
Submitted:
22 October 2024
Posted:
23 October 2024
You are already at the latest version
Abstract
Leydig cells, pivotal for male fertility, undergo significant oxidative stress due to their proximity to macrophages and reactive oxygen species (ROS) generated by extracellular structures. This stress disrupts Leydig cell function, impairing steroidogenesis and spermatogenesis, ultimately leading to male infertility. Oxidative stress triggers apoptosis and inhibits testosterone production, crucial for spermatogenesis regulation. Understanding the mechanisms of oxidative stress and antioxidation on Leydig cells is vital. Some studies highlight the role of antioxidants in mitigating oxidative damage, emphasizing critical pathway of Sirt1 in oxidative stress and Nrf2 in antioxidation. Future research should focus on screening antioxidant substances and optimizing antioxidant dosages for Leydig cell protection and testosterone restoration, thereby enhancing reproductive performance. This review provides insights into combating male infertility by addressing Leydig cell oxidative stress and antioxidation.
Keywords:
Leydig cells
; spermatogenesis
; testosterone
; oxidative stress
; antioxidants
1. Introduction
Male infertility caused by oxidative stress in Leydig cells has become a prominent problem. In the interstitial tissues adjacent to the testicular seminiferous tubules, Leydig cells serve as important endocrine cells. Their pivotal role in steroidogenesis is crucial for male fertility, as is their contribution to spermatogenesis [1,2]. Testosterone synthesized by Leydig cell exerts diverse effects on male development across the whole reproductive stages. Specifically, during the fetal stage, testosterone secretion by fetal Leydig cell is imperative for the morphogenesis of the genital tract and external genitalia, as well as the migration of the testes into the scrotum. In puberty, the testosterone plays a critical role in facilitating spermatogenesis, maintaining the structure integrity of the blood-testicular barrier, and ensuring fertility maintenance [2]. Due to the proximity of Leydig cell to macrophages in the testis, Leydig cell highly susceptible to reactive oxygen species (ROS) produced by extracellular structures [2,3]. Activated macrophages release ROS, which can oxidize adjacent tissues and cells, including sensitive Leydig cells to oxidative stress [3]. Furthermore, Leydig cell also be important components of innating immune responses by releasing of cytokines, chemokines, and other substances that defenses against pathogens and potential self-impairments [4]. when the homeostatic balance between antioxidant and oxidative stress induced by ROS is disrupted, Leydig cells become vulnerable to oxidative stress, leading to severe dysfunction in steroidogenesis and spermatogenesis, ultimately causing in male infertility [2]. The excessive production of oxygen free radicals induces oxidative stress, which overwhelms the antioxidant capabilities of cells, leading to homeostatic imbalance of ROS and antioxidation. Studies have demonstrated that oxidative stress damages the testicular cells by promoting lipid peroxide, triggering apoptosis, disrupting mitochondrial function, and reducing testosterone secretion [5]. Decreased testosterone production by Leydig cells can result in impaired steroidogenesis, compromised spermatogenesis, and ultimately male infertility [6,7].
The review summarizes the detrimental effects of oxidative stress on leydig cells, and explores the antioxidative mechanism of common antioxidants. By providing an overview of oxidative and antioxidative stress in Leydig cells, this review offers valuable insights for developing strategies to prevent male infertility resulting from oxidative stress.
1. Pathways of ROS Production in the Organism and Potential Harm to Male Fertility
Elevated ROS levels in the male reproductive tract, stemming from both endogenous and exogenous factors, disrupt the balance between oxidant production and the scavenging activity of antioxidant enzymes, thus triggering oxidative stress, as illustrated in Figure 1. Overall, ROS disturbed the regulation of metabolic hormone (Leptin, Insulin and T3), hormone (Cortisol) and immune (Proinflammatory cytokines) stress, and then indirectly impact on the regulation of hypothalamus-pituitary gland-Testes axis (H-P-T axis). Meanwhile, the ROS directly disturb the regulation of H-P-T axis. Finally, ROS is harm to male fertility. Some synthetic and natural antioxidants have been widely used to alleviate oxidative stress of Leydig cell.
Various sources of ROS originate from testicular cells, with disruptions observed in the mitochondrial electron transport chain (ETC) and mitochondrial/microsomal cytochrome P450 enzymatic reactions, representing consequences of the oxidative stress [74]. The suppression of StAR protein transcription and subsequent the reduce of cholesterol transport to the inner mitochondrial membrane for conversion into 17-hydroxypregnenolone is a consequence of oxidative stress, resulting in impaired Leydig cell steroidogenesis [75]. By suppressing the transcription of essential enzymes such as P450scc, CYP17, and 3β-HSD, ROS, particularly H2O2 originating from medullary cells and testicular macrophages, induces mitochondrial dysfunction, thereby compromising steroidogenesis [76]. Therefore, ROS disturb the process of testosterone synthesis by inhibiting the transcription of enzymes in ETC, and steroidogenic acute regulatory protein (StAR), that is a translocator protein (TSP) co-factor and participate in the formation of transduceosome to transport cholesterol into mitochondria (Figure 2). The presence of oxidative stress (OS) in spermatogonia is also correlated with diminished levels of cellular antioxidant enzymes, such as glutathione peroxidase (GPX), superoxide dismutase (SOD), malondialdehyde (MDA), and catalase (CAT), along with the progression of apoptotic processes [65,77].
3. Function of Leydig Cells
3.1. Involvement in the Regulation of Spermatogenesis
Leydig cells are in the connective tissues surrounding the seminiferous tubules of the testes, and mature Leydig cells are typical steroid hormone-secreting cells characterized by abundant slippery endoplasmic reticulum, large and numerous mitochondria, and the presence of lipid droplets in the cytoplasm, containing essential substances for steroid hormones synthesis. The hallmark enzyme of Leydig cell is 3β-hydroxysteroid dehydrogenase. Leydig cells play pivotal roles in supporting germ cells, regulating spermatogenesis, and secreting testosterone to promote sperm maturation. Beyond their roles in supporting germ cells and regulating spermatogenesis, Leydig cells serve crucial functions in reproductive physiology. They maintain the integrity of immune barrier for germ cells by promoting the tight junction of blood-testis barrier. Additionally, Leydig cells secrete testosterone that is essential for promoting the development, differentiation of reproductive organs, as well as maintaining secondary sexual characteristics and sexual function.
Spermatogenesis is a major physiological function of testes, and serves as the basis of male reproduction. Abnormalities in spermatogenesis leads to male infertility. A series of intricate and interacting processes involve in regulation in spermatogenesis [8]. Leydig cells, alongside peritubular myeloid cells and Sertoli cells, form a distinct somatic cell population responsible for orchestrating spermatogenesis, with Leydig cells playing a vital regulatory role in this process (Figure 3).
3.2. Production of Testosterone and Indirect Regulation in Spermatogenesis
Several studies have highlighted that hormones and autocrine signals play a crucial role in continuous and asynchronous sperm production [8]. Androgens secreted by Leydig cells in the testes play a fundamental role in male sexual differentiation and behavioral patterns [9,10], serving as the cornerstone for sustaining, initiating, and modulating spermatogenesis [11,12,13,14]. Testicular cells treated with androgen-specific knockdowns induced androgen receptor binding in Leydig cells to control spermatogenesis. suggesting an irreplaceable role for Leydig cell in maintaining normal male reproduction [15,16,17,18]. These studies have confirmed that normal spermatogenesis is dependent on testosterone concentration to be maintained by Leydig cells.
Testicular Leydig cells play a pivotal role in the indirect regulation of spermatogenesis, primarily mediated by steroidogenic proteins such as StAR, members of the cytochrome P450 family (P450scc, P450c17), the peripheral-type benzodiazepine receptor, hydroxysteroid dehydrogenases (17β-HSD, 3β-HSD), and the luteinizing hormone receptor (LHR), which collectively facilitate testosterone biosynthesis [36]. The synthesis of testosterone in Leydig cells of the testis commences with the mobilization of cholesterol from lipid droplets and/or plasma membranes. This cholesterol is then shuttled into mitochondria, where it is enzymatically converted into pregnenolone. Subsequently, pregnenolone is processed by enzymes present in the smooth endoplasmic reticulum (ER) to yield steroidal products [19].
The production of testosterone can be outlined in the following sequential steps: In response to the hypothalamic gonadotropin-releasing hormone (GnRH), the pituitary gland secretes luteinizing hormone (LH). LH binds to LH receptor (LHR) on the Leydig cell, activating G proteins and stimulating the production of cyclic adenosine 3',5'-monophosphate (cAMP), ultimately initiating steroidogenesis [20]. In the subsequent stages, cAMP prompts the translocation of cholesterol to the mitochondria, marking a pivotal step in steroid synthesis. Following this, pregnenolone is synthesized from the cholesterol in the mitochondria [21], which is then enzymatically transformed in the smooth endoplasmic reticulum to produce testosterone [22,23].
Testosterone diffuses from leydig cells, via androgen-binding proteins translocated to Sertoli cells where it binds to androgen receptors to regulate spermatogenesis. At least four crucial processes in spermatogenesis depend on testosterone: maintenance of BTB, meiosis, Sertoli-sperm adhesion, and sperm release [24,25,26,27,28,29]. This suggests that leydig cells play an indirect regulatory role in spermatogenesis [30].
