
Submitted:
18 October 2024
Posted:
18 October 2024
You are already at the latest version
Abstract
Native garden plants significantly contribute to the conservation of biodiversity and ecosystem functions in urban environments. This study aimed to identify the physiochemical differences among native grasses subjected to drought and salinity stress and to assess their potential as garden plants adaptable to outdoor conditions and global climate change. Physiological parameters, such as photosynthetic rate, water use efficiency, and chlorophyll and carotenoid contents, decreased in Melica grandiflora and Carex forficula with increasing drought or salinity. In contrast, Carex boottiana maintained high water use efficiency and stable chlorophyll and carotenoid levels even under se-vere drought and salinity conditions. Biochemical analyses showed that C. boottiana had lower levels of malondialdehyde and reactive oxygen species (including O2.- and H2O2) than those in the other species under stress conditions. Principal component analysis revealed clear differences in tolerance levels among the grass species. C. boottiana demonstrated high adaptability to both drought and salinity stress, indicating its potential as a sustainable and resilient garden material for urban landscapes facing severe climatic challenges.
Keywords:
drought
; garden material
; native grass
; physiochemical attributes
; salinity
1. Introduction
The global population is rapidly urbanizing; therefore, the protection of ecosystem services and biodiversity in urban areas has become increasingly vital. Vegetation plays an essential role in these environments by providing breeding habitats and altering microclimates [1]. Consequently, gardens are considered significant contributors to biological diversity [2,3,4]. In particular, native plants can positively impact urban biodiversity conservation as they are well suited to the local climate and soil conditions where they naturally thrive [5,6]. However, unlike most naturally occurring communities, urban gardens typically consist of diverse mixtures of planted and volunteer species, with a substantial proportion of non-native species [1,7,8]. Invasive non-native species are now considered significant threats to global biodiversity [9], and garden plants account for > 40% of the widely recognized invasive species [10]. With the enforcement of the Nagoya Protocol, it is increasingly urgent to reinforce biological sovereignty and reduce dependence on foreign biological resources as well as the associated royalty costs for imported ornamental plants. Among 4,000 native plant species in the Republic of Korea, approximately 600 have notable developmental potential, and very few are currently utilized [11]. Despite the emphasis on biological sovereignty, the expansion of arboretums and gardens in urban areas faces challenges. For example, the utilization of native flora remains limited owing to insufficient information regarding their adaptability to local environments.
Ornamental grasses have become increasingly popular as key landscaping elements, as modern gardens are specifically tasked with addressing climate change, biodiversity loss, and urban ecosystem services. In the Republic of Korea, interest in ornamental grasses is rapidly increasing along with a growing variety of available species [12]. These grasses play a crucial role in landscaping as they are aesthetically appealing and offer significant ecosystem services. They enhance soil health through their extensive root systems, which improve soil structure and prevent erosion [13]. Notably, many ornamental grass varieties used in other countries originated in East Asia before being introduced to Europe and North America, where selective breeding occurred before their return to the Republic of Korea [14]. However, imported grasses often face challenges adapting to unique Korean climatic conditions. Additionally, domestic native species are underutilized, largely owing to insufficient information regarding their habitat preferences and ecological characteristics [15,16]. The Korean native plant Melica grandiflora is a perennial grass that is native to the temperate regions of Asia and is primarily found in mountainous meadows and at the edges of forests [17,18]. Carex forficula is a perennial herbaceous plant that is primarily native to wetlands and lowland riparian areas in East Asia [19]. Similarly, Carex boottiana, which is found in comparable habitats, is tolerant to various soil conditions and plays a significant role in nutrient cycling. These three native grass species play crucial roles in preventing soil erosion and maintaining the health of the ecosystem [20].
Abiotic stressors, such as drought, salinity, and cold, are significant environmental factors that negatively affect plant growth, survival, and performance [21,22,23]. Remarkably, rapid changes in global climate and various projections indicate increases in aridity and salinity in semiarid regions in the near future [24]. As water evaporates from the soil, its salt concentration increases, thereby leading to drought and salinity [15]. Although plant tolerance to drought or salt has been reported, garden plants have received relatively little attention in this regard [23]. Drought stress leads to plant dehydration, stomatal closure, and restricted gas exchange, subsequently inhibiting metabolism and photosynthetic rates and ultimately resulting in plant death. However, the capacity of plants to endure stressful environments depends on the species, phase of growth, and duration and intensity of water deficit [25,26]. In the initial phase of salinity stress, plants undergo water stress, which restricts leaf expansion. The osmotic impact of salinity stress can inhibit cell development and division and cause stomatal closure [27,28]. Under prolonged salinity conditions, plants undergo ionic stress, which may result in premature senescence of mature leaves, thereby reducing the photosynthetic area required for continued growth [29]. Similarly, salt stress affects various crucial processes, such as growth and development, carbon fixation, and protein synthesis, leading to growth inhibition. However, lethal concentrations, growth reduction rates, and salt tolerance levels vary among plant species [30].
Identifying drought or salinity tolerance in plants is essential for promoting the development of urban gardens and conserving biodiversity through the use of native species. Therefore, the present study investigated the selection of outdoor garden plant species by studying the adaptability of three Korean native grasses to various drought or salinity stress conditions for providing basic information, such as appropriate irrigation intervals and lethal salt concentrations in soil, to assess whether these three native grasses are suitable as garden plants in outdoor spaces.
2. Materials and Methods
2.1. Plant Materials and Experimental Design for Drought and Salinity Stress
The experiments were conducted between June and August 2023 in the Sejong National Arboretum greenhouse in the Republic of Korea. Three native grasses, M. grandiflora Koidz. (Poaceae), C. forficula Franch. & Sav, and C. boottiana hooks. & Arn. (Cyperaceae), with high ornamental values and excellent potential for use in gardens were included in this study [12] (Figure 1). The grasses were planted in 19 cm Pixel Garden plastic pots filled with eden soil (a mixture of peat moss, cocopeat, vermiculite, zeolite, and pearlite; pH 5.5–6.5, Electrical Conductivity (EC) < 1 ds/m). Air temperature and humidity in the greenhouse were monitored for 30 days using a data logger (Spectrum Watchdog 1650 Micro Station Data Logger; Spectrum Technologies, Inc, USA). Environmental conditions were affected by weather conditions; however, temperature was maintained between 15–28 °C (Figure S1). Plants were watered daily to field moisture capacity for two weeks prior to starting the treatments. Plants were divided into five replicates in each group and subjected to five treatments, each of drought and salinity stresses, for 30 days (June–July 2023). Drought stress was applied using an automatic irrigation system (self-made), and the irrigation cycle was as follows: well-watered condition (WW, once every 72 h), moderate drought stress (MD, once every 336 h), and severe drought stress (SD, no irrigation). Drought-treated plants were irrigated with 500 mL water in all treatments by measuring the water content in the pot. In general, glycophytes die at salinity (NaCl) concentrations of 100–200 mM, whereas halophytes survive at salinity concentrations of 300–500 mM [30,31,32]. Salinity stress was induced using NaCl as follows: untreated condition (UT), moderate salinity stress (MS, 250 mM NaCl), and severe salinity stress (SS, 500 mM NaCl). Salinity-treated plants were irrigated with 500 mL saline or distilled water at every 3-day interval (Table 1).
2.2. Photosynthetic Parameters
Photosynthetic measurements were conducted on fully expanded leaves from 9:00 am to 4:00 pm using an LI-6800 portable photosynthesis system (LICOR-6800; Li-Cor Inc., USA). The CO2 concentration was maintained at 400 μmol·mol−1, with a flow rate of 600 μmol·s−1. The cuvette was maintained at a temperature and relative humidity range of 25 °C and 50–60%, respectively. Light was supplied by red-blue LEDs mounted on the LI-6800 cuvettes, providing a constant artificial irradiation of 1,000 μmol·m−2·s−1. The same leaves were used for measurements of various photosynthetic parameters, such as net photosynthetic rate (Pn), stomatal conductance (gs), and transpiration rate (Tr); leaf-level instantaneous water use efficiency (WUE, µmol CO2 per mmol H2O) was calculated as the ratio of photosynthetic rate (Pn) to transpiration rate (Tr), as described in a previous study [33].
