs80860411A>C had a significant effect on fattening performance traits, as well as on several slaughter performance traits, including: width of the loin eye, loin and ham weights without skin and backfat and carcass meat content (P ≤ 0.01, P ≤ 0.05). The obtained results indicate that the studied SNP has the potential to be included in pig selection programs, thus potentially influencing the improvement of fat-tening and slaughter traits of different pig breeds.
Keywords:
rs80860411
; GALNT15
; fattening traits
; slaughter traits
; pigs
1. Introduction
Meat quality is a very important aspect of pig breeding. The definition of quality was created centuries ago, and the first mentions in the literature indicate the ancient period. This concept is not easy to define, because it depends on many factors. In addition to genetic factors, such as breed and sex, and physiological factors (age, condition, health), meat quality is also influenced by environmental factors (animal husbandry system, nutrition, transport, slaughter and post-slaughter processing). One of the most important economic characteristics of pigs is the skeletal muscle mass of fattening pigs. Their development and growth is a factor that ensures the profitability of breeding and an adequate supply of meat consumed by humans [1,2].
Analysis of genes regulating skeletal muscle development is of great importance for understanding the molecular basis of this phenomenon. Polymorphic variants of many genes affect the differentiation of fattening efficiency. The results of genetic and genomic studies published in recent years have significantly contributed to a better understanding of many molecular mechanisms involved in the process of muscle tissue synthesis [3,4]. Genetic markers that are predictive of pork quality can be used in selection programs [5]. The recessive allele of the RYR1 gene was considered for years to be the most important genetic marker in pig breeding [6]. Currently, polymorphism in intron 3 of the IGF2 gene, encoding insulin like growth factor 2, is the most important genetic marker for fattening, slaughter and meat quality traits in pigs [7,8,9]. Mutation in the IGF2 gene has been introduced in recent years (thanks to the availability of genome editing techniques) to the gene pool in breeds in which it does not occur naturally [10]. Other important genetic polymorphisms for pig breeding have been located in the MC4R, CAST, PRKAG3 genes [5,11].
Whole genome sequencing (WGS) is an effective technique for identifying common DNA sequence variation, including genetic variants that affect various traits of farm animals [12]. Genotype is one of the main factors determining carcass composition and quality, therefore, ongoing studies for years have been aimed at identifying QTL (Quantitative Trait Loci) and QTN (Quantitative Trait Nucleotide) [13,14]. Thanks to the use of genome-wide association studies (GWAS), the identification of polymorphic variants with QTN potential has become feasible. The analysis of the occurrence of polymorphic variants (most often SNPs) is widely used to detect genotype-phenotype associations [15,16,17].
The aim of this study was to analyze the association between the rs80860411 polymorphism located in intergenic region, which was selected in genomic analyses [18] and fattening and slaughter traits of Polish Landrace, Polish Large White and Pulawska pigs.
2. Materials and Methods
Animals and Nutrition
The research was conducted using material from 426 individuals of three breeds: Polish Landrace (n = 192), Polish Large White (n = 187) and the native Pulawska breed (n = 48). The Polish Landrace animals came from 29 sires and 100 dams, the Polish Large White from 40 sires and 114 dams, and the Pulawska from 15 sires and 32 dams. The studied groups of animals were not significantly related to each other, therefore the sire effect was not included in the statistical models.
The material was obtained from Pig Performance Control Stations located in Poland, where routine pig assessment is conducted. Therefore, there was no need for the approval of the research by the bioethics committee.
The animals were fed, kept and slaughtered under standard conditions to determine their fattening and slaughter performance and to estimate their breeding value. The pigs were kept in individual pens until they reached 30 kg of body weight. During fattening, they received two types of feed (from 30 to 80 kg of body weight and from 80 to 100 kg of body weight) administered ad libitum until they reached a body weight of 100 (± 2.5) kg. The slaughter was carried out by exsanguination after stunning with high-voltage electric forceps.
Fattening Performance Test
During the experiment, the following fattening traits were recorded at the experimental stations: average daily gain from 30 to 100 kg of body weight as test daily gain (g/day), lifetime daily gain (g/day), daily feed intake (kg), feed conversion ratio (kg/kg gain), age at slaughter (days) and number of days in the test (days) [19,20].
Carcass Traits
After slaughter, the half carcasses were cut into pieces taking into account the parameters of muscle content, fat content and basic meat quality features in accordance with the station methodology procedure. The following measurements were made: carcass yield (%), middle length of carcass (cm), loin weight (kg), loin and ham weights without skin and backfat (kg), width of loin eye (cm), height of loin eye (cm), loin eye area (cm2), carcass meat content (%), average backfat thickness (cm) and weight of primary cuts (kg).
The average backfat thickness was determined using five measurements, which included: the thickest point over the shoulder blade, the place on the back above the joint located between the last thoracic vertebra and the first lumbar vertebra, and three measurement over the loin – above the rostral, middle and caudal edge of the gluteus muscle cross-section [21]. The loin eye area was determined by cutting the muscle between the last thoracic vertebra and the first lumbar vertebra. The measurement was made based on the contour made in the cephalic plane of the plane of cut.
Genotyping
Genomic DNA was isolated from samples of the longissimus dorsi muscle. The following kits were used for this purpose: Genomic Mini and Sherlock AX (A&A Biotechnology, Poland) and ReliaPrepTM gDNA Tissue Miniprep System (Promega, USA).
Genotypes were analyzed by PCR-RFLP. PCR products were amplified using two pairs of primers (Table 1) that were designed using the PRIMER3 program (https://primer3.ut.ee/). Due to the lack of PCR product in the Pulawska breed, the second pair of primers was designed. Additionally, DNA sequencing of the longer PCR product was performed to determine the cause of the lack of PCR product in the Pulawska breed (potential polymorphism at the primer(s) annealing site for the first set of primers). The existence of the A/C polymorphism was also confirmed by sequencing (Sanger method) of homozygous genotypes (AA and CC) for the rs80860411. Chromas software (https://technelysium.com.au/wp/, version 2.6.6) was used to analyze the sequencing results.
