
Submitted:
20 October 2024
Posted:
21 October 2024
You are already at the latest version
Abstract
Insects, nematodes, and aquatic animals face several biotic and abiotic stressors that can significantly affect their fitness – specifically damaging their cellular proteins' competent functional nature. In response, they have evolved sophisticated stress-responsive mechanisms. Certain endogenous proteins, known as small heat shock proteins (sHSPs), serve to maintain the stability and function of proteins under stress. Since the identification of the first sHSPs, an increasing number of sHSPs, mainly due to the new robust sequencing tools, continue to be identified and reported to play a critical role in the organism’s response to stress. This review explores and summarizes the contributions of the sHSPs implicated in stress response. Understanding their function is crucial for advancing our knowledge in agricultural pest management, climate change adaptation, and biotechnology.
Keywords:
Abiotic stress
; Biotic stress
; Cell damage
; Apoptosis
; Oxidative stress
; Resistance
; Insects
; Nematodes
; Aquatic animals
1. Introduction
Insects, nematodes and aquatic animals are regularly exposed to biotic and abiotic stressors such as increasing temperature, salinity, and heavy metal accumulation that can significantly affect their physiology, growth and development, behavior, and survival [1,2,3]. Changes in the stressors above the organism’s threshold can disrupt their intracellular proteins and consequently induce physiological stress, cell damage and death [1,4]. For example, in the fruit fly Drosophila virilis, exposure of pupae to heat stress delayed their sexual maturity, whereas in adults, it caused sterility within seven days post-exposure [5]. In the face of climate change, stressors will likely become more acute or chronic and, therefore, have more impact on organisms [6]. In a simulation experiment, increased heatwaves (frequency and severity) reduced the chemosensitivity of bumblebees, as pollinators, to sense and locate plants - as their food source [7]. This can, consequently, lead to the decline of the bumblebee population/colony and food security [7].
While stressors pose significant challenges to organisms, they also drive evolutionary adaptations. Organisms have evolved and can employ either behavioral adaptations to avoid or escape the stressor or physiological adaptations to endure the stressor better [8,9]. As a physiological response, cells synthesize small heat shock proteins (sHSPs) that function as chaperons - bind to the partially folded or denatured substrate proteins, preventing them from irreversible folding/denaturation or enhancing their correct functional folding (Figure 1A) [4,10,11,12,13]. sHSPs have both constitutive and stress-inducible members [14], with a low molecular weight (typically 12 – 42 kDa) and a highly conserved α-crystallin domain (ACD) consisting of 80 – 100 amino acids) [15]. Under stress conditions, some sHSPs can accumulate and reach up to 1% of the total cellular proteins [16]. Unlike other families of Heat shock proteins (HSP), sHSPs do not require ATP to perform their chaperone activity [17] – thus, they act fast in response to stress, and their response is less costly energy-wise.
The chaperone role of sHSPs is not only during stress exposure but also during normal development. Actually, sHSPs have been implicated in various biological processes such as cell growth, differentiation, lipid membrane polymorphism [18,19,20], diapause [21], and lifespan [22]. For instance, heat shock protein p26 in brine shrimp (Artemia sp.), with a crystalline structure as shown in Figure 1B, is reported to confer resistance to Artemia embryos exposed to stressors e.g., extreme temperatures, prolonged anoxia, and desiccation during encystment and diapause [23]. It is noteworthy that in all developmental stages, p26 is expressed exclusively in the embryo stage [24], and constitutes 10 – 15 % of the total yolk proteins. While the chaperone function/mechanism of p26 is reported to be by the formation of ~500KDa oligomers [25,26], other in vitro studies conducted with transfected mammalian cells cultured under stress show that it could be through apoptosis inhibition [27]. Transfected mammalian cells have exhibited enhanced thermotolerance [27], and resistance to oxidative damage induced by hydrogen peroxide [24].
The high evolutionary conservation observed in sHSPs across nematodes, insects, fish, and other organisms underscores their importance in fundamental cellular processes. Amino acid sequence alignment of sHSPs from nematodes, insects, and fish reveals how highly these proteins are conserved across these taxas. In nematodes, particularly in C. elegans, the conservation of sHSPs is highly evident. For instance, multiple amino acid sequence alignment of CeHsp12 [28,29] showed over 86% mean pairwise similarity across various species within the Caenorhabditis genus Figure 2.1. Similarly, in fish, such as Carassius sp., Cyprinus sp., Sinocyclochellus sp. Ctenopharyngodon sp., Culter sp, the sHSP family are well-conserved, with sHsp20 showing over 95% pairwise similarity in sequence alignment Figure 2.2. Moreover, in insects, Amyelois transitella, Manduca sexta, Leptosia nina, Zerene cesonia, Colias sp., Bombyx mori, Maniola sp., Ostrinia nubialist, Parnassius sp., Chilo sp., Loxostege sticticalis, all exhibit more than 90% pairwise similarity of sHsp19 Figure 2.3.
While this review focuses on sHSPs, other HSP families – mainly classified based on molecular weight - are excellently reviewed, among other publications [10,30]. In insects, nematodes, and aquatic animals, the first sHSPs were recognized and extracted from D. melanogaster in 1974 [30], C. elegans in 1983 [31], and brine shrimp (Artemia sp.) in 1999 [32], respectively. Since then, due to new sequencing tools such as genomics and transcriptomics, an increasing number of sHSPs has continued to be reported in several of these organisms such as Diamondback moth (Plutella xylostella), C. elegans, Meloidogyne hapla, Penaeus monodon, V. philippinarum and goldfish (Carassius auratus) [33,34,35]. Functional characterization of these genes in stress response has also been reported and excellently reviewed in papers such as [9,11,36,37]. This review summarizes several sHSPs involved in the response of insects, nematodes, and aquatic organisms to abiotic and biotic stressors. Understanding their function is crucial for advancing our knowledge in agricultural pest management, climate change adaptation, and biotechnology. In the following sections, we explore how sHSPs contribute to the resilience of these organisms under various environmental stressors, highlighting their relevance in mitigating environmental stressors and maintaining cellular homeostasis.
Figure 2.1.
Amino acid sequence alignment of CeHsp12 among different nematode species. The alignment was performed using the MAFFT algorithm in Geneious Prime software 2024.0.7. The mean pairwise identity over all pairs in the column is depicted. Green: 100% identity, greenish-brown: at least 30% identity but less than 100%, red: less than 30%.
Figure 2.1.
Amino acid sequence alignment of CeHsp12 among different nematode species. The alignment was performed using the MAFFT algorithm in Geneious Prime software 2024.0.7. The mean pairwise identity over all pairs in the column is depicted. Green: 100% identity, greenish-brown: at least 30% identity but less than 100%, red: less than 30%.

Figure 2.2.
Amino acid sequence alignment of sHsp19 among different insect species. The alignment was performed using the MAFFT algorithm in Geneious Prime software 2024.0.7. The mean pairwise identity over all pairs in the column is depicted. Green: 100% identity, greenish-brown: at least 30% identity but less than 100%, red: less than 30%.
Figure 2.2.
Amino acid sequence alignment of sHsp19 among different insect species. The alignment was performed using the MAFFT algorithm in Geneious Prime software 2024.0.7. The mean pairwise identity over all pairs in the column is depicted. Green: 100% identity, greenish-brown: at least 30% identity but less than 100%, red: less than 30%.
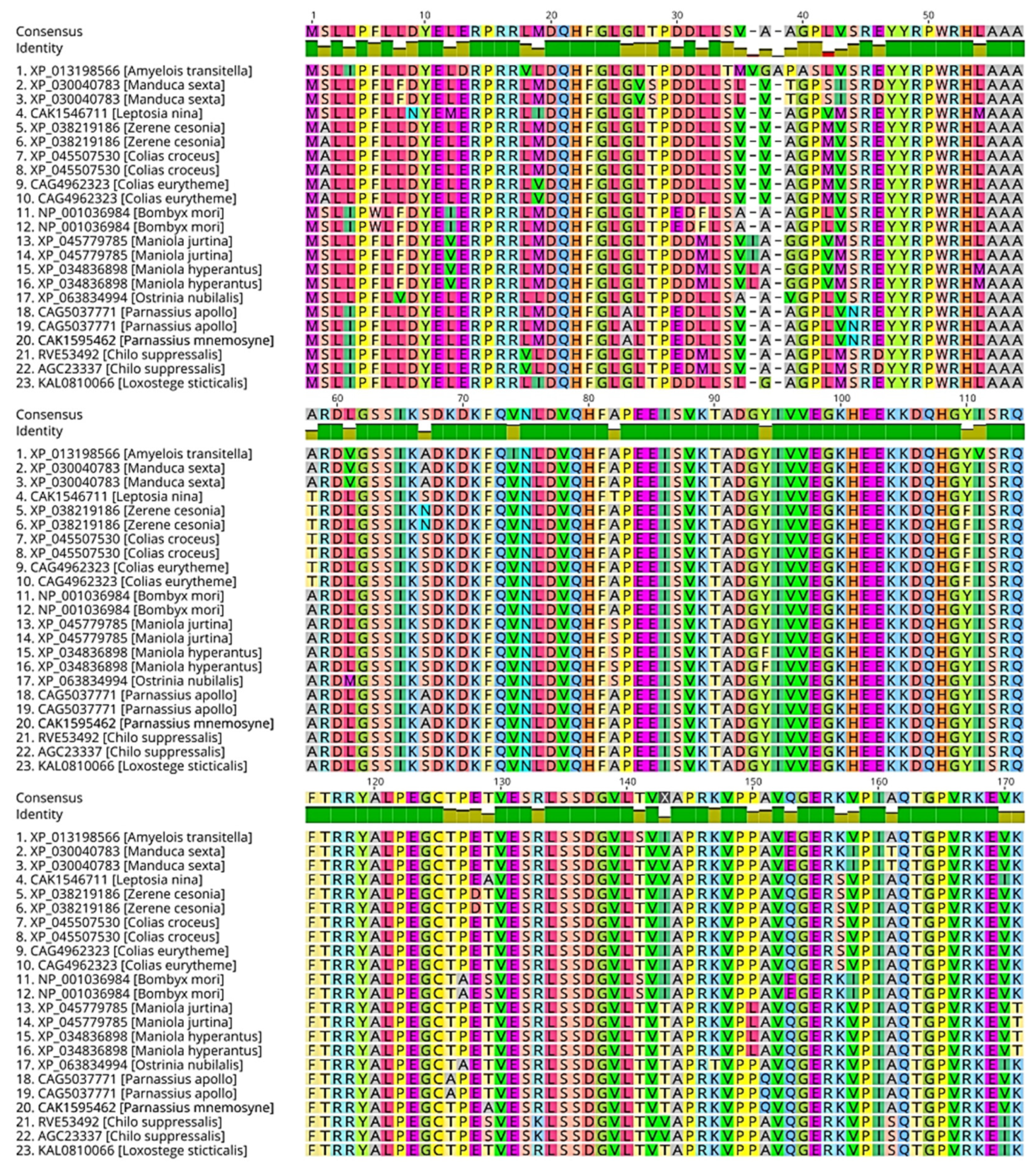
Figure 2.3.