3.3. Other Factors Regulating Spermatogenesis via Leydig Cells
There has been a large body of research that shows that Leydig cells regulate spermatogenesis through the secretion of growth factors and steroidogenesis [31,32]. Furthermore, testicular Leydig cells produce cytokines and hormones that also contribute to regulation of spermatogenesis [33], In order for hormones and cytokines to be able to regulate spermatogenesis, they must bind to specific receptors on Leydig cells (Table 1) including estrogens, interleukin 1α (IL-1α), inhibin, insulin-like peptide 3 (INSL3), insulin-like growth factor 1 (IGF1), transforming growth factor β (TGFβ), and thyroid hormones [9,10,34,35,36,37]. Studies suggest that leydig cell dysfunction adversely affects not only spermatogenesis and sex differentiation, but also gonadal development, and androgen production
4. The Impairment of Oxidative Stress on Leydig Cell Functions
4.1. Stimulation of Leydig Cell Apoptosis
Apoptosis, a crucial aspect of normal cellular functioning, involves the programmed elimination of cells, leading to a reduction in their overall number. Oxidative stress may damage the cell's DNA, leading to the activation of death receptors in the cell. Nevertheless, in response to oxidative stress, genes in apoptotic signaling may be weakened, thereby impeding the signaling required for normal apoptosis. Thus, intense oxidative stress can then lead to necrosis or cancerous transformation of cells during the course of the disease.
Exposing to H2O2 can provoke DNA damage and deteriorate the functional performance as well as hormone secretion, particularly leydig cells producing testosterone. Several studies have shown that in vitro, even at physiological concentrations, H2O2 interferes with leydig cells’ normal function and increases the abnormal levels of intracellular oxidative stress and then lead to cell apoptosis. The accumulation of excessively produced H2O2 in leydig cells in vivo can lead to oxidative stress, often resulting from hormonal disturbances or other conditions that compromise cellular function [65].
4.2. Inhibition of Testosterone Production
When the concentrations of oxygen and its derived free radicals surpass the intrinsic antioxidant defenses of a cell, it results in a state of oxidative stress. ROS comprise oxygen radicals like hydroxyl radicals and superoxide anions [3], as well as substances that, although not radicals in their nature, are capable of generating free radicals in the context of cellular and tissue environments [66].
The crucial role of luteinizing hormone in stimulating testosterone production in Leydig cells is imperative for spermatogenesis. The testes rely on adequate levels of testosterone to ensure proper spermatogenesis and the survival of germ cells. Disruptions in testosterone production, whether partial or complete, can have significant adverse effects on the spermatogenic process, potentially leading to its malfunction or complete cessation.
Multiple investigations have revealed a direct correlation between oxidative stress-mediated cellular damage and overall testosterone production. Alleviating the inhibition of Leydig cells and testosterone synthesis by oxidative stress may improve the state of testosterone production.
4.3. Other Harmful Effects
First, oxidative stress has an impact on cellular metabolism, and once oxidative stress occurs, it initiates a stress-adaptive response in the cellular life mechanism [67]. This response regulates cellular metabolism in response to the appropriate stress stimulus [68]. However, when the oxidative stress level is higher, this adaptive response is not effective in rescuing cellular damage. For example, high levels of H202 in the cell give rise to oxidative damage to proteins and DNA [69]. Similarly, the more a cell adapts to oxidative stress, the more it gradually reduces its metabolic capacity and function [70].
Second, oxidative stress affects mitochondria and thus the process of cellular esterification. Mitochondria play a major role in adenylate conversion, and red blood cells contain a large number of adenylate molecules [71]. If the cell is subjected to some stress, it may lead to mitochondrial damage, which can perturb cellular metabolism. And this perturbation will precisely trigger oxidative stress [72].
ROS impedes the phosphorylation of mTOR/AKT/ERK1/2 and LH/FSH signaling, consequently suppressing Leydig cell differentiation. This, in turn, results in reduced testosterone (T) levels and spermatogenesis due to lower FSH concentrations [73].
5. Current Status of Research on Antioxidant Stress Substances for Leydig Cells
5.1. Classification of Antioxidant Substances
The principle of antioxidant is to slow down or inhibit cellular oxidative damage by scavenging free radicals, there are a large number of varieties of antioxidants, which have different effects on the cell or the organism, the following table (Table 2) classifies them according to their sources and summarizes their functional roles [78,79].
5.2. Current Research on Antioxidant Substances against Oxidative Stress
Despite the abundance of antioxidant compounds, the research investigating the application of antioxidants to Leydig cells for combating oxidative stress and mitigating the adverse effects of ROS on male spermatogenesis and reproductive function remains conspicuously limited [97,98,99,100], and the following table (Table 3) summarizes the roles and pathways of antioxidant stress substances that have been used in leydig cells in recent years. In conclusion, Melatonin and Vitamin E is effective and affordable, and Nrf2 signal pathway is in antioxidation.
5.3. Typical Pathways against Oxidative Stress in Leydig Cell
Silent information regulator of transcription 1 (SIRT1), a critical regulator in the oxidative stress pathway, it belongs to a conserved subclass of histone deacetylases that are dependent on nicotinamide adenine dinucleotide (NAD+) [85,86,87]. Recent research has furthermore validated the pivotal role of SIRT1 in conferring resilience against oxidative stress-induced injuries [92,93], and its mechanism may be related to its deacetylation effect on different target genes and target proteins [88,89,90,91].
Oxidative stress induces apoptosis in Leydig cells, an important mechanism as contributing to male reproductive impairment. Activation of autophagy serves to protects Leydig cells [124,125], the SIRT1 pathway antagonizes oxidative stress and activates cellular autophagy. The reduction of ROS mediated by Sirt1 involves a complex signaling network, including the Sirt1/FOXOs, Sirt1/NF-κB, Sirt1/NOX, Sirt1/SOD, and Sirt1/SOD. Sirt1/SOD and Sirt1/eNO. Among these biological factors, eNOs and SOD occupy a downstream position relative to FOXOs, which activate Sirt1 and, subsequently initiate the signaling pathway via a positive feedback loop [94,95,96]. Therefore, the augmentation of various antioxidant molecules helps to mitigate the adverse effects of oxidative stress [126,127].
Several investigations have demonstrated the capacity of Sirt1 to modulate several crucial regulatory factors (Figure 4), including the nuclear factor-κB (NF-κB), the forkhead box O1 transcription factor (FOXO1), the nuclear factor erythroid 2-related factor 2 (Nrf2), the tumor suppressor P53, as well as the peroxisome proliferator-activated receptor γ coactivator 1α (PGC-1α). Additionally, it has been found to regulate the activity of the hypoxia-inducible factor-1α (HIF-1α) and the adenosine 5'-monophosphate (AMP)-activated protein kinase (AMPK), and endothelialnitric synthase (eNOS), Ku70, P66Shc, and many other target genes and target proteins, which in turn play important roles in oxidative stress injury [97,98,99,100,124,125].
6. Conclusions
Maintaining an optimal balance between oxygen-derived free radicals and antioxidants is necessary for Leydig cells to protect themselves against oxidative insults. Under normal conditions, Leydig cells rely on numerous antioxidants to uphold their antioxidant defenses, providing protection or restoration when confronted with oxidative stress. When managing oxidative stress, it is imperative to administer antioxidants at the appropriate dosage or concentration. Additionally, assessing whether the antioxidants can restore the testosterone-producing capacity of Leydig cells impaired by oxidative stress is of great importance for their potential application in livestock and poultry production. Further research on antioxidants should delve into greater depth, aiming to effectively mitigate oxidative damage to Leydig cells and enhance reproductive outcomes, such as spermatogenesis, in male animals at an affordable cost and with easy accessibility.
Author Contributions
Drafting the manuscript and Data analysis: Y.T. and H.L.; Conceptualization and Supervision: M.Z.; Reviewing the manuscript: M.Z., G.Z. and C.Z.; Project administration, M.Z.; Funding acquisition: MZ. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by National Key Research and Development Projects of China (2021YFD1600201, 2021YFD1200403), and 14th Five Year Major Special Project in Sichuan Province (2021ZDZX0008).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Leisegang, K.; Henkel, R.; Agarwal, A. The in vitro modulation of steroidogenesis by inflammatory cytokines and insulin in TM3 Leydig cells. Endocrinology 2018, 16, 26. [Google Scholar] [CrossRef]
- Riris, A. A. I. D. C.; I'tishom, R.; Khaerunnisa, S.; Juliantina, F.; Qurrota, A.; Mardhiyah, M. S. Role of antioxidant to protect Leydig cells induced by reactive oxygen species, a literature review. 2021, 1.
- Shi, Z.; Feng, Y.; Wang, J.; Zhang, H.; Ding, L.; Dai, J. Perfluorododecanoic acid-induced steroidogenic inhibition is associated with steroidogenic acute regulatory protein and reactive oxygen species in cAMP-stimulated Leydig cells. Toxicol Sci 2010, 114, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Yong, L.; Fa, W.; Zhang, R.; Liu, B. T-2 toxin induces apoptosis via the Bax-dependent caspase-3 activation in mouse primary Leydig cells. Journal of Biochemical and Molecular Toxicology.
