
Submitted:
16 October 2024
Posted:
17 October 2024
You are already at the latest version
Abstract
Cadmium (Cd) is a toxic metal primarily found as a by-product of zinc production. Cd was a proven carcinogen, and exposure to this metal has been linked to various adverse health effects, which were first reported in the mid-19th century and thoroughly investigated by the 20th century. The toxicokinetics and dynamics of Cd reveal its propensity for long biological retention and predominant storage in soft tissues. Cd pollution, primarily from industrial activities, strongly contaminates soil and water and affects human health and ecosystems. Cd enters the human body mainly through ingestion and inhalation, with food and tobacco smoke being the primary sources. It accumulates in various organs, particularly the kidney and liver, and is known to cause severe health problems, including renal dysfunction, bone diseases, cardiovascular problems, and many others. On a cellular level, Cd disrupts numerous biological processes, inducing oxidative stress generation and DNA damage. This comprehensive review explores Cd pollution, accumulation, distribution, and biological impacts on bacteria, fungi, edible mushrooms, plants, animals, and humans on a molecular level. Molecular aspects of carcinogenesis, apoptosis, autophagy, specific gene expression, stress protein synthesis, and ROS formation caused by Cd were discussed as well. This paper also summarizes how Cd is removed from contaminated environments and the human body.
Keywords:
cadmium toxicity
; pollution sources
; toxicity on the human body
; biological impact
; bacteria
; fungi
; edible mushrooms
; plants
; animals
; molecular mechanisms
; Cd removal
1. Introduction
Cadmium (Cd, atomic number: 48, atomic mass: 112.4, period number: 5, Group 12, electron configuration: 4d105s2, isotopes: 7) was discovered in 1817 by the German chemist Friedrich Stromeyer. This chemical element was named after one of the first heroes in Greek mythology, Cadmus, the legendary founder of Thebes, known for his immense strength before Heracles. The Latin word cadmia and the Greek word καδμεία, which was once used to refer to the standard zinc (Zn) ore calamine, are the sources of the name [1]. Cd was not known to be a separate element until 1817, as in nature, Cd is always found in the presence of zinc. Cd and zinc are chemically and physically similar. Cd is a by-product of zinc and lead production. The Cd/Zn ratio is 0.5% in nonferrous ores, with the actual recovery of cadmium metal estimated at 50-65% of that present in raw ore [2]. Cd is unique among metals due to its diverse toxic effects, long biological half-life, low rate of excretion, and predominant storage in soft tissues rather than bone [3]. In Table 1 are described some of Cd’s physical and chemical properties.
Cd was found to have its initial negative health consequences in 1858. Those who used polishing agents containing Cd suffered from respiratory and gastrointestinal problems. The earliest toxicological experiments were conducted in 1919. In humans, emphysema and proteinuria were first documented in the 1940s in workers exposed to Cd dust [3]. Following World War II, a bone ailment known as Itai-Itai disease—a type of Cd-induced renal osteomalacia—was reported in Japan, causing fractures and excruciating agony [3]. The toxicokinetics and toxicodynamics of Cd were then discussed, along with how it binds to the metallothionein protein [5]. In the 1970s, health advisories regarding the dangers of Cd pollution were distributed globally [6]. Early research was done on the impacts on reproduction and cancer, but there is still much doubt about a quantitative evaluation of these adverse effects in humans. The World Health Organization's International Program on Chemical Safety recognized renal failure as a critical consequence of Cd and offered a rough quantitative evaluation. In the 1990s and 2000s, various epidemiological studies found adverse health impacts in demographic groups in Japan, China, Europe, and the United States, sometimes with minimal ambient Cd exposure. The early discovery of metallothionein's important role in Cd toxicology laid the groundwork for recent studies that used biomarkers of susceptibility to the development of Cd-related renal dysfunction, such as metallothionein gene expression in peripheral lymphocytes and metallothionein autoantibodies in blood plasma [7].
2. The Origin of Cd Pollution
Cd is an environmental contaminant classified eighth on the Top 20 Hazardous Substances Priority List because of its high toxicity and sluggish metabolism [8]. The primary source of Cd is stack dust, produced during zinc purification by distillation and deposited in all fractions [9]. Plants can absorb Cd directly on the soil [10]. Phosphate fertilizers and atmospheric deposition have been the primary sources of Cd intake into soils. Phosphorite and apatite rocks mainly used in producing of phosphate fertilizers contain Cd and several other heavy metals [11]. The amount of Cd accumulated in soil due to environmental contamination depends on the magnitude of emissions, transit, and retention. The fate of heavy metal contaminants in soil is mainly determined by the balance of sorption, leaching, and plant uptake. Soil variables like pH, redox state, organic matter, clay, hydrous oxides, and free carbonates significantly impact these processes. Metal destiny varies significantly amongst soil types, including forest and heavily developed agricultural land [12]. Global production and use of Cd are significant; for instance, Cd pigments consumption surpasses 2500 tons annually [13]. For thousands of years, it has also been used as a pigment due to its ability to produce almost all the rainbow colors: brilliant yellow, orange, and red, interacting with other chemical elements. Thus, a bright orange color is made when the bright yellow pigment cadmium sulfide (CdS) is mixed with cadmium selenite (CdSe). Cadmium sulphoselenide (Cd2SSe) creates a pigment called Cadmium red. The pigments mentioned above are still used nowadays as plastic colorants. In principle, Cd is part of various chemical compounds as a divalent cation [14]. As a result, millions of people are in danger of exposure to mercury pollution due to purposeful or accidental cadmium releases into the environment. Human activities associated with Cd emissions include industrial production, crop farming, animal breeding, aquaculture, and wastewater treatment [15]. Low amounts of Cd are naturally found in the lithosphere (0.15 mg/kg in the Earth's crust and 1.1 × 10−4 mg/L in seawater) [16]. Still, various industrial processes, including mining and smelting, have increased the element's availability in the environment and increased human exposure to it. Thousands of tons of garbage polluted with Cd are dumped into the environment globally each year [16]. In Figure 1 is presented the distribution of heavy metals in the environment.
Jinding lead-zinc mine is currently the largest lead-zinc deposit proven in China, one of only a few with over 10-million-ton reserves worldwide. Its mining area is 6.8 km2, with more than 80% open-pit mining. The large-scale mining of the Jinding Pb-Zn deposit began in the 1980s, causing significant environmental problems [18]. Descriptive statistics revealed that 1.1% As, 7.3% Cd, 0.3% Pb, and 0.2% Hg contents exceeded China's Soil Environmental Quality Management Standard (GB 15618-2018, in Chinese). Furthermore, 32.8% of As, 74.4% of Cd, 89.2% of Pb, 45.0% of Cr, and 13.7% of Hg concentrations in soil samples exceeded the background soil concentrations of heavy metals in this location, with Cd and Pb having highest levels that were 11.64- and 21.47-fold the background values. On the other hand, compared to regular farming areas, industrial and mining enterprises, sewage irrigation and urban samples from those areas had considerably greater concentrations of As, Cd, Pb, Cr, or Hg in their soil [19]. The results of principal component (PC) and geostatistical studies showed that PC was highly associated with As and Cr and influenced mainly by natural factors [19]. Surface waters, especially the lakes are a significant global source of freshwater, and Cd pollution of water bodies is becoming more of an issue as industry, agriculture, and other human activities advance [20]. Cd pollution in lakes poses a significant risk to water quality, drinking water supplies, the food chain, and freshwater ecosystems. Sediments have been observed to act as both a sink and a source of Cd in water bodies. Yearlong monitoring in Meiliang Bay (northern part of Taihu Lake, National Wetland Park, eastern China) revealed that the mobility of Cd in sediments varied widely and significantly impacted the Cd pollution level in the overlying water [8]. Tourism has been shown to contribute to Cd pollution (primarily through hotel wastewater and increased traffic) and vice versa. Cd pollution of beaches, coastal waterways, food, urban parks, and other areas poses risks to tourists and increases human exposure to this poisonous metal [21].
2.1. Natural Sources of Cd
2.1.1. Cd in Soil Water and Groundwater
Generally, Cd concentration in the earth is around 0.1–0.5 ppm, and this metal mainly accumulates in sedimentary rocks. Natural activities such as erosion, weathering of rocks, volcano eruptions, and wildfires release large amounts of Cd into soils and rivers, respectively, seas and oceans. Phosphorites and marine phosphates contain high amounts of Cd, as much as 500 ppm. Maximum permissible Cd concentrations are 5 μg/g in soil and water and 1 μg/L in groundwater [20]. Large quantities of Cd (about 15,000 mt (metric tons) are transported into rivers by erosion and weathering of rock materials. Volcanic eruptions release about 820 metric tons of Cd, while forest fires release up to 70 metric tons [22]. Detailed information on Cd distribution in European top soils is available on the European Commission's official webpage: https://esdac.jrc.ec.europa.eu/search/node/cadmium [23]. As seen from Figure 2, out of the total, 72.6% of the samples have Cd values <0.07 mg kg−1, 21.6% in the range 0.07–1 mg kg−1, and the remaining 5.5% higher than the threshold of 1 mg kg−1, which is generally considered the limit for risk assessment [24].
Data about all heavy metal concentrations is permanently deposited at the European Soil Data Centre (ESDAC), esdac.jrc.ec.europa. EU, European Commission, Joint Research Centre.
The quality of surface water in Europe is regulated by the Directive 2000/60/EC. There are 100,000 surface water bodies in Europe, but only 40% are in good condition. As Kubier et al. [21] highlighted, WHO Guidelines for Drinking Water Quality recommend a value of up to 3 μg/L.
Groundwater in Pakistan has typical Cd contents of 10 μg/L from Jurassic sulfide-bearing sedimentary rocks. In contrast, in Germany, groundwater concentrations of Cd range from 0.11 μg/L in loess aquifers beneath arable land to 2.7 μg/L in sandy aquifers beneath forested regions [21].
A comparison of crucial aquifer systems reveals a link between rock type, groundwater environment, and Cd contents. Cd 90th percentile background levels in groundwater varied from less than 0.1 μg/L in Paleozoic, Triassic, and Jurassic aquifers to more than 1 μg/L in Rotliegend, Cretaceous, and Cenozoic aquifers. Aside from calcium carbonate Cretaceous aquifers, limestone-dominated aquifer basins exhibited low Cd levels in groundwater. Most of them are in alkaline aquifer systems [21]. Cd concentrations above 1 μg/L were found in groundwater in sandstone aquifers and unconsolidated sand and gravel aquifer systems in the western United States. However, in most samples collected from 3124 wells in the US Cd was below 1 μg/L.
Groundwater, but from a glacial aquifer system in the United States had Cd values ranging from 0.018 μg/L to 1.0 μg/L. However, 84% of groundwater samples (N = 847) were below the detection limit. A survey of groundwater near garbage sites in the United States revealed Cd values of up to 6000 μg/L. Municipal solid waste dumps in the European Union can produce leachates with Cd values up to 2700 μg/L. As a result, Cd concentrations that exceed the natural background can be caused by both natural and anthropogenic mechanisms [21].
2.1.2. Air
Natural Cd emissions come mainly from rock weathering, airborne soil particles from deserts, sea spray, forest fires, biogenic material, volcanoes, and hydrothermal vents. In Europe, the leading standard regulating the Cd concentration in ambient air is the Directive 2008/50/EC [25]. According to it, the maximum permission level is 5 ng/m3, valid since 2013 [25]. In the USA, this restriction is even more substantial, and the Regulatory bodies such as NAAQS (National Ambient Air Quality Standards), USEPA (United States Environmental Protection Energy Agency), and NIOSH (National Institute of Occupational Safety and Health) limited Cd in the air to 200 pg/m3. It is known that PM2.5 can transport heavy metals such as Cd, Hg, Pb, Cr, and Mn. In a research conducted in Beijing, China, the immediate impacts of PM2.5 exposure on blood Cd levels were studied. The findings revealed that the average blood Cd concentration was 0.64 μg/L. A significant correlation was observed between PM2.5 exposure and blood Cd level (P < 0.05) as authors reported [26].
Soil particles are the most common source of natural emissions into the atmosphere, followed by forest and bushfires, sea salt, volcanic emissions, and meteoric dust [27]. Wildfires increase Cd concentrations in soils and ashes. Long-term behavior analysis revealed decreasing Cd concentrations in the solid phase, as rainfall and pH decrease with time following fire, resulting in desorption and mobility of Cd and other heavy metals [21]. Wildfires in California, for example, raised the average Cd concentration in the runoff by more than two orders of magnitude. Cd concentrations in biomass ash can reach 30 mg/kg, providing an additional method for increasing Cd concentrations in soil because such ash is commonly used as fertilizer. In the short term, the bioavailable pool of Cd remains low due to an ash-induced pH increase, but as pH rises due to rainfall, Cd bioavailability increases [21].
2.2. Anthropogenic Cd Sources
Anthropogenic Cd inputs into soil, groundwater, and atmosphere come from mining, nonferrous metal manufacturing, fossil fuel combustion, phosphate fertilizer and pesticide manufacturing, iron, steel, cement production, road dust, plastics production, wildfires, and municipal and sewage sludge incineration. Environmental pollution with Cd stems from its widespread use in the production of alloys and batteries, as a pigment in plastics, paints, and ceramics, and corrosion-resistant coatings of metal products. This heavy metal is primarily found and abundant in lead, copper, and zinc ores. Over the past 50 years, anthropogenic Cd emissions have fallen by more than 90% [28]. Like uranium (U), using phosphate fertilizers with Cd as an impurity is a common cause of high Cd concentrations in soil and groundwater. This Cd addition pathway to groundwater was explored in the United States, Canada, Britain, Norway, Sweden, Finland, Denmark, Germany, Australia, and New Zealand. The findings indicate that P fertilizer application alters soil chemistry. Furthermore, Cd can enter the food chain and be hazardous to living organisms. Cd sources can be both local and diffuse [29].
Local sources such as mines, industrial sites, and abandoned mining deposits cause increased Cd concentrations, albeit generally on a limited spatial scale [30]. Atmospheric emissions, wastewater reuse, and agricultural operations can all act as diffuse sources, resulting in the widespread distribution of Cd in the environment [31].
The screens of mobile phones from different generations showed a significant decrease in the quantity of Cd (from 1.0 μg/g to undetectable levels) and of Pb (from 35.0 μg/g to 2.0 μg/g) from feature phones to smartphones [32]. Cd is also commonly present in children’s toys. As reported by Igweze et al., [33] cheap toys purchased from Port Harcourt, Nigeria stores were determined to contain three toxic metals (Pb, Cd, and As). The present heavy metals in all the toys were below the limits set by the EU [33].
Heavy metals, including arsenic, cadmium, chromium, and lead, are present in goods made from leathers, synthetic leathers used in shoe production, and textiles. Bielak et al. [34] found that children's footwear made from sheepskins contained As, Ba, Cd, Cr, Hg, Pb, Sb, and Se. In 2006, leathers used for insoles, shoe uppers, and clothing underwent testing in Turkey for Co, Cr, Cu, Pb, Ni, and Zn due to their close contact with the body. In Bielak et al.’s paper, it is described that in 2015, various types of fibers used in the textile industry in Turkey, such as cotton, acrylic, polyester, nylon, viscose, and polypropylene, were examined for the presence of Al, Cd, Co, Cr, Cu, Fe, Mn, Ni, Pb, Tl, and Zn. Additionally, in Italy in 2009, Cr was identified in a wool top [34].
The utilization of cadmium for creating affordable jewelry has recently become of concern. Kern et al. conducted a study in 2020 and found that seven out of the nine undamaged jewelry items tested released cadmium in amounts that surpassed the recommended maximum of 18 μg. They concluded that the undamaged items had a maximum extractable cadmium amount of 6230 μg, nearly three hundred times greater than the 18-μg limit [35].
3. Cd Toxicity on Living Organisms
Heavy metals, respectively Cd, exist in soils in two forms - immobilized (organically bound) and mobile (in the soil solution). The immobilized ones are considered nonbioavailable, while the mobile is considered bioavailable. The access of heavy metals to organisms is also affected by different soil characteristics such as pH, oxido-reductive balance, clay, iron oxide, and organic matter content. It is well known that the presence of Cd and other heavy metals in the environment influences the growth and survival of microorganisms and also shifts the microbial community in these habitats, resulting in poorer soil fertility [27,31,36]. As a rule, Gram-positive bacteria are more sensitive than Gram-negative bacteria [37].
3.1. Cd Accumulation and Toxicity on Bacteria
Several studies on the interactions of Cd with microorganisms were reported [38]. In some bacteria, such as Ralstonia metallidurans CH34, isolated near a zinc production plant in Belgium, a gene, called the czc-gene, responsible for the induction of resistance to Cd was found [39]. Heavy-metal resistance genes were found to be localized outside this microorganism's bacterial chromosome in two small (3-5 Kb) self-replicating circular DNA molecules called plasmids -pMOL28 and pMOL30. Overview results on bioremediation of Cd by the Gram (-) bacterium Pseudomonas aeruginosa are published by Chellaiah [38].
3.1.1. Resistance Mechanisms of Bacteria
Bacteria inhabiting high-metal environments evolved several mechanisms of resistance, fighting for survival. Usually, Cd (II) forms stable toxic complexes in the cell. The concentration of Cd ions in the cell is regulated by cellular homeostasis through these specific resistance mechanisms [40].
Resistance mechanisms of bacteria function as energy-dependent efflux of metal ions that export them out of the bacterial cell. In most cases, the resistance is carried out by plasmids that contain genes encoding resistance to Cd [32]. Besides Cd2+, the plasmid-mediated resistance system removes other metal ions such as Ag+, AsO2-, AsO43-, Co2+, CrO42-, Cu2+, Hg2+, Ni2+, Pb2+, TeO32- and Zn2+. Three mechanisms of resistance can be distinguished.
3.1.1.1. Proteins Conducting the Export of Heavy Metals
The first group of bacterial transport proteins is proteins associated with resistance, budding, and cell division (Resistance-nodulation-cell division), shortly named “RND proteins.” These proteins were first found in the widely studied multi-resistant bacterium Ralstonia metallidurans, which are polyresistant to several heavy metals. This bacterium was isolated near a Zinc smelter in Belgium and described by Mergeay et al. [39]. Besides the heavy metal efflux, RND proteins play a role in the budding of Mesorhisobium lotti and the cell division of E. coli.
RND proteins are divided into seven notable families found in organisms of all kingdoms. Most important are the families of inner-membrane, periplasmic, and outer-membrane efflux proteins. They form a single efflux - a protein complex that can evacuate Cd from the cytoplasm via the cytoplasmic membrane, cell wall and outer membrane (typical for the Gram-negatives) into the extracellular space. Unlike the ATP-dependent one, this type of transport system is energy-independent, known as the CBA-efflux transport system.
The Cd-Co-Zn- CBA-efflux system in polyresistant R. metallidurans is widely discussed. This beta-proteobacterium contains two large plasmids that determine its resistance to heavy metals, including Cd: pMOL30 and pMOL28. The first plasmid, pMOL30, encodes resistance to Co2+, Zn2+, and Cd2+ (named czc genes). When genes are expressed, they increase the MIC of Zn2+, Co2+, and Cd2+ from 7 to 50 times [32]. The structural genes responsible for Cd and other heavy metals resistance in the pMOL30 plasmid have been cloned and sequenced at least 20 years ago. Four open reading frames (ORFs) were detected in the czc-DNA sequence. They are arranged on the czc operon as follows: first is the czcC gene (encodes a CzcC protein of 346 amino acids (AA) followed by the czcB gene (CzcB protein composed of 521 AA), the czcA gene (CzcA protein composed of 1064 AA), and the czcD gene (CzcD protein consisting of 200 AA) [40]. The second plasmid, pMOL28, encodes resistance to Ni2+ and Co2+, increasing the MIC of Co2+ 16 times. This type of resistance is called the cnr gene, which encodes the CnrA protein. CzcA and CnrA proteins were the first proteins identified in the RND-family The resistance to Cd, along with cobalt and zinc, in the bacterium R. metallidurans (former name Alcaligenes eutrophus) is shown in Figure 3.
3.1.1.2. Proteins Facilitating the Transport of Heavy Metals
These proteins (Cation Diffusion Facilitators, or CDF) have been found in many prokaryotes and eukaryotes. Their substrate is predominantly cadmium but also zinc, cobalt, nickel, and iron cations [40]. Ion transport is carried out via both concentration and chemiosmotic gradients. Bacteria usually possess one or more CDF genes. Such have been found in the bacterium R. metallidurans and yeast Saccharomyces cerevisiae.