Amino acid sequence alignment of sHsp20 among different fish species. The alignment was performed using the MAFFT algorithm in Geneious Prime software 2024.0.7. The mean pairwise identity over all pairs in the column is depicted. Green: 100% identity, greenish-brown: at least 30% identity but less than 100%, red: less than 30%.
Figure 2.3.
Amino acid sequence alignment of sHsp20 among different fish species. The alignment was performed using the MAFFT algorithm in Geneious Prime software 2024.0.7. The mean pairwise identity over all pairs in the column is depicted. Green: 100% identity, greenish-brown: at least 30% identity but less than 100%, red: less than 30%.

2. Small Heat Shock Proteins in Insects
2.1. Temperature Extremes Stressor
Temperature is one of the most important determining factors of insect abundance and distribution [9,38]. Exposure to high temperatures, even for a short time, can negatively affect insect growth and development [39,40]. For instance, fruit fly Zeugodacus tau exposed to high temperatures (34°C and 38°C) for a short duration (12 h) exhibited changes in their mating behavior, antioxidant defense, and detoxifying enzymes, albeit in a sex-specific manner [41]. Notably, high temperatures promoted mating, a behavioral adaptation strategy for survival under environmental stress.
Insects produce sHSPs as one of the various physiological responses to exposure to temperature extremes [11,12]. Several sHSPs have been responsible for offering heat tolerance, heat acclimation, and hardening [42,43]. For example, in a study tracking levels of sHSPs in outdoor larvae of gall fly, Eurosta solidaginis, the sHSP αB-crystallin increased in expression levels just prior to and during cold weather (i.e., in late fall and winter), suggesting that it confers protection from coldness [44]. Although temperature extremes cause physiological damages, within specific temperature limits the damages caused are reversible, such as with the help of sHSPs [45,46]. In Drosophila melanogaster, two genes (Hsp22 and Hsp23) were upregulated during recovery from cold treatment but not during the cold stress exposure. Also, the role of sHsps in thermal tolerance has been shown through gene knockdown by RNAi. For instance, the thermal tolerance of the soil-dwelling beetle Gastrophysa atrocyanea lowered when two sHSPs genes (i.e., sHSPs 21 and 23) were silenced [47].
2.2. Desiccation Stressor
Maintaining water balance is a critical requirement for insects [48], as dehydration can lead to significant physiological injuries such as protein denaturation, ROS production, nucleic acid damage, and membrane damage through lipid peroxidation [49] and, consequently, death [50]. Due to the severity of the cellular damage resulting from dehydration, it is better prevented than repaired. Thus, it is advantageous for the insect to initiate an early response to dehydration. Several sHSPs have been identified to offset or respond to dehydration/rehydration changes – suggesting their importance in preventing possible protein denaturation. For example, the sHSP, smHsp, in the larvae of Belgica antarctica, an insect that lives in Antarctica, reacted to varying levels of dehydration and rehydration [50]. This Antarctic midge larva spends most of its time frozen in ice, but during summer, the ice melts, exposing it to potential dehydrating conditions such as intense sunlight and wind. Under fast dehydration (75% RH for 36 h) – quick loss of water, sHSP mRNA was increased, and levels continued to be high during rapid rehydration (done by submerging larvae in water for two hours), indicating involvement in preparation for stress exposure. Several other cases of sHSPs involvement in desiccation are presented in Table 1.
2.3. Pesticides and Heavy Metals Stressor
Exposure to pesticides is a significant contributor to terrestrial insect population decline [51,52]. Pesticide exposure is linked to increased ROS production in cells, which, if not neutralized or regulated to low levels, targets proteins whose amino acid residues are oxidation-sensitive and consequently can change their functional structure and/or cause aggregation [53,54,55,56]. Besides exposure to targeted insecticides, insects can also get impacted by exposure to non-insect targeted pesticides such as herbicides due to sharing a common habitat with the target pests [57,58]. For instance, Glyphosphate, the most used herbicide worldwide, negatively affects insect physiology, delays the growth and development of larvae, and disrupts the foraging behavior of worker honeybees Apis mellifera (Farina et al., 2019). Also, glyphosate impaired the habituation of mosquito larvae (Aedes aegypti) under field conditions [59].
Insects respond to pesticide exposure by overproducing sHSPs. ROS activates heat shock factor (HSF1) to stimulate expression of these sHSPs within the cells [60]. sHSPs contribute to repairing the damage from pesticide-induced oxidative stress [61], stimulate the activity of antioxidant-related genes and antioxidant enzymes, or hinder the activation of ROS transcription factors [62,63,64]. In Apis Cerana cerana, a knockdown of the sHSP gene, AccsHSP21.7, decreased its resistance to oxidative stress resulting from exposure to a commercial herbicide, glyphosate, resulting in a significantly increased mortality rate [65]. The promoter region of the AccsHSP21.7 gene was rich in binding sites for oxidative stress transcription factors CREB (cAMP Response Element-Binding Protein) and HSF (Heat Shock Factor) [65]. CREB can help the insect save energy by transitioning to diapause stage [9] e.g., under hypoxia conditions [66], whereas HSF turns on heat shock genes that subsequently encode more sHSP synthesis [67].
The buildup of a heavy metal beyond its functional range is toxic and has been linked to oxidative stress, as the buildup enables the formation of free radicals [68,69]. Also, metals such as Cd can disrupt the proteins involved in DNA repair and DNA damage signaling [70]. Besides, Cd has a high affinity for and interacts with the thiol groups in cysteine residues, disrupting/denaturing competent protein structures and consequently stimulating the expression of chaperone proteins such as sHSPs. For example, exposure of aquatic larvae of Chironomus riparius. affects the expression levels of several sHSPs [71]. In Daphnia magna, a model insect for investigating heavy metal toxicity, sHSPs like DmsHSP1 and DmsHSP5 were upregulated when the insect was exposed to heavy metals (Cd2+, Cu2+, and Zn2+), preventing potential protein agglutination and denaturation (M. Li et al., 2022a). It is noteworthy that besides metallothionein (MT), sHSPs are being explored as possible bioindicators of metal contamination because of their sensitivity to small changes in cell homeostasis and their conserved nature in many species [36,71,72].
2.4. Hypoxia/Anoxia Stressor
Several insects or parts of their life stages live in habitats with consistent or bursts of low oxygen levels, such as ground burrows, decaying organic matter, aquatic ecosystems, flood-prone soils, and high altitudes [73]. Under hypoxic conditions, organisms employ short- and long-term mechanisms to increase their access to oxygen or decrease their oxygen demand, and failure can result in tissue damage or death [74]. Mortality of adults of the flesh fly Sarcophaga crassipalpis exposed to severe hypoxia (3% oxygen) reached LD50 within seven days [75]. Therefore, the ability to survive in hypoxic or anoxic conditions is necessary for the survival of these insects. Several studies have shown evidence of induction of sHSPs when insects are subjected to hypoxia/anoxia conditions, as detailed in Table 1. For example, in the flesh fly (Sarcophaga crassipalpis), which typically burrows underground [76], three sHSPs (hsp25, hsp23, and hsp18) were upregulated within two days during exposure to hypoxia (3% oxygen), and upregulation continued 2 hrs post-exposure albeit rapidly declined thereafter, returning to control levels under normoxia conditions [75].
2.5. Crowding and Starvation Stressors
Generally, when organisms are crowded, they are at risk of being stressed [77], and consequently, their growth and development processes are affected. For instance, in the locust species Locusta migratoria L., the fecundity of solitary locusts was higher compared to that of gregarious locusts (representative of crowding) [78]. As a stress factor, population density has been shown to affect the physiology of organisms, such as inducing the production of sHSPs, as shown in Table 1. In the migratory locust, Locusta migratoria L., mRNAs of three sHSPs (Hsp20.5, Hsp20.6, and Hsp20.7) were more expressed in gregarious phases compared to solitary phases [77]. It is worth noting that insects can experience multiple stressors in a crowd, including starvation, pathogen infection, and desiccation. Therefore, given that HSP induction is in response to any stressor that causes protein unfolding [79], it is difficult to decipher the contributions of a single stressor to sHSPs induction [77]
Within the context of starvation, when an insect is deprived of food, its energy homeostasis can be damaged [80]. sHSPs have been implicated in responding to this stress sensitively (Table 1). In the 4-day-old larvae of Housefly Musca domestica, the expression of MdomHSP27 was significantly down-regulated after a six-hour starvation period (Tian et al., 2018). Silencing of TcHSP18.3 in Tribolium castaneum Herbst [81], and hsp27 gene in Drosophila melanogaster [82] reduced the respective insect’s ability to endure starvation. These findings suggested that sHSPs can be sensitively responsive to starvation in insects.
2.6. Parasitism Stressor
Parasitism can be a stressor inducing sHSPs in hosts [83], eliciting induction of sHSPs for instance in respone to the venom injected into it. Envenomation of Pieris rapae Linnaeus by the endoparasitic wasp Pteromalus puparum stimulates expression of hsp20 in the host [84]. Similarly, envenomation in flesh fly Sarcophaga crassipalpis by the ectoparasitic wasp Nasonia vitripennis stimulates upregulation of hsp23 13 h after envenomation and maintained for 48 h from post envenomation suggesting that sHSPs could be involved in immune responses to parasite insult [83]. Also, envenomation of the larvae of Plodia interpunctella. by the ectoparasitoid Bracon hebetor stimulated upregulation of sHSP (shsp) [85]
2.7. UV Radiation Stressor
Some studies have indicated a correlation between UV-induced stress and the expression of sHSPs in insects. UV radiation, such as UV-A photo-oxidize photosensitizers inside insect cells results in the production of large quantities of ROS that ultimately impair the functional competence of proteins [86,87,88]. For instance, UV-A irritation on Spodoptera frugiperda adults resulted in increased expression levels of five sHSPs genes (SfsHsp21.3, SfsHsp20, SfsHsp20.1, SfsHsp19.3, and SfsHsp29) indicating that they play a role in molecular response mechanisms [89]. Similarly, expression levels of Hsp22.6 and Hsp27.6 in the honeybee Apis cerana cerana [90,91], as well as those of Hsp27 and Hsp21.8b in the beetle Tribolium castaneum, were upregulated upon exposure to UV radiation [92,93].
2.8. Energy Cost of sHSps Response
sHSPs enhance insects’ ability to resist or recover from stressors. However, there is a potential fitness cost linked to overexpression of sHSPs [94]. For instance, in Drosophila, cells that overexpressed the large heat shock protein Hsp70 at normal temperature grew slower than normal cells – indicating that accruing Hsp70 is costly to cell growth [95]. Also in locust, solitary locusts showed larger body size, longer life span, and greater reproductive potential compared to gregarious locusts that had expressed HSPs [96]. Such costs probably originate from the stoppage of normal cell functions during the stress response, high energy expenditure, and the toxic effects of high HSP concentrations produced by the stressor. One of the strategies employed to reduce sHSPs response energy cost by insects is momentary sHSPs expression – where a decline in expression levels is observed with continuous exposure or when the stressor fades [97]. Nevertheless, the benefit of HSPs overexpression could still outweigh the potential associated costs, e.g., under density scenarios [77].

Table 1.
Small heat-shock proteins (sHSPs) in insects stimulated in response to exposure to several stressors.