- Agarwal, A.; Parekh, N.; Selvam, M. K. P.; Henkel, R.; Shah, R.; Kalyan, S.; Punab, M.; Sengupta, P. Male Oxidative Stress Infertility (MOSI), Proposed Terminology and Clinical Practice Guidelines for Management of Idiopathic Male Infertility. World Journal of Men's Health 2019, 37, 296–312. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R. J.; Smith, T. B.; Jobling, M. S.; Baker, M. A.; De Iuliis, G. N. Oxidative stress and male reproductive health. Asian Journal of Andrology 2014, 16, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Clark, B. J.; Stocco, D. M.; Tu, H.; Manna, P. R.; Wang, X. J. Cholesterol Transporters of the START Domain Protein Family in Health and Disease. Biochimica et Biophysica Acta 2015, 1851, 674–691. [Google Scholar]
- Cheng, C. Y.; Wong, E. W. P.; Yan, H. H. N.; Mruk, D. D. Regulation of spermatogenesis in the microenvironment of the seminiferous epithelium, New insights and advances. Advances in Experimental Medicine and Biology 2010, 315, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Andrabi, S. S.; Parvez, S.; & Tabassum, H.; & Tabassum, H. Ischemic stroke and mitochondria: Mechanisms and targets. Protoplasma 2020, 257, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Sharp, J. L.; Ethridge, S. B.; Ballard, S. L.; Adkins, A. E.; Moore, B. L. The effects of chronic estradiol treatment on opioid self-administration in intact female rats. Drug and Alcohol Dependence 2021, 225, 108816. [Google Scholar] [CrossRef] [PubMed]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Zientara, A.; Dzemali, O.; Odavic, D.; Roukoz, H.; Holzhey, D.; Mohr, F. W. 158-Igiant Aneurysms of Saphenous Vein Grafts, Presentation of Two Uncommon Cases and Surgical Management. Journal of Thoracic and Cardiovascular Surgery 2013, 146, S107–S107. [Google Scholar]
- Cooke, P. S.; Walker, W. H. Male fertility in mice requires classical and nonclassical androgen signaling. Cell Reports 2021, 36, 109557. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Siow, R. C.; Mann, G. E. Impaired redox signaling and antioxidant gene expression in endothelial cells in diabetes: A role for mitochondria and the nuclear factor-E2-related factor 2-Kelch-like ECH-associated protein 1 defense pathway. Antioxidants & Redox Signaling 2011, 14, 469–487. [Google Scholar]
- Zirkin BR; Papadopoulos V. Leydig cells: formation, function, and regulation. Biol Reprod.
- Manna, P. R.; Cohen-Tannoudji, J.; Counis, R.; Huhtaniemi, I.; Kraemer, F. B.; Stocco, D. M. Mechanisms of action of hormone-sensitive lipase in mouse Leydig cells, its role in the regulation of the steroidogenic acute regulatory protein. Endocrinology 2013, 154, 8505–8518. [Google Scholar]
- Zhong, L.; Sun, H.; Zhang, Y.; Liu, C.; Hu, L.; Xie, Y. Research on the steroidogenesis of proliferated Leydig cells in vitro. Molecular and Cellular Endocrinology 2013, 370, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S. O.; Pant, A. B.; Farombi, E. O. Effects of quercetin on mRNA expression of steroidogenesis genes in primary cultures of Leydig cells treated with atrazine. Toxicology in Vitro 2013, 27, 700–707. [Google Scholar] [CrossRef]
- Fard, A. H. H.; Kouchesfehani, H. M.; Jalali, H. Investigation of cholestasis-related changes in characteristics of spermatogonial stem cells in testis tissue of male Wistar rats. Cell Journal (Yakhteh) 2020, 22, e13660. [Google Scholar]
- Zirkin, B.R.; Papadopoulos, V. Leydig cells: formation, function, and regulation. Biol Reprod 2018, 99, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Leping, Y.; Ye, Z. J.; Zhang, Y.; Sun, Y.; Xu, B.; Fan, Y.; Li, S.; Liu, C.; Wang, J.; Wang, X. Inhibitors of Testosterone Biosynthetic and Metabolic Activation Enzymes. Steroids 2011, 76, 785–794. [Google Scholar]
- Zhou, R.; Wu, J.; Liu, B.; Ma, G.; Zhang, X. The roles and mechanisms of Leydig cells and myoid cells in regulating spermatogenesis. Cellular and Molecular Life Sciences 2019, 76, 2681–2695. [Google Scholar] [CrossRef] [PubMed]
- Shima, Y.; Morohashi, K.I. Leydig progenitor cells in fetal testis. Mol Cell Endocrinol. 2017, 445, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Stanton, P. G.; Sluka, P.; Foo, C. F. H.; Stephens, A. N.; Smith, A. I. Proteomic Changes in Rat Spermatogenesis in Response to In Vivo Androgen Manipulation; Impact on Meiotic Cells. PLoS ONE 2012, 7, e41739. [Google Scholar] [CrossRef] [PubMed]
- Miriam, H.; Jenny, S.; Mark, J.; King, N. J. C.; Handelsman, D. J.; Allan, C. M. Sertoli and germ cell development in hypogonadal (hpg) mice expressing transgenic follicle-stimulating hormone alone or in combination with testosterone. Endocrinology 2003, 144, 509–517. [Google Scholar]
- Takada, M.; Fukuhara, D.; Takiura, T.; Nishibori, Y.; Kotani, M.; Kiuchi, Z.; Kudo, A.; Beltcheva, O.; Ito-Nitta, N.; Nitta, K.R. Involvement of GLCCI1 in mouse spermatogenesis. FASEB J 2023, 37, e22680. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.H. Non-classical actions of testosterone and spermatogenesis. Philos Trans R Soc Lond B Biol Sci. 2010, 365, 1557–1569. [Google Scholar] [CrossRef] [PubMed]
- Chauvigné, F.; Zapater, C.; Gasol, J.M.; Cerdà, J. Germ-line activation of the luteinizing hormone receptor directly drives spermiogenesis in a nonmammalian vertebrate. Proc Natl Acad Sci USA 2014, 111, 1427–1432. [Google Scholar] [CrossRef] [PubMed]
- Liza, O. D.; Kyriakos, P.; Andrea, W.; Ulrich, G.; Müller, J.; Gabriele, L.; McLachlan, R. I.; Stanton, P. G. Transcriptional profiling of the hormone-responsive stages of spermatogenesis reveals cell-, stage-, and hormone-specific events. Endocrinology 2009, 150, 5074–5084. [Google Scholar]
- Marettová, E.; Maretta, M.; Legáth, J. Toxic effects of cadmium on testis of birds and mammals: a review. Anim Reprod Sci 2015, 155, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Wei, X.; Deng, W. Nestin-dependent mitochondria-ER contacts define stem Leydig cell differentiation to attenuate male reproductive ageing. Nat Commun 2022, 13, 4020. [Google Scholar] [CrossRef] [PubMed]
- Svechnikov, K.; Landreh, L.; Weisser, J.; Izzo, G.; Colón, E.; Svechnikov, I.; Söder, O. Origin, development and regulation of human Leydig cells. Hormone Research in Paediatrics 2010, 73, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Yuichi, S.; Kanako, M.; Shogo, H.; Tatsuhiko, A.; Hiroyuki, O.; Takashi, B.; Sawako, M.; Yurina, S.; Haruhiko, A.; Taro, T. Contribution of Leydig and Sertoli cells to testosterone production in mouse fetal testes. Molecular Endocrinology 2013, 27, 63–73. [Google Scholar]
- De Gendt, K.; Swinnen, J. V.; Saunders, P. T. K.; Atanassova, N.; Sharpe, R. M.; Smith, L. B.; Verhoeven, G. A Sertoli cell-selective knockout of the androgen receptor causes spermatogenic arrest in meiosis. Proceedings of the National Academy of Sciences 2004, 101, 1327–1332. [Google Scholar] [CrossRef] [PubMed]
- Willems, A.; Roesl, C.; Mitchell, R. T.; Milne, L.; Jeffery, N.; Smith, S. Sertoli cell androgen receptor signalling in adulthood is essential for post-meiotic germ cell development. Molecular Reproduction & Development 2015, 82, 626–627. [Google Scholar]
- Welsh, M.; Saunders, P. T. K.; Atanassova, N.; Sharpe, R. M.; Smith, L. B. Androgen action via testicular peritubular myoid cells is essential for male fertility. The FASEB Journal 2009, 23, 4218–4230. [Google Scholar] [CrossRef] [PubMed]
- O'Shaughnessy, P. J.; Verhoeven, G.; De Gendt, K.; Monteiro, A.; Abel, M. H. Direct action through the Sertoli cells is essential for androgen stimulation of spermatogenesis. Endocrinology 2010, 151, 2343–2348. [Google Scholar] [CrossRef] [PubMed]
- Qian, T.; Wang, H.; Wang, P.; Geng, L.; Mei, L.; Osakada, T.; Wang, L.; Tang, Y.; Kania, A.; Grinevich, V.; et al. A genetically encoded sensor measures temporal oxytocin release from different neuronal compartments. Nat Biotechnol 2023, 41, 944–957. [Google Scholar] [CrossRef] [PubMed]