2.3. Measurements of Chlorophyll Parameters
After the experiment, leaf samples weighing 0.1 g were collected. The leaves were submerged in 10 mL glass vials containing 80% acetone and stored in the dark at 4 °C for 7 d to ensure complete pigment extraction. The chlorophyll contents (Chla, Chlb, ChlT, carotenoid, Chla/b, and Car/ChlT) were calculated using equations according to a previous study [34]. Absorbance values at wavelengths of 663, 645, and 470 nm were then measured with a spectrophotometer (Multiskan Skyhigh, Thermo Fisher Scientific, USA).
2.4. Determination of Malondialdehyde Content for Estimating Membrane Damage
Following the experiment, 0.1 g of fresh leaf tissue was homogenized in 0.5 mL of 0.2% (w/v) trichloroacetic acid (TCA) and centrifuged at 1,500 × g for 10 min at 4 °C. The resulting supernatant was then combined with a 1.5-mL reaction mixture containing 0.5% thiobarbituric acid (TBA) in 20% TCA. The mixture was incubated at 95 °C for 20 min and subsequently cooled on ice. Absorbance was measured at 532 nm and 600 nm using a microplate reader (Multiskan Skyhigh, Thermo Fisher Scientific). The malondialdehyde (MDA) content was calculated using an extinction coefficient of ε = 155 mM-1·cm-1, as described previously [35].
2.5. Determination of O2.- and H2O2 Concentration
O2- content was measured according to a modified version of a previously described method [36]. The concentration of O2- was quantified based on its ability to reduce nitro blue tetrazolium (NBT). Leaf samples (0.1 g) were homogenized in 1.5 mL of 10 mM potassium phosphate buffer (pH 7.8) containing 0.05% NBT and 10 mM NaN3. After a 20-min incubation at room temperature (20–25 °C), a 1-mL aliquot of the reaction mixture was heated in a water bath at 85 °C for 15 min and then cooled rapidly. The O2- content was expressed as the absorbance at 580 nm per gram of fresh weight (A580·g-1 FW). The H2O2 content was determined as described previously [37]. Leaf samples (0.1 g), frozen in liquid nitrogen, were extracted with 1 mL of 0.1% TCA and centrifuged at 12,000 × g for 15 min. The resulting supernatant (0.5 mL) was mixed with 0.5 mL of 100 mM potassium phosphate buffer (pH 7.8) and 1 mL of 1 M KI. After incubating the mixture in the dark for 1 h, the absorbance at 390 nm was measured using a microplate reader (Multiskan Skyhigh, Thermo Fisher Scientific, USA), and the H2O2 content was calculated from a standard curve.
2.6. Statistical Analysis
Statistical analyses were conducted using IBM SPSS Statistics 26 (IBM Corp., Armonk, NY, USA), with statistical significance determined at p < 0.05. To assess differences among groups, a one-way Analysis of Variance (ANOVA) was employed, followed by Tukey's HSD test for chlorophyll and biochemical parameters and Scheffe’s test for photosynthetic parameters, accounting for the varying number of repetitions. An independent t-test was used to examine significant differences in photosynthetic parameters between M. grandiflora and C. forficula under different treatments, including well-watered (WW), moderate drought (MD), untreated (UT), and mild salinity (MS). Additionally, principal component analysis (PCA) was performed to identify key factors influencing drought and salinity stress and to further understand plant tolerance to the conditions. The data are expressed as mean ± standard deviation.
3. Results
3.1. Changes in Growth and Visible Injuries
The average temperature during drought and salinity treatment was 25.8 ℃. The relative humidity was high (78%) owing to continuous rainfall during the experimental period (Figure S1). Differences in growth changes and visible injuries were noticeable depending on drought or salinity treatments in the three native grasses (Figure 2). For C. boottiana, the structure of the canopy, by altering its length and width, showed little change under SD compared to that under WW. No visible necrotic areas were noticed on the leaves. In contrast, the other two species exhibited reduced growth rates and bronzing of >90% leaves under SD. In particular, M. grandiflora was the first to show noticeable leaf necrosis and browning under SD and died 24 days after the start of the experiment (DAE, data not shown). Under salt stress, there was a marked reduction in growth and visible damage (necrosis) in the two species, with the exception of C. boottiana. However, visible foliar damage to M. grandiflora was more pronounced under drought stress than under salinity stress. Conversely, C. forficula exhibited more visible foliar damage caused by salinity stress than by drought stress.
3.2. Photosynthetic Parameters
The Pn of all three native grass species significantly decreased with increased drought stress. In particular, C. boottiana showed the largest decline (9.1-fold) under SD compared to that under WW (Figure 3a). The photosynthetic efficiencies of M. grandiflora and C. forficula could not be measured owing to withering and necrosis of leaves under SD. The gs and Tr indicated similar trends in the three native grasses (Figure 3b,c). The gs and Tr of M. grandiflora did not significantly differ under WW and MD treatments. C. forficula and C. boottiana showed decreases in gs and Tr with increased drought stress (MD and SD). WUE differed depending on the species and drought treatment. WUE of M. grandiflora decreased more under MD than under WW; however, those of the other two species did not show significant differences. C. boottiana had a higher WUE under SD than under WW and MD; however, the difference was not significant (Figure 3d).
The photosynthetic efficiencies of M. grandiflora and C. forficula could not be measured owing to withering and necrosis of leaves under SS. The Pn of the three species significantly decreased with increased salinity; however, that of M. grandiflora was not significantly different under UT and MS (Figure 4a). The gs and Tr values of M. grandiflora and C. forficula were not significantly different under UT and MS. In contrast, the gs and Tr of C. boottiana rapidly decreased under salinity stress (MS and SS) (Figure 4b,c). A significant difference in WUE was noticed among the three grasses (Figure 4d). M. grandiflora had a lower WUE under MS than under UT; however, the difference was not significant. C. forficula and C. boottiana showed significant increases of 6.6-fold (under MS) and 3.6-fold (under SS), respectively, compared to that under UT.
3.3. Chlorophyll Parameters
The growth of plants was associated with substantial reductions in chlorophyll and carotenoid contents and Chl a/b ratios in fully expanded leaves of native grasses under abiotic stress (Table 2 and Table 3). The ChlT and carotenoid contents of M. grandiflora and C. forficula under SD were notably lower by 86.7% and 61.5 %, respectively than those under WW. In contrast, photosynthetic pigments (chlorophyll and carotenoids) of C. boottiana showed no pronounced differences under different drought conditions. In all species, the Chl a/b ratio gradually decreased as drought stress increased. Among them, C. boottiana showed the lowest decrease of 9.1%, whereas M. grandiflora showed the highest decrease of 67.6% under SD compared to that under WW. The Car/ChlT ratio was highest for all three species under SD and was more strongly influenced by carotenoids than by chlorophyll (Table 2). The ChlT and carotenoid levels gradually decreased in M. grandiflora and C. forficula with increasing salt concentrations. C. boottiana exhibited the highest ChlT and carotenoid levels among the three species under SD, and did not show pronounced differences in ChlT and carotenoid levels under different salt concentrations. The Chl a/b ratios of M. grandiflora and C. forficula significantly decreased with increasing salt concentrations. The Car/ChlT ratios of M. grandiflora and C. boottiana under salt treatment had stronger effects on carotenoids than on chlorophyll and were approximately 28% and 4.4% higher, respectively, under SS than under UT. In contrast, C. forficula did not show a defined trend under those treatments (Table 3).
