
Submitted:
15 October 2024
Posted:
15 October 2024
You are already at the latest version
Abstract
Drought stress is one of the most important climate-related factors affecting crop production. Tomatoes (Solanum lycopersicum L.) are economically important crop which are highly sensitive to drought. The entomopathogenic fungus Beauveria bassiana is a widely used biological insecticide,which can form symbiotic relationships with plants via endophytic colonization, increasing plant biomass and the ability to resist biotic stress. An increase in the water absorption capacity and relative water content of tomato roots as well as the number, length, and width of tomato leaves stomata were observed after B. bassiana colonization via root irrigation. Meanwhile, peroxidase activity and proline content increased significantly, while malondialdehyde reduced significantly. Expression levels of key genes related to stomatal development, as well as drought tolerance pathways such as jasmonic acid, ascorbate peroxidase, redox homeostasis, nitrogen metabolism, carotenoid biosynthesis, and fatty acid biosynthesis also increased significantly. These results indicate that colonization of B. bassiana enhances the water absorption capacity of tomato seedlings as well as the rate of transpiration via increase the number and size of stomata and improved drought tolerance in tomato seedlings significantly and increases drought tolerance in tomato via the “water spender” pathway, which provide a new strategy for improving crop resistance to drought stress.
Keywords:
entomogenous fungi
; endophytic colonization
; tomato
; drought tolerance
1. Introduction
Drought is one of the most critical abiotic factors directly affecting global crop yields. Due to global warming, the average temperature on Earth is continuing to rise, increasing the frequency of droughts. Meanwhile, the increasing demand for water in modern agriculture, shortages in freshwater resources, and continuous population growth are further exacerbating the impact of drought on crop yields, limiting normal plant growth and development[1,2]. Drought affects a series of physiological activities such as nutrient absorption, photosynthesis, and respiration, thereby affecting the overall growth rate and developmental cycle, which in turn, cause a decline in crop yield and quality[3,4]. Effective mitigation of the harm caused by drought stress is therefore an urgent issue in the field of plant protection.
Endophytes refer to all microorganisms that can form communities within plants and survive on plant organs, while causing no significant damage to the host[5]. It has also been shown that endophytes can cooperate with plants, reducing the effects of abiotic stresses such as drought via increased tolerance[6]. Beauveria bassiana is a broad-spectrum insect pathogenic fungus. In 1991, B. bassiana was also identified as an endophytic fungus in plants, possessing the ability to colonize agricultural crops[7]. The mutually beneficial symbiotic relationship between B. bassiana and plants has since been revealed[8], and the positive effects on plant resistance to biotic stress and biomass have been shown. For example, artificial inoculation of plants with B. bassiana resulted in an effective increase in the mortality rate of insect pests[9]. Colonization of B. bassiana via leaf spraying was also found to promote tomato growth and have insecticidal effects against tomato mites[10]. Previous research has also shown that endophytic colonization of B. bassiana blastospores induced resistance to various phytopathogens including sclerotinia sclerotiorum and Pythium Myriotylum[11,12], as well as increasing the number of fruit per plant and yield per unit area of tomatoes[13]. However, few reports have examined the effect of B. bassiana colonization on plant tolerance to abiotic stresses such as drought.
Long-term evolution has led to the formation of complex mechanisms of stress adaptation and resistance, with various physiological, biochemical, and molecular reactions occurring in plants under drought stress[14,15]. For example, the "drought avoidance" strategy causes changes in the root structure, allowing the stability of the root cell membrane to be maintained and sufficient water to be retained, as well as reducing evapotranspiration via closing of the stomata[16]. Increases in levels of abscisic acid and antioxidant activity also occur, together with the induction of defense mechanisms and activation of drought stress-related gene expression[17,18]. One of the most common stress resistance strategies in plants is the overproduction of different types of osmolyte, such as proline, mannitol, and sorbitol, which act to promote the stability of reactive oxygen species (ROS) detoxification, the overall structure of the cell membrane, and the natural structure of enzymes[19]. Plants also possess an antioxidant defense system, consisting of superoxide dismutase (SOD) and peroxidase (POD), as well as antioxidants such as carotenoids, all of which work together to protect plants from the destructive effects of ROS[20]. ROS can damage DNA, hinder normal protein synthesis, and cause a decrease or even inactivation of enzyme activity in plants, resulting in changes in cell membrane permeability and structure, and a subsequent increase in malondialdehyde (MDA), which is an important indicator of drought stress in plants[21].
Tomato (Solanum lycopersicum L.) is one of the most extensively studied vegetable crops as well as one of the most economically important. Drought is a critical abiotic stress, severely restricting tomato growth and yield[22]. Previous studies have shown that endophytic colonization of B. bassiana in Zea mays L. (Z. mays) stimulated root growth and flowering, thereby improving drought tolerance[23]. Research has further shown that this endophytic-induced increase in drought tolerance is achieved via two pathways: the "water saver" and "water spender" [24]. However, few reports have examined the effect of endophytic colonization of B. bassiana on drought tolerance in tomatoes. In this study, tomato seedlings were colonized with B. bassiana blastospores then subjected to drought stress in order to clarify the effect on water utilization pathways and subsequent drought tolerance. The findings provide a new strategy for improving crop resistance to drought stress.
2. Results
2.1. Colonization of B. bassiana in the Tomato Seedlings
The endophytic colonization rate of B. bassiana in the tomato seedlings was 74.19% . After being placed in a light incubator for 5-6 days, no white colony was observed in the control treatment, while under B. bassiana treatment white colonies growing around the leaf tissues were observed, the colony morphology consistent with B. bassiana (Figure S1a-c).
2.2. Effect of B. bassiana Colonization on Growth of Tomato Seedling Under Simulated Drought Conditions in Hydroponics
In the drought stress experiment, no significant differences in growth phenotypes were observed between the Control and Bb treatment groups under normal growth conditions. However, after 12 h of drought stress, the differences in growth phenotype gradually became apparent. The leaves of the PEG treatment group showed slight wilting, curling, and sagging, with significantly greater wilting than the other three treatment groups (Figure 1a). After 24 h of drought stress, leaves of plants in the PEG treatment group showed severe wilting and leaf curling, while drought symptoms were also apparent in the PB group, but to a much lesser degree (Figure 1b). After 36 h of drought stress, all seedlings experienced severe wilting in both the PEG and PB treatment groups (Figure 1c).
2.3. Effect of B. bassiana Treatment on Water Absorption
The effect of B. bassiana colonization on the water absorption capacity of the tomato seedlings was also determined. On day 4 after being placed in Hoagland nutrient solution, the water absorption capacity of plants in the Bb treatment group was significantly higher than that of the control, at 3.41±0.43 mL and 2.57±0.67 mL, respectively (p<0.05). Meanwhile, on day 8, the water absorption capacity of plants in the Bb treatment group was significantly higher than that of the Control (p<0.05), at 8.67±0.91 mL and 6.75±0.95 mL, respectively. Similar findings were also observed on day 12 (p<0.05), at 14.89±1.5 mL and 11.12±2.17 mL, respectively (Figure 2).
2.4. Effect of B. bassiana Colonization on Tomato Seedling Growth Indicators Under Drought Stress in the Pot Experiment
The growth phenotype of each treatment group was determined under simulated drought stress conditions in a pot experiment. Under normal growth conditions, the Bb treatment group showed better growth than the Control. Meanwhile, on day 1 after drought stress, the leaves of plants in the GH treatment group were narrower and wilted compared to those in the GB treatment group. On day 6 of drought stress, seedlings in the GH treatment showed slight wilting, curling, and drooping of leaves, while those in the GB treatment group showed slight drooping. On day 12 of drought stress, seedlings in the GH treatment group had dried up and wilted severely compared to those in the GB treatment group, while on day 18, both GH and GB treatment resulted in severe wilting and withering of the leaves (Figure 3a).
Significant differences in seedling height between treatments were observed on days 1, 6, 12, and 18 after drought stress (p<0.05; Figure 3b). Over time, the seedling height in the groups under drought stress was significantly lower than that of the control and Bb treatments. The height of seedlings in groups treated with B. bassiana was higher than that of those treated without B. bassiana (Bb vs. Control and GB vs. GH) under identical watering conditions, respectively. On days 1, 6 and 12 after drought stress, the height of seedlings in the GB treatment group was significantly higher than that of seedlings in the GH treatment group. Meanwhile, under Bb treatment, seedling height was significantly higher than that of the Control group at all time points. These findings indicate that root irrigation with B. bassiana caused an increase in the height of the tomato seedlings under both normal and drought stress conditions.
Similar to the results of plant height, significant differences in stem diameter were also observed between treatments on days 1, 6, 12, and 18 after drought stress (p<0.05; Figure 3c). Over time, the stem diameter in the groups under drought stress was significantly lower than that of the control and Bb treatments. The stem diameter of seedlings treated with B. bassiana was higher than that of those treated without B. bassiana (Bb vs. Control and GB vs. GH) under identical watering conditions, respectively. On days 1, 6, 12 and 18 after drought stress, the stem diameter of seedlings in the GB treatment group was significantly higher than that of plants in the GH treatment group. Meanwhile, stem diameter was also significantly higher in the Bb treatment compared to the Control group. These findings indicate that irrigation with B. bassiana caused an increase in the stem diameter of tomato seedlings under both normal and drought stress conditions.
Meanwhile, significant differences in root length were observed between treatments on day 18 of drought stress (p<0.05; Figure 3d). The root length in the groups under drought stress was significantly lower than that of the control and Bb treatment groups. Drought caused a significant reduction in root length, although the reduction was less in groups treated with B. bassiana compared to those treated without B. bassiana (Bb vs. Control and GB vs. GH) under identical watering conditions, respectively.