The PCR mixture contained ~100 ng of genomic DNA, 12 pmol of each primer, 1× PCR buffer (with 2 mM MgCl2), 0.2 mM dNTP mix, 0.3 U Taq polymerase, and deionized water (up to 12 µl). The following thermal profile was used: 94°C for 5 minutes, followed by 35 cycles of 94°C for 30 seconds, 60°C for 35 seconds, and 72°C for 35 seconds (50 seconds for the longer PCR product in the Pulawska breed), and a final extension at 72°C for 5 minutes. The obtained amplicons (424 and 842 bp) were digested with the restriction enzyme DraI (EURx) for 1 h at 37°C. The resulting DNA fragments were separated electrophoretically in 2% agarose gels stained with ethidium bromide in 1x TBE buffer (120 V, 40 min). The results were visualized under UV light and archived (Quantum, VILBER).
Statistical Analysis
In each analyzed herd of pigs, the frequencies of genotypes and alleles were estimated. The frequencies of individual genotypes within breeds were compared using the chi-square test.
In the analysis of the association between the tested genotype and fattening and slaughter performance traits, the following linear model (General Linear Model) was applied using SAS software (ver. 8.02):
Yijk = μ + bi + gj + (bg)ij + βSWk + eijk
where:
Yijk – observation;
μ – overall mean;
bi – fixed effect of i breed;
gj – fixed effect of j genotype;
(bg)ij – interaction between i breed and j genotype;
βSWk – linear effect of slaughter weight as covariate (for slaughter traits only);
eijk – random residual error.
The least squares method (LSM) was used to determine the significance of differences between genotype groups.
3. Results
In order to confirm the occurrence of the rs80860411 polymorphism, sequencing of samples designated as homozygous genotypes was performed (Figure 1).
The obtained PCR products of 424 bp length were digested with DraI enzyme. This allowed for the determination of three genotypes based on the obtained restriction fragments (Figure 2).
In the Pulawska breed, digestion of PCR products (842 bp) with DraI enzyme allowed obtaining three genotypes; the restriction fragment lengths were consistent with the design (Figure 3).
In this study, the AC genotype was found to be most frequent in Polish Landrace and Pulawska breeds. The CC genotype was most frequently observed in Polish Large White breed, while it was not found in the Polish Landrace breed. In all the breeds studied, the AA genotype was found to be least frequent (Table 2).
The Hardy-Weinberg equilibrium was checked using the chi-square test. The distribution of genotypes differed between the breeds studied. The observed and expected genotype frequencies in Pulawska and Polish Large White breeds did not differ significantly, so these herds were in Hardy-Weinberg equilibrium. However, Polish Landrace herd was not in Hardy-Weinberg equilibrium.
The presented study showed significant associations between the rs80860411 polymorphism and fattening traits, as well as slaughter traits in Polish pig breeds (Table 3 and Table 4).
A significant effect of the analyzed SNP was demonstrated on all fattening traits examined, except for daily feed intake, in the Pulawska breed. Individuals with the AC genotype were characterized by a significantly higher test daily gain (p ≤ 0.01) and lifetime daily gain (p ≤ 0.05) compared to AA homozygous individuals. However, in pigs with the AA genotype the highest number of days on test, feed conversion and age at slaughter were observed; these traits were significantly higher (p ≤ 0.01, p ≤ 0.05) than in heterozygotes and CC homozygous pigs. In the case of the remaining two breeds, no significant differences were found between the values of the analyzed traits.
The statistical analysis of slaughter traits, taking into account the right half-carcass, showed highly significant relationships (p ≤ 0.01) between the rs80860411 polymorphism and the width of the loin eye, the carcass meat content and the weight of primary cuts in the Pulawska breed. The values of these traits were significantly higher in individuals with the AA and AC genotypes compared to CC homozygotes. This polymorphism was also significantly associated (p ≤ 0.05) with the loin eye area in the Pulawska and Polish Landrace breeds. In the case of Polish Large White pigs, no significant relationships were found between the analyzed polymorphism and the traits included in the study.
4. Discussion
The main factor that determines the profitability of pig farming is the appropriate feed. It is assumed that feed costs account for about 70% of expenses. Rational feeding has a positive effect on the growth, development, health of individuals, as well as on reproductive and slaughter performance. A balanced diet also affects the better achievement of genetic potential by animals. Feed conversion describes the degree to which the animal uses the food or energy contained in the feed and other nutrients taken with it. The average feed conversion is about 2.8-3 kg/kg of body weight gain for fattening pigs. Carcass yield is the ratio of the carcass weight to the pre-slaughter weight of the animal expressed as a percentage (%), and its average value is from 75 to 78% and depends on the age and degree of fattening of individuals [22]. Feed intake and feed conversion ratio are among the main factors that influence the economic efficiency of pig production. Fatness traits are characterized by high phenotypic variability, and the heritability coefficient was estimated at about 0.5. This indicates a significant contribution of genetic background in the formation of this group of traits. Hence, it is indicated that they can be improved or modified using appropriate genetic markers [16].
Our study shows variable occurrence of alleles for rs80860411A>C between the studied breeds. Compared to the study conducted by Terman et al. [23], which aimed to estimate the association between ACLY gene polymorphism and fattening and quality traits of pork, the frequency of genotypes and alleles in our study was distributed in a similar way, but the CC genotype did not occur in the Polish Landrace breed. The heterozygous genotype occurred with the highest frequency in the Polish Landrace and Pulawska breeds. The AA genotype occurred with the lowest frequency in all studied breeds. The genotype distribution differs between the breeds studied. Polish Landrace herd was out of Hardy-Weinberg equilibrium, which this may be due to the selection carried out in the herd. The pig breeds selected in the study are among the most popular breeds in Poland, of which Polish Landrace and Polish Large White are meat breeds. The fat-meat type representative is the Pulawska breed, which is a native breed covered by the protection program. Additionally, this breed is characterized by resistance to diseases, excellent adaptation to environmental conditions, and longevity [24].
In the current literature, there is an increasing number of publications on gene searches for the detection of single SNPs (QTN). Ahmed et al. [25] in their work recorded nearly 56.5 million SNPs in genomic data of 155 cattle samples. About 25.87 million biallelic SNPs were identified in Cashmere cattle, occurring mainly in intergenic (58.20%) and intronic (37.71) regions. Only 0.85% of all SNPs were detected in exons. Additionally, 0.9% of SNPs were found in untranslated regions and 0.1% in splicing sites.
In the study conducted by Piórkowska et al. [26] the region of chromosome 15 of pigs (SSC15) was described. This region is very rich in quantitative trait loci (QTL) related to pork quality, growth performance, fat and meat content in the carcass. The study included two pig breeds, Pulawska and Polish Landrace. In the Pulawska breed, characterized by high meat quality, mutations were observed mainly in non-coding regions, such as introns and intergenic regions. An interesting panel of gene variants was indicated, which can be used to understand the impact on important production traits of pigs in further association studies.