3.4. MDA and Reactive Oxygen Species (O2.-, H2O2)
Phytotoxicity in native grasses under abiotic stress was quantified by evaluating MDA contents, an indicator of lipid peroxide ions, in leaves. As seen in Figure 5a and Figure 6a, MDA contents of M. grandiflora and C. forficula significantly increased with increasing drought and salinity stresses. M. grandiflora showed the most rapid increase (2.7-fold) under SD compared to that under WW, and C. forficula showed the most acute increase (2.9-fold) under SS compared to that under UT. However, C. boottiana showed little difference in MDA levels under abiotic stress, which increased 2.0-fold under SD compared to that under WW; MDA levels in C. boottiana increased under MS compared to that under UT and slightly decreased under SS. O2.- and H2O2, indicators to evaluate oxidative stress in plants, also showed a similar trend of change (Figure 5b,c and Figure 6b,c). The O2.- and H2O2 contents of M. grandiflora significantly increased under drought and salinity stress by 2.1-fold and 3.9-fold, respectively. C. forficula showed highly enhanced O2.- and H2O2 contents under drought stress; however, they were slightly changed under salt stress. In addition, C. boottiana showed differences in H2O2 contents (highest levels under SD and SS) under drought and salinity stresses. However, O2.- levels in C. boottiana under drought and salinity stresses were not significantly different.
3.5. Principal Component Analysis
To better understand the interaction between drought or salt stress and the three native grass species, PCA was conducted (Figure 7). In the PCA, PC1 accounted for 61.6% and 46.4% of the total variance in drought and salinity stress, respectively, whereas PC2 explained 21.7% of drought stress and 25.3% of salinity stress. The biplot analysis revealed the correlations between the stress indices calculated for all traits. Under drought stress, positive correlations were observed among Tr, gs, Pn, ChlT, carotenoids, Chl a/b, and WUE, whereas negative correlations were observed among MDA, O2-, H2O2, and Car/ChlT (Figure 7a). For salinity stress, positive correlations were noted with Pn, ChlT, and carotenoids, whereas negative correlations were observed with MDA, O2-, H2O2, and Chl a/b (Figure 7b). Additionally, the scatterplot classified the grasses into tolerant or sensitive groups based on their response to drought or salinity stress (Figure 7c,d). As a result of comparing the three species in drought or salinity treatments, differences in the degree of tolerance and sensitivity to stress were distinguished clearly.
4. Discussion
Drought and salinity are two key abiotic stressors that negatively impact plant growth and development [38,39,40]. As a result, plants initiate various physiological and biochemical responses to mitigate the effects of the stresses. The responses involve alterations in morphology, photosynthesis, and distribution of harmful ionic species, in addition to biochemical adjustments, for example, in antioxidant or metabolic activities [30,41,42,43,44,45]. In the present study, all three grass species exhibited osmotic stress under drought conditions due to the accumulation of intracellular salts and ions caused by dehydration. In contrast, salinity stress triggered ionic stress, characterized by the buildup of toxic ions in the intercellular space, resulting from osmotic stress. Consequently, the grasses showed more severe morphological damage under saline conditions than under drought stress.
Photosynthesis, a critical and complex physiological process, is impacted heavily by abiotic stresses such as drought and salinity [42]. Such stresses directly inhibit photosynthesis, leading to declines in morphological and physiological traits, leaf water potential, sap flow, and stomatal conductance [38,45,46,47,48]. In the present study, the Pn values of all grasses decreased steadily as drought and salinity levels increased. Notably, C. boottiana exhibited lower gs and Tr but showed enhanced WUE under both SD and SS conditions. Typically, plants that are tolerant to drought or salt stress improve their leaf water status by reducing transpiration through decreased gs, which minimizes water loss, enhances Pn, and ultimately boosts WUE [45,49,50,51].
Reduction in chlorophyll content under abiotic stress is commonly recognized as an indicator of oxidative stress [52], which triggers pigment photooxidation and chlorophyll degradation [53]. As such, chlorophyll content serves as a key indicator of a plant's physiological state [54]. In response to drought or salinity stress, M. grandiflora and C. forficula exhibited a decline in total chlorophyll (ChlT) content. However, C. boottiana maintained the highest levels of ChlT and carotenoids under MD, with no significant changes observed under salinity stress. Carotenoids, which play a secondary role in light harvesting, assist chlorophyll in absorbing solar energy when excess light cannot be managed by chlorophyll alone. Additionally, carotenoids help prevent the formation of reactive oxygen species (ROS) [55]. In drought- or salt-tolerant plants, carotenoid levels rise under stress, indicating enhanced photoprotection [56].
MDA is formed predominantly through the peroxidation of polyunsaturated fatty acids by ROS, and it is applied extensively as an indicator for lipid peroxidation in plant cell membranes caused by oxidative stress. The phenomenon is acknowledged broadly as an indicator of stress-induced cellular damage in plants and is often associated with potassium (K+) efflux from plant cells [57,58]. Thus, MDA can signal various acute responses to oxidative stress, reflecting damage severity. In the present study, all three grass species, excluding C. boottiana under severe salt stress (SS), exhibited elevated MDA, O2-, and H2O2 levels according to drought and salinity conditions. This aligns with previous studies that have reported an overall increase in lipid peroxidation alongside a decline in total chlorophyll and carotenoid content under prolonged stress [59]. The accumulation of MDA and ROS often correlates with a reduction in photosynthetic pigments during extended periods of stress [60]. In particular, C. boottiana showed no significant changes, or only slight variations, in MDA, O2-, and H2O2 levels between non-stress treatments (WW, UT) and stress treatments (MD, MS, and SS). This suggests that C. boottiana possesses a robust capacity to mitigate oxidative stress, maintaining stable levels of oxidative markers under stress conditions [61]. This stability likely indicates an efficient antioxidant defense system or reduced susceptibility to lipid peroxidation and ROS accumulation, which could explain its stress tolerance [62]. Moreover, plant cell membranes play a crucial role in the regulation of ion homeostasis and maintaining osmotic balance, both of which are essential for minimizing the impact of salinity stress [61]. As cell membranes are central to numerous physiological and biological processes, they are often key targets of environmental stress [63].
In general, tolerant plants show no major impact on growth or appearance. They are able to maintain gas exchange and water use at high levels while keeping oxidative stress low by effectively managing osmotic balance and nutrient levels [64,65]. Conversely, sensitive plants show low growth rates and decreased gas exchange ability due to intracellular nutrient imbalance, resulting in increased oxidative stress, ultimately leading to plant aging and death [66,67,68]. In the present study, there were different responses to drought or salinity stress in the three grass species, which were divided into sensitive and tolerant, based on the results of the analyses of factor results (Figure 7). Our findings also confirmed that the three species studied demonstrated a certain level of tolerance to drought and salinity stress; however, M. grandiflora and C. forficula were sensitive to severe abiotic stress. Notably, C. boottiana showed the highest adaptability to extreme drought and salinity conditions, exhibiting reduced gs and Tr alongside increased WUE and chlorophyll pigment contents under severe stress. In addition, MDA and ROS levels were not altered significantly under salinity stress. Drought- and salt-tolerant plants adopt strategies such as reducing stomatal conductance to minimize transpiration and water loss, thereby maintaining positive water balance under stress conditions. This adjustment also leads to improved photosynthesis rates (Pn) and overall WUE in tolerant plants [69,70,71]. Additionally, such plants activate their antioxidant defense systems to counteract excessive ROS production that results from drought and salinity stress. The balance between ROS generation and detoxification is crucial for preventing oxidative damage to cellular structures, such as proteins and lipids [45,72,73,74].
5. Conclusions
Increased tolerance to drought or salinity stress is a major aim in the genetic improvement of plants of environmental, horticultural, agricultural, and economic importance. This study highlights the importance of selecting native plant species with high adaptability to abiotic stresses for urban landscaping. C. boottiana has emerged as a promising candidate owing to its robust performance under adverse conditions, offering a sustainable solution for enhancing urban biodiversity and ecosystem services. These findings advocate for increased utilization of native species to minimize reliance on non-native plants, thereby supporting the goals of global biodiversity conservation. However, to better understand the adaptation mechanisms of C. boottiana to drought or salinity stress, further studies involving biochemical and ultra-morphological analyses are required.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Change in air temperature and relative humidity in the greenhouse during a month (June~July) of the study period. Data are represented as the daily average.
