
Submitted:
23 October 2024
Posted:
24 October 2024
You are already at the latest version
Abstract
Obesity is a complex disease increasing risk of other pathologies. Its prevention or long-term weight loss maintenance is problematic. Gut microbiome is considered a potential obesity modulator. The objective of the present study was to summarize recent findings regarding relationships between obesity, gut microbiota and diet, with emphasis on dietary fiber and resistant starch. Composition of the human gut microbiome and methods of its quantification are described. Products of the gut microbiome metabolism, such as short-chain fatty acids and secondary bile acids including their effects on the gut microbiota, intestinal barrier function and immune homeostasis are discussed in the context of obesity. Importance of dietary fiber and resistant starch is emphasized as far as effects of the host diet on the composition and function of the gut microbiome are concerned. Complex relationships between human gut microbiome and obesity are finally summarized.
Keywords:
secondary bile acids
; short-chain fatty acids
; butyrate
; dietary fiber
; resistant starch
; intestinal barrier function
; immune homeostasis
1. Introduction
The most general definition of obesity is based on the BMI (Body Mass Index; weight in kg divided by the square of height in m) over 30 kg/m2 [1]. Obesity is a consequence of effects of multiple factors, including genetics, excessive energy intake, sedentary lifestyle, socioeconomic status, poor sleep quality, parental weight or environment [2]. It is a complex disease increasing risk of many other pathologies, such as ischemic heart disease, stroke, type 2 diabetes (T2D), hypertension, hyperlipidemia or also several types of cancer [3]. However, there is no effective, well-defined, evidence-based intervention for preventing obesity [4]. Attempts to maintain weight loss long-term, including lifestyle interventions aiming to decrease energy intake on the one hand and increase physical activities on the other hand are mostly unsuccessful [5]. Nevertheless, due to its epidemic proportions (the current worldwide prevalence in women and men is nearly 15 % and nearly 11 %, respectively), the treatment of obesity is of an utmost importance [6].
Another potential obesity modulator is currently considered gut microbiome [6,7,8,9]. In this context, review articles are available regarding general aspects of the gut microbiota [10], its principal metabolites, such as short chain fatty acids (SCFAs; [11,12,13]) and bile acids (BAs; [14,15,16]), and dietary components with a potential to modulate the gut microbiome: dietary fiber (DF; [17,18] or resistant starch (RS; [19]). The present study summarizes the principal findings described in a more detail in the above-mentioned articles, with the objective to indicate possibilities to prevent obesity via DF/RS intake and modulation of the gut microbiome composition, including its metabolites SCFAs and BAs.
2. Human Gut Microbiome Composition
According to a phylogenetic tree based on the rRNA data, Bacteria are one of the three domains (apart from Archaea and Eukaria) in the hierarchical taxonomic rank domain – kingdom – phylum – class – order – family – genus – species [20]. Based on the analysis of the human gut microbiome, the most frequently represented phyla here are Bacteroidetes (synonym Bacteroidota) and Firmicutes (synonym Bacillota), which comprise 90 % of all human gut phyla; the remaining 10 % represent Actinobacteria (synonym Actinomycetota), Proteobacteria, Fusobacteria and Verrucomicrobia [10].
Firmicutes are constituted mostly by the gram-positive genera, Clostridium being the most abundant genera (95 %), followed by Lactobacillus, Bacillus, Enterococcus, and Ruminococcus [21]. On the other hand, Bacteroidetes are gram-negative bacteria with the most abundant genera Bacteroides and Prevotella followed by Parabacteroides and Alistipes [22], as far as Actinobacteria are concerned, the main representative on the genus level is Bifidobacterium [23].
Following factors can influence an individual gut microbial composition during the whole life: preterm or full term born; vaginal or caesarean delivery; nursing by human milk or a substitute [24,25,26]; BMI; exercise; diet [21,24,27,28] and past illnesses including the use of drugs and antibiotics [29]. It follows from the metagenomic data of Rothschild et al. (2018) that the diet and environment affect composition of the human gut microbiome in healthy individuals more than the host genetics [30].
Moreover, the question of the stability of the human gut microbiome in the adult individuals is still not fully resolved. Some studies identified three distinct clusters of balanced microbial states called enterotypes, dominated by Bacteroides (enterotype I), Prevotella (enterotype II) and Ruminococcus (enterotype III), respectively. The bacterial communities of the different enterotypes can possibly generate energy by either fermentation of carbohydrates (enterotype I) or from mucin glycoproteins (enterotypes I and III) and can also differ regarding level of vitamin production [23]. On the other hand, the data of Jeffery et al. (2012) or Knights et al. (2014) suggest presence of continuous gradients of variation in bacterial communities rather than the discrete enterotypes [31,32].
Stable and diverse intestinal microbiota with the host organism together represent a complex symbiotic connection when intestinal microbes positively influence the physiology, immunology and metabolism of the host [33,34]. On the other hand, imbalance in the composition and therefore function of the intestinal microbiome, defined as intestinal dysbiosis, may contribute to the development or progression of obesity and comorbidities associated with it [35,36].
3. Methods of the Gut Microbiota Identification
The number of studies analyzing microbial communities has increased rapidly due to technological advances in sequencing and metagenomics, overcoming the fact that more than 90 % of the gut bacteria cannot be detected by conventional culture methods [37]. However, the relative abundance of bacterial groups does not always correspond to the absolute abundance of bacterial taxa observed. 16S rRNA gene sequencing and whole genome sequencing (WGS) are currently two main approaches used in the species and functional analysis of the gut microbiome.
The sequence of the 16S rRNA gene encoding the genetic information of the small ribosomal subunit is highly conserved throughout the prokaryotic kingdom. Though variability of these selected segments is low on the species level and is therefore sufficient for the species identification and the phylogenetic categorization of the taxa [38,39], the accuracy of the method is assured only up to the phyla or genera level. The taxonomic classification of the data obtained is based on the comparison of 16S rRNA amplicon sequences with the reference libraries of the currently most frequently used platforms such as Illumina [40], Roche 454 or Ion Torrent [41].
The WGS is recommended (though it is relatively costly) for analysis of the metagenome in a specific biological setting; it offers high resolution down to the species or strain level. The WGS methods (Sanger sequencing, shotgun sequencing, next-generation sequencing, nanopore sequencing) provide information on the functional properties of microorganisms, including the presence of additional genes related to the study's problem [42,43,44,45].
Suitable bioinformatics tools for subsequent raw data processing were summarized by Combrink et al. [46] and Beghini et al. [47] compared the use of HUMAnN 3 and MetaPhlAn 3 software for taxonomic profiling and description of the metabolic potential of a given microbial community. Recently, meta-transcriptomic tools have also been used to assess gene expression in microorganisms on the regulation level [48]. Karcher et al. [49] or Tett et al. [50] applied the software MetaPhlAn 2 to detect evolutionarily related strains between different host communities.