Since most of the significant variants identified in genome-wide association studies are located in introns and intergenic regions [27], it is worth paying more attention to them. The rs80860411A>C polymorphism analyzed in the presented work is located relatively close to the GALNT15 gene.
The GLANT15 (GALNTL2) gene is localized on porcine chromosome 13 (NCBI, Gene ID: 100157572, updated on 17-Aug-2024). The gene product catalyses the initial reaction in O-linked oligosaccharide biosynthesis (the transfer of an N-acetyl-D-galactosamine residue to a serine or threonine residue on the protein receptor). It is able to transfer GalNAc residues to the Muc5AC peptide [28]. Protein glycosylation is an essential process in all eukaryotes. Studies have shown that a wide variety of types of protein glycosylation occur in animals, plants, and microorganisms. It plays a significant role in the process of protein folding, their transport to target sites, as well as their activity and localization within the cell. In the body, glycoproteins participate in many biological processes, and their most important functions include cell recognition, differentiation, development. They also participate in signal transduction [29].
One of the most numerous forms of protein glycosylation in animals is considered to be mucin-type O-glycosylation, consisting of glycans attached via O-linked N-acetylgalactosamine (GalNAc) to serine and threonine residues. In most cases, the initiation of glycosylation, i.e., the attachment of the first glycan to the appropriate amino acid residue in the protein, is controlled by one or two genes encoding enzymes responsible for this step. However, a large family of up to 20 homologous genes encoding UDP-GalNAc:polypeptide GalNAc-transferases (GalNAc-Ts) is involved in the control of mucin-type O-glycosylation. Therefore, this type of glycosylation is characterized by the highest potential for differential regulation in cells. The GalNAc-T family is the largest family of glycosyltransferase enzymes with a high degree of evolutionary conservation in animals. However, it has not been found in bacteria, yeast, and plants. It has been shown that single GALNT genes encoding GalNAc-T are important in animals, but their large number does not provide full functional redundancy [30].
GALNTs enzymes catalyze the addition of GalNAc sugar to Ser and Thr residues, resulting in the formation of Tn glycan. GALNTs are activated following relocation from the Golgi apparatus to the endoplasmic reticulum. This process is mediated by Src tyrosine kinase, as well as by the Arf1-COPI protein complex. The GALNTs activity constitutes a powerful mechanism of control of O-glycosylation, which has been termed the GALA pathway [31]. O-GlcNAc glycosylation, as an important mechanism of protein glycosylation coordinated by the aforementioned O-GlcNAc transferase, as well as O-GlcNAcase, helps maintain skeletal muscle structure and function. As a result of glycosylation, the conformational stability and solubility of proteins as well as the thermal stability of enzymes are significantly increased [32]. Muscle proteome analysis revealed the presence of many O-GlcNAc-modified proteins [33] and among them several key contractile proteins of striated muscles were identified [34]. A specific role of O-GlcNAcylation in acute skeletal muscle damage has also been demonstrated. It may become an important issue in the field of non-hereditary muscle diseases [32].
Valdés-Hernández et al. [35] identified potential target genes related to FA (fatty acids) metabolism, which play a role in lipid metabolism (e.g., ADIPOQ, FGFR4, PLIN1, NEU3, TFRC), carbohydrate metabolism (e.g., GALNT15, ADIPOQ, NEU3, PPP1R3D), glucose metabolism (MAFA) and ion binding (e.g., ADIPOQ, GOT1, SOD3, NUDT14). Among the listed FA-correlated genes, GALNT15 is included in the group of genes involved in meat quality. The results of this experiment may facilitate the implementation of breeding strategies based on the use of functional information and deepen the knowledge of gene regulation in pig muscle.
Many GWAS conducted in pigs focus on meat quality traits, hematological parameters and leukocyte function. There are also studies in which genetic markers were associated with slaughter traits, such as carcass meat content, which is currently one of the most important commercial traits. It is worth mentioning that ham is one of the most valuable pork products on the world markets. Thanks to the use of GWAS, 18 significant SNPs were detected in SSC2, which were associated with muscle mass traits [36]. These results were the basis for further studies for later use in marker-assisted selection programs aimed at improving muscle mass in pigs. The effect of iron overload on gene expression in mouse skeletal muscle and heart was investigated. Some of the identified genes were involved in modulation of lipid and glucose metabolism, transcription as well as responses to cellular stress. As a result of the study, a list of 14 genes, including the GALNTL2 gene, was obtained, whose expression in skeletal muscle was increased by iron [37].
It has been described that due to the increasing incidence of obesity, insulin resistance and hyperlipidemia, the number of patients with fatty liver disease is still increasing. There are differences in susceptibility to steatohepatitis, as well as in progression to cirrhosis. This is attributed to the complex interaction of extrinsic and genetic factors that affect the intracellular network. Thanks to multidisciplinary studies, a list of 208 genes co-expressed in fibroblasts from two patients with fatty liver disease was obtained. Among them was the GALNTL2 gene and other related genes, such as: DACT1, MAPK14, PCNXL2, RGN, TLR4, ZNF521 [38].
In the study by Funari et al. [39], 161 cartilage-selective genes were identified, and among them was GALNTL2 gene. These genes have been identified as highly expressed in cartilage, but with low expression and low variability in 34 non-cartilage tissues. Among these 161 genes, a number of genes specific to cartilage were identified, such as cartilage collagen genes, as well as 25 genes associated with skeletal phenotypes in humans and/or mice. In addition, many other cartilage-selective genes were identified, the role of which in cartilage has not yet been determined, or new, as yet undescribed genes.
Through the integrative analysis of GWAS, eQTL (expression quantitative trait loci) and QTT (quantitative trait transcript), GALNT15/GALNTL2 were shown to be strong candidate genes for drip loss in pork [18]. Recent studies provide valuable information on the genetic basis of qualitative and quantitative meat traits. This significantly expands the number of candidate genes that can be used for future functional analysis and genetic improvement of pork quality.