Author Contributions
Conceptualization, S.Y.W.; methodology, S.G.B.; validation, S.Y.W. and S.G.B.; formal analysis, S.G.B. and J.W.S.; investigation, S.G.B. and J.W.S.; resources, J.M.S., J.W.S. and S.G.B.; data curation, S.G.B. and J.I.N; writing—original draft preparation, S.G.B.; writing—review and editing, S.G.B.; visualization, S.G.B.; supervision, S.Y.W.; project administration, S.Y.W.; funding acquisition, J.M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Garden Business Fund of the KOREA FOREST SERVICE.
Data Availability Statement
The data presented in this study are available on request from the corresponding author, due to privacy or ethical reasons.
Acknowledgments
We would like to thank the Sejong National Arboretum under the Korea Forest Service for providing plant material and equipment support for field experiments.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the study design; collection, analyses, and interpretation of data; writing of the manuscript; or decision to publish the results.
References
- Wang, D.; Xu, P.Y.; An, B.W.; Guo, Q.P. Urban green infrastructure: Bridging biodiversity conservation and sustainable urban development through adaptive management approach. Front. Ecol. Evol. 2024, 12, 1440477. [Google Scholar] [CrossRef]
- Owen, J. The Ecology of a Garden: the First Fifteen Years; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Miotk, P. The naturalized garden – A refuge for animals? – first results. Zool. Anz. 1996, 235, 101–116. [Google Scholar]
- Mason, C.F. Thrushes now largely restricted to the built environment in eastern England. Divers. Distrib. 2000, 6, 189–194. [Google Scholar] [CrossRef]
- Bigirimana, J.; Bogaert, J.; De Cannière, C.; Bigendako, M.J.; Parmentier, I. Domestic garden plant diversity in Bujumbura, Burundi: role of the socio-economical status of the neighborhood and alien species invasion risk. Landsc. Urban Plan. 2012, 107, 118–126. [Google Scholar] [CrossRef]
- Available online: https://www.fs.usda.gov/wildflowers/Native_Plant_Materials/Native_Gardening/index.shtml.
- Thompson, K.; Austin, K.C.; Smith, R.M.; Warren, P.H.; Angold, P.G.; Gaston, K.J. Urban domestic gardens (I): putting small-scale plant diversity in context. J. Vegetation Sci. 2003, 14, 71–78. [Google Scholar] [CrossRef]
- Seitz, B.; Buchholz, S.; Kowarik, I.; Herrmann, J.; Neuerburg, L.; Wendler, J.; Wendler, J.; Egerer, M. Land sharing between cultivated and wild plants: urban gardens as hotspots for plant diversity in cities. Urban Ecosyst. 2022, 25, 927–939. [Google Scholar] [CrossRef]
- IUCN publishing division, Cambridge, UK and Gland, Switzerland. IUCN 2000. The world conservation union, gland, switzerland.
- Smith, R.M.; Thompson, K.; Hodgson, J.G.; Warren, P.H.; Gaston, K.J. Urban domestic gardens (IX): composition and richness of the vascular plant flora, and implications for native biodiversity. Biol. Conserv. 2006, 129, 312–322. [Google Scholar] [CrossRef]
- Choi, W.K.; Moon, A.R.; Song, J.H.; Kim, Y.J.; Song, Y.J.; Jeong, J.R.; Jin, H.Y. A study on revitalization strategy of wild-flower industry through status of wildflower farm investigation. Korean. J. Hortic. Sci. 2016, 34, 232–232. [Google Scholar]
- Kim, J. The current state and characteristics of ornamental grasses in South Korea. J. Korean Inst. Landsc. Archit. 2021, 49, 151–162. [Google Scholar] [CrossRef]
- Brown, R.; Green, L. Soil health and the impact of plant root systems. Environ. Sci. [Lett.] 2019, 8, 89–97. [Google Scholar]
- Darke, R. The Encyclopedia of Grasses for Livable Landscapes; Timber Press: Portland, OR, USA, 2007. [Google Scholar]
- Kim, J.W. Korean Plant Ecology Treasure Ⅱ; Seoul, 2016. Eco and nature Publishing.
- Song, K.H.; Kim, Y.J.; Kim, C.G.; Won, C.O.; Lee, J.K. A-Z Korean Garden Plants; Designpost: Goyang, 2018. [Google Scholar]
- Choi, Y.; Kim, H. The ecology of Melica grandiflora in mountainous regions. J. Plant Ecol. 2010, 34, 123–130. [Google Scholar]
- Lee, J.S. Flora of Korea: Grasses Korean National Arboretum, 2015.
- Thompson, R.; Green, P. Sedges in wetland ecosystems: A vital role. J. Wetland Ecol. 2020, 24, 134–142. [Google Scholar]
- Carter, S. The importance of Carex species in biodiversity conservation. Plant Ecol. Rev. 2021, 18, 90–101. [Google Scholar]
- Cregg, B.M. Improving drought tolerance of trees: theoretical and practical considerations. In. Acta Hortic. Acta Hortic. International horticultural congress: Nursery crops; development, evaluation, production and use 630 2004, XXVI, 147–158. [CrossRef]
- Fernàndez, J.A.; Balenzategui, L.; Bañón, S.; Franco, J.A. Induction of drought tolerance by paclobutrazol and irrigation deficit in Phillyrea angustifolia during the nursery period. Scientia Horticulturae 2006, 107, 277–283. [Google Scholar] [CrossRef]
- Rafi, Z.N.; Kazemi, F.; Tehranifar, A. Morpho-physiological and biochemical responses of four ornamental herbaceous species to water stress. Acta Physiol. Plant. 2019, 41. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Lakshmanan, G.M.A.; Gomathinayagam, M.; Panneerselvam, R. Alterations in morphological parameters and photosynthetic pigment responses of Catharanthus roseus under soil water deficits. Colloids Surf. B Biointerfaces 2008, 61, 298–303. [Google Scholar] [CrossRef]
- Tátrai, Z.A.; Sanoubar, R.; Pluhár, Z.; Mancarella, S.; Orsini, F.; Gianquinto, G. Morphological and physiological plant responses to drought stress in Thymus citriodorus. Int. J. Agro 2016, 2016, 1–8. [Google Scholar] [CrossRef]
- Flowers, T.J. Improving crop salt tolerance. J. Exp. Bot. 2004, 55, 307–319. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Cramer, G.R.; Nowak, R.S. Supplemental manganese improves the relative growth, net assimilation and photosynthetic rates of salt-stressed barley. Physiol. Plant. 1992, 84, 600–605. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: a review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef]
- Baek, S.G.; Woo, S.Y. Physiological and biochemical responses of two tree species in urban areas to different air pollution levels. Photosynthetica 2010, 48, 23–29. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Mai, V.C.; Bednarski, W.; Borowiak-Sobkowiak, B.; Wilkaniec, B.; Samardakiewicz, S.; Morkunas, I. Oxidative stress in pea seedling leaves in response to Acyrthosiphon pisum infestation. Phytochemistry 2013, 93, 49–62. [Google Scholar] [CrossRef]
- Tahjib-ul-Arif, Md.; Roy, P.R.; Al Mamun Sohag, A.A.M.; Afrin, S.; Rady, M.M.; Hossain, M.A. Exogenous calcium supplementation improves salinity tolerance in BRRI dhan28; a salt-susceptible high-yielding Oryza sativa cultivar. J. Crop Sci. Biotechnol. 2018, 21, 383–394. [Google Scholar] [CrossRef]
- Billah, M.; Aktar, S.; Brestic, M.; Zivcak, M.; Khaldun, A.B.M.; Uddin, M.S.; Bagum, S.A.; Yang, X.; Skalicky, M.; Mehari, T.G.; et al. Progressive genomic approaches to explore drought- and salt-induced oxidative stress responses in plants under changing climate. Plants (Basel) 2021, 10, 1910. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumar, V.; Chauhan, A.; Datta, S.; Wani, A.B.; Singh, N.; Singh, J. Toxicity, degradation and analysis of the herbicide atrazine. Environ. Chem. Lett. 2018, 16, 211–237. [Google Scholar] [CrossRef]
