
Submitted:
16 October 2024
Posted:
17 October 2024
You are already at the latest version
Abstract
Salt stress is an environmental factor that limits plant seed germination, growth, and survival. We performed a comparative RNA sequencing transcriptome analysis of the seeds during germination from two cultivars with contrasting salt tolerance responses. A transcriptomic comparison between salt-tolerant cotton cv Jin-mian 25 and salt-sensitive cotton cv Su-mian 3 revealed both similar and differential expression patterns between the two genotypes during salt stress. The expression of genes related to aquaporins, kinases, reactive oxygen species (ROS) scavenging, trehalose biosynthesis, and phytohormone biosynthesis and signaling that include ethylene (ET), gibberellin (GA), abscisic acid (ABA), jasmonic acid (JA), and brassinosteroid (BR) were systematically investigated between the cultivars. Despite the involvement of these genes in cotton's response to salt stress in positive or negative ways, their expression levels were mostly similar in both genotypes. Interestingly, a PXC2 gene (Ghir_D08G025150) was identified, which encodes a leucine-rich repeat receptor-like protein kinase (LRR-RLK). This gene showed an induced expression pattern after salt stress treatment only in salt-tolerant cv Jin-mian 25, but not salt-sensitive cv Su-mian 3. Our multifaceted transcriptome approach illustrated a differential response to salt stress between salt-tolerant and salt-sensitive cotton.
Keywords:
Salt Stress
Introduction
Cotton is a natural, high-quality fiber source and an important oil crop, which has a very important role in China’s economic development [1]. However, in recent years, there has been a rapid increase in population, leading to an increasing demand for cotton, which has resulted in the expansion of cotton cultivation scale [2]. The contradiction between “competing for land between cotton and grain” has become increasingly severe [3]. Human activities and global climate change have made soil salinization a more serious issue, with the global area of saline land reaching 9.5×108 hm2 and growing by more than 1 million hectares per year [4]. Currently, China is severely affected by soil salinization, with the total area of saline soil estimated at around 99 million hectares, and soil salinization is one of the important factors affecting cotton yield and quality [5], seriously affecting agricultural production and social and economic development. Screening and cultivating salt-tolerant cotton varieties is one of the most economical and effective ways to utilize salinized soil. By transferring cotton cultivation to saline and barren land, further expanding the cotton planting area, it has great significance for the cotton industry and economic development.
Cotton is a relatively salt-tolerant crop and a pioneering crop for saline land improvement, but its salt tolerance varies among developmental stages and tissues of different genotypes [6]. High salt stress inhibited cotton growth, reduced cotton yield and fiber quality [7]. Soil salt stress reduces the vitality of cotton root systems and root water absorption ability, inhibiting water transport from roots to leaves. In the NaCl simulated saline stress treatment, the increase in Cl-, K+, Ca2+, and N content in leaves is positively correlated with cotton tolerance (Ashraf and Ahmad 2000). In addition, the activities of antioxidant enzymes, enzymes involved in secondary metabolism, active oxygen content, osmotic adjustment substance content, and plant hormones play important roles in regulating cotton’s response to salt stress [8]. Researchers have found that in saline stress treatment, salt-tolerant cotton cultivars are compared with salt-sensitive cultivars, with higher activities of antioxidant enzymes and enzymes involved in secondary metabolism, lower active oxygen content, higher osmotic adjustment substance (such as proline) and secondary metabolite content [9]. Salt stress causes ion imbalance in plant tissues, with salt-sensitive cultivars accumulating more Na+ but less K+ in their leaves compared to salt-tolerant cultivars. In contrast, the changes in ion homeostasis of salt-tolerant cultivars are not obvious, and the K+/Na+ ratio increases [10]. Salt stress led to changes in the content of fatty acids and hormones in cotton leaves, such as an increase in palmitic acid, stearic acid, and oleic acid, and a decrease in linoleic acid and linolenic acid [11]. Therefore, comprehending the molecular mechanism of salt tolerance in cotton and exploring key salt-tolerance genes is crucial for breeding resilient, salt-tolerant cotton cultivars.
Currently, with the completion of cotton genomic sequencing and assembly, the advancement of cotton functional genomics has been accelerated [12,13,14,15,16]. Based on the genomic data, transcriptomics has also made some progress in revealing the mechanisms of cotton salt tolerance and candidate gene identification. Transcriptomics has revealed some important genes that respond to salt stress in cotton, including membrane receptor proteins, transporters, transcription factors, CDPK, MAPK signal cascades, and hormone biosynthesis and signal transduction [17,18]. In diploid cotton species Gossypium davidsonii with superior stress tolerance, transcriptome data elucidated that salt overly sensitive (SOS) and ROS signaling pathways were closely related. Furthermore, photosynthesis pathways and metabolism play important roles in ion homeostasis and oxidation balance in salt stress tolerance [19]. Comparative transcriptome sequencing analysis was conducted on two landrace cotton cultivars with different salt tolerance, and it was found that some biological processes such as transcriptional regulation, signal transduction, and secondary metabolism exist in a markedly different manner between the two cultivars, providing scientific evidence for explaining the differences in salt tolerance between the two cultivars [20]. Integrative transcriptomic analyses revealed that ion transport, hormone metabolism, and ROS scavenging pathways played important roles in cotton root adaptation to salt stress [21]. Using single-cell transcriptomics technology to systematically analyze the response of cotton roots to salt stress revealed that differentially expressed genes (DEGs) identified were concentrated on plant-type primary cell wall biogenesis, defense response, phenylpropanoid biosynthesis, transcription factors, and plant hormone metabolic pathways [22]. These results compared the transcriptomic expression profiles of cotton seedlings exposed to salt stress using RNA sequencing and revealed that cotton has different regulatory mechanisms in response to salt stress.
Genetic engineering and expressing of stress-resistant genes in cotton have also been proven to be an effective method for improving cotton salt stress resistance [23]. For instance, overexpressing the rice stress-resistant gene SNAC1 in cotton can significantly improved the transgenic cotton’s resistance to salt stress [24]. Overexpressing the Artemisia vulgaris stress response gene AvDH1 in cotton can reduce damage caused by salt stress and increase productivity in saline fields [25]. The GhDof1 DOF transcription factor gene of the cotton family was induced to express under salt stress, and overexpressing GhDof1 can enhance cotton’s resistance to salt stress [26]. Transgenic ApGSMT2g and ApDMT2g cotton showed higher salt tolerance and more seed cotton yield in saline fields compared to wild-type [27]. The bZIP family transcription factor participated in the regulation of cotton’s resistance to salt stress, and overexpressing the GhABF2 and GhABF3 genes in cotton can improve its tolerance to salt stress by regulating the expression of some stress-related genes and increasing the activity of antioxidant enzymes [28,29]. These results demonstrated that using biotechnology to manipulate key candidate genes may be an important strategy for enhancing the stress resistance of cotton.
Despite these efforts, the regulatory molecular network of salt tolerance genes in cotton remains unclear. Furthermore, to our knowledge, there has been no time-course comparative transcriptome analysis of the effect of salt stress in cotton. In this study, we explored the transcriptional responses of salt-tolerant and salt-sensitive cotton cultivars to salt stress. We aimed to compare the transcriptional responses associated with phytohormone signaling, regulation of gene expression related to aquaporins, kinases, and ROS scavenging, and trehalose metabolism following the salt stress of two contrasting cotton varieties at different seed germination stages to better understand the mechanism involved in the response to salt stress. Our findings exposed temporal and genotype-specific responses to salt stress and identified critical salt-tolerant metabolic pathways and promising candidate genes for the engineering of salt-tolerant cotton.
Materials & Methods
Plant Materials and Experiment Design
The salt-tolerant cotton Jin-mian 25 and salt-sensitive cotton Su-mian 3 were used in this study. Plump cotton seeds with evenly sized were selected, and sterilized by soaking in 10% hydrogen peroxide for 30 minutes. Then rinse them with sterile water 4-5 times and use filter paper to absorb moisture from the seed surface. Then, the seeds from both genotypes were selected and treated for 0h, 6h, 12h, 24h, and 72h with 150 mM NaCl solution. The seeds or the seedlings were collected for transcriptome analyses at the corresponding treatment time. All samples (30 samples) were frozen in liquid nitrogen and stored at - 80 °C. Three biological replicates were performed in this study.
Measurement of Phenotypic Parameters
The cotton seeds were placed parallelly on the rectangular filter papers soaked in distilled water and a 150mmol-1 NaCl solution, respectively. Then, place another soaked filter paper on top of the seeds and fold it in half to roll it into a cylinder, repeating this process three times and tying the bottom with an elastic band. Place the cylinders in a germination box filled with 3 cm of salt solution, with the control group being distilled water. Place the germination box in the seed germination chamber, with a temperature of 30±1°C and relative humidity of 60% under 12 h/12 h light/dark (light intensity ~ 16,000 LUX) cycle. The germination potential and germination rate of the seeds were calculated on days 3 and 7. On the seventh day, five seedlings were randomly selected to calculate the fresh weight, seedling length, root length, and hypocotyl length, with each treatment repeated three times.
RNA Extraction and Sequencing
Total RNA from 30 samples was extracted using the RNAprep pure plant kit (TIANGEN, China) according to the manufacturer’s instructions. Su-mian 3 and Jin-mian 25 seeds exposed to salt stress for 0h, 6h, 12h, 24h and 72h were named SS0/ST0, SS6/ST6, SS12/ST12, SS24/ST24, and SS72/ST72, respectively. The cDNA sequencing libraries were constructed using the AMPure XP system (Beckman Coulter, Beverly, USA). All libraries were sequenced on an Illumina NovaSeq 6000 platform using 150 bp paired-end reads. Three biological replicates were performed.
Transcriptome Data Analysis
The raw data in fastq format was processed with the FASTX-Toolkit (http://hannonlab.cshl.edu/fastx_toolkit/). Clean data (clean reads) were obtained by removing low-quality reads, reads containing adapters, and poly-N from the raw data. Concurrently, the Q30 and GC content of the clean data were calculated. All subsequent analyses were based on high-quality clean data. Clean reads from each sample were then mapped to the reference genome for the cotton cultivar TM-1 [12] using HISAT2 software [30,31]. Reads with at most one mismatch were utilized to calculate the gene expression levels. Gene expression values were calculated following the method of the previous study [32,33]. DEGs between two samples were identified using DESeq2 and presented using fragments per kilobase of transcript per million fragments mapped (FPKM) [34]. The resulting p values were adjusted using Benjamini and Hochberg’s approach for controlling the false discovery rate (FDR). Genes with an adjusted p value < 0.01 and |log2fold change|≥ 1 were defined as differentially expressed. Clean data were available from the Genome Sequence Archive in the BIG Data Center of Sciences (https://bigd.big.ac.cn/) under accession number CRA017659.
Gene Functional Annotation Analyses
The functions of DEGs were annotated using the following databases: Nr (NCBI nonredundant protein sequences, ftp://ftp.ncbi.nih.gov/blast/db/), Gene Ontology (GO) (Gene Ontology, http://www.geneontology.org/), and Kyoto Encyclopedia of Genes and Genomes (KEGG) (http://www.genome.jp/kegg/). KEGG pathway analysis of DEGs was performed with BLAST software and KEGG enrichment was analyzed using KOBAS 2.0 software with p-value < 0.05 [35]. GO analysis of DEGs was carried out using the GOseq R package [36].
Data Processing, Visualization, and Statistical Analysis
The bar diagrams were plotted using Graphpad Prism 6.0. Venn diagram was drawn using an online tool (https://bioinformatics.psb.ugent.be/webtools/Venn/). Heat maps was drawn to display the gene expression patterns of the FPKM values by using the TBtools software [37]. The R package available at https://www.r-project.org/ was utilized for the student’s t-test. The Shapiro-Wilk test was used to assess normality, confirming that the data followed a Gaussian distribution. The least significant difference (LSD) was employed to assess significance at either the 1% or 5% levels. The analysis involved a minimum of three biological replicates for each sample.
Results
Salt Sensitivity Assessment of Su-Mian 3 and Jin-Mian 25 Cotton Cultivars
The seeds of Su-mian 3 and Jin-mian 25 cotton were treated using distilled water and 150mM NaCl solution. The germination potential and germination rate of seeds as well as the fresh weight, seedling length, root length, and hypocotyl length of seedlings were determined after 7 days of treatment (Figure 1). As shown in Figure 1, there was a significant difference in the salt tolerance testing between the two cotton cultivars. Compared to the control (Figure 1A), the growth of Su-mian 3 seedlings was severely inhibited (Figure 1B) with its fresh weight (Figure 1C), seedling length (Figure 1D), root length (Figure 1E), and hypocotyl length (Figure 1F) significantly reduced by about 58%, 63%, 58%, and 73%, respectively. Meanwhile, the germination potential (Figure 1G) and germination rate (Figure 1H) of Su-mian 3 seeds were significantly decreased by about 86% and 58%. However, Jin-mian 25 grew well under 150mM salt stress treatment, and there was no obvious difference in the growth of seedlings (Figure 1I,J). Overall, only the root length was significantly reduced by about 12% (Figure 1M), and other indicators including fresh weight (Figure 1K), seedling length (Figure 1L), hypocotyl length (Figure 1N), germination potential (Figure 1O), and germination rate (Figure 1P) were hardly affected by salt stress and had no significant differences between the control and salt treatment. The above results indicated that Su-mian 3 was a salt-sensitive (SS) cultivar, while Jin-mian 25 was a salt-tolerant (ST) cultivar.
Transcriptome Profiles of Su-Mian 3 and Jin-Mian 25 Cotton Plants Exposed to Salt Stress
To study how cv Su-mian 3 and Jin-mian 25 cotton plants respond to salt stress at the gene expression level, we analyzed the transcriptome profiles of the salt-sensitive cotton cv Su-mian 3 and the salt-tolerant cotton cv Jin-mian 25 (Figure S1). It’s known that seed germination and seedling growth were the stages most affected by salt stress [38]. Hence, cv Su-mian 3 and Jin-mian 25 plants at the seed germination and seedling growth stages were used for the transcriptome profiling analyses. The seeds were completely submerged by 150mM NaCl solution for 0, 6, 12, 24, and 72h, and then sampled for expression profile analyses in response to salt stress (Figure 2A). On average, 27,002,000 paired-end reads were generated per sample, resulting in a total of 243 Gb of high-quality clean bases obtained from the 30 samples (Table S1). Using the G. hirsutum cv TM-1 reference genome [12], the average rates of mapped reads for cv Su-mian 3 and Jin-mian 25 samples were 92.70% and 92.08%, respectively (Table S2).
As these two cultivars showed great variance in response to salt stress, we evaluated gene expression at five time points (0, 6, 12, 24, and 72h) during salt treatment from cv Su-mian 3 and Jin-mian 25, hereafter referred to as SS0, SS6, SS12, SS24, SS72, ST0, ST6, ST12, ST24, and ST72, respectively. A total of 12,237 DEGs were identified across five comparisons: SS0 vs ST0, SS6 vs ST6, SS12 vs ST12, SS24 vs ST24, and SS72 vs ST72 (Figure 2B). Specifically, 444 genes were significantly upregulated in ST0 compared to SS0, while 354 genes were significantly downregulated. In comparing ST6 with SS6, 2,428 genes were significantly upregulated and 4,100 genes were significantly downregulated. Furthermore, in the comparison between ST12 and SS12, 1,691 genes were significantly upregulated and 1,580 genes were significantly downregulated. On the other hand, when compared to the SS24 group, the ST24 group exhibited 83 upregulated genes and 105 downregulated genes. Similarly, when compared to the SS72 group, the ST72 group had 715 upregulated genes and 737 downregulated genes (Figure 2B).
Pathway Enrichment of DEGs
Next, we conducted KEGG pathway enrichment analysis to investigate the biological functions of these DEGs during salt stress in both genotypes. The results showed that these DEGs were mainly involved in protein processing in the endoplasmic reticulum, starch and sucrose metabolism, plant-pathogen interaction, plant hormone signal transduction, glyoxylate and dicarboxylate metabolism, glycolysis/gluconeogenesis, MAPK signaling pathway-plant, cysteine and methionine metabolism, alpha-linolenic acid metabolism (Figure 3A-E), suggesting that these metabolic pathway genes might confer the resistance to salt stress in cv Jin-mian 25 cotton.
Additionally, the Venn diagram showed that there were 18 shared DEGs between the two genotypes across the five time points (Figure 3F). Out of these 18 shared DEGs, 8 genes had their expression levels suppressed by salt stress, and 10 genes had their expression induced (Figure 4A). This includes the PXC2 gene (Ghir_D08G025150), which encodes the leucine-rich repeat receptor-like protein kinase (LRR-RLK). This gene showed an induced expression pattern at 12, 24, and 72h time point after salt stress treatment only in cv Jin-mian 25, with higher transcripts compared to cv Su-mian 3 (Figure 4A). This finding suggests that further functional verification of this gene is needed.
Expression Patterns of Genes Related to Aquaporins, Kinases, and ROS Scavenging during Salt Stress in Both Genotypes
To further mine DEGs related to salt stress, we conducted a search for relevant pathways in databases using the keywords “salt” and “salt stress”, which helped us narrow down the list of candidate genes. The pathways we looked at included: response to salt stress (GO:0009651), negative regulation of response to salt stress (GO:1901001), regulation of response to salt stress (GO:1901000), cellular response to salt stress (GO:0071472), positive regulation of response to salt stress (GO:1901002), and response to salt (GO:1902074) from the GO database, as well as the MAPK signaling pathway-plant from the KEGG database. By using this approach, we were able to reduce the number of identified DEGs related to salt stress to 347.