5. Conclusions
The results of the studies on pigs belonging to the breeds: Polish Large White, Polish Landrace and Pulawska prove that the studied polymorphic site, rs80860411, may be responsible for fattening traits and contribute to a better understanding of the functionality of the region located near the not yet well-known GALNT15 gene. Statistically significant differences were found between the genotypes and selected fattening and slaughter traits of pigs. To sum up, our studies can be considered the first step towards determining the functionality of the rs80860411 region located near the GALNT15 gene and its effect on improving fattening and slaughter traits of polish pigs.
Funding
The study was partially funded by statutory activity of the National Research Institute of Animal Production no. 01-11-01-21.
Institutional Review Board Statement
The bioethics committee approval of this study is not necessary because the experiment was carried out on biological material derived from the Pig Testing Stations of the National Research Institute of Animal Production where pigs were fed, maintained and slaughtered in standardized conditions to estimate their performance and breeding values.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mohammadabadi M, Bordbar F, Jensen J, Du M, Guo W. Key Genes Regulating Skeletal Muscle Development and Growth in Farm Animals. Animals (Basel). 2021 Mar 16;11(3):835. [CrossRef] [PubMed]
- Yan E, Guo J, Yin J. Nutritional regulation of skeletal muscle energy metabolism, lipid accumulation and meat quality in pigs. Anim Nutr. 2023 Jun 7;14:185-192. [CrossRef]
- Rothschild MF, Hu ZL, Jiang Z. Advances in QTL mapping in pigs. Int J Biol Sci. 2007 Feb 10;3(3):192-7. [CrossRef]
- Zeng Q, Du ZQ. Advances in the discovery of genetic elements underlying longissimus dorsi muscle growth and development in the pig. Anim Genet. 2023 Dec;54(6):709-720. [CrossRef]
- Rohrer GA, Nonneman DJ, Miller RK, Zerby H, Moeller SJ. Association of single nucleotide polymorphism (SNP) markers in candidate genes and QTL regions with pork quality traits in commercial pigs. Meat Sci. 2012 Dec;92(4):511-8. [CrossRef]
- Fujii, J. , Otsu K., Zorzato F., De Leon S., Khanna V.K., Weiler J.E., O’Brien P.J., MacLennan D.H. (1991) – Identification of a mutation in porcine ryanodine receptor associated with malignant hyperthermia. Science 253, 448-451. [CrossRef]
- Nezer C, Moreau L, Brouwers B, Coppieters W, Detilleux J, Hanset R, Karim L, Kvasz A, Leroy P, Georges M. An imprinted QTL with major effect on muscle mass and fat deposition maps to the IGF2 locus in pigs. Nat Genet. 1999 Feb;21(2):155-6. [CrossRef]
- Van Laere AS, Nguyen M, Braunschweig M, Nezer C, Collette C, Moreau L, Archibald AL, Haley CS, Buys N, Tally M, Andersson G, Georges M, Andersson L. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature. 2003 Oct 23;425(6960):832-6. [CrossRef] [PubMed]
- Zhang J, Chai J, Luo Z, He H, Chen L, Liu X, Zhou Q. Meat and nutritional quality comparison of purebred and crossbred pigs. Anim Sci J. 2018 Jan;89(1):202-210. [CrossRef]
- Xiang G, Ren J, Hai T, Fu R, Yu D, Wang J, Li W, Wang H, Zhou Q. Editing porcine IGF2 regulatory element improved meat production in Chinese Bama pigs. Cell Mol Life Sci. 2018 Dec;75(24):4619-4628. [CrossRef]
- López-Buesa P, Burgos C, Galve A, Varona L. Joint analysis of additive, dominant and first-order epistatic effects of four genes (IGF2, MC4R, PRKAG3 and LEPR) with known effects on fat content and fat distribution in pigs. Anim Genet. 2014 Feb;45(1):133-7. [CrossRef]
- Li S, Lei H, Li J, Sun A, Ahmed Z, Duan H, Chen L, Zhang B, Lei C, Yi K. Analysis of genetic diversity and selection signals in Chaling cattle of southern China using whole-genome scan. Anim Genet. 2023 Jun;54(3):284-294. [CrossRef]
- Ron M, Weller JI. From QTL to QTN identification in livestock--winning by points rather than knock-out: a review. Anim Genet. 2007 Oct;38(5):429-39. [CrossRef]
- Wang M, Wang Q, Pan Y. From QTL to QTN: candidate gene set approach and a case study in porcine IGF1-FoxO pathway. PLoS One. 2013;8(1):e53452. [CrossRef]
- Salas RC, Mingala CN. Genetic Factors Affecting Pork Quality: Halothane and Rendement Napole Genes. Anim Biotechnol. 2017 Apr 3;28(2):148-155. [CrossRef]
- Ropka-Molik K, Pawlina-Tyszko K, Żukowski K, Tyra M, Derebecka N, Wesoły J, Szmatoła T, Piórkowska K. Identification of Molecular Mechanisms Related to Pig Fatness at the Transcriptome and miRNAome Levels. Genes (Basel). 2020 May 29;11(6):600. [CrossRef]
- Pocrnic I, Lourenco D, Misztal I. SNP profile for quantitative trait nucleotide in populations with small effective size and its impact on mapping and genomic predictions. Genetics. 2024 Jun 24:iyae103. [CrossRef]
- Liu X, Zhang J, Xiong X, Chen C, Xing Y, Duan Y, Xiao S, Yang B, Ma J. An Integrative Analysis of Transcriptome and GWAS Data to Identify Potential Candidate Genes Influencing Meat Quality Traits in Pigs. Front Genet. 2021 Oct 21;12:748070. [CrossRef]
- Blicharski T, Polok P, Snopkiewicz M. Results of the pig assessment in 2017 [Wyniki oceny trzody chlewnej w 2017 roku]. Polish Association of Pig Breeders and Producers POLSUS [Polski Związek Hodowców i Producentów Trzody Chlewnej POLSUS], Warsaw 2018, pp. 5-35.
- Różycki, M. , Tyra M. Results of fattening and slaughter performance assessment of pigs at control stations [Wyniki oceny użytkowości tucznej i rzeźnej świń w stacjach kontroli]. Stan Hodowli i Wyniki Oceny Świń w roku 2012. Zespół Wydawnictw i Poligrafii IZ PIB, Kraków 2013, s. 49 -72. (in polish).