Adequate experimental design, including proper selection of input parameters, such as age, gender, disease prevalence and lifestyle, is essential to ensure maximum precision of the results in studies dealing with the analysis of the gut microbiome, especially as far as longitudinal studies are concerned [51]. Moreover, correctness of all factors of the analytical process, such as storage method [40], choice of primers, identical reagent kits throughout the follow-up [46] and suitable controls [40,52,53] should also be ensured.
4. Products of the Gut Microbiome Metabolism
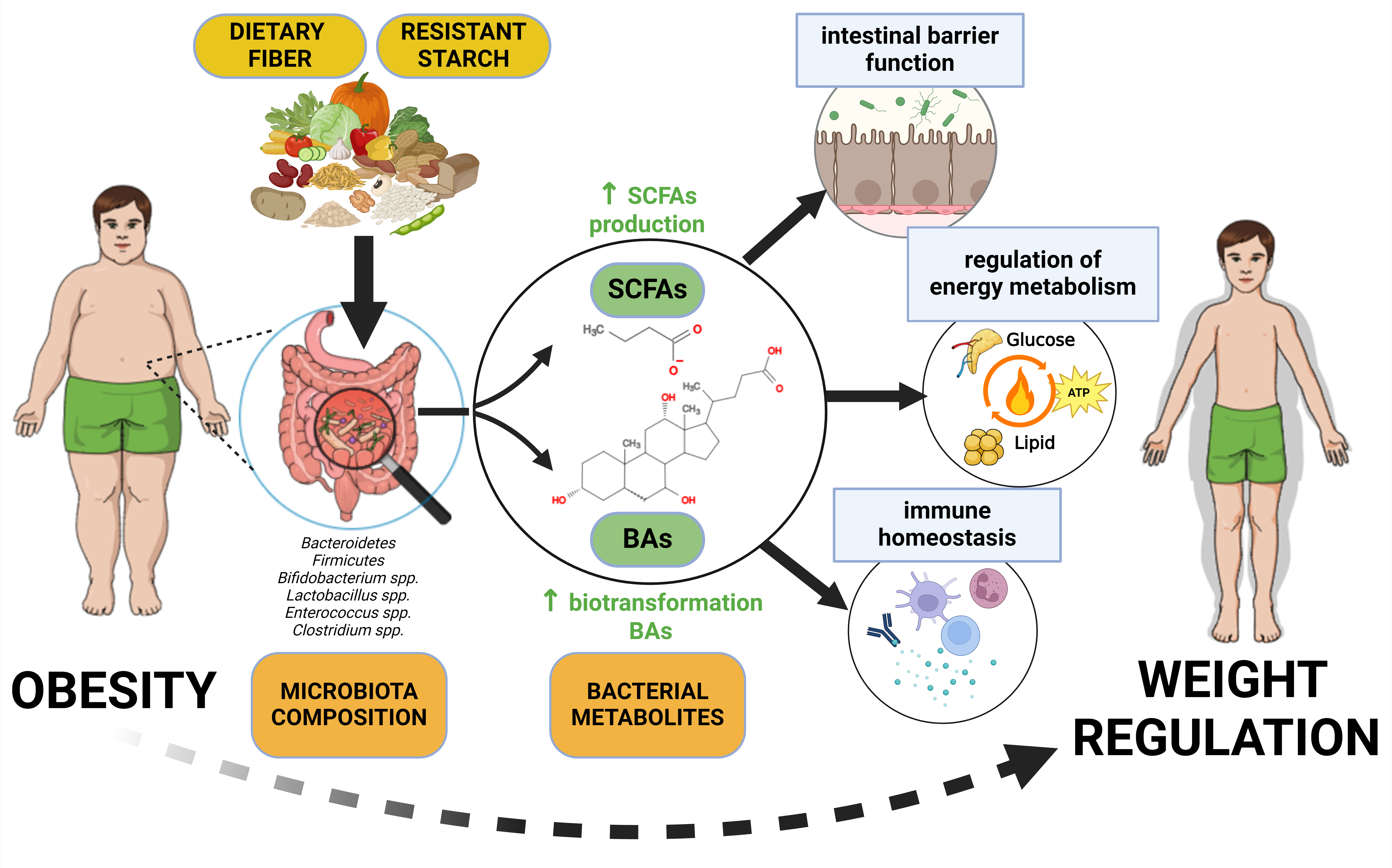
According to current knowledge, it is estimated that the intestinal microbiota includes up to 100 trillion microbial cells and contains more than 22 million microbial genes encoding many enzymes with diverse metabolic activity [54]. Intestinal microbiota can produce a large number of bacterial metabolites with a wide spectrum of biological activities for the host organism. These metabolites can be generally divided into three types: 1) metabolites produced by bacterial degradation of nutrients and other food components from a diet, such as SCFAs; 2) metabolites created by the host organism and modified by microbiota, such as secondary BAs; 3) metabolites produced de novo by the microbiota such as lipopolysaccharides (LPSs; [55]). The present article is focused especially on SCFAs and BAs, including secondary BAs, which are studied in relation to the regulation of both human obesity and intestinal microbiome [56,57].
4.1. Short-Chain Fatty Acids
4.1.1. Butyrate, Propionate and Acetate Functions
SCFAs are organic acids with the chain length < six carbons [13]. They are predominantly produced by anaerobic bacterial fermentation of indigestible food components, such as dietary fiber (DF) and resistant starch (RS; [58]), which are also collectively referred to as microbiota-accessible carbohydrates (MACs; [59,60]).
SCFAs most represented in human colon are acetate (60 % of total SCFAs), propionate (20 %) and butyrate (20 %; [61]). Lactate isomers, valerate, iso-butyrate and iso-valerate also belong to the human gut metabolome, but are present here only in low concentrations [62]. Functions of SCFAs are mediated by free fatty acid receptors (FFAR2 and FFAR3), G protein-coupled transmembrane receptors (GPCRs) located (among other things) on colonocytes [63], and by colonic epithelium SCFA proton-coupled, sodium-coupled and ATP-dependent transporters [64]. Majority of SCFAs is absorbed in the colon by passive diffusion or by active transport, and only a small fraction (about 5 %) is excreted in the feces [12].
One of the SCFAs functions in the gut is to strengthen integrity and modulate permeability of the gut barrier [13]. Butyrate upregulates expression of the genes coding for claudin-1 (CLDN-1), zonula occludens-1 (ZO-1) and occludin (OCLN), proteins composing tight junctions [65]. Butyrate also increases expression of mucin 2, strengthening the mucus layer of the gut epithelium [66].
SCFAs in general, and butyrate in particular regulate intestinal cell proliferation. The mechanism is based mainly on inhibition of histone deacetylase (HDAC) by butyrate. Histone hyperacetylation enables a steady expression of the gene coding for p21Cip1 protein (cyclin-dependent kinase inhibitor 1). Higher concentration of p21Cip1 protein increases probability of its binding to D1 and D3 cyclins. Cyclin inactivation leads to blocking of the cell cycle and cell arrest [67].
