
Submitted:
22 October 2024
Posted:
22 October 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Background/Objectives: Although the protective effects of zinc against COVID-19 are documented, its impact on the COVID-19 vaccine immunogenicity remains unknown. Methods: We conducted a prospective study involving a cohort of 79 Japanese individuals (aged 21–56 years; comprising three subcohorts) and measured their serum zinc levels pre-vaccination and anti-SARS-CoV-2 IgM/IgG levels pre- and post-vaccination over 4 months. Results: Serum zinc concentrations ranged 74–140 and 64–113 ug/dL in male and female individuals, respectively, with one male and 11 female participants exhibiting subclinical zinc deficiency (60–80 ug/dL). Mixed models for antibody titers, accounting for subcohorts, repeat measurements, and covariates (e.g., vaccine type, sex, age, height, steroid use, medical history, smoking and drinking habits, perceived stress, and sleep disturbances) showed positive effects of zinc on IgM (p = 0.012) and IgG (p = 0.013) in 45 female individuals with 255 observations. However, a similar association was not found in the 34 male participants with 162 observations. This discrepancy may be attributed to one participant being included in the subcohort with frequent repeat measurements (10 repeats in 4 months). COVID-19 mRNA vaccine immunogenicity was enhanced in the participants with high baseline blood zinc levels within the reference range. Conclusions: Our findings underscore the relevance of maintaining adequate zinc levels before vaccination, which can be achieved through a balanced diet and healthy lifestyle choices.
Keywords:
Zinc
; SARS-CoV-2
; Immunogenicity
; Humoral immune response
; Vaccination
1. Introduction
COVID-19 is a global health threat, highlighting the crucial role of the immune system in combating the SARS-CoV-2 infection. Recognized risk factors for severe infection include age, sex, and insufficient levels of micronutrients, including zinc [1,2,3,4]. Zinc, an essential trace element, plays a vital role in supporting both innate and adaptive immunity [5,6,7,8]. For instance, zinc is involved in the maturation of dendritic cells, activation of mast cells, and T cell maturation. Disruption of zinc homeostasis can profoundly affect the adaptive immune response [9], potentially impairing antibody production [10], which crucially affects the humoral immune response, also known as the antibody-mediated immune response [11]. Given its crucial role in immune function, it is essential to investigate the association between the baseline zinc levels and vaccine immunogenicity.
According to an observation of elderly individuals in nursing homes in the United States, normal baseline concentrations of blood zinc have been linked to lower overall mortality rates compared with low levels of zinc [12]. Several studies have reported low serum zinc levels in patients with COVID-19, which are associated with severe disease, death risk, and poor clinical outcomes [13,14,15]. Moreover, other studies have suggested that zinc supplementation could contribute to an additional defense against the illness [16,17,18], potentially by lowering the viral load and enhancing the immunity of patients with COVID-19. However, the association between the baseline levels of serum zinc and post-vaccination antibody titers has been scarcely studied. In an observational study of healthcare workers in Germany with a moderate level of baseline zinc who received two doses of the anti-SARS-CoV-2 vaccine (BNT162B2), the total serum zinc levels and supplementation showed no significant associations [19]. We believe that the results are inconclusive because of limitations, including participants’ ethnicity and confounding factors, such as psychological stress, sleep deprivation, and physical exercise, which could affect the zinc levels [20] and immune system function. Therefore, we performed a prospective study in the Japanese general population, measuring the serum zinc levels prior to vaccination and tracking anti-SARS-CoV-2 IgM and IgG levels before and for 4 months following vaccination. Our aim was to examine how baseline blood zinc levels affect the humoral immune response following COVID-19 mRNA vaccination.
2. Materials and Methods
This study was approved by the Ethics Committee for Clinical Research (No. R3-39) and the Ethical Review of Human Genome and Genetic Analysis Research (No. R3-4) at the School of Medicine, Saga University, Saga, Japan. Written informed consent was obtained from all participants prior to any study-related procedures.
2.1. Study Design and Participants
A total of 79 participants, comprising 11 healthcare workers, 68 students, and employees from hospitals and a university in Saga Prefecture, were enrolled in this study. Participants received either two doses of BNT162b2 (Pfizer Inc., New York, NY, USA/BioNTech SE, Mainz, Germany) at a dose of 30 µg (N = 53) or mRNA-1273 (Moderna Inc., Cambridge, MA, USA/Takeda Pharmaceutical Co., Ltd., Tokyo, Japan) at a dose of 100 µg (N = 26) (Figure 1). The total number of observations was 255 for 45 female and 162 for 34 male individuals. The first and second doses of BNT162b2 were administered between April and May 2021, with a 21-day interval between doses. The first and second doses of Moderna’s mRNA-1273 were administered in August 2021, with a 28-day interval between doses. None of the participants had a history of COVID-19 prior to vaccination. Blood samples were collected from all participants before the first vaccination (Week 0) and at various intervals up to Week 12 after the second vaccination. This included healthcare workers (Subcohort 1), university students (Subcohort 2), and university employees and students (Subcohort 3).
2.2. Laboratory Testing
Serum was extracted from the samples on the same day as blood collection and stored at -80°C until analysis. This study employed a hig-sensitivity chemiluminescent enzyme immunoassay (CLEIA) platform (Sysmex Co., Kobe, Japan) to measure three types of anti-SARS-CoV-2 antibodies: the S1 subunit of anti-spike protein (S1) IgG, anti-S1 IgM, and anti-nucleocapsid protein (N) IgG [21]. The units for anti-S1 IgG, IgM, and anti-N IgG were expressed as binding antibody units per mL (BAU/mL), Sysmex units per mL (SU/mL), and SU/mL, respectively. The BAU was calibrated according to the WHO International Standards. The total serum concentration of zinc was determined using a zinc quantification kit (Espa Zn II Kit; NIPRO, Tokyo, Japan), following the manufacturer's instructions.