There was a significant difference in FW among treatments on day 18 of drought treatment (p<0.05). Both the FW and relative WC in the groups under drought stress was significantly lower than that of the control and Bb treatment groups. Both the FW and relative WC of groups treated with B. bassiana was higher than that of those treated without B. bassiana (Bb vs. Control and GB vs. GH) under identical watering conditions, respectively. Meanwhile, the DW was lower in the B. bassiana treatment groups (Bb and GB) compared to those treated without B. bassiana (Control and GH) (Figure 3e-g). These findings indicate that treatment with B. bassiana increased the water absorption capacity of the tomato plants.
2.5. Effect of B. bassiana Treatment on Stomata Morphology in Tomato Leaves Under Drought Stress
The number of stomata under each treatment was observed and counted during different stages of drought stress using a microscope (Figure 4a). Significant differences in the number of stomata were observed in each treatment on days 1, 6, and 12 of drought stress (p<0.05). Overall, the number of stomata was higher in tomato leaves under drought stress compared to Control and Bb treatment groups, as well as higher in groups treated with B. bassiana compared to those groups treated without B. bassiana (Bb vs. Control and GB vs. GH). Furthermore, with increasing drought stress, the number of stomata increased significantly under both GH and GB treatment, with a significantly higher number under GB compared to GH treatment. On day 12 of drought stress, the number of stomata per unit area was 34.94 ± 1.51 under GB treatment compared to 30.24 ± 0.69 under GH treatment (p<0.05; Figure 4c).
Stomatal length and width under each treatment were also observed and measured on day 12 of drought stress using a microscope (Figure 4b). The length and width of stomata were higher in tomato leaves treated with B. bassiana compared to those treated without B. bassiana (Bb vs. Control and GB vs. GH). Stomatal lengths were greater under Bb and GB treatment (24.53 ± 1.58 μm and 23.57 ± 1.9 μm, respectively) compared to the Control and GH treatment (21.24 ± 1.7 μm and 21.48 ± 1.21 μm, respectively) (Figure 4d), as were stomatal widths (16.32 ± 0.96 μm and 16.18 ±0.7 μm, vs. 15.08 ± 1.42 μm and 15.14 ± 0.76 μm, respectively) (Figure 4e).
2.6. Analysis of Physicochemical Indexes in the Tomato Seedlings
Significant differences in POD activity were observed between treatments on days 1, 6, 12 and 18 of drought stress (p<0.05). Overall, activity was higher in tomato seedlings treated with B. bassiana compared to those without (Bb vs. Control and GB vs. GH) (p<0.05), suggesting that B. bassiana colonization activated POD activity (Figure 5a).
Significant increases in PRO content were also observed under GH and GB treatment compared to Bb and Control treatment on days 1, 6, 12 and 18 of drought stress (p<0.05), and on day 1of drought stress, results were significantly higher under Bb compared to Control treatment. These findings indicate that both drought and B. bassiana treatment resulted in an increase in the PRO content. Compared with GH treatment, a significant increase in PRO content was observed under GB treatment on days 1, 6, 12 and 18 of drought stress (p<0.05), suggesting that under drought conditions, B. bassiana treatment caused a further increase in PRO content in the tomato seedlings (Figure 5b).
Compared to the Control treatment, a decrease in MDA content was observed on day 1 of drought stress in plants treated with B. bassiana (p<0.05). Overall, drought stress caused a significant increase in MDA accumulation in all plants (p<0.05); however, a significant reduction was observed under GB compared to GH treatment. This indicates that treatment with B. bassiana inhibited the drought-stress induced accumulation of MDA in the tomato seedlings (Figure 5c).
2.7. Analysis of Physicochemical Indexes in the Tomato Seedlings
Compared with the Control, three genes related to signaling pathways involved in plant stomatal development and regulation (Solyc08g061560.2, Solyc12g042760.1 and Solyc09g091760.1) were up-regulated on day 1 of drought stress following treatment with B. bassiana (Bb and GB) (Figure 6a-c). On day 12 of drought treatment, expression of all three genes was up-regulated under both B. bassiana treatment and drought stress (Bb, GH, and GB). In addition, on day 1 of drought stress, treatment with B. bassiana also resulted in the upregulation of relative expression levels of five drought resistance pathway-related genes (Figure 6d-h). On day 12 of drought stress, all genes except for the PSY gene were upregulated in the Bb treatment group compared to the Control. These findings indicate that treatment with B. bassiana induced the expression of drought resistant-related genes in the tomato seedlings.
3. Discussion
Numerous studies have focused on the problem of drought in crop production; for example, in terms of the management of water conservancy facilities[25,26], breeding of drought tolerant varieties[27], use of mulching[28], moisture preservation[29], and the addition of exogenous substances to enhance plant drought tolerance[30,31]. Exogenous application of 2,4-electrobrassinolide (EBR) was found to effectively alleviate wilting of tomato seedlings and the decrease in the relative water content of leaves[32], as well as increasing the activity of SOD, POD, and CAT enzymes, and the expression of related genes (FeSOD, POD, and CAT). Meanwhile, in tobacco, increases in osmoregulatory substances such as proline and soluble sugars, and the content of plant growth hormone (IAA) and brassinolide, as well as up-regulation of drought tolerance-related genes induced by BR (BRL3 and BZR2) and IAA (YUCCA6, SAUR32, and IAA26) signaling pathways were also observed following , all of which served to enhance drought resistance[33,34,35]. Studies have also shown that endophytic microorganisms have a significant positive effect on plant drought tolerance[36,37,38]. B. bassiana is an important biological insecticide, and has been widely commercialized for the biological control of various agricultural pests[39,40,41]. Recent studies have further shown that B. bassiana can colonize plants and alter their biological characteristics, promoting growth as well as disease and insect resistance[42,43,44,45,46]. Our previous results indicated that compared to hydrophobic conidia, hydrophilic blastospores of B. bassiana not only resulted in higher resistance to Botrytis cinerea infection after endophytic colonization in tomatoes[47], but also improved tomato yield[13]. In the present study, we found that colonization of B. bassiana blastospores significantly improved drought tolerance in tomato seedlings under both potted drought stress and hydroponic PEG-simulated drought conditions. Compared to hydrophobic conidia, hydrophilic blastospores offer various advantages because of their degree of water solubility and ease of production.
Previously study has found that colonization of B. bassiana promoted the growth of maize roots, thereby improving their tolerance to drought stress[43]. In this study, endophytic colonization of B. bassiana in tomatoes promoted root growth and increased the water absorption capacity, as evidenced by the increase in FW and WC. There has already been studied suggested that plants enhance their tolerance to drought stress by changing their morphological structure; for example, by increasing their root to shoot ratio, decreasing stomatal aperture size, and increasing stomatal density, all of which are common response mechanisms during plant adaptation to drought stress[48]. A previous study revealed that colonization with different strains of B. bassiana resulted in a significant increase in the number of tobacco epidermal hairs, the stomatal density, and the stomatal index, thereby promoting the development of tobacco epidermal stomata[49]. Meanwhile, an increase in leaf stomatal density was also observed in rice and Leymus chinensis plants under mild and moderate drought stress, although a decrease in leaf stomatal density was observed under severe drought conditions[50]. The results of our study revealed that B. bassiana colonization in tomato seedlings caused an increase in the expression of key genes related to the size and number of stomata, resulting in a significant increase in both, which in turn affects the transpiration of water. Aimone et al. found that colonization by endophytic fungi improves drought tolerance via two main pathways: the “water saver” and “water spender”[24]. The results of this study suggest that endophytic colonization of B. bassiana in tomatoes enhances drought resistance via the “water spender” pathway. Namely, colonization of B. bassiana promoted water absorption via the roots, thus maintaining the stability of the plant cell membranes, and improving drought tolerance.
Plants have developed various mechanisms of drought tolerance. Of these, the clearance of free radicals (ROS) plays an important role in short-term survival under severe stress, while metabolic changes contribute to long-term protection[51]. Under drought conditions, plants regulate their enzymatic system via superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), glutathione reductase (GR), and other enzymes that act to clear ROS. Of these, POD is known to cooperate with SOD to eliminate ROS, thereby enhancing plant resistance. In this study, colonization of B. bassiana under drought stress significantly increased POD activity in the tomato seedlings, resulting in the effective removal of ROS and subsequent protection or a reduced risk of drought-induced damage. Osmotic adjustment (OA) has been shown to be an important physiological adaptation to drought stress, improving overall drought tolerance[14].
Proline is a highly water-soluble substance that helps maintain the water content of plant cells and tissues[52,53] . An increase in free proline was found to not only alleviate damage caused by drought-induced water loss in plants, but also maintain water retention in lemon leaf cells and tissues[54,55]. In this study, even under normal growth conditions, B. bassiana colonization caused a significant increase in the content of proline in the tomato seedlings. Furthermore, under drought stress, an increase in proline was observed under all treatments, although this was significantly higher in plants treated with B. bassiana. Proline dehydrogenase (PRO) plays an important role in the synthesis of proline, and in this study, a significant increase in PRO activity was observed in tomato seedlings treated with B. bassiana. This highlights the mechanism behind the increase in proline content. Meanwhile, in contrast, MDA disrupts the structure and function of proteins, thereby disrupting the normal physiological and metabolic rhythm of plants, preventing healthy growth and development. MDA plays an important role in plant membrane lipid peroxidation, changes in which can serve as an important indicator of stress-induced damage. Research has also shown a significant positive correlation between changes in proline content and MDA in plants under stress. For example, under combined drought and salt stress, the membrane permeability of rice seedlings was found to increase together with increases in contents of proline and MDA[56]. In this study, the MDA content of the tomato seedlings increased significantly following drought stress; however, a significant decrease was observed in plants colonized with B. bassiana. These findings suggest that B. bassiana colonization significantly reduced the degree of membrane lipid peroxidation in tomato plants under drought stress.