3.1.1.3. P-Type ATP-Ase
P-type ATP-ases play a protective role on the bacterial cell against heavy metals. These enzymes belong to the transport proteins that pump out the cell the metal cations using energy from ATP hydrolysis, i.e., their action is energy-dependent. Among the heavy metals, their substrates are the Cd and silver cations. P-type ATPase can import its substrate from the external environment into the cytoplasm and export it from the cytoplasm outside the cell [40]. According to this principle, P-type ATP-ases are divided into Importing and Exporting. It should be noted that P-type ATPase is precious for two reasons: it imports micronutrients such as Mg2+, but at the same time, it can also import heavy metals, and secondly, it exports heavy metal cations and thus neutralize them, driving the bacterial cell to life.
3.1.2. Bacteria in Cd-Contaminated Soil
In a study by Salam et al. Illumina shotgun sequencing of DNA extracted from two Cd-contaminated agricultural soils showed the predominance of the phyla, classes, genera, and species of Proteobacteria (37.38%), Actinobacteria (35.02%), Prevotella (6.93%), and Conexibacter woesei (8.93%) in the first sample, and Proteobacteria (50.50%), Alphaproteobacteria (22.28%), Methylobacterium (9.14%), and Methylobacterium radiotolerans (12,80%) in the second one. The study concluded that contamination with Cd has a significant impact on the structure and function of the soil microbial community, changes the resistome of heavy metals, modifies the physicochemical properties of the soil, and leads to the substantial decline of certain native community members that are not accustomed to Cd stress [41].
The presence of Cd has a crucial impact on bacterial diversity, and there are notable differences in microbial communities between areas with high Cd pollution and those with low Cd pollution. Yu et al. reported that higher concentrations of Cd significantly increased the abundance of Proteobacteria and Gemmatimonas and decreased the abundance of Nitrospirae. Members of the genera Burkholderia and Bacillus were reported to develop a resistance to Cd and may play an essential role in the bioremediation of Cd-contaminated soils [42]. In a study by Luo et al., high Cd-level sites displayed lower diversity indices than low Cd-level sites. The dominant phyla observed by the authors in paddy soil samples are Proteobacteria, Chloroflexi, Acidobacteria, Actinobacteria, Gemmatimonadetes, Verrucomicrobia, Thaumarchaeota, Firmicutes, and Nitrospirae. In this study is pointed out Actinobacteria are tolerant to Cd, whereas Proteobacteria, Verrucomicrobia, and Nitrospirae are sensitive [43].
Sun et al. [44] found that the biodiversity and composition of the soil microbial community were affected in Cd-contaminated soil, with bacterial diversity impacted more than fungal diversity. The abundance of the soil microbial community decreased. At the same time, the composition changed at the phylum level, specifically in biomarkers for bacteria such as Saccharibacteria and Gemmatimonadetes, as well as in Arenimonas, Xanthomonadales, Nitrosomonadaceae, Methylophilales, Caulobacteriales, Aeromicrobium, Chitinophagaceae, Acidimicrobiales, Nocardioidaceae, Propionibacteriales, Frankiales, and Gemmatimonadaceae, which were found to be positively correlated with the total and available Cd [44]. Khan et al. describe an overview of Cd toxicity against living organisms and microbial resistance mechanisms, emphasizing the efflux systems, antioxidant profiling, and Cd eradication potential exhibited by microorganisms when exposed to Cd+2. Cd-resistance and bioremediation potential make these microorganisms a good bioresource for green chemistry to exterminate environmental Cd+2 [45].
Heavy metals alter the soil microbial community composition, and the microorganisms that adapt to this stress increase abundance [46]. Usually, the highest bacterial diversity is detected in severely contaminated soils [47]. As reported by authors, phyla Proteobacteria and Acidobacteria are abundant in soils contaminated by Cd and other heavy metals. The prevalence of Actinobacteria, Acidobacteria, Proteobacteria, and Chloroflexi in heavy metal-polluted soils was reported by Hemmat-Jou et al. [48]. Besides, the soil pH influences the heavy metals' mobility and their toxic effects on bacterial communities. Acidic pH turns metals more bioavailable than the neutral pH.
Using minimal bacteriological media with heavy metals added and no carbon source researchers are able of growing, isolating and further studying the heavy metal-tolerant bacteria. To determine the bacterial sensitivity to heavy metals growth of the bacteria is then assessed based on different criteria such as turbidity, biomass, and enzyme activities [49].
Several studies have shown that some lactic acid bacteria (LAB), including Lactobacillus plantarum, Lactobacillus rhamnosus, Bifidobacterium breve, and Bifidobacterium lactis, can bind and remove heavy metals such as Cd and Pb in vitro [50]. There is also substantial evidence that probiotic LAB have antioxidative properties that may be effective against Cd-induced oxidative stress in humans. Based on these unique characteristics, daily LAB consumption may be a preventive dietary strategy for people exposed to Cd [50]. Table 2 shows Cd removal potential by some bacterial and yeast strains.
3.2. Cd Accumulation and Toxicity on Fungi
The primary impact of Cd’s toxicity is mainly on fungi’s growth and replication. Fungi are particularly affected, as some fungal species may be eradicated following Cd exposure in soil. There is a selection pressure for resistant strains after low-level exposure to Cd in soil.
Soil fungi have an essential role in detoxifying and improving Cd-contaminated soils. The fungal cell wall contains polysaccharides and chitin and effectively helps control Cd tolerance and its uptake while acting as a barrier against Cd2+ entry into plant cells. Functional groups like carboxylic and hydroxylic groups and amino acids in the fungal cell wall carry a negative charge, enabling the cell wall to attract positively charged metallic ions. Research has shown that species such as Absidia cylindrospora, Suillus luteus (ectomycorrhizal fungi), and the group of Neotyphodium endophytes have significant potential for Cd-tolerance and can be used effectively in remediating Cd-contaminated soils [51].
Heavy metals like Cd are known to be some of the most hazardous pollutants. In research conducted by Zheng et al., a filamentous fungus strain YZ1 was found in soil from wheat farmland. This strain, which belongs to Purpureocillium sp. based on its appearance and genetic evidence, had a minimum inhibitory concentration of 1 mM Cd and could survive in 100 mM Cd, reaching maximum biomass at 0.4 mM Cd. The strain YZ1 can be a great choice for cleaning up Cd-contaminated farmland in wheat-producing areas [52].
In another study by Fazli et al., seven fungi that were highly tolerant to Cd were tested: Aspergillus versicolor, Aspergillus fumigatus, Paecilomyces sp.9, Paecilomyces sp.G, Terichoderma sp, Microsporum sp, and Cladosporium sp [53]. The findings revealed that the fungi displayed various resistance mechanisms against Cd and could sequester Cd from liquid environments. Aspergillus versicolor showed a remarkable difference in detoxification behavior compared to the other isolated fungi. It demonstrated a strong ability to grow actively in the presence of Cd and reduce the Cd concentration to less toxic levels. The introduction of Aspergillus versicolor as a scavenger organism marks the initial step in the emergence of this fungus in bioremediation science [53].
3.3. Cd Accumulation and Toxicity on Edible Mushrooms
The accumulation of heavy metals in the fruiting bodies of mushrooms is extensively studied. The mean content of Cd in analyzed mushrooms ranges from 0.370 to 2.151 mg/kg d.w., while Pb is found at the level of 0.243–0.424 mg/kg d.w. Heavy metals are believed to cause pronounced toxicological harm to human health when contaminated mushrooms, even at low metal concentrations, are consumed [54]. Furthermore, consuming mushrooms contaminated with heavy metals can lead to damage to the kidneys and heart, as well as the impairment of the digestive, immunological, skeletal, and nervous systems [54]. Mushrooms can gather high amounts of minerals, even when grown in soils with low metal content. This is attributed to the species' genetic characteristics, including many transport genes and binding ligands [55].
Mushrooms can absorb specific forms of heavy metals, such as Cd2+, Cd6+, Hg2+, As5+, etc., into their fruiting bodies. In connection, metals' intracellular speciation and uptake are typically regulated by metallothioneins and GT-complexes that are directly linked to fungal physiology [56]. Species with the ability to accumulate Cd include Agaricus bisporus, A. campestris, A. macrosporus, Armillaria mellea, Amanita muscaria and A. allies, Boletus edulis, Cantharellus cibarius, Cystoderma carcharias, Macrolepiota procera, Xerocomus badius, and Tricholoma matsutake. The concentration of Cd in edible mushrooms can go up to 1.3925 mg/kg and readily accumulate in more significant quantities within the caps.
The highest allowable levels of Cd for Agaricus bisporus (common mushroom), Pleurotus ostreatus (oyster mushroom), Lentinula edodes (shiitake mushroom) are set at 0.20 mg/kg. For other mushroom species, it is 1.0 mg/kg. The mentioned maximum levels are applicable after washing the mushrooms and separating the edible part [57].
As the number of wild mushroom consumers continues to rise, monitoring the presence of toxic elements and their potential risks has become important. In this regard, the elevated Cd levels in wild edible mushrooms can pose health risks to consumers, especially as several species of Tricholoma mushrooms are consumed in fresh or processed form [56].
3.4. Cd Accumulation and Toxicity on Plants
In plants, Cd predominantly accumulates in the roots, with lesser amounts in the leaves [58]. Cd exhibits toxicity to a broad range of plants, but its harmful effects are mitigated by sediment, high concentrations of dissolved salts, or organic matter. Cd adversely affects plant growth in experimental settings, although no field effects have been reported. Plants more readily take the metal from nutrient solutions than from soil, with most studies demonstrating its effects in nutrient solution cultures. Cd has been shown to impact stomatal opening, transpiration, and photosynthesis in plants grown in nutrient solutions [58].
Chlorosis and stunted growth in plants are easily identifiable indicators of Cd poisoning. Higher toxicity slows plant growth and causes necrosis [59]. Cd toxicity harms plants by limiting carbon fixation and reducing chlorophyll concentration and photosynthetic activity [60]. Cd accumulation in plants can result in various physiological, biochemical, and structural alterations. Its accumulation modifies mineral nutrient intake, slows stomatal opening by interfering with plant water balance, disrupts Calvin cycle enzymes, photosynthesis, and carbohydrate metabolism, changes antioxidant metabolism, and reduces agricultural output [61].
Exposure to Cd in the soil causes osmotic stress in plants by reducing leaf-relative water content, stomatal conductance, and transpiration, resulting in physiological damage [62]. Cd poisoning generates an overproduction of reactive oxygen species (ROS), which damages plant membranes and destroys cell proteins and organelles [60].
As reported by Huybrechts et al., exposing Trigonella foenum-graecum seeds to Cr and Pb, and Cd solutions revealed that Cd at 10 mg L−1 showed the most significant inhibitory effect on germination, the highest concentration tested. Similarly, Triticum aestivum required less Cd than Pb to hinder seed germination. However, it's important to note that there can be significant variations between plant species [59].
Cd is recognized for inhibiting seed germination through various mechanisms [63]. In Vigna unguiculata seeds, the inhibitory effect of Cd was suggested to be caused by impaired water uptake, limiting the water availability for the developing embryo. In addition to a restricted water supply, inhibition of starch mobilization from the endosperm, combined with an impaired translocation of soluble sugars to the embryonic axis, can further starve the embryonic axis. A decrease in hydrolyzing enzymes, including α-amylase, proteases, and acid phosphatases, in Sorghum bicolor seeds was proposed to be responsible for the reduced storage mobilization. Calcium is crucial for amylase activity, and replacing the chemically similar Cd ion could disrupt normal enzyme functioning. Furthermore, radish seeds experienced direct competition for Ca-calmodulin binding sites between Ca and Cd ions [59].
Cd exposure is widely known to cause DNA damage. The mechanisms behind this Cd-induced DNA damage involve the ROS-induced formation of 8-hydroxyguanosine and the inhibition of DNA repair systems. Table 3 describes Cd’s effect on different plant species.
The adverse effect of Cd on the cell cycle is presented in Table 4.
The structure that provides the primary defense for a plant cell against pathogen attacks and adverse environmental conditions such as drought and metals is the cell wall. Preventing excess Cd from entering the cytoplasm is crucial because it can cause damage to macromolecules, proteins, and DNA due to Cd-induced oxidative stress. Roots in direct contact with Cd from the soil have cell walls that play a crucial role in this process. Research indicates that most plant species store the majority of Cd within the cell walls of roots [59]. When the capacity of the cell wall is surpassed, Cd may form complexes with PCs and then can finally be sequestered within vacuoles by transferring ABC transporters through the tonoplast.
For a plant to go through its life cycle, it needs to start the reproductive phase, which in higher plants involves the development of flowers, pollination, and fertilization, followed by seed production. When Brassica campestris plants were exposed to Cd during the flowering stage, the content of glutathione (GSH) and ascorbate (AsA) decreased the most, suggesting that the reproductive phase was highly vulnerable [59]. Cd has been demonstrated to harm pollen germination and disrupt pollen tube morphology in various plant species [64].
3.5. Cd Accumulation and Toxicity on Animals
Cd toxicity causes a wide range of health issues, including some of the deadliest diseases, such as heart disease [65,66], kidney disease [67,68], liver disease [67], cancer, and diabetes [65]. Practically every system in an animal's body can be harmed by Cd. Over an extended period, Cd accumulates in the kidney and liver [69]. Farm animals can come into contact with contaminated water, soil, vegetation, and car and industrial emissions [70,71]. Foods of all kinds contain a lot of different types of Cd. Foods made from grains, leafy vegetables like spinach, and mainstays like potatoes have relatively high levels of Cd, ranging from 30 to 150 ppb [72]. Sunflower, soybean, and peanut seeds all have naturally high levels of Cd, seemingly with no adverse health effects. Fish and meat typically have lower levels of Cd, ranging from 5 to 40 ppb [73]. Offal organs where Cd accumulates, like the kidney and liver, can contain remarkably high Cd levels—up to 1000 ppb [73].
Numerous organisms readily accumulate Cd, notably mollusks, where bioconcentration factors can reach several thousand. Soil invertebrates also exhibit significant Cd accumulation. In contrast, most organisms display low to moderate concentration factors, typically under 100 [74]. Cd in tissues is often bound to proteins, including specific heavy-metal-binding proteins known as metallothioneins, which have been isolated from organisms exposed to Cd [75]. The highest Cd concentrations are found in the kidney, gills, and liver (or their equivalents). The primary route of Cd elimination in organisms is likely through the kidney, although, in crustaceans, substantial amounts can also be shed via the exoskeleton.
The acute toxicity of Cd to aquatic organisms varies considerably, even among closely related species, and is influenced by the free ionic concentration of the metal. Cd interferes with calcium metabolism in animals, causing hypocalcemia in fish by inhibiting calcium uptake from the water. However, high calcium concentrations in the water can protect fish from Cd uptake by competing at the uptake sites [76]. Zinc exacerbates Cd toxicity in aquatic invertebrates. Sublethal effects on aquatic invertebrates include impaired growth and reproduction and structural damage to gills. Fish, particularly salmonids, show variable sensitivity to Cd, with sublethal effects, including spinal malformations [77]. Embryos and early larvae are the most susceptible life stages, while eggs are the least affected. There is no consistent interaction between cadmium and zinc in fish.
In a recent study by Djedjibegovic et al. Cd content (mean concentration) was tested in seafood samples in Bosnia and Herzegovina. The team concluded that mercury and cadmium were detected in all analyzed samples (100%), while lead was detected in 33 samples (89.2%). Metals content was in order Hg > Cd > Pb in most of the species, except blue mussel (Pb > Cd > Hg) and Indian white prawn (Hg > Pb > Cd). Cadmium content was close to the corresponding MRL in two samples of Patagonian squid (0.918 and 0.896 mg kg−1). It was also relatively high in the other three samples of the same species (0.591, 0.425, and 0.391 mg kg−1) [78].
A study involving long-term Cd exposure in rodents found that Cd leads to loss of bone in the jaw that supports the teeth [79]. In a study conducted in the Czech Republic by Drapal et al., higher average levels of Cd were found in the liver (0.10 mg/kg) and kidney (0.62 mg/kg) of cattle over 2 years old compared to lower levels in the liver (0.06 mg/kg) and kidney (0.24 mg/kg) of cattle under 2 years old. Ruminants are exposed to Cd contamination by consuming pasture during the summer and preserved feed, including cereals, during the winter [80,81]. According to Chirinos-Peinado et al., Cd and Pb can accumulate in fresh cow’s milk [82]. Fay et al. discussed that Cd nephrotoxicity is associated with altered microRNA expression in the rat renal cortex [83].
3.6. Cd Accumulation and Toxicity in Humans
Cd poses a significant health danger to people, even at low concentrations. The body's ability to adapt to Cd exposure is limited due to its inability to undergo metabolic degradation and poor excretion [14]. Cd toxicity affects various animal organs, including the liver, kidney, lungs, testes, prostate, heart, skeletal, neurological, and immunity. Prolonged exposure to Cd can accumulate in the body and cause illnesses primarily affecting the lungs and kidneys [84]. It is believed that chronic lung diseases, high blood pressure, cancer, leukemia, genetic toxicity, damage to human organs, abdominal pain, burning sensations, nausea, vomiting, and various cancers may be related to slow poisoning by Cd in small doses. Additionally, Cd exposure may be associated with diseases related to the central nervous system, cognitive and behavioral functions, chronic diseases, teratogenic effects, cardiovascular disease, lung function abnormalities, and damage to the kidneys [85].
Acute Cd poisoning symptoms typically develop after 24 hours and include shortness of breath, weakness, and fever. It can lead to pulmonary edema, pneumonia, and, in severe cases, respiratory failure and death. Cadmium's distribution in the body is determined by its chemical form. Exposure to Cd in the form of inorganic salts (e.g., CdCl2) leads to a higher accumulation of Cd2+ ions in the liver, kidneys, and bones than Cd combined with metallothionein. CdCl2 primarily accumulates in the liver and CdMT in the kidney [86]. Cd is deposited in various organs, including the liver, kidney, testis, spleen, heart, lungs, thymus, salivary glands, epididymis, and prostate. However, because of their high MT content, the liver and kidney store around 50% of the accumulated Cd [87]. Cd can accumulate in the pancreas, lungs, central nervous system, and testes in men. Cd particles are carried through the primary olfactory neurons [88].
3.6.1. Cd Distribution in the Human Body
Cd, a subject of increasing scientific and medical interest due to its detrimental effects on health, presents a pressing need for a comprehensive understanding of its distribution in human tissues and organs. This knowledge forms the basis for crucial in vitro and in vivo animal studies [89]. The specific amounts of Cd in various cell types and tissue layers, in particular, have not been thoroughly explored. This underscores the necessity for further investigation [90]. Only a handful of researchers have delved into the total Cd content in the human body [89]. Figure 4 below presents the primary sources of Cd and its effects on the human body.
3.6.1.1. Ingestion
Cd tends to persist and accumulate in soil, eventually entering plant metabolism. Cd enters the food chain as it accumulates in edible plant parts such as fruits and seeds [91]. This accumulation rises when soil pH decreases; therefore, acid rain has the effect of raising Cd concentrations in plants. Cereals and bread account for 34% of daily Cd intake in Western countries, followed by leafy vegetables, particularly spinach, among adults (20%), potatoes (11%), legumes and nuts (7%), stem/root vegetables (6%), and fruits (5%) [92]. Mei et al. studied the accumulation and molecular mechanisms of Cd in cereal crops and tobacco [93]. Wheat can also accumulate high levels of Cd in the grain [94]. In Eastern countries, fish and shellfish are the most common Cd sources, followed by grains and vegetables, particularly rice [95]. Rice is one of the most important food crops worldwide, feeding over 5 billion people. Thus, Cd pollution in rice has attracted great attention [96]. Although dietary Cd exposure poses a minor health risk in most Eastern countries, it remains a concern for specific subgroups. Cd is dispersed over the earth, and there are locations with extremely high concentrations of Cd in the soil. Crop uptake of Cd in these locations can result in high food exposures for those living nearby. For example, in Japan's Jinzu and Kakehashi river basins, soil is extensively contaminated with Cd from industrial waste [97,98]. Locals who habitually ingested rice cooked with Cd-contaminated water experienced a severe kidney and bone illness known as "Itai-Itai" sickness, which was marked by bone distortion and many fractures, particularly in women [99]. The decrease in Cd intake over the last 50 years could be ascribed to less sewage sludge escaping into agricultural soil due to improved control and environmental awareness in industrialized countries. These processes primarily transferred Cd from plants into the food chain, increasing human exposure to the metal. Some aquatic creatures can also be significantly impacted by Cd deposition at levels above the regulatory limits [77].