2.3. Self-Administered Questionnaire
A self-administered questionnaire was used to collect information on sex, age, height, weight, smoking status, alcohol intake, exercise habits, perceived stress, sleep disturbance, and medical history. The participants’ smoking habits remained unchanged during the preceding year. Ethanol intake was calculated based on the amount of alcohol consumed in the previous six months, adjusted per 60 kg of body weight, and then categorized into four groups: <1 g/day, ≥1 g/day, <20 g/day, and ≥20 g/day. Exercise habits were assessed by asking, "Do you usually exercise?", with possible answers including: no habit, <1 day/week, 1–3 days/week, and ≥3 days/week. Perceived stress was evaluated on a 5-point scale, with responses ranging from no (0), mostly to (1), unsure (2), quite often (3), and yes (4). Sleep disturbance was classified as “yes” for positive indications and “no” for no indications. Participants receiving steroids at the time of the study were classified as "yes" for steroid use, while those who had not used steroids within the preceding 3 years were considered as "no." Dyslipidemia was similarly categorized: participants with concurrent dyslipidemia were marked as "yes," while those without a history of dyslipidemia in the previous three years were marked as "no."
2.4. Statistical Analyses
Statistical analyses were performed using SAS 9.4 TS Level 1M5 for Windows (SAS Institute, Cary, NC, USA) with an alpha value of 0.05. The primary outcomes, anti-S1-IgG (BAU/mL) and anti-S1-IgM (SU/mL) levels, along with the serum zinc level (μg/dL) as the primary explanatory variable, were log-transformed to achieve an approximate normal distribution. Male and female individuals were examined separately for the following reasons: 1) the estimation was possible for the whole observation period for 45 female individuals (255 observation points), while data for 34 male participants (162 observation points) were non-estimable at 1–3 weeks after the second dose; and 2) there was a significant difference in the zinc levels according to sex (Figure 2).
2.4.1. Covariates
In addition to sex, age, vaccine type, and weeks post-vaccination, the following covariates were included in the analysis: height, smoking status, ethanol intake, exercise habits, perceived stress, sleep disturbances, steroid use, and dyslipidemia. These attributes are associated with vaccine efficacy, immune response, and zinc levels, as reported in previous studies [23,24,25,26,27,28,29].
2.4.2. Mixed Model
Mixed model analysis was conducted to estimate log-transformed antibody titers, accounting for subcohorts, repeat measurements, and covariates in three models. Model 1 included the effects of post-vaccination week, vaccine type, age, and zinc levels. Model 2 included lifestyle habits and current medical history factors, such as body height, steroid use, dyslipidemia, allergic disease, exercise habits, cigarette smoking, ethanol intake, perceived stress, and sleep disturbance.
2.4.3. Least Squares Geometric Means
The non-linear dose-response relationships between zinc, anti-S1-IgG, and anti-S1-IgM are depicted as least-squares geometric means and standard errors for the categorized zinc levels. Anti-S1 IgG and IgM levels corresponding to T1–T3 groups are visualized using Model 1, with modification for zinc. The term "log-transformed zinc" was replaced by "T1–T3 × time course" to reflect the interaction between zinc levels (T1–T3 groups) and the time course.
2.4.4. Sensitivity Analysis
Among the 45 female participants, one with dyslipidemia was excluded from the sensitivity analysis, resulting in 44 participants with a total of 245 data points. When conducting additional analyses, we considered the body weight instead of height.
3. Results
3.1. Baseline Characteristics
During the observation period, all participants had anti-N IgG levels below 1.2 SU/m, indicating no previous exposure to the virus or vaccination. Table 1 shows the baseline characteristics of the 79 participants. The study group comprised 45 female (57%) and 34 male participants (43%) with an age range of 21–56 years. The baseline characteristics of the participants are presented in Table 1, which categorizes them into three quantiles (T) based on baseline serum zinc levels by sex. The distribution of exercise habits and perceived stress did not differ among the tertiles (female, p >0.6; male, p >0.8, Fisher’s exact test). Two male participants in the T3 group reported steroid use, whereas one female participant in the T1 group reported dyslipidemia.
3.2. Effect of Baseline Zinc on Anti-S1 IgM Post-Vaccination
Although no association between zinc and anti-S1 IgM was found in male participants, a positive effect was observed in female participants. The partial regression coefficient (β) for log-transformed zinc on log-transformed anti-S1 IgM (SU/mL) was 2.2 (p<0.001) and 1.5 (p = 0.012) in Models 1 and 2, respectively (Table 2). As shown in Figure 3 (left panel), anti-S1 IgM levels exhibited the most significant differences at 1–3 weeks after the second dose. Participants in the highest zinc tertile (T3; 93–113 μg/dL) displayed the highest anti-S1 IgM levels compared to those in the lower tertiles (T1 and T2; 64–92 μg/dL).
3.3. Effect of Baseline Zinc on Anti-S1 IgG Post-Vaccination
Similar to the IgM, the positive fixed effect was estimated in females with a β of 1.7 (p<0.001) in Model 1 (Table 3). The positive effect of zinc in female individuals remained after the inclusion of lifestyle habits and current medical history (β = 1.1, p = 0.013 in Model 2). Similar to the IgG response, anti-S1 IgG levels differed most significantly 1–3 weeks after the second dose (Figure 3). The highest zinc tertile displayed the highest anti-S1 IgG levels compared to those in the lower tertiles, suggesting a beneficial effect of higher pre-vaccination zinc levels on the post-vaccination humoral immune response.