A previous study revealed that colonization of B. bassiana caused an increase in resistance to biotic stress and significant increases in expression levels of key genes related to plant resistance pathways, such as the salicylic acid pathway, jasmonic acid (JA) pathway, and ethylene pathway[49]. B. bassiana colonization was also found to induce up-regulation of key genes related to these pathways in tomatoes, thereby increasing resistance to B. cinerea and Sclerotinia sclerotiorum[12,13]. Plant tolerance to drought is known to be closely related to various physiological pathways, including the JA pathway, ascorbate peroxidase (APX) pathway, nitrogen metabolism pathway, carotenoid biosynthesis pathway, and ABA biosynthesis pathway[57].When subjected to drought stress, expression levels of associated genes are up-regulated, thereby activating the above-mentioned pathways. In line with this, endophytic colonization of B. bassiana significantly increased expression levels of key genes in the tomato seedlings under drought stress, thereby activating related pathways, nevertheless the biochemical levels such as endogenous JA and nitrogen need to be identified in further study(Figure 7).
4. Materials and Methods
The tomato species BEAUTY was bred by Jilin Dalu Seed Industry Co., Ltd. (Gongzhuling, Jilin Province, China). Peat soil was obtained from Pindstrup Mosebrug A/S (Ryomgard, Denmark) and used for the drought stress pot experiments. B. bassiana strain D1-5[13]was isolated and identified from diseased larvae of Ostrinia furnacalis in the field by the Institute of Plant Protection, Jilin Academy of Agricultural Sciences, Changchun, Jilin Province, China.
B. bassiana conidia were scraped off then cultured in sterilized Sabouraud Dextrose Broth with Yeast Extract (SDY) medium for 96 h in a shaker at 26 ℃ and 200 r/min. They were then centrifuged and filtered to obtain clean blastospores[58]. The suspension was diluted using an appropriate amount of 0.05% (v/v) Tween-80 then mixed evenly to a final concentration of 1×108 spores/mL as determined using a hemocytometer under a microscope. The samples were then stored at 4 ℃ until use[47]. Healthy tomato seeds of uniform size were selected for inoculation. The plant surfaces were disinfected with 1% sodium hypochlorite (NaClO) before sowing in pots (height: 18.5 cm, diameter: 15.2 cm) containing peat soil. When seedlings reached a height of 10 cm, root irrigation with B. bassiana blastospores was carried out. As a control, seedlings were irrigated with 0.05% (v/v) Tween-80. A total of three replicates were prepared with 10 plants per replicate. The inoculation amount per plant was 40 mL, with a 24-h interval between each treatment. To caculated the colonization rate of B. bassiana, we used colonization rate equation:
PEG treatment was employed to induce drought stress under hydroponic conditions[59]. Briefly, tomato seedlings irrigated with B. bassiana or 0.05% (v/v) Tween-80 and showing consistent growth were held under running water to remove any soil from their roots. They were then washed three times with distilled water and placed in 500-mL triangular bottles containing Hoagland nutrient solution for 12 h to eliminate damage caused during transplantation.
Four treatments were examined in this experiment. Treatment 1: tomato seedlings root irrigated with 0.05% (v/v) Tween-80 then placed in Hoagland nutrient solution (Control); Treatment 2: tomato seedlings root irrigated with B. bassiana then placed in Hoagland nutrient solution (Bb); Treatment 3: tomato seedlings root irrigated with 0.05%(v/v) Tween-80 then placed in Hoagland nutrient solution with a final concentration of 8% (w/v) PEG-6000 (PEG); Treatment 4: tomato seedlings root irrigated with B. bassiana then placed in Hoagland nutrient solution with a final concentration of 8% PEG-6000 (PB). Morphological changes in the tomato seedlings were observed and recorded at 0, 12, 24, and 36 h after treatment, and photos were obtained by an electronic camera. Each treatment involved 10 plants, all of which were placed in a greenhouse at a constant temperature of 26 ℃.
A hydroponics experiment was conducted to determine the effects of B. bassiana colonization on the water absorption ability of the tomato seedlings. Tomato seedlings irrigated with either 0.05% (v/v) Tween-80 or B. bassiana and showing consistent growth were selected. Rhizosphere soil was removed as described above then all seedlings were placed in tubes containing 50-mL Hoagland nutrient solution for 12 days. Two treatments were examined in this experiment, with 10 seedlings per treatment. Treatment 1: tomato seedlings root irrigated with 0.05% (v/v) Tween-80 root then placed in Hoagland nutrient solution (Control); Treatment 2: tomato seedlings root irrigated with B. bassiana then placed in Hoagland nutrient solution (Bb). The amount of water remaining in each tube was observed and recorded every 2 days to determine the rate of absorption[60].
A pot experiment was carried out to examine the effect of drought stress on tomato seedling biomass. Four treatments were examined, with three replicates of 10 seedlings per treatment. Treatment 1: tomato seedlings root irrigated with 0.05% (v/v) Tween-80 root irrigation (Control); Treatment 2: tomato seedlings root irrigated with B. bassiana (Bb); Treatment 3: tomato seedlings root irrigated with 0.05% (v/v) Tween-80 then exposed to natural drought (GH); Treatment 4: tomato seedlings root irrigated with B. bassiana then exposed to natural drought (GB). Under treatments 1 and 2, plants root irrigated with Tween-80 or B. bassiana were irrigated with an equal amount of distilled water three days after treatment and every 3 days thereafter, while no water was provided under treatments 3 and 4. Evaluation of plant drought stress was determined based on the surface soil is dry in the pots. Height and stem diameters were recorded and measured days 1, 6, 12, and 18 after observation of drought stress, while Root length, fresh weight (FW), and dry weight (DW) were measured on day 18. The length of the seedling from soil level to the highest point of growth was determined using a ruler as a measure of plant height, while the diameter of the stem one-third below the cotyledon was determined using a vernier caliper as a measure of stem diameter. To determine root length, underground parts were removed and washed then surface moisture was absorbed before measuring the length using a vernier caliper. FW was determined using a TP-114 thousandth analytical balance (Beijing Sadolis Instrument System Co., Ltd, Beijing, China), while DW was determined after drying. Briefly, whole seedlings with soil removed were placed in a 105 ℃ oven for 30 min to induce withering then in an oven set to 65 ℃ for 24 h. The final weight was determined as a measure of DW[61]. The relative water content (WC) of the plants was then calculated as follows: WC (%) = (FW - DW) / FW × 100.
On day 12 of natural drought stress, 10 tomato seedlings from the drought stress pot experiment were randomly selected from each of the three replicates in each treatment group. Samples were then taken from the same position of the third fully expanded leaf from the top in each seedling. Dust and dirt on the leaf surface were gently removed using sterile cotton balls then a thin layer of nail polish was applied on the front of the leaf. The leaf surface was then allowed to dry naturally for 10 - 20 min. After drying, the nail polish was removed from the leaf, spread onto a slide and covered. A confocal microscope Leica SP8 laser scanning confocal microscope (Leica Microsystems (Shanghai) Trading Co., Ltd, Germany) was then used to obtain images of the stomata under five different fields of view. The number of stomata in each field was determined under 20× magnification then one stoma from three images in each replicate was randomly selected for measurement of the length and width at 40× magnification [62].
Leaves were collected from each of the four treatment groups in the drought stress pot experiments for analysis of physicochemical properties and gene expression levels. Five randomly selected seedlings from each replicate were sampled on days 1, 6, 12, and 18 of drought stress. The top three leaves were then removed and stored at -80 ℃ until use. A Peroxidase assay kit (A084-3-1), Proline detection kit (A107-1-1), and Plant Malondialdehyde Detection Kit (A003-3-1) (Nanjing Jiancheng Biotechnology Company, Nanjing, China) were used for analysis of POD, PRO, and MDA contents, respectively, based on the manufacturer’s instructions.
Analysis of the following stress resistance-related genes was also carried out: the AOS gene, which is involved in jasmonic acid (JA) synthesis and the signal response pathway[63]; the APX2 gene, which is related to ascorbate peroxidase and redox homeostasis[64]; the NIR1 gene, which is associated with nitrite reductase related to the nitrogen metabolism pathway[65]; the PSY gene, which is associated with the limiting enzyme of carotenoid and abscisic acid (ABA) biosynthesis[66]; and the FAB2 gene, which is associated with fatty acid (FA) biosynthesis[67]. Total RNA was extracted from the tomato leaf samples using TRIzol reagent (Nanjing Jiancheng Biotechnology Company, Nanjing, China) according to the manufacturer’s instructions then reverse transcription was performed using the total RNA as a template for synthesis of cDNA[68]. Real-time quantitative RT-PCR(qPCR) was then performed to determine gene expression levels under different treatments using the 2-ΔΔCT relative quantification method[69]. mRNA sequences of the eight candidate genes were determined in the NCBI (https://www.ncbi.nlm.nih.gov/) database[70,71], and the Actin 7 gene (ACT) was used as a reference[72]. The specific primers, sample cDNA synthesis reaction system, and qPCR conditions are listed in Supplementary Table S1. The primers were synthesized by Sangong Biotechnology Co., Ltd. (Shanghai, China).
All experimental results represent the average of all replicates. Multiple comparison tests were performed on all datasets using IBM SPSS Statistics 26 software to evaluate significant differences between treatments. GraphPad 8.0.2 software was used to plot the results.
5. Conclusions
Endophytic colonization of B. bassiana was found to promote root growth in tomato seedlings under drought stress as well as upregulating the expression of key genes related to stomatal formation. The number and size of stomata per unit area also increased, thereby promoting the water utilization efficiency. These findings suggest that colonization of B. bassiana in tomato seedlings increased drought resistance via the "water spender" pathway. Endophytic colonization of B. bassiana also promoted enzyme activity, as well as expression levels of various key genes related to drought resistance (Figure 7). Overall, these findings provide a new strategy for improving the survival ability of tomato plants grown under drought stress.
