Submitted:
12 October 2024
Posted:
14 October 2024
You are already at the latest version
Abstract
Russula, a prominent genus of ectomycorrhizal fungi, is notably abundant and diverse in China. We present here the findings from studies on various Chinese Russula collections. Two notable species within subg. Heterophyllinae, namely Russula leucoviridis and R. subswatica, were described and illustrated based on comprehensive morphological characteristics and molecular evidence. Morphologically, Russula leucoviridis is characterized by its small to medium-sized basidiomata with a cuticle that cracks and breaks into small green patches near the center after mature, a pastel green to green pileus center with light yellow spots, and a white to greenish white pileus margin; whereas R. subswatica is distinguished by an infundibuliform pileus with a deeply depressed center after mature, a light orange to grayish orange pileus center, and a yellowish white to light orange margin with a purplish to purplish-black hue, and a white to yellowish white stipe that exhibits a purplish hue at the junction with the pileus. The phylogenetic analyses were constructed using a combined dataset of ITS, nrLSU, RPB2 and mtSSU. Relying on both morphological characteristics and multigene phylogeny, the former species is classified within subsect. Virescentinae, while the latter is affiliated with subsect. Griseinae. In this study, we contribute new data on infrageneric phylogenetic relations and enrich the species diversity of Subg. Heterophyllinae in China.
Keywords:
Basidiomycota
; ectomycorrhizal fungi
; Russulaceae
; new species
; China
1. Introduction
The genus Russula Pers., a type genus of the family Russulaceae, is recognized as the largest genus within the phylum Basidiomycota [1,2]. It comprises approximately 1,100 described species, with estimates suggesting the existence of at least 3,000 species [3,4]. Species of this genus typically form ectomycorrhizal symbiotic associations with a wide range of host trees primarily including Pinus spp., Castanopsis spp., Picea spp., Larix spp., Fagus spp. and Castanea spp., as well as with various herbs, sedges, and shrubs [5,6,7]. These fungi inhabit a diverse array of ecosystems, ranging from Arctic tundra to tropical forests, and play a crucial role in forest development and maintaining ecological balance [8,9]. Numerous species of Russula are renowned as wild edible mushrooms in various regions around the world and are commonly traded commercially [9,10,11]. Investigations and statistics on the resource diversity of Chinese macrofungi indicate that approximately 78 species of Russula are edible in China, with the Russula griseocarnosa complex and the R. virescens complex being particularly popular [10,12,13]. However, the genus Russula also includes several toxic species, making precise identification essential for the safe utilization of these fungi. The rapid advancements in molecular biology techniques have greatly advanced the study of the systematics and phylogeny of Russula. Recent molecular phylogenetic studies have indicated that the taxa within the genus Russula are divided into nine subgenera, namely: R. subg. Glutinosae Buyck & X.H. Wang, R. subg. Archaeae Buyck & V. Hofst., R. subg. Compactae (Fr.) Bon, R. subg. Brevipedum Buyck & V. Hofst., R. subg. Crassotunicatae Buyck & V. Hofst., R. subg. Heterophyllidiae Romagnesi, R. subg. Cremeoochraceae Buyck & X.H. Wang, R. subg. Malodorae Buyck & V. Hofst., and R. subg. Russula Per. [14,15,16]. The infrageneric classification system in this study followed Buyck et al. [14,15,16].
Subgenus Heterophyllidia is one of the most complex and diverse groups within the genus Russula. The subgenus is mainly characterized by the mostly medium to large basidiomata, rarely very small basidiomata, almost all possible pileus colors, equal lamellae, mild to strongly acrid taste, mostly white or cream basidiospore print, basidiospores with inamyloid or partly amyloid suprahilar spot, mostly abundant gloeocystidia that are typically mucronate to obtuse-rounded, and pileipellis without primordial hyphae [14]. In China, numerous new species of subg. Heterophyllidia have been continuously described and reported in recent studies [17,18,19,20,21,22], underscoring the region as a hotspot for the discovery of previously unknown species. During surveys on the species diversity and geographic distribution of the subgenus Heterophyllidia, the first author of this paper discovered that many species, including the R. virescens complex, R. cyanoxantha complex, R. heterophylla complex, and several undescribed species, were being consumed and sold. Therefore, it is essential to clarify the species diversity within this subgenus to ensure the safe and sustainable consumption of these wild fungi. In this study, several new collections of subg. Heterophyllidia from China were examined using morphological characteristics and phylogenetic analyses, resulting in detailed descriptions and illustrations of two notable new species, along with an updated phylogenetic tree to confirm their distinct placements. This study significantly enhances our understanding of species diversity within the subg. Heterophyllidia in China.
2. Materials and Methods
2.1. Morphological Studies
Fresh basidiomata were collected and photographed in their natural forest habitat. All specimens were dried at temperatures ranging from 45 to 55 °C and subsequently deposited in the herbarium of the Research Institute of Tropical Forestry, Chinese Academy of Forestry (RITF). The terminology used to describe the morphological characteristics follows the guidelines established by Adamčík et al. [3]. Macroscopic characteristics were documented through meticulous notes and high-resolution photographs. Color codes primarily adhere to the standards set by Kornerup and Wanscher [23]. Microscopic examination of the specimen sections was carried out using a ZEISS Imager M2 microscope. Estimates of spore ornamentation density were conducted in accordance with the methodology described by Adamčík and Marhold [24]. The estimates of hymenial cystidia density were performed referring to Buyck [25]. The basidiospores were observed and measured in Melzer’s reagent from a lateral perspective, with ornamentation excluded. Following pretreatment of dried specimens with 5% potassium hydroxide (KOH), additional micromorphological characteristics were identified and measured in Congo red. The coloration of the cystidia contents was observed in a sulfovanillin (SV) solution [26]. The pileipellis were examined using cresyl blue to verify the presence of ortho- or metachromatic reactions [27]. The structure and ornamentation of the basidiospores were illustrated using a JEOL JSM-6510 scanning electron microscope (SEM). Basidiospore measurements are presented in the format (Min–)AV-SD–AV–AV+SD(–Max), where Min denotes the minimum value, Max denotes the maximum value, AV represents the average value, SD stands for the standard deviation, and Q indicates the length-to-width ratio of the basidiospores.
2.2. Molecular Study
Total genomic DNA was extracted from dry specimens using an optimized CTAB protocol [28]. The following four regions were amplified and sequenced using standard primer sets for enhanced accuracy and fluency: 600 base pairs of the ITS region of rDNA with the primers ITS1 and ITS4 [29]; 900 base pairs of the nuclear ribosome large subunit (nrLSU) with the primers LR0R and LR5 [30]; 700 base pairs of the second largest subunit of RNA polymerase II (RPB2) using the primers bRPB2-6f and fRPB2-7cr [31,32]; 600 base pairs of the ribosomal mitochondrial small subunit (mtSSU) using primers MS1 and MS2 [29]. The successful PCR products were sequenced using an ABI 3730 DNA analyzer with the ABI BigDye 3.1 terminator cycle sequencing kit (Shanghai Sangon Biological Engineering Technology and Services Co., Ltd, Shanghai, China). The newly generated sequences have been submitted to the GenBank database.
2.3. Phylogenetic Analyses
For phylogenetic analyses, species from the subgenus Heterophyllidia that exhibit high similarity to our new species were selected, along with representative species closely related to subsect. Griseinae Jul. Schäff. and subsect. Virescentinae Singer. Russula maguanensis J. Wang, X.H. Wang, Buyck & T. Bau and R. substriata J. Wang, X.H. Wang, Buyck & T. Bau were utilized as the outgroup. The NCBI accession numbers for the sequences employed in the phylogenetic tree are detailed in Table 1. Initial sequence alignment was conducted using the online version MAFFT 7.0 (http://mafft.cbrc.jp/alignment/server/). To ensure reliable and accurate results, manual evaluations and adjustments were made in BioEdit as necessary [33]. Maximum likelihood (ML) and Bayesian analysis (BA) methods were employed for conducting the phylogenetic analyses. Maximum likelihood (ML) analysis was performed using RAxML-HPC2 on XSEDE (8.2.12) via the CIPRES Science Gateway (www.phylo.org). The ML analysis utilized the rapid bootstrap algorithm with 1000 replicates to ensure the robustness of the results under the GAMMA model. Bootstrap support (BS) values of 70% or higher on the final tree were considered significant. Bayesian analysis (BA) was conducted using MrBayes 3.2.7a on XSEDE through the CIPRES Science Gateway (www.phylo.org) under the GTR model. A total of 50 million generations were executed across four independent Markov chains, with tree sampling occurring every 100 generations. For each analysis, the initial 25% of the trees were excluded as part of the burn-in phase. The 50% majority-rule consensus trees were used to obtain Bayesian posterior probability (PP) values, and nodes with PP values of 0.95 or higher were deemed to have significant support.