Cd intake from meals in humans ranges from 8 to 25 µg per day, with only 0.5 to 1.0 µg retained in the body. Factors affecting this form of absorption include dose, exposure period, dietary components, nutritional status, age, and gender [88]. As reported by Raikwar et al., the safe recommended intake of Cd is 15-50 µg/day for adults and 2-25 µg/day for children [69].
3.6.1.2. Inhalation
Cd air levels can be hundreds of times higher in the workplace than in the broader environment [100]. The Occupational Safety and Health Administration (OSHA) sets the permissible exposure limit (PEL) for Cd fume or Cd oxide in the workplace at 0.1 mg/m3. However, Cd concentrations in ambient air range from 1 × 10-6 mg/m3 in non-industrialized areas to 4 × 10-5 mg/m3 in urban areas. Non-occupational Cd exposure from the air is unlikely to cause harmful health effects. Cd air levels are typically insufficient to create health issues in the general population. Even in places with substantial industrial Cd emissions, the average atmospheric concentration does not exceed 35 ng Cd/m3 of air [101]. Tobacco smokers have four to five times greater levels of Cd in their blood and two to three times higher amounts in their kidneys than nonsmokers (Figure 5) [102,103]. According to Gray et al., the liquids used in electronic cigarettes also contain Cd with a concentration of 0.108 µg/g [104].
3.6.1.3. Permeation
The skin absorbs tiny amounts of Cd. Hence, it is not considered a critical route of exposure. However, recent research has highlighted the environmental significance of photosensitive CdS and CdSe pigments and nano semiconductors, whose oxidized products (cadmium sulfate (CdSO4), and cadmium selenite (CdSeO4) are significantly more soluble and bioavailable, and thus potentially more dangerous [105]. An in vitro investigation employing human full-thickness skin as a model to describe the impact of Cd exposure on skin revealed that the metal only enters the epidermis; previously, it was demonstrated that its solubility into the stratum corneum layer is a rate-limiting process (Figure 6) [87].
3.6.2. Harmful Effects on Human Organs and Systems
3.6.2.1. Effects on the Blood and Circulatory System
Smokers have higher levels of Cd in their blood and are more likely to develop atherosclerosis, specifically peripheral vascular disease, than non-smokers [106]. Research suggests that low-dose Cd exposure increases the incidence of peripheral arterial disease (PAD). This element also has a detrimental effect on the cardiovascular system [107]. Cd's harmful effects on the vasculature of many organs, particularly endothelial cells, have been demonstrated through experimental research. Cd damages endothelium and smooth muscle cells, leading to the production of atherosclerotic plaques [108]. This is supported by epidemiological and clinical research. Cd can also influence homocysteine metabolism. High levels of homocysteine can lead to heart disease, stroke, peripheral vascular disease, and cognitive impairment [109].
3.6.2.2. Effects on the Reproductive System
Cd can impair reproductive functioning. Exposure to Cd poisoning predominantly impacts testicular function [110]. The toxic effects of Cd in the testis include damage to the vascular endothelium. Cells experience morphological and functional changes, including inhibition of testosterone synthesis and impaired spermatogenesis due to oxidative stress, impaired antioxidant defense mechanisms, and inflammatory response severity. Cd can disrupt prostate function, affecting hormonal activity, secretion, and fertility in men [88]. Exposure to Cd harms human male reproductive organs/systems, impairing spermatogenesis, semen quality, particularly sperm motility, and hormone synthesis/release [111]. According to experimental and human investigations, it also disrupts female fertility, reproductive hormone balance, and menstrual cycles [112]. Based on the available data, it is possible to conclude that low-dose Cd exposure harms both male and female reproduction and impacts pregnancy or its outcome [113]. Furthermore, maternal prenatal Cd exposure may have a distinct effect on male and female offspring, particularly female offspring. As a result, measures must be taken to limit exposure to Cd [114]. The testicles are the primary target of Cd-induced acute toxicity [115]. The effect is quick. The testicles begin to shrink, followed by inflammation, edema, acute bleeding, and necrosis in less than 24 hours after a single dose of cadmium [116].
3.6.2.3. Effects on the Respiratory System
Inhalation produces respiratory distress and damages the respiratory tract [117]. High Cd concentrations in contaminated air have been linked to conditions like emphysema, anosmia, and chronic rhinitis. Lampe et al. [118] studied the impact of Cd exposure on lung function among 96 males with one to three lung function tests between 1994 and 2002. Researchers discovered that smoking was linked to lower forced expiratory volume in 1 second (a measure of lung function) and higher urine cadmium levels [120]. Inhaling Cd can cause respiratory distress syndrome [88].
3.6.2.4. Effects on the Kidney System and Bones
Long-term exposure to high doses of Cd results in Itai-Itai disease, primarily affecting women. This disease is marked by severely compromised tubular and glomerular function, generalized osteomalacia, and osteoporosis, leading to multiple bone fractures [119]. Prolonged exposure to low doses of Cd has been associated with tubular damage, resulting in reduced reabsorptive capacity for nutrients, vitamins, and minerals, nephropathy, and proteinuria [72,102]. Non-absorbed molecules include zinc or copper bound to metallothionein (MT), glucose, amino acids, phosphate, calcium, and low-molecular-weight (LMW) proteins like 2-microglobulin (2-M) and 1-microglobulin (1-M), also known as protein HC, retinol-binding protein (RBP), and uric acid, resembling Fanconi syndrome, a genetic disorder of renal tubular transport. Urinary markers for Cd exposure include Cd itself, LMW proteins (2-M, 1-M), and enzymes of renal tubular origin, such as N-acetyl glucosaminidase (NAG). Generally, urinary Cd levels indicate long-term body burden before kidney damage develops, while blood Cd levels indicate recent exposure [120]. Cd disrupts the metabolism of calcium, magnesium, iron, zinc, and copper in cells, leading to demineralization, osteomalacia, osteoporosis, and bone disorders, necessitating the replacement of these ions. Cd's competitive displacement of calcium ions weakens bone structure, often causing fractures, especially in children and postmenopausal women. Cd also inhibits the activity of 1-hydroxycholecalciferol hydroxylase, an enzyme essential for converting 25(OH)D3 to the active form of vitamin D3, 1,25(OH)2D3, in the kidney [86]. This active form is crucial for calcium absorption in the intestine [88]. Research of 31 to 60-year-old men from Pakistan's growing industrial nation found that diabetic males (N = 196) had considerably higher Cd levels in their blood and urine than non-diabetic males (N = 238). These disparities were observed in both smokers (n = 209) and non-smokers (n = 225) [121]. Although Cd is known for its renal toxicity, the dose-response relationship at low environmental exposure levels remains unclear. The Centers for Disease Prevention and Control (CDC) and European health agencies acknowledge this uncertainty and the potential public health impact of low-level Cd exposure, urging further research.
The Avonmouth lead/zinc smelter in southwest England closed in early 2003 and was the UK's largest source of atmospheric Cd emissions. It emitted 978 kg of Cd in its final year, accounting for nearly 30% of the UK's point source emissions. The site had produced zinc for over 70 years, emitting other nephrotoxic metals like lead, mercury, and arsenic. Given Cd's high nephrotoxicity and long biological half-life, this study focused on Cd exposure. With approximately 50,000 people living within 5 km of the smelter, there is concern about increased Cd exposure from inhalation contaminated air and ingesting homegrown vegetables and house dust. 180 volunteers (74 men and 106 women) were studied for health problems caused by Cd. Among the participants, 109 (40 men and 69 women; 61%) were never-smokers, 36 (19 men and 17 women; 20%) were former smokers, and 32 (13 men and 19 women; 18%) were current smokers. Results obtained by urine analyses showed early kidney damage [122].
3.6.2.5. Effects on the Central Nervous System
Accumulating evidence over the years suggests that Cd may be a potential cause of neurodegenerative diseases. This could be related to the excessive production of free radicals, which damage organs depending on their defense mechanisms [123]. Since Cd is a toxic agent affecting various cell types, this study aimed to investigate Cd's effects and consequences on different organs in mice. To test the hypothesis of concentration-dependent Reactive Oxygen Species (ROS) generation and DNA damage, Agnihotri et al. [124] assessed the serum of 4–5 week-old male Swiss albino mice, which were treated with cadmium chloride (CdCl2), added to their drinking water for 30 days. The expression of Bcl-2-associated X protein (Bax), an apoptotic marker, was twice as high in the brain compared to the liver at an exposure level of 0.5 mg L−1 CdCl2 [124]. Additionally, correlation and linkage data analysis of the antioxidant defense system showed rapid changes in the brain compared to other organs in the study. The results indicate that even at low doses, Cd impairs the brain due to its lipid peroxidase sensitivity, which promotes Cd-induced oxidative injury in the brain [125]. Human studies indicate that exposure to Cd can lead to aberrant behavior and lower IQ in children and adults. The blood-brain barrier is susceptible to Cd. Cd has direct harmful effects when it crosses the blood-brain barrier, which can happen in young children or with certain medical conditions. The choroid plexus epithelium can accumulate significant levels of Cd, which reduces concentrations in other body regions [126].
Cd toxicity can cause cell injury, cell death, and organ failure through various chemical pathways, the most common of which is oxidative stress, in numerous body compartments and tissues, including the central nervous system [127]. In this context, many in vivo and in vitro research studies have provided evidence showing Cd-induced neurotoxicity in the CNS and Cd’s involvement in many compartments and cells of the CNS [128]. Cd-treated embryos developed a smaller head with unclear boundaries between the brain subdivisions, particularly in the mid-hindbrain region. Embryos display normal anterior-to-posterior regionalization; however, the commitment of neural progenitor cells was affected by Cd [76]. Cd has been shown to produce free radicals in the brain, potentially damaging neurons and oligodendrocytes (OLG). OLGs are the glial cells which myelinate axons in the CNS. An early study reported that Cd toxicity affected CNS white matter [129], and one laboratory demonstrated that OLGs are direct targets of this structure [130]. Experimental studies have shown that Cd can also be a potent neurotoxicant for the peripheral nervous system [131]. Moreover, Cd has a half-life of more than 15 years in humans. Elderly workers may be more susceptible to an increased Cd body burden and may develop peripheral polyneuropathy (PNP) over time [132].
3.6.2.6. Effects on the Immune System
Chronic Cd exposure can negatively impact the immune system [133]. Cd targets several cells, including T cells, macrophages, B cells, and natural killer cells. Cd's direct immunotoxicity alters immunological responses, including cell-mediated and humoral immunity. Cd toxicity may cause anemia and eosinophilia, according to Sarkar et al. [88]. The effect of Cd on the immune system has been investigated utilizing experimental animals, particularly rodents. Cd can interact directly with immune system cells, causing modifications in their status and functionality that can be evaluated in vitro. Direct consequences include cell death and signaling pathways interference, altered cytokine production, cell surface marker expression, cell activation, and differentiation [134]. It has been demonstrated that Cd exposure impairs immune cells' ability to operate normally. When administered orally to mice, Cd increases vulnerability to herpes simplex virus type 2, reduces T and B cell production, and enhances macrophage phagocytosis [135]. It has been shown that school-age children exposed to Cd experience a drop in sensitivity and IgG antibody titers. Stress is thought to be the cause of the induction of apoptosis, as is the case with many other immunotoxic drugs. Cd also has an impact on cytokine production [136].
4. Molecular Mechanisms of Cd Toxicity
Cd affects cellular proliferation, differentiation, and apoptosis. The International Agency for Research on Cancer (IARC) classed it as a proven carcinogen, a member of Group No1. However, it has a low genotoxic potential. Cd's indirect effects cause reactive oxygen species (ROS) production and DNA damage [137,138]. In vitro studies indicate that Cd has several activities that still need to be fully understood. Chronic heavy metal exposure leads to increased expression of stress proteins (such as heat shock protein 70 and metallothioneins), which can cause apoptosis, growth inhibition, proliferation, or carcinogenicity in animal cells, depending on factors like amount, timing, cell line, and presence of other chemicals. Cd carcinogenesis is primarily caused by oxidative stress, DNA repair inhibition, and altered apoptosis rates [88,139]. A recent study describes the effects of Cd on signaling through Ca2+, NO, c-AMP, nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), and developmental pathways such as Wnt signaling, as well as kinases [125]. There are well-established effects of Cd on kinase activation and downstream events of immediate early response oncogene induction, as these events are likely involved in cancer promotion and progression and cell survival [140]. Mitochondria regulate cell homeostasis, proliferation, motility, senescence, and death. Cell and tissue aging, as well as numerous illnesses, such as Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, and even cancer, are associated with mitochondrial dysfunction [141].
Cd can change the expression of various genes, including immediate early response genes, stress response genes, transcriptional factors, and translational factors. It activates the c-jun N-terminal kinase (JNK) pathway, leading to the over-expression of genes responsible for the synthesis of metallothionines and heat shock proteins. Cd also affects transcription factors such as metal-regulatory transcription factor, nuclear factor-κB, and NF-E2-related factor 2, as well as translational factors like TIF3 and TEF-1δ. These changes can provoke the development and progression of tumors [142].
The mechanisms by which Cd disrupts gene expression include changes in Ca intracellular level, ROS generation, effect on cell kinases, and DNA methylation. Cadmium increases intracellular Ca content, affecting gene expression directly by binding to target sites in different genes or indirectly through activation of kinases. Additionally, it mimics Ca and activates Ca-dependent genes. Changes in Ca level lead to ROS production and an increase in gene expression. Cadmium activates cellular protein kinases, leading to increased phosphorylation of transcription factors and an increase in certain gene expression [142]. Additionally, cadmium exposure inhibits DNA-methyltransferase-1 (DNMT1) and decreases DNA methylation, which can be associated with cell transformation and hyperproliferation. Cadmium exposure inhibits DNA repair and damages the genome, leading to potential cell transformation. It interferes with repair processes and inhibits DNA repair genes expression, transcription factor activity, and protein function. Additionally, cadmium can replace zinc in proteins, leading to nonfunctional enzymes. Zinc supplementation can help correct some of the DNA damage [142].
Autophagy, a process of self-degradation that plays a crucial role in eliminating proteins and clearing damaged organelles, is increasingly acknowledged to be involved in Cd toxicity. Cd can function as both a protector and a promoter of cell death [146]. The conflicting impact on cell fate is determined by the appropriate level of autophagy needed to sustain cell survival. Cd exposure disrupts normal cellular autophagy, and both excessive autophagy and its absence can lead to cell death [141]. The effects of Cd on the autophagy process are observed as either stimulation or disruption, likely based on gene expression [141]. Autophagy following Cd exposure appears to either suppress or trigger apoptosis, as the increased accumulation of ROS can activate both autophagy and apoptosis. Moreover, Cd-induced elevation of intracellular Ca leads to ROS induction, initiating cell apoptosis due to the interaction between Ca signaling and ROS in normal and pathological conditions (Figure 7) [142].
Cd triggers cell death by modifying the behavior of protein kinases such as mitogen-activated protein kinase (MAPK). Cd enhances the activity of p38-mitogen-activated protein kinase (p38 MAPK), leading to increased expression of genes related to inflammation and cell death, ultimately resulting in tissue necrosis and kidney damage in rats. Furthermore, recent findings show that Cd prompts cell death in TM3 Leydig cells by generating reactive oxygen species (ROS) and promoting phosphorylation via the JNK pathway. Consequently, this causes a reduction in the levels of the anti-cell death protein Bcl-2, followed by the activation of caspase-3 and cell demise [142].
Exposure to Cd impacts the functioning of glutamate, acetylcholine, GABA (gamma-aminobutyric acid), and dopamine neurotransmitter receptors in the brain. N-methyl-D-aspartate receptor (NMDAR) voltage-dependent calcium channels facilitate neuronal uptake of Cd, which leads to increased Cd influx following stimulation with glutamate or N-methyl-D-aspartate (NMDA) and glycine. Additionally, Cd interacts with muscarinic acetylcholine receptors, leading to cell death in primary cholinergic neurons from the basal forebrain by suppressing the muscarinic receptor M1. A subsequent study by the same authors demonstrated that oxidative stress caused Cd-induced muscarinic receptor disruption [143].
Early research suggests that Cd inhibits the neuronal GABAA receptor channel complex through a binding site different from the recognition sites for GABA and other drugs. Moreover, exposure to Cd alters the expression of GABAA receptors in animal studies. Specifically, altered protein expression levels of GABAARα5 and GABAARδ were observed in the hippocampus of mice offspring following Cd exposure during pregnancy and lactation, indicating that GABAARα5 is more susceptible to environmental pollutants during puberty and young adulthood. Conversely, GABAARδ may reflect the accumulation of environmental contaminants in adulthood [143].
Cd-induced neurotoxicity causes impairments in movement due to Cd's specific impact on DA receptors. Cd exposure reduces the production of mRNA and proteins associated with dopamine (DA)-D2 receptors in the stratum of rat brains. In contrast, the levels of expression for DA-D1 receptors remain unchanged. Additionally, experiments using molecular docking have shown that Cd may directly attach itself to the competitive site of dopamine on DA-D2 receptors (Figure 8) [143].
CdCl2 leads to the disassembly of the cytoskeleton in various cultured neuronal cells, affecting both the actin and microtubule networks. In primary rat cortical neurons, Cd causes the destruction of microtubules and reduces acetylated tubulin levels. Furthermore, Cd down-regulates the gene expression of microtubule dynamics and microtubule motor-based proteins in a neuronal human cellular model [143,145].
The toxic effects of Cd on human health are widespread and are caused by various biochemical and molecular mechanisms. The main ways in which Cd causes harm include inducing oxidative stress, disrupting Ca2+ signaling, interfering with cellular signaling pathways, and making epigenetic modifications. Cd interacts with cellular components such as mitochondria and DNA, and causes extensive damage at both cellular and tissue levels [146]. Cd induces oxidative stress, which is a crucial mechanism behind its toxicity, and thus disrupts the balance between oxidants and antioxidants, leading to cellular damage and apoptosis. Furthermore, Cd negatively impacts signaling pathways such as Mitogen-Activated Protein Kinase (MAPK), Nuclear Factor kappa-light-chain-enhancer of activated B cells (NF-κB), and Tumor Protein 53 (p53) pathways. Cd’s interference with these pathways contributes to pathological conditions and carcinogenesis. The epigenetic effects of Cd, include DNA methylation and histone modifications, and causes long-term impact on gene expression and disease manifestation (Figure 9) [146].
5. Methods for Cd Removal
5.1. From the Environment
5.1.1. Chemical Precipitation
Chemical precipitation is the most efficient and widely used technology for removing Cd+2 from wastewater because it is simple, inexpensive, and easy to apply. In this procedure, substances react with Cd+2 to produce insoluble precipitates [147]. Precipitants such as hydroxides and sulfides are often employed to precipitate Cd+2, resulting in insoluble Cd(OH)2 and CdS residues, respectively. Simple sedimentation or filtration procedures could remove insoluble precipitates from water before they are released into the environment or reused [45].
5.1.2. Adsorption
Adsorption is another excellent way to remove Cd+2 from wastewater. Physiological interactions transfer metal ions from the aqueous to the solid phase [148]. Mineral clay, mineral ores, agricultural wastes, synthetic polymers, and industrial leftovers are all examples of low-cost absorbent materials [45].
5.1.3. Ion-Exchange
This approach uses the ion exchange principle to adsorb Cd+2 on solid surfaces. Metal ions in solution are replaced by ions bonded to the surface of an insoluble matrix, commonly known as an ion exchanger, and covalently bound to negative or positive functional groups. As a result, these functional groups are connected to oppositely charged ions, which are then replaced by the same charged ions found in the solution due to their higher affinity for the functional groups. Many inexpensive materials, such as waste iron, fly ash, resins, zeolites, and silicate minerals, have been widely used as ion exchangers [45]. pH and temperature are the most important parameters influencing ion exchangers' uptake of metal ions.
5.1.4. Membrane Filtration
Membrane filtration is a practical approach for removing hazardous metal ions from polluted water. Several methods use membranes to remove metal ions from effluent, including nanofiltration, ultrafiltration, electrodialysis, and reverse osmosis. Generally, metal-containing wastewater is passed over membranes with varying pore diameters depending on the nature and kind of metal ions to be extracted using selective pressure or electric current (for example, in electrodialysis). The membrane's pore size is smaller than the metal ion, preventing metal ions from passing through and decontaminating the effluent.