- Hernández, J.A.; Ferrer, M.A.; Jiménez, A.; Barceló, A.R.; Sevilla, F. Antioxidant systems and O2/H2O2 production in the apoplast of Pisum sativum L. leaves: its relation with NaCl-induced necrotic lesions in minor veins. Plant Physiol. 2001, 127, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: an overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Díaz-Vivancos, P.; Álvarez, S.; Fernández-García, N.; Sánchez-Blanco, M.J.; Hernández, J.A. NaCl-induced physiological and biochemical adaptative mechanisms in the ornamental Myrtus communis L. plants. J. Plant Physiol. 2015, 183, 41–51. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Díaz-Vivancos, P.; Álvarez, S.; Fernández-García, N.; Sánchez-Blanco, M.J.; Hernández, J.A. Physiological and biochemical mechanisms of the ornamental Eugenia myrtifolia L. plants for coping with NaCl stress and recovery. Planta 2015, 242, 829–846. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy. 2017, 7, 18. [CrossRef]
- Li, L.; Gu, W.; Li, J.; Li, C.; Xie, T.; Qu, D.; Meng, Y.; Li, C.; Wei, S. Exogenously applied spermidine alleviates photosynthetic inhibition under drought stress in maize (Zea mays L.) seedlings associated with changes in endogenous polyamines and phytohormones. Plant Physiol. Biochem. 2018, 129, 35–55. [Google Scholar] [CrossRef]
- Wang, X.-M.; Wang, X.-K.; Su, Y.-B.; Zhang, H.-X. Land pavement depresses photosynthesis in urban trees especially under drought stress. Sci. Total Environ. 2019, 653, 120–130. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.-G.; Yoon, T.-M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Scientia Horticulturae. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Moutinho-Pereira, J.M.; Gonçalves, B.C.; Ferreira, H.F.; Correia, C.M. Changes in growth, gas exchange, xylem hydraulic properties and water use efficiency of three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2007a, 60, 183–192. [Google Scholar] [CrossRef]
- Moutinho-Pereira, J.M.; Magalhães, N.; Gonçalves, B.; Bacelar, E.; Brito, M.; Correia, C. Gas exchange and water relations of three Vitis vinifera L. cultivars growing under Mediterranean climate. Photosynthetica 2007, 45, 202–207. [Google Scholar] [CrossRef]
- Rivero, R.M.; Kojima, M.; Gepstein, A.; Sakakibara, H.; Mittler, R.; Gepstein, S.; Blumwald, E. Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc. Natl Acad. Sci. U. S. A. 2007, 104, 19631–19636. [Google Scholar] [CrossRef] [PubMed]
- Faisal, S.; Mujtaba, S.M.; Asma, M.; Mahboob, W. Polyethylene glycol mediated osmotic stress impacts on growth and biochemical aspects of wheat (Triticum aestivum L.). J. Crop Sci. Biotechnol. 2019, 22, 213–223. [Google Scholar] [CrossRef]
- Nezhadahmadi, A.; Prodhan, Z.H.; Faruq, G. Drought tolerance in wheat. ScientificWorldJournal 2013, 2013, 610721. [Google Scholar] [CrossRef]
- Marques, M.C.; Nascimento, C.W.A.; da Silva, A.J.; da Silva Gouveia-Neto, A. Tolerance of an energy crop (Jatropha curcas L.) to zinc and lead assessed by chlorophyll fluorescence and enzyme activity. S. Afr. J. Bot. 2017, 112, 275–282. [Google Scholar] [CrossRef]
- Knox, J.P.; Dodge, A.D. Singlet oxygen and plants. Phytochemistry 1985, 24, 889–896. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. Changes in carotenoids, tocopherols and diterpenes during drought and recovery, and the biological significance of chlorophyll loss in Rosmarinus officinalis plants. Planta 2000, 210, 925–931. [Google Scholar] [CrossRef]
- Campos, P.S.; Quartin, V.; Ramalho, J.C.; Nunes, M.A. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of Coffea sp. plants. J. Plant Physiol. 2003, 160, 283–292. [Google Scholar] [CrossRef]
- Demidchik, V.; Straltsova, D.; Medvedev, S.S.; Pozhvanov, G.A.; Sokolik, A.; Yurin, V. Stress-induced electrolyte leakage: the role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 2014, 65, 1259–1270. [Google Scholar] [CrossRef]
- Sairam, R.K.; Shukla, D.S.; Saxena, D.C. Stress induced injury and antioxidant enzymes in relation to drought tolerance in wheat genotypes. Biologia Plant. 1997, 40, 357–364. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, B. Drought and heat stress injury to two cool-season turfgrasses in relation to antioxidant metabolism and lipid peroxidation. Crop Sci. 2001, 41, 436–442. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Leshem, Y.Y. Plant Membranes: A Biophysical Approach to Structure, Development and Senescence; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1992; p. 266. [Google Scholar]
- He, R.; Liu, Y.; Song, C.; Feng, G.; Song, J. Osmotic regulation beyond nitrate nutrients in plant resistance to stress: a review. Plant Growth Regul. 2024, 103, 1–8. [Google Scholar] [CrossRef]
- Choudhary, S.; Wani, K.I.; Naeem, M.; Khan, M.M.A.; Aftab, T. Cellular responses, osmotic adjustments, and role of osmolytes in providing salt stress resilience in higher plants: polyamines and nitric oxide crosstalk. J. Plant Growth Regul. 2023, 42, 539–553. [Google Scholar] [CrossRef]
- Tewari, R.K.; Yadav, N.; Gupta, R.; Kumar, P. Oxidative stress under macronutrient deficiency in plants. J. Soil Sci. Plant Nutr. 2021, 21, 832–859. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Plant oxidative stress: biology, physiology and mitigation. Plants (Basel) 2022, 11, 1185. [Google Scholar] [CrossRef]
- Da Cruz Ferreira, R.L.; de Mello Prado, R.; de Souza Junior, J.P.; Gratão, P.L.; Tezotto, T.; Cruz, F.J.R. Oxidative stress, nutritional disorders, and gas exchange in lettuce plants subjected to two selenium sources. J. Soil Sci. Plant Nutr. 2020, 20, 1215–1228. [Google Scholar] [CrossRef]
- Koyro, H.W. Effect of salinity on growth, photosynthesis, water relations and solute composition of the potential cash crop halophyte Plantago coronopus (L.). Environ. Exp. Bot. 2006, 56, 136–146. [Google Scholar] [CrossRef]
- Stepien, P.; Johnson, G.N. Contrasting responses of photosynthesis to salt stress in the glycophyte Arabidopsis and the halophyte Thellungiella: role of the plastid terminal oxidase as an alternative electron sink. Plant Physiol. 2009, 149, 1154–1165. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Mu, X.; Shao, H.; Wang, H.; Brestic, M. Global plant responding mechanisms to salt stress: Physiological and Molecular Levels and Implications in Biotechnology. Crit. Rev. Biotechnol. 2015, 35, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Genes and salt tolerance: bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Armando Massange-Sánchez, J.; Vanessa Sánchez-Hernández, C.; Mireya Hernández-Herrera, R.; Andrea Palmeros-Suárez, P. The Biochemical mechanisms of salt tolerance in plants. In Plant Stress Physiol. Perspect. Agric, 2021. [CrossRef]
- Iqbal, M.J. Role of osmolytes and antioxidant enzymes for drought tolerance in wheat. In Global Wheat Production, Shah, F., Abdul, B., Muhammad, A., Eds.; IntechOpen, 2021.
Figure 1.
List of Korean collection sites for the three native species.

Figure 2.
Images showing changes in growth and visible injuries of the three native grasses following 30 days of drought [well-watered condition (WW), irrigation once every 72 h; moderate drought stress (MD), irrigation once every 336 h; severe drought stress (SD), no irrigation for 30 days] and salinity [UT, untreated condition; moderate salinity stress (MS), 250 mM NaCl; severe salinity stress (SS), 500 mM NaCl] stresses.