3. Results
3.1. Phylogeny
Both the Maximum Likelihood (ML) and Bayesian Analysis (BA) of the combined ITS, nrLSU, RPB2 and mtSSU sequence dataset produced consistent tree topologies, with only the ML tree depicted in Figure 1. Posterior probabilities from the Bayesian analysis (BA) are also indicated along the branches. The samples of our two new species each formed a strongly supported clade, nested within subsect. Virescentinae and Griseinae. They are distinctly separate from any known and sequenced species within subg. Heterophyllidia. Phylogenetic analyses also confirmed that both subsections Virescentinae and Griseinae are monophyletic groups, each with robust support indicated by bootstrap values (BS) of 100% and posterior probabilities (PP) of 1, while the monophyly of the remaining four subsections within subgenus Heterophyllidia was also significantly supported.
Russula leucoviridis is phylogenetically positioned as the sister taxon to a clade consisting of two Chinese species R. prasina G.J. Li & R.L. Zhao and R. xanthovirens Y. Song & L.H. Qiu, with 100% bootstrap support and 1.00 posterior probabilities. Our second species, R. subswatica clustered with Pakistani species R. swatica Sarwar & Hanif, demonstrating robust phylogenetic support with 100% bootstrap support and 1.00 posterior probabilities.
3.2. Taxonomy
MycoBank: MB855842
Diagnosis: Pileus small to medium-sized; cuticle adhering many small frosty particles near the center when young, cracking and broken into small green patches near the center after mature, patches crowded towards the center; pastel green to green towards the center with light yellow spots, white to greenish white near the margin.
Holotype: China, Hainan Province, Wuzhishan City, Shuiman Town, Wuzhi Mountain National Reserve;18°51′59.50″ N, 109°40′30.59″ E, 800m asl., in evergreen broad-leaved forest; 23 June 2016, leg. JXM105 (RITF3405).
Etymology: Leuco (Latin) = white; viridis (Latin) = green; the species name refers to white pileus margin and green center.
Description: Pileus small to medium-sized, 30–55 mm in diameter; initially hemispheric when young, expanding to planeconvex with a depressed center after mature; margin incurved, not cracked, striation up to the 1/3 of the radius; cuticle dry, glabrous, peeling to 1/4 of the radius, many small adhering frosty particles near the center when young, cracking and broken into small green patches near the center after mature, patches crowded towards the center; pastel green (27A4) to green (27A7) towards the center with light yellow (3A5) spots, white (1A1) to greenish white (28A2) near the margin. Lamellae adnate to almost free, 3–5 mm deep, 8–10 at 1 cm near the pileus margin, white (1A1) to cream; lamellulae sometimes present and irregular in length; furcations occasional near the stipe; edge entire and concolor. Stipe 20–55 × 7–16 mm, cylindrical, slightly inflated towards the base, white (1A1) to almond, medulla initially stuffed when young, becoming hollow after mature. Context 2–3 mm thick in half of the pileus radius, white (1A1), unchanging when bruised, taste mild, odor inconspicuous. Spore print white (1A1) to cream.
Basidiospores (5.7–)6.0–6.9–7.8(–8.8) × (4.8–)5.3–5.9–6.5(–7.0) μm, Q = 1.03–1.15–1.27(–1.36), subglobose to broadly ellipsoid; ornamentation of medium-sized, moderately distant [5–6(–8) in a 3 μm diameter circle] amyloid warts or spines, 0.3–0.5 μm high, locally reticulate, occasionally fused in short chains [0–2(–3) in the circle], occasionally to frequently connected by line connections [2–3(–4) in the circle]; suprahilar spot small, amyloid. Basidia (35.0–)39.0–46.0–53.0(–56.2) × (8.4–)9.3–10.3–11.4 μm, mostly 4-spored, sometimes 2- and 3-spored, clavate; basidiola subcylindrical or clavate, ca. 5.5–9.5 μm wide. Hymenial gloeocystidia on lamellae sides dispersed to moderately numerous, (46.0)48.6–57.7–66.8(76.0) × (7.4–)8.4–10.1–11.9(12.7) μm, typically fusiform, occasionally clavate, apically mainly obtuse, occasionally acute, sometimes with 4–8 μm long appendage, thin-walled; contents granulose or heteromorphous, turning reddish black in SV. Hymenial gloeocystidia on lamellae edges often narrower, (38.7–)44.8–55.7–66.5(–75.2) × (5.6–)6.3–7.8–9.2(–9.8) μm, typically clavate, sometimes fusiform, apically mainly obtuse, rarely mucronate, sometimes with 4–8 μm long appendage, thin-walled; contents heteromorphous, turning reddish black in SV. Marginal cells (16.7–)18.4–22.8–27.2(–30.6) × (3.2–)3.8–4.5–5.1 μm, clavate or lageniform, sometimes flexuous. Pileipellis orthochromatic in cresyl blue, not sharply delimited from the underlying context, 240–320 μm deep, two-layered; suprapellis 180–200 μm deep, composed of loose, ascending or erect hyphal terminations; subpellis 80–140 μm deep, composed of intricate, dense, 3–6 μm wide hyphae. Hyphal terminations near the pileus margin occasionally unbranched, sometimes flexuous, thin-walled; terminal cells (10.8–)13.2–22.3–31.4(–37.4) × (4.0–)4.5–5.0–5.7(–6.2) μm, mainly narrowly lageniform, occasionally cylindrical, apically attenuated or constricted, sometimes obtuse; subterminal cells often shorter and wider, ca. 5–8 μm wide, rarely branched. Hyphal terminations near the pileus center similar; terminal cells (8.0–)10.0–14.1–18.2(–25.0) × (2.6–)3.1–3.7–4.3(–4.7) μm, mainly lageniform or subcylindrical, apically attenuated or constricted; subterminal cells often shorter and wider, rarely branched, ca. 4–6 μm wide. Pileocystidia near the pileus margin always one-celled, (30.9–)32.3–37.5–42.8(–47.5) × (2.8–)3.8–5.0–6.2(–6.8) μm, mainly clavate, occasionally fusiform or lanceolate, apically typically obtuse, occasionally acute, sometimes with 2–5 μm long, round or ellipsoid appendage, thin-walled; contents granulose or heteromorphous, turning reddish black in SV. Pileocystidia near the pileus center similar, always one-celled, (29.6–)31.4–36.5–41.6(–44.6) × (3.3–)3.7–4.4–5.2(–5.4) μm, thin-walled, mainly clavate or cylindrical, apically obtuse, contents granulose or heteromorphous, turning reddish black in SV.
Additional specimens examined: China, Hainan Province, Wuzhishan City, Shuiman Town, Wuzhi Mountain National Reserve; 18°51′59.50″ N, 109°40′30.59″ E, 800m asl., in evergreen broad-leaved forest; 23 June 2016, leg. JXM106 (RITF3406).
Notes: Russula leucoviridis is positioned within the subsection Virescentinae based on a combination of its morphological characteristics and phylogenetic analysis. In phylogenetic analysis, R. leucoviridis is the sister taxon to a clade consisting of R. prasina and R. xanthovirens, both originating from China, with strong support evidenced by 100% bootstrap values and 1.00 posterior probability. However, R. prasina often has the larger basidiomata (43–130 mm), a grass green pileus center and pale yellowish green margin, pale ocher lamellae and ochraceous yellow stipe base [34]; R. xanthovirens differs from R. leucoviridis in having the larger basidiomata (50–75 mm), a yellowish green to deep green pileus center with cracked yellowish margin when age, whitish lamellae with reddish hue, white stipe with green tinge, often shorter and wider basidia [(29–)30–51.5(–60) × (9.5–)10–9.5–14.5(–15) μm] and often longer pileocystidia (40–69.5 × 4–9 μm) [35].