Table 1.
Small heat-shock proteins (sHSPs) in insects stimulated in response to exposure to several stressors.
| Insect species | Small heat shock protein/gene | Expression pattern | Reference |
|---|---|---|---|
| TEMPERATURE stressor | |||
| Corn stalk borer (Sesamia nonagrioides larvae) | SnoHsp19.5 and SnoHsp20.8 | Both genes were upregulated (within 15 min) by heat shock at 40 °C and when larvae recovered after cold shock. |
Gkouvitsas et al. [21] |
| Leaf beetle (Gastrophysa atrocyanea) | sHsps 21 and sHsps 23 | RNAi knockdown of both genes decreased viability and lowered heat resistance. | Atungulu et al. [47] |
| Liriomyza sativae pupae |
ls-hsp19.5, ls-hsp20.8 and ls-hsp21.7 |
All significantly induced by cold treatment with ls-hsp20.8 displayed the greatest sensitivity. This suggests that different sHsps may be responsive to various stressor intensities. |
Huang et al. [98] |
| Chaperone proteins aid winter survival of freeze-tolerant gall fly larvae Eurosta solidaginis. | α- crystallins and β-crystallins | The sHSPs αB-crystallin increased in levels just prior to and during cold weather (i.e., in late fall and winter). Both αA and αB crystallin were highly induced in response to freeze/thaw conditions. |
Zhang et al. [44] |
| Silkworm (Bombyx mori) | shsp19.9, shsp20.1, shsp20.4, shsp20.8, shsp21.4, shsp23.7 and shsp21.4. | All genes were upregulated by heat stress except shsp21.4, which was downregulated. |
Sakano et al. [99] Li et al. [100] |
| Red flour beetle (Tribolium castaneum) | Tchsp18.3 | Gene was upregulated in response to heat stress but not to cold stress. | Xie et al. [81] |
| Drosophila melanogaster | Hsp22 and Hsp23 | The removal of the genes mRNA by RNAi interrupted recovery (time to recover and mobility following recovery) from chill injury thus showing that upregulation of the genes is required for recovery, but not during the cold stress itself. | Colinet et al. [101] |
| flesh fly (Sarcophaga cras-sipalpis) | Hsp23 | Deletion of genes’ mRNA reduced cold hardiness. | Rinehart et al. [102] |
| Western flower thrip (Frankliniella occidentalis) | FoHSP11.6 and FoHSP28.0 | Both generes were induced by both low and high temperature with maximum expression levels attained after 0.5 – 1 h of temperature stress exposure. Also, thermotolerance reduced when both genes were silenced by RNAi. | Yuan et al. [103] |
| Chilo suppressalis (Walker) | Cshsp19.0 | Gene was upregulated as a response to heat and cold stress exposure for 2 h. | Dong et al. [104] |
| Spodoptera frugiperda | SfsHsp21.3, SfsHsp20, SfsHsp20.1, SfsHsp19.3, and SfsHsp29. | All genes were significantly upregulated at both temperature extremes (42°C and 4°C) with the exception of two genes (SfsHsp20.1 and SfsHsp19.3) in the adult males that did not respond to the 4°C treatment. | Yang et al. [89] |
| PESTICIDE and heavy metal stressor | |||
| Apis cerana cerana | AccsHSP21.7 | A knockdown of the sHSP gene decreased the insect’s resistance to a commercial herbicide glyphosate, resulting in significant mortality. |
Huang et al. [65] |
| Fall armyworm (FAW) (Spodoptera frugiperda) | sHsp19.07, sHsp20.7 and sHsp19.74. | All genes were upregulated following exposure to the Chlorantraniliprole pesticide. Though sHsp19.74 reached maximum mRNA expression levels faster (8 h after exposure) than the rest (12h), its levels plummeted at 12 h after exposure, suggesting a momentary responsiveness of sHSPS to pesticide treatment. |
Samanta et al. [105] |
| Diamondback moth (Plutella xylostella L) | Fourteen sHSPs (sHSP27.5, sHSP28.9, sHSP21.6, sHSP18.8, sHSP19.22, sHSP21.8, sHSP21.9, sHSP22.1, sHSP23.4, sHSP19.5, sHSP20.06, sHSP20.09, sHSP19.23, sHSP20.1) | Fourth instar larvae were exposed to various pesticides and heavy metals for 24 hr. sHSPs responses were as below. Beta-cypermethrin pesticide significantly upregulated all except sHSP20.09, whereas chlorfenapyr pesticide downregulated all except sHSP28.9. Expression responses to Indoxacarb and Cantharidin were irregular. Exposure to H2O2 for 24 h downregulated five sHSPs (sHSP19.22, sHSP19.23, sHSP21.6, sHSP22.1, and sHSP23.4) Copper (Cu2+) downregulated three sHSPs (sHSP20.1 sHSP22.1, sHSP28.9) and upregulated seven sHSPs (sHSP19.22, sHSP19.23, sHSP20.06, sHSP20.09, sHSP21.8, sHSP21.9, sHSP27.5). Manganese (Mn2+) upregulated four sHSPs (sHSP20.1, sHSP21.6, sHSP22.1, sHSP28.9) and upregulated all the rest. Nickel (Ni2+) upregulated (sHSP19.22, sHSP19.5, sHSP20.06, sHSP20.09), not induced (sHSP20.1, sHSP21.8, sHSP21.9), and the rest were downregulated. Gene expression response to Lead (Pb2+) was irregular. |
Chen & Zhang [33] |
| Daphnia magna | eleven sHSP genes (termed DmsHSP1 - DmsHSP11) | Insect exposure to heavy metals (Cd2+, Cu2+, and Zn2+) upregulated DmsHSP1 and DmsHSP5. RNAi knockdown of genes DmsHSP1–21, except DmsHSP11–12.8, increased susceptibility to heavy metal stress exposure. |
Li et al. [56] |
| Acquatic larvae of Chironomus riparius. |
hsp17, hsp21, hsp22, hsp23, hsp24, hsp27, and hsp34) | Following acute exposure to Cadmium (Cd), hsp23, hsp24, hsp27, and hsp34 were upregulated, whereas levels of hsp17 and hsp21 remained unaltered. This indicates that sHSPs have diverse roles during response to Cd. | Martín-Folgar & Martínez-Guitarte [71] |
| Chinese rice grasshopper (Oxya chinensis) | OcGrp78, OcHsp70, OcHsp90, and OcHsp40 | Following exposure to Cadmium (Cd), mRNA expression levels of all genes increased, reaching a maximum within a short period (6 h), albeit decreasing significantly after 12 h. |
Zhang et al. [106] |
| HYPOXIA OR ANOXIA stressor | |||
| Flesh fly (Sarcophaga crassipalpis) | hsp25, hsp23, and hsp18 | hsp25, hsp23, and hsp18 were upregulated by at least 10-fold within two days of hypoxia (3 % oxygen) treatment application. Upregulation was maintained for the whole treatment period (10 days) and during recovery - 2 h post-treatment – after which expression levels declined. | Michaud et al. [75] |
| Gall fly larvae (Eurosta solidaginis) | αA and αB crystallin | Both sHSPs increased in response to anoxia (exposure period of 24 h under N2 gas at 15 °C) |
Zhang et al. [44] |
| CROWDING stressor | |||
| Migratory locusts (Locusta migratoria L) | Hsp20.5, Hsp20.6, and Hsp20.7 | mRNAs of all sHPS were more expressed in gregarious phases (representing high population density) compared to solitary phases (representing low population density) | Wang et al. [77] |
| fifth-instar nymphs of the Australian plague locust (Chortoicetes terminifera) | Hsp20.5 and Hsp20.7 | Crowding (during the gregarious phase) resulted in a 2 – 3-fold significant upregulation of both genes. | Chapuis et al. [107] |
| STARVATION stressor | |||
| Mulberry pyralid caterpillar (Glyphodes pyloalis) | GpHSP19.5, 20, 20.2, and 21.6 GpHSP21.8 and GpHSP21.4 |
Genes were upregulated time-dependently, reaching maximum levels on the sixth day of food deprivation. On the contrary, expression levels of two GpsHSPs (GpHSP21.8 and GpHSP21.4) demonstrated intermittent downregulation in comparison to the control at 2 or 4 days following the starvation period. | Chu et al. [80] |
| Fruitfly (Drosophila melanogaster) |
Hsp27 | sHPS was knocked out, and flies showed a significant decrease in resistance to starvation. | Hao et al. [82] |
| 4-day-old larvae of Housefly (Musca domestica) | MdomHSP27 MdomHSP10, MdomHSP27.1, and MdomHSP27.2 |
the expression of MdomHSP27 was significantly downregulated after a 6h starvation period, whereas the other 3 MdomHSPs (MdomHSP10, MdomHSP27.1 and MdomHSP27.2) were not significantly affected |
Tian et al. [108] |
| Red flour beetle (Tribolium castaneum) | Tchsp18.3 | When sHPS was knocked down, the lifespan of adult beetles was reduced by 15.8% (they died within 18 days after starvation) compared to the control group. |
Xie et al. [81] |
| Parasitoid wasp (Pteromalus puparum Linnaeus) | PpHSP20 | Gene expression increased significantly after 6 h of starvation but declined after 24 h |
Wang et al. [109] |
| Diamondback moth (Plutella xylostella L.) | sHSP20.1, sHSP21.6, sHSP22.1, and sHSP28.9, | Expression levels of all sHSPs were significantly downregulated following food starvation for 21 h. | Chen & Zhang [33] |
3. Small Heat Shock Proteins in Nematodes
3.1. Heat Stressor
Heat stress is a major environmental challenge for nematodes, particularly with the increasing impact of global climate change. Increasing temperatures can cause protein denaturation and malfunction, which in turn activates the heat shock response [110]. During this process, sHSPs are upregulated to aid in the refolding of damaged proteins, thus boosting the thermotolerance of nematodes [111]. Research has demonstrated that C. elegans relies on sHSPs for survival under high-temperature conditions, highlighting their critical role in protecting against heat-induced stress [112]. For instance, CeHsp16.1 and CeHsp16.2 are among the first proteins to be upregulated in response to heat shock at 35°C, and their deletion reduces survival under prolonged heat stress [113]. Other nematodes also exhibit a robust heat shock response. For example, in Meloidogyne hapla, sHSPs MhHsp12.2, MhHsp6, MhHsp1, and MhHsp43, are induced by heat stress with higher expression levels at 35-40°C [34]. In Bursaphelenchus xylophilus, Bx-sHSP16A, Bx-sHSP16B, Bx-sHSP21, and BxsHSP25 are upregulated in response to both heat and cold stress, playing a dual role in temperature adaptation [37]. In Strongyloides ratti, the expression of SrHsp17 is significantly elevated during heat stress and when exposed to pathogens, particularly in the infective larval stage whereas sHSPs 18 and sHSPs 22 are up-regulated at 37°C In third-stage larvae of filarial nematode Brugia pahangi [114,115]. These broad responses across multiple nematode species demonstrate the vital function of sHSPs in maintaining cellular integrity during heat stress.