- Tyra M, Żak G. Analysis of relationships between fattening and slaughter performance of pigs and the level of intramuscular fat (IMF) in longissimus dorsi muscle. Ann Anim Sci. 2012 Apr 12;12(2):169-178. [CrossRef]
- Karpesiuk, K. Current problems and new challenges in pig nutrition [Aktualne problemy i nowe wyzwania w w zakresie żywienia świń] in Basic issues in the field of pig breeding and husbandry – current problems and new challenges [Podstawowe zagadnienia w zakresie chowu i hodowli trzody chlewnej – aktualne problemy i nowe wyzwania]; Pawłowski R., Ed.; Publisher Warmińsko-Mazurski Ośrodek Doradztwa Rolniczego in Olsztyn, Poland; 2020; pp. 41-59.
- Terman, A. , Woźniak-Męch K., Korpal A., Polasik D., Tyra M., Szyndler-Nędza, M., Żak G., Rybarczyk A., Dybus A. Association Between ATP Citrate Lyase (ACLY) Gene Polymorphism and Fattening, Slaughter and Pork Quality Traits in Polish Pigs. Ann Anim Sci. 2021 Oct 28;21(4):1301-1313. [CrossRef]
- Polasik D, Tyra M, Szyndler-Nędza M, Korpal A, Woźniak-Męch K, Terman A. Relationship between VRTN gene polymorphism and growth, slaughter and meat quality traits in three polish pig breeds. Cienc Agrotec. 2018; 42(5):540-549. [CrossRef]
- Ahmed Z, Xiang W, Wang F, Nawaz M, Kuthu ZH, Lei C, Xu D. Whole-genome resequencing deciphers patterns of genetic diversity, phylogeny, and evolutionary dynamics in Kashmir cattle. Anim Genet. 2024 May 10. [CrossRef]
- Piórkowska K, Żukowski K, Ropka-Molik K, Tyra M. Deep sequencing of a QTL-rich region spanning 128-136Mbp of pig chromosome 15. Gene. 2018 Mar 20;647:268-275. [CrossRef]
- Zhang J, Chen W, Chen G, Flannick J, Fikse E, Smerin G, Degner K, Yang Y, Xu C; Consortium AMP-T2D-GENES; Li Y, Hanover JA, Simonds WF. Ancestry-specific high-risk gene variant profiling unmasks diabetes-associated genes. Hum Mol Genet. 2024 Apr 8;33(8):655-666. [CrossRef]
- Cheng L, Tachibana K, Iwasaki H, Kameyama A, Zhang Y, Kubota T, Hiruma T, Tachibana K, Kudo T, Guo JM, Narimatsu H. Characterization of a novel human UDP-GalNAc transferase, pp-GalNAc-T15. FEBS Lett. 2004 May 21;566(1-3):17-24. [CrossRef] [PubMed]
- Lin B, Qing X, Liao J, Zhuo K. Role of Protein Glycosylation in Host-Pathogen Interaction. Cells. 2020 Apr 20;9(4):1022. [CrossRef]
- Bennett EP, Mandel U, Clausen H, Gerken TA, Fritz TA, Tabak LA. Control of mucin-type O-glycosylation: a classification of the polypeptide GalNAc-transferase gene family. Glycobiology. 2012 Jun;22(6):736-56. [CrossRef]
- Chia J, Tay F, Bard F. The GalNAc-T Activation (GALA) Pathway: Drivers and markers. PLoS One. 2019 Mar 19;14(3):e0214118. [CrossRef]
- Dang K, Jiang S, Gao Y, Qian A. The role of protein glycosylation in muscle diseases. Mol Biol Rep. 2022 Aug;49(8):8037-8049. [CrossRef]
- Cieniewski-Bernard C, Montel V, Berthoin S, Bastide B (2012) Increasing O-GlcNAcylation level on organ culture of soleus modulates the calcium activation parameters of muscle fibers. PLoS One 7:e48218. [CrossRef]
- Cieniewski-Bernard C, Lambert M, Dupont E, Montel V, Stevens L, Bastide B (2014) O-GlcNAcylation, contractile protein modifications and calcium affinity in skeletal muscle. Front Physiol 5:421. [CrossRef]
- Valdés-Hernández J, Folch JM, Crespo-Piazuelo D, Passols M, Sebastià C, Criado-Mesas L, Castelló A, Sánchez A, Ramayo-Caldas Y. Identification of candidate regulatory genes for intramuscular fatty acid composition in pigs by transcriptome analysis. Genet Sel Evol. 2024 Feb 12;56(1):12. Erratum in: Genet Sel Evol. 2024 Feb 26;56(1):14. doi: 10.1186/s12711-024-00885-8. [CrossRef]
- Liu X, Wang LG, Luo WZ, Li Y, Liang J, Yan H, Zhao KB, Wang LX, Zhang LC. Genome-wide SNP scan in a porcine Large White×Minzhu intercross population reveals a locus influencing muscle mass on chromosome 2. Anim Sci J. 2014 Dec;85(12):969-75. [CrossRef]
- Rodriguez A, Hilvo M, Kytömäki L, Fleming RE, Britton RS, Bacon BR, Parkkila S. Effects of iron loading on muscle: genome-wide mRNA expression profiling in the mouse. BMC Genomics. 2007 Oct 19;8:379. [CrossRef]
- Jozefczuk J, Kashofer K, Ummanni R, Henjes F, Rehman S, Geenen S, Wruck W, Regenbrecht C, Daskalaki A, Wierling C, Turano P, Bertini I, Korf U, Zatloukal K, Westerhoff HV, Lehrach H, Adjaye J. A Systems Biology Approach to Deciphering the Etiology of Steatosis Employing Patient-Derived Dermal Fibroblasts and iPS Cells. Front Physiol. 2012 Sep 3;3:339. [CrossRef]
- Funari VA, Day A, Krakow D, Cohn ZA, Chen Z, Nelson SF, Cohn DH. Cartilage-selective genes identified in genome-scale analysis of non-cartilage and cartilage gene expression. BMC Genomics. 2007 Jun 12;8:165. [CrossRef]
Figure 1.
Sequencing results for the polymorphic site rs80860411A>C (A – genotype CC; B – genotype AA).
Figure 1.
Sequencing results for the polymorphic site rs80860411A>C (A – genotype CC; B – genotype AA).