- O'Hara, L.; Smith, L. B. Androgen receptor roles in spermatogenesis and infertility. Best Practice & Research Clinical Endocrinology & Metabolism 2015, 29, 595–605. [Google Scholar]
- Knobloch, J.; Reichert, J.; Strauch, J.; Linnemann, L.; Schmidt, M.; Marienfeld, R.; Hennies, S.; Merk, H. F.; Hipfel, R.; Broëer, S. Inhibition of the androgen receptor by antisense oligonucleotides regulates the biological activity of androgens in SZ95 sebocytes. Hormone and Metabolic Research 2013, 45, 149–156. [Google Scholar]
- De Gendt, K.; Verhoeven, G.; Swinnen, J. V.; Saunders, P. T. K.; Atanassova, N.; Sharpe, R. M.; Smith, L. B. Tissue- and cell-specific functions of the androgen receptor revealed through conditional knockout models in mice. Molecular and Cellular Endocrinology 2012, 352, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Chen, W.; Cheng, Z.; Wu, S.; He, J.; Han, L.; He, Z.; Qin, W. Novel Gene Regulation in Normal and Abnormal Spermatogenesis. Cells 2021, 10, 666. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Feng, H. L.; Sandlow, J. I.; Cohen, P.; Moul, J. W.; Schiff, J. D. Comparing Expression of Progesterone and Estrogen Receptors in Testicular Tissue From Men With Obstructive and Nonobstructive Azoospermia. Journal of Andrology 2009, 30, 127–133. [Google Scholar] [CrossRef]
- Farghaly, L. M.; Mohamed, S. F.; Mostafa, R. M. . Expression of estrogen receptors-α in testicular biopsies from nonobstructive azoospermic patients: a histological and immunohistochemical study. Egyptian Journal of Histology 2012, 35, 541–552. [Google Scholar] [CrossRef]
- Hofmann, M.C.; McBeath, E. Sertoli Cell-Germ Cell Interactions Within the Niche: Paracrine and Juxtacrine Molecular Communications. Front Endocrinol (Lausanne) 2022, 13, 897062. [Google Scholar] [CrossRef] [PubMed]
- Ivell, R.; Hartung, S.; Anand-Ivell, R. Insulin-Like Factor 3, Where Are We Now? Endocrine Reviews 2014, 35, 446–477. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, H.; Shiraishi, K.; Sano, N.; Shiraishi, K.; Iwamoto, T.; Watanabe, T.; Maruyama, H.; Uchida, K.; Makino, Y.; Okada, H. Gonadotropin actions on spermatogenesis and hormonal therapies for spermatogenic disorders. Endocrine Journal 2017, 64, 123–131. [Google Scholar]
- Herrera-Luna, C. V.; Scarlet, D.; Walter, I.; Aurich, C. Effect of stallion age on the expression of LH and FSH receptors and aromatase P450 in equine male reproductive tissues. Reproduction, Fertility and Development 2016, 28, 2016–2026. [Google Scholar] [CrossRef]
- Huhtaniemi, I. A short evolutionary history of FSH-stimulated spermatogenesis. Hormones (Athens) 2015, 14, 468–478. [Google Scholar] [CrossRef]
- Michael, S. D.; Curley, C. L.; Han, Z.; Zhu, H.; Wang, R.; McMahon, S. D.; Schauer, D. B. Leukemia Inhibitory Factor-Receptor is Dispensable for Prenatal Testis Development but is Required in Sertoli cells for Normal Spermatogenesis in Mice. Scientific Reports 2018, 8, 11532. [Google Scholar]
- Godfrey, M.S.; Friedman, L.N. Tuberculosis and Biologic Therapies: Anti-Tumor Necrosis Factor-α and Beyond. Clin Chest Med 2019, 40, 721–739. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Kowalewski, M.; Reichler, I. M.; Boos, A.; Kowalewski, M. P.; Reichler, I. G.; Gram, A.; Reichler, I. B. Different expression of leptin and IGF1 in the adult and prepubertal testis in dogs. Reproduction in Domestic Animals 2017, 52, 74–81. [Google Scholar] [CrossRef]
- Yoon, M. J.; Berger, T.; Roser, J. F. Localization of insulin-like growth factor-I (IGF-I) and IGF-I receptor (IGF-IR) in equine testes. Reproduction in Domestic Animals 2011, 46, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Dorji, L.; Ohkubo, T.; Miyoshi, K.; Maeda, T.; Fukui, Y. Gene Expression Differences in Oocytes Derived From Adult and Prepubertal Japanese Black Cattle during In Vitro Maturation. Reproduction in Domestic Animals 2011, 46, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Fan, T.; Xiao, C. TGF-β signaling in health, disease, and therapeutics. Signal Transduct Target Ther 2024, 9, 61. [Google Scholar] [PubMed]
- Weng, B.; Ran, M.; Chen, B.; He, C.; Dong, L.; Peng, F. Genome-wide analysis of long non-coding RNAs and their role in postnatal porcine testis development. Genomics 2017, 109, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Lecker, L.S.M.; Berlato, C.; Maniati E; Delaine-Smith R. ; Pearce O.M.T.; Heath O.; Nichols S.J.; Trevisan C.; Novak M.; McDermott J.; Brenton J.D.; Cutillas P.R.; Rajeeve V.; Hennino A.; Drapkin R.; Loessner D.; Balkwill F.R. TGFBI Production by Macrophages Contributes to an Immunosuppressive Microenvironment in Ovarian Cancer. Cancer Res 2021, 81, 5706–5719. [Google Scholar]
- Chauvigné, F. O.; Parhi, J.; Ollé, J.; Peñaranda, D. S.; Weltzien, F. A.; Wong, T. T.; Pérez, L.; Alvarado, M. V. Dual estrogenic regulation of the nuclear progestin receptor and spermatogonial renewal during gilthead seabream (Sparus aurata) spermatogenesis. Comparative Biochemistry and Physiology Part A, Molecular & Integrative Physiology.
- Wang, Y.; Chen, F.; Ye, L.; Xu, Y.; Li, H.; Xie, Y.; Zhang, Y.; Zhang, L.; Li, X. Steroidogenesis in Leydig cells, effects of aging and environmental factors. Reproduction 2017, 154, R111–r122. [Google Scholar] [CrossRef]
- Braun, B. C.; Okuyama, M. W.; Müller, K.; Ulrich, R.; Rindler, T.; Lux, F.; Eulenburg, V.; Gärtner, U. Steroidogenic enzymes, their products and sex steroid receptors during testis development and spermatogenesis in the domestic cat (Felis catus). Journal of Steroid Biochemistry and Molecular Biology 2018, 178, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Buehr, M.; McLaren, A.; Bartley, A.; Darling, S.; Wylie, C. Proliferation and migration of primordial germ cells in We/We mouse embryos. Cell and Tissue Research 2010, 361, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ge, R.S.; Zirkin, B.R. Leydig cells: from stem cells to aging. Mol Cell Endocrinol 2009, 306, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Beattie, M.; Chen, H.; Fan, J.; Papadopoulos, V.; Miller, P.; Zirkin, B.R. Aging and luteinizing hormone effects on reactive oxygen species production and DNA damage in rat Leydig cells. Biol Reprod 2013, 88, 100. [Google Scholar] [CrossRef]
- Oh, Y. S.; Song, S. Y.; Sohn, H. E.; Ahn, H. C.; Kim, J. Y.; Choe, Y. S.; Suh, Y. H.; Moon, Y. S.; Kim, J. H.; Song, J. H. Expression of cubilin in mouse testes and Leydig cells. Andrologia 2015, 48, 325–332. [Google Scholar] [CrossRef]
- Shi, L.; Wang, X.; Duan, Y.; Li, K.; Ren, Y. Antagonistic effects of selenium on lead-induced oxidative stress and apoptosis of Leydig cells in sheep. Theriogenology 2022, 185, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Groeger, G.; Quiney, C.; Cotter, T. G. Hydrogen peroxide as a cell-survival signaling molecule. Antioxidants & Redox Signaling 2009, 11, 2655–2671. [Google Scholar]
- Morielli, T.; O'Flaherty, C. Oxidative stress impairs function and increases redox protein modifications in human spermatozoa. Fertility and Sterility 2015, 103, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Jian, Z.; Lingling, Z.; Zheng, L.; Rui, Y.; Xiaoyan, W.; Qiyuan, Y. Leptin level and oxidative stress contribute to obesity-induced low testosterone in murine testicular tissue. Oxidative Medicine and Cellular Longevity 2014, 2014, 190945. [Google Scholar]
- Vladimir, E.; Anna, M.; Anatoly, V.; Natalia, V.; Pavel, S. Formation of long-lived reactive species of blood serum proteins induced by low-intensity irradiation of helium-neon laser and their involvement in the generation of reactive oxygen species. Journal of Photochemistry and Photobiology B, Biology.