Butyrate is also a primary energy source (70 %) for colonocytes [68]. Acetate stimulates secretion of glucagon-like peptide 1 (GLP-1) and peptide YY (PYY) and so participates on regulation of appetite [69] and reduction of body weight [70]. Due to the suppression of hepatic gluconeogenesis (by propionate; [71]) and lipogenesis (by acetate and butyrate; [72]) modulate SCFAs both glucose and lipid metabolism. SCFAs are also able to activate G protein-coupled receptor GPR41 and so stimulate production of leptin [73].
SCFAs also lower the pH in the colon, which can inhibit the growth of pathogenic bacteria and conversely promote the growth of beneficial bacterial genera, such as Bifidobacterium and Lactobacillus [74].
Impact of SCFAs on innate and adaptive immunity is based, among other things, on decreased production of reactive oxygen species (ROS) in neutrophils [75] and on suppressed expression of monocyte chemoattractant protein 1 (MCP-1), vascular cell adhesion molecule 1 (VCAM-1) and chemokine signaling [76,77]. Moreover, butyrate is able to suppress signaling pathway of nuclear factor kappa B (NF-κB), reducing thereby gut inflammation [78].
Butyrate signaling is based on three G-protein coupled receptors (GPCRs: GPR43, GPR41, GPR109A) expressed, among other things, on colonocytes; the signaling inhibits cAMP-dependent pathways and therefore promotes anti-inflammatory response in colon [79]. Butyrate also activates aryl hydrocarbon receptor (AhR), peroxisome proliferator-activated receptor gamma (PPARγ) and hypoxia-inducible factor 1 (HIF-1). As a consequence, hypoxic conditions in the gut lumen are induced, which supports gut homeostasis, including suppression of the growth of facultative anaerobes [11].
4.1.2. Production of Short-Chain Fatty Acids by the Gut Microbiota
Formation of SCFAs and their effects on the host organism are summarized in Figure 1. Abundances of particular intestinal SCFAs-producing bacteria depend on the stages of human life [13]. Original dominance by Enterobacteriaceae is being gradually replaced by Bifidobacteriaceae in early life [80]. Counts of Firmicutes are growing in the later stages [81] and abundances of Enterobacteriaceae increase again in older age [82].
Species within Firmicutes families Lactobacillaceae, Ruminococcaceae and Lachnospiraceae produce SCFAs by breaking down complex polysaccharides [83]. Bifidobacteria species such as B. bifidum, B. infantis and B. breve dominate the infant gut microbiome and utilize human milk oligosaccharides producing predominantly acetate [84]. Firmicutes, especially Clostridia class start to increase when the more diverse food is available [85] increasing gut concentration of propionate via the succinate pathway (converting succinyl-CoA to propionate), acrylate pathway (converting lactate to propionate) and propanediol pathway (processing of deoxy sugars; [86]).
As far as butyrate is concerned, its synthesis proceeds from the condensation of two molecules of acetyl-CoA to acetoacetyl-CoA to reduction to butyryl-CoA and finally to transformation to butyrate via either butyryl-CoA:acetate-CoA-transferase or phosphotransbutyrylase/butyrate kinase [87]. Though some species belonging to the Actinobacteria, Fusobacteria and Proteobacteria phyla are also able to generate butyrate [88], most butyrate-producing bacteria belong to the phylum Firmicutes [13]. Species of the families Lachnospiraceae and Ruminococcaceae are important butyrate producers [89,90], but one of the most abundant butyrate-producing bacteria, representing up to 5 % of the colon microbiota of healthy adults, is Faecalibacterium prauznitzii, member of the Ruminococcaceae family [91,92].
For maintaining a healthy intestinal environment, the butyrate-producing bacterial communities are instrumental [13]. Production of butyrate as the substrate for generation of energy for colonocytes enables these epithelial cells to increase oxygen consumption and maintain an anaerobic environment in the gut [93]. As a consequence, growth of the opportunistic aerobic pathogens such as Salmonella and Escherichia coli can be suppressed [94].
SCFAs produced by the gut microbiota are rapidly absorbed by the colonic epithelium and can be used by the epithelial cells themselves (butyrate is a primary energy source for the colonocytes) or within the host metabolism. Acetate is engaged in cholesterol synthesis and, as a precursor for biosynthesis of long-chain fatty acids, in lipid metabolism [19]. Propionate is, among other things, a substrate for gluconeogenesis in the liver and thus contributes to glucose homeostasis. As a part of the gut-brain-axis SCFAs (especially propionate and acetate) modulates production of the host appetite-regulating hormones and consequently satiety and energy intake [95]. Butyrate, except for the above-mentioned energy source for gut epithelial cells, participates in these cells in regulation of the gene expression as an inhibitor of HDAC with a possible consequence of stimulation of apoptosis and therefore decreasing risk of colorectal cancer [96].
4.1.3. SCFAs and Human Obesity
Most experiments and meta-analyses report increased fecal SCFAs in obese individuals compared to the healthy lean individuals [97,98]. The main mechanisms of increased concentration of SCFAs in stool of obese people may be: 1) increased bacterial production and shifts in cross-feeding between bacterial producers of SCFAs; 2) disorders of mucosal absorption; or 3) speed of consumed food transit [99]. On the other hand, serum and plasma levels of propionate and butyrate may be reduced in individuals with a higher BMI [100,101]. However, current knowledge of changes in blood SCFAs concentrations during the development and progression of obesity is incomplete, because most studies deal with the measurement of fecal SCFAs concentration in the stool, where are SCFAs more detectable [98].
4.2. Bile Acids
Sensu stricto, only secondary bile acids (secondary BAs) are products of the gut microbiota metabolism; primary bile acids (primary BAs) are synthetized in the host liver and are mentioned in the following text only as precursors of secondary bile acids.
4.2.1. Structure, Synthesis and Metabolism of Primary Bile Acids
In general, the molecule of a human bile acid (BA) includes 24 carbons forming a four-ring steroid nucleus (three six-membered rings A, B and C, and one five-membered ring D), an aliphatic side chain and two methyl groups attached at the C-18 and C-19 positions of the main carbon skeleton ([16]; Figure 2). Human primary BAs comprise cholic acid (CA; 3α,7α,12α-trihydroxy-5β-cholan-24-oic acid) and chenodeoxycholic acid (CDCA; 3α-7α-dihydroxy-5β-cholan-24-oic acid), together with their respective amino acid conjugates: the lipophilic CA and CDCA are conjugated in the liver with taurine and glycine to the more hydrophobic taurocholic acid (TCA) and taurochenodeoxycholic acid (TCDCA), and glycocholic acid (GCA) and glycochenodeoxycholic acid (GCDCA), respectively. Human secondary BAs consist of deoxycholic acid (DCA; 3α,12α-dihydroxy-5β-cholan-24-oic acid) and lithocholic acid (LCA; 3α-hydroxy-5β-cholan-24-oic acid; Figure 2). Amphipathic nature of BAs is given by the hydrophilic hydroxyl and carboxyl groups and hydrophobic methyl groups, respectively, possessing surface activity with an ability to form micelles [102].