In a recent study, it was found that a Gγ protein regulated the phosphorylation of aquaporins. These aquaporins were channels that can transport hydrogen peroxide to help alleviate oxidative stress caused by salt stress [39]. Based on this finding, we investigated the expression patterns of genes related to aquaporins, kinases, and the scavenging of ROS during salt stress in both genotypes. 21 aquaporin transcripts were detected in our data set, including 11 PIP, 7 TIP, 2 NIP and 1 SIP family genes (Figure 4B). In both genotypes, 19 aquaporin genes showed low to high expression levels, especially showing an induced expression at 72h time point after salt stress. However, the expression levels of two TIP3-2 family genes were inhibited by salt stress in both genotypes (Figure 4B). These results indicated the diverged expression patterns of aquaporin genes in response to salt stress.
The process of protein phosphorylation was typically carried out by kinases [40]. We identified 24 kinase genes that responded to salt stress in our data set, which included eight receptor-like protein kinase (RLK), four leucine-rich repeat receptor-like protein kinase (LRR-RLK), one proline-rich receptor-like protein kinase (PERK), two CBL (calcineurin B-like)-interacting serine/threonine-protein kinase 1 (CIPK1), five CIPK6, and four CIPK9 family genes (Figure 4C, D). When compared to the 0h time point, the expression levels of eight RLK genes were induced by salt stress in both genotypes, with the highest levels observed at 72h time point (Figure 4C). Among the four LRR-RLK genes that positively responded to salt stress at different time points, the PXC2 gene which was expressed much higher in cv Jin-mian 25 than in Su-mian 3, and was induced at all four time points after salt stress in cv Jinmian 25 (Figure 4C). Simultaneously, the expression level of a PERK9 gene decreased gradually with the duration of salt stress in both genotypes, indicating that it negatively regulated salt stress (Figure 4C).
It has been reported in both Arabidopsis and cotton that CIPK6 kinase acted as a positive regulator of stress-responsive genes under salt stress [41,42,43]. The five CIPK6 genes identified here all exhibited the induced expression patterns at 12, 24 or 72h time points after salt stress treatment in both genotypes (Figure 4D). In contrast, the expression levels of CIPK1 and CIPK9 genes were induced only at the 72h time point of salt stress in both genotypes (Figure 4D).
ROS serves as a signal molecule and participates in various plant physiological processes, but its content must be maintained relatively stable. Salt stress, however, causes the accumulation of active oxygen in plants, leading to oxidative stress [44]. When the accumulation of ROS occurs in plants, they initiate the antioxidant system to eliminate the accumulated ROS, thus maintaining the steady state of ROS levels under salt stress [45,46]. In our data, we identified 11 ROS-scavenging enzyme genes whose expression levels changed after salt stress, including one catalase (CAT), one glutathione S-transferase (GST), two superoxide dismutase (SOD), and seven peroxidase (POD) genes (Figure 4E). In comparison to the 0h time point, the expression levels of these 11 ROS scavenging enzyme genes were induced by salt stress in both genotypes and increased gradually with the prolongation of salt stress, indicating that these genes were positively regulated by salt stress (Figure 4E).
Expression Patterns of Genes Related to Ethylene Biosynthesis and Signaling during Salt Stress in Both Genotypes
It has been shown that phytohormones not only regulate plant growth and development under normal conditions, but also mediate various environmental stresses, including salt stress, to help plants adapt to challenging conditions [47]. To investigate the genes related to hormone biosynthesis and signaling that are regulated during salt stress, we compared the expression levels of hormone biosynthesis and signaling genes between cv Jin-mian 25 and Su-mian 3 plants.
Previous studies proved that ethylene is biosynthetic from its precursor, S-adenosylMet (SAM), through two steps [48,49]. The first step involves the conversion of SAM to 1-aminocyclopropane-1-carboxylic acid (ACC) by aminocyclopropane-1-carboxylate synthase (ACS), while the second step is the conversion of ACC to ethylene by ACC oxidase (ACO) [50,51] (Figure 5A). Two ACS gene transcripts were detected in our data set, showing an induced expression pattern at 72h time point after salt stress treatment in both genotypes with higher transcripts in cv Su-mian 3 than in cv Jin-mian 25 (Figure 5B). In contrast, eleven ACO transcripts (four ACO1, two ACO3, two ACO4 and three ACO-like genes) were detected. In both genotypes, most ACO genes showed low to high expression levels, especially showing an induced expression at 72h time point after salt stress treatment, while one ACO1 was firstly high expressed at 6 and 12h but very low levels at 72h time point (Figure 5C). The above data showed that the important enzymes ACS and ACO, involved in ethylene synthesis, might be crucial in the cotton plant’s response to salt stress.
The ethylene signal transduction process in the model plant Arabidopsis has been studied quite clearly [49]. The components of the ethylene signal pathway include ethylene receptors (ETR1, ETR2, ERS1, ERS2 and EIN4) located on the endoplasmic reticulum, signal pathway inhibitors CTR1, EIN2, transcription factor EIN3/EIL1, downstream transcription factors and functional genes [52] (Figure 5A). 14 ethylene receptors gene transcripts were detected. Among them, four ERS1, three ETR2 and one EIN4 membrane receptor genes showed the induced expression pattern only at 72h time point after salt stress treatment in both genotypes (Figure 5D). Two ETR1 genes presented high transcripts at 6, 12 and 24h but low levels at 72h time point (Figure 5D). In addition, the ethylene signal pathway inhibitor CTR1 genes showed diverged expression profiles after salt stress, among which five genes were induced, while the other eight genes were repressed only at 72h time point (Figure 5E). In contrast, most EIN2 and EIN3 gene transcripts was induced at 6, 12 or 24h time point but reduced after 72h salt stress treatment in both genotypes (Figure 5F, G). Generally, the expression patterns of genes involved in ethylene biosynthesis and signaling were mostly similar in both cv Su-mian 3 and Jin-mian 25 plants (Figure 5), which indicated that ethylene signaling pathway was involved in the salt stress response in both genotypes.
Expression Patterns of Genes Related to Gibberellin (GA) Biosynthesis and Signaling during Salt Stress in Both Genotypes
GA was also involved in the salt stress response in plants [53]. The GA biosynthetic mutant ga1-3 showed higher tolerance to high-salinity stress [54] and salinity stress would reduce endogenous GA levels via upregulating GA 2-oxidase gene [55]. The biosynthesis of gibberellins (GA) begins with the transformation of trans-geranyl-geranyl diphosphate via four enzymatic steps to produce GA12 [56] (Figure 6A). GA12 is then converted to GA53 by the enzyme GA13 oxidase (GA13ox), and both GA12 and GA53 can be transformed into GA9 and GA20, respectively, by following two different pathways catalyzed by GA20ox [57]. Finally, GA9 and GA20 are converted into the bioactive GA forms GA4 and GA1 by the enzyme GA3ox [58] (Figure 6A). In our data, six GA biosynthetic metabolic enzymes including ent-copalyl diphosphate synthase (CPS), ent-kaurene synthase (KS), ent-kaurene oxidase (KO), ent-kaurenoic acid oxidase (KAO), Gibberellin 20 oxidase (GA20ox), and GA 2-oxidase (GA2ox) were detected (Figure 6B-G).
Six CPS and five KAO enzyme genes were identified and both two were repressed at 72 h after salt stress in both genotypes (Figure 6B, E). Among the KS and KO enzyme genes, their transcripts showed a slight increase at 72 h after salt stress in both genotypes, but both expressed at low levels (less than 2 fpkm for KS or 12 fpkm for KO) during salt stress (Figure 6C, D). The transcripts of four GA20ox genes were detected in our data set, and one GA20ox1 gene was repressed at 72 h after salt stress, while two GA20ox2 genes was induced markedly during salt stress in both genotypes (Figure 6F). The bioactive form GA was inactivated by GA2ox enzymes [59]. The transcripts of four GA2ox genes was detected, and the expression patterns were slightly diverged between the two genotypes. Among the four putative GA2ox genes detected, two genes (GA2ox2 and GA2ox8) were repressed at all time course while another two (GA2ox1) were induced at 6h but repressed at other time course during salt stress in cv Jin-mian 25 (Figure 6G). In contrast, all four GA2ox gene transcripts were repressed during salt stress in cv Su-mian 3 (Figure 6G). The transcripts of GA13ox and GA3ox enzymes genes were not detected in our data set.
We further investigated the expression profiles of GA signaling-related genes during salt stress and found that GA receptor GID1 and the GA signal pathway inhibitor DELLA genes was expressed after salt stress in both genotypes. Seven GA receptor gene transcripts were detected. Among them, one GID1A and two GID1C receptor genes showed the induced expression pattern at 72h time point after salt stress treatment in both genotypes (Figure 6H). Two GID1B and another two GID1C genes presented higher transcripts at 6, 12 or 24h but lower levels at 72h compared with 0h (Figure 6H). In addition, six GA signal pathway inhibitor DELLA genes, including two GAI, two SLR1 and two RGA, showed the induced expression patterns at 24 or 72h time points after salt stress in both genotypes (Figure 6I). Generally, the expression patterns of genes involved in GA biosynthesis and signaling were mostly similar in both cv Su-mian 3 and Jin-mian 25 plants (Figure 6), which indicated both genotypes required reduced bioactive GA levels or signaling for plant salt stress tolerance.
Expression Patterns of Genes Related to Abscisic Acid (ABA) Biosynthesis and Signaling during Salt Stress in Both Genotypes
ABA is a growth inhibitor with imposition to salt stress [60]. In higher plants, β-carotene, the precursor of ABA, is synthesized via the plastidial methylerythritol phosphate (MEP) pathway from isopentenyl pyrophosphate [61,62]. Then, β-carotene is catalyzed by six enzymes and eventually converted into ABA through several steps (Figure 7A). In our data, seven ABA biosynthetic metabolic enzymes including β-carotene hydroxylase (CHY-β), zeaxanthin epoxidase (ZEP), neoxanthin synthase (NSY/ABA4), 9-cis-epoxycarotenoid dioxygenase (NCED), xanthoxin dehydrogenase (ABA2), molybdenum cofactor sulfurase (ABA3) and abscisic-aldehyde oxidase (AAO3) were detected (Figure 7A).
Two CHY-β enzyme genes were identified and both were repressed at 24 or 72 h after salt stress in both genotypes (Figure 7B). Among the four ZEP and two ABA4 genes, their transcripts were induced markedly at 72 h after salt stress in both genotypes (Figure 7B, C). NCED is a rate-limiting enzyme in ABA biosynthesis [63], and five NCED genes were detected in our data set, but all expressed at low levels (less than 4 fpkm) during salt stress (Figure 7D). The transcripts of NCED3 and NCED5 showed a slight increase at 72 h after salt stress in cv Su-mian 3, while two NCED4 genes showed the upregulated transcripts at 72 h after salt stress in cv Jin-mian 25 (Figure 7D). The transcripts of three ABA2 genes were induced markedly during salt stress in both genotypes (Figure 7E). Among the three ABA3 and three AAO3 genes, only one ABA3 expression was induced at 72 h, but the other five gene transcripts were repressed at 72 h after salt stress in both genotypes (Figure 7F). ABA is inactivated by the action of ABA 8’-hydroxylase (ABA8Ox), which is one of the key steps in modulating ABA level [64]. The transcripts of ABA8ox1 and ABA8ox4 genes, were substantially induced at 72h of salt stress treatment (Figure 7G). Additionally, the expression of two ABA8ox2 genes was reduced at 72h of salt stress treatment in both genotypes (Figure 7G).
The expression profiles of ABA signaling pathway genes during salt stress were also investigated in both genotypes. 16 ABA receptor gene transcripts were detected. Among them, two PLY1 receptor genes showed a reduced expression pattern at 72h of salt stress treatment in both genotypes (Figure 7H). PLY4, PLY8, PLY9 and PLY11 receptor genes presented induced transcripts at 6, 12, 24 or 72h compared with 0h (Figure 7H). Overall, the transcripts of the ABA receptor genes (PLY family) were induced by salt stress treatment.
In addition, key players in ABA signaling transduction including PP2C, SnRK2, ABI3, BAM1, and ABFs genes were also detected in our data. The transcripts of nine PP2C genes were suppressed at 12, 24 or 72h after salt stress in both genotypes (Figure 7I). However, eight SnRK2 genes showed up-regulated transcripts at 12, 24 or 72h after salt stress in both genotypes (Figure 7J). We also found that ABI3 gene expression was repressed at 72h of salt stress treatment in both genotypes (Figure 7K). In addition, four BAM1 genes showed reduced expression patterns after salt stress, on the contrary, two ABF2 and two ABF4 showed induced expression patterns at 72h time points after salt stress in both genotypes (Figure 7K). Generally, the genes involved in ABA biosynthesis and signaling were expressed similarly in both cv Su-mian 3 and Jin-mian 25 plants (Figure 7). Our data indicated that enhanced ABA-activated SnRKs signaling module might be involved in the salt stress regulation in cotton plants. Meanwhile, ABA exhibited complicated signaling crosstalk to control cotton plant resistance against salt stress.
Expression Patterns of Genes Related to Jasmonic Acid (JA) Biosynthesis and Signaling during Salt Stress in Both Genotypes
JA controls the responses to salt stress tolerance in plants [65,66]. Jasmonates (JAs) originate from α-linolenic acid (Figure 8A), which is produced from chloroplast membrane galactolipids catalyzed by fatty acid desaturase (FAD) and phospholipase A1 (PLA1) or defective anther dehiscence 1 (DAD1) enzymes [67]. Then, several enzymes, including lipoxygenase (LOX), allene oxide synthase (AOS), allene oxide cyclase (AOC), 12-oxophytodienoic acid reductase 3 (OPR3), and OPC-8:0 CoA ligase 1 (OPCL1), convert α-linolenic acid to OPC-8-CoA [68,69], which subsequently undergoes three rounds of β-oxidation to yield jasmonoyl-CoA and then cleaved to (+)-7-iso-jasmonoyl by thioesterase (Figure 8A). The three key enzymes in β-oxidation are acyl-CoA oxidase (ACX), a multifunctional protein (MFP), and 3-keto-acyl-CoA thiolase (KAT). Finally, (+)-7-iso-jasmonoyl-Ile is produced by the catalyzation of the jasmonate resistant1 (JAR1) enzyme (Figure 8A). In our data, all 12 JA biosynthetic metabolic enzymes were detected (Figure 8A).
Eight FAD enzyme genes were identified, among them, the expression levels of four FAD3 and two FAD2 were markedly induced at 72 h while two FAD12 were repressed at 72 h after salt stress in both genotypes (Figure 8B). Among the four PLA1 and three DAD1 genes, most of their expression levels were induced markedly at 72 h after salt stress in both genotypes (Figure 8C). Five LOX genes were detected in our data set, and the expression levels of four LOX genes showed a significantly increase at 72 h after salt stress in both genotypes, while one LOX5 gene showed the down-regulated expression at 72 h after salt stress (Figure 8D). The transcripts of AOS (Figure 8E), AOC (Figure 8F), OPR3 (Figure 8G) and OPCL1 (Figure 8H) genes all were induced markedly during salt stress in both genotypes. Among the 17 ACX genes, 12 of their expression levels (four ACX1, two ACX2, four ACX3, and two ACX4) were induced during salt stress, but the other five ACX1 and ACX2 gene expression were repressed at 72 h after salt stress in both genotypes (Figure 9A). The transcripts of MFP (Figure 9B), KAT (Figure 9C), thioesterase (Figure 9D) and JAR1 (Figure 9E) genes all were induced markedly during salt stress in both genotypes. In summary, the expression levels of the JA biosynthetic metabolic enzyme genes were induced by salt stress treatment in both genotypes. JA-Ile is inactivated by the activity of the members of the CYP94 family through hydroxylation (Figure 8A). Two CYP94B3 genes detected were not expressed (less than 1 fpkm) during salt stress (Figure 9F). The expression levels of two CYP94C1 showed a slight increase at 72 h after salt stress in both genotypes, but both expressed at low levels (less than 7 fpkm) (Figure 9F).
The expression profiles of JA signaling pathway genes were also investigated during salt stress in both genotypes. COI1 and JAZ co-receptor gene transcripts were detected. Four COI1 receptor genes showed an induced expression pattern during salt stress treatment in both genotypes (Figure 9G), and 12 JAZ protein genes presented significantly induced expression profiles at 72h compared with 0h (Figure 9H). Overall, the expression levels of the JA co-receptor genes were induced by salt stress treatment in both genotypes. In addition, MYC2 genes, as the key transcription factor of JA signaling transduction, were also detected in our data. The expression levels of two MYC2 genes were higher at 72h after salt stress in both genotypes (Figure 9I). Generally, the genes involved in JA biosynthesis and signaling were expressed similarly in both cv Su-mian 3 and Jin-mian 25 plants (Figure 8 and 9), which indicated that JA biosynthesis and signaling pathways were positively involved in the salt stress response in both genotypes.