- Recknagel, R. O.; Glende, E. A.; Britton, R. S. Free Radical Damage and Lipid Peroxidation. Hepatotoxicology 2020, 2, 165–196. [Google Scholar]
- Zhang, X.; Wang, T.; Deng, T.; Wang, L.; Jing, W.; Wang, R. Damaged spermatogenic cells induce inflammatory gene expression in mouse Sertoli cells through the activation of Toll-like receptors 2 and 4. Molecular and Cellular Endocrinology 2013, 365, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Winters, S. J.; Moore, J. P.; Clark, B. J. Leydig cell insufficiency in hypospermatogenesis, a paracrine effect of activin–inhibin signaling? Andrology 2018, 6, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Sampath, H. Mitochondrial DNA Integrity, Role in Health and Disease. Cells 2019, 8, 100. [Google Scholar] [CrossRef]
- Hanukoglu, I. Antioxidant Protective Mechanisms against Reactive Oxygen Species (ROS) Generated by Mitochondrial P450 Systems in Steroidogenic Cells. Drug Metabolism Reviews 2017, 49, 88–110. [Google Scholar] [CrossRef]
- Camacho, L.; Latendresse, J. R.; Muskhelishvili, L.; Ton, T. V.; Bishop, J. B.; Delclos, K. B. Effects of acrylamide exposure on serum hormones, gene expression, cell proliferation, and histopathology in male reproductive tissues of Fischer 344 rats. Toxicology Letters 2012, 211, 135–143. [Google Scholar] [CrossRef]
- Midzak, A. S.; Akula, N.; Huang, X.; Papadopoulos, V. Mitochondrial regulation of Leydig cell steroid synthesis. Implications for the decline in male testosterone production with advancing age. Molecular and Cellular Endocrinology 2009, 300, 89–96. [Google Scholar]
- Monageng, E.; Offor, U.; Takalani, N. B.; Makgotlho, E.; Ngole, E. M.; Shai, L. J.; Aigbe, F. R.; Omoruyi, S. I.; Oladimeji, O.; Obi, P. A. A Review on the Impact of Oxidative Stress and Medicinal Plants on Leydig Cells. Antioxidants (Basel) 2023, 12, 1151. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Hu, G.; Dong, B.; Zhang, H.; Zhang, Z.; Wang, W.; Li, Y.; Yang, F.; Chen, Z. Roscovitine protects murine Leydig cells from lipopolysaccharide-induced inflammation. Experimental and Therapeutic Medicine 2017, 13, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.J.; Chen, J.B.; Cao, J.P.; Li, X.; Sun, C.D. Citrus flavonoids and their antioxidant evaluation. Crit Rev Food Sci Nutr 2022, 62, 3833–3854. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, J.; Yu, H.; Zhuang, J.; Lu, Y.; Wang, Z. Elemental selenium at nano size possesses lower toxicity without compromising the fundamental effect on selenoenzymes, comparison with selenomethionine in mice. Free Radical Biology and Medicine 2007, 42, 1524–1533. [Google Scholar] [CrossRef] [PubMed]
- Hu, X. T.; Ding, C.; Zhou, N.; Xu, W.; Li, J.; Luo, Y.; Wu, X. Quercetin protects gastric epithelial cell from oxidative damage in vitro and in vivo. Journal of Cellular Biochemistry 2015, 116, 754–764. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B.; Chen, Z.; Hu, H.; Huang, J.; Lu, Z. Plant flavonoids, Classification, distribution, biosynthesis, and antioxidant activity. Food Chemistry 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Asensi-Fabado, M. A.; Munne-Bosch, S. Vitamins in plants, occurrence, biosynthesis and antioxidant function. Trends in Plant Science 2010, 15, 582–592. [Google Scholar] [CrossRef]
- Marmitt, D. J.; Bitencourt, S.; da Silva, G. R.; da Silva, B. S.; Correa, M. C.; Goncalves, W. F.; Cunha, M. E.; Bresciani, T.; Gasperin, B. G.; Dalsenter, P. R. Traditional plants with antioxidant properties in clinical trials-A systematic review. Phytotherapy Research 2021, 35, 5647–5667. [Google Scholar] [CrossRef]
- Alharbi, S. A.; Asad, M.; Abdelsalam, K. E. A.; Saeed, M. M.; Majid, S.; Al-Sheikh, Y. A.; Bukhari, S. I.; Siddiq, A.; Alroqi, F. R. Frankincense extract protects against testicular damage through augmentation of antioxidant defense mechanisms and modulation of apoptotic genes expression. Scientific Reports 2022, 12, 12625. [Google Scholar] [CrossRef]
- Roy, N. K.; Parama, D.; Banik, K.; Bordoloi, D.; Devi, A. K.; Thakur, K. K.; Padmavathi, G.; Venugopal, R.; Fan, L.; Sethi, G. An Update on Pharmacological Potential of Boswellic Acids against Chronic Diseases. International Journal of Molecular Sciences 2019, 20, 4101. [Google Scholar] [CrossRef] [PubMed]
- Okano, S.; Honda, Y.; Kodama, T.; Yasuda, M.; Masuko, K.; Tanaka, M.; Morimoto, C.; Sakagami, H.; Hatano, T.; Nakano, M. The Effects of Frankincense Essential Oil on Stress in Rats. Journal of Oleo Science 2019, 68, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Li, M. Y.; Zhu, X. L.; Zhao, B. X.; Wang, H. X.; Cheng, W. J.; Wei, H. Adrenomedullin alleviates the pyroptosis of Leydig cells by promoting autophagy via the ROS-AMPK-mTOR axis. Cell Death & Disease 2019, 10, 489. [Google Scholar]
- Gao, F.; Li, G.; Liu, C.; Gao, L.; Wang, H.; Liu, W.; Ding, Z. Autophagy regulates testosterone synthesis by facilitating cholesterol uptake in Leydig cells. Journal of Cell Biology 2018, 217, 2103–2119. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Song, A.; Hu, W.; Dai, M.; Hou, C.; Jin, Y.; Xia, W.; Zhang, X.; Liu, Z.; Song, X. The Anti-atherosclerotic Effect of Paeonol against Vascular Smooth Muscle Cell Proliferation by Up-regulation of Autophagy via the AMPK/mTOR Signaling Pathway. Frontiers in Pharmacology 2017, 8, 948. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Shi, L.; Li, M. Y.; Zhang, L. N.; Cheng, W. J.; Wei, H. Adrenomedullin protects Leydig cells against lipopolysaccharide-induced oxidative stress and inflammatory reaction via MAPK/NF-κB signalling pathways. Scientific Reports 2017, 7, 16479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fang, Y.; Tang, D.; Kang, R.; Yao, X.; Meng, X.; Xiao, X. Activation of MT1/MT2 to Protect Testes and Leydig Cells against Cisplatin-Induced Oxidative Stress through the SIRT1/Nrf2 Signaling Pathway. Cells 2022, 11, 1615. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; He, X.; Zhuang, M.; Niu, R.; Song, Y.; Shen, Z.; Wang, Y.; Zou, W. Melatonin Ameliorates Busulfan-Induced Spermatogonial Stem Cell Oxidative Apoptosis in Mouse Testes. Antioxidants & Redox Signaling 2018, 28, 385–400. [Google Scholar]
- Xiong, Y.; Li, J.; He, S. Zinc Protects against Heat Stress-Induced Apoptosis via the Inhibition of Endoplasmic Reticulum Stress in TM3 Leydig Cells. Biological Trace Element Research 2022, 200, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Liu, X.; Pan, Z.; Xia, L.; Zhang, Y.; Liu, S.; Zheng, Y.; Yu, Z. Zinc deficiency and cellular oxidative stress, prognostic implications in cardiovascular diseases. Acta Pharmacologica Sinica 2018, 39, 1120–1132. [Google Scholar] [CrossRef]
- Santos, H. O.; Teixeira, F. J. Use of medicinal doses of zinc as a safe and efficient coadjutant in the treatment of male hypogonadism. Aging Male 2020, 23, 669–678. [Google Scholar] [CrossRef]
- Wagner, M. S.; Wajner, S. M.; Maia, A. L. The role of thyroid hormone in testicular development and function. Journal of Endocrinology 2008, 199, 351–365. [Google Scholar] [CrossRef] [PubMed]
- O'Shaughnessy, P.J.; Monteiro, A.; Verhoeven, G.; De Gendt, K.; Abel, M.H. Effect of FSH on testicular morphology and spermatogenesis in gonadotrophin-deficient hypogonadal mice lacking androgen receptors. Reproduction.
- Gao Y, Lee WM, Cheng CY. Thyroid hormone function in the rat testis. Front Endocrinol (Lausanne) 2014, 5, 188. [Google Scholar]
- Manna, P. R.; Kero, J.; Tena-Sempere, M.; Pakarinen, P.; Stocco, D. M.; Huhtaniemi, I. Regulation of the steroidogenic acute regulatory protein, steroidogenesis, and luteinizing hormone receptor function by thyroid hormone in mouse Leydig cells. Endocrinology 2001, 142, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Mitra, S.; Lakhera, P. C.; Sen, P. N-acetylcysteine effectively mitigates cadmium-induced oxidative damage and cell death in Leydig cells in vitro. Drug and Chemical Toxicology 2016, 39, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, S.; Lin, S.; Li, L.; Lu, X.; Chen, W.; Wang, Y.; Zhang, L.; Zhao, R.; Wang, H. Melatonin ameliorates diabetic hyperglycaemia-induced impairment of Leydig cell steroidogenic function through activation of SIRT1 pathway. Reproductive Biology and Endocrinology 2022, 20, 117. [Google Scholar] [CrossRef] [PubMed]
- Balmik, A. A.; Chinnathambi, S. Multi-Faceted Role of Melatonin in Neuroprotection and Amelioration of Tau Aggregates in Alzheimer’s Disease. Journal of Alzheimer's Disease.