3.2. Cold Stressor
Although often less discussed, cold stress can also impact nematodes, particularly in environments with seasonal temperature fluctuations. Cold exposure can affect the fluidity of cellular membranes and the stability of proteins [116]. sHSPs are involved in the cold shock response of organisms, helping to preserve protein stability under low temperatures [117]. Evidence suggests that sHSPs are upregulated in nematodes exposed to cold environments, aiding in their ability to maintain cellular functions and protect proteins from cold-induced damage [118]. For example, in C. elegans, heat shock factor 1 (hsf-1) is triggered inducing the nematode into an arrested state of development (diapause) at 4 °C and this is essential for maintaining cellular stability under cold temperatures [119]. Similarly, in M. hapla, MhHsp12.2, plays a dual role, being upregulated both during heat and cold shock. The expression of this gene at low temperatures (-20 °C) enhances cold tolerance, allowing the nematode to survive periods of cold exposure during the winter season [34]. Additionally, in Panagrolaimus davidi, a cold-adapted nematode from Antarctic environments, sHSPs are crucial in ensuring survival at subzero temperatures, by maintaining protein stability and membrane fluidity under freezing conditions [120].
3.3. Desiccation Stressor
Desiccation, or dehydration, also poses a serious threat to nematode survival. During dehydration, cellular structures, including proteins, are at risk of damage. sHSPs play a vital role in enhancing desiccation tolerance by stabilizing proteins and cellular structures and preventing irreversible damage during water loss [26]. Nematodes can survive extreme desiccation, which helps them maintain cellular integrity during periods of water scarcity [121]. In Caenorhabditis elegans, the dauer larvae stage is particularly adapted to survive harsh desiccation conditions and exhibits high levels of sHSP p16 [122]. In this way, sHSPs enable nematodes to thrive in environments where water availability is unpredictable, making them essential for desiccation tolerance. Similarly, in Panagrolaimus superbus, a model species for desiccation research, both PsHSP17.1 and PsHSP20 are upregulated during desiccation and dehydration. These proteins are essential for anhydrobiosis, a process that allows nematodes to survive in completely dry environments [123].
3.4. Anhydrobiosis and Hypoxia Stressors
In extreme cases, such as anhydrobiosis (a state of suspended animation in response to complete desiccation) and hypoxia (low oxygen conditions), sHSPs have been found to play a protective role. During anhydrobiosis, where nematodes experience extreme dehydration, sHSPs help stabilize cellular components, preventing damage during the transition to and from the dried and hypoxic state [124]. In Heterorhabditis indica, Dnj-13 which is a class B J-protein belonging to hsp40 family is upregulated when subjected to anhydrobiotic conditions [125] allowing the nematode to survive complete dehydration for extended periods. Similarly, under hypoxic conditions, sHSPs contribute to cellular protection by maintaining protein homeostasis. For example, In C. elegans, the heat shock proteins hsp-16.1 and hsp-16.2 were upregulated following a one-hour exposure to hypoxic conditions in specialized hypoxia chambers [126].
3.5. Chemical Stressor
Nematodes, like many organisms, are frequently exposed to a variety of chemical stressors in their environments, including heavy metals, pesticides, and other toxic compounds. Heavy metal exposure is a common environmental stressor that triggers oxidative stress by generating reactive oxygen species (ROS) [127]. Oxidative stress arises when there is an imbalance between the production of reactive oxygen species (ROS) and the organism’s ability to detoxify them. ROS can lead to significant cellular damage, including oxidation of proteins, lipids, and DNA [128]. In C. elegans, sHSPs are often co-expressed with other antioxidant enzymes, such as superoxide dismutase and catalase, providing a coordinated defense against oxidative damage [129]. Ezemaduka et al. [130], showed that the expression of CeHSP17 increases when C. elegans is exposed to toxic heavy metals like cadmium and zinc. These metals can cause protein misfolding, and sHSPs act to prevent the aggregation of damaged proteins. Pesticides and other xenobiotic compounds are widely present in agricultural soils, posing a significant chemical stress challenge to nematodes. sHSPs have been implicated in the defense against pesticide-induced stress. For example, exposure to the pesticide cypermethrin leads to oxidative damage and protein misfolding in C. elegans. In response, at non-lethal concentrations, sHSPs HSP-16 are upregulated to prevent the aggregation of damaged proteins, promoting cellular recovery and survival under chemical stress [131].
4. Small Heat Shock Proteins in Aquatic Animals
4.1. Heavy Metal, Temperature, and Salinity Stress
Poor disposal of industrial wastes often leads to contamination and accumulation of heavy metals (HMs) in aquatic ecosystems [132,133]. Cadmium (Cd) considered the most dangerous HM to aquatic animals is the most common contaminant from industrial wastes [134]. Even at low concentrations, this HM can accumulate in aquatic animals’ vital organs, leading to death [135,136]. HM stress leads to oxidative stress in exposed tissues due to the accumulation of reactive oxygen species (ROS) [137,138]. Moreover, HMs and metalloids negatively impact protein homeostasis and the viability of cells by interfering with the protein folding processes in cells [139,140]. sHSPs have been reported to play a vital role in alleviating the stress effects of HMs in certain aquatic animals. MnHSP28.6 was found to predominately accumulate in the muscles of the oriental river prawn (Macrobrachium nipponense) upon exposure to Cd and copper (Cu) ions hence conferring HM resistance in stressed animals. Moreover, in vitro expression of this sHSP in Escherichia coli conferred host cells impressive protection against hydrogen peroxide [141]. In another study, Zhang et al. [142] demonstrated that exposing razor clam (Sinonovacula constricta) to HM stress (Cd and lead (Pb)) led to an increased accumulation of heat shock protein ScsHSP in haemocytes, gonads, and mantle hence triggering tolerance to HM toxicity stress in S. constricta. Likewise, Yang et al. [143] observed increased expression of sHSP LcHSP27 in the liver and brain of yellow croaker (Larimichthys crocea) exposed to Cd ions, which improved stress tolerance. Moreover, LcHSP27 expression was enhanced in the liver under high and low temperatures thus indicating its role in protecting the liver from multiple stress factors. In another study, Li et al. [144] reported improvement in the level of expression of VpsHSP-2 in Manila clam (Venerupis philippinarum) exposed to Cd ion stress. A previous study on hard clam (Meretrix meretrix) showed that exposing the animal to Cd stress led to increased expression of mRNA levels of Mm-HSP 20 in the digestive gland and hemocytes hence indicating its role in HM detoxification and protection against oxidative stress [145]. Li et al. [56] exposed Daphnia magna to different levels of heavy metals (Cd, Cu, and zinc [Zn]), and the authors observed the expression of eleven sHSPs (DmsHSP1 – DmsHSP11) under HM stress. Exposing the animals to Cu2+ strongly induced the expression of DmsHSP1 while it was moderately induced under Cd2+ and Zn2+ exposure. The authors also observed that DmsHSP5 was induced in all the HM stress.
Temperature stress is one of the major abiotic stress factors affecting the survival of aquatic animals. Changes in the environmental temperatures below or above the organism’s thermal threshold cause oxidative stress that could hinder the survival and general well-being of the organism [1]. The expression of sHSPs in tissues of stressed animals could aid in the alleviation of temperature stress effects in the organisms. For instance, Bildik et al. [146] investigated changes in the expression of heat shock proteins (sHSP 30) in gilt-head bream (Sparus aurata) exposed to high and low temperatures and observed a two-fold increase in the expression of sHSP30 under higher temperatures (27°C) which led to improved thermal tolerance in fish. In another study, Chen et al. [35] observed an increase in the relative expression of sHSP20 in the spleen of goldfish (Carassius auratus) exposed to high temperatures (32°C) which correlated with the fish’s thermal tolerance. Currie [147] demonstrated that acclimatization of rainbow trout (Oncorhynchus mykiss) to higher temperatures (25°C) led to increased expression of sHSP30 in all tissues except blood. Likewise, Liu et al. [148] reported increased expression of sHSP30 in the gills of O. mykiss exposed to temperature stress (25°C) hence indicating its role in heat stress tolerance in fish. A previous study on pool barb (Puntius sophore) showed increased expression of sHSP47 exposed to temperature stress (41°C).
Salinity stress in aquatic animals leads to the accumulation of ROS whose over-accumulation in tissues leads to cell damage and apoptosis. Expression of certain sHSPs in aquatic animals exposed to high or low salinity stress has been shown to enhance the stress tolerance of the organism. Shekhar et al. [149] investigated stress responses of P. monodon exposed to low salinity stress and observed increased expression of sHSP21 in tissues thus suggesting its role in salinity stress adaptive mechanism in shrimp. A recent study by Zarei et al. [150] has shown that Sterlet Sturgeon (Acipenser ruthenus) can adapt to low salinity stress through the expression of sHSP27 which is known to confer stress resistance in organisms.
4.3. Pathogenic Infection
sHSPs induced by biotic stress play a significant role in protein folding, enhancement of immune responses, and protection against diseases caused by various pathogenic organisms and viruses [151]. Although several reports have previously reported on the induction of sHSPs by abiotic stress factors, little information is known about the induction of sHSPs by abiotic stress factors. Certain studies have suggested that members of the sHSP20, and sHSP37 among others have been identified in crustaceans and several fish species. Chen et al. [35] conducted a study on goldfish (C. auratus) to understand how pathogen infection can induce the expression of sHSP in animal tissues. The authors observed that intraperitoneal injection of lipopolysaccharide (LPS) or polyinosinic-polycytidylic acid (Poly I:C) induced the expression of sHSP20 in the spleen thus improving the fish’s tolerance to stress. Arockiaraj et al. [152] reported that the expression of MrHSP37 in freshwater prawns (Macrobrachium rosenbergii) triggered an immune response upon exposure to pathogen infection (hypodermal and hematopoietic necrosis virus). Li et al. [144] demonstrated that exposing Manila clam (V. philippinarum) to a pathogen challenge (Vibrio anguillarum) induced the upregulation of VpsHP-1in the haemocytes and increased 1.5-fold and 9.9-fold 6 h and 96 h post-infection respectively. Likewise, the expression level of VpsHP-2 increased by 8.7-fold compared to the control group 24 h post-infection.
Certain studies have, however, shown a down-regulation of sHSPs in tissues of aquatic animals exposed to biotic stress factors. For instance, Huang et al. [153] demonstrated that Penaeus monodon when challenged with the white spot syndrome virus (WSSV) has a significantly decreased expression of sHSP 21 thus indicating that the gene regulation of sHSP 21 was negatively affected by WSSV.