3.4. Sensitivity Analysis
Sensitivity analysis, excluding one female participant with dyslipidemia (44 female individuals; 245 observations), resulted in a similar estimation (AIC = 659, β = 1.5, p = 0.013 in Model 2 for IgM, and AIC = 512, β = 1.1, p = 0.013 in Model 2 for IgG). A slightly modified Model 2, including weight instead of height, produced similar estimates (AIC = 681, β = 1.4, p = 0.018 in Model 2 for IgM, and AIC = 536, β = 1.1, p = 0.013 in Model 2 for IgG).
4. Discussion
Our prospective study investigated the impact of baseline blood zinc levels on the humoral immune response and production of anti-S1 IgG and IgM antibodies following COVID-19 mRNA vaccination in a Japanese cohort. Using mixed models considering repeated measures and covariates, we found higher post-vaccine IgG/IgM production in female participants with higher baseline zinc levels within the normal range, suggesting a positive effect of adequate zinc levels on optimal immune system activity.
Zinc may increase antibody titers via various pathways [30]. For example, zinc may increase the downstream signal interleukin-2 by maintaining the three-dimensional structure of NFκB, thereby increasing the production of IgG [31] and IgM [32]. Additionally, zinc is crucial for regulating T follicular helper (Tfh) cell production and plays a vital role in humoral immunity [9]. The zinc-dependent transcription factor, B-cell lymphoma 6 (BCL6), is particularly important, as it requires zinc for structural integrity to support B-cell maturation within germinal centers. BCL6 regulates B cell and Tfh cell interactions, facilitates antibody class switching, and promotes humoral immunity by upregulating key downstream proteins [33].
Our finding of a significant correlation between baseline zinc levels and antibody response to mRNA vaccination is inconsistent with that of a previous study conducted on a German population [19]. One of the advantages of our design over that of this study is the inclusion of multiple covariates; however, multivariate adjustment did not affect this association. The strong impact observed 1–3 weeks after the second dose in our study suggests that the frequent observation points played a crucial role in detecting significant associations. In contrast, the previous study lacked observations within the first month post-vaccination [19], whereas we conducted four observations within this period.
4.1. Limitations
This study had a relatively small sample size, particularly among male participants. Additionally, the significant findings were primarily observed within one month after the second vaccination, which may limit the generalizability of the results. Given the small sample size, these findings need to be validated in larger cohorts. However, with the widespread spread of COVID-19, conducting similar prospective studies in human populations may no longer be feasible. Future efforts should focus on leveraging remaining samples from existing cohorts and conducting confirmatory animal studies to further investigate the relationship between baseline zinc levels and the immune response to COVID-19 vaccination.
5. Conclusions
Our prospective study reported a positive association between baseline blood zinc levels and humoral immune response to COVID-19 vaccination in a Japanese population. This finding suggests the importance of assessing blood zinc prior to vaccination, as managing zinc concentrations through appropriate dietary and lifestyle modifications may enhance vaccine efficacy.
Author Contributions
Conceptualization, M. H., A. M., and Y. H.; Methodology, M. H. and A. M.; Formal Analysis, M. S. A.; Investigation, M. H., A. M., M. T., and G. Y.; Resources, M. H. and A. M.; Data Curation, M. H. and A. M.; Writing—Original Draft Preparation, M. S. A. and A. M.; Writing—Review and Editing, all authors; Visualization, M. S. A.; Supervision, M. H. and A. M.; Project Administration, M. H.; Funding Acquisition, Y. H. and M. H. All the authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by a research grant for Research on Emerging and Re-emerging Infectious Diseases, Health, and Labour Science Research Grants from the Ministry of Health, Labour and Welfare, Japan (R2-SHINKOGYOSEI-SHITEI-003 and 20HA2001).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Ethics Committee for Clinical Research of the School of Medicine at Saga University (approval no. R2-44 and R3-9 date of approval: Month date, 2021).
Informed Consent Statement
Informed consent was obtained from all participants involved in the study. Written informed consent was obtained from the patients prior to the publication of this paper.
Data Availability Statement
Data presented in this study are available upon request from the corresponding author (A.M.). The data are not publicly available due to privacy concerns.
Acknowledgments
We thank Miyuki Fuchigami, Mikako Horita, Nobuko Sato, Ayako Ajishi, and Miwako Koga for their administrative and technical support. We also thank Mizuho A. Kido, Histology and Neuroanatomy, Department of Anatomy and Physiology, Faculty of Medicine, Saga University, for educational assistance. Finally, we acknowledge the students, healthcare workers, and employees of Saga University for participating in the present study and for donating blood samples to evaluate humoral immune responses after vaccination.