Author Contributions
Conceptualization, Z.Z. and L.S.; methodology, W.G.; software, W.G.; validation, Y.L., Q.L. and S.D.; formal analysis, X.Z.; investigation, X.Z.; resources, Z.Z.; data curation, S.D.; writing—original draft preparation, W.G. and Z.Z.; writing—review and editing, L.S.; visualization, Y.L.; supervision, Q.L.; project administration, Z.Z.; funding acquisition, Y.L. and Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
Jilin Provincial Scientifc and Technological Development Program, Grant Number 20230203175SF and 20230508011RC.
Acknowledgments
We would like to thank the native English speaking scientists of the Elixigen Company (Huntington Beach, California, USA) for editing our manuscript.
Conflicts of Interest
The authors have no conflicts of interest to declare.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data availability statement
Data available on request from the corresponding authors.
References
- Zytynska, S. Embracing the complexity of plant-microbe-insect interactions under a changing climate for sustainable agriculture. Curr Opin Insect Sci. 2021, 44, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Krishna, R.; Ansari, W.; Soumia, P.; Yadav, A.; Jaiswal, D.; Kumar, S.; Singh, A.; Singh, M.; Verma, J. Biotechnological Interventions in Tomato (Solanum lycopersicum) for Drought Stress Tolerance: Achievements and Future Prospects. BioTech. 2022, 11, 48. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Eti, F.; Ahmed, R.; Gupta, S.; Jhan, P.; Islam, T.; Bhuiyan, M.; Rubel, M.; Khayer, A. Drought-responsive genes in tomato: meta-analysis of gene expression using machine learning. Scientific reports. 2023, 13, 19374. [Google Scholar] [CrossRef]
- González, E. Drought Stress Tolerance in Plants. International journal of molecular sciences. 2023, 24, 6562. [Google Scholar] [CrossRef]
- Yan, L.; Zhu, J.; Zhao, X.; Shi, J.; Jiang, C.; Shao, D. Beneficial effects of endophytic fungi colonization on plants. Applied microbiology and biotechnology. 2019, 103, 3327–3340. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.; Shurigin, V.; Hashem, A.; Abd, A.; Endophytic Bacteria Improve Plant Growth. Symbiotic Performance of Chickpea (Cicer arietinum L.) and Induce Suppression of Root Rot Caused by Fusarium solani under Salt Stress. Front Microbiology. 2017, 28, 1887. [Google Scholar]
- Bing, L.; Lewis, L. Suppression of Ostrinia nubilalis (Hübner)(Lepidoptera: Pyralidae) by endophytic Beauveria bassiana (Balsamo) Vuillemin. Environmental entomology. 1991, 20, 1207. [Google Scholar] [CrossRef]
- Půža, V.; Tarasco, E. Interactions between Entomopathogenic Fungi and Entomopathogenic Nematodes. Microorganisms. 2023, 8, 163. [Google Scholar] [CrossRef]
- Bamisile, B. S.; Dash, C. K.; Akutse, K. S.; Qasim, M.; Ramos Aguila, L. C.; Wang, F.; Keppanan, R.; Wang, L. Endophytic Beauveria bassiana in Foliar-Treated Citrus limon Plants Acting as a Growth Suppressor to Three Successive Generations of Diaphorina citri Kuwayama (Hemiptera: Liviidae). Insects. 2019, 10, 176. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, Y.; Zhang, J.; Liu, X.; Ju, Z.; Shi, H.; Mendoza-Mendoza, A.; Zhou, W. ; Dual role of endophytic entomopathogenic fungi: induce plant growth and control tomato leafminer Phthorimaea absoluta. Pest Manag Sci. 2023, 79, 4557. [Google Scholar] [CrossRef]
- Sui, L.; Lu, Y.; Yang, H.; et al. Endophytic Beauveria bassiana promotes sunflower growth and confers resistance against sclerotinia disease. BioControl. 2024. [Google Scholar] [CrossRef]
- Deb, L.; Dutta, P. Antagonistic potential of Beauveria bassiana (Balsamo) Vuillemin against Pythium myriotylum causing damping off of tomato. Indian Phytopathology. 2021, 74, 715. [Google Scholar] [CrossRef]
- Sui, L.; Lu, Y.; Zhou, L.; Li, N.; Li, Q.; Zhang, Z. Endophytic Beauveria bassiana promotes plant biomass growth and suppresses pathogen damage by directional recruitment. Front Microbiol. 2023. 16, 1227269. [CrossRef]
- Ozturk, M.; Turkyilmaz, U.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol Plant. 2021, 172, 1321. [Google Scholar] [CrossRef] [PubMed]
- Parrotta, L.; Aloisi, I.; Faleri, C.; Romi, M.; Del Duca, S.; Cai, G. Chronic heat stress affects the photosynthetic apparatus of Solanum lycopersicum L. cv Micro-Tom. Plant physiology and biochemistry. 2020, 154, 463. [Google Scholar] [CrossRef]
- Zuo, H.; Chen, J.; Lv, Z.; Shao, C.; Chen, Z.; Zhou, Y.; Shen, C. Tea-Derived Polyphenols Enhance Drought Resistance of Tea Plants (Camellia sinensis) by Alleviating Jasmonate-Isoleucine Pathway and Flavonoid Metabolism Flow. International journal of molecular sciences. 2024, 25, 3817. [Google Scholar] [CrossRef]
- Zhu, Y.; Yan, J.; Liu, W.; Liu, L.; Sheng, Y.; Sun, Y.; Li, Y.; Scheller, H. V.; Jiang, M.; Hou, X.; Ni, L.; Zhang, A. Phosphorylation of a NAC Transcription Factor by a Calcium/Calmodulin-Dependent Protein Kinase Regulates Abscisic Acid-Induced Antioxidant Defense in Maize. Plant physiology. 2016, 171, 1651–1664. [Google Scholar] [CrossRef]
- Ramachandra, R.; Chaitanya, K.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J Plant Physiol. 2004, 161, 1189. [Google Scholar]
- Alhaithloul, H. Impact of Combined Heat and Drought Stress on the Potential Growth Responses of the Desert Grass Artemisia sieberi alba: Relation to Biochemical and Molecular Adaptation. Plants (Basel, Switzerland). 2019, 8, 416. [Google Scholar] [CrossRef]
- Zhu, S.; Zhao, W.; Sheng, L.; Yang, X.; Mao, H.; Sun, S.; Chen, Z. Integrated transcriptome and metabolomics analyses revealed key functional genes in Canna indica under Cr stress. Scientific reports. 2024, 14, 14090. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, F.; Li, S.; Feng, Y.; Yang, J.; Zhang, N.; Si, H. Calcium-Dependent Protein Kinase 28 Maintains Potato Photosynthesis and Its Tolerance under Water Deficiency and Osmotic Stress. International journal of molecular sciences. 2022, 23, 8795. [Google Scholar] [CrossRef] [PubMed]
- Conti, V.; Parrotta, L.; Romi, M.; Del, D.; Cai, G. Tomato Biodiversity and Drought Tolerance: A Multilevel Review. Int J Mol Sci. 2023, 24, 10044. [Google Scholar] [CrossRef] [PubMed]
- Kuzhuppillymyal, L.; Tamez, P.; Gomez, R.; Rodriguez, M.; Ek-Ramos, M. Endophytic Beauveria bassiana promotes drought tolerance and early flowering in corn. World J Microbiol Biotechnol. 2020, 36, 47. [Google Scholar] [CrossRef] [PubMed]
- Aimone, C.; Giauque, H.; Hawkes, C. Fungal symbionts generate water-saver and water-spender plant drought strategies via diverse effects on host gene expression. Phytobiomes Journal. 2023, 7, 2471–2906. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, Y.; Wang, L.; Xie, W.; Song, L.; Zhang, H.; Bi, H.; Zheng, Y.; Zhang, Y.; Zhang, X.; Li, Y.; Lv, Z. Analysis of 22-year Drought Characteristics in Heilongjiang Province Based on Temperature Vegetation Drought Index. Computational intelligence and neuroscience. 2022, 1003243. [Google Scholar] [CrossRef]
- Shah, S.; Tiwari, A.; Song, X.; Talchabahdel, R.; Habiyakare, T.; Adhikari, A. Drought index predictability for historical and future periods across the Southern plain of Nepal Himalaya. Environ Monit Assess. 2022, 194, 642. [Google Scholar] [CrossRef]
- Sun, X.; Xiong, H.; Jiang, C.; Zhang, D.; Yang, Z.; Huang, Y.; Zhu, W.; Ma, S.; Duan, J.; Wang, X.; Liu, W.; Guo, H.; Li, G.; Qi, J.; Liang, C.; Zhang, Z.; Li, J.; Zhang, H.; Han, L.; Zhou, Y.; Peng, Y.; Li, Z. Natural variation of DROT1 confers drought adaptation in upland rice. Nat Commun. 2022, 13, 4265. [Google Scholar] [CrossRef]
- Ren, XL.; Zhang, P.; Chen, X.; Jia, ZK. Impacts of ridge-furrow rainfall concentration systems and mulches on corn growth and yield in the semiarid region of China. Journal of the Science of Food and Agriculture. 2016, 96, 3882–3889. [Google Scholar] [CrossRef]