Given cracking pileus surface, R. virescens (Schaeff.) Fr., R. parvovirescens Buyck, D. Mitch. & Parrent and R. viridirubrolimbata J. Z. Ying and R. crustosa Peck of subsect. Virescentinae resemble R. leucoviridis. However, R. virescens, originally described from Europe, can be distinguished by usually larger basidiomata [40–90(125) mm], a green to yellowish green pileus and shorter basidia (40–52 × 6–9.2 μm) [36]. The American species R. parvovirescens has a greenish brown to metallic bluish green pileus with green patches, often larger basidiospores (6.7–9.1 × 5.7–7.2 μm) and smaller Basidia (38–45 × 8–9 mm) [37]. R. viridirubrolimbata, (originally reported from China) is distinct in a light yellowish olive to yellowish olive pileus center with a pinkish red to light jasper red margin, smaller basidia (31−43 × 7−10μm), larger hymenial gloeocystidia on lamellae sides (78–107 × 8–10 μm) and absence of hymenial gloeocystidia on lamellae edges [13,38]. Russula crustosa, initially described in North America, possesses a brownish-yellow, greenish or subolivaceous pileus, shorter basidia [(29–)30–32–33.5(–35) × (7.5–)8–9.5–10.5(–11) μm], longer hymenial gloeocystidia on lamellae sides [(59–)63–69–75(–80) × (7–)8–9.5–11(–13) μm] and absence of hymenial gloeocystidia on the lamellar edges [39].
MycoBank: MB855843
Diagnosis: Pileus medium-sized to large; initially flat hemispheric when young, expanding and infundibuliform with a deeply depressed center after mature; light orange to grayish orange near the center, yellowish white to light orange towards the margin, partially with a purplish to purplish-black hue. Stipe white to yellowish white, exhibiting a purplish hue at the junction with the pileus.
Holotype: China. Xizang Autonomous Region, Nyingchi City, Motuo County; 29°22′13.43″ N, 95°26′58.75″ E, 900m asl., in mixed forest dominated by Quercus aquifolioides Rehder & E. H. Wilson; 15 July 2014, leg. T25530 (RITF2994).
Etymology: The species name refers to its morphological resemblance to Russula swatica.
Discription: Pileus medium-sized to large, 55–70 mm in diameter; initially flat hemispheric when young, soon expanding and infundibuliform with a deeply depressed center after mature; margin incurved, sometimes cracked after mature, striation short and inconspicuous; cuticle smooth, dry, glabrous, peeling to 1/3 of the radius; light orange (5A4) to grayish orange (6B5) near the center, yellowish white (2A2) to light orange (5A4) towards the margin, partially with a purplish to purplish-black hue. Lamellae deccurent, 2–4 mm deep, 8–11 at 1 cm near the pileus margin, white (1A1) to cream; lamellulae present and irregular in length; furcations occasional near the stipe; edge entire and concolor. Stipe 60–70 × 12–15 mm, cylindrical, slightly inflated towards the base, white (1A1) to yellowish white (2A2), exhibiting a purplish hue at the junction with the pileus, medulla initially stuffed when young, becoming hollow after mature. Context 3–5 mm thick in half of the pileus radius, white (1A1), unchanging when bruised, taste mild, odor inconspicuous. Spore print white (1A1) to yellowish white (2A2).
Basidiospores (5.7–)6.0–7.0–8.0(–8.5) × (4.7–)5.1–6.2–7.2(–8.3) μm, Q = 1.03–1.15–1.27(–1.36), subglobose to broadly ellipsoid; ornamentation of medium-sized, dense [9–11(–12) in a 3 μm diameter circle] amyloid warts, 0.2–0.8 μm high, occasionally to frequently fused in short or long chains [2–3(–4) in the circle], occasionally connected by line connections [0–1(–2) in the circle]; suprahilar spot medium-sized, amyloid. Basidia (36.6–)39.8–44.0–48.0(–54.0) × (6.4–)8.5–9.6–10.7(–11.0) μm, mostly 4-spored, sometimes 2- and 3-spored, clavate; basidiola clavate or subcylindrical, ca. 6–9.5 μm wide. Hymenial gloeocystidia on lamellae sides dispersed to moderately numerous, (55.3)60.5–72.4–84.3(97.0) × (10.2)7.7–8.8–9.9(10.5) μm, typically clavate or fusiform, occasionally lanceolate, apically mainly mucronate, occasionally obtuse, often with 4–10 μm long appendage, thin-walled; contents heteromorphous or granulose, turning yellowish brown in SV. Hymenial gloeocystidia on lamellae edges often shorter, (44.8–)52.4–68.3–84.2(–95.8) × (5.5–)7.8–9.6–11.4 μm, typically clavate or subcylindrical, occasionally fusiform, apically mainly obtuse, occasionally mucronate, usually with 3–6 μm long appendage thin-walled; contents heteromorphous, turning reddish black in SV. Marginal cells (15.8–)19.6–24.2–28.8(–35.5) × (3.6–)3.8–5.3–6.7(–9.0) μm, subcylindrical or clavate, sometimes flexuous. Pileipellis orthochromatic in cresyl blue, not sharply delimited from the underlying context, 200–260 μm deep, two-layered; suprapellis 140–180 μm deep, composed of loose, ascending or erect hyphal terminations; subpellis 80–100 μm deep, composed of relatively dense, intricate, 3–5 μm wide hyphae. Hyphal terminations near the pileus margin rarely unbranched, occasionally flexuous, thin-walled; terminal cells (9.2–)12.4–19.3–26.3(–35.2) × (4.3–)4.9–5.6–6.3(–7.0) μm, typically lageniform, occasionally clavate or subcylindrical, apically attenuated or constricted, sometimes obtuse; subterminal cells frequently shorter and wider, ca. 5–7 μm wide, typically unbranched. Hyphal terminations near the pileus center similar; terminal cells (13.7–)16.8–20.7–25.3(–27.7) × (3.7–)4–4.5–5(–6.7) μm, mainly lageniform or subcylindrical, apically attenuated or constricted; subterminal cells often shorter and wider, unbranched, ca. 4–6.5 μm wide. Pileocystidia near the pileus margin always one-celled, (45.5–)49.4–59.5–69.6(–77.4) × (5.8–)6.0–6.8–7.6(–10.0) μm, mainly clavate, occasionally subcylindrical, apically obtuse, occasionally mucronate, sometimes with 3–5 μm long, round or ellipsoid appendage, thin-walled; contents heteromorphous, turning yellowish brown in SV. Pileocystidia near the pileus center often shorter and narrower, always one-celled, (38.3–)42.6–52.6–62.5(–70.8) × (4.8–)5.5–6.3–7.0 μm, thin-walled, mainly clavate or fusiform, apically obtuse or mucronate, sometimes with 3–6 μm long appendage, contents heteromorphous or granulose, turning r yellowish brown in SV.
Additional specimens examined: China, Xizang Autonomous Region, Nyingchi City, Motuo County; 29°22′13.43″ N, 95°26′58.75″ E, 900m asl., in mixed forest dominated by Quercus aquifolioides Rehder & E. H. Wilson; 15 July, 2014, leg. T25531 (RITF2995).
Notes: The integration of morphological characteristics and phylogenetic analysis positions R. subswatica within the subsection Griseinae. In the phylogenetic tree, R. subswatica is clustered with Pakistani species R. swatica, exhibiting strong phylogenetic support with 100% bootstrap support and 1.00 posterior probabilities. However, R. swatica is distinguished from R. subswatica by its light brown to gray pileus, narrower basidiospores (6–8 × 4–6 μm), and often shorter and wider hymenial gloeocystidia (59–70 × 10–15 μm) [40].