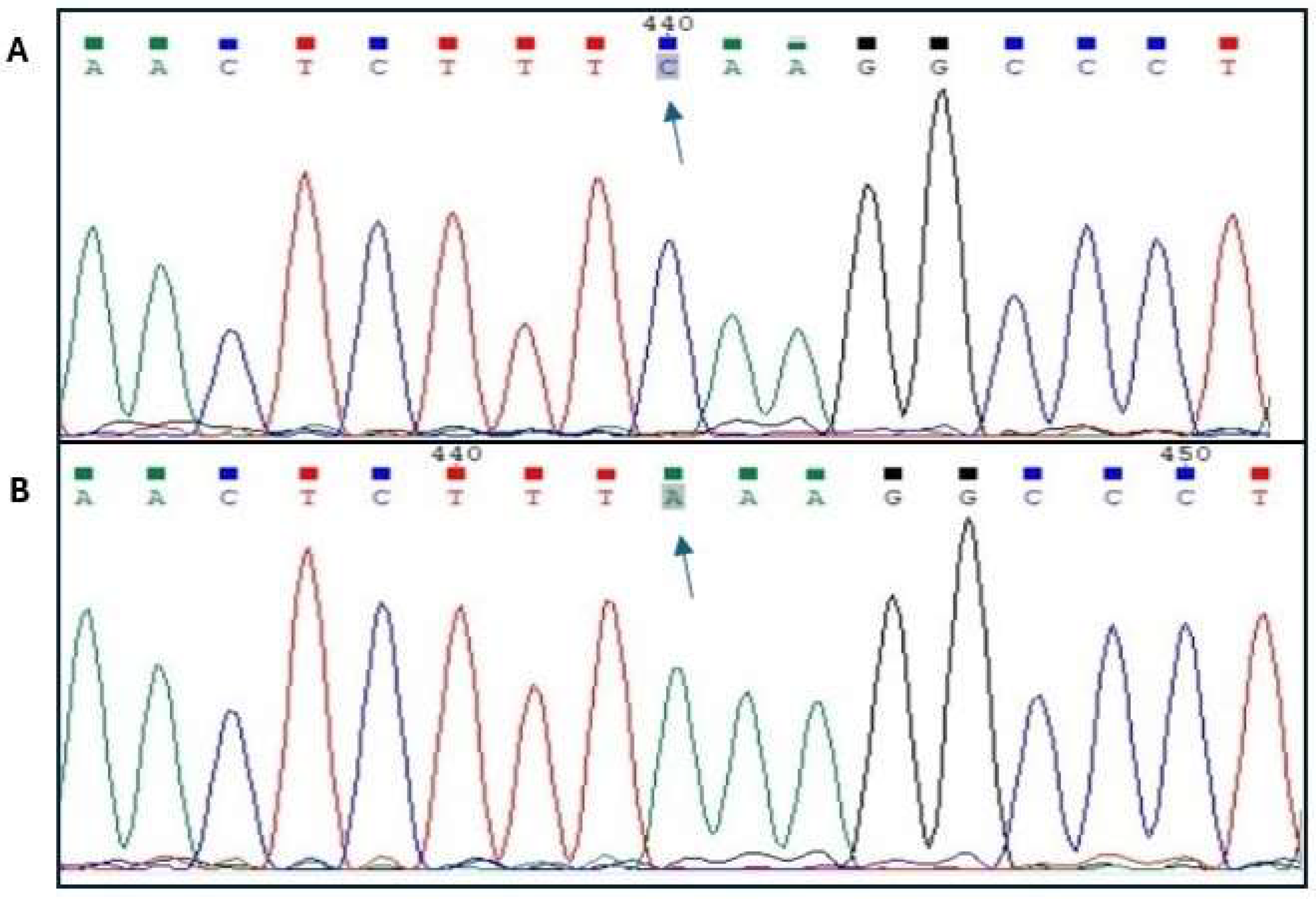
Figure 2.
Results of rs80860411A>C genotyping by PCR-RFLP method; from left: pUC19/MspI DNA marker (Thermo Scientific, Waltham, MA, USA); lanes 2, 6, 9 – AC genotype, lanes 3, 4, 5 and 8 – CC genotype, lane 7 – AA genotype).
Figure 2.
Results of rs80860411A>C genotyping by PCR-RFLP method; from left: pUC19/MspI DNA marker (Thermo Scientific, Waltham, MA, USA); lanes 2, 6, 9 – AC genotype, lanes 3, 4, 5 and 8 – CC genotype, lane 7 – AA genotype).
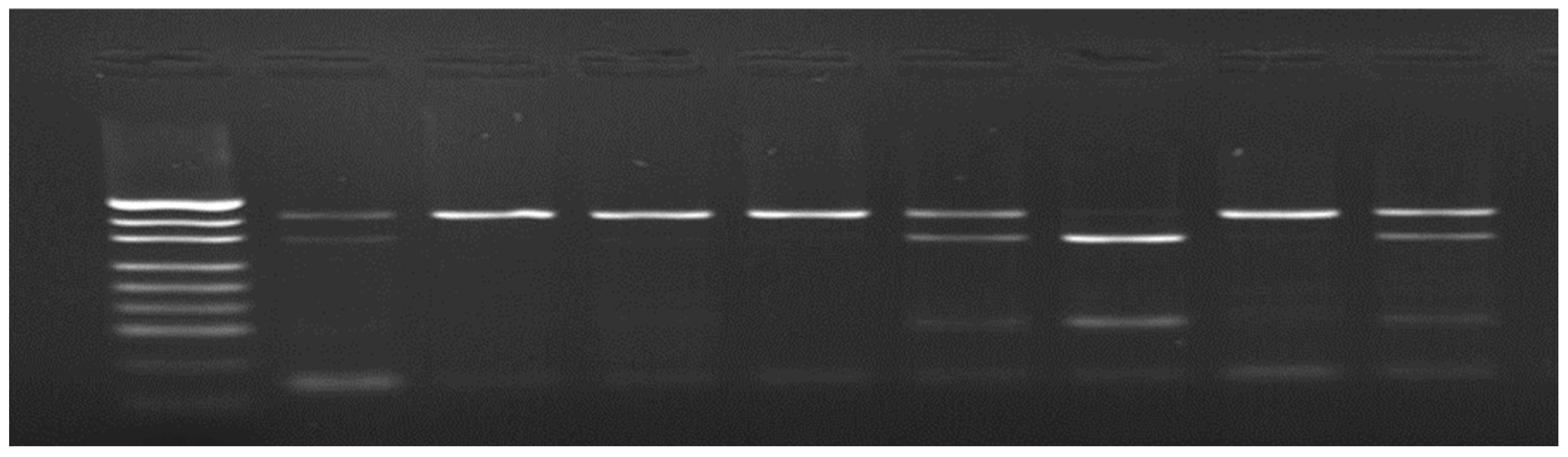
Figure 3.