Figure 2.
Images showing changes in growth and visible injuries of the three native grasses following 30 days of drought [well-watered condition (WW), irrigation once every 72 h; moderate drought stress (MD), irrigation once every 336 h; severe drought stress (SD), no irrigation for 30 days] and salinity [UT, untreated condition; moderate salinity stress (MS), 250 mM NaCl; severe salinity stress (SS), 500 mM NaCl] stresses.

Figure 3.
(a) Photosynthetic rate (Pn), (b) stomatal conductance (gs), (c) transpiration rate (Tr), and (d) water use efficiency (WUE) of the three native grasses under drought. The values for C. boottiana were analyzed by Scheffe’s test (as lowercase letters), and those for M. grandiflora and C. forficula were analyzed using t-test (as asterisks) for significant differences under drought treatments (∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001; ns, not significant). Bars in each graph indicate mean ± standard deviation (n = 5). There is no significant difference between bars marked with the same letters (p > 0.05).
Figure 3.
(a) Photosynthetic rate (Pn), (b) stomatal conductance (gs), (c) transpiration rate (Tr), and (d) water use efficiency (WUE) of the three native grasses under drought. The values for C. boottiana were analyzed by Scheffe’s test (as lowercase letters), and those for M. grandiflora and C. forficula were analyzed using t-test (as asterisks) for significant differences under drought treatments (∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001; ns, not significant). Bars in each graph indicate mean ± standard deviation (n = 5). There is no significant difference between bars marked with the same letters (p > 0.05).
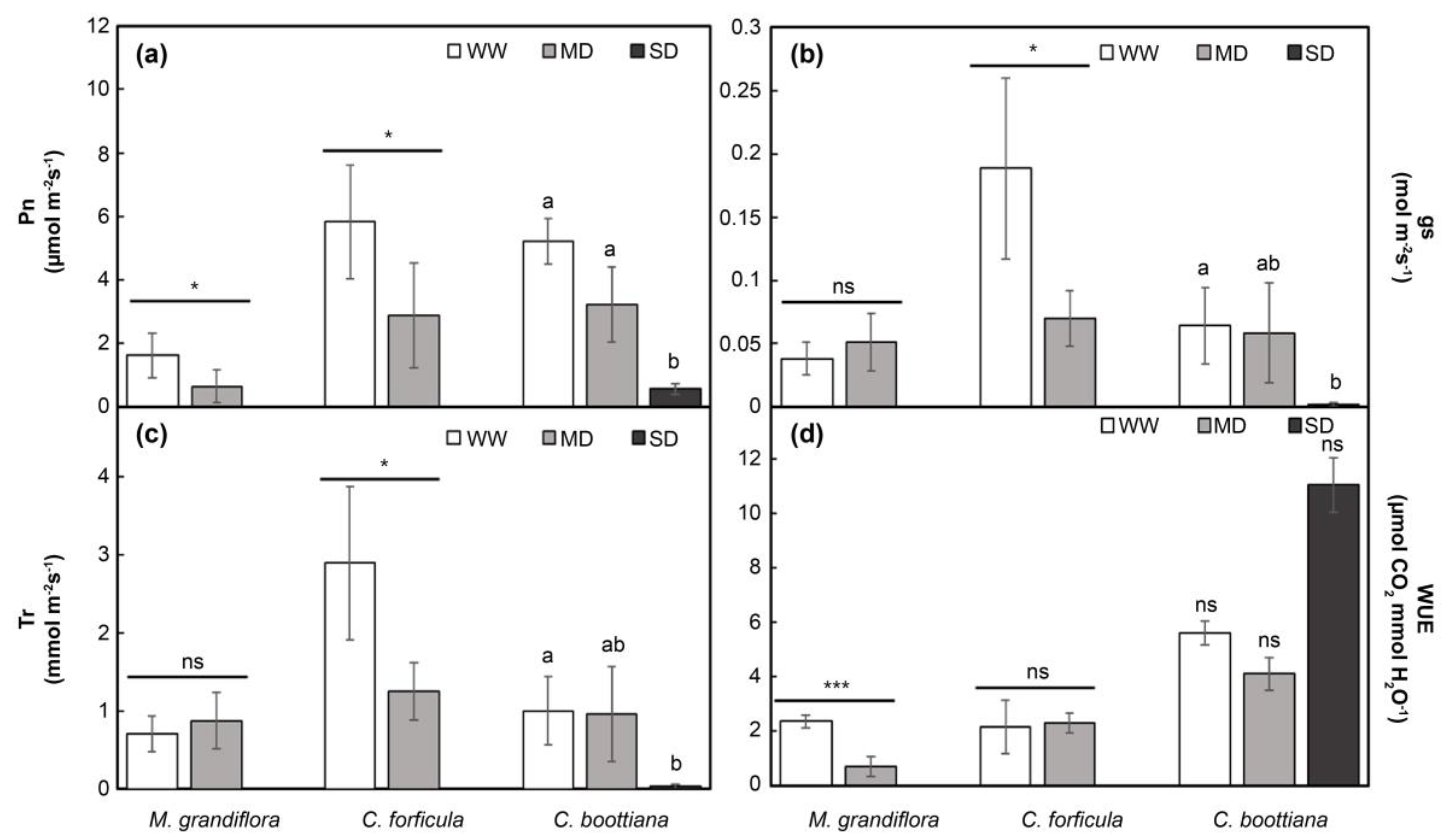
Figure 4.
(a) Pn, (b) gs, (c) Tr, and (d) WUE in three native grasses under salinity stress. The data of C. boottiana were analyzed by Scheffe’s test (as lowercase letters), and those of M. grandiflora and C. forficula were analyzed using t-test (as asterisks) to determine significant differences under different salinity treatments (∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001; ns, not significant). Bars in each graphic indicate mean ± standard deviation (n = 5). There is no significant difference between bars marked with the same letters (p > 0.05).
Figure 4.
(a) Pn, (b) gs, (c) Tr, and (d) WUE in three native grasses under salinity stress. The data of C. boottiana were analyzed by Scheffe’s test (as lowercase letters), and those of M. grandiflora and C. forficula were analyzed using t-test (as asterisks) to determine significant differences under different salinity treatments (∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001; ns, not significant). Bars in each graphic indicate mean ± standard deviation (n = 5). There is no significant difference between bars marked with the same letters (p > 0.05).