Conflicts of Interest
The authors declare no conflicts of interest. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Cai, Y.; Kim, D.J.; Takahashi, T.; Broadhurst, D.I.; Ma, S.; Rattray, N.J.W.; Casanovas-Massana, A.; Israelow, B.; Klein, J.; Lucas, C.; et al. Kynurenic acid underlies sex-specific immune responses to COVID-19. medRxiv 2021, 14, eabf8483. [Google Scholar]
- Bechmann, N.; Barthel, A.; Schedl, A.; Herzig, S.; Varga, Z.; Gebhard, C.; Mayr, M.; Hantel, C.; Beuschlein, F.; Wolfrum, C.; et al. Sexual dimorphism in COVID-19: potential clinical and public health implications. Lancet Diabetes Endocrinol. 2022, 10, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Arnold, C.G.; Libby, A.; Vest, A.; Hopkinson, A.; Monte, A.A. Immune mechanisms associated with sex-based differences in severe COVID-19 clinical outcomes. Biol. Sex. Differ. 2022, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Galmes, S.; Serra, F.; Palou, A. Current State of Evidence: Influence of Nutritional and Nutrigenetic Factors on Immunity in the COVID-19 Pandemic Framework. Nutrients 2020, 12, 2738. [Google Scholar] [CrossRef] [PubMed]
- de Faria Coelho-Ravagnani, C.; Corgosinho, F.C.; Sanches, F.F.Z.; Prado, C.M.M.; Laviano, A.; Mota, J.F. Dietary recommendations during the COVID-19 pandemic. Nutr. Rev. 2021, 79, 382–393. [Google Scholar] [CrossRef]
- Richardson, D.P.; Lovegrove, J.A. Nutritional status of micronutrients as a possible and modifiable risk factor for COVID-19: a UK perspective. Br. J. Nutr. 2021, 125, 678–684. [Google Scholar] [CrossRef]
- Schubert, C.; Guttek, K.; Reinhold, A.; Grüngreiff, K.; Reinhold, D. The influence of the trace element zinc on the immune system. LaboratoriumsMedizin 2015, 39. [Google Scholar] [CrossRef]
- Maares, M.; Haase, H. Zinc and immunity: An essential interrelation. Arch. Biochem. Biophys. 2016, 611, 58–65. [Google Scholar] [CrossRef]
- Hojyo, S.; Fukada, T. Roles of Zinc Signaling in the Immune System. J. Immunol. Res. 2016, 2016, 6762343. [Google Scholar] [CrossRef]
- Gorji, A.; Khaleghi Ghadiri, M. Potential roles of micronutrient deficiency and immune system dysfunction in the coronavirus disease 2019 (COVID-19) pandemic. Nutrition 2021, 82, 111047. [Google Scholar] [CrossRef]
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 2018, 14, 49. [Google Scholar] [CrossRef] [PubMed]
- Meydani, S.N.; Barnett, J.B.; Dallal, G.E.; Fine, B.C.; Jacques, P.F.; Leka, L.S.; Hamer, D.H. Serum zinc and pneumonia in nursing home elderly. Am. J. Clin. Nutr. 2007, 86, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Heller, R.A.; Sun, Q.; Hackler, J.; Seelig, J.; Seibert, L.; Cherkezov, A.; Minich, W.B.; Seemann, P.; Diegmann, J.; Pilz, M.; et al. Prediction of survival odds in COVID-19 by zinc, age and selenoprotein P as composite biomarker. Redox Biol. 2021, 38, 101764. [Google Scholar] [CrossRef]
- Jothimani, D.; Kailasam, E.; Danielraj, S.; Nallathambi, B.; Ramachandran, H.; Sekar, P.; Manoharan, S.; Ramani, V.; Narasimhan, G.; Kaliamoorthy, I.; et al. COVID-19: Poor outcomes in patients with zinc deficiency. Int. J. Infect. Dis. 2020, 100, 343–349. [Google Scholar] [CrossRef]
- Bagher Pour, O.; Yahyavi, Y.; Karimi, A.; Khamaneh, A.M.; Milani, M.; Khalili, M.; Sharifi, A. Serum trace elements levels and clinical outcomes among Iranian COVID-19 patients. Int. J. Infect. Dis. 2021, 111, 164–168. [Google Scholar] [CrossRef]