5. Conclusions
Environmental stressors pose significant challenges to the survival, fitness, and distribution of insects, nematodes, and aquatic organisms, particularly in the context of climate change and increased anthropogenic pressures. Small heat shock proteins (sHSPs) play an essential role in the resilience of these organisms. This review highlights the broad and highly conserved role of sHSPs across species in responding to various biotic and abiotic stressors discussed above. Future research should aim to unravel further the molecular mechanisms and pathways regulated by sHSPs, particularly in the context of multiple simultaneous stressors. Integrating genomics, proteomics, and transcriptomics approaches will be key to uncovering the full spectrum of sHSP functions. Moreover, exploring the evolutionary conservation of sHSPs across taxa will provide deeper insights into their potential as universal stress-resilience mechanisms. Ultimately, enhancing our understanding of sHSPs will not only advance fundamental biological knowledge but also inform practical applications in fields such as agriculture, conservation biology, and environmental monitoring, helping to safeguard biodiversity and ecosystem stability in an era of unprecedented environmental change
Author Contributions
M.M; Conceptualization., J.B, J.A, and M.M; writing the draft, J.B; reviewing. All authors read and approved the final draft
Funding
Not applicable
Data Availability Statement
All datasets used in this study are available from the corresponding author upon reasonable request
Acknowledgments
The authors wish to thank Mr. Fahad Kimera, Manager of Agriculture CARES research for professional support during the writing of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest
References
- Mugwanya, M.; Dawood, M.A.O.; Kimera, F.; Sewilam, H. Anthropogenic Temperature Fluctuations and Their Effect on Aquaculture: A Comprehensive Review. Aquac Fish 2022, 7, 223–243. [Google Scholar] [CrossRef]
- Viitasalo, M.; Bonsdorff, E. Global Climate Change and the Baltic Sea Ecosystem: Direct and Indirect Effects on Species, Communities and Ecosystem Functioning. Earth System Dynamics 2022, 13, 711–747. [Google Scholar] [CrossRef]
- Häder, D.-P.; Banaszak, A.T.; Villafañe, V.E.; Narvarte, M.A.; González, R.A.; Helbling, E.W. Anthropogenic Pollution of Aquatic Ecosystems: Emerging Problems with Global Implications. Science of The Total Environment 2020, 713, 136586. [Google Scholar] [CrossRef] [PubMed]
- Basha, E.; O’Neill, H.; Vierling, E. Small Heat Shock Proteins and α-Crystallins: Dynamic Proteins with Flexible Functions. Trends Biochem Sci 2012, 37, 106–117. [Google Scholar] [CrossRef]
- Walsh, B.S.; Parratt, S.R.; Mannion, N.L.M.; Snook, R.R.; Bretman, A.; Price, T.A.R. Plastic Responses of Survival and Fertility Following Heat Stress in Pupal and Adult Drosophila Virilis. Ecol Evol 2021, 11, 18238–18247. [Google Scholar] [CrossRef]
- Guillén, L.; Pascacio-Villafán, C.; Osorio-Paz, I.; Ortega-Casas, R.; Enciso-Ortíz, E.; Altúzar-Molina, A.; Velázquez, O.; Aluja, M. Coping with Global Warming: Adult Thermal Thresholds in Four Pestiferous Anastrepha Species Determined under Experimental Laboratory Conditions and Development/Survival Times of Immatures and Adults under Natural Field Conditions. Front Physiol 2022, 13. [Google Scholar] [CrossRef]
- Nooten, S.S.; Korten, H.; Schmitt, T.; Kárpáti, Z. The Heat Is on: Reduced Detection of Floral Scents after Heatwaves in Bumblebees. Proceedings of the Royal Society B: Biological Sciences 2024, 291, 20240352. [Google Scholar] [CrossRef]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in Fluctuating Thermal Environments. Annu Rev Entomol 2015, 60, 123–140. [Google Scholar] [CrossRef]
- King, A.M.; MacRae, T.H. Insect Heat Shock Proteins During Stress and Diapause. Annu Rev Entomol 2015, 60, 59–75. [Google Scholar] [CrossRef]
- Sørensen, J.G.; Kristensen, T.N.; Loeschcke, V. The Evolutionary and Ecological Role of Heat Shock Proteins. Ecol Lett 2003, 6, 1025–1037. [Google Scholar] [CrossRef]
- González-Tokman, D.; Córdoba-Aguilar, A.; Dáttilo, W.; Lira-Noriega, A.; Sánchez-Guillén, R.A.; Villalobos, F. Insect Responses to Heat: Physiological Mechanisms, Evolution and Ecological Implications in a Warming World. Biological Reviews 2020, 95, 802–821. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Hofmann, G.E. Heat-Shock Proteins, Molecular Chaperones, and the Stress Response: Evolutionary and Ecological Physiology. Annu Rev Physiol 1999, 61, 243–282. [Google Scholar] [CrossRef] [PubMed]
- Gething, M.-J.; Sambrook, J. Protein Folding in the Cell. Nature 1992, 355, 33–45. [Google Scholar] [CrossRef]
- Colinet, H.; Lee, S.F.; Hoffmann, A. Temporal Expression of Heat Shock Genes during Cold Stress and Recovery from Chill Coma in Adult Drosophila Melanogaster. FEBS J 2010, 277, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.M.; Aquilina, J.A. Evidence for Specific Subunit Distribution and Interactions in the Quaternary Structure of A-crystallin. Proteins: Structure, Function, and Bioinformatics 2010, 78, 2546–2553. [Google Scholar] [CrossRef]
- Sun, W.; Montagu, M. Van; Verbruggen, N. Small Heat Shock Proteins and Stress Toler.Pdf. Plant Physiology and Biochemistry 2002, 1577, 1–9. [Google Scholar]
- Miernyk, J.A. Protein Folding in the Plant Cell. Plant Physiol 1999, 121, 695–703. [Google Scholar] [CrossRef]
- Sun, Y.; MacRae, T.H. Small Heat Shock Proteins: Molecular Structure and Chaperone Function. Cellular and Molecular Life Sciences 2005, 62, 2460–2476. [Google Scholar] [CrossRef]
- Eisenhardt, B.D. Small Heat Shock Proteins: Recent Developments. Biomol Concepts 2013, 4, 583–595. [Google Scholar] [CrossRef]
- Tsvetkova, N.M.; Horváth, I.; Török, Z.; Wolkers, W.F.; Balogi, Z.; Shigapova, N.; Crowe, L.M.; Tablin, F.; Vierling, E.; Crowe, J.H.; et al. Small Heat-Shock Proteins Regulate Membrane Lipid Polymorphism. Proc Natl Acad Sci U S A 2002, 99, 13504–13509. [Google Scholar] [CrossRef]
- Gkouvitsas, T.; Kontogiannatos, D.; Kourti, A. Differential Expression of Two Small Hsps during Diapause in the Corn Stalk Borer Sesamia Nonagrioides (Lef. ). J Insect Physiol 2008, 54, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Morrow, G.; Battistini, S.; Zhang, P.; Tanguay, R.M. Decreased Lifespan in the Absence of Expression of the Mitochondrial Small Heat Shock Protein Hsp22 in Drosophila. J Biol Chem 2004, 279, 43382–43385. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Bossier, P.; Wang, X.; Bojikova-Fournier, S.; MacRae, T.H. Diversity, Structure, and Expression of the Gene for P26, a Small Heat Shock Protein from Artemia. Genomics 2006, 88, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Willsie, J.K.; Clegg, J.S. Nuclear P26, a Small Heat Shock/α-Crystallin Protein, and Its Relationship to Stress Resistance in Artemia Franciscana Embryos. Journal of Experimental Biology 2001, 204, 2339–2350. [Google Scholar] [CrossRef]
- Willsie, J.K.; Clegg, J.S. Small Heat Shock Protein P26 Associates with Nuclear Lamins and HSP70 in Nuclei and Nuclear Matrix Fractions from Stressed Cells. J Cell Biochem 2002, 84, 601–614. [Google Scholar] [CrossRef]
- Hibshman, J.D.; Carra, S.; Goldstein, B. Tardigrade Small Heat Shock Proteins Can Limit Desiccation-Induced Protein Aggregation. Commun Biol 2023, 6, 121. [Google Scholar] [CrossRef]
- Villeneuve, T.S.; Ma, X.; Sun, Y.; Oulton, M.M.; Oliver, A.E.; MacRae, T.H. Inhibition of Apoptosis by P26: Implications for Small Heat Shock Protein Function during Artemia Development. Cell Stress Chaperones 2006, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol Biol Evol 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.-I.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform.
- Tissières, A.; Mitchell, H.K.; Tracy, U.M. Protein Synthesis in Salivary Glands of Drosophila Melanogaster: Relation to Chromosome Puffs. J Mol Biol 1974, 84, 389–398. [Google Scholar] [CrossRef]
- Russnak, R.H.; Peter, E.; Candido, M. Nucleic Acids Research Aoning and Analysis of CDNA Sequences Coding for Two 16 Kilodalton Heat Shock Proteins (Hsps) in Caenorhabditis Elegans: Homology with the Small Hsps of Drosophila; 1983; Vol. 1;
- Liang, P.; MacRae, T.H. The Synthesis of a Small Heat Shock/α-Crystallin Protein in Artemia and Its Relationship to Stress Tolerance during Development. Dev Biol 1999, 207, 445–456. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y. Identification of Multiple Small Heat-Shock Protein Genes in Plutella Xylostella (L.) and Their Expression Profiles in Response to Abiotic Stresses. Cell Stress Chaperones 2015, 20, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Flis, Ł.; Malewski, T.; Dobosz, R. Temperature Effects on Expression Levels of Hsp Genes in Eggs and Second-Stage Juveniles of Meloidogyne Hapla Chitwood, 1949. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, M.; Luo, X.; Zhang, Z.; Hu, X. Molecular Characterization of Heat Shock Protein 20 (Hsp20) in Goldfish (Carassius Auratus) and Expression Analysis in Response to Environmental Stresses. Aquac Rep 2022, 24, 101106. [Google Scholar] [CrossRef]
- Denlinger, D.L.; Rinehart, J.P.; Yocum, G.D. Stress Proteins. In Insect Timing: Circadian Rhythmicity to Seasonality; Elsevier, 2001; pp. 155–171 ISBN 978-0-444-50608-5.
- Wang, F.; Li, D.; Chen, Q.; Ma, L. Genome-Wide Survey and Characterization of the Small Heat Shock Protein Gene Family in Bursaphelenchus Xylophilus. Gene 2016, 579, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in Global Climate Change Research: Direct Effects of Rising Temperature on Insect Herbivores. Glob Chang Biol 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Abdullah, M. BEHAVIOURAL EFFECTS OF TEMPERATURE ON INSECTS.; 1961.
- Gilbert, N.; Raworth, D.A. FORUM: INSECTS AND TEMPERATURE—A GENERAL THEORY. Can Entomol 1996, 128, 1–13. [Google Scholar] [CrossRef]
- Li, M.; Wei, X.M.; Li, J.; Wei, S.M.; Zhang, J.L.; Chen, G.H.; Zhang, X.M. Effect of Short-Term Exposure to High Temperatures on the Reproductive Behavior and Physiological Enzyme Activities in the Fruit Fly Zeugodacus Tau (Walker). Front Physiol 2023, 14, 1036397. [Google Scholar] [CrossRef]
- Huang, L.-H.; Chen, B.; Kang, L. Impact of Mild Temperature Hardening on Thermotolerance, Fecundity, and Hsp Gene Expression in Liriomyza Huidobrensis. J Insect Physiol 2007, 53, 1199–1205. [Google Scholar] [CrossRef]
- Dahlgaard, J.; Loeschcke, V.; Michalak, P.; Justesen, J. Induced Thermotolerance and Associated Expression of the Heat-shock Protein Hsp70 in Adult Drosophila Melanogaster. Funct Ecol 1998, 12, 786–793. [Google Scholar] [CrossRef]
- Zhang, G.; Storey, J.M.; Storey, K.B. Chaperone Proteins and Winter Survival by a Freeze Tolerant Insect. J Insect Physiol 2011, 57, 1115–1122. [Google Scholar] [CrossRef]
- Chown, S.L.; Terblanche, J.S. Physiological Diversity in Insects: Ecological and Evolutionary Contexts. Adv In Insect Phys 2006, 33, 50–152. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E. A Primer on Insect Cold-Tolerance. In Low Temperature Biology of Insects; Denlinger, D.L., Lee Richard, E., J., *!!! REPLACE !!!*, Eds.; Cambridge University Press: Cambridge, 2010; ISBN 9780521886352. [Google Scholar]
- Atungulu, E.; Tanaka, H.; Fujita, K.; Yamamoto, K.; Sakata, M.; Sato, E.; Hara, M.; Yamashita, T.; Suzuki, K. A Double Chaperone Function of the SHsp Genes against Heat-Based Environmental Adversity in the Soil-Dwelling Leaf Beetles. J Insect Biotechnol Sericology 2006, 75, 15–22. [Google Scholar]
- Hadley, N.F. Water Relations of Terrestrial Arthropods.; 1994.