Expression Patterns of Genes Related to Brassinosteroids (BRs) Biosynthesis and Signaling during Salt Stress in Both Genotypes
BRs are a type of steroid phytohormones found in plants, and it has been widely reported that BRs can enhance salt stress tolerance in various plants [70,71]. The detailed study of the biosynthesis of brassinolide, which is a C28 BR, showed that there are two parallel pathways, the early and late C-6 oxidation pathways [72,73]. These pathways are interconnected at multiple steps and are also linked to the early C-22 oxidation pathway [74,75,76]. Therefore, the biosynthetic pathways of BR are highly connected. Additionally, information about the enzymes and genes involved in BR biosynthesis, as well as their regulation, have been identified, including CYP90A1 (CPD), CYP90B1 (DWF4), CYP90C1 (ROT3), CYP90D1, DET2, brassinosteroid-6-oxidase 1/2 (BR6OX1/2, also known as CYP85A1/2), membrane-localized receptors BRASSINOSTEROID-INSENSITIVE 1 (BRI1), and the coreceptor BAK1 (Figure 10A).
Three DWF4 enzyme genes were identified and their expression levels were markedly induced at 24 or 72 h after salt stress in both genotypes (Figure 10B). Among the four DET2 genes, most of their expression levels were induced markedly at 72 h after salt stress in both genotypes (Figure 10C). Four CPD genes were detected in our data set, and their expression levels showed a significantly increase at 24 or 72 h after salt stress in both genotypes (Figure 10D). Among the two ROT3 genes, one of the expression levels was induced, but the other one was repressed during salt stress in both genotypes (Figure 10E). The transcripts of CYP90D1 (Figure 10F) and CYP85A1 (Figure 10G) genes all were induced slightly during salt stress in both genotypes and both expressed at low levels (less than 12 fpkm). In summary, the expression levels of the BR biosynthetic metabolic enzyme genes were induced by salt stress treatment in both genotypes.
The expression profiles of BR signaling pathway genes were also investigated during salt stress in both genotypes. BRI1 and BAK1 co-receptor gene transcripts were detected. Three BRI1 receptor genes showed an induced expression pattern during salt stress treatment in both genotypes (Figure 10H), and only one BAK1 gene presented significantly induced expression profiles at 72h compared with 0h (Figure 10 H). Overall, the expression levels of the BR receptor genes were induced by salt stress treatment in both genotypes. In addition, BIN2 genes, as the critical negative component of BR signaling, were also detected in our data. The expression levels of five BIN2 genes were higher at 12, 24 or 72h after salt stress in both genotypes (Figure 10I). Generally, the genes involved in BR biosynthesis and signaling were expressed similarly in both cv Su-mian 3 and Jin-mian 25 plants (Figure 10), which indicated that BR biosynthesis and signaling pathways were positively involved in the salt stress response in both genotypes.
We conducted a systematic investigation into the expression patterns of genes related to five phytohormone biosynthesis and signaling pathways in both cv Su-mian 3 and Jin-mian 25 under salt stress. Despite the involvement of these hormones in cotton’s response to salt stress in both positive or negative ways, the expression levels of these genes in both genotypes are similar. Therefore, these genes are not the genetic basis of the diverged salt tolerance in both genotypes.
Expression Patterns of Genes Related to Trehalose Metabolism Pathway during Salt Stress in Both Genotypes
We delved further into the expression of genes in other relevant pathways to comprehend the metabolic adaptation of salt-tolerant cotton under salt stress [77,78]. As a disaccharide that functioned in carbohydrate transport and metabolism, trehalose alleviated salt stress in plants by regulating the trehalose metabolic pathway [79,80]. In plants, trehalose is produced from UDP-Glc and Glc-6-P by trehalose-6-phosphate synthases (TPS) and trehalose-6-phosphate phosphatases (TPP), and degraded by trehalase (TRE) (Figure 11A).
In our data set, the expression levels of 12 TPS, 12 TPP, and three TRE genes were detected. Notably, nine TPS genes (four TPS1, two TPS6, one TPS7, and two TPS11) were strongly induced at 6h time point after salt stress in cv Jin-mian 25 plants but not in cv Su-mian 3 plants (Figure 11B). Three TPS genes (two TPS5, and one TPS9) were markedly induced in response to salt stress in both genotypes (Figure 11B). Ten of 12 TPP genes showed elevated expression levels at 72h time point in response to salt stress with expression at relatively low levels in both genotypes (Figure 11C). The expression of three TRE1 genes also were up-regulated after salt stress, and the expression patterns were similar in both genotypes (Figure 11D).
Discussion
Salt stress is a significant environmental factor that impacts the growth and development of plants [81]. As plants are immobile, they must develop appropriate mechanisms to adapt to high-salt environments [82]. Salt stress significantly affects plant growth and development with significant inhibition of seed germination. In this study, cotton seeds were exposed to increasing time of salt stress and the growth of Su-mian 3 (salt-sensitive) was more severely inhibited than Jin-mian 25 (salt-tolerant). This study aimed to understand the transcriptional responses associated with the salt stress of the salt-tolerant cotton cv Jin-mian 25 and the salt-sensitive cotton cv Su-mian 3. Cataloging the entire transcriptome using an RNA-Seq approach revealed differences in gene expression between salt-tolerant cotton (cv Jin-mian 25) and salt-sensitive cotton (cv Su-mian 3) plants, as well as unique and common responses to salt stress in both genotypes. The results of our thorough analysis of the transcriptome contribute to the discovery of key genes that regulate metabolic pathways during salt stress in cotton.
Previous studies have demonstrated that the phytohormones including ethylene (ET), gibberellin (GA), abscisic acid (ABA), jasmonic acid (JA), and brassinosteroid (BR) modulate salt stress resistance in plants [47,53]. Ethylene signaling can influence the salt response at various levels, including membrane receptors, cytoplasmic components, and nuclear transcription factors within the pathway [83,84]. Three gain-of-function mutations of the ethylene receptors CpETR1B, CpETR1A, and CpETR2B in Cucurbita pepo all reduced ethylene sensitivity, but enhanced salt tolerance during seed germination and plant growth [85]. The enhanced salt tolerance response of the mutants was associated with the change of cytoplasmic components, presenting a reduced accumulation of Na+, but a higher accumulation of proline and total carbohydrates, and anthocyanin. In addition, the expression levels of many membrane transporter genes, including Na+/H+ and K+/H+ exchangers, K+ efflux antiporters, high-affinity K+ transporters, and K+ uptake transporters were significantly upregulated under salt stress in mutants in comparison with WT [85]. Expression data indicated that two tomato ERFs were more strongly induced in salt-tolerant genotypes than in salt-sensitive genotypes, and over-expressing SpERF.B7 in Arabidopsis exhibited enhanced salt tolerance, proving that ERF transcription factors were involved in the salt response in tomato [86].
Many reports indicated that ethylene and related components were involved in plant salt stress response by the clearance of ROS [87]. For example, ethylene can improve Arabidopsis thaliana tolerance to salinity by regulating RBOHF-mediated ROS and Na/K balance [88]. Ethylene can also stabilize EIN3/EIL1 to enhance the expression of SIEDs and PODs, thereby clearing the accumulation of ROS in plant cells [89]. Meanwhile, ethylene treatment can recover the germination rate of alfalfa seeds under salt stress by reducing the accumulation of MDA and H2O2 and increasing POD activity [90]. A recent study also showed that at 150-mM NaCl stress, 2-mM ethephon treatment significantly increased the germination rate and potential of cotton seeds by more than 70%. Subsequent evidence proved that exogenous ethylene improved the salt tolerance and germination rate of cotton seeds by boosting antioxidant enzyme activity, increasing proline content, and reducing membrane lipid peroxidation [91]. These findings suggest that ethylene can mediate the balance of ROS in plant cells to affect plant salt tolerance.
By contrast, the GA-deficient mutant exhibited remarkable tolerance to salt stress [92]. Consistently, salt stress-induced plant DELLA accumulation elevated the expression levels of genes encoding ROS-detoxification enzymes, to reduce ROS levels thus delaying cell death and promoting tolerance [93]. In essence, these results indicated that reduced bioactive GA levels are required for plant tolerance to salt stress. ABA and GA signaling pathways appear to be interacting in the regulation of seed germination and seedling development under salt stress [94]. OsNCED5, a rice NCED gene, was induced by exposure to salt stress. Overexpression of OsNCED5 increased ABA levels and enhanced salt stress tolerance, while nced5 mutants reduced ABA levels and decreased tolerance to salt stress. Therefore, OsNCED5 might regulate plant development and salt stress resistance by controlling ABA biosynthesis [63]. Thus, endogenous ABA levels increased under salinity stress in plants and enhanced ABA signaling activated SnRK2s [95]. SnRK2s played critical roles in ROS production in ABA signaling networks [96]. Indeed, two SnRK2 kinases, SnRK2.4 and SnRK2.10, were involved in the regulation of ROS homeostasis in response to salinity in A. thaliana [97]. Together, ABA and ROS exhibit close signaling crosstalk to regulate plant resistance to salt stress.
In Arabidopsis, JA impaired plant salt stress tolerance by repressing Catalase2 expression, which led to lower CAT activity, and higher ROS accumulation [66]. In wheat, exogenous JA was found to enhance the salt tolerance of wheat seedlings by reducing membrane lipid oxidation, up-regulating the expression levels of antioxidative defense system genes, and increasing the activities of SOD, POD, CAT and APX [65]. Our analysis of the transcript indicates that salt stress triggers the expression of JA biosynthesis-related genes. Application of JA exogenously significantly alleviates salt-induced damage by increasing antioxidative enzyme activities and maintaining Na+/K+ balance [98,99]. As a stress-related hormone, JA signaling has been proven to play an essential role in plant salt tolerance.
BRs have also been widely reported to enhance salt stress tolerance in various plants [100], such as tomato [101], rice [102], and mustard [103]. Exogenous BR application decreased Na+ accumulation and increased K+ content to relieve salt toxicity by regulating the expression levels of Na+(K+)/H + antiporter genes [104]. Under saline conditions, BR can greatly reduce the generation of ROS by increasing antioxidant capacity [104,105]. These results highlight the potential roles of BR mediated ROS homeostasis in plant salt resistance.
In cotton, significant progress has been made in understanding the genetic basis of salt tolerance in the last 10-15 years. Numerous salt-responsive genes have been identified by using genomic, transcriptomic methods or transgenic approaches for enhancement of salt stress tolerance [106,107]. RNA-seq experiments showed that cotton GRF genes exhibited decreased expression in leaves under NaCl treatment [107]. Aquaporins genes in response to the salt stresses also were identified by transcriptome analysis. GhPIP2;7-silenced plants exhibited decreased SOD and POD activity under 400 mM NaCl treatment. GhTIP2;1-overexpressed plants showed reduced H2O2 and malondialdehyde accumulation but higher proline content under salt stress [108]. A recent study used VIGS to silence GhSAMC gene in cotton, which resulted in increased sensitivity to salt stress, with more ROS accumulation in leaves than in the control, suggesting that GhSAMC regulates ROS accumulation to enhance cotton’s salt tolerance [109]. For other plants, GmGSTU23, a tau-like glutathione transferase family gene, mediated the scavenging of ROS by enhancing the activity of glutathione transferase, thus conferring enhanced tolerance to salt stress in Glycine max [46]. Moreover, trehalose also enhanced antioxidant activities and the expression of stress-responsive proteins and genes, providing salt tolerance in plants [110,111]. Furthermore, a G protein γ subunit involved in salt response mainly affected the phosphorylation of aquaporins to modulate the distribution of H2O2 in sorghum, millet, rice, and maize [39]. It is evident that exposure to salt stress can cause plant cells to accumulate large amounts of ROS, necessitating an increase in the presence of reducing enzymes and substances to eliminate them.
Conclusions
We used comparative transcriptomics to analyze the DEGs in the two cultivars under salt stress to elucidate the molecular mechanisms of the differences in salt tolerance between Su-mian 3 and Jin-mian 25. Through assessment of the germination potential and germination rate of seeds as well as the fresh weight, seedling length, root length, and hypocotyl length under salt stress, Su-mian 3 was identified as a salt-sensitive cultivar, while Jin-mian 25 was a salt-tolerant cultivar. Then, transcriptome profiles of Su-mian 3 and Jin-mian 25 cotton seeds exposed to salt stress were performed. KEGG pathway enrichment analysis showed that DEGs were mainly involved in plant hormone signal transduction metabolism. Jin-mian 25 had 10 unique DEGs induced by salt stress, which might confer the salt stress tolerance. Based on this finding, we investigated the expression patterns of genes related to aquaporins, kinases, and the scavenging of ROS during salt stress in both genotypes. Most aquaporins, kinases, and the scavenging of ROS genes showed an induced expression after salt stress. Moreover, expression patterns of genes related to phytohormone ethylene biosynthesis and signaling during salt stress in both genotypes were also investigated. Thereinto, ET, ABA, JA, and BR were positively involved in the salt stress response, however, reduced GA levels were required for salt stress tolerance in both genotypes. Our transcriptome analysis also showed that trehalose alleviated salt stress by regulating the trehalose metabolic pathway in both genotypes. These results indicated that complicated signaling crosstalk existed to control cotton plant resistance against salt stress. The candidate genes identified from those multiple metabolic pathways should be further examined to determine their roles in the responses of cotton plants to salt stress.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
HL: LL, YY, and ZWC conceived and designed this experiment. HL, LL, XHK, XWW, AJS and JW collected samples and performed the study. HL, LL, CEG, JJ, and ZWC participated in the acquisition and analysis of the data. HL, LL and ZWC wrote the manuscript. LL and YY participated in the discussion draft of the manuscript. ZWC revised the final manuscript. All authors read and approved the final manuscript.
Funding
The work reported in this publication was supported by the National Key Research and Development Program of China (No. 2022YFD1200300), the earmarked fund for XJARS-Cotton (XJARS-03), Tianshan Talent Innovation Science and Technology Team (2022TSYCTD0022), and Major science and technology projects of the third Division (KY2022ZD02). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgments
We are deeply indebted to Professor Lida Zhang for helpful suggestions and comments on bioinformatic analyses, and we thank Professor Yi Huang for valuable comments on previous versions of the manuscript. We are also grateful to two anonymous reviewers for their helpful suggestions and comments.
Abbreviations
DEGs, differentially expressed genes; FPKM, fragments per kilobase of transcript per million mapped fragments; G. hirsutum, Gossypium hirsutum; A. thaliana, Arabidopsis thaliana; GO: Gene Ontology; KEGG: Kyoto Encyclopedia of Genes and Genomes; ROS, reactive oxygen species; ET, ethylene; GA, gibberellin; ABA, abscisic acid; JA, jasmonic acid; BR, brassinosteroid; LRR-RLK, leucine-rich repeat receptor-like protein kinase; cv, cultivar; POD, peroxidase; SOD, superoxide dismutase; CAT, catalase
References
- Wen, X.; Chen, Z.; Yang, Z.; Wang, M.; Jin, S.; Wang, G.; Zhang, L.; Wang, L.; Li, J.; Saeed, S.; He, S.; Wang, Z.; Wang, K.; Kong, Z.; Li, F.; Zhang, X.; Chen, X.; Zhu, Y. A comprehensive overview of cotton genomics, biotechnology and molecular biological studies. Sci China Life Sci 2023, 66((10)), 2214–2256. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, D.; Hua, J.; Kong, X.; Wang, X.; Wang, J.; Si, A.; Zhao, F.; Liu, W.; Yu, Y.; Chen, Z. Genetic and morpho-physiological differences among transgenic and no-transgenic cotton cultivars. Plants 2023, 12((19)), 3437. [Google Scholar] [CrossRef]
- Huang, W.; Wu, F.; Han, W.; Li, Q.; Han, Y.; Wang, G.; Feng, L.; Li, X.; Yang, B.; Lei, Y.; Fan, Z.; Xiong, S.; Xin, M.; Li, Y.; Wang, Z. Carbon footprint of cotton production in China: Composition, spatiotemporal changes and driving factors. Science of The Total Environment 2022, 821, 153407. [Google Scholar] [CrossRef]
- Hassani, A.; Azapagic, A.; Shokri, N. Global predictions of primary soil salinization under changing climate in the 21st century. Nature Communications 2021, 12((1)), 6663. [Google Scholar] [CrossRef]
- Li, D.; Yang, Y.; Zhao, Y.; Zhou, X.; Han, Q.; Liu, H.; Li, M. Optimizing cotton yield and soil salinity management: Integrating brackish water leaching and freshwater drip irrigation with subsurface drainage. Field Crops Research 2024, 314, 109454. [Google Scholar] [CrossRef]
- Sun, H.; Meng, M.; Yan, Z.; Lin, Z.; Nie, X.; Yang, X. Genome-wide association mapping of stress-tolerance traits in cotton. The Crop Journal 2019, 7((1)), 77–88. [Google Scholar] [CrossRef]
- Ashraf, M.; Ahmad, S. Influence of sodium chloride on ion accumulation, yield components and fibre characteristics in salt-tolerant and salt-sensitive lines of cotton (Gossypium hirsutum L.). Field Crops Research 2000, 66, 115–127. [Google Scholar] [CrossRef]
- Ibrahim, W.; Qiu, C. W.; Zhang, C.; Cao, F.; Shuijin, Z.; Wu, F. Comparative physiological analysis in the tolerance to salinity and drought individual and combination in two cotton genotypes with contrasting salt tolerance. Physiol Plant 2019, 165((2)), 155–168. [Google Scholar] [CrossRef]
- Ibrahim, W.; Zhu, Y. M.; Chen, Y.; Qiu, C. W.; Zhu, S.; Wu, F. Genotypic differences in leaf secondary metabolism, plant hormones and yield under alone and combined stress of drought and salinity in cotton genotypes. Physiol Plant 2019, 165((2)), 343–355. [Google Scholar] [CrossRef]
- Wang, N.; Qi, H.; Qiao, W.; Shi, J.; Xu, Q.; Zhou, H.-W.; Yan, G.; Huang, Q. Cotton (Gossypium hirsutum L.) genotypes with contrasting K+/Na+ ion homeostasis: implications for salinity tolerance. Acta Physiologiae Plantarum 2017, 39, 1-10.