Results of rs80860411A>C genotyping by PCR-RFLP method; from left: Gene Ruler 100 bp Plus DNA Ladder DNA Marker (Thermo Fisher Scientific, Waltham, MA, USA); lanes 2, 4, 5, 7, 8, 10, 11, 14, and 15 – CC genotype, lanes 3, 6, 9, 12, 13, and 16 – AC genotype, lanes 17, 18 – AA genotype.
Figure 3.
Results of rs80860411A>C genotyping by PCR-RFLP method; from left: Gene Ruler 100 bp Plus DNA Ladder DNA Marker (Thermo Fisher Scientific, Waltham, MA, USA); lanes 2, 4, 5, 7, 8, 10, 11, 14, and 15 – CC genotype, lanes 3, 6, 9, 12, 13, and 16 – AC genotype, lanes 17, 18 – AA genotype.
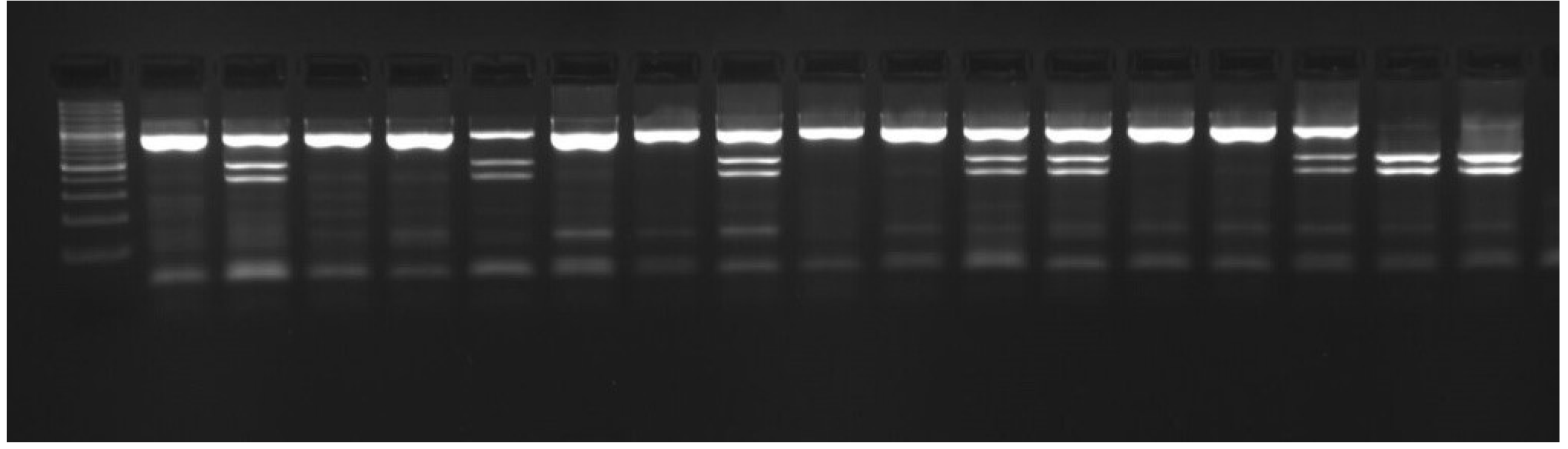

Table 1.
Primer sequences and restriction fragment lengths (A/C, rs80860411).
| Breed | Polish Large White Polish Landrace |
Pulawska |
| Forward Reverse |
5’-GTGCCTTCCTAGTGTCCCTT-3’ 5’-TCATGGACCACACACTCTAACA -3’ |
5’- CCACCCCAGACCTCTTGAAT-3’ 5’- GACTCTAGACTGAAGGCCCC-3’ |
| Amplicon length (bp) | 424 | 842 |
| Restriction fragments lengths (bp) |
AA – 316 and 108 AC – 424, 316 and 108 CC – 424 (no digestion) |
AA – 474 and 368 AC – 842, 474 and 368 CC – 842 (no digestion) |
Table 2.
Genotype and allele frequencies of rs80860411A>C SNP and calculation of Hardy-Weinberg equilibrium.
Table 2.
Genotype and allele frequencies of rs80860411A>C SNP and calculation of Hardy-Weinberg equilibrium.
| Breed | N | Genotype | Allele | HWE | ||||
|---|---|---|---|---|---|---|---|---|
| AA | AC | CC | A | C | ꭓ2 | p | ||
| Polish Landrace | 192 | 0.27 (n = 52) |
0.73 (n = 140) |
- | 0.36 | 0.64 | 63.2083 | ≤0.0001 |
| Polish Large White | 187 | 0.04 (n = 7) |
0.42 (n = 79) |
0.54 (n = 101) |
0.25 | 0.75 | 3.1896 | 0.0741 |
| Pulawska | 48 | 0.12 (n = 6) |
0.48 (n = 23) |
0.40 (n = 19) |
0.36 | 0.64 | 0.0561 | 0.8127 |
n – number of individuals, HWE – Hardy-Weinberg equilibrium.
Table 3.
Association between gene polymorphism and fattening performance traits in the analyzed breeds (LSM ± SE, least squares mean ± standard error).
Table 3.
Association between gene polymorphism and fattening performance traits in the analyzed breeds (LSM ± SE, least squares mean ± standard error).