- Hansen, J.M.; Go, Y.-M.; Jones, D.P. Nuclear and Mitochondrial Compartmentation of Oxidative Stress and Redox Signaling. Annu Rev Pharmacol Toxicol 2006, 46, 215–234. [Google Scholar] [CrossRef]
- Lopez-Martinez, G.; Elnitsky, M.A.; Benoit, J.B.; Lee, R.E.; Denlinger, D.L. High Resistance to Oxidative Damage in the Antarctic Midge Belgica Antarctica, and Developmentally Linked Expression of Genes Encoding Superoxide Dismutase, Catalase and Heat Shock Proteins. Insect Biochem Mol Biol 2008, 38, 796–804. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide Decline of the Entomofauna: A Review of Its Drivers. Biol Conserv 2019, 232, 8–27. [Google Scholar] [CrossRef]
- van Klink, R.; Bowler, D.E.; Gongalsky, K.B.; Swengel, A.B.; Gentile, A.; Chase, J.M. Meta-Analysis Reveals Declines in Terrestrial but Increases in Freshwater Insect Abundances. Science 2020, 368, 417–420. [Google Scholar] [CrossRef]
- Kannan, K.; Jain, S. Oxidative Stress and Apoptosis. Pathophysiology 2000, 7, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Martelli, F.; Zhongyuan, Z.; Wang, J.; Wong, C.-O.; Karagas, N.E.; Roessner, U.; Rupasinghe, T.; Venkatachalam, K.; Perry, T.; Bellen, H.J.; et al. Low Doses of the Neonicotinoid Insecticide Imidacloprid Induce ROS Triggering Neurological and Metabolic Impairments in Drosophila. Proc Natl Acad Sci U S A 2020, 117, 25840–25850. [Google Scholar] [CrossRef] [PubMed]
- Reichmann, D.; Voth, W.; Jakob, U. Maintaining a Healthy Proteome during Oxidative Stress. Mol Cell 2018, 69, 203–213. [Google Scholar] [CrossRef]
- Li, M.; Tang, T.; Yuan, F.; Zhang, Y.; Li, F.; Liu, F. Protective Effects of Small Heat Shock Proteins in Daphnia Magna against Heavy Metal Exposure. Science of The Total Environment 2022, 848, 157565. [Google Scholar] [CrossRef]
- Cheng, S.; Lin, R.; Wang, L.; Qiu, Q.; Qu, M.; Ren, X.; Zong, F.; Jiang, H.; Yu, C. Comparative Susceptibility of Thirteen Selected Pesticides to Three Different Insect Egg Parasitoid Trichogramma Species. Ecotoxicol Environ Saf 2018, 166, 86–91. [Google Scholar] [CrossRef] [PubMed]
- D’Ávila, V.A.; Barbosa, W.F.; Guedes, R.N.C.; Cutler, G.C. Effects of Spinosad, Imidacloprid, and Lambda-Cyhalothrin on Survival, Parasitism, and Reproduction of the Aphid Parasitoid Aphidius Colemani. J Econ Entomol 2018, 111, 1096–1103. [Google Scholar] [CrossRef]
- Baglan, H.; Lazzari, C.R.; Guerrieri, F.J. Glyphosate Impairs Learning in Aedes Aegypti Mosquito Larvae at Field-Realistic Doses. J Exp Biol 2018, 221, jeb187518. [Google Scholar] [CrossRef]
- Rajak, P.; Roy, S. Heat Shock Proteins and Pesticide Stress. In Regulation of Heat Shock Protein Responses; Asea, A.A.A., Kaur, P., Eds.; Springer International Publishing: Cham, 2018; ISBN 978-3-319-74714-9. [Google Scholar]
- Otaka, M.; Odashima, M.; Watanabe, S. Role of Heat Shock Proteins (Molecular Chaperones) in Intestinal Mucosal Protection. Biochem Biophys Res Commun 2006, 348, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Somensi, N.; Brum, P.O.; de Miranda Ramos, V.; Gasparotto, J.; Zanotto-Filho, A.; Rostirolla, D.C.; da Silva Morrone, M.; Moreira, J.C.F.; Pens Gelain, D. Extracellular HSP70 Activates ERK1/2, NF-KB and Pro-Inflammatory Gene Transcription Through Binding with RAGE in A549 Human Lung Cancer Cells. Cell Physiol Biochem 2017, 42, 2507–2522. [Google Scholar] [CrossRef]
- Yu, L.-M.; Zhang, W.-H.; Han, X.-X.; Li, Y.-Y.; Lu, Y.; Pan, J.; Mao, J.-Q.; Zhu, L.-Y.; Deng, J.-J.; Huang, W.; et al. Hypoxia-Induced ROS Contribute to Myoblast Pyroptosis during Obstructive Sleep Apnea via the NF- κ B/HIF-1 α Signaling Pathway. Oxid Med Cell Longev 2019, 2019, 1–19. [Google Scholar] [CrossRef]
- Shan, Y.; Yan, S.; Hong, X.; Zha, J.; Qin, J. Effect of Imidacloprid on the Behavior, Antioxidant System, Multixenobiotic Resistance, and Histopathology of Asian Freshwater Clams (Corbicula Fluminea). Aquatic Toxicology 2020, 218, 105333. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhang, Y.; Niu, X.; Sun, Y.; Wang, H.; Guo, X.; Xu, B.; Wang, C. AccsHSP21. 7 Enhances the Antioxidant Capacity of Apis Cerana Cerana. J Sci Food Agric 2023, 103, 5401–5411. [Google Scholar] [CrossRef]
- Wang, X.X.; Geng, S.L.; Zhang, X.S.; Xu, W.H. P-S6K Is Associated with Insect Diapause via the ROS/AKT/ S6K/CREB/HIF-1 Pathway in the Cotton Bollworm, Helicoverpa Armigera. Insect Biochem Mol Biol 2020, 120. [Google Scholar] [CrossRef]
- Brunquell, J.; Morris, S.; Lu, Y.; Cheng, F.; Westerheide, S.D. The Genome-Wide Role of HSF-1 in the Regulation of Gene Expression in Caenorhabditis Elegans. BMC Genomics 2016, 17, 559. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, Mechanism and Health Effects of Some Heavy Metals. Interdiscip Toxicol 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Valko, M. Advances in Metal-Induced Oxidative Stress and Human Disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef]
- Hartwig, A.; Asmuss, M.; Blessing, H.; Hoffmann, S.; Jahnke, G.; Khandelwal, S.; Pelzer, A.; Bürkle, A. Interference by Toxic Metal Ions with Zinc-Dependent Proteins Involved in Maintaining Genomic Stability. Food Chem Toxicol 2002, 40, 1179–1184. [Google Scholar] [CrossRef]
- Martín-Folgar, R.; Martínez-Guitarte, J.-L. Cadmium Alters the Expression of Small Heat Shock Protein Genes in the Aquatic Midge Chironomus Riparius. Chemosphere 2017, 169, 485–492. [Google Scholar] [CrossRef]
- Morales, M.; Planelló, R.; Martínez-Paz, P.; Herrero, O.; Cortés, E.; Martínez-Guitarte, J.L.; Morcillo, G. Characterization of Hsp70 Gene in Chironomus Riparius: Expression in Response to Endocrine Disrupting Pollutants as a Marker of Ecotoxicological Stress. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2011, 153, 150–158. [Google Scholar] [CrossRef]
- Hoback, W.W.; Stanley, D.W. Insects in Hypoxia. J Insect Physiol 2001, 47, 533–542. [Google Scholar] [CrossRef]
- Liu, G.; Roy, J.; Johnson, E.A. Identification and Function of Hypoxia-Response Genes in Drosophila Melanogaster. Physiol Genomics 2006, 25, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Michaud, M.R.; Teets, N.M.; Peyton, J.T.; Blobner, B.M.; Denlinger, D.L. Heat Shock Response to Hypoxia and Its Attenuation during Recovery in the Flesh Fly, Sarcophaga Crassipalpis. J Insect Physiol 2011, 57, 203–210. [Google Scholar] [CrossRef]
- Basson, C.H.; Terblanche, J.S. Metabolic Responses of Glossina Pallidipes (Diptera: Glossinidae) Puparia Exposed to Oxygen and Temperature Variation: Implications for Population Dynamics and Subterranean Life. J Insect Physiol 2010, 56, 1789–1797. [Google Scholar] [CrossRef]
- Wang, H. -S.; Wang, X. -H.; Zhou, C. -S.; Huang, L. -H.; Zhang, S. -F.; Guo, W.; Kang, L. CDNA Cloning of Heat Shock Proteins and Their Expression in the Two Phases of the Migratory Locust. Insect Mol Biol 2007, 16, 207–219. [Google Scholar] [CrossRef]
- ALBRECHT, F.O.; VERDIER, M.; BLACKITH, R.E. Maternal Control of Ovariole Number in the Progeny of the Migratory Locust. Nature 1959, 184, 103–104. [Google Scholar] [CrossRef]
- Parsell, D.A.; Lindquist, S. The Function of Heat-Shock Proteins in Stress Tolerance: Degradation and Reactivation of Damaged Proteins. Annu Rev Genet 1993, 27, 437–496. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Jiang, D.; Yan, M.; Li, Y.; Wang, J.; Wu, F.; Sheng, S. Identifications, Characteristics, and Expression Patterns of Small Heat Shock Protein Genes in a Major Mulberry Pest, Glyphodes Pyloalis (Lepidoptera: Pyralidae). Journal of Insect Science 2020, 20. [Google Scholar] [CrossRef]
- Xie, J.; Hu, X.-X.; Zhai, M.-F.; Yu, X.-J.; Song, X.-W.; Gao, S.-S.; Wu, W.; Li, B. Characterization and Functional Analysis of Hsp18.3 Gene in the Red Flour Beetle, Tribolium Castaneum. Insect Sci 2019, 26, 263–273. [Google Scholar] [CrossRef]
- Hao, X.; Zhang, S.; Timakov, B.; Zhang, P. The Hsp27 Gene Is Not Required for Drosophila Development but Its Activity Is Associated with Starvation Resistance. Cell Stress Chaperones 2007, 12, 364. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Denlinger, D.L.; Rivers, D.B. Upregulation of Transcripts Encoding Select Heat Shock Proteins in the Flesh Fly Sarcophaga Crassipalpis in Response to Venom from the Ectoparasitoid Wasp Nasonia Vitripennis. J Invertebr Pathol 2002, 79, 62–63. [Google Scholar] [CrossRef]