- Matos, M. H.; Jenny, S.; Mark, J.; King, N. J. C.; Handelsman, D. J.; Allan, C. M. Melatonin protects against cisplatin-induced ovarian damage in mice via the MT1 receptor and antioxidant activity. Biology of Reproduction 2017, 96, 1244–1255. [Google Scholar]
- Jang, H.; Lee, O. H.; Lee, Y.; Kwon, J. Melatonin prevents cisplatin-induced primordial follicle loss via suppression of PTEN/AKT/FOXO3a pathway activation in the mouse ovary. Journal of Pineal Research 2016, 60, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; He, X.; Zhuang, M.; Niu, R.; Song, Y.; Shen, Z.; Wang, Y.; Zou, W. Melatonin Ameliorates Busulfan-Induced Spermatogonial Stem Cell Oxidative Apoptosis in Mouse Testes. Reproductive Sciences 2017, 24, 240–252. [Google Scholar] [CrossRef]
- Jambor, T.; Zajickova, T.; Arvay, J.; Kisucka, L.; Ondicova, K.; Lovasova, E.; Capcarova, M. Exceptional Properties of Lepidium sativum L. Extract and Its Impact on Cell Viability, ROS Production, Steroidogenesis, and Intracellular Communication in Mice Leydig Cells In Vitro. Molecules 2022, 27, 5146. [Google Scholar] [CrossRef]
- Lai, X.; Cao, M.; Song, X.; Zhu, W.; Liu, Y.; Wei, X.; Feng, Z. Resveratrol promotes recovery of immune function of immunosuppressive mice by activating JNK/NF-κB pathway in splenic lymphocytes. Canadian Journal of Physiology and Pharmacology 2017, 95, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Donald, E. L.; Stojanovska, L.; Apostolopoulos, V.; Nurgali, K. Resveratrol alleviates oxidative damage in enteric neurons and associated gastrointestinal dysfunction caused by chemotherapeutic agent oxaliplatin. Maturitas 2017, 105, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y. H.; Zhou, L. Y.; Chen, Q. Z.; Yang, Y. J.; Liu, L. Resveratrol inactivates PI3K/Akt signaling through upregulating BMP7 in human colon cancer cells. Oncology Reports 2017, 37, 3572–3582. [Google Scholar] [CrossRef] [PubMed]
- Maier-Salamon, A.; Böhmdorfer, M.; Thalhammer, T.; Reznicek, G.; Kunert, O.; Szekeres, T.; Riha, J.; Jaeger, W. Hepatic Glucuronidation of Resveratrol, Interspecies Comparison of Enzyme Kinetic Profiles in Human, Mouse, Rat, and Dog. Drug Metabolism and Pharmacokinetics 2011, 26, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Kalantari, H.; Das, D. K. Physiological effects of resveratrol. Biofactors 2010, 36, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Emilia, J. M.; Eulalia, G. P.; Thais, M.; Manuel, C.; María, M. G.; Mariana, M.; Marta, R. F. trans-Resveratrol, a natural antioxidant from grapes, increases sperm output in healthy rats. Journal of Nutrition 2010, 140, 757–763. [Google Scholar]
- Zeng, Q.; Xiong, Q.; Zhou, M.; Tian, X.; Yue, K.; Li, Y.; Shu, X.; Ru, Q. Resveratrol attenuates methamphetamine-induced memory impairment via inhibition of oxidative stress and apoptosis in mice. J Food Biochem 2021, 45, e13622. [Google Scholar] [CrossRef]
- Ourique, G. M.; Finamor, I. A.; Saccol, E. M. H.; Schneider, J. A.; Gutierrez, K.; Ferreira, M. R. Resveratrol improves sperm motility, prevents lipid peroxidation and enhances antioxidant defenses in the testes of hyperthyroid rats. Reproductive Toxicology 2013, 37, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Bucak, M. N.; Tuncer, P. B.; Sariözkan, S.; Ulutas, P. A.; Akcay, E.; Tekin, N.; Buyukleblebici, S.; Tasdemir, U.; Coyan, K. Lycopene and resveratrol improve post-thaw bull sperm parameters, sperm motility, mitochondrial activity and DNA integrity. Andrologia 2014, 46, 785–791. [Google Scholar] [CrossRef]
- Liu, S.; Sun, Q.; Li, Z. Resveratrol protects Leydig cells from nicotine-induced oxidative damage through enhanced autophagy. Clinical and Experimental Pharmacology and Physiology 2017, 44, 316–324. [Google Scholar] [CrossRef]
- Marti, N.; Bouchoucha, N.; Sauter, K. S.; Flück, C. E. Resveratrol inhibits androgen production of human adrenocortical H295R cells by lowering CYP17 and CYP21 expression and activities. PLoS ONE 2017, 12, e0174224. [Google Scholar] [CrossRef] [PubMed]
- Svechnikov, K.; Spatafora, C.; Svechnikova, I.; Tringali, C.; Söder, O. Effects of resveratrol analogs on steroidogenesis and mitochondrial function in rat Leydig cells in vitro. Journal of Applied Toxicology 2010, 30, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Silva, E. C. B.; Cajueiro, J. F. P.; Silva, S. V.; Soares, P. C.; Guerra, M. M. P. Effect of antioxidants resveratrol and quercetin on in vitro evaluation of frozen ram sperm. Theriogenology 2012, 77, 1722–1726. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y. C.; Nagpal, M. L.; Stocco, D. M.; Lin, T. Effects of genistein, resveratrol, and quercetin on steroidogenesis and proliferation of MA-10 mouse Leydig tumor cells. Journal of Endocrinology 2007, 192, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, M. Z.; Shen, Y.; Tang, R.; Xiao, L.; Zhao, J. Lycopene Prevents DEHP-Induced Leydig Cell Damage with the Nrf2 Antioxidant Signaling Pathway in Mice. Journal of Agricultural and Food Chemistry 2020, 68, 2031–2040. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, J.; Luo, L.; Baumbach, G. L.; Shi, R.; Zirkin, B. R. Vitamin E, aging and Leydig cell steroidogenesis. Experimental Gerontology 2005, 40, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Omolaoye, T. S.; Cyril, A. C.; Radhakrishnan, R.; Adegoke, E. O.; Otulana, J. O.; Du Plessis, S. S. The Effect of Statins on Male Reproductive Parameters, A Mechanism Involving Dysregulation of Gonadal Hormone Receptors and TRPV1. International Journal of Molecular Sciences 2023, 24, 10322. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, A.; Sun, Y.; Wei, X.; Zhu, X.; Jin, Y.; Wang, L. Sirt1 exerts anti-inflammatory effects and promotes steroidogenesis in Leydig cells. Fertility and Sterility 2012, 98, 194–199. [Google Scholar] [CrossRef]
- Cormier, M.; Ghouili, F.; Roumaud, P.; Martin, L. J. Influences of flavones on cell viability and cAMP-dependent steroidogenic gene regulation in MA-10 Leydig cells. Cell Biology and Toxicology 2018, 34, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Zhu, X.; Zhang, X.; Jiang, H.; Li, B.; Wang, Z.; Li, D.; Jin, Y. Alpha-naphthoflavone attenuates non-alcoholic fatty liver disease in oleic acid-treated HepG2 hepatocytes and in high fat diet-fed mice. Biomed Pharmacother 2019, 118, 109287. [Google Scholar] [CrossRef]
Figure 1.
Both exogenous and endogenous ROS disturbed the regulation of metabolic hormone, hormonal or immune stress, and H-P-T axis, and then impact on Leydig cell function. TLRs: Toll-like receptors; NLRs: Nod-like receptors; ROS: reactive oxygen species; GnRH: Gonadotropin releasing hormone; FSH: Follicle stimulating hormone; LH: Luteinizing hormone; T3: Tri-iodothyronine; H-P-T axis: Hypothalamus-pituitary gland-Testes axis; Nrf: Nucleus factor-E2 related factor; Stir1: Silent information regulator of transcription 1; MAPK: Mitogen activated protein kinase.
Figure 1.
Both exogenous and endogenous ROS disturbed the regulation of metabolic hormone, hormonal or immune stress, and H-P-T axis, and then impact on Leydig cell function. TLRs: Toll-like receptors; NLRs: Nod-like receptors; ROS: reactive oxygen species; GnRH: Gonadotropin releasing hormone; FSH: Follicle stimulating hormone; LH: Luteinizing hormone; T3: Tri-iodothyronine; H-P-T axis: Hypothalamus-pituitary gland-Testes axis; Nrf: Nucleus factor-E2 related factor; Stir1: Silent information regulator of transcription 1; MAPK: Mitogen activated protein kinase.
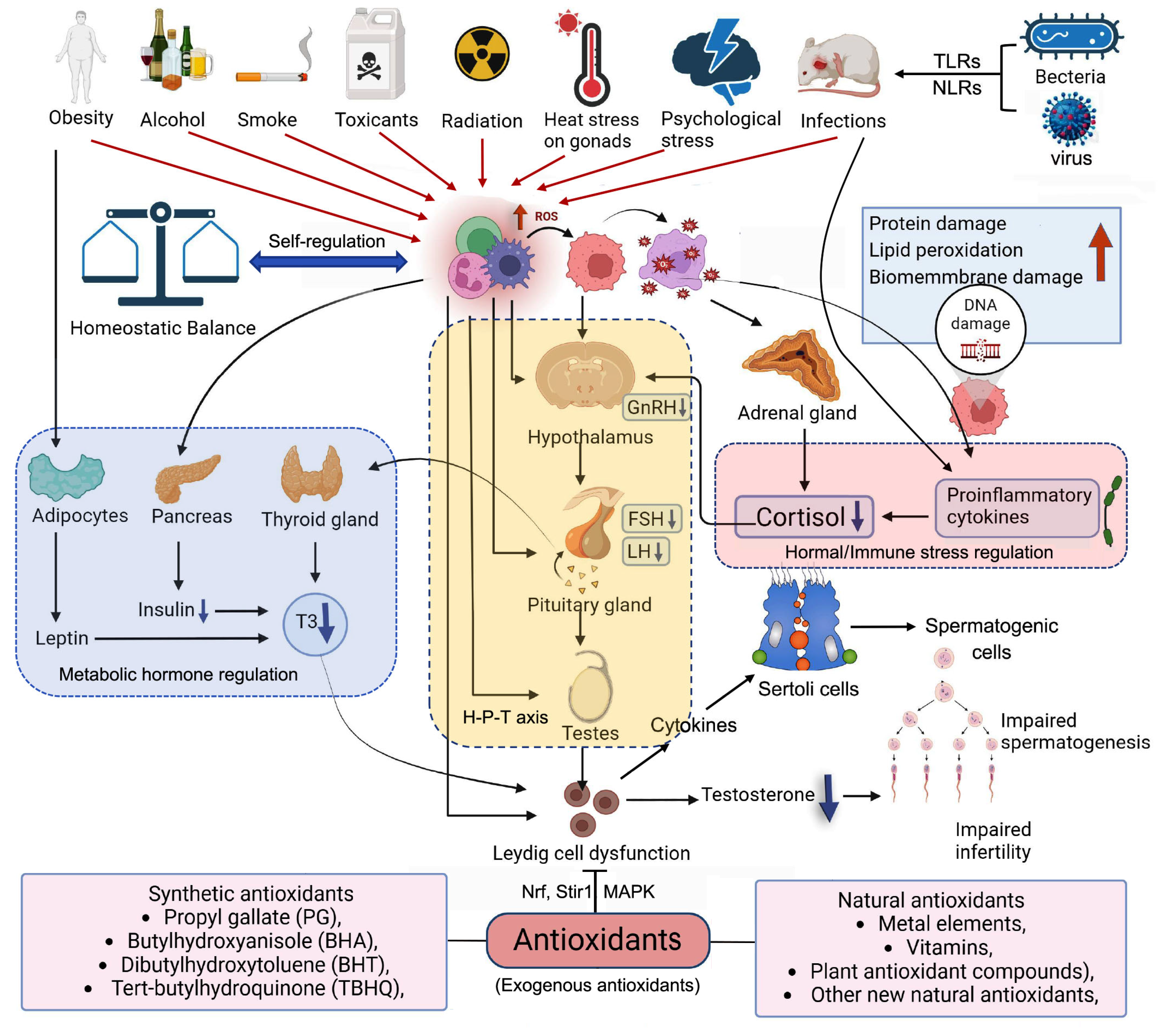
Figure 2.