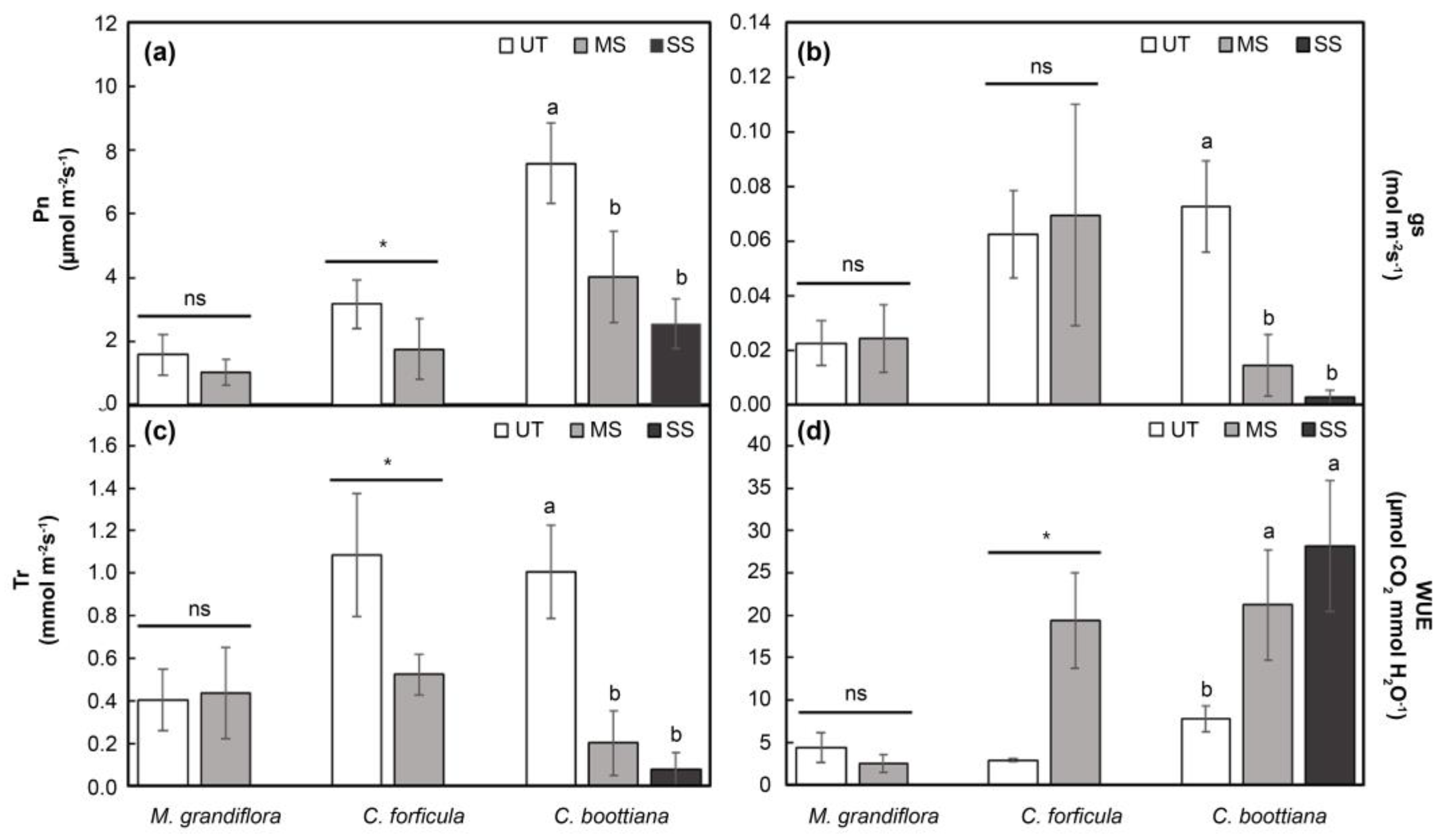
Figure 5.
(a) Malondialdehyde (MDA), (b) O2.-, and (c) H2O2 contents in three native grasses under drought stress. Significant differences, based on Tukey's HSD test, are denoted by lowercase letters. The data are presented as mean ± standard deviation (n = 4). Bars marked with the same letters are not significantly different (p < 0.05).
Figure 5.
(a) Malondialdehyde (MDA), (b) O2.-, and (c) H2O2 contents in three native grasses under drought stress. Significant differences, based on Tukey's HSD test, are denoted by lowercase letters. The data are presented as mean ± standard deviation (n = 4). Bars marked with the same letters are not significantly different (p < 0.05).
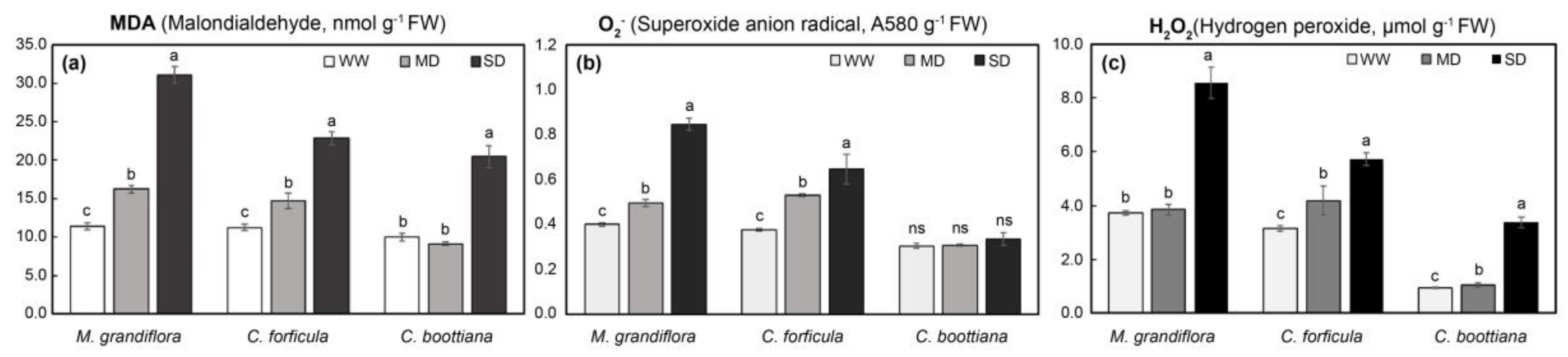
Figure 6.
(a) MDA, (b) O2.-, and (c) H2O2 contents in three native grasses under salinity stress. Significant differences, based on Tukey's HSD test, are denoted by lowercase letters. The data are presented as mean ± standard deviation (n = 4). Bars marked with the same letters are not significantly different (p < 0.05).
Figure 6.
(a) MDA, (b) O2.-, and (c) H2O2 contents in three native grasses under salinity stress. Significant differences, based on Tukey's HSD test, are denoted by lowercase letters. The data are presented as mean ± standard deviation (n = 4). Bars marked with the same letters are not significantly different (p < 0.05).
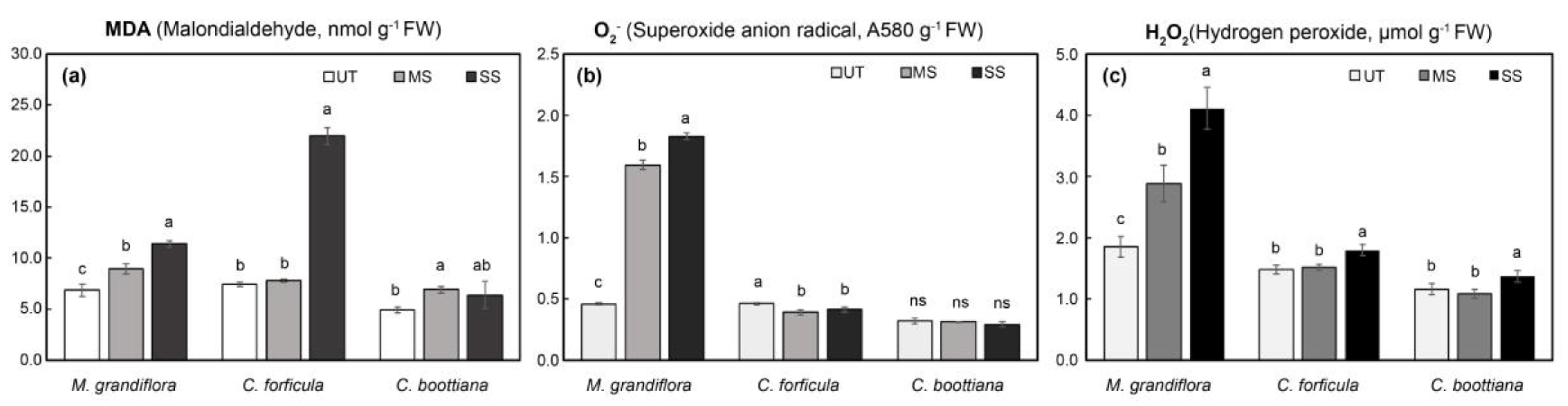
Figure 7.
Principal component analysis (PCA) of physiochemical parameters in three native grasses with distinct degrees of tolerance to drought (a, c) and salinity (b, d) stresses. (a, b) PCA-associated loading plots. (c, d) PCA-associated scatter plots.
Figure 7.
Principal component analysis (PCA) of physiochemical parameters in three native grasses with distinct degrees of tolerance to drought (a, c) and salinity (b, d) stresses. (a, b) PCA-associated loading plots. (c, d) PCA-associated scatter plots.