- Zhu, J.-Y.; Wu, G.-X.; Ye, G.-Y.; Hu, C. Heat Shock Protein Genes (Hsp20, Hsp75 and Hsp90) from Pieris Rapae: Molecular Cloning and Transcription in Response to Parasitization by Pteromalus Puparum. Insect Sci 2013, 20, 183–193. [Google Scholar] [CrossRef]
- Shim, J.-K.; Ha, D.-M.; Nho, S.-K.; Song, K.-S.; Lee, K.-Y. Upregulation of Heat Shock Protein Genes by Envenomation of Ectoparasitoid Bracon Hebetor in Larval Host of Indian Meal Moth Plodia Interpunctella. J Invertebr Pathol 2008, 97, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Sage, E.; Douki, T. Ultraviolet Radiation-Mediated Damage to Cellular DNA. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2005, 571, 3–17. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B.; Kashiwagi, T.; Eguchi, E. Selective Photoreceptor Damage in Four Species of Insects Induced by Experimental Exposures to UV-Irradiation. Micron 2002, 33, 23–31. [Google Scholar] [CrossRef]
- Meng, J.-Y.; Zhang, C.-Y.; Zhu, F.; Wang, X.-P.; Lei, C.-L. Ultraviolet Light-Induced Oxidative Stress: Effects on Antioxidant Response of Helicoverpa Armigera Adults. J Insect Physiol 2009, 55, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-L.; Meng, J.-Y.; Zhou, L.; Yao, M.-S.; Zhang, C.-Y. Identification of Five Small Heat Shock Protein Genes in Spodoptera Frugiperda and Expression Analysis in Response to Different Environmental Stressors. Cell Stress Chaperones 2021, 26, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Guo, X.; Li, Y.; Gao, H.; Guo, X.; Xu, B. SHsp22.6, an Intronless Small Heat Shock Protein Gene, Is Involved in Stress Defence and Development in Apis Cerana Cerana. Insect Biochem Mol Biol 2014, 53, 1–12. [Google Scholar] [CrossRef]
- Liu, Z.; Xi, D.; Kang, M.; Guo, X.; Xu, B. Molecular Cloning and Characterization of Hsp27.6: The First Reported Small Heat Shock Protein from Apis Cerana Cerana. Cell Stress Chaperones 2012, 17, 539–551. [Google Scholar] [CrossRef]
- Xie, J.; Xiong, W.; Hu, X.; Gu, S.; Zhang, S.; Gao, S.; Song, X.; Bi, J.; Li, B. Characterization and Functional Analysis of Hsp21.8b: An Orthologous Small Heat Shock Protein Gene in Tribolium Castaneum. Journal of Applied Entomology 2018, 142, 654–666. [Google Scholar] [CrossRef]
- Sang, W.; Ma, W.-H.; Qiu, L.; Zhu, Z.-H.; Lei, C.-L. The Involvement of Heat Shock Protein and Cytochrome P450 Genes in Response to UV-A Exposure in the Beetle Tribolium Castaneum. J Insect Physiol 2012, 58, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Cartaño, N. V; Milos, L.; Krebs, R.A.; Lindquist, S.L. Effect of Engineering Hsp70 Copy Number on Hsp70 Expression and Tolerance of Ecologically Relevant Heat Shock in Larvae and Pupae of Drosophila Melanogaster. J Exp Biol 1996, 199, 1837–1844. [Google Scholar] [CrossRef]
- Feder, J.H.; Rossi, J.M.; Solomon, J.; Solomon, N.; Lindquist, S. The Consequences of Expressing Hsp70 in Drosophila Cells at Normal Temperatures. Genes Dev 1992, 6, 1402–1413. [Google Scholar] [CrossRef]
- Pener, M.P. Locust Phase Polymorphism and Its Endocrine Relations. In; 1991; pp. 1–79.
- Wang, H.; Dong, S.-Z.; Li, K.; Hu, C.; Ye, G. A Heat Shock Cognate 70 Gene in the Endoparasitoid, Pteromalus Puparum, and Its Expression in Relation to Thermal Stress. BMB Rep 2008, 41, 388–393. [Google Scholar] [CrossRef]
- Huang, L.-H.; Wang, C.-Z.; Kang, L. Cloning and Expression of Five Heat Shock Protein Genes in Relation to Cold Hardening and Development in the Leafminer, Liriomyza Sativa. J Insect Physiol 2009, 55, 279–285. [Google Scholar] [CrossRef]
- Sakano, D.; Li, B.; Xia, Q.; Yamamoto, K.; Banno, Y.; Fujii, H.; Aso, Y. Genes Encoding Small Heat Shock Proteins of the Silkworm, Bombyx Mori. Biosci Biotechnol Biochem 2006, 70, 2443–2450. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Moghaddam, S.H.H.; Du, X.; Zhong, B.-X.; Chen, Y.-Y. Comparative Analysis on the Expression of Inducible HSPs in the Silkworm, Bombyx Mori. Mol Biol Rep 2012, 39, 3915–3923. [Google Scholar] [CrossRef] [PubMed]
- Colinet, H.; Lee, S.F.; Hoffmann, A. Knocking down Expression of Hsp22 and Hsp23 by RNA Interference Affects Recovery from Chill Coma in Drosophila Melanogaster. Journal of Experimental Biology 2010, 213, 4146–4150. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Li, A.; Yocum, G.D.; Robich, R.M.; Hayward, S.A.L.; Denlinger, D.L. Up-Regulation of Heat Shock Proteins Is Essential for Cold Survival during Insect Diapause. Proceedings of the National Academy of Sciences 2007, 104, 11130–11137. [Google Scholar] [CrossRef]
- Yuan, J.-W.; Song, H.-X.; Chang, Y.-W.; Yang, F.; Xie, H.-F.; Gong, W.-R.; Du, Y.-Z. Identification, Expression Analysis and Functional Verification of Two Genes Encoding Small Heat Shock Proteins in the Western Flower Thrips, Frankliniella Occidentalis (Pergande). Int J Biol Macromol 2022, 211, 74–84. [Google Scholar] [CrossRef]
- Dong, C.-L.; Zhu, F.; Lu, M.-X.; Du, Y.-Z. Characterization and Functional Analysis of Cshsp19. 0 Encoding a Small Heat Shock Protein in Chilo Suppressalis (Walker). Int J Biol Macromol 2021, 188, 924–931. [Google Scholar] [CrossRef]
- Samanta, S.; Barman, M.; Chakraborty, S.; Banerjee, A.; Tarafdar, J. Involvement of Small Heat Shock Proteins (SHsps) in Developmental Stages of Fall Armyworm, Spodoptera Frugiperda and Its Expression Pattern under Abiotic Stress Condition. Heliyon 2021, 7, e06906. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Zhang, J.; Guo, Y.; Ma, E. Molecular Cloning and MRNA Expression of Heat Shock Protein Genes and Their Response to Cadmium Stress in the Grasshopper Oxya Chinensis. PLoS One 2015, 10, e0131244. [Google Scholar] [CrossRef] [PubMed]
- Chapuis, M.-P.; Simpson, S.J.; Blondin, L.; Sword, G.A. Taxa-Specific Heat Shock Proteins Are over-Expressed with Crowding in the Australian Plague Locust. J Insect Physiol 2011, 57, 1562–1567. [Google Scholar] [CrossRef]
- Tian, L.; Wang, X.; Wang, X.; Lei, C.; Zhu, F. Starvation-, Thermal- and Heavy Metal- Associated Expression of Four Small Heat Shock Protein Genes in Musca Domestica. Gene 2018, 642, 268–276. [Google Scholar] [CrossRef]
- Wang, H.; Li, K.; Zhu, J.-Y.; Fang, Q.; Ye, G.-Y.; Wang, H.; Li, K.; Zhu, J.-Y. Cloning and Expression Pattern of Heat Shock Protein Genes from the Endoparasitoid Wasp, Pteromalus Puparum in Response to Environmental Stresses. Arch Insect Biochem Physiol 2012, 79, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Kayastha, P.; Wieczorkiewicz, F.; Pujol, M.; Robinson, A.; Michalak, M.; Kaczmarek, Ł.; Poprawa, I. Elevated External Temperature Affects Cell Ultrastructure and Heat Shock Proteins (HSPs) in Paramacrobiotus Experimentalis Kaczmarek, Mioduchowska, Poprawa, & Roszkowska, 2020. Sci Rep 2024, 14. [Google Scholar] [CrossRef]
- Kyriakou, E.; Taouktsi, E.; Syntichaki, P. The Thermal Stress Coping Network of the Nematode Caenorhabditis Elegans. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Zevian, S.C.; Yanowitz, J.L. Methodological Considerations for Heat Shock of the Nematode Caenorhabditis Elegans. Methods 2014, 68, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Strayer, A.; Wu, Z.; Christen, Y.; Link, C.D.; Luo, Y. Expression of the Small Heat-shock Protein Hsp-16-2 in Caenorhabditis Elegans Is Suppressed by Ginkgo Biloba Extract EGb 761. The FASEB Journal 2003, 17, 2305–2307. [Google Scholar] [CrossRef]
- Jecock, R.M.; Devaney, E. Expression of Small Heat Shock Proteins by the Third-Stage Larva of Brugia Pahangi; 1992; Vol. 56;
- Younis, A.E.; Geisinger, F.; Ajonina-Ekoti, I.; Soblik, H.; Steen, H.; Mitreva, M.; Erttmann, K.D.; Perbandt, M.; Liebau, E.; Brattig, N.W. Stage-Specific Excretory-Secretory Small Heat Shock Proteins from the Parasitic Nematode Strongyloides Ratti - Putative Links to Host’s Intestinal Mucosal Defense System. FEBS Journal 2011, 278, 3319–3336. [Google Scholar] [CrossRef]
- Fonseca, F.; Pénicaud, C.; Tymczyszyn, E.E.; Gómez-Zavaglia, A.; Passot, S. Factors Influencing the Membrane Fluidity and the Impact on Production of Lactic Acid Bacteria Starters. Appl Microbiol Biotechnol 2019, 103, 6867–6883. [Google Scholar] [CrossRef]
- Sadura, I.; Janeczko, A. Are Heat Shock Proteins Important in Low-Temperature-Stressed Plants? A Minireview. Agronomy 2024, 14. [Google Scholar] [CrossRef]
- Peter, E.; Candido, M. Caenorhabditis Elegans: Structure, Regulation and Biology.