The rate-controlling enzyme in the “classical” pathway of primary BAs (CA, CDCA) synthesis is cytochrome P450 cholesterol 7α-hydroxylase (CYP7A1); sterol 12α-hydroxylase (CYP8B1) is further required for production of CA. CDCA alone is produced via the “alternative” pathway using sterol 27α-hydroxylase (CYP27A1) and 25-hydroxycholesterol 7α-hydroxylase (CYP7B1; [14]). Amino acid conjugation of BAs proceeds in the liver by means of bile acid CoA:amino acid N-acyltransferase [103].
Quinn et al. (2020) recently reported an ability of the gut microbiota to conjugate CA with amino acids independently of the host liver synthesis. However, according to Guzior and Quinn (2021), the reason for bacterial BAs conjugation with hydrophobic amino acids is currently unclear, because the conjugation affects the detergent, signaling and antimicrobial properties of BAs [103,104].
After food intake, enteroendocrine cells of the duodenum produce hormone cholecystokinin that stimulates secretion of conjugated primary BAs from the gallbladder into the small intestine [105], where these BAs facilitate the digestion and absorption of dietary lipids, cholesterol, lipophilic vitamins and other hydrophobic food components due to their surface-active properties [106].
Conjugated primary BAs which are secreted to the duodenum, proceed through the small intestine, are reabsorbed in the terminal ileum by the apical sodium-dependent BA transporter (ASBT) and subsequently transported back to the liver within the enterohepatic circulation [107]. Small amount of the primary BAs proceeds to the colon, where these BAs are converted to the more hydrophobic secondary BAs (DCA and LCA). The secondary BAs are reabsorbed from the colon passively. Altogether 95 % of BAs is reabsorbed from the ileum and colon through the portal vein [108].
BAs are ligands of the farnesoid X receptor (FXR), CDCA being the most potent FXR agonist. Moreover, GPCRs such as Takeda G protein-coupled receptor 5 (TGR5) and sphingosine-1-phosphate receptor 2 (S1PR2) can also be activated by BAs. It is interesting that activation of these two last-mentioned receptors has opposite effects on the liver: activation of S1PR2 can lead to a pro-inflammatory mode, activation of TGR5 results in anti-inflammatory (and anti-cholestatic and anti-fibrotic) effects [103].
FXR is a major regulator of the BAs' synthesis, transport, secretion and absorption [109]. After BAs ligation, FXR dimerizes in enterocytes with the retinoid X receptor (RXR), activating transcription of the genes coding for proteins performing transcellular transport of BAs and for fibroblast growth factor 19 (FGF19) that enters the portal circulation. The FGF19 signaling stimulates in the liver phosphorylation of extracellular signal-regulated kinases (ERKs), which inhibits expression of the CYP7A1 gene coding for cholesterol-7α-hydroxylase, thereby reducing BAs synthesis [110]. The data of Friedman et al. (2018) indicate that the FXR activation modulates the gut microbiota composition via the BAs-dependent mechanism, suppressing BAs synthesis in the liver and stimulating the proliferation of the Gram-positive species, such as Streptococcus thermophilus, Lactobacillus casei, Lactobacillus paracasei, Bifidobacterium breve or Lactococcus lactis [111].
Conjugated BAs are also able to bind to dietary fibre (DF). Diet rich in whole grains significantly increased concentrations of primary conjugated BAs taurocholic, glycocholic and taurolithocholic acids with a possible resultant activation of FXR and TGR5, affecting glucose homeostasis [112].
4.2.2. Production of Secondary Bile Acids
During their intestinal transit, bound primary (conjugated) BAs undergo further structural modifications and transformations to secondary BAs by the gut microbiota, after previous deconjugation and subsequent 7α-dehydroxylation [113,114].
Deconjugation reactions are catalyzed by the microbial bile salt hydrolases (BSHs) encoded by the bsh genes that are present in all major phyla of the gut bacteria (Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria), including the gut archaea [115]. Bacterial genera capable to cleave off the amino acid (taurine, glycine) moiety comprise Gram-positives Bifidobacterium, Lactobacillus, Clostridium, Enterococcus and Listeria, but also Gram-negatives, such as Stenotrophomonas, Bacteroides and Brucella [103]. Deconjugation is a necessary first step for further modifications [115].
Deconjugated primary BAs, as signaling molecules, are able to modify the total BAs pool. Because deconjugation increases amounts of antimicrobially effective BAs (CA, CDCA), the gut microbiota is supposed to evolve the deconjugation mechanism as a means of competition; deconjugation is therefore considered an essential function of the gut bacteria [103]. Due to the fact that all major phyla of the gut bacteria are endowed with enzymes able to catalyze deconjugation reactions, horizontal transfer of the genes coding for these enzymes is supposed [116].
The three major groups of bacterial enzymes converting primary BAs into secondary BAs in the gut comprise the above-mentioned BSHs and further hydroxysteroid dehydrogenases (HSDHs) and the enzymes responsible for dehydroxylation of unconjugated BAs in the colon, located in the BA-inducible bai operon [14]. The BSHs enzymes deconjugate primary BAs by splitting the bond between the BA and glycine or taurine moieties in the C-24 position at the side chain of the BA molecules [117].
Deconjugated primary BAs undergo 7α-dehydroxylation just by several bacterial species such as Clostridium scindens, C. hylemonae and C. hiranonis (now classified as Peptacetobacter hiranonis; [118]). Although many bacteria are able to perform dehydroxylation reactions, only enzymes inducing 7α-dehydroxylation encoded by the genes located in the bai operon of the above-mentioned bacteria lead to the formation of secondary BAs [119], specifically deoxycholic acid (DCA) from CA and lithocholic acid (LCA) from CDCA, respectively, which are the most abundant secondary BAs in humans [15].
Secondary BAs, but also primary BAs, can reversibly undergo oxidation and epimerization reactions at position C-3, C-7 or C-12, from α-orientation through an oxo-intermediate to β-orientation by microbial α/β-HSDHs. Epimerized BAs have a specific nomenclature: BAs containing 3β-hydroxyl groups are called iso-BAs, while 7β- and 12β-BAs with hydroxyl groups are called epi-BAs [118,120]. For example, CA can be epimerized to ursocholic acid (UCA), isocholic acid (iCA) or 12-epicholic acid (12-ECA) while CDCA can epimerize to derivatives such as ursodeoxycholic acid (UDCA) or isochenodeoxycholic acid (iCDCA; [103]). Many intestinal bacteria including the genera Bacteroides, Clostridium, Eubacterium, Ruminococcus, Peptostreptococcus, Escherichia and Eggerthella contain HSDHs [118].