- Zhang, L.; Zhang, G.; Wang, Y.; Zhou, Z.; Meng, Y.; Chen, B. Effect of soil salinity on physiological characteristics of functional leaves of cotton plants. J Plant Res 2013, 126((2)), 293–304. [Google Scholar] [CrossRef]
- Wang, M.; Tu, L.; Yuan, D.; Zhu, D.; Shen, C.; Li, J.; Liu, F.; Pei, L.; Wang, P.; Zhao, G.; Ye, Z.; Huang, H.; Yan, F.; Ma, Y.; Zhang, L.; Liu, M.; You, J.; Yang, Y.; Liu, Z.; Huang, F.; Li, B.; Qiu, P.; Zhang, Q.; Zhu, L.; Jin, S.; Yang, X.; Min, L.; Li, G.; Chen, L.-L.; Zheng, H.; Lindsey, K.; Lin, Z.; Udall, J. A.; Zhang, X. Reference genome sequences of two cultivated allotetraploid cottons, Gossypium hirsutum and Gossypium barbadense. Nature Genetics 2019, 51((2)), 224–229. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, J.; Qi, Z.; Long, Y.; Pei, L.; Huang, X.; Grover, C. E.; Du, X.; Xia, C.; Wang, P.; Liu, Z.; You, J.; Tian, X.; Ma, Y.; Wang, R.; Chen, X.; He, X.; Fang, D. D.; Sun, Y.; Tu, L.; Jin, S.; Zhu, L.; Wendel, J. F.; Zhang, X. Genomic innovation and regulatory rewiring during evolution of the cotton genus Gossypium. Nature Genetics 2022, 54((12)), 1959–1971. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Fang, L.; Zhang, Z.; Ma, W.; Niu, Y.; Ju, L.; Deng, J.; Zhao, T.; Lian, J.; Baruch, K.; Fang, D.; Liu, X.; Ruan, Y.-l.; Rahman, M.-u.; Han, J.; Wang, K.; Wang, Q.; Wu, H.; Mei, G.; Zang, Y.; Han, Z.; Xu, C.; Shen, W.; Yang, D.; Si, Z.; Dai, F.; Zou, L.; Huang, F.; Bai, Y.; Zhang, Y.; Brodt, A.; Ben-Hamo, H.; Zhu, X.; Zhou, B.; Guan, X.; Zhu, S.; Chen, X.; Zhang, T. Gossypium barbadense and Gossypium hirsutum genomes provide insights into the origin and evolution of allotetraploid cotton. Nature Genetics 2019, 51((4)), 739–748. [Google Scholar] [CrossRef]
- Huang, G.; Wu, Z.; Percy, R. G.; Bai, M.; Li, Y.; Frelichowski, J. E.; Hu, J.; Wang, K.; Yu, J. Z.; Zhu, Y. Genome sequence of Gossypium herbaceum and genome updates of Gossypium arboreum and Gossypium hirsutum provide insights into cotton A-genome evolution. Nature Genetics 2020, 52((5)), 516–524. [Google Scholar] [CrossRef]
- Chen, Z. J.; Sreedasyam, A.; Ando, A.; Song, Q.; De Santiago, L. M.; Hulse-Kemp, A. M.; Ding, M.; Ye, W.; Kirkbride, R. C.; Jenkins, J.; Plott, C.; Lovell, J.; Lin, Y.-M.; Vaughn, R.; Liu, B.; Simpson, S.; Scheffler, B. E.; Wen, L.; Saski, C. A.; Grover, C. E.; Hu, G.; Conover, J. L.; Carlson, J. W.; Shu, S.; Boston, L. B.; Williams, M.; Peterson, D. G.; McGee, K.; Jones, D. C.; Wendel, J. F.; Stelly, D. M.; Grimwood, J.; Schmutz, J. Genomic diversifications of five Gossypium allopolyploid species and their impact on cotton improvement. Nature Genetics 2020, 52((5)), 525–533. [Google Scholar] [CrossRef]
- Peng, Z.; He, S.; Gong, W.; Sun, J.; Pan, Z.; Xu, F.; Lu, Y.; Du, X. Comprehensive analysis of differentially expressed genes and transcriptional regulation induced by salt stress in two contrasting cotton genotypes. BMC Genomics 2014, 15((1)), 760. [Google Scholar] [CrossRef]
- Yuan, Z.; Zhang, C.; Zhu, W.; Yan, G.; Chen, X.; Qiu, P.; Ruzimurod, B.; Ye, W.; Qaraevna, B. Z.; Yin, Z. Molecular mechanism that underlies cotton response to salt and drought stress revealed by complementary transcriptomic and iTRAQ analyses. Environmental and Experimental Botany 2023, 209, 105288. [Google Scholar] [CrossRef]
- Zhang, F.; Zhu, G.; Du, L.; Shang, X.; Cheng, C.; Yang, B.; Hu, Y.; Cai, C.; Guo, W. Genetic regulation of salt stress tolerance revealed by RNA-Seq in cotton diploid wild species, Gossypium davidsonii. Sci Rep 2016, 6, 20582. [Google Scholar] [CrossRef]
- Guo, J.; Shi, G.; Guo, X.; Zhang, L.; Xu, W.; Wang, Y.; Su, Z.; Hua, J. Transcriptome analysis reveals that distinct metabolic pathways operate in salt-tolerant and salt-sensitive upland cotton varieties subjected to salinity stress. Plant Sci 2015, 238, 33–45. [Google Scholar] [CrossRef]
- Ju, F.; Pang, J.; Sun, L.; Gu, J.; Wang, Z.; Wu, X.; Ali, S.; Wang, Y.; Zhao, W.; Wang, S.; Zhou, Z.; Chen, B. Integrative transcriptomic, metabolomic and physiological analyses revealed the physiological and molecular mechanisms by which potassium regulates the salt tolerance of cotton (Gossypium hirsutum L.) roots. Industrial Crops and Products 2023, 193, 116177.
- Li, P.; Liu, Q.; Wei, Y.; Xing, C.; Xu, Z.; Ding, F.; Liu, Y.; Lu, Q.; Hu, N.; Wang, T.; Zhu, X.; Cheng, S.; Li, Z.; Zhao, Z.; Li, Y.; Han, J.; Cai, X.; Zhou, Z.; Wang, K.; Zhang, B.; Liu, F.; Jin, S.; Peng, R. Transcriptional landscape of cotton roots in response to salt stress at single-cell resolution. Plant Communications 2024, 5((2)), 100740. [Google Scholar] [CrossRef]
- Hao, Y.; Lu, G.-q.; Wang, L.-h.; Wang, C.-l.; Guo, H.; Li, Y.-F.; Cheng, H. Overexpression of AmDUF1517 enhanced tolerance to salinity, drought, and cold stress in transgenic cotton. Journal of Integrative Agriculture 2018. [Google Scholar] [CrossRef]
- Liu, G.; Li, X.; Jin, S.; Liu, X.; Zhu, L.; Nie, Y.; Zhang, X. Overexpression of rice NAC gene SNAC1 improves drought and salt tolerance by enhancing root development and reducing transpiration rate in transgenic cotton. PLoS One 2014, 9((1)), e86895. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wan, S.; Liu, H.; Fan, S.; Zhang, Y.; Wang, W.; Xia, M.; Yuan, R.; Deng, F.; Shen, F. Overexpression of an Apocynum venetum DEAD-box helicase gene (AvDH1) in cotton confers salinity tolerance and increases yield in a saline field. Front Plant Sci 2015, 6, 1227. [Google Scholar] [CrossRef]
- Su, Y.; Liang, W.; Liu, Z.; Wang, Y.; Zhao, Y.; Ijaz, B.; Hua, J.-p. Overexpression of GhDof1 improved salt and cold tolerance and seed oil content in Gossypium hirsutum. Journal of plant physiology 2017, 218, 222–234. [Google Scholar] [CrossRef]
- Song, J.; Zhang, R.; Yue, D.; Chen, X.; Guo, Z.; Cheng, C.; Hu, M.; Zhang, J.; Zhang, K. Co-expression of ApGSMT2g and ApDMT2g in cotton enhances salt tolerance and increases seed cotton yield in saline fields. Plant Science 2018, 274, 369–382. [Google Scholar] [CrossRef]
- Liang, C.; Meng, Z.; Meng, Z.; Malik, W.; Yan, R.; Lwin, K. M.; Lin, F.; Wang, Y.; Sun, G.; Zhou, T.; Zhu, T.; Li, J.; Jin, S.; Guo, S.; Zhang, R. GhABF2, a bZIP transcription factor, confers drought and salinity tolerance in cotton (Gossypium hirsutum L.). Sci Rep 2016, 6, 35040.
- Zhang, H.; Mao, L.; Xin, M.; Xing, H.; Zhang, Y.; Wu, J.; Xu, D.; Wang, Y.; Shang, Y.; Wei, L.; Cui, M.; Zhuang, T.; Sun, X.; Song, X. Overexpression of GhABF3 increases cotton(Gossypium hirsutum L.) tolerance to salt and drought. BMC Plant Biology 2022, 22, (1), 313.
- Kim, D.; Langmead, B.; Salzberg, S. L. HISAT: a fast spliced aligner with low memory requirements. Nat Methods 2015, 12((4)), 357–60. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G. M.; Leek, J. T.; Salzberg, S. L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat Protoc 2016, 11((9)), 1650–67. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, J.; Qiao, J.; Li, W.; Guan, Z.; Liu, Z.; Bai, X.; Xing, B.; Zhang, J.; Li, J.; Yin, W.; Zhu, H. Graphene-mediated antioxidant enzyme activity and respiration in plant roots. ACS Agricultural Science & Technology 2022, 2, (3), 646-660.
- Liu, L.; Grover, C. E.; Kong, X.; Jareczek, J.; Wang, X.; Si, A.; Wang, J.; Yu, Y.; Chen, Z. Expression profile analysis of cotton fiber secondary cell wall thickening stage. PeerJ 2024, 12, e17682. [Google Scholar] [CrossRef]
- Love, M. I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 2014, 15((12)), 550. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J. G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21((19)), 3787–3793. [Google Scholar] [CrossRef]
- Young, M. D.; Wakefield, M. J.; Smyth, G. K.; Oshlack, A. Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biol 2010, 11((2)), R14. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H. R.; Frank, M. H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol Plant 2020, 13((8)), 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E. A. Seed priming to alleviate salinity stress in germinating seeds. Journal of Plant Physiology 2016, 192, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yu, F.; Xie, P.; Sun, S.; Qiao, X.; Tang, S.; Chen, C.; Yang, S.; Mei, C.; Yang, D.; Wu, Y.; Xia, R.; Li, X.; Lu, J.; Liu, Y.; Xie, X.; Ma, D.; Xu, X.; Liang, Z.; Feng, Z.; Huang, X.; Yu, H.; Liu, G.; Wang, Y.; Li, J.; Zhang, Q.; Chen, C.; Ouyang, Y.; Xie, Q. A Gγ protein regulates alkaline sensitivity in crops. Science 2023, 379((6638)), eade8416. [Google Scholar] [CrossRef]
- Ubersax, J. A.; Ferrell Jr, J. E. Mechanisms of specificity in protein phosphorylation. Nature Reviews Molecular Cell Biology 2007, 8((7)), 530–541. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Q. Q.; Zhou, L.; Ren, F.; Li, D. D.; Li, X. B. Arabidopsis CBL-interacting protein kinase (CIPK6) is involved in plant response to salt/osmotic stress and ABA. Mol Biol Rep 2013, 40((8)), 4759–67. [Google Scholar] [CrossRef]
- Su, Y.; Guo, A.; Huang, Y.; Wang, Y.; Hua, J. GhCIPK6a increases salt tolerance in transgenic upland cotton by involving in ROS scavenging and MAPK signaling pathways. BMC Plant Biology 2020, 20((1)), 421. [Google Scholar] [CrossRef]
- Tripathi, V.; Parasuraman, B.; Laxmi, A.; Chattopadhyay, D. CIPK6, a CBL-interacting protein kinase is required for development and salt tolerance in plants. The Plant Journal 2009, 58((5)), 778–790. [Google Scholar] [CrossRef]
- Castro, B.; Citterico, M.; Kimura, S.; Stevens, D. M.; Wrzaczek, M.; Coaker, G. Stress-induced reactive oxygen species compartmentalization, perception and signalling. Nature Plants 2021, 7((4)), 403–412. [Google Scholar] [CrossRef]
- Kesawat, M. S.; Satheesh, N.; Kherawat, B. S.; Kumar, A.; Kim, H. U.; Chung, S. M.; Kumar, M. Regulation of reactive oxygen species during salt stress in plants and their crosstalk with other signaling molecules-current perspectives and future directions. Plants (Basel) 2023, 12, (4).
- Li, X.; Pang, Y.; Zhong, Y.; Cai, Z.; Ma, Q.; Wen, K.; Nian, H. GmGSTU23 encoding a Tau class glutathione s-transferase protein enhances the salt tolerance of Soybean (Glycine max L.). Int J Mol Sci 2023, 24, (6), 5547.
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How plant hormones mediate salt stress responses. Trends Plant Sci 2020, 25((11)), 1117–1130. [Google Scholar] [CrossRef]
- S F Yang, a.; Hoffman, N. E. Ethylene biosynthesis and its regulation in higher plants. Annual Review of Plant Physiology 1984, 35((1)), 155–189. [Google Scholar] [CrossRef]
- Wang, K. L.; Li, H.; Ecker, J. R. Ethylene biosynthesis and signaling networks. Plant Cell 2002, 14 Suppl (Suppl), S131–51. [Google Scholar] [CrossRef] [PubMed]
- Ruduś, I.; Sasiak, M.; Kępczyński, J. Regulation of ethylene biosynthesis at the level of 1-aminocyclopropane-1-carboxylate oxidase (ACO) gene. Acta Physiologiae Plantarum 2013, 35((2)), 295–307. [Google Scholar] [CrossRef]
- Pattyn, J.; Vaughan-Hirsch, J.; Van de Poel, B. The regulation of ethylene biosynthesis: a complex multilevel control circuitry. New Phytol 2021, 229((2)), 770–782. [Google Scholar] [CrossRef]
- Guo, H.; Ecker, J. R. The ethylene signaling pathway: new insights. Curr Opin Plant Biol 2004, 7((1)), 40–9. [Google Scholar] [CrossRef]
- Xiao, F.; Zhou, H. Plant salt response: Perception, signaling, and tolerance. Front Plant Sci 2022, 13, 1053699. [Google Scholar] [CrossRef]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. dwarf and delayed-flowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor. The Plant Journal 2004, 37((5)), 720–729. [Google Scholar] [CrossRef]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. The DDF1 transcriptional activator upregulates expression of a gibberellin-deactivating gene, GA2ox7, under high-salinity stress in Arabidopsis. Plant J 2008, 56((4)), 613–26. [Google Scholar] [CrossRef]
- Richards, D. E.; King, K. E.; Ait-Ali, T.; Harberd, N. P. How gibberellin regulates plant growth and development: a molecular genetic analysis of gibberellin signaling. Annu Rev Plant Physiol Plant Mol Biol 2001, 52, 67–88. [Google Scholar] [CrossRef]
- Magome, H.; Nomura, T.; Hanada, A.; Takeda-Kamiya, N.; Ohnishi, T.; Shinma, Y.; Katsumata, T.; Kawaide, H.; Kamiya, Y.; Yamaguchi, S. CYP714B1 and CYP714B2 encode gibberellin 13-oxidases that reduce gibberellin activity in rice. Proc Natl Acad Sci U S A 2013, 110((5)), 1947–52. [Google Scholar] [CrossRef]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu Rev Plant Biol 2008, 59, 225–51. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Miura, K.; Itoh, H.; Tatsumi, T.; Ueguchi-Tanaka, M.; Ishiyama, K.; Kobayashi, M.; Agrawal, G. K.; Takeda, S.; Abe, K.; Miyao, A.; Hirochika, H.; Kitano, H.; Ashikari, M.; Matsuoka, M. An overview of gibberellin metabolism enzyme genes and their related mutants in rice. Plant Physiol 2004, 134((4)), 1642–53. [Google Scholar] [CrossRef] [PubMed]
- Waśkiewicz, A.; Beszterda, M.; Goliński, P. ABA: role in plant signaling under salt stress. In Salt Stress in Plants: Signalling, Omics and Adaptations, Ahmad, P.; Azooz, M. M.; Prasad, M. N. V., Eds. Springer New York: New York, NY, 2013; pp 175-196.