ROS disturb the process of testosterone synthesis. CYP: cytochrome P; 3β-HSD2:3-beta-hydroxy steroid dehydrogenases 3; 17β-HSD3:17-beta-hydroxy steroid dehydrogenases; StAR: steroidogenic acute regulatory protein; TSPc: Close status of translocator protein; TSPo: Open status of translocator protein. ↓:ROS inhibit the transcript of StAR;↓:ROS inhibit the transcript of CYP11/17A1(NADPH);↓:ROS inhibit the transcript of 3β-HSD2(NAD+).
Figure 2.
ROS disturb the process of testosterone synthesis. CYP: cytochrome P; 3β-HSD2:3-beta-hydroxy steroid dehydrogenases 3; 17β-HSD3:17-beta-hydroxy steroid dehydrogenases; StAR: steroidogenic acute regulatory protein; TSPc: Close status of translocator protein; TSPo: Open status of translocator protein. ↓:ROS inhibit the transcript of StAR;↓:ROS inhibit the transcript of CYP11/17A1(NADPH);↓:ROS inhibit the transcript of 3β-HSD2(NAD+).
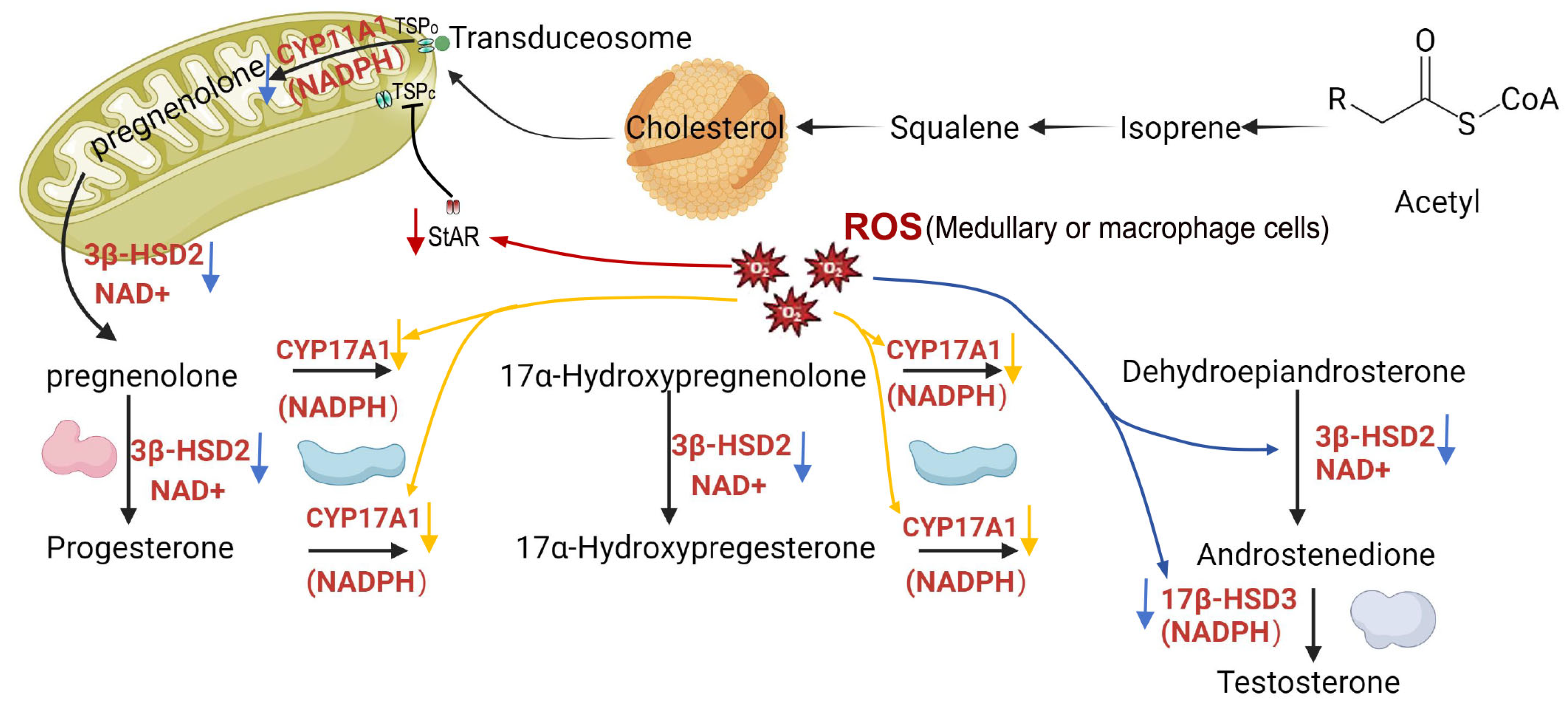
Figure 3.
Endocrine and paracrine secretion of Leydig cell paly a important role in the biological process of spermatogenesis. AR: Androgen receptor; GATA4: GATA binding protein 4; GNDF: glial cell derived neurotrophic factor; INSL3: Insulin-like factor 3; PMOC: Pro-opiomelanocortin; CSF: Colony stimulated factor; FSH: Follicle simulated factor; LH: luteinizing hormone; LIF: Leukemia inhibitory factor; Tc: The tight conjunction between Sertoli and Sertoli cell; Gc: Gap conjunction between Sertoli and Sertoli cel.
Figure 3.
Endocrine and paracrine secretion of Leydig cell paly a important role in the biological process of spermatogenesis. AR: Androgen receptor; GATA4: GATA binding protein 4; GNDF: glial cell derived neurotrophic factor; INSL3: Insulin-like factor 3; PMOC: Pro-opiomelanocortin; CSF: Colony stimulated factor; FSH: Follicle simulated factor; LH: luteinizing hormone; LIF: Leukemia inhibitory factor; Tc: The tight conjunction between Sertoli and Sertoli cell; Gc: Gap conjunction between Sertoli and Sertoli cel.
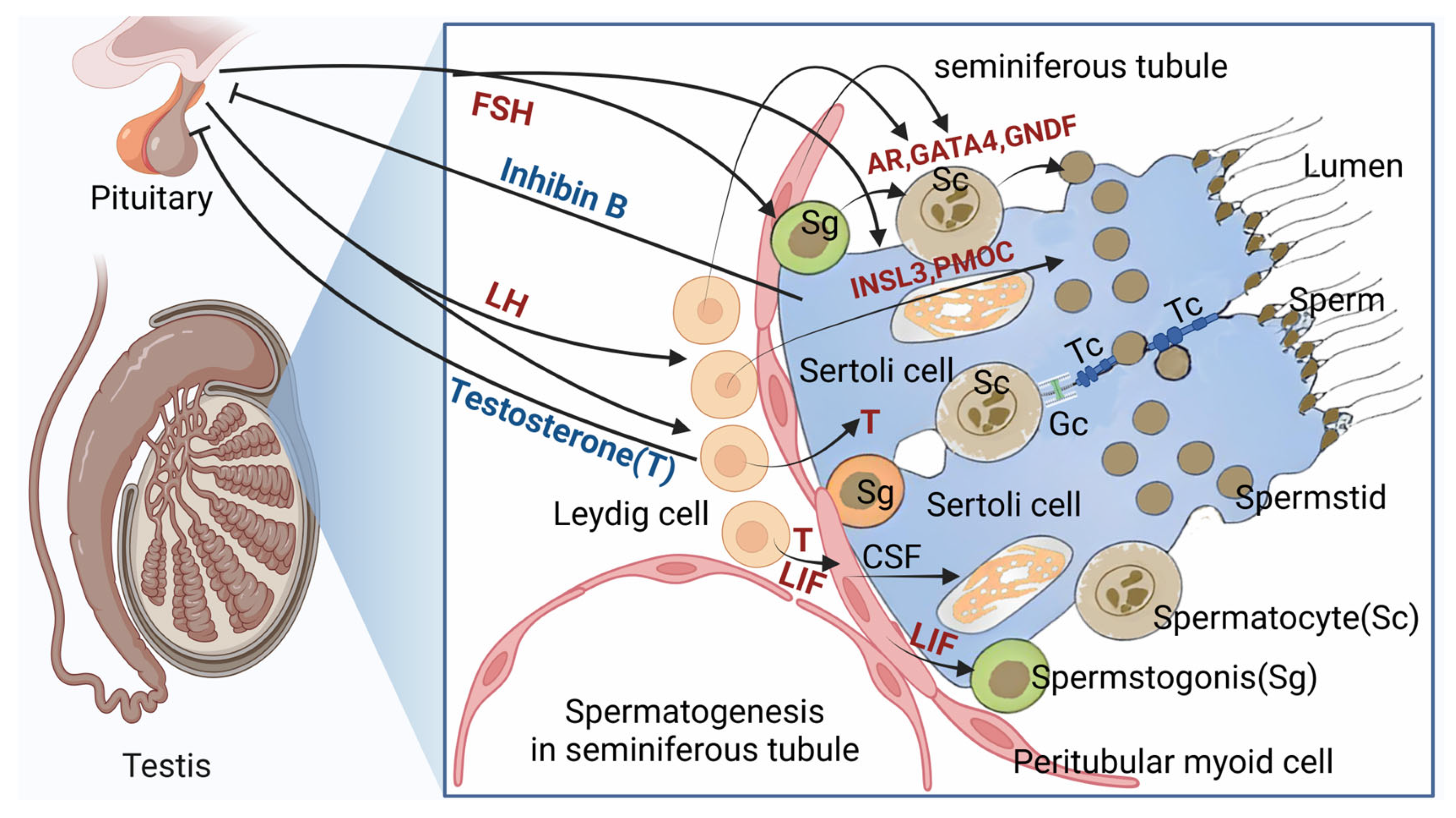
Figure 4.