- Mossink, J.P. Zinc as nutritional intervention and prevention measure for COVID-19 disease. BMJ Nutr. Prev. Health 2020, 3, 111–117. [Google Scholar] [CrossRef]
- Rahman, M.T.; Idid, S.Z. Can Zn Be a Critical Element in COVID-19 Treatment? Biol. Trace Elem. Res. 2021, 199, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Razzaque, M.S. COVID-19 pandemic: Can zinc supplementation provide an additional shield against the infection? Comput. Struct. Biotechnol. J. 2021, 19, 1371–1378. [Google Scholar] [CrossRef]
- Chillon, T.S.; Maares, M.; Demircan, K.; Hackler, J.; Sun, Q.; Heller, R.A.; Diegmann, J.; Bachmann, M.; Moghaddam, A.; Haase, H.; et al. Serum Free Zinc Is Associated With Vaccination Response to SARS-CoV-2. Front. Immunol. 2022, 13, 2738. [Google Scholar] [CrossRef]
- Lopresti, A.L. The Effects of Psychological and Environmental Stress on Micronutrient Concentrations in the Body: A Review of the Evidence. Adv. Nutr. 2020, 11, 103–112. [Google Scholar] [CrossRef]
- Ashenagar, M.S.; Matsumoto, A.; Sakai, H.; Tokiya, M.; Hara, M.; Hirota, Y. Comparison of CLEIA and ELISA for SARS-CoV-2 Virus Antibodies after First and Second Dose Vaccinations with the BNT162b2 mRNA Vaccine. Vaccines 2022, 10, 487. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Hara, M.; Ashenagar, M.S.; Tokiya, M.; Sawada, T.; Iwasaka, C.; Furukawa, T.; Kitagawa, K.; Miyake, Y.; Hirota, Y. Variant Allele of ALDH2, rs671, Associates with Attenuated Post-Vaccination Response in Anti-SARS-CoV-2 Spike Protein IgG: A Prospective Study in the Japanese General Population. Vaccines 2022, 10, 1035. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, T.; Ikeda, K.; Tanaka, S.; Taniguchi, T.; Igari, H.; Onouchi, Y.; Kaneda, A.; Matsushita, K.; Hanaoka, H.; Nakada, T.A.; et al. Antibody responses to BNT162b2 mRNA COVID-19 vaccine and their predictors among healthcare workers in a tertiary referral hospital in Japan. Clin. Microbiol. Infect. 2021, 27, e1861. [Google Scholar] [CrossRef]
- Zheng, C.; Shao, W.; Chen, X.; Zhang, B.; Wang, G.; Zhang, W. Real-world effectiveness of COVID-19 vaccines: a literature review and meta-analysis. Int. J. Infect. Dis. 2022, 114, 252–260. [Google Scholar] [CrossRef]
- Deb, P.; Zannat, K.E.; Talukder, S.; Bhuiyan, A.H.; Jilani, M.S.A.; Saif-Ur-Rahman, K.M. Association of HLA gene polymorphism with susceptibility, severity, and mortality of COVID-19: A systematic review. HLA 2022, 99, 281–312. [Google Scholar] [CrossRef] [PubMed]
- Shimanoe, C.; Matsumoto, A.; Hara, M.; Akao, C.; Nishida, Y.; Horita, M.; Nanri, H.; Higaki, Y.; Tanaka, K. Perceived stress, depressive symptoms, and cortisol-to-cortisone ratio in spot urine in 6878 older adults. Psychoneuroendocrinology 2021, 125, 105125. [Google Scholar] [CrossRef]
- Salimans, L.; Liberman, K.; Njemini, R.; Kortekaas Krohn, I.; Gutermuth, J.; Bautmans, I. The effect of resistance exercise on the immune cell function in humans: A systematic review. Exp. Gerontol. 2022, 164, 111822. [Google Scholar] [CrossRef]
- De Spiegeleer, A.; Bronselaer, A.; Teo, J.T.; Byttebier, G.; De Tre, G.; Belmans, L.; Dobson, R.; Wynendaele, E.; Van De Wiele, C.; Vandaele, F.; et al. The Effects of ARBs, ACEis, and Statins on Clinical Outcomes of COVID-19 Infection Among Nursing Home Residents. J. Am. Med. Dir. Assoc. 2020, 21, 909–914 e902. [Google Scholar] [CrossRef]