- Robinson, D.; Jones, S.; Lebron, I.; Reinsch, S.; Domínguez, M.; Smith, A.; Jones, D.; Marshall, M.; Emmett, B. Experimental evidence for drought induced alternative stable states of soil moisture. Scientific Reports. 2016, 6, 20018. [Google Scholar] [CrossRef]
- Wang, J.; Fu, B. Z.; Li, S. X.; Wang, X.; Song, W. X.; Ye, Y. N.; Hu, P. F.; Wang, T. R. Effects of exogenous melatonin on growth and physiological characteristics of Agropyron mongolicum seedlings under drought stress. The journal of applied ecology. 2023, 34, 2947–2957. [Google Scholar]
- Zhao, M.; Li, J.; Shi, X.; Sanaullah, M.; Quan, Y.; Guo, D.; Wang, L.; Wang, S. Effects of exogenous plant regulators on growth and development of "Kyoho" grape under salt alkali stress. Frontiers in Plant Science. 2023, 14, 1274684. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.; Tang, Y.; Yuan, Y.; Sun, J.; Zhong, M.; Guo, S. The role of 24-epibrassinolide in the regulation of photosynthetic characteristics and nitrogen metabolism of tomato seedlings under a combined low temperature and weak light stress. Plant Physiology Biochemistry. 2016, 107, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Ma, X.; Hussain, Q.; Asim, M.; Iqbal, A.; Ren, X.; Shah, S.; Chen, K.; Shi, Y. Application of 2,4-Epibrassinolide Improves Drought Tolerance in Tobacco through Physiological and Biochemical Mechanisms. Biology (Basel). 2022, 11, 1192. [Google Scholar] [CrossRef]
- Chen, Y.; Yao, Z.; Sun, Y.; Wang, E.; Tian, C.; Sun, Y.; Liu, J.; Sun, C.; Tian, L. Current Studies of the Effects of Drought Stress on Root Exudates and Rhizosphere Microbiomes of Crop Plant Species. Int. J. Mol. Sci. 2022, 23, 2374. [Google Scholar] [CrossRef]
- Guo, C.; Zhang, K.; Sun, H.; Zhu, L.; Zhang, Y.; Wang, G.; Li, A.; Bai, Z.; Liu, L.; Li, C. Root Cortical Senescence Enhances Drought Tolerance in Cotton. Plant, cell & environment 2024. [Google Scholar] [CrossRef]
- Xu, L.; He, J.; Meng, Y.; Zheng, Y.; Lu, B.; Zhang, J.; Zhou, Y. Enhancing drought resistance in Pinus tabuliformis seedlings through root symbiotic fungi inoculation. Frontiers in Plant Science. 2024, 15, 1446437. [Google Scholar] [CrossRef]
- Javed, J.; Rauf, M.; Arif, M.; Hamayun, M.; Gul, H.; Ud-Din, A.; Ud-Din, J.; Sohail, M.; Rahman, M.; Lee, I. Endophytic Fungal Consortia Enhance Basal Drought-Tolerance in Moringa oleifera by Upregulating the Antioxidant Enzyme (APX) through Heat Shock Factors. Antioxidants (Basel). 2022, 11, 1669. [Google Scholar] [CrossRef] [PubMed]
- Santra, H.; Banerjee, D. Production, Optimization, Characterization and Drought Stress Resistance by β-Glucan-Rich Heteropolysaccharide From an Endophytic Fungi Colletotrichum alatae LCS1 Isolated From Clubmoss (Lycopodium clavatum). Front Fungal Biology. 2022, 2, 796010. [Google Scholar] [CrossRef]
- Zhu, S.; Feng, X.; Liu, Y.; Jin, D.; Luo, X.; Fan, Y. Expression of a viral ecdysteroid UDP-glucosyltransferase enhanced the insecticidal activity of the insect pathogenic fungus Beauveria bassiana. Pest Manag Sci. 2024, 80, 4915–4923. [Google Scholar] [CrossRef]
- Basso, V.; Pinheiro, D.; Toldi, M.; Gonçalves, K.; Vicenço, B. Beauveria bassiana submerged spores for control of two-spotted spider mite (Tetranychus urticae Koch (Acari: Tetranychidae)): production, stability, and virulence. Archieves of Microbiology. 2023, 206, 23. [Google Scholar] [CrossRef]
- Aak, A.; Hage, M.; Rukke, B. Biological control of Cimex lectularius with Beauveria bassiana: Effects of substrate, dosage, application strategy, and bed bug physiology. Pest Management Science. 2023, 79, 4599–4606. [Google Scholar] [CrossRef] [PubMed]
- Daza, F.; Roman, G.; Rodriguez, M.; Vargas, I.; Heano, H.; Cereda, M.; Mulet, R. Spores of Beauveria bassiana and Trichoderma lignorum as a bioinsecticide for the control of Atta cephalotes. Biological Research. 2019, 52, 51. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, Y.; Wang, B. Entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae play roles of maize (Zea mays) growth promoter. Scientific Reports. 2022, 12, 15706. [Google Scholar] [CrossRef] [PubMed]
- Raad, M.; Glare, T.; Brochero, H.; Müller, C.; Rostás, M. Transcriptional Reprogramming of Arabidopsis thaliana DefencePathways by the Entomopathogen Beauveria bassiana Correlates With Resistance Against a Fungal Pathogen but Not Against Insects. Frontiers Microbiology. 2019, 10, 615. [Google Scholar]
- Sui, L.; Lu, Y.; Xu, M.; Liu, J.; Zhao, Y.; Li, Q.; Zhang, Z. Insect hypovirulence-associated mycovirus confers entomopathogenic fungi with enhanced resistance against phytopathogens. Virulence. 2024, 15, 2401978. [Google Scholar] [CrossRef]
- Idrees, A.; Afzal, A.; Qadir, Z.; Li, J. Bioassays of Beauveria bassiana Isolates against the Fall Armyworm, Spodoptera frugiperda. Journal of Fungi(Basel). 2022, 8, 717. [Google Scholar] [CrossRef]
- Sui, L.; Lu, Y.; Zhu, H.; Wan, T.; Li, Q.; Zhang, Z. Endophytic blastospores of Beauveria bassiana provide high resistance against plant disease caused by Botrytis cinerea. Fungal Biol. 2022, 126, 528–533. [Google Scholar] [CrossRef]
- Galmés, J.; Medrano, H.; Flexas, J. Photosynthetic limitations in response to water stress and recovery in Mediterranean plants with different growth forms. New phytologist. 2007, 175, 81–93. [Google Scholar] [CrossRef]
- Qin, X.; Zhao, X.; Huang, S.; Deng, J.; Li, X.; Luo, Z.; Zhang, Y. Pest management via endophytic colonization of tobacco seedlings by the insect fungal pathogen Beauveria bassiana. Pest Manag Sci. 2021, 77, 2007–2018. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. Journal of experimental botany. 2008, 59, 3317–3325. [Google Scholar] [CrossRef]
- Zu, X.; Lu, Y.; Wang, Q.; et al. Increased Drought Resistance 1 Mutation Increases Drought Tolerance of Upland Rice by Altering Physiological and Morphological Traits and Limiting ROS Levels. Plant Cell Physiol. 2021, 62, 1168–1184. [Google Scholar] [CrossRef] [PubMed]
- Živanović, B.; Milić Komić, S.; Tosti, T.; Vidović, M.; Prokić, L.; Veljović, J. Leaf Soluble Sugars and Free Amino Acids as Important Components of Abscisic Acid-Mediated Drought Response in Tomato Plants. Plants(Basel). 2020, 9, 1147. [Google Scholar]
- Zou, Y. N. , Wu, Q. S., Huang, Y. M., Ni, Q. D., He, X. H. Mycorrhizal-mediated lower proline accumulation in Poncirus trifoliata under water deficit derives from the integration of inhibition of proline synthesis with increase of proline degradation. PloS one. 2013, 8, e80568. [Google Scholar] [CrossRef]
- Gurrieri, L.; Merico, M.; Trost, P.; Forlani, G.; Sparla, F. Impact of Drought on Soluble Sugars and Free Proline Content in Selected Arabidopsis Mutants. Biology (Basel). 2020, 9, 367. [Google Scholar] [CrossRef]
- Pouris, J.; Levizou, E.; Karatassiou, M.; Meletiou-Christou, M.; Rhizopoulou, S. The Influence of the Partitioning of Sugars, Starch, and Free Proline in Various Organs of Cyclamen graecum on the Biology of the Species and Its Resistance to Abiotic Stressors. Plants (Basel.). 2022, 11, 1254. [Google Scholar] [CrossRef]
- Tu, Y.; Jiang, A.; Gan, L.; Hossain, M.; Zhang, J.; Peng, B.; Xiong, Y.; Song, Z.; Cai, D.; Xu, W.; Zhang, J.; He, Y. Genome duplication improves rice root resistance to salt stress. Rice. 2014, 7, 15. [Google Scholar] [CrossRef]