SIRT1 is a hub in these antioxidative stress pathways. P65/53: lymphocyte protein tyrosine kinase56/53; NF-kB: Nuclear factor-k-gene binding; TSC2: Tuberous sclerosis complex 2; LKB1: Liver kinase B1; mTOR: Mechanistic target of rapamycin; AMPK: (AMP)-activated protein kinase; PGC1α: peroxisome proliferator-activated receptor γ coactivator 1α; FOXO: forkhead box O.
Figure 4.
SIRT1 is a hub in these antioxidative stress pathways. P65/53: lymphocyte protein tyrosine kinase56/53; NF-kB: Nuclear factor-k-gene binding; TSC2: Tuberous sclerosis complex 2; LKB1: Liver kinase B1; mTOR: Mechanistic target of rapamycin; AMPK: (AMP)-activated protein kinase; PGC1α: peroxisome proliferator-activated receptor γ coactivator 1α; FOXO: forkhead box O.
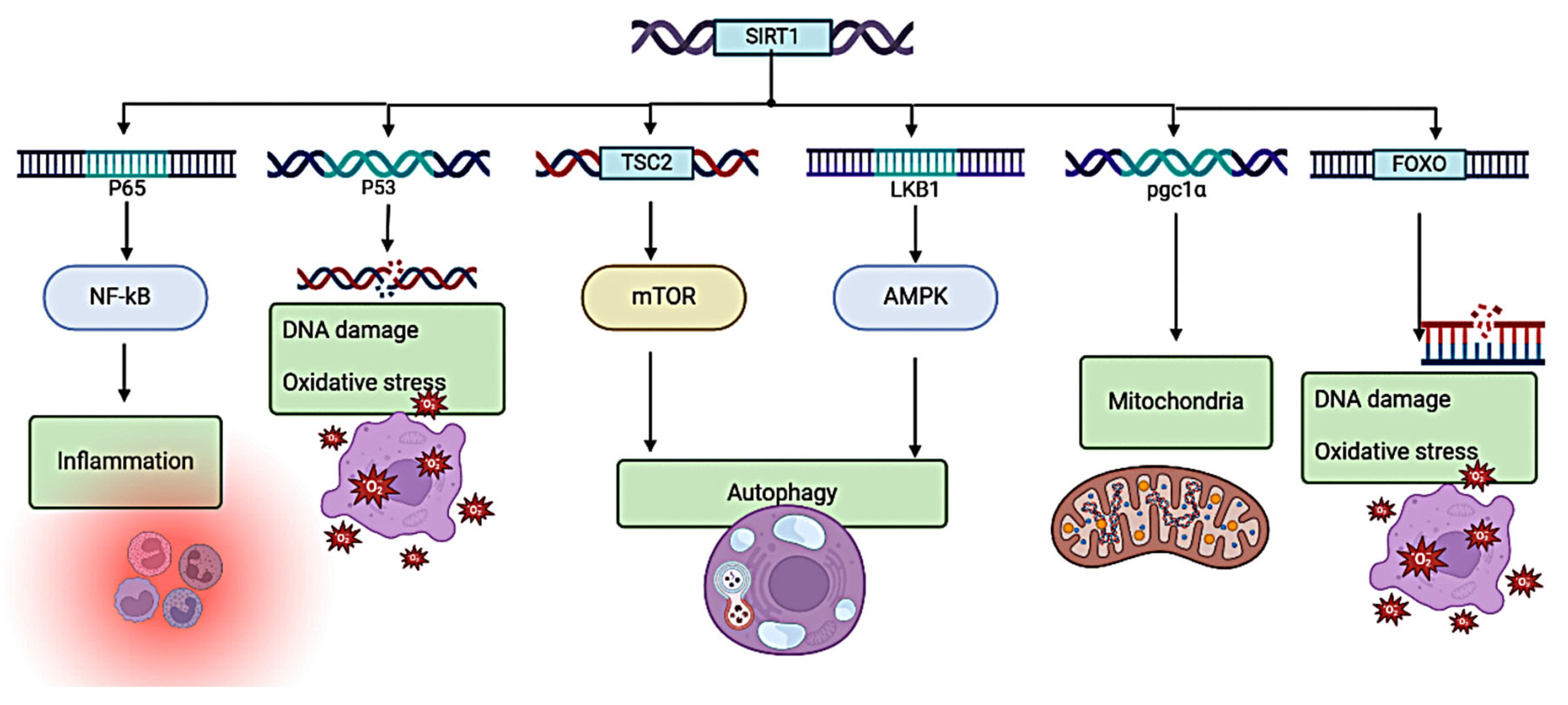

Table 1.
The receptors on Leydig cell associated with spermatogenesis.
| Receptors/ Cytokines |
Species | Characteristic | cited document |
| AR | Humans, Rats, Dogs | Synergizes with androgens to regulate spermatogenesis | [36,38,39,40,41] |
| ER | Human, Rat | Maintains sperm count and function | [42,43,44] |
| LGRS | Rats | Facilitates the stimulatory effect of INSL3 on cAMP production during spermatogenesis. | [45,46] |
| OTR | Human, monkey | Modulates oxytocin levels within the male reproductive tract. | [38] |
| LHR | Human, rat, flounder | Synergizes with LH to promote testosterone production | [47] |
| FSHR | Human, Rat, Stallion | Mediates the effects of follicle stimulating hormone on spermatogenesis | [47,48,49] |
| LIFR | Rats | Maintenance of germ cell numbers, sperm motility, seminiferous tubule structure and steroid formation | [50] |
| TNFR | Rats | Harmonizes spermatogenesis | [51] |
| IGFR | Rats, Horses, Dogs | Regulates testicular function and germ cell development | [52,53,54] |
| TGF-βR | Human, Rat, Boar | Controls meiosis and early spermatogenesis | [55,56,57] |
| PGR | Humans, cats, sea bream | Regulates the role of progesterone in spermatogenesis | [58,59,60] |
| KIT | Rats | Promotes proliferation, migration and postnatal spermatogenesis of primordial germ cells | [61,62,63] |
| Cubn | Rats | Facilitates endocytosis and nutrient uptake in both germ cells and somatic cells, essential for the sustenance of spermatogenesis. | [64] |
Table 2.
Classification of antioxidant substances.
| Source | Species | Characteristic | Reference |
| Synthetic antioxidants | Propyl gallate (PG), Butylhydroxyanisole (BHA), Dibutylhydroxytoluene (BHT), tert-butylhydroquinone (TBHQ), |
Although it has a strong antioxidant capacity, it is capable of producing hydrogen peroxide at physiological pH and temperature. Hydrogen peroxide itself is highly oxidizing and quite toxic to cells. | [80] |
| Natural antioxidants | Metal elements: copper, iron, zinc and manganese | They are cofactors of a series of important antioxidant enzymes, binding with enzymes to make enzymes with strong catalytic activity, by catalyzing superoxide radicals, thus eliminating ROS and preventing their damage to proteins, DNA and lipids. | [81,82] |
| Vitamins: Mainly vitamins A, B, C, E and K. | It not only has strong antioxidant ability, but also has a good protective effect on the structure of biological membranes. | [83,84] | |
| plant antioxidant compounds: mainly polysaccharides, alkaloids, polyphenols, flavonoids and saponins five kinds of compounds. | [85,86,87,88,89,90,91] [92,93] |
||
| Other new natural antioxidants: Astaxanthin and antioxidant peptides | Antioxidant substances capable of traversing the blood-brain barrier can neutralize free radicals within the body, thereby rejuvenating cellular function and promoting overall organism vitality. | [94,95,96] |
Table 3.
Antioxidative stress substances for Leydig cells.
| Antioxidant | Oxidative inducer | Animals | Effect | Pathway | Reference | |
| In Vivo Experiments | Adrenomedullin, N-Acetylcysteine |
LPS | Rats | Promoted cellular autophagy and attenuated focal death of LPS-exposed leydig cells | ROS-AMPK -mTOR |
[88,101] |
| Adrenomedullin, N-Acetylcysteine |
LPS | Rats | Reduction of LPS-induced ROS overproduction | MAPK / NF-κB |
[78,79,88,101] | |
| Melatonin | Cis-platinum | Mouse | Through the MT1/MT2 melatonin receptors, melatonin modulates the Sirt1/Nrf2 signaling axis, serving as a pivotal protective agent against cisplatin-induced apoptosis in mouse mesangial cells. | Sirt1 / Nrf2 |
[92,102,103,104,105,106] | |
| Lepidium sativum L | Aluminum oxide | Rats | Significantly reduced levels of ROS production and increased sperm motility and viability in mice | — | [107] | |
| Resveratrol | H2O2 | Mice | Resveratrol safeguards against H2O2-triggered oxidative stress, thereby mitigating the decline in Leydig cell viability and functional proficiency. Lower concentrations of resveratrol had cytoprotective effects on oxidatively stressed leydig cells. | Nrf2- ARE |
[108,109,110,111,112,113,114,115,116,117,118,119,120,121] | |
| Lycopene | Diethylphosphite | Mice | Lycopene is capable of enhancing antioxidant potential by modulating the Nrf2 signaling pathway, subsequently alleviating DEHP-induced damage to Leydig cells. | Nrf2 pathway |
[122] | |
| Cellular experiments | Vitamin E | Environment | Rats | Vitamin E exhibits a notable protective effect against oxidant-mediated lipid peroxidation in cultured Leydig cells, as well as preserving the cells' capacity to produce testosterone in vitro. | — | [123] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