The circulating pool of modified BAs can be further diversified by other biotransformation reactions, such as sulfation, esterification and even reconjugation by the gut microbiota [15]. Further research is necessary to clarify physiological effect of these BA derivatives, which may be important in intracellular signaling and regulation of homeostasis in the human organism [104]. Bacterial taxa mediating bile acid biotransformation reactions are summarized in Table 1; overview of the synthesis, reabsorption, biotransformation and physiological effects of BAs in the human body is shown in Figure 3.
4.2.3. Effects of Bile Acids on the Gut Microbiota, Intestinal Barrier Function and Immune Homeostasis
From the physicochemical viewpoint are BAs detergents, acting on the membrane of enterocytes and enabling thereby absorption of fatty acids in the small intestine. From the same reason are BAs also able to disrupt bacterial membranes. However, the effect differs between conjugated and non-conjugated BAs. Primary BAs conjugated with glycine or taurine are fully ionized at physiological pH, which prevents them to significantly interact with bacterial membranes. On the other hand, non-conjugated BAs (CA, CDCA) are able to disrupt bacterial membranes and damage the bacterial cell [124]. Sannasiddappa et al. (2017) or Kurdi et al. (2006) reported that non-conjugated BAs decreased viability of the tested bacterial species (Staphylococcus aureus, Lactobacillus spp., Bifidobacterium spp.) more than their conjugated counterparts [125,126]. From this viewpoint, deconjugation can be seen as a means of competition between gut bacteria and is it an essential function of the gut microbiota [103].
As an example, Bifidobacterium bifidum is able to inhibit a toxicogenic activity of Vibrio cholerae by metabolizing primary BAs into secondary BAs (CA into DCA; [127]). Conversion of primary BAs to secondary BAs by commensal bacteria provides colonization resistance against Clostridioides difficile: primary BAs induce spore germination, whilst secondary BAs are toxic for vegetative cells of C. difficile [128]. Moreover, secondary BAs (LCA and DCA) can exert a direct antifungal activity against Candida albicans [129].
BAs composition and pool size exert antimicrobial effect both directly, by their detergent properties, and indirectly, via stimulating production of antimicrobial peptides (AMPs) and modulating the host immunity [130,131]. BAs also modulate the microbiome composition: CA supports, but DCA inhibits abundances of the species of the Firmicutes phylum, DCA on the other hand stimulates the proportion of Bacteroidetes.

Table 1.
Overview of bacterial taxa mediating bile acid biotransformation reactions.
| Biotransformation reactions | Phylum | Genera | Reference |
|---|---|---|---|
| deconjugation | Bacteroidetes | Bacteroides Parabacteroides Barnesiella Alistipes |
[45,121] |
| Firmicutes | Clostridium Eubacterium Ruminococcus Lachnospira Roseburia Lactobacillus Enterococcus |
[14,15,121,125] | |
| Actinobacteria | Bifidobacterium Eggerthella |
[15,121] | |
| 7-α dehydroxylation | Firmicutes | Clostridium spp. (C. scindens) (C. hylemonae) Peptacetobacter hiranonis |
[14,118,122] |
| oxidation / epimerization | Firmicutes | Clostridium Eubacterium Collinsella |
[14] |
| Bacteroidetes | Bacteroides | ||
| sulfation | Firmicutes | Clostridium Fusobacterium Peptococcus |
[15] |
| Proteobacteria | Pseudomonas | ||
| esterification | Firmicutes | Eubacterium Lactobacillus |
[15,123] |
| Bacteroidetes | Bacteroides |
BAs concurrently modulate intestinal barrier function and immune homeostasis [15]. Primary BAs improve intestinal barrier function by inducing expression of ZO-1 and concurrently decreasing expression of tumor necrosis factor alpha (TNF-α) and interleukin 6 (IL-6; [132]). As FXR agonists, primary BAs stimulate expression of the tight junction proteins [133], contrary to secondary DCA that impairs BAs deconjugation, thus inhibiting FXR activation with a consequence of impairment of the mucosal barrier function [134]. On the other hand, DCA stimulates expression and secretion of human β-defensins 1 and 2, improving maintenance of intestinal homeostasis [135]. Moreover, BAs promote epithelial regeneration via TGR5 signaling [136].
As far as BAs effects on intestinal innate and adaptive immunity are concerned, BAs stimulation of TGR5 promotes differentiation of monocytes into tolerogenic dendritic cells [137] and activation of FXR inhibits signaling pathway of NF-κB [138]. Moreover, BAs promote polarization of macrophages toward the anti-inflammatory phenotype M2 [139] and activate TGR5-cAMP (cyclic adenosine monophosphate)-PKA (protein kinase A) signaling pathway, thus limiting IL-1β and IL-18 and stimulating IL-10 production [140].
4.2.5. BAs Analysis
Methods of BAs analysis recently summarized Singh et al. (2019; [16]). 1H and 13C nuclear magnetic resonance spectroscopy (NMR) was useful for elucidating the diverse structures of conjugated BAs (GCA, GCDCA, TCA, TCDCA; [153]). Gas chromatography/mass spectroscopy (GC/MS) with electrospray ionisation (ESI) provided spectral data on the forms of conjugation and BAs structure [154]. Singh et al. (2019) suggested higher sensitivity and selectivity in BAs analysis of more recent methods, such as high-resolution mass spectroscopy (HR-MS), collision-induced decomposition (CID), hybrid quadrupole time-of-flight (TOF) multiple reaction monitoring (MRM) or selected reaction monitoring (SRM; [16]). BAs analyses, reviewed for biological fluids and tissues by Griffiths and Sjövall (2010), comprise BAs extraction, group separation (in the case of complex BAs mixtures or a presence of many types of conjugates), cleavage of conjugates, derivatization of polar groups, and determination by GC/MS, liquid chromatography/mass spectroscopy (LC/MS), or enzymatic assays and immunoassays [155].
5. Host Diet and Gut Microbiota
Diet is an important factor influencing the intestinal microbiota, which can have a potential therapeutic effect for adjusting of composition, diversity and stability of the intestinal microbiome [156]. Short-term diets and specific dietary interventions have been shown to rapidly alter gut diversity. However, these changes are often transient and do not persist after the diet and the intervention have ended [157]. Long-term eating habits and a typical diet across populations from different geographical areas correspond with the microbial composition and are thus associated with the stability of a given enterotype of an individual or group of individuals [158,159].