- Zhang, M.; Cui, G.; Bai, X.; Ye, Z.; Zhang, S.; Xie, K.; Sun, F.; Zhang, C.; Xi, Y. Regulatory Network of Preharvest Sprouting Resistance Revealed by Integrative Analysis of mRNA, Noncoding RNA, and DNA Methylation in Wheat. Journal of agricultural and food chemistry. 2021, 69, 4018–4035. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fu, B.; Lin, Q.; Riley, I. .; Ding, S.; Chen, L.; Cui, J.; Yang, L.; Li, H. Colonization of Beauveria bassiana 08F04 in root-zone soil and its biocontrol of cereal cyst nematode (Heterodera filipjevi). PLoS One 2020, 15, e0232770. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Javed, R.; Adeel, M.; Rizwan, M.; Yang, Y. PEG 6000-Stimulated Drought Stress Improves the Attributes of In Vitro Growth, Steviol Glycosides Production, and Antioxidant Activities in Stevia rebaudiana Bertoni. Plants (Basel). 2020, 9, 1552. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhang, Y.; Li, Z.; Dong, F.; Chen, L. Two grasses differ in their absorptive root physiological traits and rooting depth under drought in an alpine steppe. Annals of botany. 2024, mcae151. [Google Scholar] [CrossRef]
- Jordan, W.; Ritchie, J. Influence of soil water stress on evaporation, root absorption, and internal water status of cotton. Plant Physiol. 1971, 48, 783. [Google Scholar] [CrossRef]
- Vidović, M.; Ćuković, K. Isolation of high-quality RNA from recalcitrant leaves of variegated and resurrection plants. 3 Biotech. 2020, 10, 286. [Google Scholar] [CrossRef]
- Zhou, R.; Song, Y.; Xue, X.; Xue, R.; Jiang, H.; Zhou, Y.; Qi, X.; Wang, Y. Differential Transcription Profiling Reveals the MicroRNAs Involved in Alleviating Damage to Photosynthesis under Drought Stress during the Grain Filling Stage in Wheat. International journal of molecular sciences. 2024, 25, 5518. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jin, M.; Xu, L.; Xi, H.; Wang, B.; Du, S.; Liu, H.; Wen, Y. Effects of ketoprofen on rice seedlings: Insights from photosynthesis, antioxidative stress, gene expression patterns, and integrated biomarker response analysis. Environmental pollution (Barking, Essex : 1987). 2020, 263, 114533. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, J.; Han, Z.; Han, Z.; Li, S.; Zhang, J.; Ma, H.; Han, Y. Screening of differentially expressed microRNAs and target genes in two potato varieties under nitrogen stress. BMC plant biology. 2022, 22, 478. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Meng, C.; Yuan, Y.; Nath, U. K.; Zhao, Y.; Wang, Z.; Yang, S.; Li, L.; Niu, L.; Yao, Q.; Wei, F.; Zhang, X. CaPSY1 gene plays likely the key role in carotenoid metabolism of pepper (Capsicum annuum) at ripening. Functional plant biology : FPB. 2021, 48, 141–155. [Google Scholar] [CrossRef]
- Park, M.; Lee, H.; Kim, I.; Kim, H. Characterization of fab2 T-DNA insertion mutants in terms of fatty acid composition and plant phenotype. Plant signaling & behavior. 2023, 18, 2213937. [Google Scholar]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods (San Diego, Calif.). 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Liu, X.; Dong, X.; Liu, Z.; Shi, Z.; Jiang, Y.; Qi, M.; Xu, T.; Li, T. Repression of ARF10 by microRNA160 plays an important role in the mediation of leaf water loss. Plant molecular biology. 2016, 92, 313–336. [Google Scholar] [CrossRef]
- Egea, I.; Albaladejo, I.; Meco, V.; Morales, B.; Sevilla, A.; Bolarin, M.; Flores, F. The drought-tolerant Solanum pennellii regulates leaf water loss and induces genes involved in amino acid and ethylene/jasmonate metabolism under dehydration. Scientific reports. 2018, 8, 2791. [Google Scholar] [CrossRef]
- Bokhale, M.; Mwaba, I.; Allie, F. Real-time PCR data for reference candidate gene selection in tomato infected with Tomato curly stunt virus. Data in Brief. 2020, 31, 105750. [Google Scholar] [CrossRef] [PubMed]
- Jensen, N.; Ottosen, C.; Zhou, R. Exogenous Melatonin Alters Stomatal Regulation in Tomato Seedlings Subjected to Combined Heat and Drought Stress through Mechanisms Distinct from ABA Signaling. Plants (Basel). 2023, 12, 1156. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of B. bassiana colonization on the phenotype of hydroponically grown tomato seedlings at different stages of drought. Seedlings treated with (a) PEG-6000 for 12 h, (b) PEG-6000 for 24 h, and (c) PEG-6000 for 36 h. From left to right are Control, Bb, GH, GB treatment. Control seedlings were placed in Hoagland nutrient solution after root irrigation with 0.05% (v/v) Tween-80. Bb seedlings were placed in Hoagland nutrient solution after root irrigation with B. bassiana. PEG seedlings were placed in Hoagland nutrient solution containing 8%PEG-6000 after root irrigation with 0.05% (v/v) Tween-80 irrigation. PB seedlings were placed in Hoagland nutrient solution containing 8%PEG-6000 after root irrigation with B. bassiana.
Figure 1.
Effect of B. bassiana colonization on the phenotype of hydroponically grown tomato seedlings at different stages of drought. Seedlings treated with (a) PEG-6000 for 12 h, (b) PEG-6000 for 24 h, and (c) PEG-6000 for 36 h. From left to right are Control, Bb, GH, GB treatment. Control seedlings were placed in Hoagland nutrient solution after root irrigation with 0.05% (v/v) Tween-80. Bb seedlings were placed in Hoagland nutrient solution after root irrigation with B. bassiana. PEG seedlings were placed in Hoagland nutrient solution containing 8%PEG-6000 after root irrigation with 0.05% (v/v) Tween-80 irrigation. PB seedlings were placed in Hoagland nutrient solution containing 8%PEG-6000 after root irrigation with B. bassiana.

Figure 2.
Effect of B. bassiana colonization on the water absorption ability of tomato seedlings. (a) effect of B. bassiana colonization on water absorption 4, 8 and 12 days after drought treatment; (b) phenotypic effects of B. bassiana colonization on water absorption 12 days after drought treatment. Means represent the average of all replications. p values indicate a significant difference based on one-way ANOVA, respectively.
Figure 2.
Effect of B. bassiana colonization on the water absorption ability of tomato seedlings. (a) effect of B. bassiana colonization on water absorption 4, 8 and 12 days after drought treatment; (b) phenotypic effects of B. bassiana colonization on water absorption 12 days after drought treatment. Means represent the average of all replications. p values indicate a significant difference based on one-way ANOVA, respectively.
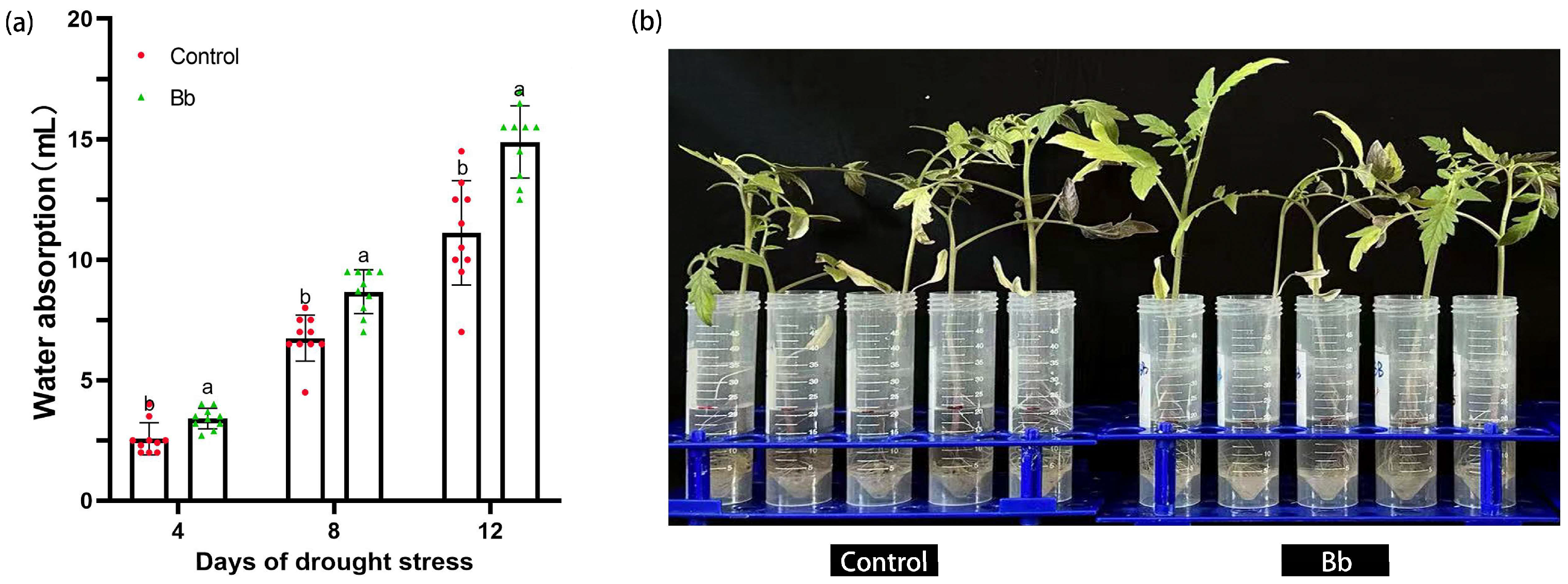
Figure 3.
Effects of B. bassiana on growth phenotypes, plant height, stem diameter, root length, fresh and dry weights, and the relative water content of the tomato seedlings at different stages of drought stress. (a) effects on overall growth after 1, 6, 12 and 18 days of drought stress. Control plants were root irrigated with 0.05% (v/v) Tween-80 root; Bb plants were root irrigated with B. bassiana; GH plants were treated with natural drought after root irrigation with 0.05% (v/v) Tween-80; GB plants were root irrigated with B. bassiana then subjected to natural drought treatment. (b) tomato seedling height, and (c) stem diameter at different stages of drought stress, (d)the root length, (e) fresh weight, (f) dry weight and (g) relative water content on day 18 of drought stress. Means represent the average of all replications. p values indicate a significant difference using one-way ANOVA, respectively.
Figure 3.
Effects of B. bassiana on growth phenotypes, plant height, stem diameter, root length, fresh and dry weights, and the relative water content of the tomato seedlings at different stages of drought stress. (a) effects on overall growth after 1, 6, 12 and 18 days of drought stress. Control plants were root irrigated with 0.05% (v/v) Tween-80 root; Bb plants were root irrigated with B. bassiana; GH plants were treated with natural drought after root irrigation with 0.05% (v/v) Tween-80; GB plants were root irrigated with B. bassiana then subjected to natural drought treatment. (b) tomato seedling height, and (c) stem diameter at different stages of drought stress, (d)the root length, (e) fresh weight, (f) dry weight and (g) relative water content on day 18 of drought stress. Means represent the average of all replications. p values indicate a significant difference using one-way ANOVA, respectively.