- Daniels, L.B.; Sitapati, A.M.; Zhang, J.; Zou, J.; Bui, Q.M.; Ren, J.; Longhurst, C.A.; Criqui, M.H.; Messer, K. Relation of Statin Use Prior to Admission to Severity and Recovery Among COVID-19 Inpatients. Am. J. Cardiol. 2020, 136, 149–155. [Google Scholar] [CrossRef]
- Haase, H.; Rink, L. Zinc signals and immune function. Biofactors 2014, 40, 27–40. [Google Scholar] [CrossRef]
- Kawano, Y.; Noma, T. Role of interleukin-2 and interferon-gamma in inducing production of IgG subclasses in lymphocytes of human newborns. Immunology 1996, 88, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Sauerwein, R.W.; Van der Meer, W.G.; Drager, A.; Aarden, L.A. Interleukin 2 induces T cell-dependent IgM production in human B cells. Eur. J. Immunol. 1985, 15, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Mascle, X.; Albagli, O.; Lemercier, C. Point mutations in BCL6 DNA-binding domain reveal distinct roles for the six zinc fingers. Biochem. Biophys. Res. Commun. 2003, 300, 391–396. [Google Scholar] [CrossRef]
Figure 1.
Study design. All participants, including healthcare workers, university students, and employees, received two doses of the Pfizer and Moderna (BioNTech) COVID-19 vaccines. Blood samples were collected at Week 0 and at various intervals up to Week 12 after the second vaccination from healthcare workers (subcohort 1), university students (subcohort 2), and university employees and students (subcohort 3).
Figure 1.
Study design. All participants, including healthcare workers, university students, and employees, received two doses of the Pfizer and Moderna (BioNTech) COVID-19 vaccines. Blood samples were collected at Week 0 and at various intervals up to Week 12 after the second vaccination from healthcare workers (subcohort 1), university students (subcohort 2), and university employees and students (subcohort 3).
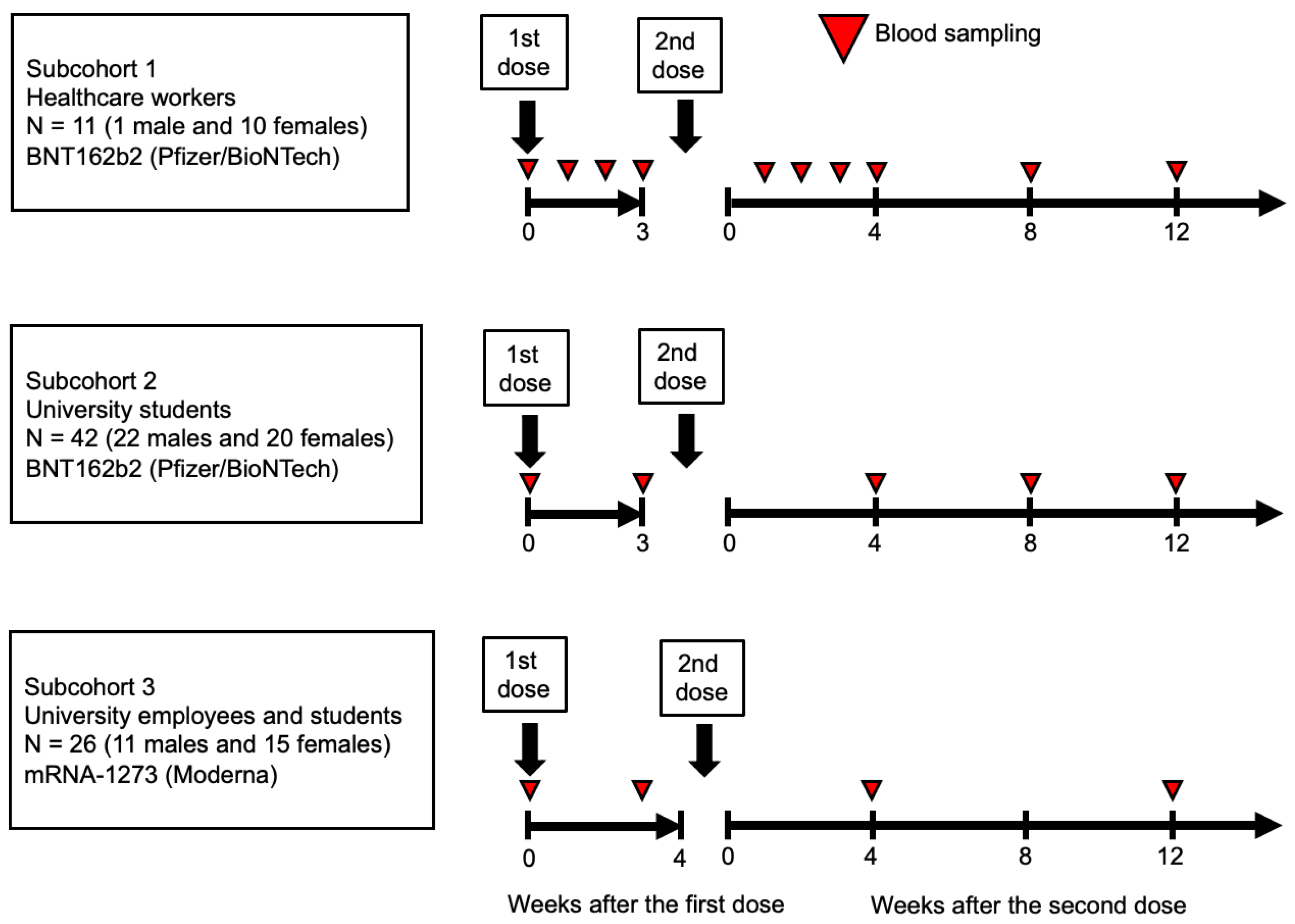
Figure 2.
Zinc levels according to sex and distribution. The total blood zinc levels are displayed as dots for each individual, categorized by sex (male and female). The bars represent the median zinc levels, and the error bars indicate the interquartile range. The p-value was used for the Spearman's rank correlation between sex and zinc levels (a). The histograms (b) represent the distribution of zinc levels separately for male and female individuals. The gray-shaded area corresponds to zinc deficiency (<80 μg/dL)) according to the general standards in Japanese hospitals, allowing comparison of the zinc deficiency prevalence between sexes.
Figure 2.
Zinc levels according to sex and distribution. The total blood zinc levels are displayed as dots for each individual, categorized by sex (male and female). The bars represent the median zinc levels, and the error bars indicate the interquartile range. The p-value was used for the Spearman's rank correlation between sex and zinc levels (a). The histograms (b) represent the distribution of zinc levels separately for male and female individuals. The gray-shaded area corresponds to zinc deficiency (<80 μg/dL)) according to the general standards in Japanese hospitals, allowing comparison of the zinc deficiency prevalence between sexes.
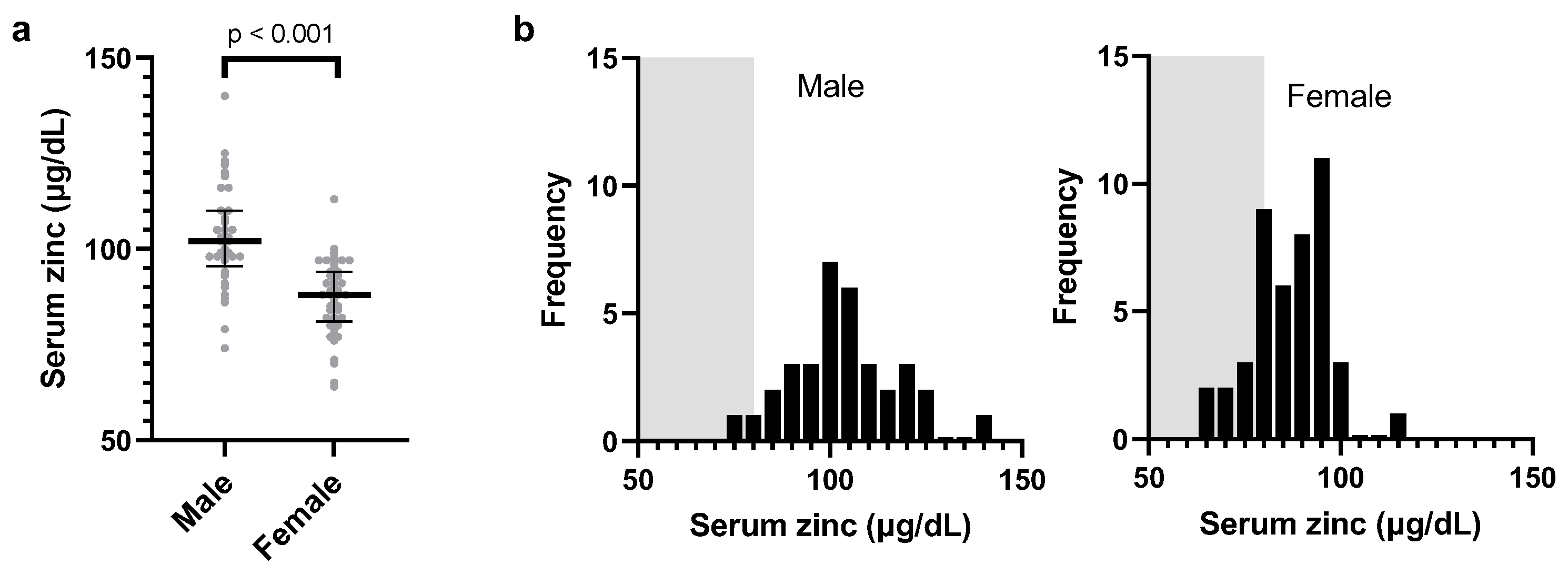
Figure 3.