- Jia, K.-P.; Mi, J.; Ali, S.; Ohyanagi, H.; Moreno, J. C.; Ablazov, A.; Balakrishna, A.; Berqdar, L.; Fiore, A.; Diretto, G.; Martínez, C.; de Lera, A. R.; Gojobori, T.; Al-Babili, S. An alternative, zeaxanthin epoxidase-independent abscisic acid biosynthetic pathway in plants. Molecular Plant 2022, 15((1)), 151–166. [Google Scholar] [CrossRef] [PubMed]
- Endo, A.; Okamoto, M.; Koshiba, T. ABA Biosynthetic and Catabolic Pathways. In Abscisic Acid: Metabolism, Transport and Signaling, Zhang, D.-P., Ed. Springer Netherlands: Dordrecht, 2014; pp 21-45.
- Huang, Y.; Jiao, Y.; Xie, N.; Guo, Y.; Zhang, F.; Xiang, Z.; Wang, R.; Wang, F.; Gao, Q.; Tian, L.; Li, D.; Chen, L.; Liang, M. OsNCED5, a 9-cis-epoxycarotenoid dioxygenase gene, regulates salt and water stress tolerance and leaf senescence in rice. Plant Sci 2019, 287, 110188. [Google Scholar] [CrossRef]
- Shiono, K.; Yoshikawa, M.; Kreszies, T.; Yamada, S.; Hojo, Y.; Matsuura, T.; Mori, I. C.; Schreiber, L.; Yoshioka, T. Abscisic acid is required for exodermal suberization to form a barrier to radial oxygen loss in the adventitious roots of rice (Oryza sativa). New Phytologist 2022, 233((2)), 655–669. [Google Scholar] [CrossRef]
- Zhu, M.; Liu, Y.; Cai, P.; Duan, X.; Sang, S.; Qiu, Z. Jasmonic acid pretreatment improves salt tolerance of wheat by regulating hormones biosynthesis and antioxidant capacity. Front Plant Sci 2022, 13, 968477. [Google Scholar] [CrossRef]
- Song, R. F.; Li, T. T.; Liu, W. C. Jasmonic acid impairs Arabidopsis seedling salt stress tolerance through MYC2-mediated repression of CAT2 expression. Front Plant Sci 2021, 12, 730228. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann Bot 2013, 111((6)), 1021–58. [Google Scholar] [CrossRef]
- Wasternack, C.; Song, S. Jasmonates: biosynthesis, metabolism, and signaling by proteins activating and repressing transcription. J Exp Bot 2017, 68((6)), 1303–1321. [Google Scholar] [CrossRef]
- Koo, A. J. K.; Chung, H. S.; Kobayashi, Y.; Howe, G. A. Identification of a peroxisomal acyl-activating enzyme involved in the biosynthesis of jasmonic acid in Arabidopsis*♦. Journal of Biological Chemistry 2006, 281((44)), 33511–33520. [Google Scholar] [CrossRef]
- Zhang, H.-Y.; Wang, X.; Wang, X.-N.; Liu, H.-F.; Zhang, T.-T.; Wang, D.-R.; Liu, G.-D.; Liu, Y.-Q.; Song, X.-h.; Zhang, Z.; You, C. Brassinosteroids biosynthetic gene MdBR6OX2 regulates salt stress tolerance in both apple and Arabidopsis. Plant Physiology and Biochemistry 2024, 212, 108767. [Google Scholar] [CrossRef] [PubMed]
- Rattan, A.; Kapoor, D.; Ashish; Kapoor, N.; Bhardwaj, R.; Sharma, A., Chapter 12 - Involvement of brassinosteroids in plant response to salt stress. In Brassinosteroids in Plant Developmental Biology and Stress Tolerance, Ahammed, G. J.; Sharma, A.; Yu, J., Eds. Academic Press: 2022; pp 237-253.
- Zebosi, B.; Vollbrecht, E.; Best, N. B. Brassinosteroid biosynthesis and signaling: Conserved and diversified functions of core genes across multiple plant species. Plant Communications 2024, 100982. [Google Scholar] [CrossRef] [PubMed]
- Bajguz, A.; Chmur, M.; Gruszka, D. Comprehensive overview of the brassinosteroid biosynthesis pathways: substrates, products, inhibitors, and connections. Front Plant Sci 2020, 11, 1034. [Google Scholar] [CrossRef]
- Vriet, C.; Russinova, E.; Reuzeau, C. From squalene to brassinolide: the steroid metabolic and signaling pathways across the plant kingdom. Mol Plant 2013, 6((6)), 1738–57. [Google Scholar] [CrossRef]
- Zhao, B.; Li, J. Regulation of brassinosteroid biosynthesis and inactivation. J Integr Plant Biol 2012, 54((10)), 746–59. [Google Scholar] [CrossRef]
- Fujioka, S.; Yokota, T. Biosynthesis and metabolism of brassinosteroids. Annual Review of Plant Biology 2003, 54(2003), 137–164. [Google Scholar] [CrossRef]
- Lunn, J. E.; Delorge, I.; Figueroa, C. M.; Van Dijck, P.; Stitt, M. Trehalose metabolism in plants. Plant J 2014, 79((4)), 544–67. [Google Scholar] [CrossRef]
- Minami, A.; Yano, K.; Gamuyao, R.; Nagai, K.; Kuroha, T.; Ayano, M.; Nakamori, M.; Koike, M.; Kondo, Y.; Niimi, Y.; Kuwata, K.; Suzuki, T.; Higashiyama, T.; Takebayashi, Y.; Kojima, M.; Sakakibara, H.; Toyoda, A.; Fujiyama, A.; Kurata, N.; Ashikari, M.; Reuscher, S. Time-course transcriptomics analysis reveals key responses of submerged deepwater rice to flooding. Plant Physiol 2018, 176((4)), 3081–3102. [Google Scholar] [CrossRef]
- Yao, Y.; Yang, Y.; Pan, Y.; Liu, Z.; Hou, X.; Li, Y.; Zhang, H.; Wang, C.; Liao, W. Crucial roles of trehalose and 5-azacytidine in alleviating salt stress in tomato: Both synergistically and independently. Plant Physiology and Biochemistry 2023, 203, 108075. [Google Scholar] [CrossRef]
- Yang, Y.; Yao, Y.; Li, J.; Zhang, J.; Zhang, X.; Hu, L.; Ding, D.; Bakpa, E. P.; Xie, J. Trehalose alleviated salt stress in tomato by regulating ROS metabolism, photosynthesis, osmolyte synthesis, and trehalose metabolic pathways. Front Plant Sci 2022, 13, 772948. [Google Scholar] [CrossRef]
- Zhou, H.; Shi, H.; Yang, Y.; Feng, X.; Chen, X.; Xiao, F.; Lin, H.; Guo, Y. Insights into plant salt stress signaling and tolerance. Journal of Genetics and Genomics 2024, 51((1)), 16–34. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int J Mol Sci 2021, 22, (9).
- Cao, Y. R.; Chen, S. Y.; Zhang, J. S. Ethylene signaling regulates salt stress response: An overview. Plant Signal Behav 2008, 3((10)), 761–3. [Google Scholar] [CrossRef]
- Licausi, F.; Ohme-Takagi, M.; Perata, P. APETALA2/Ethylene Responsive Factor (AP2/ERF) transcription factors: mediators of stress responses and developmental programs. New Phytologist 2013, 199((3)), 639–649. [Google Scholar] [CrossRef] [PubMed]
- Cebrián, G.; Iglesias-Moya, J.; García, A.; Martínez, J.; Romero, J.; Regalado, J. J.; Martínez, C.; Valenzuela, J. L.; Jamilena, M. Involvement of ethylene receptors in the salt tolerance response of Cucurbita pepo. Horticulture Research 2021, 8((1)), 73. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Liu, J.; Lin, Z.; Sun, W.; Wu, Z.; Hu, H.; Zhang, Y. ERF transcription factors involved in salt response in tomato. Plant Growth Regulation 2018, 84((3)), 573–582. [Google Scholar] [CrossRef]
- Zhang, M.; Smith, J. A. C.; Harberd, N. P.; Jiang, C. The regulatory roles of ethylene and reactive oxygen species (ROS) in plant salt stress responses. Plant Molecular Biology 2016, 91((6)), 651–659. [Google Scholar] [CrossRef]
- Jiang, C.; Belfield, E. J.; Cao, Y.; Smith, J. A. C.; Harberd, N. P. An Arabidopsis soil-salinity–tolerance mutation confers ethylene-mediated enhancement of sodium/potassium homeostasis. The Plant Cell 2013, 25((9)), 3535–3552. [Google Scholar] [CrossRef]
- Peng, J.; Li, Z.; Wen, X.; Li, W.; Shi, H.; Yang, L.; Zhu, H.; Guo, H. Salt-induced stabilization of EIN3/EIL1 confers salinity tolerance by deterring ROS accumulation in Arabidopsis. PLoS Genet 2014, 10((10)), e1004664. [Google Scholar] [CrossRef]
- Wang, Y.; Diao, P.; Kong, L.; Yu, R.; Zhang, M.; Zuo, T.; Fan, Y.; Niu, Y.; Yan, F.; Wuriyanghan, H. Ethylene enhances seed germination and seedling growth under salinity by reducing oxidative stress and promoting chlorophyll content via ETR2 pathway. Front Plant Sci 2020, 11, 1066. [Google Scholar] [CrossRef]
- Li, H.; Sun, H.; Ping, W.; Liu, L.; Zhang, Y.; Zhang, K.; Bai, Z.; Li, A.; Zhu, J.; Li, C. Exogenous ethylene promotes the germination of cotton seeds under salt stress. Journal of Plant Growth Regulation 2023, 42((6)), 3923–3933. [Google Scholar] [CrossRef]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. dwarf and delayed-flowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor. Plant J 2004, 37((5)), 720–9. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Renou, J. P.; Berthomé, R.; Harberd, N. P.; Genschik, P. Plant DELLAs restrain growth and promote survival of adversity by reducing the levels of reactive oxygen species. Curr Biol 2008, 18((9)), 656–60. [Google Scholar] [CrossRef] [PubMed]
- Yuan, K.; Rashotte, A. M.; Wysocka-Diller, J. W. ABA and GA signaling pathways interact and regulate seed germination and seedling development under salt stress. Acta Physiologiae Plantarum 2011, 33((2)), 261–271. [Google Scholar] [CrossRef]
- Zhu, J. K. Abiotic stress signaling and responses in plants. Cell 2016, 167((2)), 313–324. [Google Scholar] [CrossRef]
- Yang, G.; Yu, Z.; Gao, L.; Zheng, C. SnRK2s at the crossroads of growth and stress responses. Trends Plant Sci 2019, 24((8)), 672–676. [Google Scholar] [CrossRef]
- Szymańska, K. P.; Polkowska-Kowalczyk, L.; Lichocka, M.; Maszkowska, J.; Dobrowolska, G. SNF1-related protein kinases SnRK2.4 and SnRK2.10 modulate ROS homeostasis in plant response to salt stress. Int J Mol Sci 2019, 20, (1).
- Delgado, C.; Mora-Poblete, F.; Ahmar, S.; Chen, J. T.; Figueroa, C. R. Jasmonates and plant salt stress: molecular players, physiological effects, and improving tolerance by using genome-associated tools. Int J Mol Sci 2021, 22, (6).
- Gao, Z.; Gao, S.; Li, P.; Zhang, Y.; Ma, B.; Wang, Y. Exogenous methyl jasmonate promotes salt stress-induced growth inhibition and prioritizes defense response of Nitraria tangutorum Bobr. Physiologia Plantarum 2021, 172((1)), 162–175. [Google Scholar] [CrossRef]
- Nolan, T. M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: multidimensional regulators of plant growth, development, and stress responses. Plant Cell 2020, 32((2)), 295–318. [Google Scholar] [CrossRef]
- Jia, C.; Zhao, S.; Bao, T.; Zhao, P.; Peng, K.; Guo, Q.; Gao, X.; Qin, J. Tomato BZR/BES transcription factor SlBZR1 positively regulates BR signaling and salt stress tolerance in tomato and Arabidopsis. Plant Sci 2021, 302, 110719. [Google Scholar] [CrossRef]
- Özdemir, F.; Bor, M.; Demiral, T.; Türkan, İ. Effects of 24-epibrassinolide on seed germination, seedling growth, lipid peroxidation, proline content and antioxidative system of rice (Oryza sativa L.) under salinity stress. Plant Growth Regulation 2004, 42, (3), 203-211.
- Wani, A. S.; Ahmad, A.; Hayat, S.; Tahir, I. Epibrassinolide and proline alleviate the photosynthetic and yield inhibition under salt stress by acting on antioxidant system in mustard. Plant Physiol Biochem 2019, 135, 385–394. [Google Scholar] [CrossRef]
- Su, Q.; Zheng, X.; Tian, Y.; Wang, C. Exogenous Brassinolide Alleviates Salt Stress in Malus hupehensis Rehd. by Regulating the Transcription of NHX-Type Na(+)(K(+))/H(+) Antiporters. Front Plant Sci 2020, 11, 38.
- Xia, X. J.; Chen, Z.; Yu, J. Q. ROS mediate brassinosteroids-induced plant stress responses. Plant Signal Behav 2010, 5((5)), 532–4. [Google Scholar] [CrossRef]
- Abdelraheem, A.; Esmaeili, N.; O’Connell, M.; Zhang, J. Progress and perspective on drought and salt stress tolerance in cotton. Industrial Crops and Products 2019, 130, 118–129. [Google Scholar] [CrossRef]
- Cao, J.-F.; Huang, J.-Q.; Liu, X.; Huang, C.-C.; Zheng, Z.-S.; Zhang, X.-F.; Shangguan, X.-X.; Wang, L.-J.; Zhang, Y.-G.; Wendel, J. F.; Grover, C. E.; Chen, Z.-W. Genome-wide characterization of the GRF family and their roles in response to salt stress in Gossypium. BMC Genomics 2020, 21((1)), 575. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Hao, J.; Su, Y.; Li, B.; Zhao, N.; Zhu, M.; Huang, Y.; Tian, B.; Shi, G.; Hua, J. Two aquaporin genes, GhPIP2;7 and GhTIP2;1, positively regulate the tolerance of upland cotton to salt and osmotic stresses. Front Plant Sci 2021, 12, 780486. [Google Scholar] [CrossRef]
- Yang, L.; Wang, X.; Zhao, F.; Zhang, X.; Li, W.; Huang, J.; Pei, X.; Ren, X.; Liu, Y.; He, K.; Zhang, F.; Ma, X.; Yang, D. Roles of S-adenosylmethionine and its derivatives in salt tolerance of cotton. International Journal of Molecular Sciences 2023, 24((11)), 9517. [Google Scholar] [CrossRef]
- Nawaz, M.; Hassan, M. U.; Chattha, M. U.; Mahmood, A.; Shah, A. N.; Hashem, M.; Alamri, S.; Batool, M.; Rasheed, A.; Thabit, M. A.; Alhaithloul, H. A. S.; Qari, S. H. Trehalose: a promising osmo-protectant against salinity stress—physiological and molecular mechanisms and future prospective. Molecular Biology Reports 2022, 49((12)), 11255–11271. [Google Scholar] [CrossRef]
- Shahzad, A. N.; Qureshi, M. K.; Ullah, S.; Latif, M.; Ahmad, S.; Bukhari, S. A. H. Exogenous trehalose improves cotton growth by modulating antioxidant defense under salinity-induced osmotic stress. Pakistan Journal of Agricultural Research 2020, 33((2)), 270–279. [Google Scholar] [CrossRef]
Figure 1.
Salt sensitivity assessment of Su-mian 3 and Jin-mian 25 cotton plants. A: The phenotype of Su-mian 3 (SS) seedlings grown in distilled water (CK) for seven days; B: The phenotype of Su-mian 3 (SS) seedlings grown in 150mM NaCl solution (150) for seven days; C: Fresh weight of Su-mian 3 seedlings between CK and salt stress; D: Length of Su-mian 3 seedlings between CK and salt stress; E: Root length of Su-mian 3 seedlings between CK and salt stress; F: Hypocotyl length of Su-mian 3 seedlings between CK and salt stress; G: Germination potential of Su-mian 3 seeds; H: Germination rate of Su-mian 3 seeds; I: The phenotype of Jin-mian 25 (ST) seedlings grown in distilled water (CK) for seven days; J: The phenotype of Jin-mian 25 (ST) seedlings grown in 150mM NaCl solution (150) for seven days; K: Fresh weight of Jin-mian 25 seedlings between CK and salt stress; L: Length of Jin-mian 25 seedlings between CK and salt stress; M: Root length of Jin-mian 25 seedlings between CK and salt stress; N: Hypocotyl length of Jin-mian 25 seedlings between CK and salt stress; O: Germination potential of Jin-mian 25 seeds; P: Germination rate of Jin-mian 25 seeds. Note: Su-mian 3 is a salt-sensitive cultivar (SS-Salt Sensitive), and Jin-mian 25 is a salt-tolerant cultivar (ST-Salt Tolerant). Values are mean ± SD, n = 5, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, Student’s t-test.
Figure 1.