- Horikawa, M.; Fukuyama, M.; Antebi, A.; Mizunuma, M. Regulatory Mechanism of Cold-Inducible Diapause in Caenorhabditis Elegans. Nat Commun 2024, 15. [Google Scholar] [CrossRef]
- Thorne, M.A.S.; Kagoshima, H.; Clark, M.S.; Marshall, C.J.; Wharton, D.A. Molecular Analysis of the Cold Tolerant Antarctic Nematode, Panagrolaimus Davidi. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Adhikari, B.N.; Wall, D.H.; Adams, B.J. Effect of Slow Desiccation and Freezing on Gene Transcription and Stress Survival of an Antarctic Nematode. Journal of Experimental Biology 2010, 213, 1803–1812. [Google Scholar] [CrossRef] [PubMed]
- Erkut, C.; Vasilj, A.; Boland, S.; Habermann, B.; Shevchenko, A.; Kurzchalia, T. V Molecular Strategies of the Caenorhabditis Elegans Dauer Larva to Survive Extreme Desiccation. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Tyson, T.; O’Mahony Zamora, G.; Wong, S.; Skelton, M.; Daly, B.; Jones, J.T.; Mulvihill, E.D.; Elsworth, B.; Phillips, M.; Blaxter, M.; et al. A Molecular Analysis of Desiccation Tolerance Mechanisms in the Anhydrobiotic Nematode Panagrolaimus Superbus Using Expressed Sequenced Tags. BMC Res Notes 2012, 5. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, J. Hypoxia-Inducible Factor-1 (HIF-1)-Independent Hypoxia Response of the Small Heat Shock Protein Hsp-16.1 Gene Regulated by Chromatin-Remodeling Factors in the Nematode Caenorhabditis Elegans. Journal of Biological Chemistry 2013, 288, 1582–1589. [Google Scholar] [CrossRef]
- Balakumaran, M.; Chidambaranathan, P.; Tej Kumar, J.P.J.P.; Sirohi, A.; Kumar Jain, P.; Gaikwad, K.; Iyyappan, Y.; Rao, A.R.; Sahu, S.; Dahuja, A.; et al. Deciphering the Mechanism of Anhydrobiosis in the Entomopathogenic Nematode Heterorhabditis Indica through Comparative Transcriptomics. PLoS One 2022, 17. [Google Scholar] [CrossRef]
- Hong, M.; Kwon, J.Y.; Shim, J.; Lee, J. Differential Hypoxia Response of Hsp-16 Genes in the Nematode. J Mol Biol 2004, 344, 369–381. [Google Scholar] [CrossRef]
- Kim, B.M.; Rhee, J.S.; Jeong, C.B.; Seo, J.S.; Park, G.S.; Lee, Y.M.; Lee, J.S. Heavy Metals Induce Oxidative Stress and Trigger Oxidative Stress-Mediated Heat Shock Protein (Hsp) Modulation in the Intertidal Copepod Tigriopus Japonicus. Comparative Biochemistry and Physiology Part - C: Toxicology and Pharmacology 2014, 166, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive Oxygen Species, Toxicity, Oxidative Stress, and Antioxidants: Chronic Diseases and Aging. Arch Toxicol 2023, 97, 2499–2574. [Google Scholar]
- Sampayo, J.N.; Olsen, A.; Lithgow, G.J. Oxidative Stress in Caenorhabditis Elegans: Protective Effects of Superoxide Dismutase/Catalase Mimetics. Aging Cell 2003, 2, 319–326. [Google Scholar] [CrossRef]
- Ezemaduka, A.N.; Wang, Y.; Li, X. Expression of CeHSP17 Protein in Response to Heat Shock and Heavy Metal Ions; 2017; Vol. 49;
- Shashikumar, S.; Rajini, P.S. Cypermethrin Elicited Responses in Heat Shock Protein and Feeding in Caenorhabditis Elegans. Ecotoxicol Environ Saf 2010, 73, 1057–1062. [Google Scholar] [CrossRef]
- Sonone, S.S.; Jadhav, S.; Sankhla, M.S.; Kumar, R. Water Contamination by Heavy Metals and Their Toxic Effect on Aquaculture and Human Health through Food Chain. Letters in Applied NanoBioScience 2021, 10, 2148–2166. [Google Scholar] [CrossRef]
- Gaur, V.K.; Sharma, P.; Sirohi, R.; Awasthi, M.K.; Dussap, C.G.; Pandey, A. Assessing the Impact of Industrial Waste on Environment and Mitigation Strategies: A Comprehensive Review. J Hazard Mater 2020, 398, 123019. [Google Scholar] [CrossRef]
- Khan, Z.; Elahi, A.; Bukhari, D.A.; Rehman, A. Cadmium Sources, Toxicity, Resistance and Removal by Microorganisms-A Potential Strategy for Cadmium Eradication. Journal of Saudi Chemical Society 2022, 26, 101569. [Google Scholar] [CrossRef]
- Moiseenko, T.I.; Gashkina, N.A. Distribution and Bioaccumulation of Heavy Metals (Hg, Cd and Pb) in Fish: Influence of the Aquatic Environment and Climate. Environmental Research Letters 2020, 15. [Google Scholar] [CrossRef]
- Das, S.; Kar, I.; Patra, A.K. Cadmium Induced Bioaccumulation, Histopathology, Gene Regulation in Fish and Its Amelioration – A Review. Journal of Trace Elements in Medicine and Biology 2023, 79, 127202. [Google Scholar] [CrossRef]
- Tamás, L.; Mistrík, I.; Zelinová, V. Heavy Metal-Induced Reactive Oxygen Species and Cell Death in Barley Root Tip. Environ Exp Bot 2017, 140, 34–40. [Google Scholar] [CrossRef]
- Georgiadou, E.C.; Kowalska, E.; Patla, K.; Kulbat, K.; Smolińska, B.; Leszczyńska, J.; Fotopoulos, V. Influence of Heavy Metals (Ni, Cu, and Zn) on Nitro-Oxidative Stress Responses, Proteome Regulation and Allergen Production in Basil (Ocimum Basilicum L.) Plants. Front Plant Sci 2018, 9. [Google Scholar] [CrossRef]
- Tamás, M.J.; Sharma, S.K.; Ibstedt, S.; Jacobson, T.; Christen, P. Heavy Metals and Metalloids as a Cause for Protein Misfolding and Aggregation. Biomolecules 2014, 4, 252–267. [Google Scholar] [CrossRef]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic Mechanisms of Five Heavy Metals: Mercury, Lead, Chromium, Cadmium, and Arsenic. Front Pharmacol 2021, 12, 1–19. [Google Scholar] [CrossRef]
- Yuan, F.; Yang, Z.; Tang, T.; Xie, S.; Liu, F. A 28.6-KD Small Heat Shock Protein (MnHSP28.6) Protects Macrobrachium Nipponense against Heavy Metal Toxicity and Oxidative Stress by Virtue of Its Anti-Aggregation Activity. Fish Shellfish Immunol 2019, 95, 635–643. [Google Scholar] [CrossRef]
- Zhang, A.; Lu, Y.; Li, C.; Zhang, P.; Su, X.; Li, Y.; Wang, C.; Li, T. A Small Heat Shock Protein (SHSP) from Sinonovacula Constricta against Heavy Metals Stresses. Fish Shellfish Immunol 2013, 34, 1605–1610. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.-L.; Yao, C.-L.; Wang, Z.-Y. Acute Temperature and Cadmium Stress Response Characterization of Small Heat Shock Protein 27 in Large Yellow Croaker, Larimichthys Crocea. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2012, 155, 190–197. [Google Scholar] [CrossRef]
- Li, C.; Wang, L.; Ning, X.; Chen, A.; Zhang, L.; Qin, S.; Wu, H.; Zhao, J. Identification of Two Small Heat Shock Proteins with Different Response Profile to Cadmium and Pathogen Stresses in Venerupis Philippinarum. Cell Stress Chaperones 2010, 15, 897–904. [Google Scholar] [CrossRef]
- Li, H.; Liu, S.; He, C.; Gao, X.; Yuan, X. Identification of a Small HSP Gene from Hard Clam Meretrix Meretrix and Its Potential as an Environmental Stress Biomarker. Aquat Biol 2013, 18, 243–252. [Google Scholar] [CrossRef]
- Bildik, A.E.; Ekren, G.S.A.; Akdeniz, G.; Kiral, F.K. Effect of Enviromental Temperature on Heat Shock Proteins (HSP30, HSP70, HSP90) and IGF-I MRNA Expression in Sparus Aurata. Iran J Fish Sci 2019, 18, 1014–1024. [Google Scholar]
- Currie, S. The Effects of Heat Shock and Acclimation Temperature on Hsp70 and Hsp30 MRNA Expression in Rainbow Trout: In Vivo and in Vitro Comparisons. J Fish Biol 2000, 56, 398–408. [Google Scholar] [CrossRef]
- Liu, X.; Shi, H.; Liu, Z.; Kang, Y.; Wang, J.; Huang, J. Effect of Heat Stress on Heat Shock Protein 30 (Hsp 30) MRNA Expression in Rainbow Trout (Oncorhynchus Mykiss).; 2019.
- Shekhar, M.S.; Kiruthika, J.; Ponniah, A.G. Identification and Expression Analysis of Differentially Expressed Genes from Shrimp (Penaeus Monodon) in Response to Low Salinity Stress. Fish Shellfish Immunol 2013, 35, 1957–1968. [Google Scholar] [CrossRef] [PubMed]
- Zarei, S.; Ghafouri, H.; Vahdatiraad, L.; Heidari, B. The Influence of HSP Inducers on Salinity Stress in Sterlet Sturgeon (Acipenser Ruthenus): In Vitro Study on HSP Expression, Immune Responses, and Antioxidant Capacity. Cell Stress Chaperones 2024, 29, 552–566. [Google Scholar] [CrossRef]
- Muthusamy, S.K.; Dalal, M.; Chinnusamy, V.; Bansal, K.C. Genome-Wide Identification and Analysis of Biotic and Abiotic Stress Regulation of Small Heat Shock Protein (HSP20) Family Genes in Bread Wheat. J Plant Physiol 2017, 211, 100–113. [Google Scholar] [CrossRef]
- Arockiaraj, J.; Vanaraja, P.; Easwvaran, S.; Singh, A.; Othman, R.Y.; Bhassu, S. Gene Expression and Functional Studies of Small Heat Shock Protein 37 (MrHSP37) from Macrobrachium Rosenbergii Challenged with Infectious Hypodermal and Hematopoietic Necrosis Virus (IHHNV). Mol Biol Rep 2012, 39, 6671–6682. [Google Scholar] [CrossRef]
- Huang, P.; Kang, S.; Chen, W.; Hsu, T.; Lo, C.; Liu, K.; Chen, L. Identification of the Small Heat Shock Protein, HSP21, of Shrimp Penaeus Monodon and the Gene Expression of HSP21 Is Inactivated after White Spot Syndrome Virus (WSSV) Infection. Fish Shellfish Immunol 2008, 25, 250–257. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A) Effect of abiotic and biotic stress on insect, nematode, fish, molluscs, and crustacean health and small heat shock protein response. (B) The three-dimensional structure was established using the SWISS-MODEL program based on the p26 of Artemia sinica as a template.
Figure 1.
A) Effect of abiotic and biotic stress on insect, nematode, fish, molluscs, and crustacean health and small heat shock protein response. (B) The three-dimensional structure was established using the SWISS-MODEL program based on the p26 of Artemia sinica as a template.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