| Trait | Genotype | Polish Large White | Polish Landrace | Pulawska | Whole population |
|---|---|---|---|---|---|
| Number of days on test [days] |
AA AC CC |
79.354 ± 5.39 83.738 ± 3.65 82.628 ± 3.72 |
77.804 ± 1.54 78.660 ± 1.15 - |
100.333 ± 3.40A 85.217 ± 1.74B 88.105 ± 1.91B |
82.599 ± 1.68 81.960 ± 1.17 81.108 ± 1.39 |
| Daily feed intake [kg] |
AA AC CC |
2.396 ± 0.15 2.319 ± 0.10 2.365 ± 0.10 |
2.536 ± 0.04 2.477 ± 0.03 - |
2.138 ± 0.06 2.277 ± 0.03 2.244 ± 0.03 |
2.438 ± 0.04 2.398 ± 0.03 2.439 ± 0.04 |
| Lifetime daily gain [g/day] |
AA AC CC |
592.653 ± 36.73 566.227 ± 24.86 574.518 ± 25.33 |
603.930 ± 8.46 595.056 ± 6.29 - |
498.833 ± 23.20a 569.130 ± 11.85b 535.684 ± 13.04b |
577.517 ± 10.58 576.256 ± 7.34 579.110 ± 8.77 |
| Test daily gain [g/day] |
AA AC CC |
883.968 ± 58.63 858.058 ± 39.69 866.307 ± 40.44 |
924.189 ± 16.93 913.801 ± 12.60 - |
721.167 ± 27.90Aa 829.30 ± 14.25B 805.158 ± 15.64b |
873.856 ± 17.88 877.083 ± 12.41 884.28 ± 14.82 |
| Feed conversion [kg/kg gain] |
AA AC CC |
2.708 ± 0.11 2.710 ± 0.08 2.737 ± 0.08 |
2.762 ± 0.03 2.724 ± 0.02 - |
2.988 ± 0.07a 2.750 ± 0.04b 2.798 ± 0.04a |
2.811 ± 0.03 2.749 ± 0.02 2.774 ± 0.03 |
| Age at slaughter [days] |
AA AC CC |
169.980 ± 11.07 180.444 ± 7.49 176.930 ± 7.64 |
168.592 ± 2.55 170.854 ± 1.90 - |
204.833 ± 7.98a 178.304 ± 4.08b 189.105 ± 4.48b |
177.185 ± 3.25 177.056 ± 2.26 175.537 ± 2.69 |
Values marked with different letters indicate statistically significant differences: p ≤ 0.05 (a, b) or p ≤ 0.01 (A, B).
Table 4.
Association between rs80860411 polymorphism and slaughter traits, taking into account the right half-carcass, in the analyzed breeds (LSM ± SE, least squares mean ± standard error).
Table 4.
Association between rs80860411 polymorphism and slaughter traits, taking into account the right half-carcass, in the analyzed breeds (LSM ± SE, least squares mean ± standard error).
| Trait | Genotype | Polish Large White | Polish Landrace | Pulawska | Whole population |
|---|---|---|---|---|---|
| Carcass yield [%] |
AA AC CC |
75.973 ± 0.40 76.031 ± 0.27 76.097 ± 0.28 |
76.885 ± 0.13 76.861 ± 0.10 - |
76.230 ± 0.30 76.420 ± 0.15 76.393 ± 0.17 |
76.479 ± 0.13 76.450 ± 0.09 76.516 ± 0.11 |
| Middle length of carcass [cm] |
AA AC CC |
77.603 ± 1.04 78.764 ± 0.70 78.491 ± 0.71 |
79.209 ± 0.74 79.665 ± 0.55 - |
78.625 ± 0.85 78.452 ± 0.44 78.203 ± 0.48 |
79.133 ± 0.57 79.464 ± 0.39 79.220 ± 0.47 |
| Loin weight [kg] |
AA AC CC |
7.439 ± 0.27 7.327 ± 0.18 7.355 ± 0.19 |
7.765 ± 0.09 7.648 ± 0.07 - |
7.272 ± 0.23 7.700 ± 0.12 7.545 ± 0.13 |
7.681 ± 0.09 7.611 ± 0.06 7.617 ± 0.08 |
| Loin weight without skin and backfat [kg] |
AA AC CC |
5.882 ± 0.26 5.865 ± 0.17 5.855 ± 0.18 |
6.121 ± 0.08 6.041 ± 0.06 - |
5.736 ± 0.20 6.079 ± 0.10 5.840 ± 0.11 |
6.019 ± 0.09 5.987 ± 0.06 5.939 ± 0.07 |
| Ham weight without skin and backfat [kg] |
AA AC CC |
9.538 ± 0.26 9.294 ± 0.17 9.310 ± 0.18 |
9.272 ± 0.09 9.302 ± 0.07 - |
9.184 ± 0.22 9.214 ± 0.11 8.823 ± 0.12 |
9.223 ± 0.09 9.201 ± 0.06 9.119 ± 0.07 |
| Loin eye area [cm2] |
AA AC CC |
53.631 ± 2.35 51.196 ± 1.58 51.518 ± 1.61 |
49.993 ± 0.70a 51.661 ± 0.52b - |
52.217 ± 2.36a 53.972 ± 1.20a 49.587 ± 1.32b |
51.042 ± 0.78 51.971 ± 0.54 51.114 ± 0.65 |
| Width of loin eye [cm] |
AA AC CC |
10.188 ± 0.35 10.195 ± 0.23 10.295 ± 0.24 |
10.091 ± 0.11 10.307 ± 0.08 - |
10.431 ± 0.32A 10.554 ± 0.16A 9.778 ± 0.18B |
10.239 ± 0.12 10.357 ± 0.08 10.262 ± 0.10 |
| Height of loin eye [cm] |
AA AC CC |
6.741 ± 0.28 6.562 ± 0.18 6.608 ± 0.19 |
6.894 ± 0.09 6.851 ± 0.07 - |
6.977 ± 0.25 6.924 ± 0.13 6.772 ± 0.14 |
6.898 ± 0.09 6.834 ± 0.06 6.837 ± 0.08 |
| Average backfat thickness of five measurements [cm] |
AA AC CC |
1.123 ± 0.14 1.122 ± 0.09 1.177 ± 0.10 |
1.341 ± 0.04 1.292 ± 0.03 - |
1.295 ± 0.13 1.371 ± 0.07 1.527 ± 0.07 |
1.339 ± 0.05 1.311 ± 0.03 1.396 ± 0.04 |
| Carcass meat content [%] |
AA AC CC |
63.155 ± 1.35 61.939 ± 0.90 62.117 ± 0.92 |
61.766 ± 0.44 61.890 ± 0.33 - |
61.339 ± 1.18A 62.276 ± 0.60A 59.254 ± 0.66B |
61.798 ± 0.46 61.687 ± 0.32 61.155 ± 0.38 |
| Weight of primary cuts [kg] |
AA AC CC |
24.261 ± 0.53 23.797 ± 0.35 23.859 ± 0.36 |
23.995 ± 0.17 24.049 ± 0.13 - |
23.621 ± 0.45A 23.993 ± 0.23A 22.838 ± 0.25B |
23.862 ± 0.18 23.828 ± 0.12 23.618 ± 0.15 |
Values marked with different letters indicate statistically significant differences: p ≤ 0.05 (a, b) or p ≤ 0.01 (A, B).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