Salt sensitivity assessment of Su-mian 3 and Jin-mian 25 cotton plants. A: The phenotype of Su-mian 3 (SS) seedlings grown in distilled water (CK) for seven days; B: The phenotype of Su-mian 3 (SS) seedlings grown in 150mM NaCl solution (150) for seven days; C: Fresh weight of Su-mian 3 seedlings between CK and salt stress; D: Length of Su-mian 3 seedlings between CK and salt stress; E: Root length of Su-mian 3 seedlings between CK and salt stress; F: Hypocotyl length of Su-mian 3 seedlings between CK and salt stress; G: Germination potential of Su-mian 3 seeds; H: Germination rate of Su-mian 3 seeds; I: The phenotype of Jin-mian 25 (ST) seedlings grown in distilled water (CK) for seven days; J: The phenotype of Jin-mian 25 (ST) seedlings grown in 150mM NaCl solution (150) for seven days; K: Fresh weight of Jin-mian 25 seedlings between CK and salt stress; L: Length of Jin-mian 25 seedlings between CK and salt stress; M: Root length of Jin-mian 25 seedlings between CK and salt stress; N: Hypocotyl length of Jin-mian 25 seedlings between CK and salt stress; O: Germination potential of Jin-mian 25 seeds; P: Germination rate of Jin-mian 25 seeds. Note: Su-mian 3 is a salt-sensitive cultivar (SS-Salt Sensitive), and Jin-mian 25 is a salt-tolerant cultivar (ST-Salt Tolerant). Values are mean ± SD, n = 5, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, Student’s t-test.
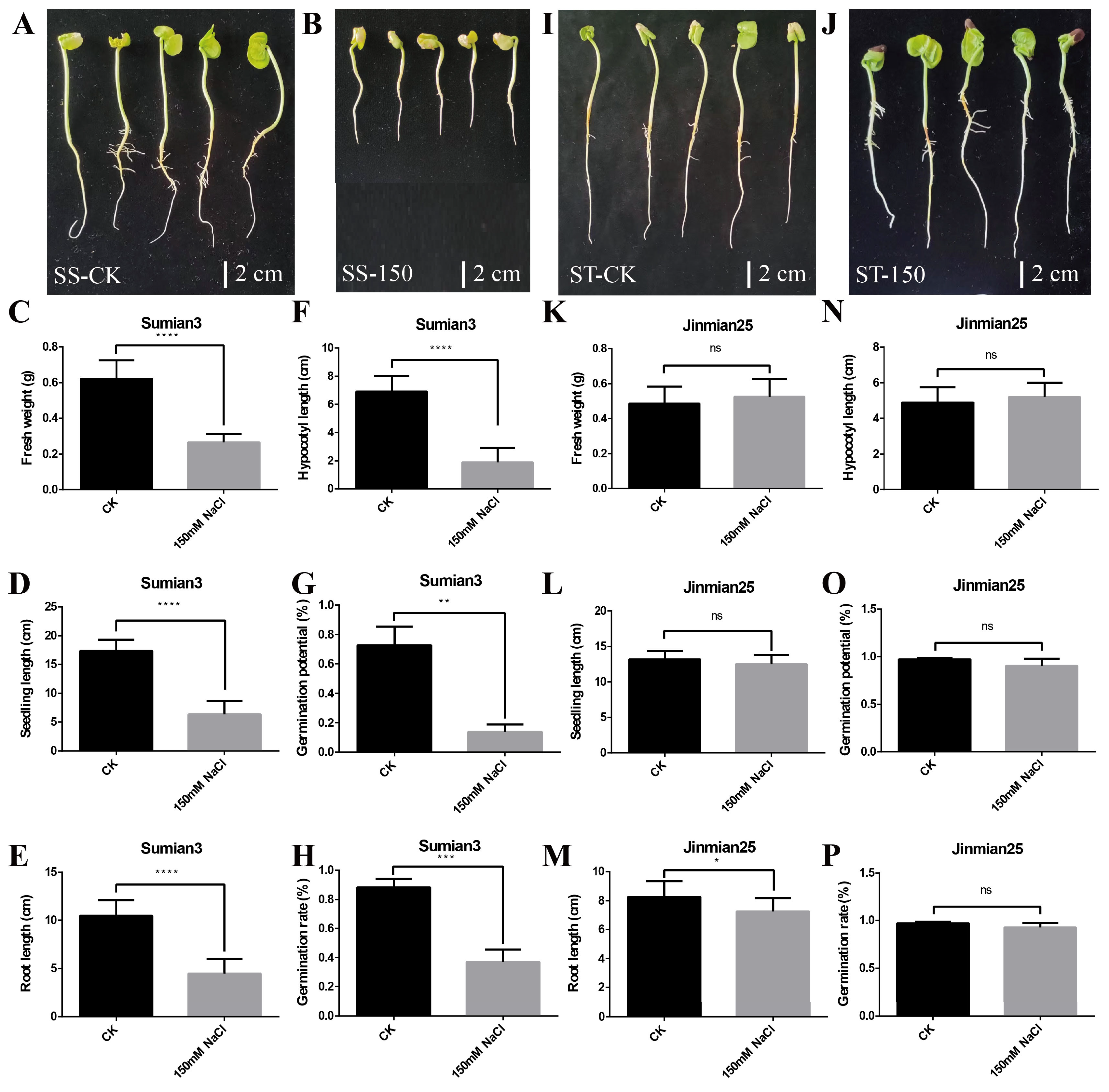
Figure 2.
Seeds of cv Su-mian 3 and Jin-mian 25 treated by 150mM NaCl solution for 0, 6, 12, 24, and 72h for expression profile analyses and identification of DEGs. A: Seed germination and seedling growth of cv Su-mian 3 and Jin-mian 25 under salt stress. B: Number of DEGs between the comparisons of SS0 vs ST0, SS6 vs ST6, SS12 vs ST12, SS24 vs ST24, and SS72 vs ST72 groups.
Figure 2.
Seeds of cv Su-mian 3 and Jin-mian 25 treated by 150mM NaCl solution for 0, 6, 12, 24, and 72h for expression profile analyses and identification of DEGs. A: Seed germination and seedling growth of cv Su-mian 3 and Jin-mian 25 under salt stress. B: Number of DEGs between the comparisons of SS0 vs ST0, SS6 vs ST6, SS12 vs ST12, SS24 vs ST24, and SS72 vs ST72 groups.
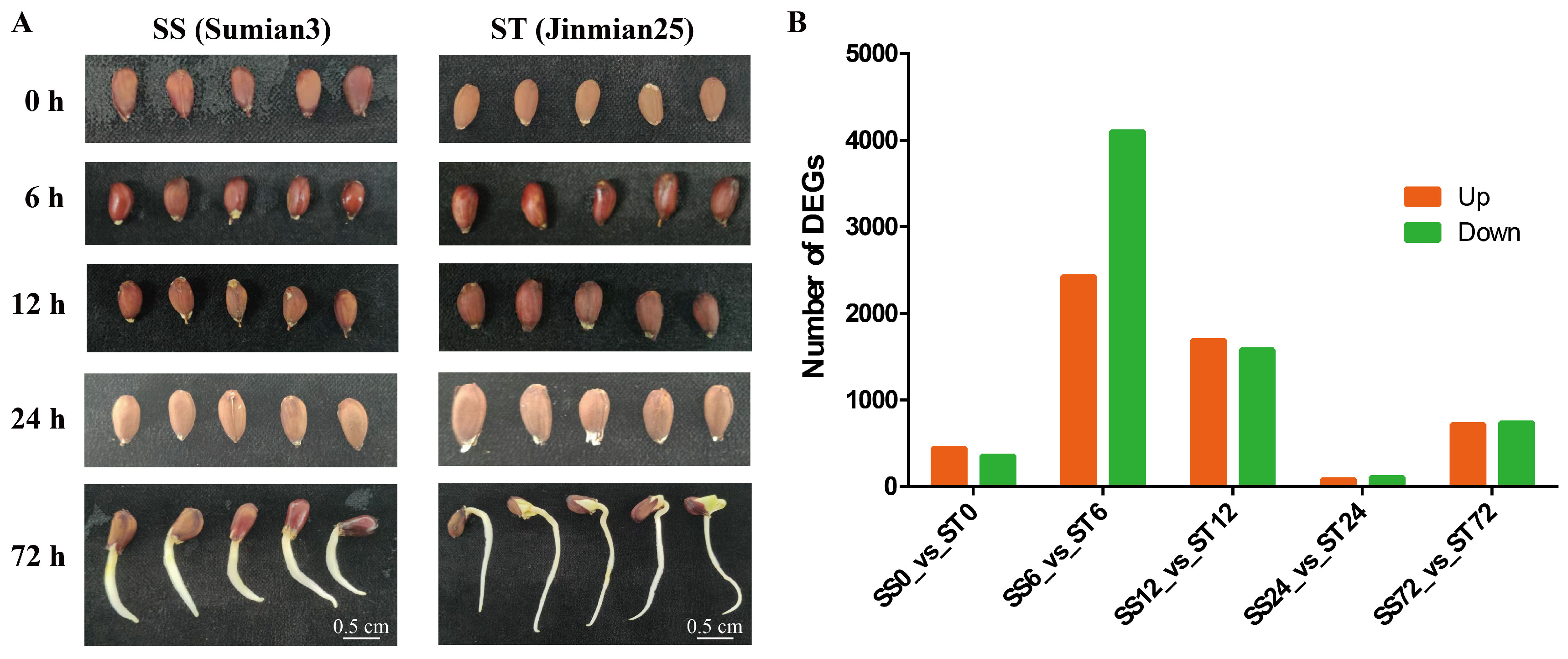
Figure 3.
KEGG pathway analysis of enriched DEGs. A: Top 20 pathways of significantly enriched DEGs from SS0 vs ST0; B: Top 20 pathways of significantly enriched DEGs from SS6 vs ST6; C: Top 20 pathways of significantly enriched DEGs from SS12 vs ST12; D: Top 20 pathways of significantly enriched DEGs from SS24 vs ST24; E: Top 20 pathways of significantly enriched DEGs from SS72 vs ST72; F: Venn diagram comparison of DEGs from five time points after 150 mM NaCl stress between two cotton cultivars.
Figure 3.
KEGG pathway analysis of enriched DEGs. A: Top 20 pathways of significantly enriched DEGs from SS0 vs ST0; B: Top 20 pathways of significantly enriched DEGs from SS6 vs ST6; C: Top 20 pathways of significantly enriched DEGs from SS12 vs ST12; D: Top 20 pathways of significantly enriched DEGs from SS24 vs ST24; E: Top 20 pathways of significantly enriched DEGs from SS72 vs ST72; F: Venn diagram comparison of DEGs from five time points after 150 mM NaCl stress between two cotton cultivars.
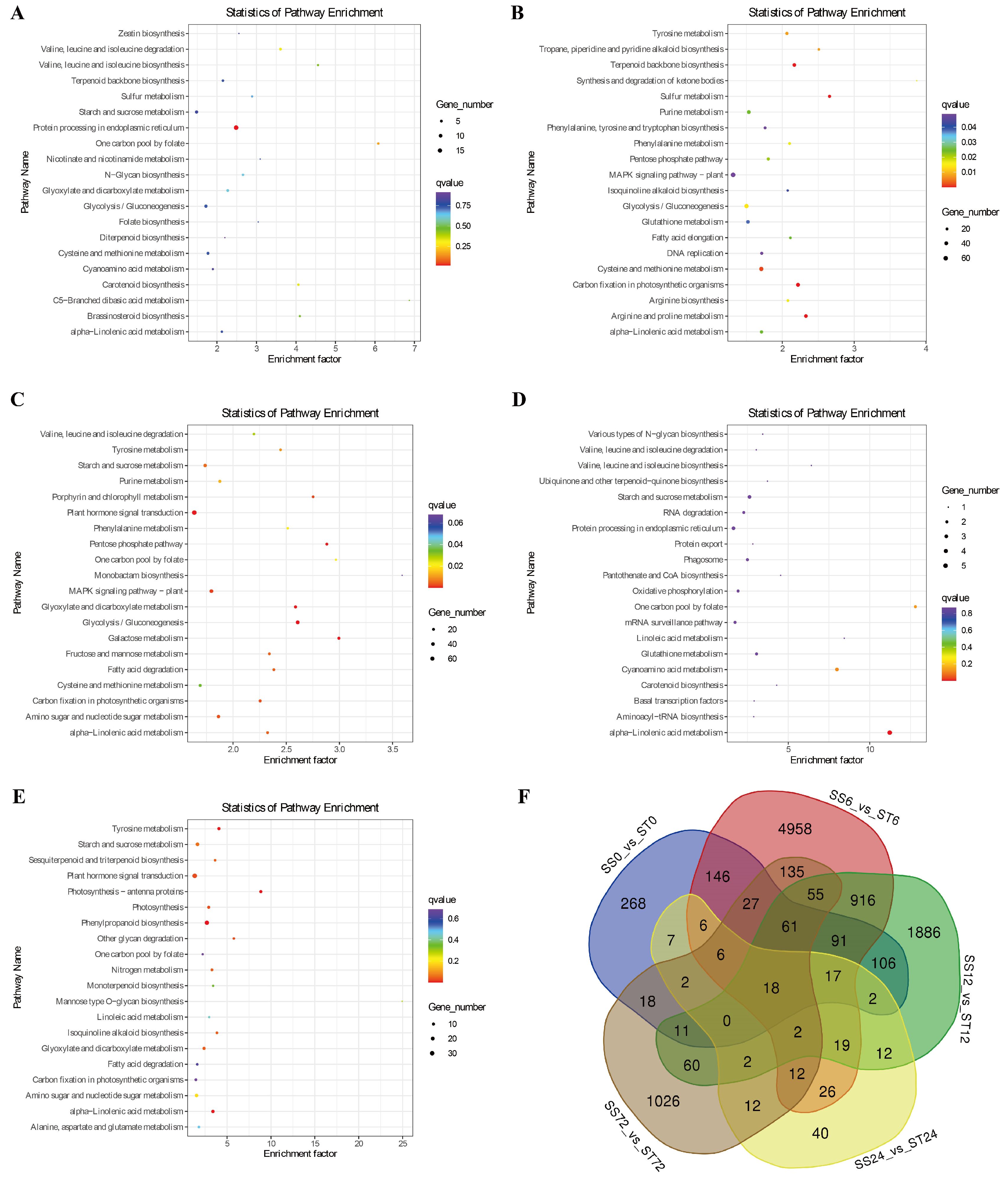
Figure 4.
Expression patterns of different pathway genes during salt stress in both genotypes of cotton. A: Shared DEGs of two cotton cultivars from five-time points after salt stress; B: Aquaporin genes; C: RLK genes; D: CIPK genes; E: ROS scavenging enzyme genes.
Figure 4.
Expression patterns of different pathway genes during salt stress in both genotypes of cotton. A: Shared DEGs of two cotton cultivars from five-time points after salt stress; B: Aquaporin genes; C: RLK genes; D: CIPK genes; E: ROS scavenging enzyme genes.
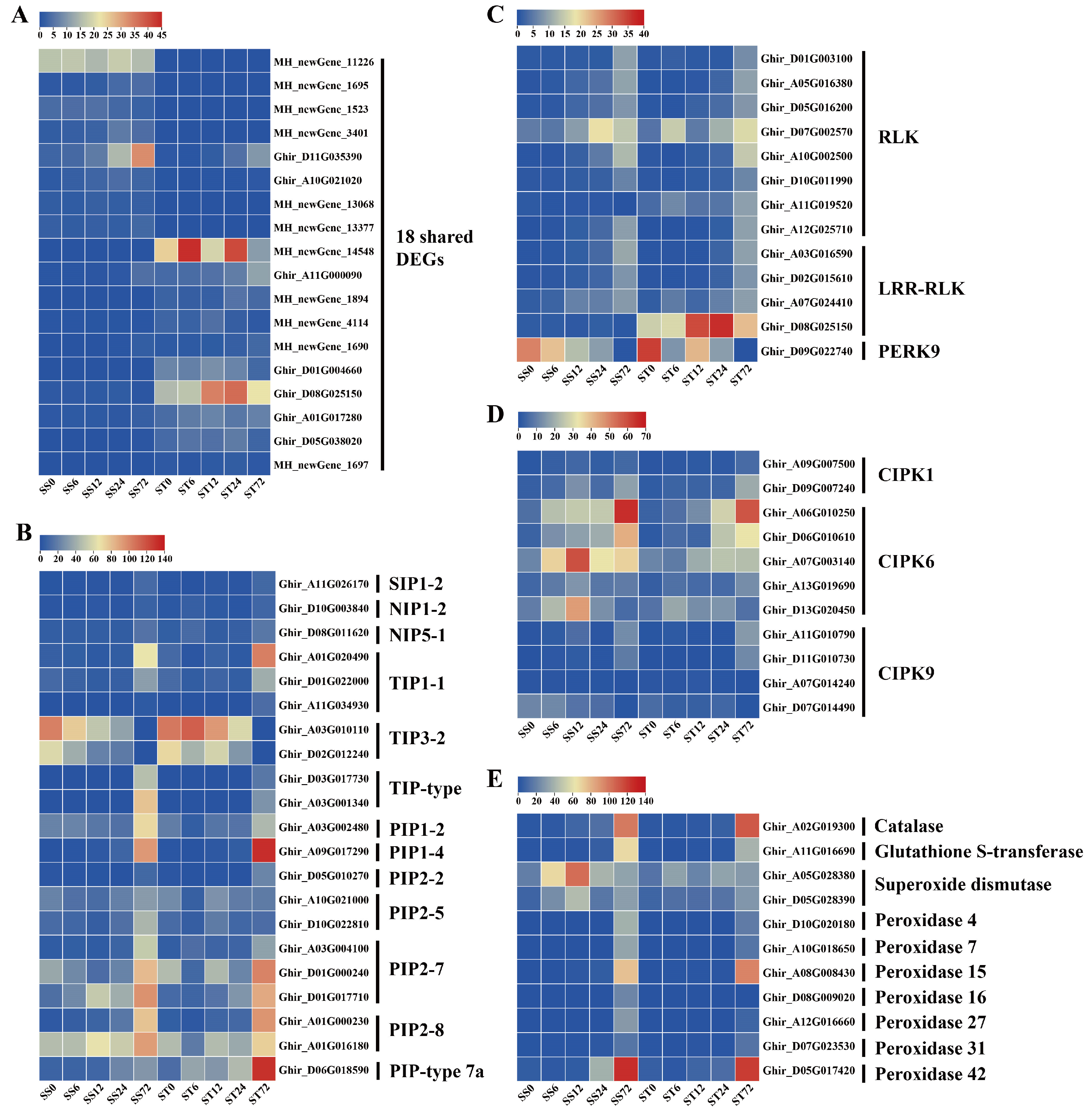
Figure 5.