Estimated anti-S1 IgM and IgG levels after the second dose in female participants. Least-squares geometric means and standard errors were calculated using mixed models that account for vaccine type, age, and the interaction between zinc levels (T1, T2, and T3 as class variables) and the number of weeks post-vaccination (as a class variable). The models include random effects based on repeated measures and the three cohorts. Binding antibody units (BAU) were calibrated against the WHO international standard, and the x-axis is displayed on a log-2 scale for improved visualization. SU, Sysmex unit.
Figure 3.
Estimated anti-S1 IgM and IgG levels after the second dose in female participants. Least-squares geometric means and standard errors were calculated using mixed models that account for vaccine type, age, and the interaction between zinc levels (T1, T2, and T3 as class variables) and the number of weeks post-vaccination (as a class variable). The models include random effects based on repeated measures and the three cohorts. Binding antibody units (BAU) were calibrated against the WHO international standard, and the x-axis is displayed on a log-2 scale for improved visualization. SU, Sysmex unit.
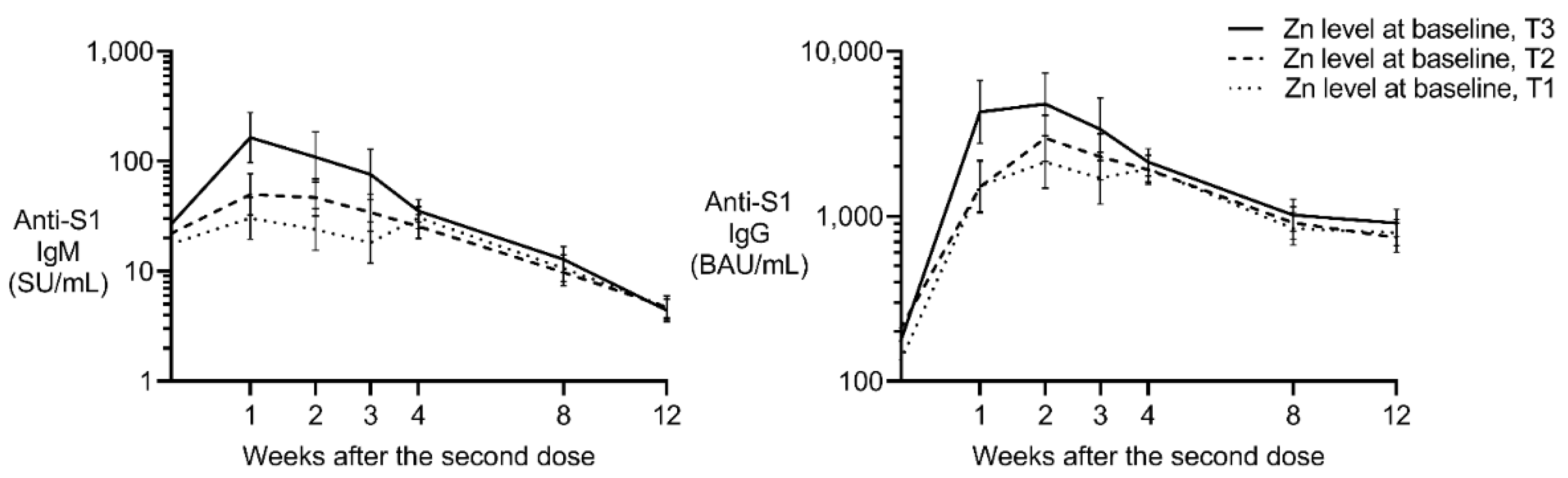

Table 1.
Baseline characteristics of the participants.
| Male | Female | ||||||
| Serum zinc categories | T1 | T2 | T3 | T1 | T2 | T3 | |
| Median (μg/dL) | 90.5 | 103 | 119 | 78.5 | 88.5 | 97 | |
| Interquartile range (μg/dL) | 74–98 | 99–107 | 108–140 | 64–82 | 83–92 | 93–113 | |
| Number | 12 | 11 | 11 | 16 | 14 | 15 | |
| Age, years | |||||||
| Median | 23 | 22 | 23 | 37.7 | 22.2 | 22 | |
| Interquartile range | 20.7–63.0 | 20.5–47.2 | 20.9–46.8 | 22.0–59.0 | 21.3–54.0 | 20.7–59.6 | |
| Occupation | |||||||
| Healthcare worker | 1 | 0 | 0 | 3 | 5 | 2 | |
| Student | 7 | 7 | 8 | 6 | 6 | 8 | |
| University worker | 4 | 4 | 3 | 7 | 3 | 5 | |
| Smoking, yes | 0 | 1 | 0 | 0 | 0 | 0 | |
| Steroid use, yes | 0 | 0 | 2 | 0 | 0 | 0 | |
| Dyslipidemia | 0 | 0 | 0 | 1 | 0 | 0 | |
| Perceived stress* | |||||||
| 0/1/2/3/4 | 3/1/3/5/0 | 5/1/1/3/1 | 5/1/3/2/0 | 2/2/5/5/2 | 4/1/4/4/1 | 6/1/2/5/1 | |
| rs671 variant allele** | |||||||
| 0/1/2 | 8/3/1 | 3/5/3 | 4/5/2 | 10/6/0 | 7/6/1 | 10/4/1 | |
| Ethanol, g/day/weight60kg | |||||||
| <1/ ≥1,<20/ ≥20 | 3/8/1 | 6/4/1 | 6/5/0 | 12/4/0 | 7/5/2 | 11/4/0 | |
| Exercise habit*** | |||||||
| 0/1/2/3 | 2/4/4/2 | 2/5/2/2 | 3/0/5/3 | 7/1/5/3 | 6/0/5/3 | 10/1/3/1 | |
| Allergic disease, yes | 2 | 0 | 2 | 4 | 1 | 1 | |
| Sleep disturbance | 0 | 2 | 0 | 0 | 1 | 1 | |
*No (0) to yes (4). **ALDH2*1/*1 (0), ALDH2*1/*2 (1), ALDH2*2/*2 (2). ***No habit (0), less than once a week (1), 1 to 3 days/week (2), and more than 3 days/week (3).