All conventional treatment methods have disadvantages, such as waste byproducts, sludge production, membrane fouling, low selectivity and efficiency, and high capital and operating costs [149]. Thus, using living and dead microbial biomass to remove heavy metal ions from the environment has gained popularity due to its benefits, such as cost-effectiveness, efficiency, and environmental friendliness [150].
5.2. From the Human Body
Nano selenium-enriched probiotics have been investigated recently for their potential in addressing Cd liver toxicity [151]. The presence of Cd and other heavy metals like Pb in the gut and food-derived microbes can actively or passively affect the bioavailability of these toxins within the gut [152]. Probiotic treatment can alleviate Cd-induced cytotoxicity, reduce oxidative stress and inflammation, restore tight-junction integrity, and lower gut permeability in intestinal epithelial cells and animals. This safeguarding of the gut barrier leads to higher levels of fecal Cd and reduced Cd accumulation in mouse tissues, indicating probiotics' ability to inhibit intestinal Cd absorption [153]. Capriglione et al. studied the impact of CdCl2 on immortalized, non-tumorigenic thyroid cells Nthy-ori-3-1 and the protective influence of quercetin against CdCl2-induced damage [154]. Nutritional trace metals such as Zn and Se have the ability to alleviate Cd-induced mitochondrial toxicity. Furthermore, Se supplementation can reduce Cd-induced oxidative stress and the mitochondrial apoptosis pathway [155]. A study reported by Smereczanski et al. found that an extract from Aronia melanocarpa L. berries has a protective effect against Cd in a rat model. The co-administration of that extract significantly reduces the harmful effect of Cd-induced kidney damage [156]. Resveratrol has also been found to have a protective effect against hepatotoxicity, according to Al-Baqami et al. [157]. Magnesium chloride was reported to diminish Cd toxicity via drinking water with a concentration of 500 mg Mg/L [158]. Asperuloside, an iridoid monoterpenoid glycoside found in many medicinal plants, is described as a substance attenuating Cd-induced toxicity [159].
6. Cd Resistance Mechanisms
Some microbes, such as bacteria, absorb heavy metals such as Cd+2 and necessary metal ions, helping to eliminate hazardous metal ions from the aquatic environment [152]. Heavy metal ions within the cell can generate ROS and thus disorder bacterial metabolism by damaging DNA, RNA, and proteins. Hyperaccumulation of Cd+2 may cause bacterial respiratory proteins to disorganize, disrupting the physiological functioning of the cell. To battle the fatal effects of heavy metals, bacteria have evolved various metal resistance methods, including biosorption, efflux transport, intracellular and extracellular sequestration, transformation, and physiological adaptations [160].
Organisms have mechanisms to detoxify Cd, including chelation, compartmentalization, and efflux. In yeast S. cerevisiae, Yeast Cadmium Factor 1 (YCF1) provides resistance to Cd by sequestering glutathione-conjugated cadmium, bis(glutathionato)Cd [161]. Efflux and sequestration of heavy metals via metallothioneins and other molecules containing thiol groups are well-known metal resistance mechanisms in bacteria against Cd+2, briefly described below [45].
6.1. Biosorption
Bacteria and yeasts have the intrinsic ability to bioabsorb metal ions due to their unique cell envelope, which limits cellular intake of harmful metal ions to preserve homeostasis [162]. When dealing with microbial organisms such as yeast or bacteria, the cell wall is the first structure interacting with metal ions. Several studies have indicated the existence of critical functional groups on biomass/biomaterial surfaces, such as hydroxyl, thiol, carboxyl, and amino groups, which play a crucial role in metal ion biosorption [163]. Nonetheless, the specific process of Cd+2 biosorption is unknown and varies depending on factors such as biomass type, heavy metal properties, co-metal ion presence, pH, and medium temperature. The most influential factor is the composition of biomaterials' surfaces [45].
Understanding biosorption's mechanism requires a better understanding of the cell surface's structure and chemical makeup. Gram-negative bacteria's cell wall comprises peptidoglycan, phospholipids, and lipopolysaccharides. The lipopolysaccharides have a negative charge, contributing to the cell wall's anionic character. The cell wall of Gram-positive bacteria contains about equal amounts of peptidoglycan and teichoic acids (TAs). While both carry an anionic charge at neutral pH, TAs (anionic, phosphate-based linear polymers) are critical in keeping a net negative charge on the bacterial surface [45].
6.2. Efflux Transport Systems
Heavy metals enter microbial cells via critical metal ion absorption pathways earlier described in this review. Cd uses manganese and magnesium transport systems in Gram-positive and Gram-negative bacteria. Cd hyperaccumulation causes the cell to produce efflux systems for its removal to maintain homeostasis. The efflux systems may be chromosomal or plasmid-controlled. Three distinct efflux systems, notably resistance nodulation cell division (RND), P-type ATPases, and cation diffusion facilitator (CDF), have been discovered in bacteria to remove heavy metal divalent cations such as Cd+2 from the cells [45].
7. Conclusions
Cd's toxic effects, environmental impact, sources, health effects, and biological impacts make it a pressing issue. This review highlights its toxic nature, widespread pollution, entry into living organisms’ systems, including the human body, through ingestion, inhalation, and permeation, and its accumulation in organs, leading to severe health issues. Cd exposure leads to extensive environmental and health harm. Cd is highly toxic and can be absorbed by plants and crops from the soil, enters the animal body leading to potential exposure for humans through the food chain. It accumulates in various organs, particularly the kidneys and liver, and is known to cause severe health problems, including renal dysfunction, bone diseases, cardiovascular problems, and many others. On a cellular level, Cd disrupts numerous biological processes, inducing oxidative stress generation and DNA damage, and is classified as a carcinogen by the International Agency for Research on Cancer (IARC). Preventing the use of products containing Cd is essential to minimize its adverse effects on humans and other living beings. The current Cd usage trend will lead to more severe consequences if it continues. To avoid Cd toxicity, restoring Cd-contaminated sites and appropriately disposing of materials containing Cd is essential.
Author Contributions
Conceptualization, Supervision, Writing - review & editing, Data curation, Project administration, Funding Acquisition – GS; Writing - original draft, Data curation, Visualization – SD, VM; Data curation, Visualization – SD, VM; Supervision, Writing - review & editing, Data curation – GS. All authors have read and agreed to the published version of the manuscript.
Funding
APC was funded by Grant NBU-GFSR-28/2024, Central Fund for Strategic Development - New Bulgarian University, Sofia, Bulgaria.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used during the current study are available from the corresponding author upon request.
Acknowledgments
This work was conducted at UPIZ Educational and Research Laboratory of Biology-MF-NBU, New Bulgarian University, Sofia, Bulgaria.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Peana, M.; Pelucelli, A.; Chasapis, C.T.; Perlepes, S.P.; Bekiari, V.; Medici, S.; Zoroddu, M.A. Biological Effects of Human Exposure to Environmental Cadmium. Biomolecules 2023, 13, 36. [Google Scholar] [CrossRef] [PubMed]
- Thomas O. Llewellyn IC 9380 Cadmium (Materials Flow). The National Institute for Occupational Safety and Health (NIOSH), 1994.
- Goering, P. L.; Waalkes, M. P.; Klaassen, C. D. Toxicology of Cadmium. Toxicology of Metals 1995, 189–214. [Google Scholar] [CrossRef]
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The Effects of Cadmium Toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Feng, J.; Gao, Y.; Liu, X.; Qu, L.; Zhu, L. Physiologically Based Toxicokinetic and Toxicodynamic (PBTK-TD) Modelling of Cd and Pb Exposure in Adult Zebrafish Danio Rerio: Accumulation and Toxicity. Environmental Pollution 2019, 249, 959–968, Nordberg, G. F. Historical Perspectives on Cadmium Toxicology. Toxicology and Applied Pharmacology 2009, 238 (3), 192–200. https://doi.org/10.1016/j.taap.2009.03.015. [Google Scholar] [CrossRef] [PubMed]
- Fatima, G.; Raza, A. M.; Hadi, N.; Nigam, N.; Mahdi, A. A. Cadmium in Human Diseases: It’s More than Just a Mere Metal. Indian Journal of Clinical Biochemistry 2019, 34, 371–378. [Google Scholar] [CrossRef]
- Nordberg, G. F. Historical Perspectives on Cadmium Toxicology. Toxicology and Applied Pharmacology 2009, 238, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Ding, S.; Li, C.; Tang, Y.; Fan, X.; Xu, H.; Daniel C.W., Tsang; Zhang, C. High Cadmium Pollution from Sediments in a Eutrophic Lake Caused by Dissolved Organic Matter Complexation and Reduction of Manganese Oxide. Water Research 2021, 190, 116711–116711. [Google Scholar] [CrossRef]
- Sienko, M. J. Chemistry, Principles and Applications; New York: McGraw-Hill, 1979. [Google Scholar]
- Friberg, L. T.; Carl-Gustaf Elinder; Tord Kjellstrom; Nordberg, G. F. Cadmium and Health; CRC Press, 2019.
- Lindén, K. Andersson, A. Os, A. Cadmium in Organic and Conventional Pig Production. Archives of Environmental Contamination and Toxicology 2001, 40, 425–431. [Google Scholar] [CrossRef]
- Suciu, N. A.; Devivo, R.; Rizzati, N.; Capri, E. Cd Content in Phosphate Fertilizer: Which Potential Risk for the Environment and Human Health? Current Opinion in Environmental Science & Health 2022, 100392. [Google Scholar] [CrossRef]
- Alloway, B. J.; Steinnes, E. Anthropogenic Additions of Cadmium to Soils. Cadmium in Soils and Plants 1999, 97–123. [Google Scholar] [CrossRef]
- Satchanska, G. Toxicity of heavy metals and pesticides on living world. NBU Publishing house, Sofia, Bulgaria. Monograph. 2024 ISBN 978-619-233-322-5.
- Yuan, Z.; Luo, T.; Liu, X.; Hua, H.; Zhuang, Y.; Zhang, X.; Zhang, L.; Zhang, Y.; Xu, W.; Ren, J. Tracing Anthropogenic Cadmium Emissions: From Sources to Pollution. Science of The Total Environment 2019, 676, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhang, K.; Jiang, S.; Liu, D.; Zhou, H.; Zhong, R.; Zeng, Q.; Cheng, L.; Miao, X.; Tong, Y.; Lu, Q. Association of Co-Exposure to Heavy Metals with Renal Function in a Hypertensive Population. Environment international 2018, 112, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Goncharuk, E.A.; Zagoskina, N.V. Heavy Metals, Their Phytotoxicity, and the Role of Phenolic Antioxidants in Plant Stress Responses with Focus on Cadmium: Review. Molecules 2023, 28, 3921. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Zhang, Y.; Cloquet, C.; Zhu, C.; Fan, H.; Luo, C. Tracing Sources of Pollution in Soils from the Jinding Pb–Zn Mining District in China Using Cadmium and Lead Isotopes. Applied Geochemistry 2015, 52, 147–154. [Google Scholar] [CrossRef]
- Jiang, Y.; Guo, X. Multivariate and Geostatistical Analyses of Heavy Metal Pollution from Different Sources among Farmlands in the Poyang Lake Region, China. Journal of Soils and Sediments 2019, 19, 2472–2484. [Google Scholar] [CrossRef]
- Mikhailenko, A.V.; Ruban, D.A.; Ermolaev, V.A.; van Loon, A.J. Cadmium Pollution in the Tourism Environment: A Literature Review. Geosciences 2020, 10, 242. [Google Scholar] [CrossRef]
- Kubier, A.; Wilkin, R. T.; Pichler, T. Cadmium in Soils and Groundwater: A Review. Applied Geochemistry 2019, 108, 104388. [Google Scholar] [CrossRef]
- Hayat, M. T.; Nauman, M.; Nazir, N.; Ali, S.; Bangash, N. Environmental Hazards of Cadmium: Past, Present, and Future. Cadmium Toxicity and Tolerance in Plants 2019, 163–183. [Google Scholar] [CrossRef]
- European Commission. Europa.eu. https://esdac.jrc.ec.europa.eu/search/node/cadmium (accessed 2024-08-21).
- Ballabio, C.; Jones, A.; Panagos, P. Cadmium in Topsoils of the European Union – an Analysis Based on LUCAS Topsoil Database. Science of The Total Environment 2024, 912, 168710. [Google Scholar] [CrossRef]
- Directive 2008/50/EC of the European Parliament and of the Council of 21 May 2008 on ambient air quality and cleaner air for Europe.
- Wu, Y.; Yang, X.; Wang, H.; Jia, G.; Wang, T. Relationship between Ambient PM2.5 Exposure and Blood Cadmium Level in Children under 14 Years in Beijing, China. Journal of Hazardous Materials 2021, 403, 123871. [Google Scholar] [CrossRef]
- Scaccabarozzi, D.; Castillo, L.; Aromatisi, A.; Milne, L.; Búllon Castillo, A.; Muñoz-Rojas, M. Soil, Site, and Management Factors Affecting Cadmium Concentrations in Cacao-Growing Soils. Agronomy 2020, 10, 806. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Haider, F.U.; Maqsood, M.F.; Mohy-Ud-Din, W.; Shabaan, M.; Ahmad, M.; Kaleem, M.; Ishfaq, M.; Aslam, Z.; Shahzad, B. Recent Advances in Microbial-Assisted Remediation of Cadmium-Contaminated Soil. Plants 2023, 12, 3147. [Google Scholar] [CrossRef]
- Satchanska, G.; Ivanova, I.; Veneta Groudeva; Pentcheva, E. N.; Kerestedjian, T.; Evgeny Golovinsky. Culture-Dependent Approach for Determining Microbial Diversity in Soils from KCM/AGRIA Region. Comptes Renduе of Academy Bulgarian of Science 2005, 58, 409–416. [Google Scholar]
- Surenbaatar, U.; Lee, S.; Kwon, J.-Y.; Lim, H.; Kim, J.-J.; Kim, Y.-H.; Hong, Y.-S. Bioaccumulation of Lead, Cadmium, and Arsenic in a Mining Area and Its Associated Health Effects. Toxics 2023, 11, 519. [Google Scholar] [CrossRef]
- Satchanska, G. Bioindicatory and biotransformation activity of microorganisms obtained from xenobiotics polluted soils and waters. PHD Thesis, A book. Edd. PSSE, Sofia, Bulgaria. 2022, 146 p. ISBN 978-954-749-126-7.
- Gómez, M.; Grimes, S.; Qian, Y.; Feng, Y.; Fowler, G. Critical and Strategic Metals in Mobile Phones: A Detailed Characterisation of Multigenerational Waste Mobile Phones and the Economic Drivers for Recovery of Metal Value. Journal of Cleaner Production 2023, 419, 138099. [Google Scholar] [CrossRef]
- Igweze, Z. N.; Ekhator, O. C.; Orisakwe, O. E. A Pediatric Health Risk Assessment of Children’s Toys Imported from China into Nigeria. Heliyon 2020, 6. [Google Scholar] [CrossRef]
- Bielak, E.; Marcinkowska, E. Heavy Metals in Leathers, Artificial Leathers, and Textiles in the Context of Quality and Safety of Use. Scientific Reports 2022, 12. [Google Scholar] [CrossRef]
- Kern, M. S.; Boron, M. L.; Weidenhamer, J. D. Buyer Beware: Inexpensive, High Cadmium Jewelry Can Pose Severe Health Risks. Science of The Total Environment 2020, 142926. [Google Scholar] [CrossRef]
- Satchanska, G.; Pentcheva, E. N.; Atanasova, R.; Groudeva, V.; Trifonova, R.; Golovinsky, E. Microbial Diversity in Heavy-Metal Polluted Waters. Biotechnology & Biotechnological Equipment 2005, 19, 61–67. [Google Scholar] [CrossRef]
- Cordoba-Novoa, H. A.; Jeimmy Cáceres-Zambrano; Torres-Rojas, E. Isolation of Native Cadmium-Tolerant Bacteria and Fungi from Cacao (Theobroma Cacao L.) - Cultivated Soils in Central Colombia. Heliyon 2023, 9, e22489–e22489. [Google Scholar] [CrossRef]
- Chellaiah, E. R. Cadmium (Heavy Metals) Bioremediation by Pseudomonas aeruginosa: A Minireview. Applied Water Science 2018, 8. [Google Scholar] [CrossRef]