Expression of genes related to ethylene biosynthesis and signaling during salt stress. A: Schematic overview of ethylene biosynthesis and signaling pathway. Expression patterns of ethylene biosynthesis genes in both genotypes during salt stress, including ACS genes (B), and ACO genes (C). Expression patterns of ethylene signaling pathway genes in two genotypes during salt stress, including receptors genes (D), inhibitor CTR1 genes (E), inhibitor EIN2 genes (F) and transcription factor EIN3 genes (G). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow while the highest expression is indicated in red.
Figure 5.
Expression of genes related to ethylene biosynthesis and signaling during salt stress. A: Schematic overview of ethylene biosynthesis and signaling pathway. Expression patterns of ethylene biosynthesis genes in both genotypes during salt stress, including ACS genes (B), and ACO genes (C). Expression patterns of ethylene signaling pathway genes in two genotypes during salt stress, including receptors genes (D), inhibitor CTR1 genes (E), inhibitor EIN2 genes (F) and transcription factor EIN3 genes (G). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow while the highest expression is indicated in red.
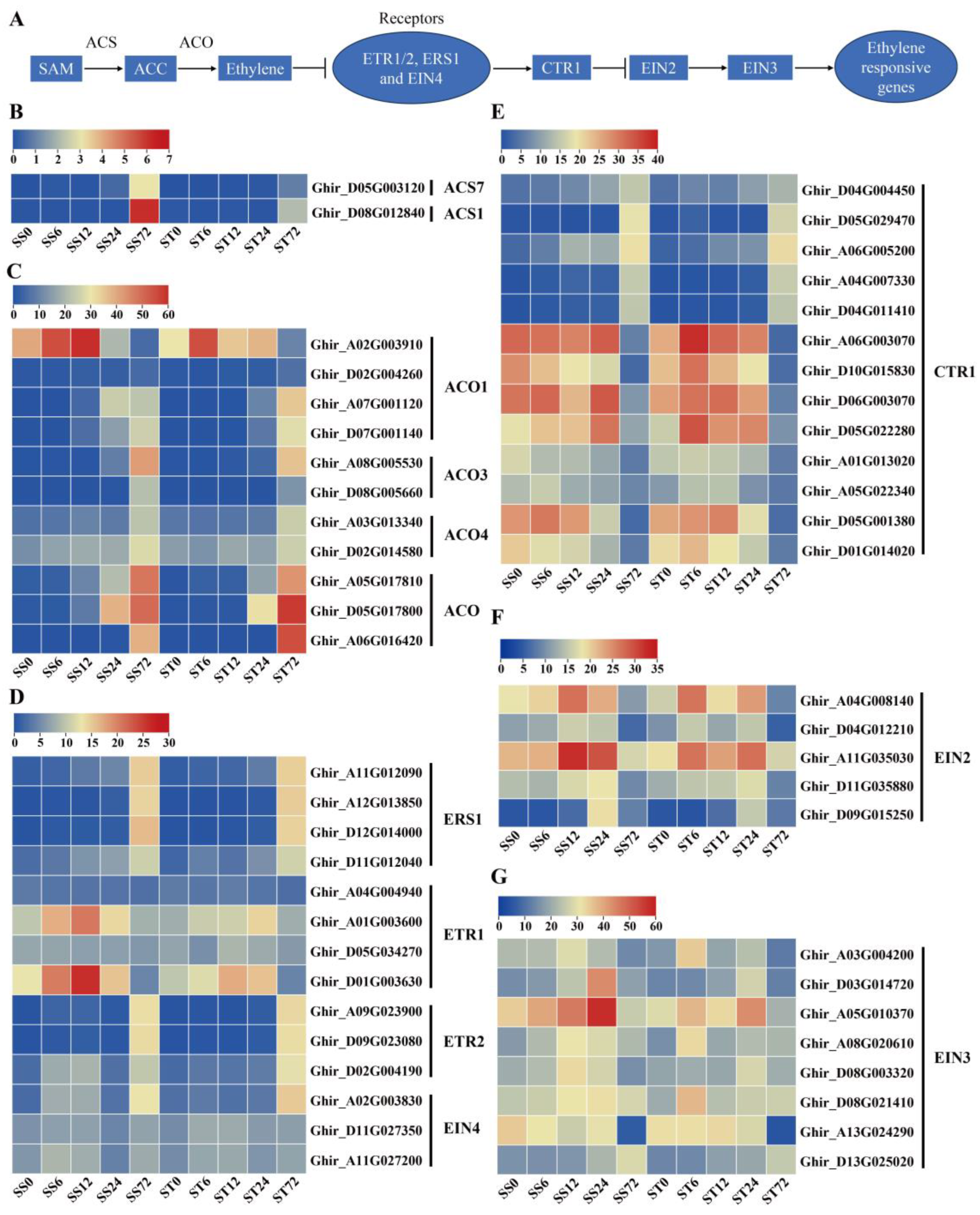
Figure 6.
Expression of genes related to GA biosynthesis and signaling during salt stress. A: Schematic overview of GA biosynthesis and signaling pathway. Expression patterns of GA biosynthesis genes in both genotypes during salt stress, including CPS genes (B), KS genes (C), KO genes (D), KAO genes (E), GA20ox genes (F). Expression patterns of GA signaling pathway genes in two genotypes during salt stress, including GA2ox genes (G), receptor GID genes (H), and inhibitor DELLA genes (I). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow while the highest expression is indicated in red.
Figure 6.
Expression of genes related to GA biosynthesis and signaling during salt stress. A: Schematic overview of GA biosynthesis and signaling pathway. Expression patterns of GA biosynthesis genes in both genotypes during salt stress, including CPS genes (B), KS genes (C), KO genes (D), KAO genes (E), GA20ox genes (F). Expression patterns of GA signaling pathway genes in two genotypes during salt stress, including GA2ox genes (G), receptor GID genes (H), and inhibitor DELLA genes (I). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow while the highest expression is indicated in red.
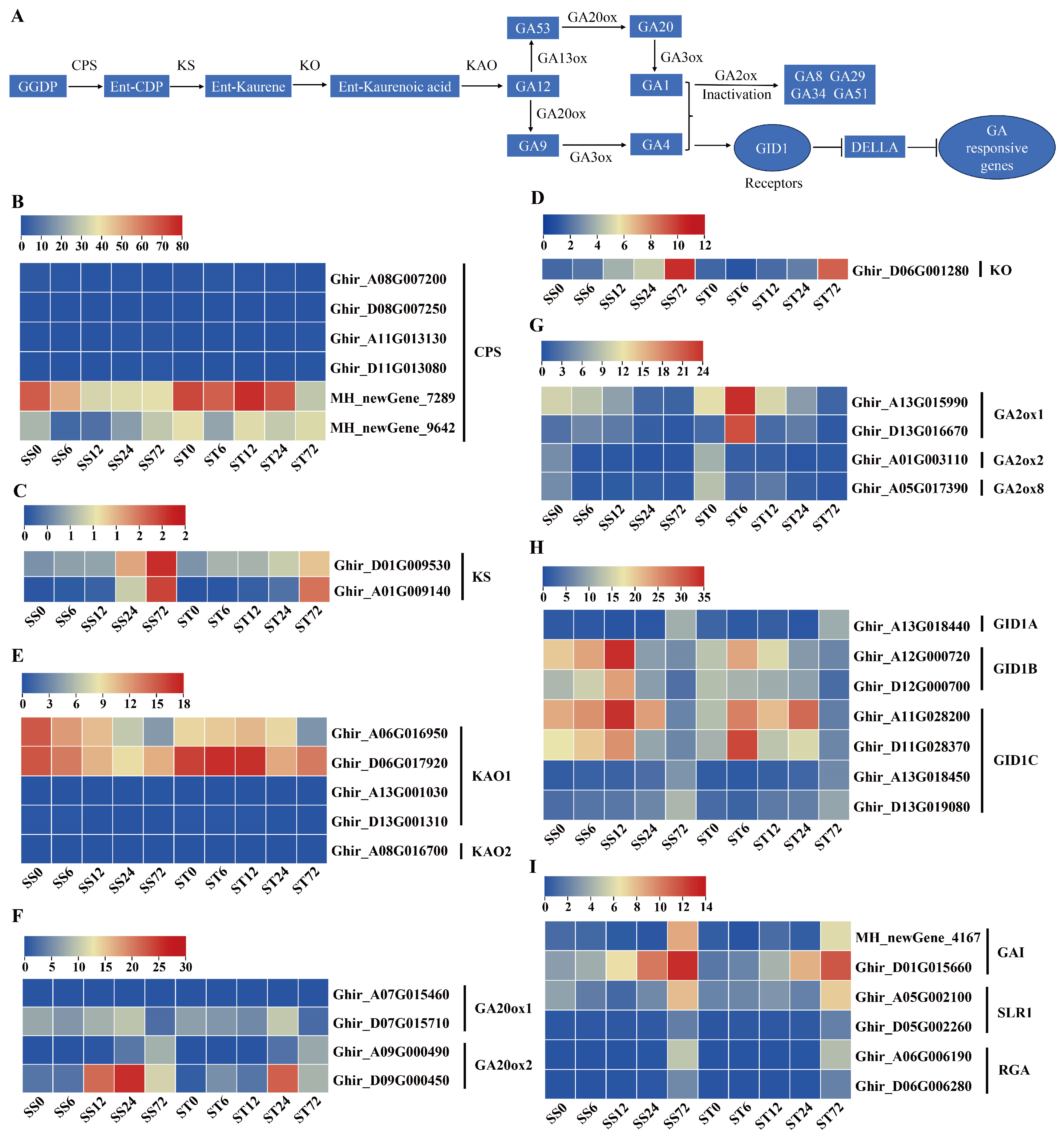
Figure 7.
Expression of genes related to ABA biosynthesis and signaling during salt stress. A: Schematic overview of ABA biosynthesis and signaling pathway. Expression patterns of ABA biosynthesis genes in both genotypes during salt stress, including CHY-β and ZEP genes (B), ABA4 genes (C), NCED genes (D), ABA2 genes (E), ABA3 and AAO3 genes (F). Expression patterns of ABA signaling pathway genes in two genotypes during salt stress, including ABA8ox genes (G), PLY receptor genes (H), PP2C genes (I), SnRK2 genes (J), and ABI3, ABFs or BAM1 genes (K). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow while the highest expression is indicated in red.
Figure 7.
Expression of genes related to ABA biosynthesis and signaling during salt stress. A: Schematic overview of ABA biosynthesis and signaling pathway. Expression patterns of ABA biosynthesis genes in both genotypes during salt stress, including CHY-β and ZEP genes (B), ABA4 genes (C), NCED genes (D), ABA2 genes (E), ABA3 and AAO3 genes (F). Expression patterns of ABA signaling pathway genes in two genotypes during salt stress, including ABA8ox genes (G), PLY receptor genes (H), PP2C genes (I), SnRK2 genes (J), and ABI3, ABFs or BAM1 genes (K). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow while the highest expression is indicated in red.
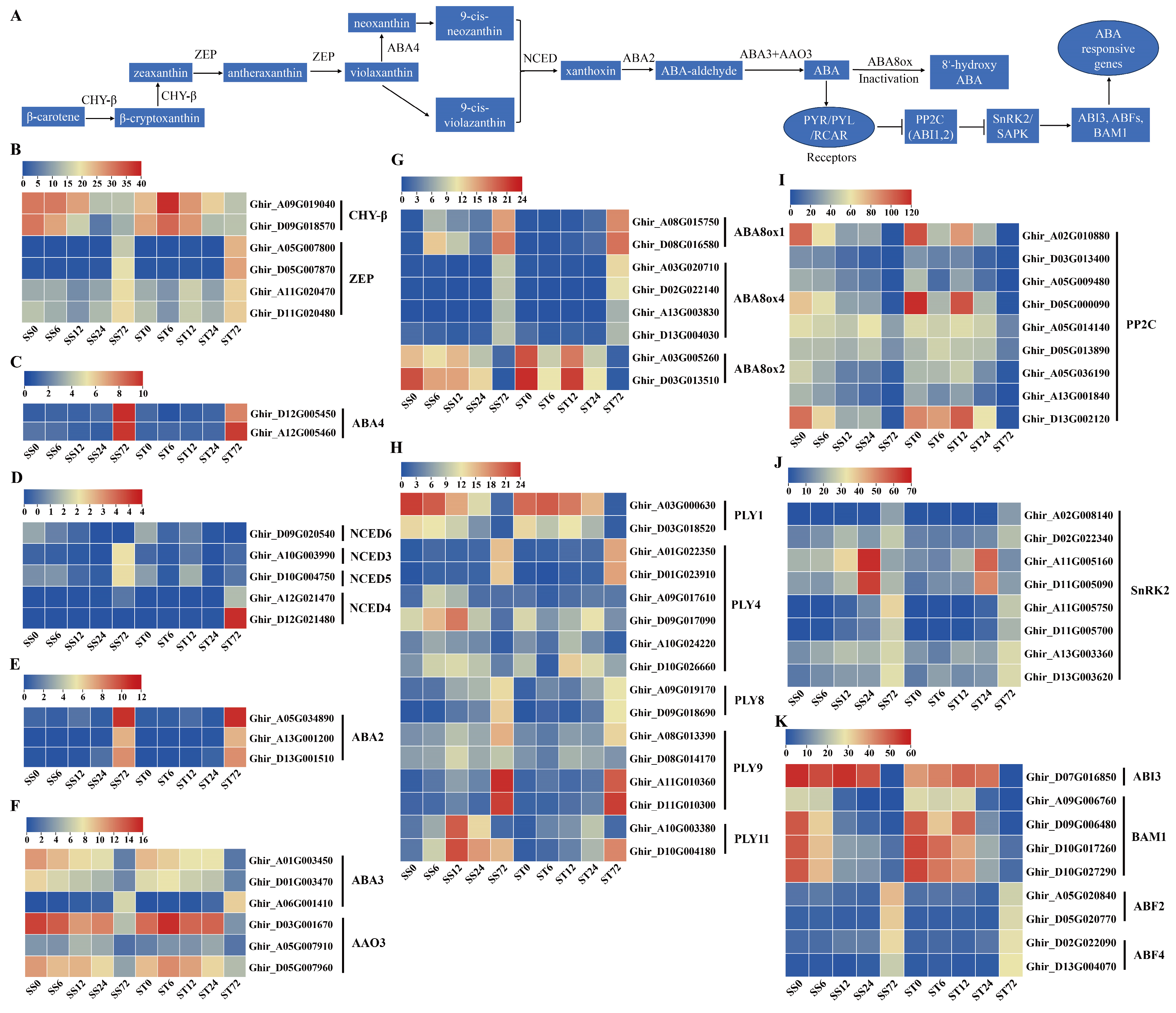
Figure 8.
Expression of genes related to JA biosynthesis and signaling during salt stress. A: Schematic overview of JA biosynthesis and signaling pathway. Expression patterns of JA biosynthesis genes in both genotypes during salt stress, including FAD genes (B), PLA1 and DAD1 genes (C), LOX genes (D), AOS genes (E), AOC genes (F), OPR3 genes (G), and OPCL1 genes (H). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow and the highest expression is indicated in red.
Figure 8.
Expression of genes related to JA biosynthesis and signaling during salt stress. A: Schematic overview of JA biosynthesis and signaling pathway. Expression patterns of JA biosynthesis genes in both genotypes during salt stress, including FAD genes (B), PLA1 and DAD1 genes (C), LOX genes (D), AOS genes (E), AOC genes (F), OPR3 genes (G), and OPCL1 genes (H). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow and the highest expression is indicated in red.