Morphologically, R. subswatica exhibits certain similarities with R. medullata Romagn., and R. shawarensis Kiran & Khalid, both of which belong to subsect. Griseinae. However, R. medullata, originally report from Europe, can be distinguished by a lilac, flesh-pink, pinkish brown, and pinkish grey pileus, ochraceous lamellae after mature and ochre spore print [36,41]. The Pakistani species R. shawarensis has a light pinkish brown to grey buff pileus, very rare to absent lamellulae, shorter basidia [(31.5–)34–39.5(–41) × (7–)9–10.5(–11) μm], often larger hymenial gloeocystidia on lamellae sides [(66–)72–92.5(–105) × (9.5–)10–13(–14) μm], longer the terminal cells of the hyphae near both the pileus margin (av. 48.2 × 3.8 μm) and the pileus center (av. 41.5 × 4.2 μm), and longer pileocystidia near the pileus center (av. 58 × 7 μm) [42].
4. Discussion
In this study, we have described and named two new species of Russula subg. Heterophyllinae: R. leucoviridis and R. subswatica. The identification and characterization of these species enhance our understanding of the biodiversity and phylogenetic relationships within the subg. Heterophyllinae in China. These findings highlight the richness and diversity of ectomycorrhizal fungi in the region, significantly contributing to both taxonomic and ecological studies. Russula leucoviridis is notable for its small to medium-sized basidiomata with a cuticle that cracks and breaks into small green patches near the center after mature, a pastel green to green pileus center with light yellow spots, and a white to greenish white pileus margin. In contrast, R. subswatica is characterized by its infundibuliform pileus, a light orange to grayish orange pileus center, and a yellowish white to light orange margin with a purplish to purplish-black hue, and a white to yellowish white stipe that exhibits a purplish hue at the junction with the pileus. These unique morphological characteristics are crucial for differentiating these new species from closely related ones, and their distinctiveness is further confirmed by multi-gene phylogenetic analysis. Phylogenetic studies based on ITS, nrLSU, RPB2, and mtSSU regions, offer robust support for the classification of these species. The assignment of R. leucoviridis to subsection Virescentinae and R. subswatica to subsection Griseinae suggests distinct evolutionary pathways and adaptation strategies within the subg. Heterophyllinae. In conclusion, the combination of molecular evidence and morphological characteristics provides strong support for the recognition and delineation of these new species. The addition of R. leucoviridis and R. subswatica not only enriches the taxonomic diversity of the subgenus but also highlights the ongoing need for meticulous taxonomic research to uncover the hidden diversity of fungi.
Author Contributions
Conceptualization, B.C. and J.L.; methodology, B.C.; software, B.C. and F.Y.; validation, B.C. and J.L.; formal analysis, B.C.; investigation, B.C. and F.Y.; resources, J.L.; data curation, J.L.; writing—original draft preparation, B.C.; writing—review and editing, B.C. and J.L.; visualization, B.C.; supervision, J.L.; project administration, B.C., J.L. and F.Y.; funding acquisition, B.C., J.L. and F.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (No. 32300010, 32370018 and 32101499) and the Guangzhou Science and Technology Plan Project (2024A04J3421).
Data Availability Statement
All data obtained and analyzed in this study have been included in
this article.
Acknowledgments
We are grateful to Qi Zhao (Chinese Academy of Sciences) and Xumeng Jiang (Northwest A&F University) for providing the samples.
Conflicts of Interest
The authors declare no conflict of interests.
References
- Buyck, B.; Zoller, S.; Hofstetter, V. Walking the thin line… ten years later: the dilemma of above- versus below-ground features to support phylogenies in the Russulaceae (Basidiomycota). Fungal Divers. 2018, 89, 267–292. [Google Scholar] [CrossRef]
- He, MQ.; Cao, B.; Liu, F.; Boekhout, T.; Denchev, T.T.; Schoutteten, N.; Denchev, C.M.; Kemler, M.; Gorjón, S.P.; et al. Phylogenomics, divergence times and notes of orders in Basidiomycota. Fungal Divers. 2024, 89, 127–406. [Google Scholar]
- Adamčík, S.; Looney, B.; Caboň, M.; Jančovičová, S.; Adamčíková, K.; Avis, P.G.; Barajas, M.; Bhatt, R.P.; Corrales, A.; Das, K.; Hampe, F.; Ghosh, A.; et al. The quest for a globally comprehensible Russula language. Fungal Divers. 2019, 99, 369–449. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; et al. Outline of Fungi and fungus-like taxa–2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Hackel, J.; Henkel, T.W.; Moreau, P.A.; De Crop, E.; Verbeken, A.; Sà, M.; Buyck, B.; Neves, M.A.; Vasco-Palacios, A.; Wartchow, F.; Schimann, H.; Carriconde, F.; et al. Biogeographic history of a large clade of ectomycorrhizal fungi, the Russulaceae, in the Neotropics and adjacent regions. New Phytol. 2022, 236, 698–713. [Google Scholar] [CrossRef] [PubMed]
- Looney, B.; Miyauchi, S.; Morin, E.; Drula, E.; Courty, P.E.; Kohler, A.; Kuo, A.; Labutti, K.; Pangilinan, J.; Lipzen, A.; et al. Evolutionary transition to the ectomycorrhizal habit in the genomes of a hyperdiverse lineage of mushroom-forming fungi. New Phytol. 2022, 233, 2294–2309. [Google Scholar] [CrossRef]
- Noffsinger, C.R.; Adamčikova, K.; Eberhardt, U.; Caboň, M.; Bazzicalupo, A.; Buyck, B.; Kaufmann, H.; Weholt, Ø.; Looney, B.P.; Matheny, P.B.; Berbee, M.L.; Tausan, D.; Adamčik, S. Three new species in Russula subsection Xerampelinae supported by genealogical and phenotypic coherence. Mycologia 2024, 116, 322–349. [Google Scholar] [CrossRef]
- Knudsen, H.; Borgen, T. Russulaceae in Greenland. In Arctic and Alpine Mycology 1; University of Washington Press: Seattle, WA, USA, 1982; pp. 1–559. [Google Scholar]
- Looney, B.P.; Meidl, P.; Piatek, M.J.; Miettinen, O.; Martin, F.M.; Matheny, P.B.; Labbé, J.L. Russulaceae: a new genomic dataset to study ecosystem function and evolutionary diversification of ectomycorrhizal fungi with their tree associates. New Phytol. 2018, 218, 54–65. [Google Scholar] [CrossRef]
- Wang, X.H. Taxonomic comments on edible species of Russulaceae. Mycosystema 2020, 39, 1617–1639. [Google Scholar]
- Huang, L.H.; Wang, J.; Li, S.H.; Qin, W.Q.; Li, H.J.; Wang, X.H. Clarifying the identity of marketed edible Huotanjun (Burnt Charcoal Mushroom) in southern China. Mycol. Prog. 2023, 22, 80. [Google Scholar] [CrossRef]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource diversity of Chinese macrofungi: edible, medicinal and poisonous species. Fungal Diversi. 2019, 98, 1–76. [Google Scholar]