- Mergeay, M.; Monchy, S.; Vallaeys, T.; Auquier, V.; Benotmane, A.; Bertin, P.; Taghavi, S.; Dunn, J.; van der Lelie, D.; Wattiez, R. Ralstonia Metallidurans, a Bacterium Specifically Adapted to Toxic Metals: Towards a Catalogue of Metal-Responsive Genes. FEMS Microbiology Reviews 2003, 27, (2–3). [Google Scholar] [CrossRef] [PubMed]
- Osman, G. E. H.; Abulreesh, H. H.; Elbanna, K.; Shaaban, M. R.; Samreen, S.; Ahmad, I. Recent Progress in Metal-Microbe Interactions: Prospects in Bioremediation. Journal of Pure and Applied Microbiology 2019, 13, 13–26. [Google Scholar] [CrossRef]
- Salam, L. B.; Obayori, O. S.; Ilori, M. O.; Amund, O. O. Effects of Cadmium Perturbation on the Microbial Community Structure and Heavy Metal Resistome of a Tropical Agricultural Soil. Bioresources and Bioprocessing 2020, 7. [Google Scholar] [CrossRef]
- Yu, X.; Zhao, J.; Liu, X.; Sun, L.; Tian, J.; Wu, N. Cadmium Pollution Impact on the Bacterial Community Structure of Arable Soil and the Isolation of the Cadmium Resistant Bacteria. Frontiers in Microbiology 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Xie, L.; Jin, D.; Mi, B.; Wang, D. H.; Li, X. F.; Dai, X.; Zou, X.; Zhang, Z.; Ma, Y.; Liu, F. Bacterial Community Response to Cadmium Contamination of Agricultural Paddy Soil. Applied Soil Ecology 2019, 139, 100–106. [Google Scholar] [CrossRef]
- Sun, H.; Shao, C.; Jin, Q.; Li, M.; Zhang, Z.; Liang, H.; Lei, H.; Qian, J.; Zhang, Y. Effects of Cadmium Contamination on Bacterial and Fungal Communities in Panax Ginseng-Growing Soil. BMC Microbiology 2022, 22. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Elahi, A.; Bukhari, D. A.; Rehman, A. Cadmium Sources, Toxicity, Resistance and Removal by Microorganisms-A Potential Strategy for Cadmium Eradication. Journal of Saudi Chemical Society 2022, 26. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Oliverio, A. M.; Brewer, T. E.; Benavent-González, A.; Eldridge, D. J.; Bardgett, R. D.; Maestre, F. T.; Singh, B. K.; Fierer, N. A Global Atlas of the Dominant Bacteria Found in Soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef]
- Li, X.; Meng, D.; Li, J.; Yin, H.; Liu, H.; Liu, X.; Cheng, C.; Xiao, Y.; Liu, Z.; Yan, M. Response of Soil Microbial Communities and Microbial Interactions to Long-Term Heavy Metal Contamination. Environmental Pollution 2017, 231, 908–917. [Google Scholar] [CrossRef]
- Hemmat-Jou, M. H.; Safari-Sinegani, A. A.; Mirzaie-Asl, A.; Tahmourespour, A. Analysis of Microbial Communities in Heavy Metals-Contaminated Soils Using the Metagenomic Approach. Ecotoxicology 2018, 27, 1281–1291. [Google Scholar] [CrossRef] [PubMed]
- Rathnayake, I. V. N.; Megharaj, M.; Krishnamurti, G. S. R.; Bolan, N. S.; Naidu, R. Heavy Metal Toxicity to Bacteria – Are the Existing Growth Media Accurate Enough to Determine Heavy Metal Toxicity? Chemosphere 2013, 90, 1195–1200. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Yin, R.; Yu, L.; Zhao, J.; Chen, W.; Yu, R.; Liu, X.; Chen, Y.; Zhang, H.; Chen, W. Screening of Lactic Acid Bacteria with Potential Protective Effects against Cadmium Toxicity. 2015, 54, 23–30. [Google Scholar] [CrossRef]
- Sidhu, G. P. S.; Bali, A. S.; Bhardwaj, R. Use of Fungi in Mitigating Cadmium Toxicity in Plants. 2019, 397–426. [Google Scholar] [CrossRef]
- Zheng, X.; Yang, S.; Chen, L.; Kimotho, R. N.; Chen, M.; Chen, J.; Zhang, J.; Li, X. A Newly-Isolated Cd-Loving Purpureocillium Sp. Strain YZ1 Substantially Alleviates Cd Toxicity to Wheat. Plant and Soil 2021. [Google Scholar] [CrossRef]
- Fazli, M. M.; Soleimani, N.; Mehrasbi, M.; Darabian, S.; Mohammadi, J.; Ramazani, A. Highly Cadmium Tolerant Fungi: Their Tolerance and Removal Potential. Journal of Environmental Health Science and Engineering 2015, 13. [Google Scholar] [CrossRef]
- Dowlati, M.; Sobhi, H. R.; Esrafili, A.; FarzadKia, M.; Yeganeh, M. Heavy Metals Content in Edible Mushrooms: A Systematic Review, Meta-Analysis and Health Risk Assessment. Trends in Food Science & Technology 2021, 109, 527–535. [Google Scholar] [CrossRef]
- Gałgowska, M.; Pietrzak-Fiećko, R. Cadmium and Lead Content in Selected Fungi from Poland and Their Edible Safety Assessment. Molecules 2021, 26, 7289. [Google Scholar] [CrossRef]
- Širić, I.; Kumar, P.; Eid, E.M.; Bachheti, A.; Kos, I.; Bedeković, D.; Mioč, B.; Humar, M. Occurrence and Health Risk Assessment of Cadmium Accumulation in Three Tricholoma Mushroom Species Collected from Wild Habitats of Central and Coastal Croatia. J. Fungi 2022, 8, 685. [Google Scholar] [CrossRef]
- Orywal, K.; Socha, K.; Nowakowski, P.; Zoń, W.; Kaczyński, P.; Mroczko, B.; Łozowicka, B.; Perkowski, M. Health Risk Assessment of Exposure to Toxic Elements Resulting from Consumption of Dried Wild-Grown Mushrooms Available for Sale. PLOS ONE 2021, 16. [Google Scholar] [CrossRef]
- Brima, E.I. Toxic Elements in Different Medicinal Plants and the Impact on Human Health. Int. J. Environ. Res. Public Health 2017, 14, 1209. [Google Scholar] [CrossRef] [PubMed]
- Huybrechts, M.; Cuypers, A.; Deckers, J.; Iven, V.; Vandionant, S.; Jozefczak, M.; Hendrix, S. Cadmium and Plant Development: An Agony from Seed to Seed. Int. J. Mol. Sci. 2019, 20, 3971. [Google Scholar] [CrossRef] [PubMed]
- Haider, F. U.; Liqun, C.; Coulter, J. A.; Cheema, S. A.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M. Cadmium Toxicity in Plants: Impacts and Remediation Strategies. Ecotoxicology and Environmental Safety 2021, 211, 111887. [Google Scholar] [CrossRef] [PubMed]
- Nazar, R.; Iqbal, N.; Masood, A.; Khan, M. I. R.; Syeed, S.; Khan, N. A. Cadmium Toxicity in Plants and Role of Mineral Nutrients in Its Alleviation. American Journal of Plant Sciences 2012, 03, 1476–1489. [Google Scholar] [CrossRef]
- Ismael, M. A.; Elyamine, A. M.; Moussa, M. G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in Plants: Uptake, Toxicity, and Its Interactions with Selenium Fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef]
- Shaari, N. E. M.; Tajudin, M. T. F. M.; Khandaker, M. M.; Majrashi, A.; Alenazi, M. M.; Abdullahi, U. A.; Mohd, K. S. Cadmium Toxicity Symptoms and Uptake Mechanism in Plants: A Review. Brazilian Journal of Biology 2024, 84. [Google Scholar] [CrossRef]
- Pan, J.; Li, D.; Shu, Z.; Jiang, X.; Ma, W.; Wang, W.; Zhu, J.; Wang, Y. CsPDC-E1α, a Novel Pyruvate Dehydrogenase Complex E1α Subunit Gene from Camellia Sinensis, Is Induced during Cadmium Inhibiting Pollen Tube Growth. Canadian Journal of Plant Science 2017. [Google Scholar] [CrossRef]
- Nguyen, J.; Patel, A.; Gensburg, A.; Bokhari, R.; Lamar, P.; Edwards, J. Diabetogenic and Obesogenic Effects of Cadmium in Db/Db Mice and Rats at a Clinically Relevant Level of Exposure. Toxics 2022, 10, 107. [Google Scholar] [CrossRef]
- Liu, Y.; Lin, X.; Hao, Z.; Yu, M.; Tang, Y.; Teng, X.; Sun, W.; Kang, L. Cadmium Exposure Caused Cardiotoxicity in Common Carps (Cyprinus Carpio L.): MiR-9-5p, Oxidative Stress, Energetic Impairment, Mitochondrial Division/Fusion Imbalance, Inflammation, and Autophagy. Fish & Shellfish Immunology 2023, 138, 108853–108853. [Google Scholar] [CrossRef]
- Andjelkovic, M.; Buha Djordjevic, A.; Antonijevic, E.; Antonijevic, B.; Stanic, M.; Kotur-Stevuljevic, J.; Spasojevic-Kalimanovska, V.; Jovanovic, M.; Boricic, N.; Wallace, D.; et al. Toxic Effect of Acute Cadmium and Lead Exposure in Rat Blood, Liver, and Kidney. Int. J. Environ. Res. Public Health 2019, 16, 274. [Google Scholar] [CrossRef]
- Cui, J.; Liu, Y.; Hao, Z.; Liu, Y.; Qiu, M.; Kang, L.; Teng, X.; Tang, Y. Cadmium Induced Time-Dependent Kidney Injury in Common Carp via Mitochondrial Pathway: Impaired Mitochondrial Energy Metabolism and Mitochondrion-Dependent Apoptosis. Aquatic Toxicology 2023, 261, 106570–106570. [Google Scholar] [CrossRef] [PubMed]
- Raikwar, M.; Kumar, P.; Singh, M. Toxic Effect of Heavy Metals in Livestock Health. Veterinary World 2008, 28. [Google Scholar] [CrossRef]
- Bąkowska, M.; Pilarczyk, B.; Tomza-Marciniak, A.; Pilarczyk, R.; Udała, J. Cadmium in Selected Organs of Game Animals from Areas with Different Degrees of Industrialisation and Its Intake by Human Consumers. Animals 2024, 14, 305. [Google Scholar] [CrossRef] [PubMed]
- Acheampong, S. Heavy Metals’ Poisoning in Farm Animals. IntechOpen eBooks 2023. [Google Scholar] [CrossRef]
- Satarug, S. Cadmium Sources and Toxicity. Toxics 2019, 7, 25. [Google Scholar] [CrossRef]
- Sethy, K.; Pati, S.; Jena, D.; Panda, S.; Pradhan, S.; Mishra, C. Heavy Metal Toxicity in Animals: A Review. The Pharma Innovation Journal 2020, 9, 134–137. [Google Scholar]
- Dobson, S.; World Health Organization. Cadmium, Environmental Aspects; 1992, ISBN 92 4 157135 7.
- Nordberg, M.; Nordberg, G.F. Metallothionein and Cadmium Toxicology—Historical Review and Commentary. Biomolecules 2022, 12, 360. [Google Scholar] [CrossRef]
- Baldisserotto, B.; Kamunde, C.; Matsuo, A.; Wood, C. M. Acute Waterborne Cadmium Uptake in Rainbow Trout Is Reduced by Dietary Calcium Carbonate. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2004, 137, 363–372. [Google Scholar] [CrossRef]
- Mahjoub, M.; Fadlaoui, S.; El Maadoudi, M.; Smiri, Y. Mercury, Lead, and Cadmium in the Muscles of Five Fish Species from the Mechraâ-Hammadi Dam in Morocco and Health Risks for Their Consumers. 2021, 2021, 1–10. [Google Scholar] [CrossRef]
- Djedjibegovic, J.; Marjanovic, A.; Tahirovic, D.; Caklovica, K.; Turalic, A.; Lugusic, A.; Omeragic, E.; Sober, M.; Caklovica, F. Heavy Metals in Commercial Fish and Seafood Products and Risk Assessment in Adult Population in Bosnia and Herzegovina. Scientific Reports 2020, 10. [Google Scholar] [CrossRef]
- Browar, A.W.; Koufos, E.B.; Wei, Y.; Leavitt, L.L.; Prozialeck, W.C.; Edwards, J.R. Cadmium Exposure Disrupts Periodontal Bone in Experimental Animals: Implications for Periodontal Disease in Humans. Toxics 2018, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Drapal, J.; Steinhauser, L.; Stastny, K.; Faldyna, M. Cadmium Concentration in Cattle Tissues in the Czech Republic. Veterinární Medicína 2021, 66, 369–375. [Google Scholar] [CrossRef]
- Tahir, I.; Alkheraije, K. A. A Review of Important Heavy Metals Toxicity with Special Emphasis on Nephrotoxicity and Its Management in Cattle. 2023, 10. [Google Scholar] [CrossRef] [PubMed]
- Chirinos-Peinado, D.; Castro-Bedriñana, J.; Ríos-Ríos, E.; Mamani-Gamarra, G.; Quijada-Caro, E.; Huacho-Jurado, A.; Nuñez-Rojas, W. Lead and Cadmium Bioaccumulation in Fresh Cow’s Milk in an Intermediate Area of the Central Andes of Peru and Risk to Human Health. Toxics 2022, 10, 317. [Google Scholar] [CrossRef] [PubMed]
- Fay, M.J.; Alt, L.A.C.; Ryba, D.; Salamah, R.; Peach, R.; Papaeliou, A.; Zawadzka, S.; Weiss, A.; Patel, N.; Rahman, A.; et al. Cadmium Nephrotoxicity Is Associated with Altered MicroRNA Expression in the Rat Renal Cortex. Toxics 2018, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Smereczański, N.M.; Brzóska, M.M. Current Levels of Environmental Exposure to Cadmium in Industrialized Countries as a Risk Factor for Kidney Damage in the General Population: A Comprehensive Review of Available Data. Int. J. Mol. Sci. 2023, 24, 8413. [Google Scholar] [CrossRef]
- Charkiewicz, A.E.; Omeljaniuk, W.J.; Nowak, K.; Garley, M.; Nikliński, J. Cadmium Toxicity and Health Effects—A Brief Summary. Molecules 2023, 28, 6620. [Google Scholar] [CrossRef]
- Skipper, A.; Sims, J.N.; Yedjou, C.G.; Tchounwou, P.B. Cadmium Chloride Induces DNA Damage and Apoptosis of Human Liver Carcinoma Cells via Oxidative Stress. Int. J. Environ. Res. Public Health 2016, 13, 88. [Google Scholar] [CrossRef]
- Chavatte, L.; Juan, M.; Mounicou, S.; Leblanc Noblesse, E.; Pays, K.; Nizard, C.; Bulteau, A.-L. Elemental and Molecular Imaging of Human Full Thickness Skin after Exposure to Heavy Metals. Metallomics 2020, 12, 1555–1562. [Google Scholar] [CrossRef]
- Sarkar, A., Ravindran, G. and Krishnamurthy, V. A brief review on the effect of cadmium toxicity: from cellular to organ level. Int J Biotechnol Res 2013, 3(1), 17–36. [Google Scholar]
- Egger, A. E.; Grabmann, G.; Gollmann-Tepeköylü, C.; Pechriggl, E. J.; Artner, C.; Türkcan, A.; Hartinger, C. G.; Fritsch, H.; Keppler, B. K.; Brenner, E.; Grimm, M.; Messner, B.; Bernhard, D. Chemical Imaging and Assessment of Cadmium Distribution in the Human Body. Metallomics 2019, 11, 2010–2019. [Google Scholar] [CrossRef] [PubMed]
- Satchanska, G. Cadmium Toxicity on Human: A Review. Journal of Balkan Ecology http://en.ecobalk.com/. 2012, 15, 349–357. [Google Scholar]
- Chavez, E.; He, Z. L.; Stoffella, P. J.; Mylavarapu, R. S.; Li, Y. C.; Moyano, B.; Baligar, V. C. Concentration of Cadmium in Cacao Beans and Its Relationship with Soil Cadmium in Southern Ecuador. Science of The Total Environment 2015, 533, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; C. Gobe, G.; A. Vesey, D.; Phelps, K.R. Cadmium and Lead Exposure, Nephrotoxicity, and Mortality. Toxics 2020, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Mei, S.; Lin, K.; Williams, D.V.; Liu, Y.; Dai, H.; Cao, F. Cadmium Accumulation in Cereal Crops and Tobacco: A Review. Agronomy 2022, 12, 1952. [Google Scholar] [CrossRef]
- Zhou, M.; Li, Z. Recent Advances in Minimizing Cadmium Accumulation in Wheat. Toxics 2022, 10, 187. [Google Scholar] [CrossRef]
- Kim, K.; Melough, M.M.; Vance, T.M.; Noh, H.; Koo, S.I.; Chun, O.K. Dietary Cadmium Intake and Sources in the US. Nutrients 2019, 11, 2. [Google Scholar] [CrossRef]
- Ai, H.; Wu, D.; Li, C.; Hou, M. Advances in Molecular Mechanisms Underlying Cadmium Uptake and Translocation in Rice. Frontiers in Plant Science 2022, 13. [Google Scholar] [CrossRef]
- Nogawa, K.; Suwazono, Y.; Nishijo, M.; Sakurai, M.; Ishizaki, M.; Morikawa, Y.; Watanabe, Y.; Kido, T.; Nakagawa, H. Increase of Lifetime Cadmium Intake Dose-Dependently Increased All Cause of Mortality in Female Inhabitants of the Cadmium-Polluted Jinzu River Basin, Toyama, Japan. Environmental Research 2018, 164, 379–384. [Google Scholar] [CrossRef]
- Nishijo, M.; Nogawa, K.; Suwazono, Y.; Kido, T.; Sakurai, M.; Nakagawa, H. Lifetime Cadmium Exposure and Mortality for Renal Diseases in Residents of the Cadmium-Polluted Kakehashi River Basin in Japan. Toxics 2020, 8, 81. [Google Scholar] [CrossRef]
- Uetani, M.; Kobayashi, E.; Suwazono, Y.; Kido, T.; Nogawa, K. Cadmium Exposure Aggravates Mortality More in Women than in Men. International journal of environmental health research 2006, 16, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Vergara-Murillo, F.; Martinez-Yanez, K.; Fortich-Revollo, A.; Paternina-Caicedo, A.; Johnson-Restrepo, B. Biochemical and Hematological Markers in Workers with Chronical Exposure to Lead and Cadmium in Colombia. Toxics 2022, 10, 524. [Google Scholar] [CrossRef] [PubMed]
- Conterato, G.M.M.; Bulcão, R.P.; Sobieski, R.; Moro, A. M.; Charão, M.F.; Monteiro, F.; Maria, F.; Moreira, A.; Roehrs, M.; Tonello, R.; Batista, B.L.; Grotto, D.; Barbosa, F.; Garcia, S.C.; Emanuelli, T. Blood Thioredoxin Reductase Activity, Oxidative Stress and Hematological Parameters in Painters and Battery Workers: Relationship with Lead and Cadmium Levels in Blood. 2013, 33, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; Moore, M. R. Adverse Health Effects of Chronic Exposure to Low-Level Cadmium in Foodstuffs and Cigarette Smoke. Environmental Health Perspectives 2004, 112, 1099–1103. [Google Scholar] [CrossRef]
- Barregard, L.; Fabricius-Lagging, E.; Lundh, T.; Mölne, J.; Wallin, M.; Olausson, M.; Modigh, C.; Sallsten, G. Cadmium, Mercury, and Lead in Kidney Cortex of Living Kidney Donors: Impact of Different Exposure Sources, Environmental Research 2010, 110, 47–54. [Google Scholar] [CrossRef]