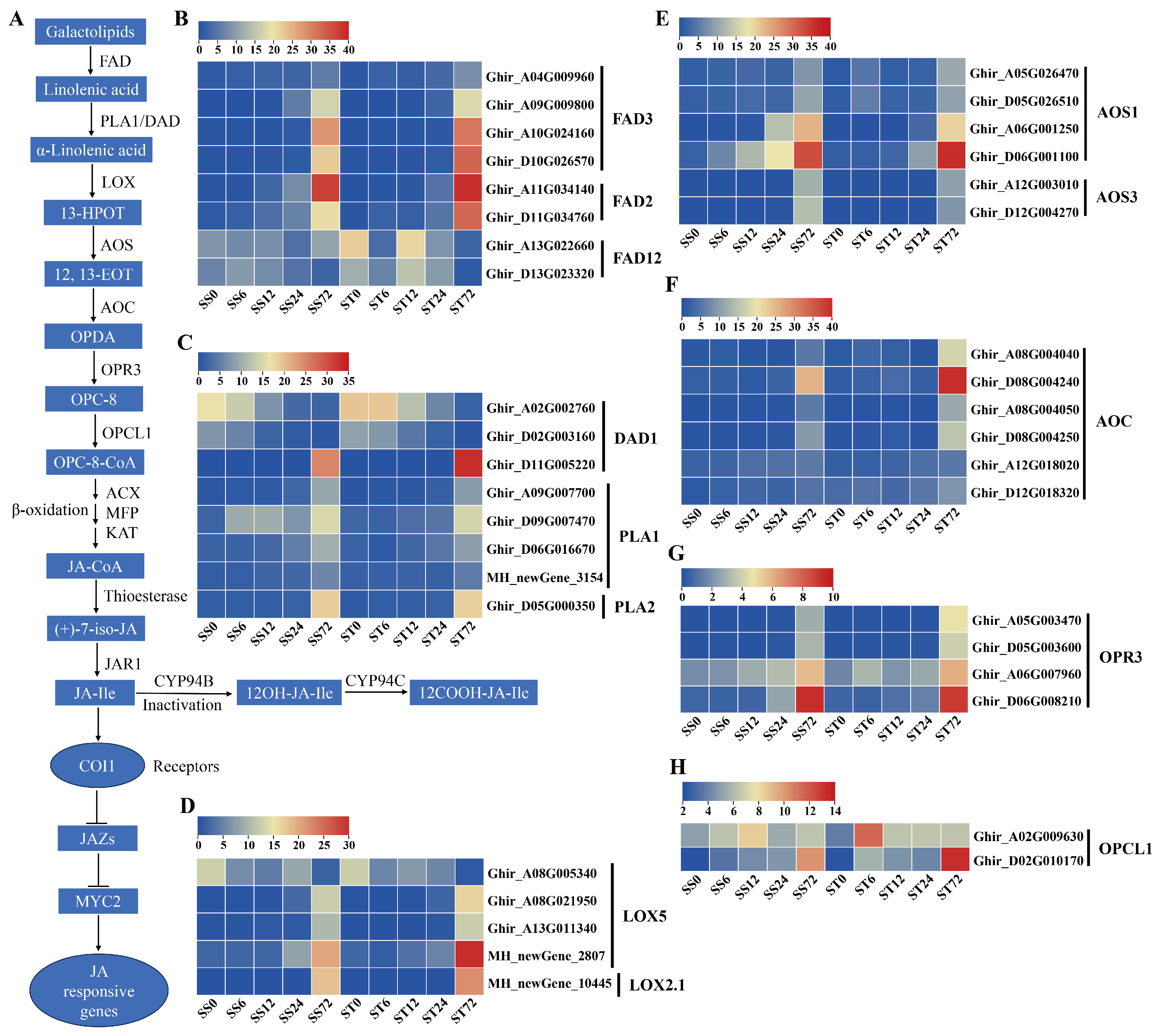
Figure 9.
Expression of genes related to JA biosynthesis and signaling during salt stress. A: Expression patterns of JA biosynthesis genes in both genotypes during salt stress, including ACX genes (A), MFP genes (B), KAT genes (C), Thioesterase genes (D), and JAR1 genes (E). Expression patterns of JA signaling pathway genes in two genotypes during salt stress, including CYP94 genes (F), COI1 genes (G), JAZ genes (H), and MYC2 genes (I). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower expression levels are represented in blue, higher expression is indicated in yellow and the highest expression is indicated in red.
Figure 9.
Expression of genes related to JA biosynthesis and signaling during salt stress. A: Expression patterns of JA biosynthesis genes in both genotypes during salt stress, including ACX genes (A), MFP genes (B), KAT genes (C), Thioesterase genes (D), and JAR1 genes (E). Expression patterns of JA signaling pathway genes in two genotypes during salt stress, including CYP94 genes (F), COI1 genes (G), JAZ genes (H), and MYC2 genes (I). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower expression levels are represented in blue, higher expression is indicated in yellow and the highest expression is indicated in red.
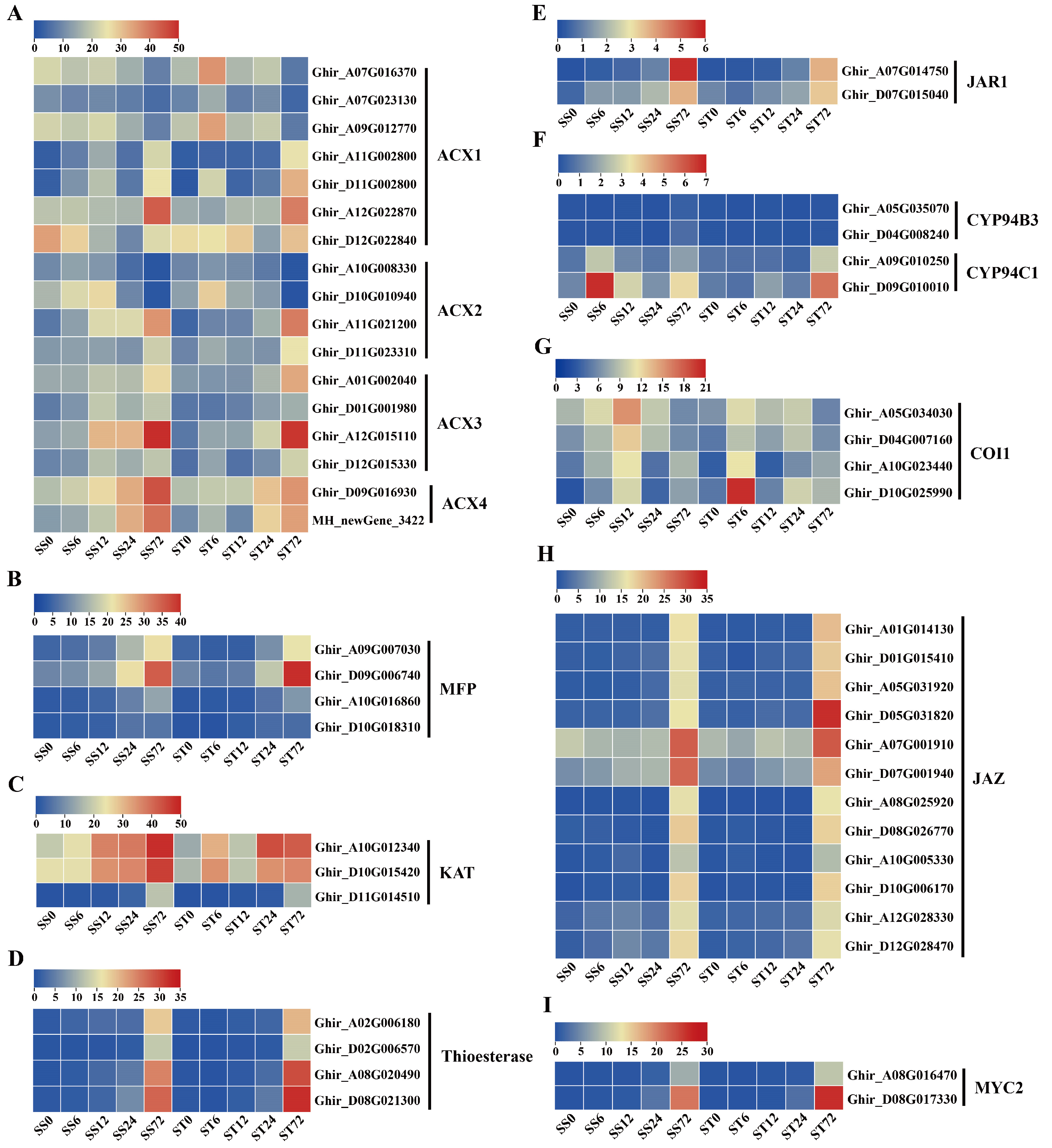
Figure 10.
Expression of genes related to BR biosynthesis and signaling during salt stress. A: Schematic overview of BR biosynthesis and signaling pathway. Expression patterns of BR biosynthesis genes in both genotypes during salt stress, including DWF4 genes (B), DET2 genes (C), CPD genes (D), ROT3 genes (E), CYP90D1 genes (F), and CYP85A1 genes (G). Expression patterns of BR signaling pathway genes in two genotypes during salt stress, including BRI and BAK1 receptor genes (H), and BIN2 genes (I). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow and the highest expression is indicated in red.
Figure 10.
Expression of genes related to BR biosynthesis and signaling during salt stress. A: Schematic overview of BR biosynthesis and signaling pathway. Expression patterns of BR biosynthesis genes in both genotypes during salt stress, including DWF4 genes (B), DET2 genes (C), CPD genes (D), ROT3 genes (E), CYP90D1 genes (F), and CYP85A1 genes (G). Expression patterns of BR signaling pathway genes in two genotypes during salt stress, including BRI and BAK1 receptor genes (H), and BIN2 genes (I). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow and the highest expression is indicated in red.
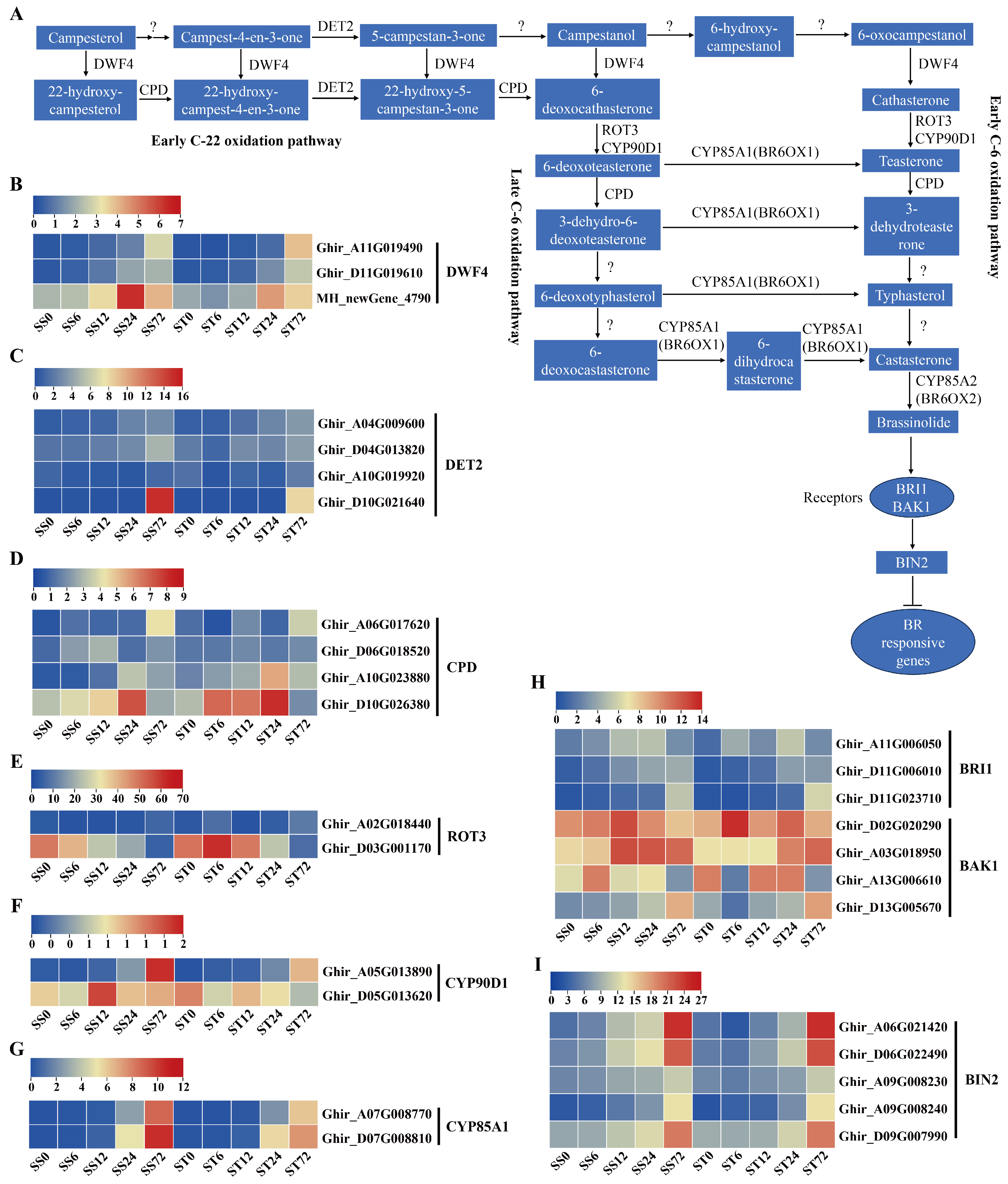
Figure 11.
Expression of genes related to trehalose metabolism during salt stress. A: Schematic overview of trehalose biosynthesis and degradation pathway. Expression patterns of trehalose biosynthesis and degradation genes during salt stress in both genotypes, including TPS genes (B), TPP genes (C), and TRE1 genes (D). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow and the highest expression is indicated in red.
Figure 11.
Expression of genes related to trehalose metabolism during salt stress. A: Schematic overview of trehalose biosynthesis and degradation pathway. Expression patterns of trehalose biosynthesis and degradation genes during salt stress in both genotypes, including TPS genes (B), TPP genes (C), and TRE1 genes (D). Each row represents one gene, columns represent samples from different salt stress time points and colors represent gene expression levels as FPKM values. Lower levels of expression are represented in blue, higher expression is indicated in yellow and the highest expression is indicated in red.
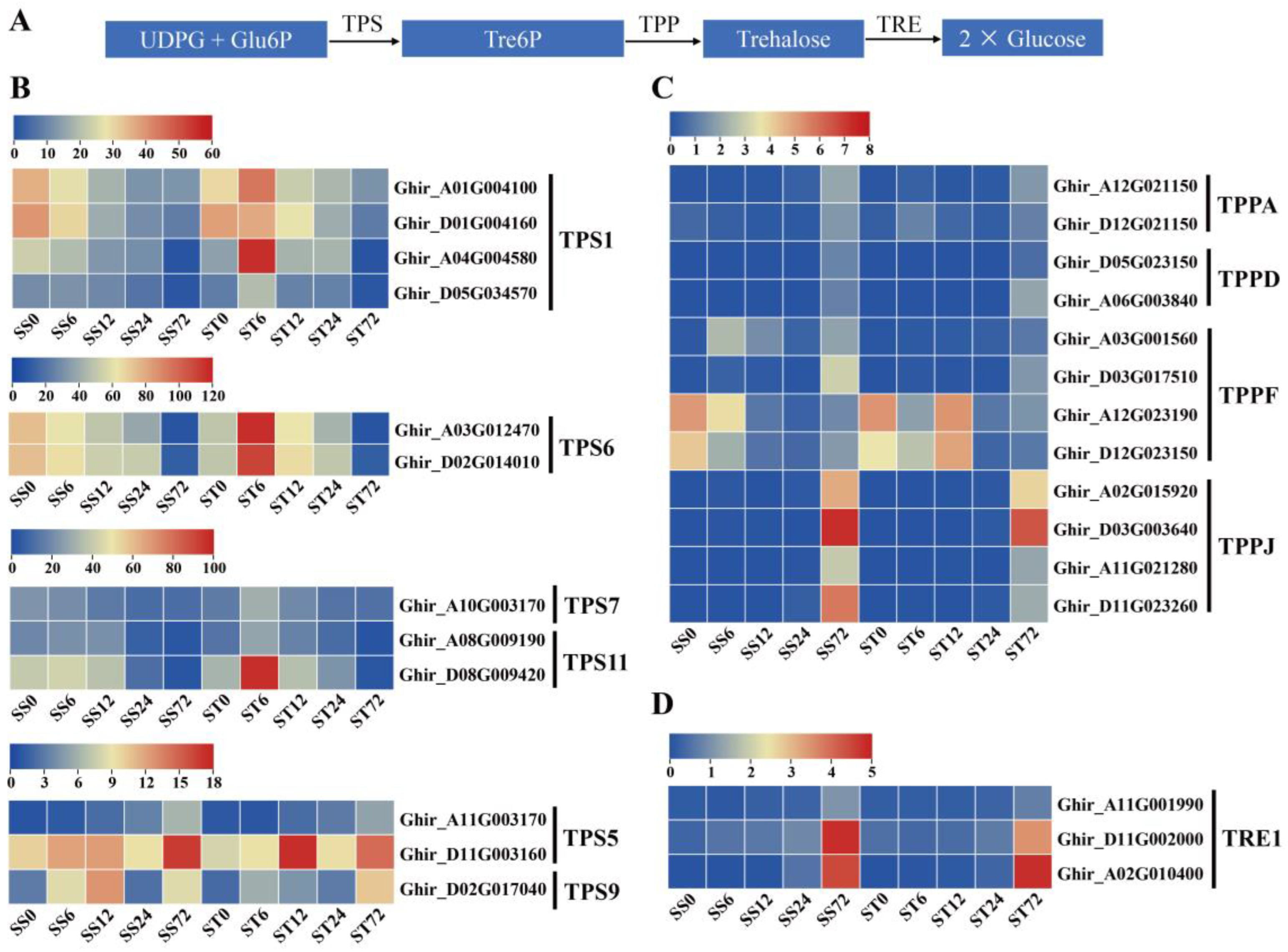
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