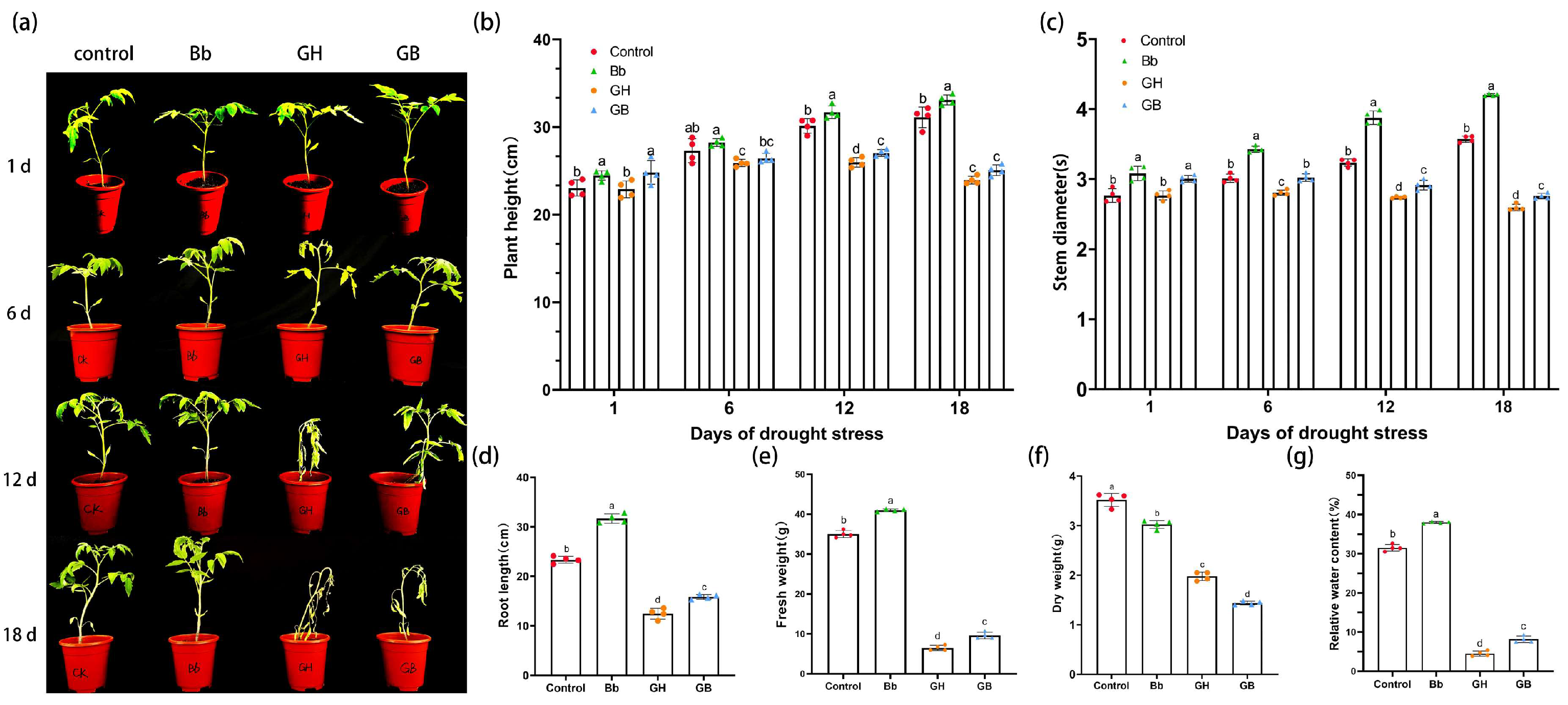
Figure 4.
Changes in visual field with 40×and stomatal length, width, and number in tomato leaves on day 12 of drought stress in each treatment. (a) changes in stomatal number under microscope 20×field of view and (b) stomatal size under microscope 20×field of view on day 12 of drought stress. (c) effects of colonization of B. bassiana on stomatal number(d) stomatal length and (e) stomatal width on day 12 of drought stress in each treatment. Treatments are as described in Figure 2. Means represent the average of all replications. p values indicate a significant difference using one-way ANOVA, respectively.
Figure 4.
Changes in visual field with 40×and stomatal length, width, and number in tomato leaves on day 12 of drought stress in each treatment. (a) changes in stomatal number under microscope 20×field of view and (b) stomatal size under microscope 20×field of view on day 12 of drought stress. (c) effects of colonization of B. bassiana on stomatal number(d) stomatal length and (e) stomatal width on day 12 of drought stress in each treatment. Treatments are as described in Figure 2. Means represent the average of all replications. p values indicate a significant difference using one-way ANOVA, respectively.
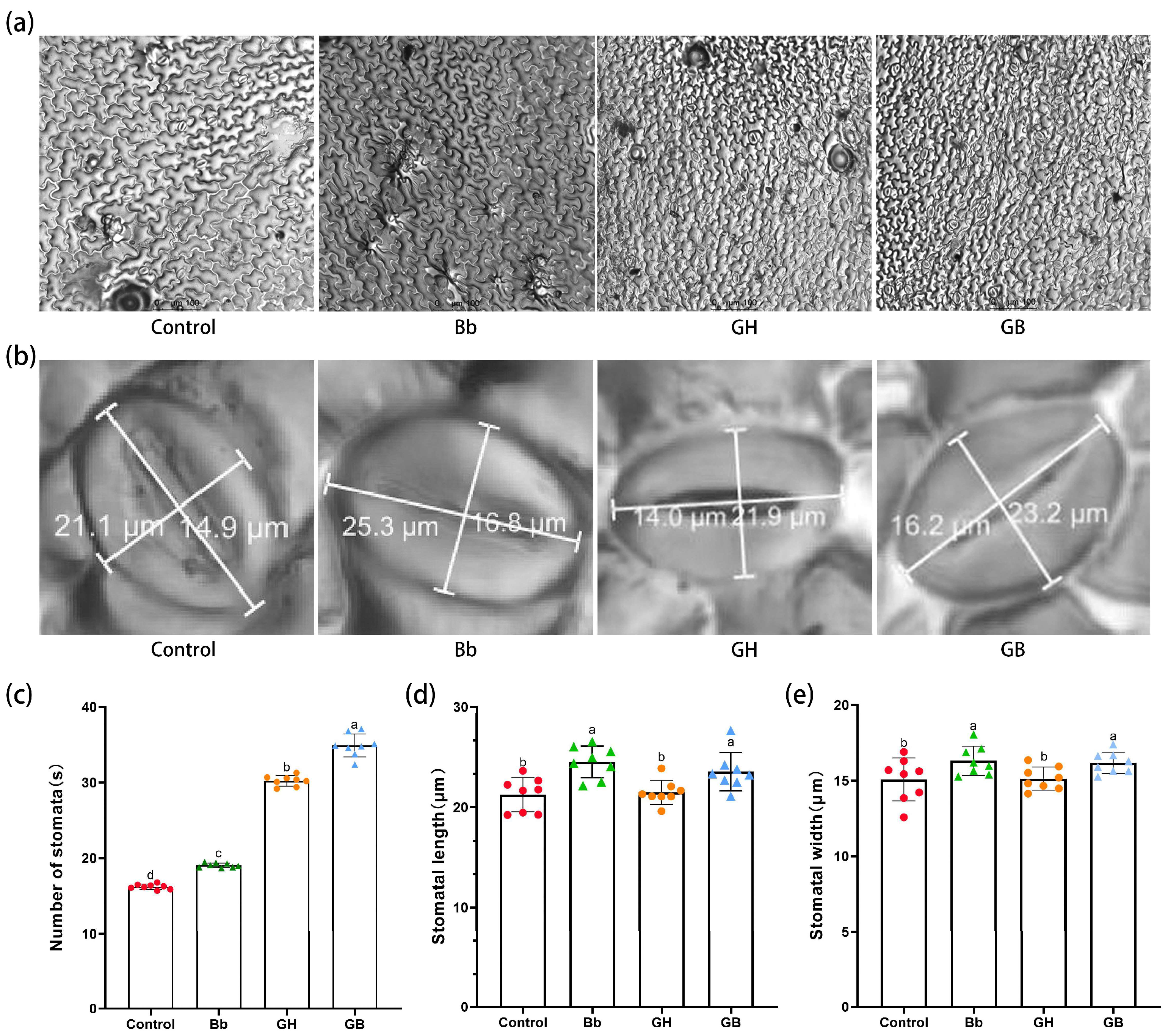
Figure 5.
Effect of colonization of B. bassiana on (a) POD, (b) PRO and (c) MDA activity in tomato seedlings at different stages of drought stress. Treatments are as described in Figure 2. Means represent the average of all replications. p values indicate a significant difference usin ANOVA, respectively.
Figure 5.
Effect of colonization of B. bassiana on (a) POD, (b) PRO and (c) MDA activity in tomato seedlings at different stages of drought stress. Treatments are as described in Figure 2. Means represent the average of all replications. p values indicate a significant difference usin ANOVA, respectively.