- Deng, C.Y.; Shi, L.Y.; Wang, J.; Xiang, Z.; Li, S.M.; Li, G.J.; Yang, H. Species diversity of Russula virescens complex “qingtoujun” in southern China. Mycosystema 2020, 39, 1–23. [Google Scholar]
- Buyck, B.; Zoller, S.; Hofstetter, V. Walking the thin line... ten years later: The dilemma of above- versus below-ground features to support phylogenies in the Russulaceae (Basidiomycota). Fungal Divers. 2018, 89, 267–292. [Google Scholar] [CrossRef]
- Buyck, B.; Wang, X.H.; Adamčíková, K.; Caboň, M.; Jančovičová, S.; Hofstetter, V.; Adamčík, S. One step closer to unravelling the origin of Russula: subgenus Glutinosae subg. nov. Mycosphere 2020, 11, 285–304. [Google Scholar] [CrossRef]
- Buyck, B.; Horak, E.; Cooper, J.A.; Wang, X.H. Russula subgen. Cremeoochraceae sub gen. nov.: A very small and ancient lineage sharing with Multifurca (Russulaceae) an identical, largely circum-Pacific distribution pattern. Fungal Syst. Evol. 2024, 14, 109–126. [Google Scholar]
- Li, G.J.; Li, S.M.; Buyck, B.; Zhao, S.Y.; Xie, X.J.; Shi, L.Y.; Deng, C.Y.; Meng, Q.F.; Sun, Q.B.; Yan, J.Q.; Wang, J.; Li, M. Three new Russula species in sect. Ingratae (Russulales, Basidiomycota) from southern China. MycoKeys 2021, 84, 103–139. [Google Scholar]
- Chen, B.; Song, J.; Chen, Y.L.; Zhang, J.H.; Liang, J.F. Morphological and phylogenetic evidence for two new species of Russula subg. Heterophyllidia from Guangdong Province of China. MycoKeys 2021, 82, 139–157. [Google Scholar] [PubMed]
- Song, Y. Species of Russula subgenus Heterophyllidiae (Russulaceae, Basidiomycota) from Dinghushan biosphere reserve. Eur. J. Taxon. 2022, 826, 1–32. [Google Scholar] [CrossRef]
- Han, Y.X.; Liang, Z.Q.; Zeng, N.K. Notes on four species of Russula subgenus Heterophyllidiae (Russulaceae, Russulales) from southern China. Front. Microbiol 2023, 14, 1140127. [Google Scholar] [CrossRef]
- Chen, Y.L.; Chen, B.; Liang, R.X.; Wang, S.K.; An, M.Y.; Zhang, J.H.; Liang, J.Y.; Wang, Y.X.; Gao, X.L.; Liang, J.F. Four new species of Russula subsect. Cyanoxanthinae from China (Russulales, Russulaceae). MycoKeys 2024, 107, 21–50. [Google Scholar]
- Chen, Y.L.; Liang, J.F.; Wang, S.K.; Chen, B.; Zhang, J.H.; An, M.Y.; Li, X.; Gao, X.L.; Chen, X.; Liao, J.P. Notes on two new species of Russula subsect. cyanoxanthinae (Russulaceae, Russulales) from southern china. Mycol. Prog 2024, 23, 52. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Taschenlexikon der Farben, 3rd ed.; Muster-Schmidt Verlag: Göttingen, Germany, 1981. [Google Scholar]
- Adamčík, S.; Marhold, K. Taxonomy of the Russula xerampelina group. I. Morphometric study of the Russula xerampelina group in Slovakia. Mycotaxon 2000, 76, 463–480. [Google Scholar]
- Buyck, B. The study of microscopic features in Russula 2. Sterile cells of the hymenium. Russulales News 1991, 1, 62–85. [Google Scholar]
- Caboň, M.; Eberhardt, U.; Looney, B.; Hampe, F.; Kolařík, M.; Jančovičová, S.; Verbeken, A.; Adamčík, S. New insights in Russula subsect. Rubrinae: Phylogeny and the quest for synapomorphic characters. Mycol. Prog. 2017, 16, 877–892. [Google Scholar]
- Buyck, B. Valeur taxonomique du bleu de crésyl pour le genre Russula. Bull. De La Société Mycol. De Fr. 1989, 105, 1–6. [Google Scholar]
- Zhou, L.L.; Liang, J.F. An improved protocol for extraction of DNA from macrofungi. Guangdong Fore. Sci. Tech. 2011, 27, 13–16. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protocols: A Guide to Methods and Applications; Academic Press: New York, NY, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Hall, B.D. Body plan evolution of ascomycetes, as inferred from an RNA polymerase II, phylogeny. Proc. Natl. Acad. Sci. USA 2004, 13, 4507–4512. [Google Scholar] [CrossRef]
- Matheny, P.B. Improving phylogenetic inference of mushrooms with RPB1 and RPB2 nucleotide sequences (Inocybe; Agaricales). Mol. Phyl. Evol. 2005, 35, 1–20. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp Ser 1999, 41, 95–98. [Google Scholar]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; et al. Fungal diversity notes 1036-1150: taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Song, Y.; Li, J.W.; Buyck, B.; Zheng, J.F.; Qiu, L.H. Russula verrucospora sp. nov. and R. xanthovirens sp. nov., two novel species of Russula (Russulaceae) from southern China. Cryptogamie Mycol 2018, 39, 129–142. [Google Scholar] [CrossRef]
- Sarnari, M. Monografia illustrate de genere Russula in Europa; Secondo, T., Ed.; AMB, Centro Studi Micologici: Trento, Italy, 1998. [Google Scholar]
- Buyck, B.; Mitchell, D.; Parrent, J. Russula parvovirescens sp. nov., a common but ignored species in the eastern United States. Mycologia 2006, 98, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Ying, J.Z. A study on Russula viridi-rubrolimata sp. nov. and its related species of subsection Virescentinas. Acta Mycol Sin 1983, 2, 34–37. [Google Scholar]
- Adamčík, S.; Jančovičová, S.; Buyck, B. The Russulas described by Charles Horton Peck. Cryptogamie Mycol 2018, 39, 3–108. [Google Scholar] [CrossRef]
- Sarwar, S.; Aziz, T.; Hanif, M.; Ilyas, S.; Shaheen, S. Russula swatica: A new species of Russula based on molecular, light microscopy, and scanning electron microscopy analyses from Swat Valley of Khyber Pakhtunkhwa province of Pakistan. Microsc. Res. Tech. 2019, 82, 1700–1705. [Google Scholar] [CrossRef]
- Mleczko, P. ; M. Kozak, and S. Zubek. Russula medullata (russulales, basidiomycota): A new species in the mycobiota or Poland. Pol. Bot. J. 2010; 55, 483–488. [Google Scholar]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Larsson, E.; Angelini, C.; Brandrud, T.E.; Dearnaley, J.D.W.; et al. Fungal Planet description sheets: 1112-1181. Persoonia 2020, 45, 251–409. [Google Scholar] [CrossRef]
Figure 1.
The phylogenetic tree based on the ITS-nrLSU-RPB2-mtSSU dataset. Species within the subgenus Heterophyllidia that exhibit high similarity to our new species, as well as representative species closely related to subsections Griseinae and Virescentinae, were selected. Support values are shown with Bootstrap support (BS) ≥70% and Bayesian Posterior Probabilities (PP) ≥0.95.
Figure 1.
The phylogenetic tree based on the ITS-nrLSU-RPB2-mtSSU dataset. Species within the subgenus Heterophyllidia that exhibit high similarity to our new species, as well as representative species closely related to subsections Griseinae and Virescentinae, were selected. Support values are shown with Bootstrap support (BS) ≥70% and Bayesian Posterior Probabilities (PP) ≥0.95.