Table 2.
Estimation of fixed effects on log-transformed anti-S1 IgM (SU/mL).
| Males | Females | ||||||||
| Model 1 | Model 2 | Model 1 | Model 2 | ||||||
| AIC = 427.1 | AIC = 419.6 | AIC = 682 | AIC = 680 | ||||||
| 162 observations | 162 observations | 255 observations | 255 observations | ||||||
| 34 participants | 34 participants | 45 participants | 45 participants | ||||||
| Fixed effect | β | p | β | p | Β | p | β | p | |
| BNT162b2 (reference) | |||||||||
| mRNA-1273 | −0.05036 | 0.7928 | −0.2172 | 0.3295 | 0.251 | 0.1028 | 0.239 | 0.2248 | |
| Age (per year of age) | −0.01927 | 0.0327 | −0.00379 | 0.6982 | −0.00958 | 0.0573 | −0.005 | 0.4755 | |
| Body height (per cm) | −0.01749 | 0.2796 | −0.012 | 0.3177 | |||||
| Steroid use, yes | −1.1015 | 0.0055 | — | — | |||||
| Dyslipidemia, yes | — | — | −1.3556 | 0.0002 | |||||
| Allergic disease, yes | 0.1747 | 0.3552 | 0.005 | 0.9729 | |||||
| Exercise habit (per category) | 0.01473 | 0.8767 | 0.028 | 0.6047 | |||||
| Cigarette smoke, yes | −0.07372 | 0.9281 | — | — | |||||
| Ethanol intake (per category) | −0.5076 | 0.0008 | 0.2721 | 0.0139 | |||||
| Perceived stress (per category) | −0.02271 | 0.7359 | 0.0276 | 0.6176 | |||||
| Sleep disturbance, yes | −0.3611 | 0.5297 | −0.7167 | 0.0124 | |||||
| Log zinc (μg/dL) | −0.7695 | 0.1856 | −0.4191 | 0.4909 | 2.2135 | <0.0001 | 1.5435 | 0.0122 | |
| The effects of baseline characteristics were computed using mixed models to account for repeated measurements and the random effect of the subpopulation. All models include the fixed effects of post-vaccination week as a categorical variable and variables listed in the table. The results are expressed in sysmex unit per mL (SU/mL). β, partial correlation coefficient. | |||||||||
Table 3.
Estimation of fixed effects on log-transformed anti-S1 IgG (BAU/mL).
| Males | Females | ||||||||
| Model 1 | Model 2 | Model 1 | Model 2 | ||||||
| AIC = 258 | AIC = 263 | AIC = 529 | AIC = 535 | ||||||
| 162 observations | 162 observations | 255 observations | 255 observations | ||||||
| 34 participants | 34 participants | 45 participants | 45 participants | ||||||
| Fixed effect | β | p | β | p | β | p | β | p | |
| BNT162b2(reference) | |||||||||
| mRNA-1273 | 0.4599 | 0.0062 | 0.3233 | 0.0094 | 0.8347 | <0.0001 | 0.7555 | <0.0001 | |
| Age (per years of age) | −0.012 | 0.017 | −0.005 | 0.3195 | −0.005 | 0.1384 | −0.0038 | 0.4312 | |
| Body height (per cm) | −0.0165 | 0.0659 | −0.002 | 0.8262 | |||||
| Steroid use, yes | −0.4357 | 0.0455 | — | — | |||||
| Dyslipidemia, yes | — | — | −0.6725 | 0.0107 | |||||
| Allergic disease, yes | 0.1581 | 0.131 | −0.2301 | 0.0196 | |||||
| Exercise habit (per category) | 0.0388 | 0.46 | −0.0102 | 0.7973 | |||||
| Cigarette smoke, yes | −1.2462 | 0.0064 | — | — | |||||
| Ethanol intake (per category) | −0.1627 | 0.0487 | −0.0068 | 0.9326 | |||||
| Perceived stress (per category) | 0.07533 | 0.0444 | 0.044 | 0.2769 | |||||
| Sleep disturbance, yes | −0.7196 | 0.0246 | −0.388 | 0.064 | |||||
| Log zinc(μg/dL) | −0.3268 | 0.3113 | −0.056 | 0.8676 | 1.609 | <0.0001 | 1.124 | 0.0131 | |
| The effects of baseline characteristics were computed using mixed models to account for repeated measurements and the random effect of the subpopulation. All models include the fixed effects of post-vaccination week as a categorical variable and variables listed in the table. BAU, binding antibody units, calibrated using the WHO International Standard. β, partial correlation coefficient. | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