- Gray, N.; Halstead, M.; Gonzalez-Jimenez, N.; Valentin-Blasini, L.; Watson, C.; Pappas, R.S. Analysis of Toxic Metals in Liquid from Electronic Cigarettes. Int. J. Environ. Res. Public Health 2019, 16, 4450. [Google Scholar] [CrossRef]
- Liu, H.; Gao, H.; Long, M.; Fu, H.; Alvarez, P. J. J.; Li, Q.; Zheng, S.; Qu, X.; Zhu, D. Sunlight Promotes Fast Release of Hazardous Cadmium from Widely-Used Commercial Cadmium Pigment. Environmental Science & Technology 2017, 51, 6877–6886. [Google Scholar] [CrossRef]
- Tellez-Plaza, M.; Guallar, E.; Fabsitz, R. R.; Howard, B. V.; Umans, J. G.; Francesconi, K. A.; Goessler, W.; Devereux, R. B.; Navas-Acien, A. Cadmium Exposure and Incident Peripheral Arterial Disease. Circulation. Cardiovascular quality and outcomes 2013, 6, 626–633. [Google Scholar] [CrossRef]
- Qu, F.; Zheng, W. Cadmium Exposure: Mechanisms and Pathways of Toxicity and Implications for Human Health. Toxics 2024, 12, 388. [Google Scholar] [CrossRef]
- Tinkov, A. A.; Filippini, T.; Ajsuvakova, O. P.; Skalnaya, M. G.; Aaseth, J.; Bjørklund, G.; Gatiatulina, E. R.; Popova, E. V.; Nemereshina, O. N.; Huang, P.-T.; Vinceti, M.; Skalny, A. V. Cadmium and Atherosclerosis: A Review of Toxicological Mechanisms and a Meta-Analysis of Epidemiologic Studies. Environmental Research 2018, 162, 240–260. [Google Scholar] [CrossRef]
- Ledda, C.; Cannizzaro, E.; Lovreglio, P.; Vitale, E.; Stufano, A.; Montana, A.; Li Volti, G.; Rapisarda, V. Exposure to Toxic Heavy Metals Can Influence Homocysteine Metabolism? Antioxidants 2020, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Massányi, M.; Satarug, S.; Madeddu, R.; Stawarz, R.; Massányi, P. Evidence for Ovarian and Testicular Toxicities of Cadmium and Detoxification by Natural Substances. Stresses 2022, 2, 1–16. [Google Scholar] [CrossRef]
- Saha, R.; S. Roychoudhury; Varghese, A.; Nandi, P.; Kar, K.; A. Paul Choudhury; Kalita, J. C.; Lukac, N.; P. Massanyi; A. Kolesarova. Cadmium Induced Deterioration of Sperm Quality: Protective Role of Coenzyme Q10 in Rats. CRC Press eBooks 2019, 127–132. [CrossRef]
- Massányi, P.; Massányi, M.; Madeddu, R.; Stawarz, R.; Lukáč, N. Effects of Cadmium, Lead, and Mercury on the Structure and Function of Reproductive Organs. Toxics 2020, 8, 94. [Google Scholar] [CrossRef]
- Dettwiler, M.; Flynn, A.C.; Rigutto-Farebrother, J. Effects of Non-Essential “Toxic” Trace Elements on Pregnancy Outcomes: A Narrative Overview of Recent Literature Syntheses. Int. J. Environ. Res. Public Health 2023, 20, 5536. [Google Scholar] [CrossRef]
- Kumar, S.; Sharma, A. Cadmium Toxicity: Effects on Human Reproduction and Fertility. Reviews on Environmental Health 2019, 34, 327–338. [Google Scholar] [CrossRef]
- Marini, H.R.; Micali, A.; Squadrito, G.; Puzzolo, D.; Freni, J.; Antonuccio, P.; Minutoli, L. Nutraceuticals: A New Challenge against Cadmium-Induced Testicular Injury. Nutrients 2022, 14, 663. [Google Scholar] [CrossRef]
- Taylor, B. A.; Heiniger, H. J.; Meier, H. Genetic Analysis of Resistance to Cadmium-Induced Testicular Damage in Mice. Experimental Biology and Medicine 1973, 143, 629–633. [Google Scholar] [CrossRef]
- Xiong, R.; Wu, Q.; Trbojevich, R.; Levan Muskhelishvili; Davis, K.; Bryant, M.; Richter, P.; Cao, X. Disease-Related Responses Induced by Cadmium in an in Vitro Human Airway Tissue Model. Toxicology Letters 2019, 303, 16–27. [Google Scholar] [CrossRef]
- Lampe, B. J.; Park, S. K.; Robins, T.; Mukherjee, B.; Litonjua, A. A.; Amarasiriwardena, C.; Weisskopf, M.; Sparrow, D.; Hu, H. Association between 24-Hour Urinary Cadmium and Pulmonary Function among Community-Exposed Men: The va Normative Aging Study. Environmental Health Perspectives 2008, 116, 1226–1230. [Google Scholar] [CrossRef]
- Sasaki, T.; Hyogo Horiguchi; Matsukawa, T.; Kobayashi, M.; Omori, Y.; Oguma, E.; Atsushi Komatsuda. A Suspected Case of “Itai-Itai Disease” in a Cadmium-Polluted Area in Akita Prefecture, Japan. Environmental Health and Preventive Medicine 2024, 29, 40–40. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; Ruangyuttikarn, W.; Nishijo, M.; Ruiz, P. Urinary Cadmium Threshold to Prevent Kidney Disease Development. Toxics 2018, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Afridi, H. I.; Kazi, T. G.; Kazi, N.; Jamali, M. K.; Arain, M. B.; Jalbani, N.; Baig, J. A.; Sarfraz, R. A. Evaluation of Status of Toxic Metals in Biological Samples of Diabetes Mellitus Patients. Diabetes Research and Clinical Practice 2008, 80, 280–288. [Google Scholar] [CrossRef]
- Thomas, L. D. K.; Hodgson, S.; Nieuwenhuijsen, M.; Jarup, L. Early Kidney Damage in a Population Exposed to Cadmium and Other Heavy Metals. Environmental Health Perspectives 2009, 117, 181–184. [Google Scholar] [CrossRef]
- Tobwala, S.; Wang, H.-J.; Carey, J.W.; Banks, W.A.; Ercal, N. Effects of Lead and Cadmium on Brain Endothelial Cell Survival, Monolayer Permeability, and Crucial Oxidative Stress Markers in an in Vitro Model of the Blood-Brain Barrier. Toxics 2014, 2, 258–275. [Google Scholar] [CrossRef]
- Agnihotri, S. K.; Agrawal, U.; Ghosh, I. Brain Most Susceptible to Cadmium Induced Oxidative Stress in Mice. Journal of Trace Elements in Medicine and Biology 2015, 30, 184–193. [Google Scholar] [CrossRef]
- Leal, R. B.; Rieger, D. K.; Peres, T. V.; Lopes, M. W.; Gonçalves, C. A. S. Cadmium Neurotoxicity and Its Role in Brain Disorders. Metal Ion in Stroke 2012, 751–766. [Google Scholar] [CrossRef]
- Zheng, W. Toxicology of Choroid Plexus: Special Reference to Metal-Induced Neurotoxicities. Microscopy Research and Technique 2001, 52, 89–103. [Google Scholar] [CrossRef]
- Oggiano, R.; Pisano, A.; Sabalic, A.; Farace, C.; Fenu, G.; Lintas, S.; Forte, G.; Bocca, B.; Madeddu, R. An Overview on Amyotrophic Lateral Sclerosis and Cadmium. Neurological Sciences 2020, 42, 531–537. [Google Scholar] [CrossRef]
- Branca, J.J.V.; Fiorillo, C.; Carrino, D.; Paternostro, F.; Taddei, N.; Gulisano, M.; Pacini, A.; Becatti, M. Cadmium-Induced Oxidative Stress: Focus on the Central Nervous System. Antioxidants 2020, 9, 492. [Google Scholar] [CrossRef]
- Fern, R.; Black, J. A.; Ransom, B. R.; Waxman, S. G. Cd(2+)-Induced Injury in CNS White Matter. Journal of Neurophysiology 1996, 76, 3264–3273. [Google Scholar] [CrossRef] [PubMed]
- Almazan, G.; Liu, H.-N.; Khorchid, A.; Sundararajan, S.; Martinez-Bermudez, A. K.; Chemtob, S. Exposure of Developing Oligodendrocytes to Cadmium Causes HSP72 Induction, Free Radical Generation, Reduction in Glutathione Levels, and Cell Death. Free radical biology & medicine 2000, 29, 858–869. [Google Scholar] [CrossRef]
- Arruebarrena, M.A.; Hawe, C.T.; Lee, Y.M.; Branco, R.C. Mechanisms of Cadmium Neurotoxicity. Int. J. Mol. Sci. 2023, 24, 16558. [Google Scholar] [CrossRef] [PubMed]
- Viaene, M. K.; Roels, H. A.; Leenders, J.; De Groof, M.; Swerts, L. J.; Lison, D.; Masschelein, R. Cadmium: A Possible Etiological Factor in Peripheral Polyneuropathy. Neurotoxicology 1999, 20, 7–16. [Google Scholar] [PubMed]
- Zhong, Q.; Zhou, W.; Lin, J.; Sun, W.; Qin, Y.; Li, X.; Xu, H. Independent and Combined Associations of Blood Manganese, Cadmium and Lead Exposures with the Systemic Immune-Inflammation Index in Adults. Toxics 2023, 11, 659. [Google Scholar] [CrossRef]
- Mirkov, I.; Popov Aleksandrov, A.; Ninkov, M.; Tucovic, D.; Kulas, J.; Zeljkovic, M.; Popovic, D.; Kataranovski, M. Immunotoxicology of Cadmium: Cells of the Immune System as Targets and Effectors of Cadmium Toxicity. Food and Chemical Toxicology 2021, 149, 112026. [Google Scholar] [CrossRef]
- Thomas, P. T.; Ratajczak, H. V.; Aranyi, C.; Gibbons, R. D.; Fenters, J. D. Evaluation of Host Resistance and Immune Function in Cadmium-Exposed Mice. Toxicology and Applied Pharmacology 1985, 80, 446–456. [Google Scholar] [CrossRef]
- Dakeshita, S.; Kawai, T.; Uemura, H.; Hiyoshi, M.; Oguma, E.; Horiguchi, H.; Kayama, F.; Aoshima, K.; Shirahama, S.; Rokutan, K.; Arisawa, K. Gene Expression Signatures in Peripheral Blood Cells from Japanese Women Exposed to Environmental Cadmium. Toxicology 2009, 257, 25–32. [Google Scholar] [CrossRef]
- Bertin, G.; Averbeck, D. Cadmium: Cellular Effects, Modifications of Biomolecules, Modulation of DNA Repair and Genotoxic Consequences (a Review). Biochimie 2006, 88, 1549–1559. [Google Scholar] [CrossRef]
- Teschke, R. Aluminum, Arsenic, Beryllium, Cadmium, Chromium, Cobalt, Copper, Iron, Lead, Mercury, Molybdenum, Nickel, Platinum, Thallium, Titanium, Vanadium, and Zinc: Molecular Aspects in Experimental Liver Injury. Int. J. Mol. Sci. 2022, 23, 12213. [Google Scholar] [CrossRef]
- Luevano, J.; Damodaran, C. A Review of Molecular Events of Cadmium-Induced Carcinogenesis. Journal of Environmental Pathology, Toxicology and Oncology 2014, 33, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Templeton, D. M.; Liu, Y. Multiple Roles of Cadmium in Cell Death and Survival. Chemico-Biological Interactions 2010, 188, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Ortega, R.; Carmona, A. Neurotoxicity of Environmental Metal Toxicants: Special Issue. Toxics 2022, 10, 382. [Google Scholar] [CrossRef] [PubMed]
- Đukić-Ćosić, D.; Baralić, K.; Javorac, D.; Djordjevic, A. B.; Bulat, Z. An Overview of Molecular Mechanisms in Cadmium Toxicity. Current Opinion in Toxicology 2020, 19, 56–62. [Google Scholar] [CrossRef]
- Carmona, A.; Roudeau, S.; Ortega, R. Molecular Mechanisms of Environmental Metal Neurotoxicity: A Focus on the Interactions of Metals with Synapse Structure and Function. Toxics 2021, 9, 198. [Google Scholar] [CrossRef]
- Hansen, K. B.; Yi, F.; Perszyk, R. E.; Furukawa, H.; Wollmuth, L. P.; Gibb, A. J.; Traynelis, S. F. Structure, Function, and Allosteric Modulation of NMDA Receptors. The Journal of General Physiology 2018, 150, 1081–1105. [Google Scholar] [CrossRef]
- Forcella, M.; Lau, P.; Oldani, M.; Melchioretto, P.; Bogni, A.; Gribaldo, L.; Fusi, P.; Urani, C. Neuronal Specific and Non-Specific Responses to Cadmium Possibly Involved in Neurodegeneration: A Toxicogenomics Study in a Human Neuronal Cell Model. NeuroToxicology 2020, 76, 162–173. [Google Scholar] [CrossRef]
- Qu, F.; Zheng, W. Cadmium Exposure: Mechanisms and Pathways of Toxicity and Implications for Human Health. Toxics 2024, 12, 388. [Google Scholar] [CrossRef]
- Satchanska, G. Growing Environmental Bacterium Biofilms in PEO Cryogels for Environmental Biotechnology Application. IntechOpen eBooks 2022. [Google Scholar] [CrossRef]
- Chwastowski, J.; Bradło, D.; Żukowski, W. Adsorption of Cadmium, Manganese and Lead Ions from Aqueous Solutions Using Spent Coffee Grounds and Biochar Produced by Its Pyrolysis in the Fluidized Bed Reactor. Materials 2020, 13, 2782. [Google Scholar] [CrossRef]
- Satchanska, G., Topalova, Y., Dimkov, R., Petrov, P., Tsvetanov, C., Selenska-Pobell, S., Gorbovskaya, A., Golovinsky, E. Phenol biodegradation by two xenobiotics-tolerant bacteria immobilized in polyethylene oxide cryogels. Comptes Rendus de L'Academie Bulgare des Sciences 2009, 62, 957–964, http://www.proceedings.bas.bg/.
- Alfadaly, R.A.; Elsayed, A.; Hassan, R.Y.A.; Noureldeen, A.; Darwish, H.; Gebreil, A.S. Microbial Sensing and Removal of Heavy Metals: Bioelectrochemical Detection and Removal of Chromium(VI) and Cadmium(II). Molecules 2021, 26, 2549. [Google Scholar] [CrossRef] [PubMed]
- Vicas, S.I.; Laslo, V.; Timar, A.V.; Balta, C.; Herman, H.; Ciceu, A.; Gharbia, S.; Rosu, M.; Mladin, B.; Chiana, L.; et al. Nano Selenium—Enriched Probiotics as Functional Food Products against Cadmium Liver Toxicity. Materials 2021, 14, 2257. [Google Scholar] [CrossRef] [PubMed]
- George, F.; Mahieux, S.; Daniel, C.; Titécat, M.; Beauval, N.; Houcke, I.; Neut, C.; Allorge, D.; Borges, F.; Jan, G.; et al. Assessment of Pb(II), Cd(II), and Al(III) Removal Capacity of Bacteria from Food and Gut Ecological Niches: Insights into Biodiversity to Limit Intestinal Biodisponibility of Toxic Metals. Microorganisms 2021, 9, 456. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Tian, F.; Zhao, J.; Zhang, H.; Narbad, A.; Chen, W. Oral Administration of Probiotics Inhibits Absorption of the Heavy Metal Cadmium by Protecting the Intestinal Barrier. Applied and Environmental Microbiology 2016, 82, 4429–4440. [Google Scholar] [CrossRef]
- Capriglione, F.; Maiuolo, J.; Celano, M.; Damante, G.; Russo, D.; Bulotta, S.; Maggisano, V. Quercetin Protects Human Thyroid Cells against Cadmium Toxicity. Int. J. Mol. Sci. 2021, 22, 6849. [Google Scholar] [CrossRef]
- Cheng, H.; Yang, B.; Ke, T.; Li, S.; Yang, X.; Aschner, M.; Chen, P. Mechanisms of Metal-Induced Mitochondrial Dysfunction in Neurological Disorders. Toxics 2021, 9, 142. [Google Scholar] [CrossRef]
- Smereczański, N.M.; Brzóska, M.M.; Rogalska, J.; Hutsch, T. The Protective Potential of Aronia melanocarpa L. Berry Extract against Cadmium-Induced Kidney Damage: A Study in an Animal Model of Human Environmental Exposure to This Toxic Element. Int. J. Mol. Sci. 2023, 24, 11647. [Google Scholar] [CrossRef]
- Al-Baqami, N.M.; Hamza, R.Z. Protective Effect of Resveratrol against Hepatotoxicity of Cadmium in Male Rats: Antioxidant and Histopathological Approaches. Coatings 2021, 11, 594. [Google Scholar] [CrossRef]
- Grosicki, A.; Małagocki, P.; Kycko, A.; Monkiewicz, J.; Korol, W. Magnesium Supplements Affect Selected Cadmium Toxic Actions and Uptake of Repeated Doses of Cadmium. Bulletin of the Veterinary Institute in Pulawy 2015, 59, 541–546. [Google Scholar] [CrossRef]
- Kong, Z.; Liu, C.; Opeyemi Joshua Olatunji. Asperuloside Attenuates Cadmium-Induced Toxicity by Inhibiting Oxidative Stress, Inflammation, Fibrosis and Apoptosis in Rats. Scientific Reports 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Satchanska, G., Selenska-Pobell S. Bacterial diversity in heavy metal polluted soil sample KCM-B collected near Pb-Zn smelter KCM, Bulgaria. Acta Microbiologica Bulgarica 2023, 37, 350–358. [Google Scholar] [CrossRef]
- Wei, W.; Smith, N.; Wu, X.; Kim, H.; Seravalli, J.; Khalimonchuk, O.; Lee, J. YCF1-Mediated Cadmium Resistance in Yeast Is Dependent on Copper Metabolism and Antioxidant Enzymes. Antioxidants & Redox Signaling 2014, 21, 1475–1489. [Google Scholar] [CrossRef]
- Camacho-Chab, J.C.; Castañeda-Chávez, M.D.R.; Chan-Bacab, M.J.; Aguila-Ramírez, R.N.; Galaviz-Villa, I.; Bartolo-Pérez, P.; Lango-Reynoso, F.; Tabasco-Novelo, C.; Gaylarde, C.; Ortega-Morales, B.O. Biosorption of Cadmium by Non-Toxic Extracellular Polymeric Substances (EPS) Synthesized by Bacteria from Marine Intertidal Biofilms. Int. J. Environ. Res. Public Health 2018, 15, 314. [Google Scholar] [CrossRef]
- Satchanska, G.; Topalova, Y.; Ivanov, I.; Golovinsky, E. Xenobiotic Biotransformation Potential Of Pseudomonas Rhodesiae KCM-R5 and Bacillus Subtilis KCM-RG5, Tolerant to Heavy Metals and Phenol Derivatives. Biotechnology & Biotechnological Equipment 2006, 20, 97–102. [Google Scholar] [CrossRef]
Figure 1.
Distribution of heavy metals in the environment [17].
Figure 1.
Distribution of heavy metals in the environment [17].
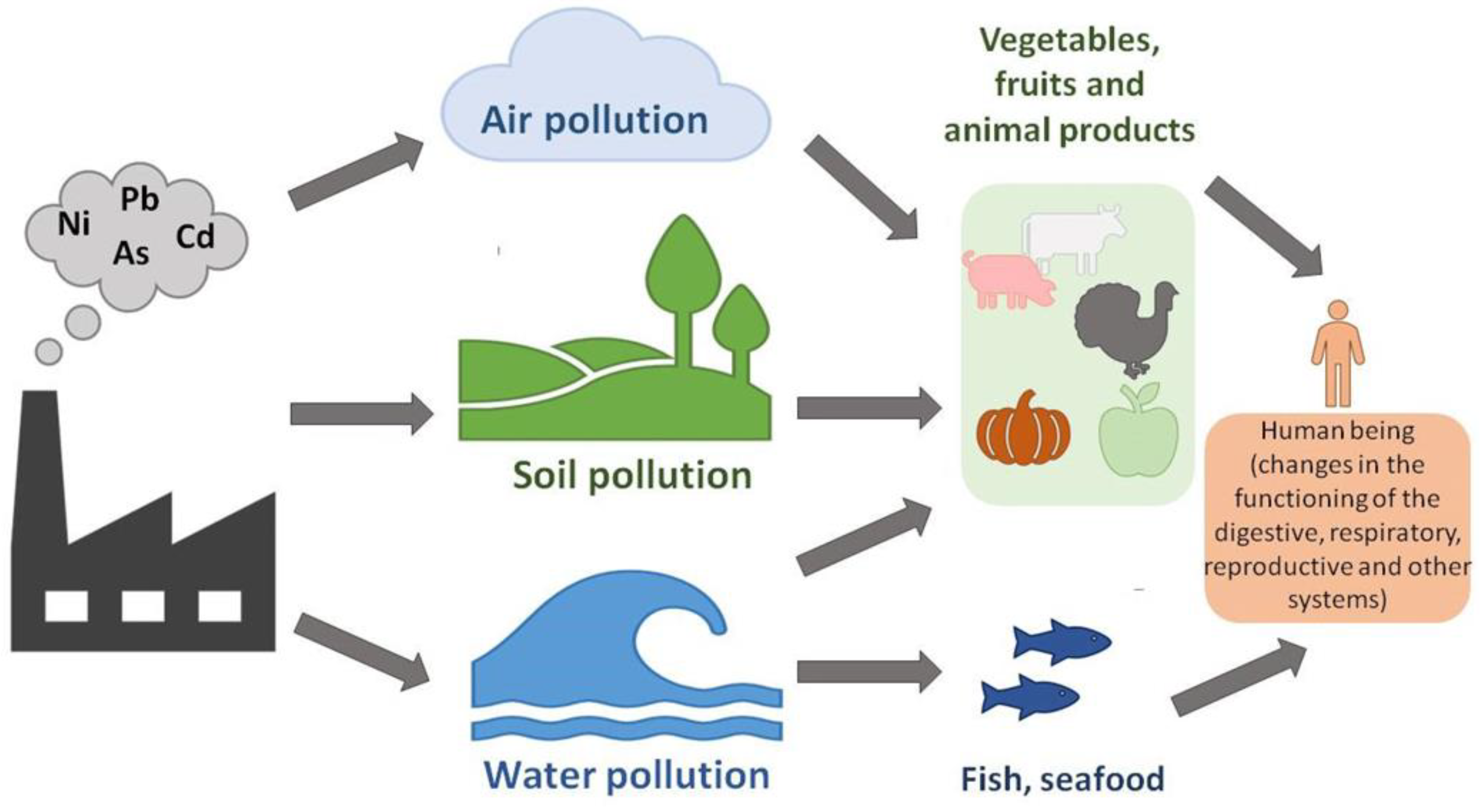
Figure 2.
Cd concentration for 2024 in Europe with the outliers overlaid as black circles. The dimension of the circle indicates the concentration for the given outlier; concentrations above 5 mg/kg−1 are also indicated by their numerical value [24]. As seen from the figure the highest Cd concentration was registered in North Spain.
Figure 2.
Cd concentration for 2024 in Europe with the outliers overlaid as black circles. The dimension of the circle indicates the concentration for the given outlier; concentrations above 5 mg/kg−1 are also indicated by their numerical value [24]. As seen from the figure the highest Cd concentration was registered in North Spain.