Diets rich in protein and fat on the one hand and carbohydrates on the other hand are often associated with Bacteroides-abundant (enterotype I) and Prevotella-driven (enterotype II) microbiota profile, respectively [10]. It follows from the data of Wu et al. (2011) that Bacteroidetes and Actinobacteria positively correlated with dietary fat, but negatively with dietary fibre (DF); the opposite behavior was reported for Firmicutes and Proteobacteria [158]. DF increased gut abundance of Prevotella, while bile resistant genera like Bilophila and Bacteroides correlated with a high-fat animal-based diet [157].
The abundances of dominant species of colonic microbiota can be modified rapidly and at the same time reversibly by dietary intervention, which demonstrated Walker et al. (2011) in a cohort of overweight men on a diet high in resistant starch (RS). Despite predominant inter-individual differences, the RS-diet rapidly changed abundances of the specific bacterial groups, with the most conspicuous increase in Ruminococcus bromii and Eubacterium rectale; however, the changes were rapidly reversed by the subsequent diet [160].
De Filippo et al. (2010), when comparing diets of children from rural Africa and Europe, respectively, found out that fibre-rich diets stimulate genera within the Bacteroidetes phyla (specifically Prevotella and Xylanibacter), but animal-based and sugar-rich diets increase amounts of Firmicutes and Enterobacteriaceae [161].
Vegetarians generally tend to have higher bacterial diversity and higher ratios of Prevotella to Bacteroides and reduced numbers of Enterobacteriaceae including Escherichia coli compared to omnivores [162]. Compared to omnivores, vegetarians and vegans have also higher abundance of Lachnospiraceae (genera Roseburia, Anaerostipes, Blautia) and Ruminococcaceae (genera Ruminococcus and Faecalibacterium prauznitzii), and lower abundance of Bacteroides, Parabacteroides and Alistipes [163].
Intake of vegetable proteins based on whey or peas can increase abundance of the genera Bifidobacterium and Lactobacillus [164,165]. On the other hand, consumption of animal proteins leads to increased abundances of Bacteroides, Alistipes and Bilophila, but also of some species of the genera Clostridium [164]. Shoonakker et al. (2024) reported a decrease in the family Lactobacillaceae and Enterobacteriaceae after a dietary protein restriction [166]. Generally speaking, bacteria with proteolytic and amino acid degrading activity comprise genera Clostridium, Fusobacterium, Peptostreptococcus, Veillonella and Bacteroides [167]. It should be mentioned however, that controlled human studies examining exclusively the effect of different dietary protein sources are limited and most of the evidence comes from model animal studies [168].
As far as dietary lipids are concerned, results of intervention studies show reduced bacterial diversity owing to the high-fat diet (HFT) and reduced total numbers of resident bacteria, including Firmicutes [165]. On the other hand, HFT typically increases genera such as Alistipes and Bacteroides, while dietary fat restriction leads to increased numbers of beneficial Faecalibacterium [166,169].
The Western-type diet, characterized by a high protein and fat (especially saturated fat) intake and associated with higher incidence of metabolic disorders (T2D), cardiovascular diseases and obesity [170,171], increases abundances of Bacteroides, Alistipes and Bilophila and decreases genera Lactobacillus, Roseburia, Eubacterium and Enterococcus [172,173]. In contrast, the Mediterranean diet, based on dietary fibre (cereals, legumes, vegetables, fruits and nuts), unsaturated fatty acids (fish, vegetable oils) and antioxidants (flavonoids, polyphenols; [174,175,176]), increases overall microbial diversity, including families of Clostridiaceae and Lactobacillaceae and specifically genera such as Bacteroides, Bifidobacterium, Prevotella, Roseburia, Clostridium, Lactobacillus and Faecalibacterium) and decreases abundances of Proteobacteria [175,177,178,179].
Boosting fibre consumption to 40 g/day in healthy adults increased faecal abundance of a group of Lachnospiraceae (ND3007), SCFA-producing family of bacteria with a concomitant tendency to increase the butyrate-producing capability [180].
5.1. Dietary Fiber
Dietary fiber (DF) is possible to sub-divide into insoluble dietary fibre, including prebiotics, and soluble dietary fibre [17]. To the main sources of insoluble dietary fiber (cellulose, hemicelluloses and fructans; [181]) belong whole wheat flour, brown rice, nuts, beans and vegetables [182]. Prebiotics, a special class of insoluble fibre, are defined as “non-digestible compounds that, when consumed, induce changes in composition and/or activity of the gastrointestinal bacteria, thus causing benefit(s) upon host health” [183].
Prebiotics are classified based on their ability to withstand passage through the gastrointestinal tract (resistance to gastric acidity and to the host hydrolytic enzymes, gastrointestinal absorption), on their suitability for fermentation by intestinal microbiota and on their ability to stimulate the growth and activity of intestinal bacteria with positive effects on the host health [184]. Selective fermentation, modulation of intestinal pH, bulk of stool, growth inhibition of pathogenic bacteria and host protection against toxic metabolites of the putrefactive bacteria are usually listed as the principal attributes of prebiotics. Fructooligosaccharides, galactooligosaccharides, xylooligosaccharides, arabinooligosaccharides, oligofructose, inulin, β-glucan, guar gum, resistant starch and maltodextrin are considered most important types of prebiotics [17].
Soluble fiber, contrary to insoluble fiber, are viscous gel-like substances able to slow absorption of nutrients in the intestine [185]. Pectin, guar gum and some types of inulin, present in whole grains, legumes, seeds and nuts, and some fruits and vegetables, are the most widely used representatives of soluble fibre [186].
The positive effects of DF on the gut microbiome and on the gut epithelium comprise production of SCFAs [182], including SCFAs-mediated modulation of the epithelial barrier functions [187] and the host innate immune defenses [188]. Therefore, one of possible dietary interventions in obesity is to support in the gut the carbohydrate-associated enterotype [10] by securing sufficient intake of DF, including also resistant starch [6].
From this viewpoint is it necessary to view the gut microbiota as a hugely diverse ecosystem [18]. As far as diversity is concerned, alpha diversity (within-community diversity) and beta diversity (measure of similarity between communities) are usually used when diversity indicators are calculated [189]. Richness (number of taxonomic groups), evenness (taxa abundances distribution) and phylogenetic diversity (phylogenetic distance regarding the branch lengths in the phylogenetic tree) are indispensable tools as far as alpha diversity metrics is concerned [190].
Intake of DF is able to change rapidly composition of the human gut microbiome [191]. However, dietary interventions using fermentable DF aiming to promote gut alpha diversity are often ineffective and their results are contradictory as follows from a meta-analysis of pertinent clinical trials [192]. Moreover, Zhang et al. [193] demonstrated alleviation of children obesity with concomitantly reduced gut alpha diversity.