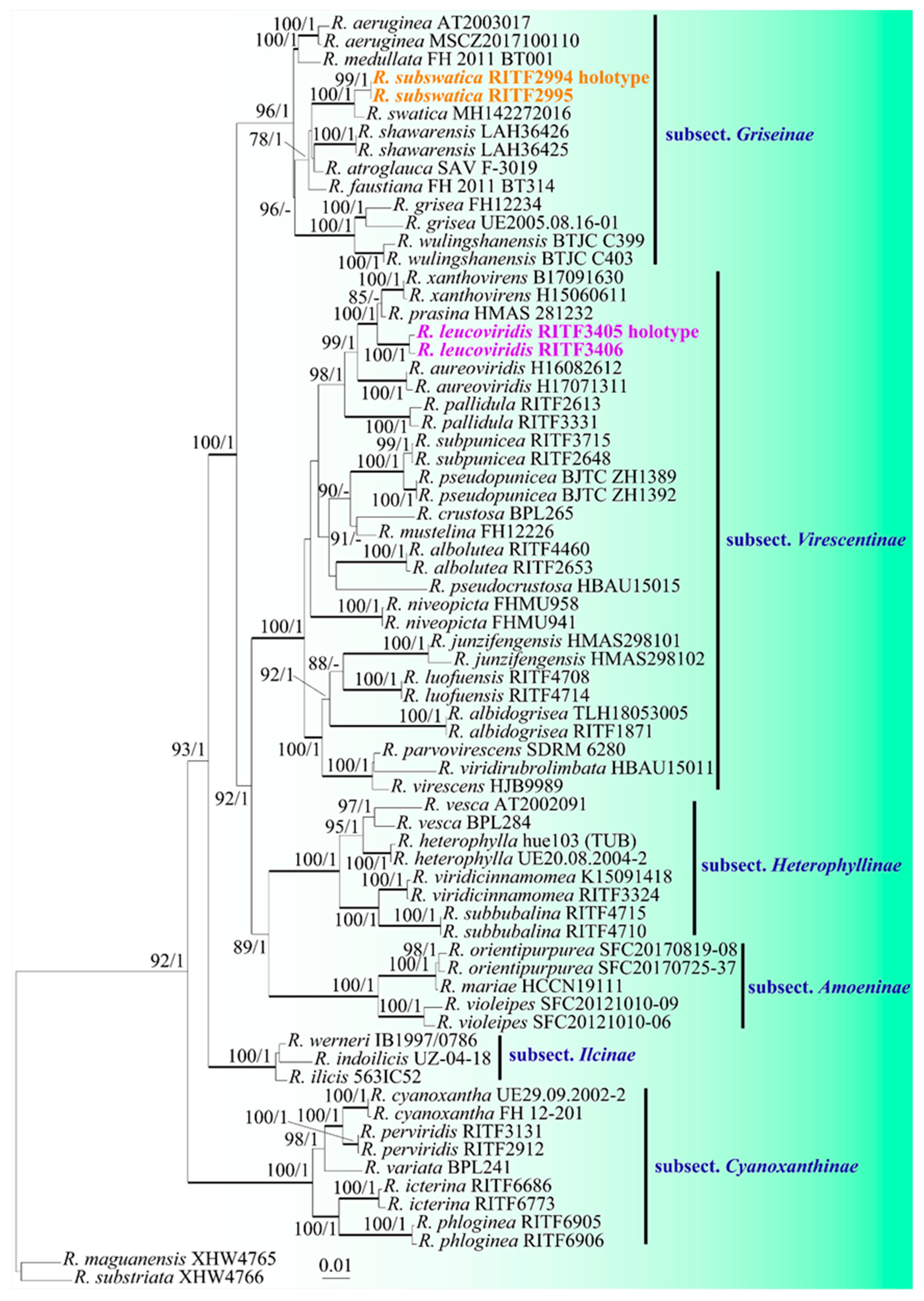
Figure 2.
Figure 2. Basidiomata and microscopic structures of Russula leucoviridis. (A–D) Basidiomata. (E,F) Basidiospores in SEM (JSM-6510LV). Scale bars: 10mm (A–D), 2 μm (E), 1μm (F).
Figure 2.
Figure 2. Basidiomata and microscopic structures of Russula leucoviridis. (A–D) Basidiomata. (E,F) Basidiospores in SEM (JSM-6510LV). Scale bars: 10mm (A–D), 2 μm (E), 1μm (F).

Figure 3.
Microscopic structures of Russula leucoviridis. (A) Basidia. (B) Basidiola. (C) Marginal cells. (D) Hymenial gloeocystidia on lamellae sides. (E) Hymenial gloeocystidia on lamellae edges. Scale bar: 10 μm.
Figure 3.
Microscopic structures of Russula leucoviridis. (A) Basidia. (B) Basidiola. (C) Marginal cells. (D) Hymenial gloeocystidia on lamellae sides. (E) Hymenial gloeocystidia on lamellae edges. Scale bar: 10 μm.
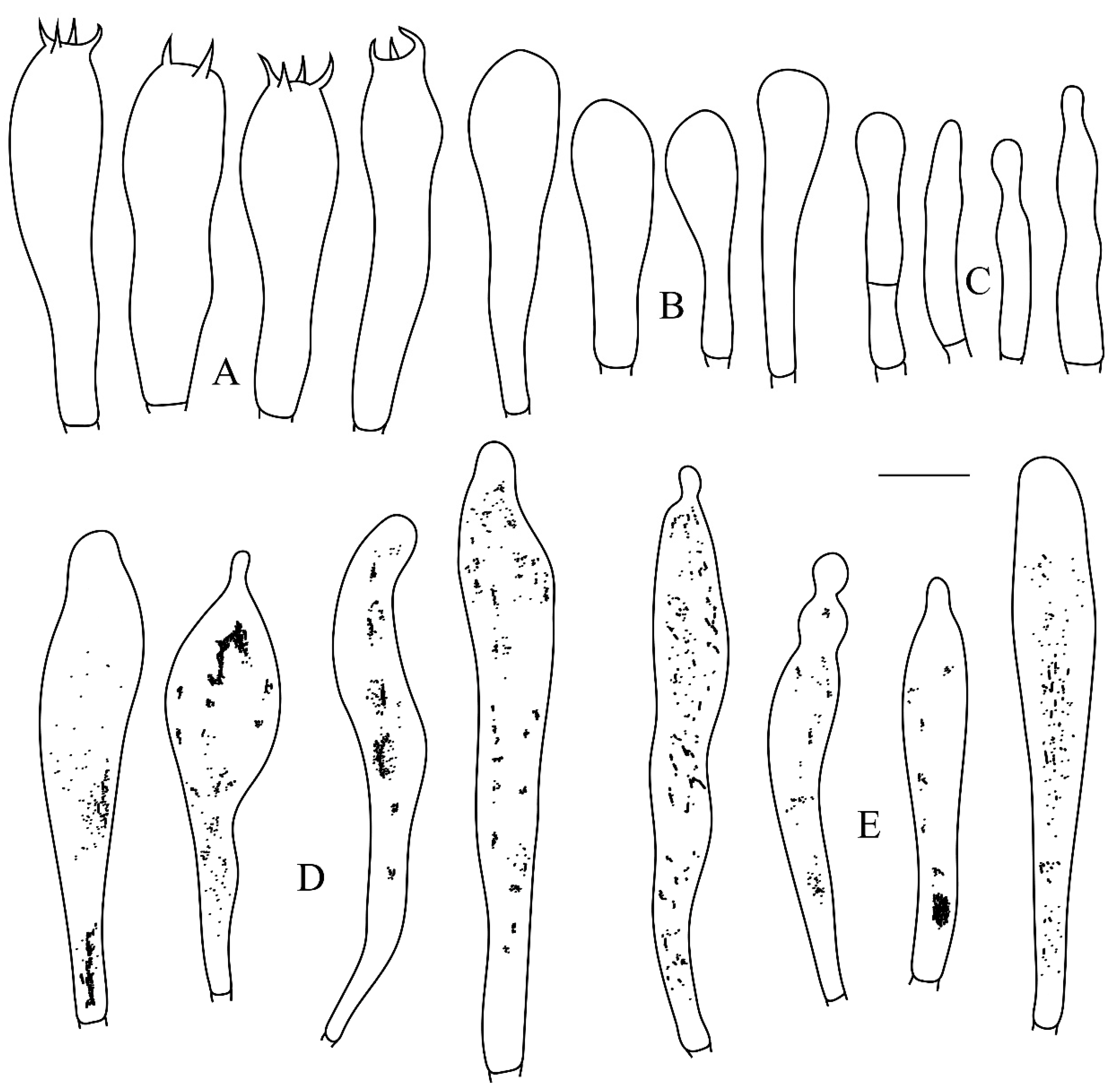
Figure 4.
Microscopic structures of Russula leucoviridis. (A) Pileocystidia near the pileus margin. (B) Pileocystidia near the pileus center. (C) Hyphal terminations near the pileus margin. (D) Hyphal terminations near the pileus center. Scale bar: 10 μm.
Figure 4.
Microscopic structures of Russula leucoviridis. (A) Pileocystidia near the pileus margin. (B) Pileocystidia near the pileus center. (C) Hyphal terminations near the pileus margin. (D) Hyphal terminations near the pileus center. Scale bar: 10 μm.