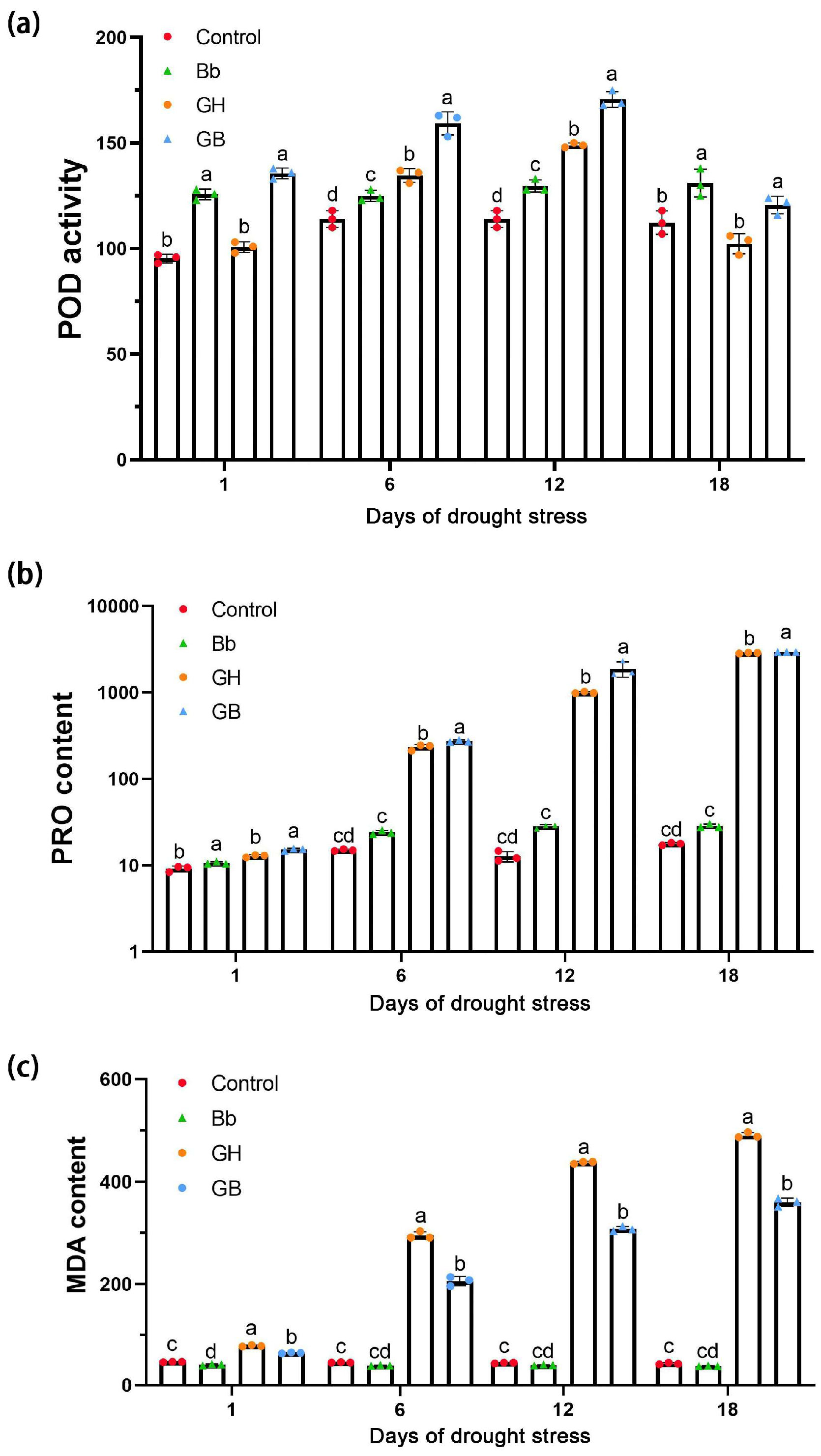
Figure 6.
Expression levels of eight key genes related to stomatal development and drought resistance in tomato seedlings at different stages of drought stress. Treatments are as described in Figure 2. Values were determined using qRT-PCR.
Figure 6.
Expression levels of eight key genes related to stomatal development and drought resistance in tomato seedlings at different stages of drought stress. Treatments are as described in Figure 2. Values were determined using qRT-PCR.
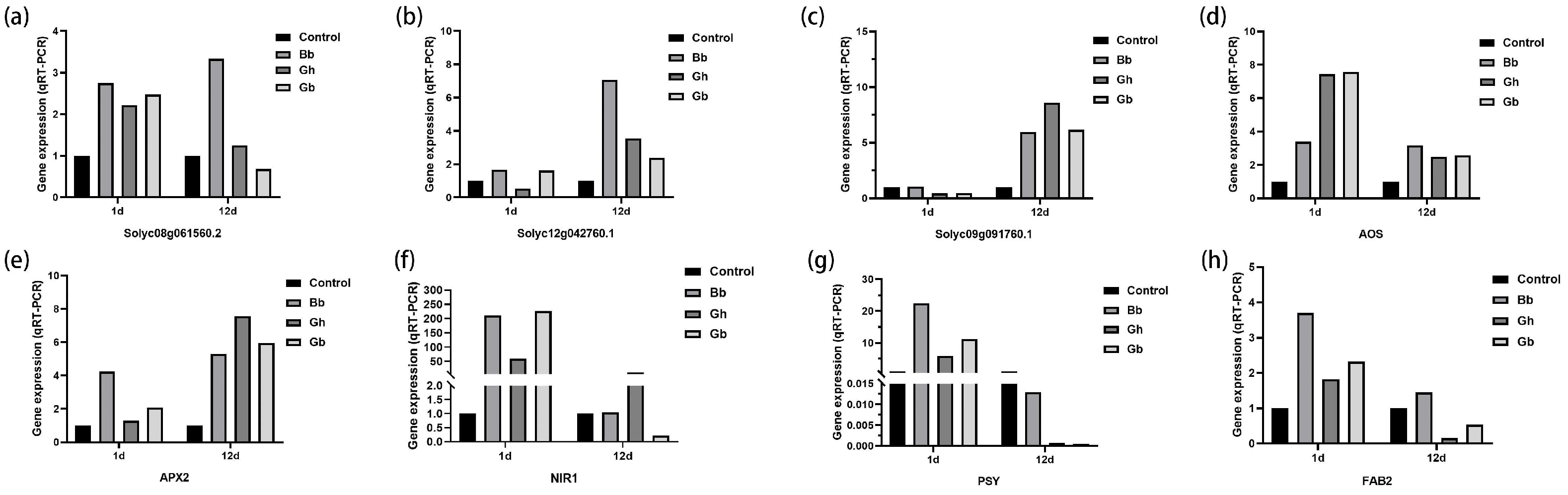
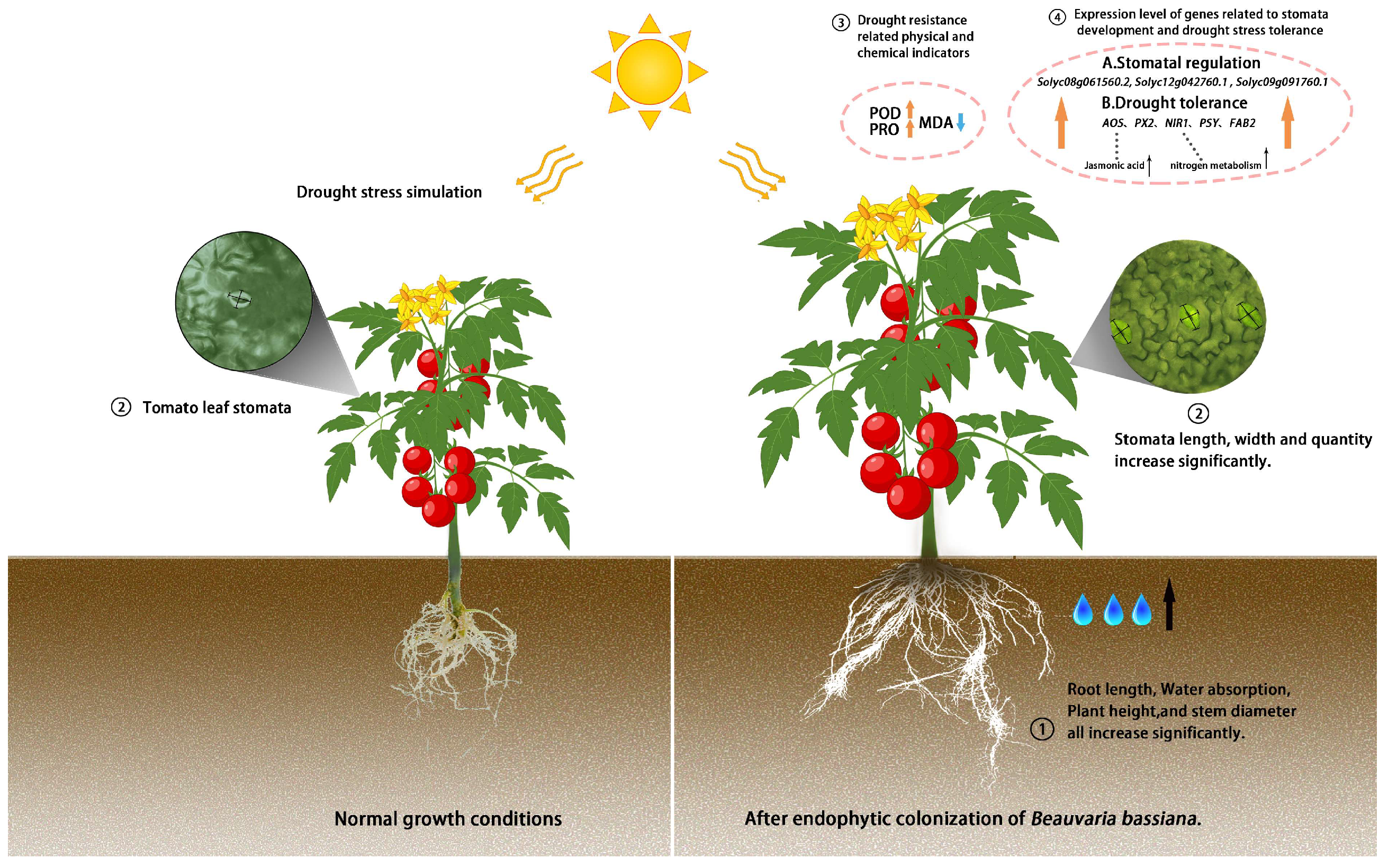
Figure 7.
Schematic representation of the effect of endophytic colonization of B. bassiana on drought resistance in tomato via the "water spender" pathway.
Figure 7.
Schematic representation of the effect of endophytic colonization of B. bassiana on drought resistance in tomato via the "water spender" pathway.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