As far as the above-mentioned richness is concerned, it can be increased by a long-term intake of DF [194]. Wang et al. [180]even reported higher fecal alpha diversity in a short-term (2 weeks) two-phase randomized cross-over trial swapping white bread for high-fibre bread (fibre intake 40 g/day). However, DF intake is mostly associated with a reduction in richness of the gut microbiota [195,196]. The likely reason relates to the substrate-specific abilities of the gut microbiota to utilize the available substrates [197]. Therefore, limited overall substrate availability or changing the type of substrate can cause loss of gut bacteria and reduce richness [198].
Changes in the parameter of evenness (equitability of the proportions of the bacteria composing a given community) are dependent on the abundances of bacteria sharing the fibre substrate and on the initial composition of the gut microbiome [199]. Therefore, decreased evenness within the highly competitive gut environment after supplementation of different types of DF [198], can be explained by a greater chance of the initially highly abundant bacteria (provided they are the substrate utilizers) as compared to the low abundant bacteria to utilize the supplemented substrate [199]. Consequently, the last-mentioned authors Cantu-Jungles and Hamaker [199] conclude that despite still achieved health-related improvement after the fibre supplementation, reduction in alfa diversity is possible, even expected.
In order to DF provide health-beneficial metabolites in the gut, microbial communities must be present that are able to express specific genes coding for enzymes that degrade and metabolize complex carbohydrates [200]. These so-called carbohydrate-active enzymes (CAZymes; [201]) can be divided into several classes, such as glycoside hydrolases (GHs), glycosyltransferases (GTs), polysaccharide lyases (PLs), carbohydrate esterases (CEs) and carbohydrate-binding modules [202]. As an example of a relatively versatile fibre-utilizing genera, Bifidobacterium has at its disposal CAZymes for metabolizing arabinoxylan-derived oligosaccharides and inulin-type fructans [203,204]. On the other hand, Monoglobus pectinilyticus is a highly specialized pectin degrader whose genes code for special CEs and PLs [205]. Lacticaseibacillus paracasei was able to express an extracellular exo-inulinase in the presence of a β-(2,1)-fructan, which increased availability of fructooligosaccharides for other bacteria [206].
Supplementation by dietary fibre rich in arabinans and galacturonans increased pool of genes encoding for arabinofuranosidases, β-glucanases, β-xylanases and polysaccharide lyases [207]. Rivière et al. (2016) demonstrated importance of the cross-feeding reactions between Bifidobacterium and the butyrate producing genera Roseburia and Anaerostipes for the production of the host-beneficial metabolites [208].
Kok et al. [18] conclude that fibre utilization in the human gut can be successfully realized either as a particular bacteria-substrate interaction or via cross-feeding interactions with participation of several taxa. However, for a successful fibre utilization are necessary not only the above-mentioned CAZymes, but also regulatory elements, transporters and binding proteins comprising carbohydrate gene clusters [201]. Following the discovery of the starch utilization system in Bacteroides thetaiotaomicron, the glycan utilization system called polysaccharide utilization loci (PULs) was characterized in genera Bacteroides in general [209]. On the other hand, species within Firmicutes and Actinobacteria degrade glycans using various transport systems, and the presence of PULs encoding for polysaccharide-degrading enzymes, oligosaccharide transporters and transcriptional regulators were reported in strains of the Firmicutes phylum [210].
According to Kok et al. [18], a response of the gut microbiome to DF supplementation is highly personalized. Rodriguez et al. [211] distinguished after 3 months of inulin supplementation in a cohort of obese patients a sub-group of responders (BMI decrease), showing higher abundances of Akkermansia and Butyricicoccus and a lower abundance of Anaerostipes. Therefore, though Kok et al. [18] admit a possibility for a modulation of the gut microbiome by dietary interventions, the authors concurrently recommend to employ microbiome-derived biomarkers for the subjects' classification [18]. Chen et al. [23] proposed in this context an employment of the above-mentioned term of enterotypes namely Prevotella-, Bacteroides- and Ruminococcus-dominant enterotype [212]. Treated with the high-fibre diet, individuals with Prevotella enterotype reduced weight more efficiently than individuals with Bacteroides enterotype [213]. Similarly, better fibre-utilizing ability, including higher SCFAs concentration was reported with Prevotella enterotypes as compared to Bacteroides enterotypes [212].
One of suitable biomarkers regarding fibre metabolism is Bifidobacterium spp., an example of a single taxonomic enrichment after supplementation of the prebiotic fibers [214]. Kok et al. [18] summarized results of the studies differentiating responders and non-responders by observing enrichment of Bifidobacterium spp. after providing different types of DF, including galactooligosaccharides or agave inulin. Responders, as compared to non-responders, were often individuals with less stable gut microbiome [18,215]. Higher abundances of Bacteroidetes and Firmicutes at baseline were reported in responders and non-respondents, respectively [18]. Magne et al. [216] suggested a ratio of Firmicutes/Bacteroidetes as a suitable marker for evaluating gut dysbiosis in obese patients.
However, effects of the fibre-rich dietary interventions based exclusively on the enterotype-dominance should be considered with caution in general population due to confounding factors such as a baseline diet, exercise or use of antibiotics [217]. Probands classified as high-dietary fibre consumers at a baseline, in comparison with low-dietary fibre consumers, showed increased abundances of Bifidobacterium and Faecalibacterium and decreased amounts of Coprococcus, Dorea and Ruminococcus after dietary inulin intervention, demonstrating a broader inulin-conditioned taxonomic response [218].
5.2. Resistant Starch
Starches in general are macromolecules based on glucose, stored in many plants as a primary source of energy [219]. However, from the viewpoint of providing energy to the human organism, a substantial difference exists between various types of starches. Most types of starch are easily hydrolyzed in the small intestine by the human saccharolytic enzymes (α-amylase, maltase and isomaltase; [220]). On the other hand, resistant starch (RS) escape hydrolysis in the small intestine and proceed undigested into the large intestine, where they are broken down by enzymes produced by the gut microbiota [221].
The glucose units in starches are arranged in linear chains of amylose (α-1,4-glycosidic linkage) and a branched-chained amylopectin (α-1,6-glycosidic bonds; [222]). The digestibility/resistance of the starches is primarily determined by the amylose/amylopectin ratio and their organization within starch granules. The more densely and more tightly packed are these components in the starch granules, the less accessible they are to the human digestive enzymes [223].
Grains (whole grains of barley, oats and wheats), legumes (lentils, chickpeas, beans), tubers (potatoes and yams; their cooking and subsequent cooling induces starch retrogradation) and some kinds of the processed foods (whole grain breads, pasta underwent to the extrusion cooking) belong to the most important food sources of RS.