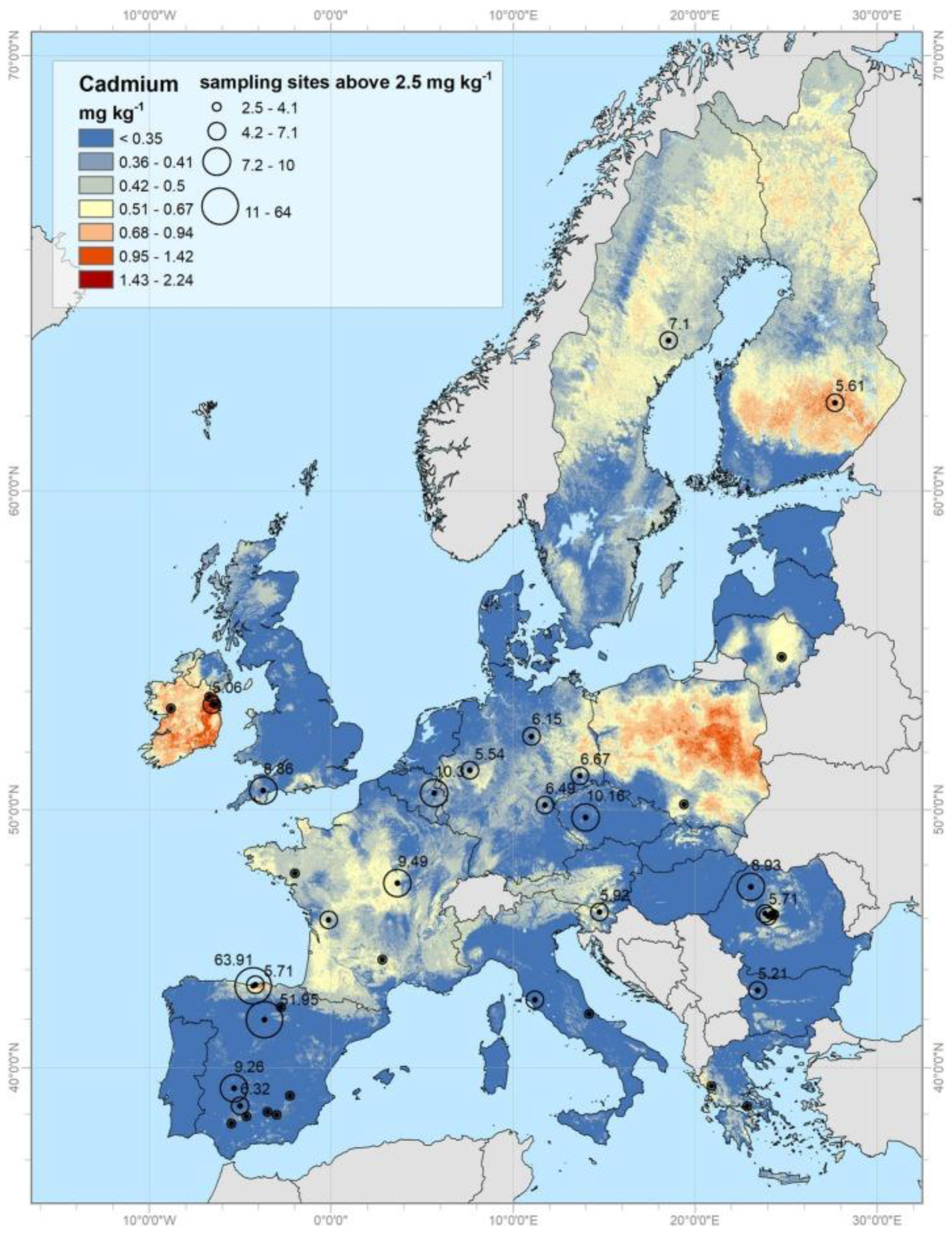
Figure 3.
Czc model for Cd2+, Zn2+, and Co2+ efflux system functioning as proton/cation antiporter consisting of inner membrane (CzcA), outer membrane (CzcC), and membrane fusion (CzcB) proteins functioning as a dimer [40].
Figure 3.
Czc model for Cd2+, Zn2+, and Co2+ efflux system functioning as proton/cation antiporter consisting of inner membrane (CzcA), outer membrane (CzcC), and membrane fusion (CzcB) proteins functioning as a dimer [40].
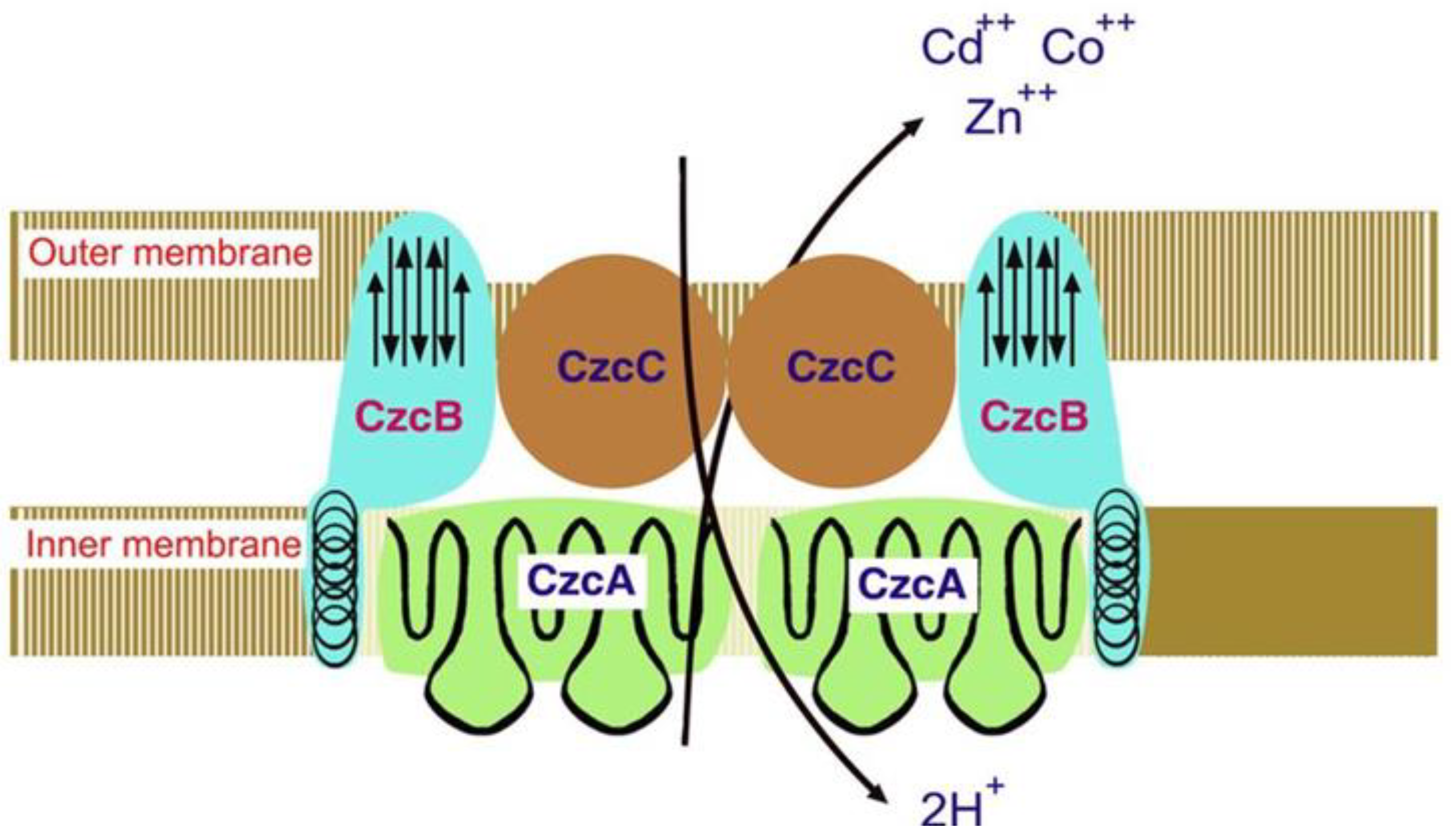
Figure 4.
Sources of Cd and its most significant effects on different parts of the human body [85].
Figure 4.
Sources of Cd and its most significant effects on different parts of the human body [85].
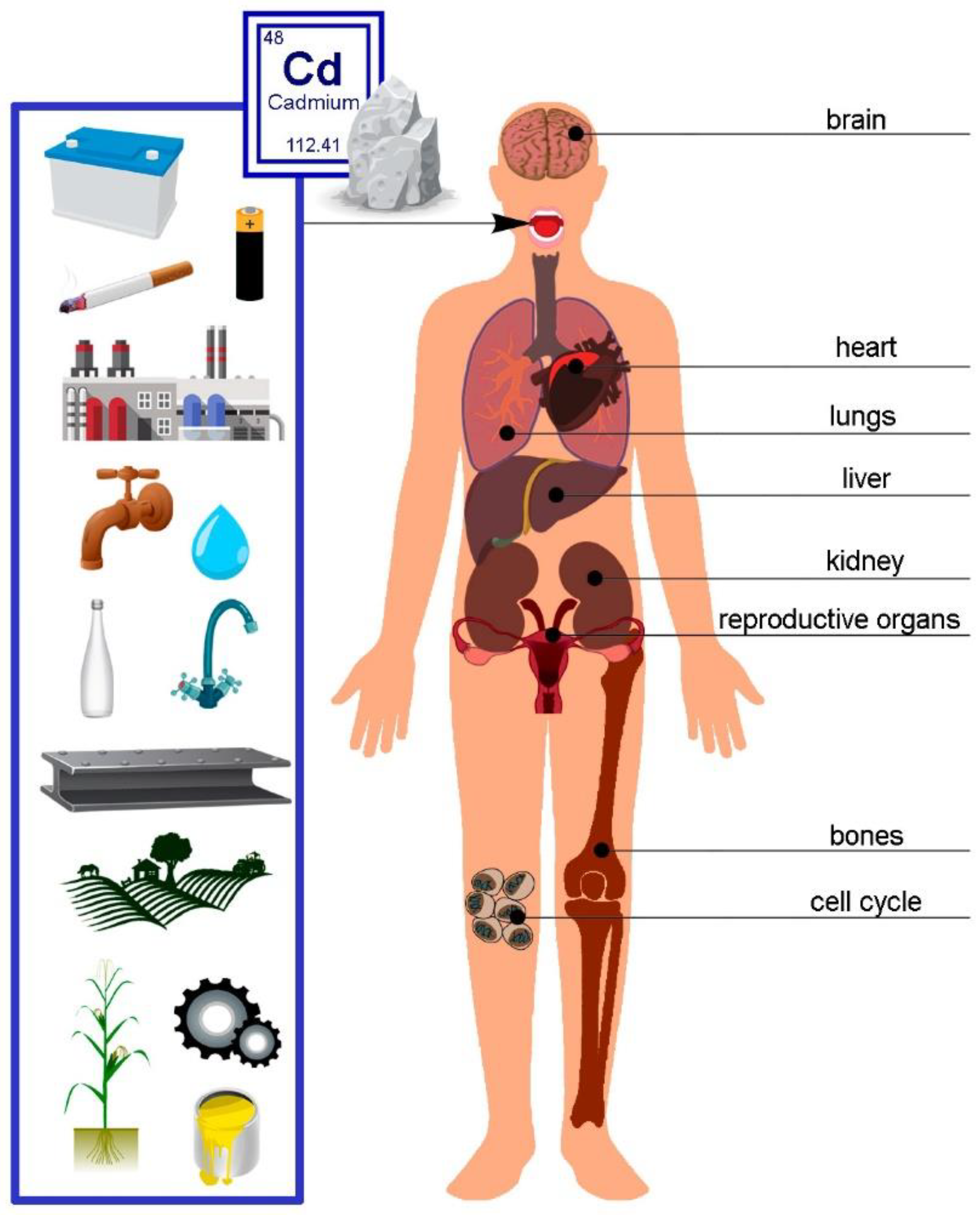
Figure 5.
Cd exposure leads to the development of smoking-related lung diseases [85].
Figure 5.
Cd exposure leads to the development of smoking-related lung diseases [85].
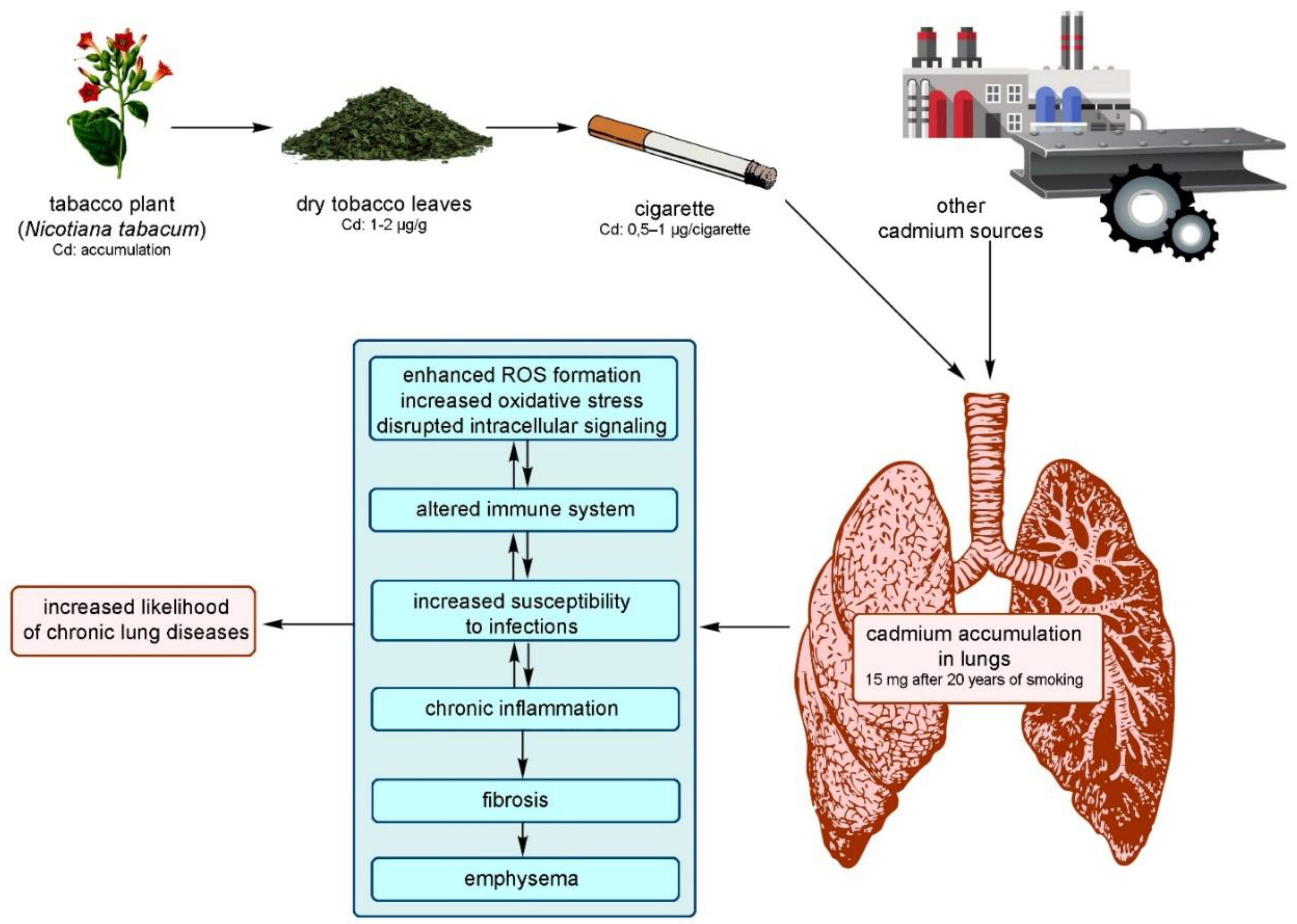
Figure 6.
Primary outcomes in health effects following chronic Cd exposure [1].
Figure 6.
Primary outcomes in health effects following chronic Cd exposure [1].
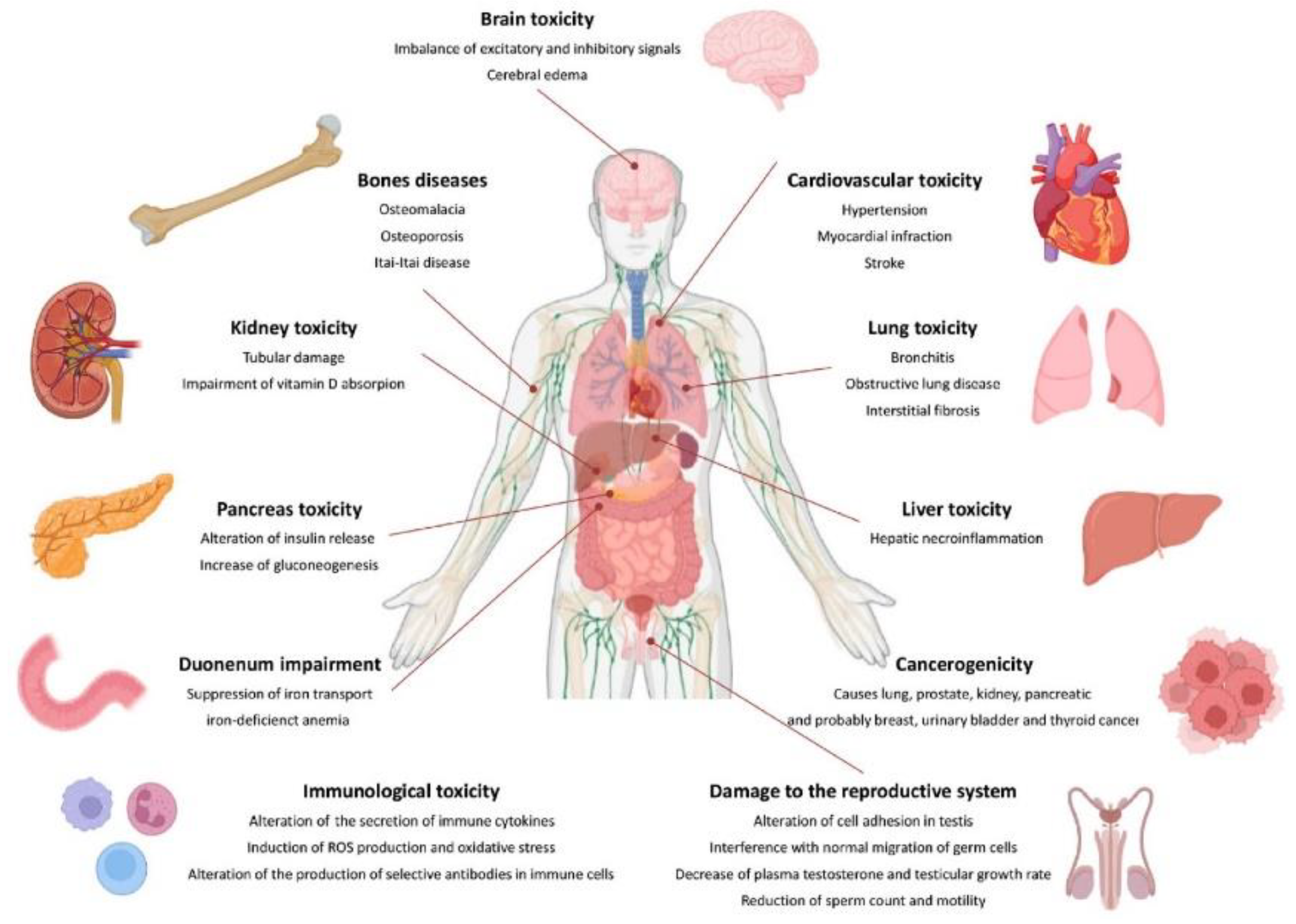
Figure 7.
General mechanisms and specific molecular pathways of Cd toxicity. The figure shows general mechanisms (gene regulation, apoptosis, autophagy, oxidative stress, and interaction with bioelements) along with the specific molecular pathways of Cd toxicity [142].
Figure 7.
General mechanisms and specific molecular pathways of Cd toxicity. The figure shows general mechanisms (gene regulation, apoptosis, autophagy, oxidative stress, and interaction with bioelements) along with the specific molecular pathways of Cd toxicity [142].