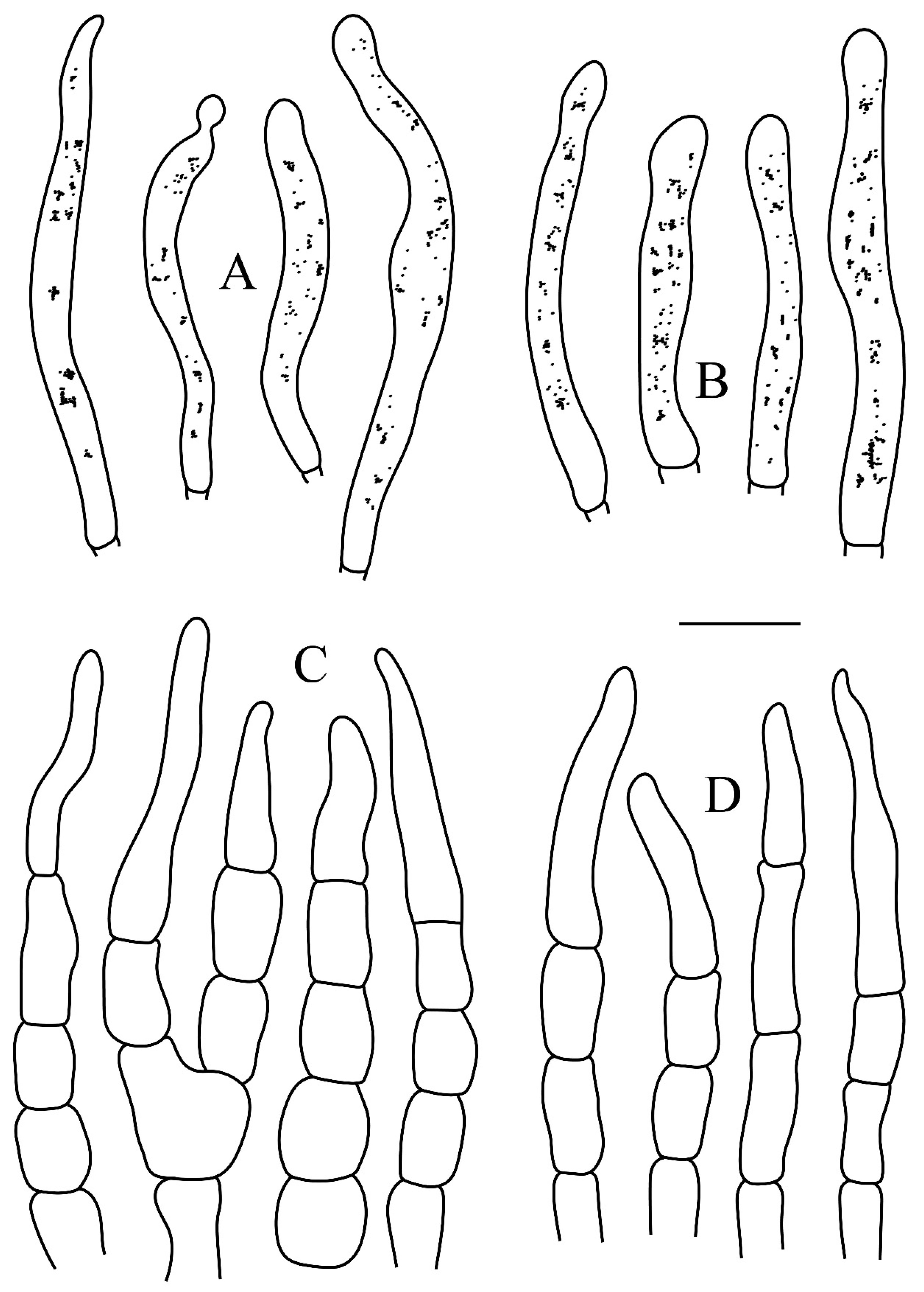
Figure 5.
Basidiomata and microscopic structures of Russula subswatica. (A) Basidiomata. (B–D) Basidiospores in SEM (JSM-6510LV). Scale bars: 15mm (A), 1μm (B–D).
Figure 5.
Basidiomata and microscopic structures of Russula subswatica. (A) Basidiomata. (B–D) Basidiospores in SEM (JSM-6510LV). Scale bars: 15mm (A), 1μm (B–D).

Figure 6.
Microscopic structures of Russula subswatica. (A) Basidia. (B) Basidiola. (C) Marginal cells. (D) Hymenial gloeocystidia on lamellae sides. (E) Hymenial gloeocystidia on lamellae edges. Scale bar: 10 μm.
Figure 6.
Microscopic structures of Russula subswatica. (A) Basidia. (B) Basidiola. (C) Marginal cells. (D) Hymenial gloeocystidia on lamellae sides. (E) Hymenial gloeocystidia on lamellae edges. Scale bar: 10 μm.

Figure 7.
Russula subswatica of Russula subswatica. (A) Pileocystidia near the pileus margin. (B) Pileocystidia near the pileus center. (C) Hyphal terminations near the pileus margin. (D) Hyphal terminations near the pileus center. Scale bar: 10 μm.
Figure 7.
Russula subswatica of Russula subswatica. (A) Pileocystidia near the pileus margin. (B) Pileocystidia near the pileus center. (C) Hyphal terminations near the pileus margin. (D) Hyphal terminations near the pileus center. Scale bar: 10 μm.
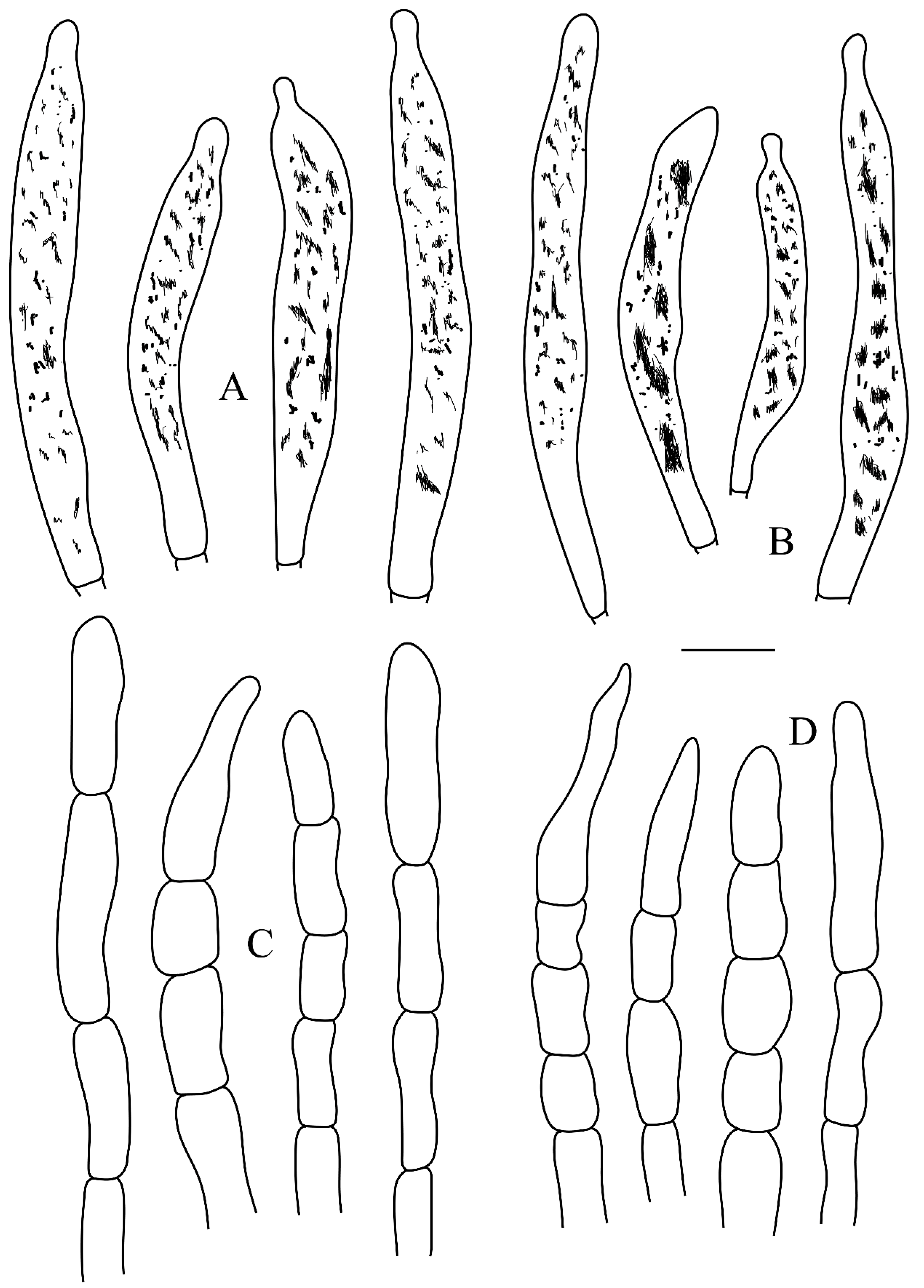

Table 1.