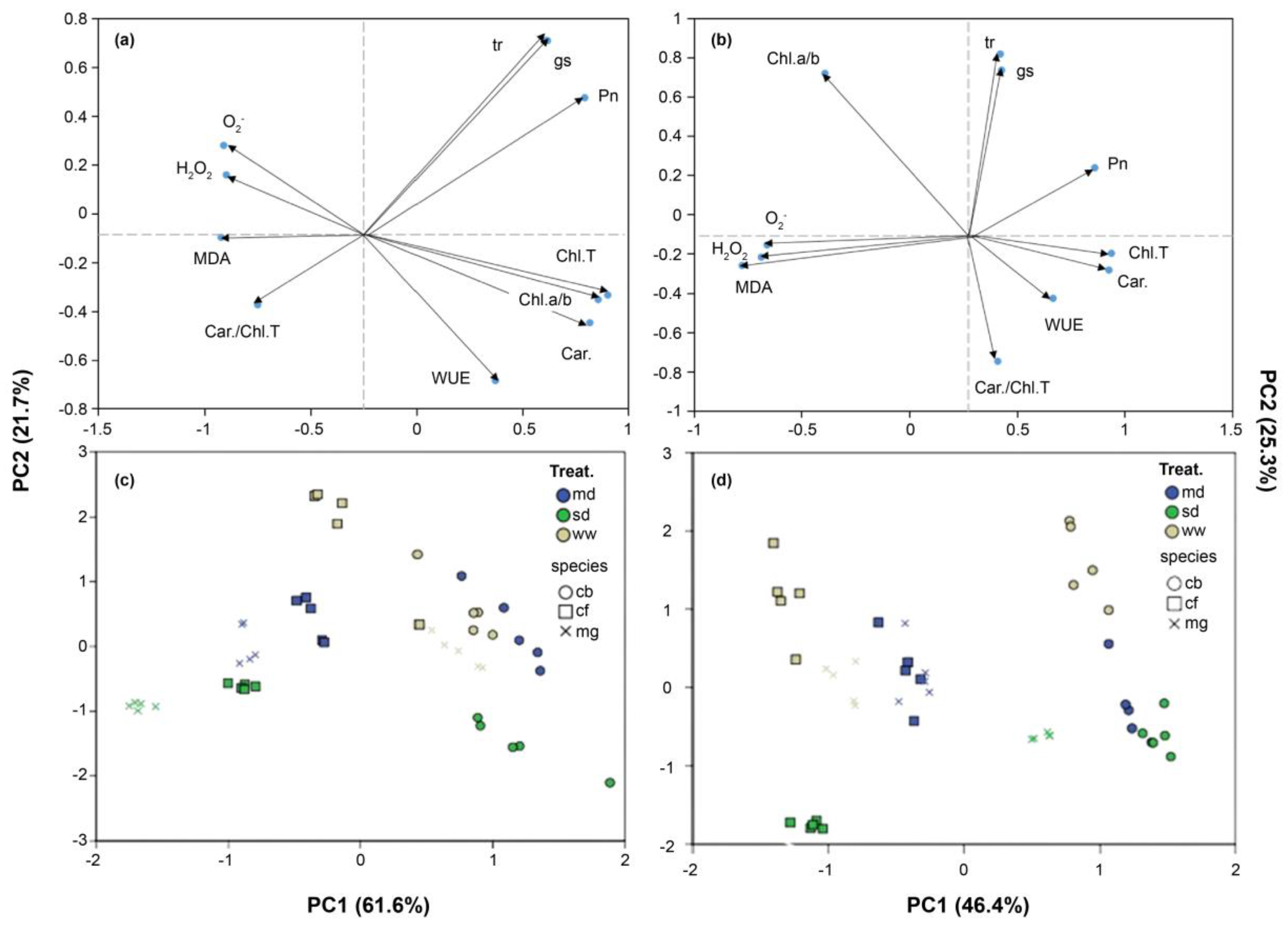

Table 1.
Experimental design for the three treatments of drought and salinity stresses.
| Experimental design | |||
|---|---|---|---|
| Drought stress | Salinity stress | ||
| Well-watered (irrigation once every 72 h) |
WW | Untreated (no NaCl treatment) |
UT |
| Moderate drought treated (irrigation once every 336 h) |
MD | Moderate salinity-treated (250 mM NaCl) |
MS |
| Severe drought treated (no irrigation for 30 days) |
SD | Severe salinity-treated (500 mM NaCl) |
SS |
Table 2.
ChlT and carotenoid contents, and Chla/b and Car/ChlT ratios in three native grasses under drought stress.
Table 2.
ChlT and carotenoid contents, and Chla/b and Car/ChlT ratios in three native grasses under drought stress.
| Chlorophyll (mg∙g-1 FW) |
Drought level |
M. grandiflora |
C. forficula |
C. boottiana |
| ChlT | WW1 | 4.01±0.90 a | 3.20±0.28 a | 2.94±0.27 b |
| MD2 | 0.73±0.14 b | 1.85±0.16 b | 4.51±0.12 a | |
| SD3 | 0.53±0.04 b | 1.23±0.04 c | 3.22±0.24 b | |
| Carotenoid | WW | 8.13±0.84 a | 6.48±0.50 a | 5.81±0.48 c |
| MD | 1.63±0.28 b | 3.93±0.24 b | 8.85±0.19 a | |
| SD | 1.26±0.06 b | 2.70±0.10 c | 7.86±0.47 b | |
| Chl a/b ratio | WW | 2.38±0.04 a | 2.05±0.04 a | 2.52±0.07 a |
| MD | 1.02±0.11 b | 1.83±0.04 b | 2.31±0.06 b | |
| SD | 0.77±0.03 c | 1.10±0.04 c | 2.29±0.05 b | |
| Car/ChlT ratio | WW | 2.03±0.02 c | 1.58±0.88 c | 1.54±0.86 b |
| MD | 2.24±0.06 b | 1.66±0.93 b | 1.96±0.02 b | |
| SD | 2.39±0.06 a | 2.19±0.02 a | 2.44±0.05 a |
The data were analyzed using Tukey's HSD test and are expressed as mean ± standard deviation (n = 5). Means with the same letters are not significantly different (p < 0.05).
Table 3.
ChlT and carotenoid contents, and Chla/b and Car/ChlT ratios in three native grasses under salinity stress.
Table 3.
ChlT and carotenoid contents, and Chla/b and Car/ChlT ratios in three native grasses under salinity stress.
| Chlorophyll (mg∙g-1 FW) |
Salt level |
M. grandiflora |
C. forficula |
C. boottiana |
| ChlT | UT1 | 2.80±0.05 a | 3.37±0.24 a | 7.19±0.10 ns |
| MS2 | 1.42±0.09 b | 1.47±0.28 b | 7.02±0.11 ns | |
| SS3 | 0.42±0.02 c | 0.79±0.04 c | 6.92±0.23 ns | |
| Carotenoid | UT | 5.61±0.38 a | 6.40±0.52 a | 17.75±0.09 a |
| MS | 3.11±0.16 b | 3.20±0.59 b | 17.53±0.13 b | |
| SS | 1.09±0.03 c | 1.53±0.10 c | 17.82±0.09 a | |
| Chl a/b ratio | UT | 2.28±0.05 a | 2.38±0.06 a | 0.73±0.06 ns |
| MS | 1.83±0.07 b | 1.31±0.06 b | 0.81±0.06 ns | |
| SS | 1.06±0.05 c | 1.16±0.06 c | 0.88±0.15 ns | |
| Car/ChlT ratio | UT | 2.00±0.02 c | 1.89±0.03 b | 2.47±0.03 b |
| MS | 2.19±0.03 b | 2.17±0.01 a | 2.50±0.02 ab | |
| SS | 2.56±0.05 a | 1.94±0.06 b | 2.58±0.07 a |
The data were analyzed using Tukey's HSD test and are expressed as mean ± standard deviation (n = 5). Means with the same letters are not significantly different (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