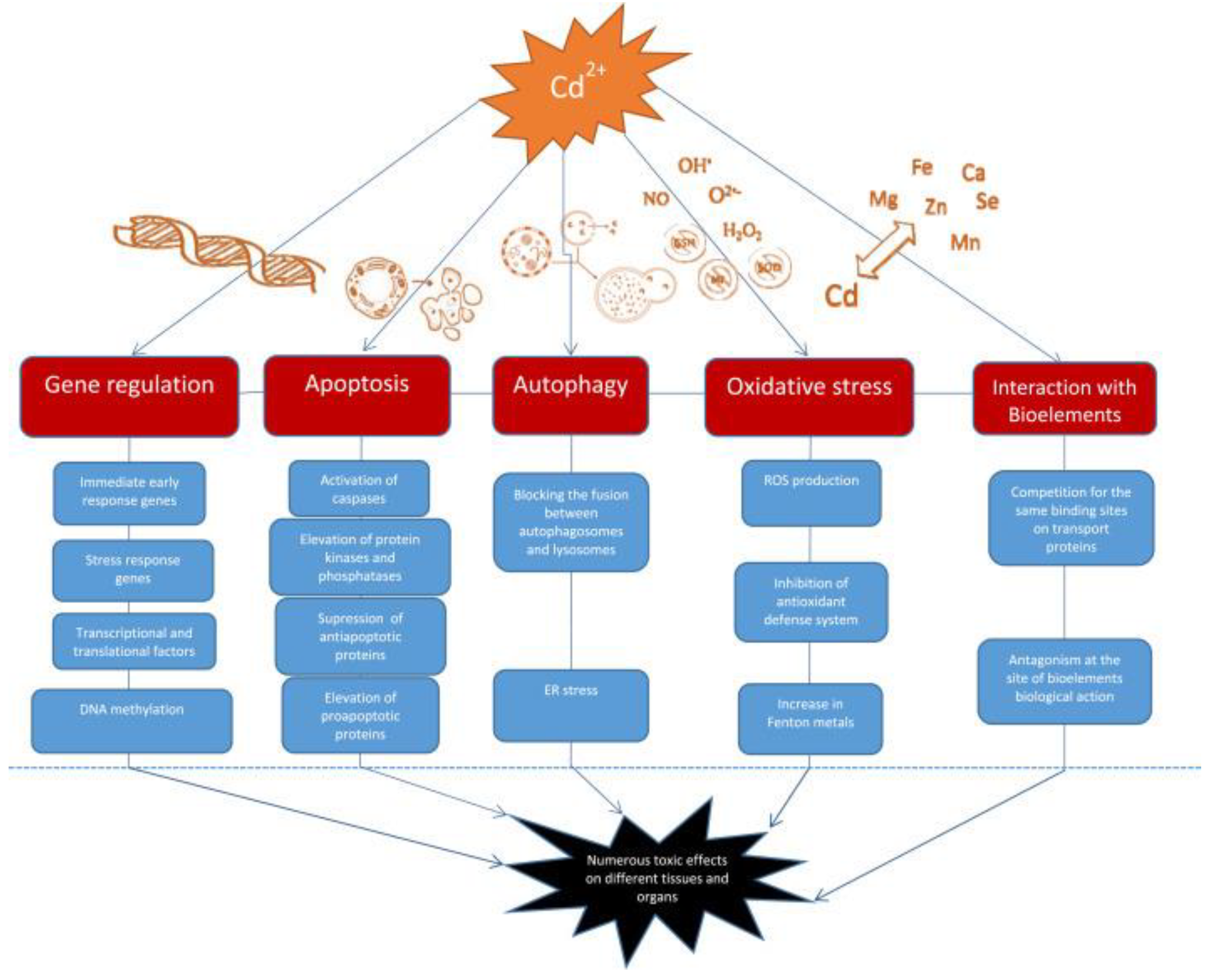
Figure 8.
Interaction sites of environmental metal toxicants with the NMDA receptor are as follows: Cd2+ facilitates neuronal uptake after receptor stimulation. Similarly, Mn2+ can enter the plasma membrane through the NMDAR. Cd can directly attach to the DRPEER sequence in the extracellular domain (in the agonist binding domain (ABD)/ transmembrane domain (TMD) linker) of the GluN1 subunit, inhibiting NMDA-mediated current. Pb competes with zinc for the zinc-binding site of the GluN2 subunit, thereby affecting receptor function. As for Hg, there are indications of interactions with cysteine -SH groups that regulate NMDAR activity, although this suggestion still requires experimental evidence. In most instances, metals may induce ROS, which can interact with the –SH groups of the NMDAR [143,144].
Figure 8.
Interaction sites of environmental metal toxicants with the NMDA receptor are as follows: Cd2+ facilitates neuronal uptake after receptor stimulation. Similarly, Mn2+ can enter the plasma membrane through the NMDAR. Cd can directly attach to the DRPEER sequence in the extracellular domain (in the agonist binding domain (ABD)/ transmembrane domain (TMD) linker) of the GluN1 subunit, inhibiting NMDA-mediated current. Pb competes with zinc for the zinc-binding site of the GluN2 subunit, thereby affecting receptor function. As for Hg, there are indications of interactions with cysteine -SH groups that regulate NMDAR activity, although this suggestion still requires experimental evidence. In most instances, metals may induce ROS, which can interact with the –SH groups of the NMDAR [143,144].
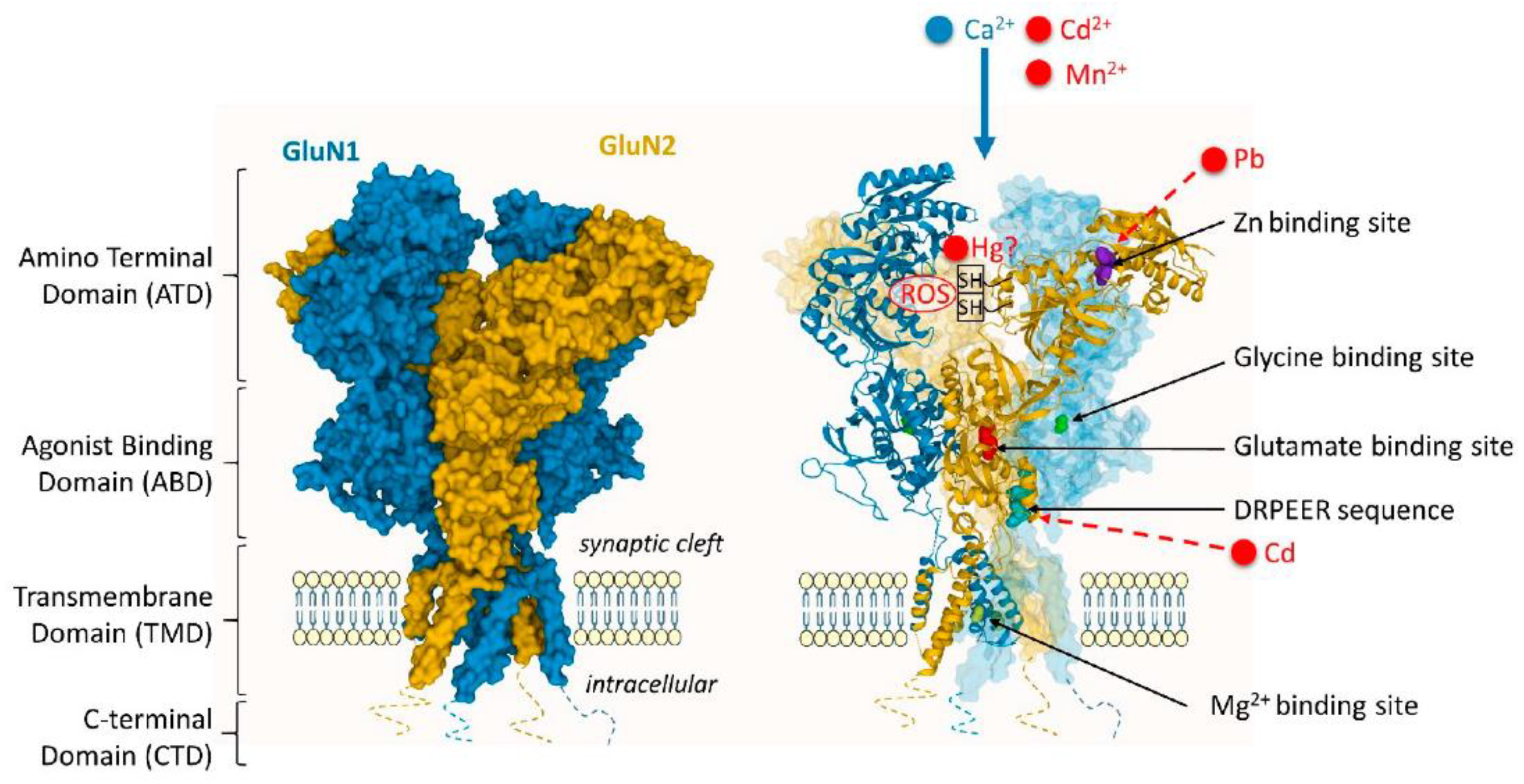
Figure 9.
Cd’s effect on MAPK pathways disrupts cellular signaling and health. The figure demonstrates how Cd2+ disrupts the MAPK signaling pathways, highlighting direct and indirect impacts on cellular function. This leads to abnormal cell responses and an increased risk of disease. Cd's role in activating NF-κB signaling is a pathway to increased inflammation. Cd2+ activates the NF-κB pathway and interacts with ROS, receptors, and kinases, resulting in heightened inflammation and potential chronic conditions. Cd2+ also affects the p53 pathway, which leads to DNA damage responses, including apoptosis and cell-cycle control. The role of p53 is in protecting cellular health. Cd2+ disrupts Ca2+ homeostasis by mimicking and interfering with Ca2+ in cells. The figure depicts the effects of Cd on calcium channels and pumps and the subsequent risks to cell function and activity. The figure also demonstrates the pathways through which Cd2+ influences epigenetic processes, impacting DNA methylation, histone modification, and non-coding RNA activity. These changes lead to significant alterations in gene expression, affecting cellular functions and contributing to disease progression, particularly in cancer and other severe health conditions [146].
Figure 9.
Cd’s effect on MAPK pathways disrupts cellular signaling and health. The figure demonstrates how Cd2+ disrupts the MAPK signaling pathways, highlighting direct and indirect impacts on cellular function. This leads to abnormal cell responses and an increased risk of disease. Cd's role in activating NF-κB signaling is a pathway to increased inflammation. Cd2+ activates the NF-κB pathway and interacts with ROS, receptors, and kinases, resulting in heightened inflammation and potential chronic conditions. Cd2+ also affects the p53 pathway, which leads to DNA damage responses, including apoptosis and cell-cycle control. The role of p53 is in protecting cellular health. Cd2+ disrupts Ca2+ homeostasis by mimicking and interfering with Ca2+ in cells. The figure depicts the effects of Cd on calcium channels and pumps and the subsequent risks to cell function and activity. The figure also demonstrates the pathways through which Cd2+ influences epigenetic processes, impacting DNA methylation, histone modification, and non-coding RNA activity. These changes lead to significant alterations in gene expression, affecting cellular functions and contributing to disease progression, particularly in cancer and other severe health conditions [146].
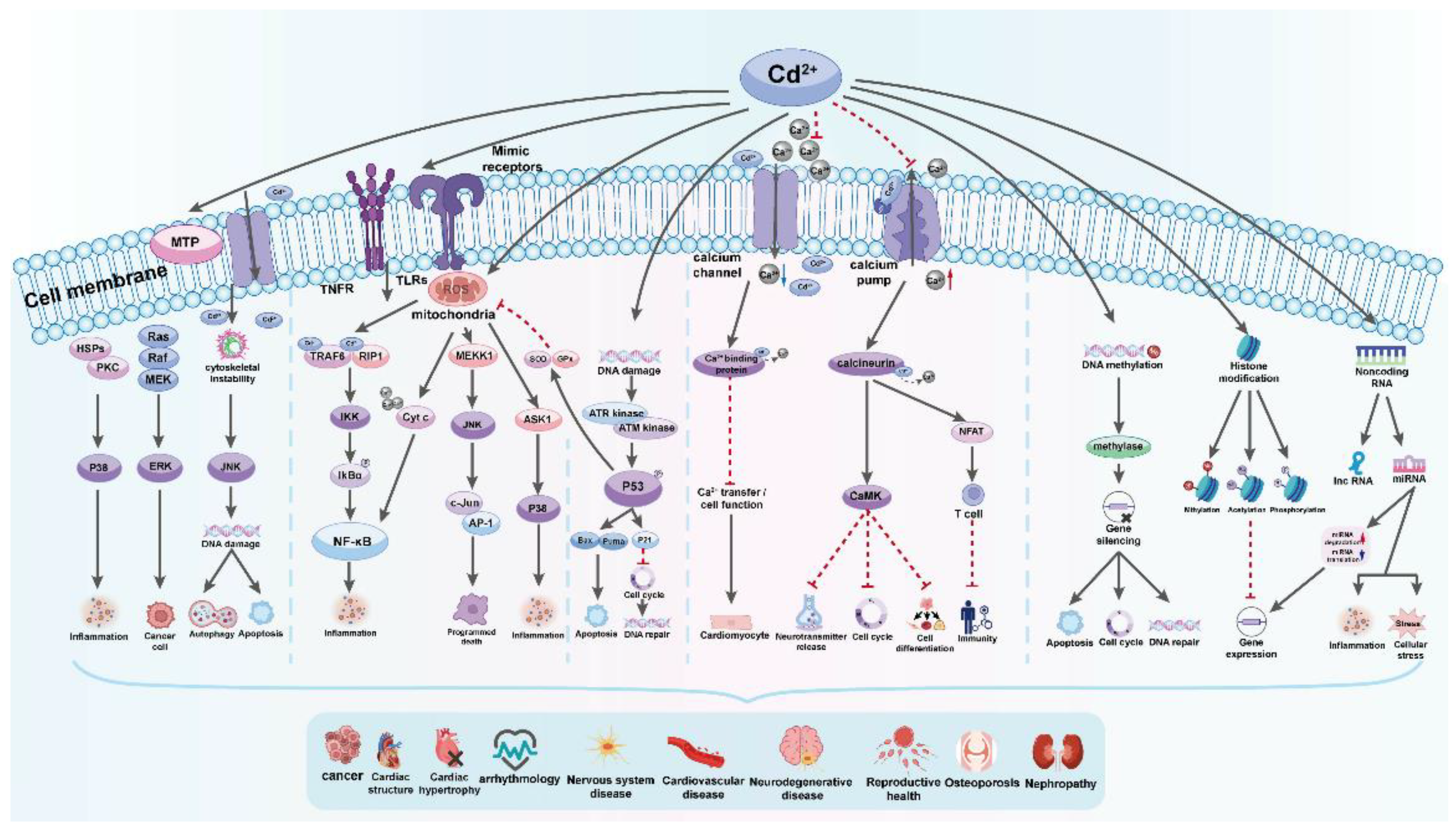

Table 1.
Physical and chemical properties of Cd [4].
Table 1.
Physical and chemical properties of Cd [4].
| Atomic number | 48 |
| Atomic weight | 112.41 u |
| Atomic radius | 155 pm |
| Electronic configuration | [Kr]4d105s2 |
| Melting point | 321.07 °C |
| Boiling point | 767.3 °C |
| Density at 20 °C | 8.65 g/cm3 |
| Reduction potential Cd2+ + 2e− → Cd(s) | −0.40 E° |
| Heat of fusion | 6.21 kJ/mol |
| Heat of vaporization | 99.6 kJ/mol |
| Electronegativity (Pauling scale) | 1.69 |
| First ionization energy | 867.8 kJ/mol |
| Second ionization energy | 1631.4 kJ/mol |
Table 2.
Cd+2 removal (%)/uptake (mM/g) potential in some bacterial and yeast strains [45].
Table 2.
Cd+2 removal (%)/uptake (mM/g) potential in some bacterial and yeast strains [45].
| Organism | Cd+2 resistance (mM) |
Cd+2 removal (%) uptake (mM/g) |
|---|---|---|
| Klebsiella pneumoniae | 13.3 | 57.4 |
| Escherichia coli P4 | 10.6 | 56 |
| Salmonella enterica 43C | 13.3 | 22 |
| Bacillus sp. | – | 50 |
| Rhodobacter sphaeroides | – | 30.7 |
| Microbacterium oxydans CM3 | – | 43 |
| Candida tropicalis | 25.0 | 92 |
| Pichia hampshirensis 4Aer | 24.0 | 28 |
| Candida tropicalis 3Aer | 25.1 | 31 |
| Trametes versicolor | 5.0 | 0.300 |
| Trichosporon ashii | 10 | 78 |
| Pichia kudriavzevii | 15 | 61 |
Table 3.
Plant species show varying responses to Cd-induced DNA damage. This damage includes DNA strand breaks, chromosomal aberrations, and micronuclei formation. Cd also impacts the expression of DNA repair genes and causes alterations in amplified fragment length polymorphism (AFLP), inter-simple sequence repeat (ISSR), random amplified polymorphic DNA (RAPD), sequence-related amplified polymorphism (SRAP), and simple sequence repeat (SSR) profiles, leading to a reduction in genomic template stability (GTS). The symbols ↑ and ↓ indicate increases and decreases, respectively [59].
Table 3.
Plant species show varying responses to Cd-induced DNA damage. This damage includes DNA strand breaks, chromosomal aberrations, and micronuclei formation. Cd also impacts the expression of DNA repair genes and causes alterations in amplified fragment length polymorphism (AFLP), inter-simple sequence repeat (ISSR), random amplified polymorphic DNA (RAPD), sequence-related amplified polymorphism (SRAP), and simple sequence repeat (SSR) profiles, leading to a reduction in genomic template stability (GTS). The symbols ↑ and ↓ indicate increases and decreases, respectively [59].
| Species | Organ | Cd Concentration |
Exposure Duration |
Effect |
|---|---|---|---|---|
| Allium cepa | Root tip | 50–200 µM | 2 h + 24 h recovery |
Micronucleus formation Chromosomal aberrations |
| Root tip | 25 µM | 48 h | ||
| Root tip | 25 µM | 48 h | % tail DNA ↑ | |
| Arabidopsis thaliana | Root tip | 0.125–2.5 mg L−1 | 5 d | Altered expression DNA repair genes |
| Root | 1.25–4 mg L−1 | 5 d | Altered RAPD profile Altered expression DNA repair genes |
|
| Leaf | 0.5–5 mg L−1 | 16 d | Altered AFLP profile | |
| Leaf | 0.25–8 mg L−1 | 15 d | Microsatellite instability Altered RAPD profile |
|
| Leaf | 5 µM | 72 h | Altered expression DNA repair genes | |
| Brassica chinensis | Leaf | 15–120 mg kg−1 soil | 30 d | Altered RAPD profile |
| Brassica oleracea | Root | 2.5–20 mg kg−1 soil | 3–56 d | Altered % tail intensity |
| Capsicum annuum | Root tip | 20–100 ppm | 24 h | Chromosomal aberrations |
| Leaf | 20–100 ppm | 24 h | Altered RAPD profile | |
| Hordeum vulgare | Root tip | 75–225 µM | 7 d | Altered RAPD profile (GTS ↓) |
| Leaf | 5 µM | 15 d | DNA damage ↑ | |
| Ipomoea aquatica | Entire seedling | 15–120 mg kg−1 soil | 21 d | Altered RAPD profile (GTS ↓) |
| Lactuca sativa | Root tip | 25 µM | 48 h | Chromosomal aberrations Micronucleus formation % DNA damage ↑ |
| Lathyrus sativus | Root tip | 5–50 µM | 3–7 d | Chromosomal aberrations Micronucleus formation |
| Leucaena leucocephala | Leaf | 50 mg L−1 | 15 d | Altered RAPD profile |
| Nicotiana tabacum | Root and leaf | 10–15 µM | 7 d | % tail DNA ↑ |
| Oryza sativa | Root tip | 50–200 µM | 48–96 h | Altered SRAP profile (GTS ↓) |
| Sphagnum palustre | Shoot | 0.1–10 µM | 24–48 h | Altered ISSR profile (GTS ↓) |
Table 4.
An analysis of the effects of Cd on cell cycle-related parameters organized by plant species. Exposure to Cd decreases the mitotic index (i.e., the ratio of cells undergoing mitosis to the total number of cells) and changes in nuclear ploidy levels. It influences the expression of cell cycle-related genes in various plant species. The symbols ↑ and ↓ represent increases and decreases, respectively [59].
Table 4.
An analysis of the effects of Cd on cell cycle-related parameters organized by plant species. Exposure to Cd decreases the mitotic index (i.e., the ratio of cells undergoing mitosis to the total number of cells) and changes in nuclear ploidy levels. It influences the expression of cell cycle-related genes in various plant species. The symbols ↑ and ↓ represent increases and decreases, respectively [59].
| Species | Organ | Cd Concentration |
Exposure Duration |
Effect |
|---|---|---|---|---|
| Allium cepa | Root tip | 50–200 µM | 2 h + 24 h recovery |
Mitotic index ↓ |
| Root tip | 25 µM | 48 h | Mitotic index ↓ | |
| Arabidopsis thaliana | Root tip | 0.125–2.5 mg L−1 | 5 d | 2C ↓, 4C ↑, 8C ↑ Altered cell cycle phase distribution Altered expression cell cycle-related genes |
| Root | 1.25–4 mg L−1 | 5 d | 2C ↓, 4C ↑ Altered expression of cell cycle-related genes |
|
| Leaf | 5 µM | 3–12 d | Endoreduplication factor ↓ Epidermal cell number and cell surface area ↓ Altered expression of cell-cycle related genes |
|
| Capsicum annuum | Root tip | 20–100 ppm | 24 h | Mitotic index ↓ |
| Lactuca sativa | Root tip | 25 µM | 48 h | Mitotic index ↓ |
| Lathyrus sativus | Root tip | 5–50 µM | 3–7 d | Mitotic index ↓ |
| Oryza sativa | Root | 200 µM | 7 d | Cortex cell length in the elongation zone ↓ Cortex cell number in the elongation zone ↓ Altered expression of cell cycle-related genes |
| Sorghum bicolor | Root tip | 50–200 µM | 5 d | Inhibition of S phase progression |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