GenBank accession numbers for the sequences used in the phylogenetic analysis are provided, with newly generated sequences highlighted in bold.
Table 1.
GenBank accession numbers for the sequences used in the phylogenetic analysis are provided, with newly generated sequences highlighted in bold.
| Taxon | Voucher | Location | ITS | LSU | RPB2 | mtSSU |
| R. aeruginea | AT2003017 | Sweden | DQ421999 | DQ421999 | DQ421946 | – |
| R. aeruginea | MSCZ2017100110 | China | MH422576 | MK881944 | – | MK882072 |
| R. albidogrisea | TLH18053005 | China | MN275581 | MN839562 | MT085637 | MN839610 |
| R. albidogrisea | RITF1871 | China | MW397095 | MW397128 | – | MW403841 |
| R. albolutea | RITF2653 | China | MT672478 | MW397120 | MW411340 | MW403833 |
| R. albolutea | RITF4460 | China | MT672475 | MW397121 | MW411341 | MW403834 |
| R. atroglauca | SAV F-3019 | Slovakia | MT738270 | MT738248 | MT732156 | – |
| R. aureoviridis | H17071311 | China | MN275564 | MN839561 | MT085636 | MN839609 |
| R. aureoviridis | H16082612 | China | KY767809 | MK881920 | – | MK882048 |
| R. crustosa | BPL265 | USA | KT933966 | KT933826 | KT933898 | – |
| R. cyanoxantha | FH 12-201 | Germany | KR364093 | KR364225 | KR364341 | – |
| R. cyanoxantha | UE29.09.2002-2 | France | DQ422033 | DQ422033 | DQ421970 | – |
| R. faustiana | FH 2011 BT314 | Germany | MT738277 | MT738253 | MT732162 | – |
| R. grisea | FH12234 | Germany | KT934006 | KT933867 | KT933938 | – |
| R. grisea | UE2005.08.16-01 | Germany | DQ422030 | DQ422030 | DQ421968 | – |
| R. heterophylla | hue103 (TUB) | Europe | AF418609 | AF325309 | – | – |
| R. heterophylla | UE20.08.2004-2 | Sweden | DQ422006 | DQ422006 | DQ421951 | – |
| R. icterina | RITF6686 | China | PP700452 | PP700462 | PP707790 | PP700443 |
| R. icterina | RITF6773 | China | PP700453 | PP700463 | PP707792 | PP700445 |
| R. ilicis | 563IC52 | Europe | AY061682 | – | – | – |
| R. indoilicis | UZ-04-18 | India | MW547505 | – | – | – |
| R. junzifengensis | HMAS298101 | China | OR826832 | OR826833 | OR915864 | OR941507 |
| R. junzifengensis | HMAS298102 | China | OR880061 | OR880054 | OR915865 | OR941508 |
| R. leucoviridis | RITF3405 | China | PQ373872 | PQ373868 | PQ381291 | PQ373876 |
| R. leucoviridis | RITF3406 | China | PQ373873 | PQ373869 | PQ381292 | PQ373877 |
| R. luofuensis | RITF4708 | China | MW646975 | MW646987 | – | MW646998 |
| R. luofuensis | RITF4714 | China | MW646977 | MW646989 | – | MW647000 |
| R. maguanensis | XHW4765 | China | MH724918 | MH714537 | MH939989 | OQ586179 |
| R. mariae | HCCN19111 | South Korea | KF361762 | KF361812 | KF361712 | – |
| R. medullata | FH 2011 BT001 | Germany | MT738280 | MT738256 | MT732166 | – |
| R. mustelina | FH12226 | Germany | KT934005 | KT933866 | KT933937 | – |
| R. niveopicta | FHMU958 | China | OP837461 | OP837453 | – | – |
| R. niveopicta | FHMU941 | China | OP837462 | OP837454 | – | – |
| R. orientipurpurea | SFC20170819-08 | South Korea | MT017550 | – | MT199638 | MT196926 |
| R. orientipurpurea | SFC20170725-37 | South Korea | MT017548 | – | MT199639 | MT196927 |
| R. pallidula | RITF2613 | China | MH027958 | MH027960 | MH091698 | MW403845 |
| R. pallidula | RITF3331 | China | MH027959 | MH027961 | MH091699 | MW403846 |
| R. parvovirescens | SDRM 6280 | USA | MK532789 | – | – | – |
| R. perviridis | RITF3131 | China | OR907098 | OR907072 | OR914548 | OR934523 |
| R. perviridis | RITF2912 | China | OR907100 | OR907070 | OR914547 | OR934522 |
| R. phloginea | RITF6905 | China | OR907116 | OR907051 | OR914555 | OR934516 |
| R. phloginea | RITF6906 | China | OR907117 | OR907055 | OR914558 | OR934520 |
| R. prasina | HMAS 281232 | China | MH454351 | – | – | – |
| R. pseudocrustosa | HBAU15015 | China | MT337520 | – | – | – |
| R. pseudopunicea | BJTC ZH1392 | China | OP133164 | OP133204 | OP156853 | OP143937 |
| R. pseudopunicea | BJTC ZH1389 | China | OP133163 | OP133203 | OP156852 | OP143936 |
| R. shawarensis | LAH36425 | Pakistan | MT738293 | MT738267 | MT732176 | – |
| R. shawarensis | LAH36426 | Pakistan | MT738292 | MT738268 | MT732177 | – |
| R. subbubalina | RITF4710 | China | MW646978 | MW646990 | – | MW647001 |
| R. subbubalina | RITF4715 | China | MW646979 | MW646991 | – | MW647002 |
| R. subpunicea | RITF3715 | China | MN833635 | MW397124 | MW411344 | MW403837 |
| R. subpunicea | RITF2648 | China | MN833638 | MW397125 | MW411345 | MW403838 |
| R. substriata | XHW4766 | China | MH724921 | MH714540 | MH939992 | OQ371394 |
| R. subswatica | RITF2994 | China | PQ373870 | PQ373866 | – | PQ373874 |
| R. subswatica | RITF2995 | China | PQ373871 | PQ373867 | – | PQ373875 |
| R. swatica | MH142272016 | Pakistan | MK389374 | – | – | – |
| R. variata | BPL241 | USA | KT933959 | KT933818 | KT933889 | – |
| R. vesca | AT2002091 | Sweden | DQ422018 | DQ422018 | DQ421959 | – |
| R. vesca | BPL284 | USA | KT933978 | KT933839 | KT933910 | – |
| R. violeipes | SFC20121010-06 | South Korea | KF361808 | KF361858 | KF361758 | – |
| R. violeipes | SFC20121010-09 | South Korea | KF361807 | KF361857 | KF361757 | – |
| R. virescens | HJB9989 | Belgium | DQ422014 | DQ422014 | DQ421955 | – |
| R. viridicinnamomea | RITF3324 | China | MW397098 | MW397130 | MW411348 | MW403847 |
| R. viridicinnamomea | K15091418 | China | MK049972 | MK881938 | – | MK882066 |
| R. viridirubrolimbata | HBAU15011 | China | MT337526 | – | – | – |
| R. werneri | IB1997/0786 | Sweden | DQ422021 | – | – | – |
| R. wulingshanensis | BTJC C403 | China | OP133166 | OP133206 | OP156855 | OP143939 |
| R. wulingshanensis | BTJC C399 | China | OP133165 | OP133205 | OP156854 | OP143938 |
| R. xanthovirens | H15060611 | China | MG786056 | MN839560 | T085635 | MN839608 |
| R. xanthovirens | B17091630 | China | MG786055 | MK881928 | – | MK882056 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