According to their origin and main properties, RS is usually divided into four types, RS type 1 to RS type 4 [19]. RS type 1 (RS1) is physically inaccessible to the digestive enzymes due to a specific polysaccharide-protein structure forming the protective barrier. Whole grains and seeds are the common sources [224]. The native granular form of certain raw foods, such as raw potatoes or green bananas, is typical for RS type 2 (RS2). The tightly packed granular structure of amylose, present in high concentration in RS2, limits access of digestive enzymes [225]. The above-mentioned cooking and subsequent cooling of certain food matrices (tubers) elicits realignment and recrystallization of the starch molecules that became resistant to saccharolytic enzymes. Cooked and cooled potatoes, pasta and rice thus contain considerable amounts of this RS type 3 (RS3; [221]). Chemically modified starches not naturally occurring in foods are categorized as RS type 4 (RS4). Their resistance to digestion is mainly based on the cross-linking of the starch macromolecules introduced by various industrial processes [226].
Particular microbial taxa or the species within the taxa might prefer a certain type of RS consumed by the host. Typical example is a predilection of Ruminococcus bromii for RS2 from high-amylose maize or of some Bacteroides species for RS3 from retrograded starch [19]. Accessibility and fermentability of RS can be influenced not only by the presence of other types of dietary fibers in the host gut, but also by intake of basic nutrients (proteins, lipids) and micronutrients [227].
As an example, diet rich in soluble fibers can increase the abundance of Lactobacillus spp., supporting the positive prebiotic effect of RS. On the other hand, a protein-rich diet might steer the colonic bacteria to protein fermentation with a negative outcome of ammonia production. Another important factor affecting gut microbiota composition is a duration of the RS intake. One-time consumption of RS can shift the colonic microbiota abundances relatively rapidly. However, an establishment in the gut of the microbial community more diversified and thus more resilient and more resistant to dysbiosis can be achieved by a long-time RS intake [19].
Similarly to dietary fibre (see part 5.1), RS is hydrolyzed by saccharolytic bacteria metabolizing complex carbohydrates and subsequently producing SCFAs, mainly acetate, propionate and butyrate [87]. Though both Bacteroidetes and Firmicutes are able to produce all these volatile fatty acids, representatives of the Bacteroidetes phylum produce predominantly propionate, species within the Firmicutes phylum are more specialized to butyrate production, Faecalibacterium prauznitzii and Eubacterium rectale being the principal producers [228].
As far as mechanisms of RS utilization in the colon are concerned, quantitatively and qualitatively most important RS fermenting bacteria belong to Bacteroidetes and Firmicutes phyla [160]. Bacteroides spp. within the Bacteroidetes phylum or Ruminococcus spp. within the phylum of Firmicutes are especially proficient to breakdown RS (among other complex carbohydrates) to produce SCFAs. Ruminococcus bromii, in particular, is instrumental in the first stages of RS degradation enabling other microbiota constituents to access the degradation products for further fermentation [19].
Actinobacteria phylum also contributes to RS fermentation in the colon, namely the genera of Bifidobacterium efficiently degrade RS to produce SCFAs with a consequence of lowering the colon pH, which renders this environment unfavourable for pathogenic bacteria [229].
Bacteria is not the only domain engaged in the RS fermentation in the colon. Methanobrevibacter smithii, methanogenic member of the domain Archaea, is able to incorporate hydrogen produced by other microorganisms during RS fermentation into methane, preventing hydrogen accumulation and suppression of the fermentation processes in the gut.
The most frequently mentioned positive effects of RS on the gut concern gut barrier function, modulation of inflammation and functioning of the gut immune system [19]. RS supports colonocyte turnover, thereby enhancing the epithelial barrier. RS fermentation products, SCFAs, contribute to upregulation of the expression of the tight junction proteins that are essential for maintaining the gut barrier integrity [230]. As far as modulation of inflammation is concerned, butyrate inhibits production of TNF-α and IL-6, pro-inflammatory cytokines contributing significantly to propagation of the inflammatory processes [231]. Moreover, butyrate positively affects immune cell differentiation, especially differentiation of naïve T-cells into regulatory T-cells (Tregs). Tregs are instrumental in maintaining gut immune homeostasis [232].
The three main types of current methods for RS determination comprise enzymatic, chromatographic and spectroscopic ones [19]. Megazyme resistant starch assay, an example of the advanced enzymatic techniques, simulates processes within the human digestive tract by an application of a series of relevant enzymes that remove accompanying components in the food sample matrix; remaining RS is quantified spectrophotometrically. The method provides data useful in nutritional labelling [223].
Chromatographic (high-performance liquid chromatography) separation and determination of the sample components after preceding enzymatic hydrolysis is able to measure molecular size and structure and thus to distinguish RS from other DF [234]. As far as spectroscopic methods are concerned, nuclear magnetic resonance spectroscopy (NMR) provides data regarding detailed RS structures on the molecular level and is therefore able to differentiate between particular types of RS [235].
6. Human Gut Microbiome and Obesity
Gut microbiome of the obese population is often associated with the intestinal dysbiosis characterized by a loss of microbial gene richness and thus a change in composition and metabolic activities of the intestinal microbiota [236,237,238]. Dysbiosis is related to the loss of commensal bacteria, excessive growth of pathogenic/conditionally pathogenic bacteria and loss of total microbial diversity [239,240]. For example, authors Liu et al. [36], Ciobârcă et al. [241] and Heiss and Olofsson [242] reported reduced gut microbiome diversity in obese individuals.
Obesity is often associated with increased Firmicutes/Bacteroidetes ratio, including low abundance of Bacteroidetes [6,243]. This finding confirmed Kasai et al. [244] and Koliada et al. [245] in Japanese or Ukrainian obese adult population, respectively. Increased levels of Firmicutes and decreased abundance of Bacteroidetes in obese individuals reported also Ley et al. [246]. Stojanov et al. [247] reported a substantially higher likelihood of the overweight with a Firmicutes/Bacteroidetes ratio of ≥ 1 in comparison with Firmicutes/Bacteroidetes ratio of < 1.
However, Breton et al. [7] point out that operating only on the level of phyla alone is imprecise. This statement is based, among other things, on findings of Cani et al. [248] that, despite some Firmicutes genera including Clostridium, Lactobacillus and Ruminococcus are increased in obesity, one of the most abundant Firmicutes species Faecalibacterium prauznitzii is decreased in this state. Similarly, increased abundance of Bacilli and decreased abundance of Clostridia, but not an association with Firmicutes/Bacteroidetes ratio was found out in American adults [249]. Similarly, Hu et al. [250] reported significant association of Bacteroides and Prevotella, but not Firmicutes/Bacteroidetes ratio, with obesity in Korean adolescents. Duncan et al. [251] also reported no relationships between BMI, body weight loss and Bacteroidetes abundance in obese and non-obese groups. Moreover, children obesity was associated with increased abundance of Proteobacteria, including positive correlation between Proteobacteria and fat intake in the study of Mendez-Salazar et al. [252].
Breton et al. [7] mention in this context discrepancies even within particular bacterial genera. Decreased levels of Lactobacillus paracasei, but increased abundances of Lactobacillus reuteri and Lactobacillus gasseri were found out in the stool of obese subjects in comparison with the lean ones [253]. So, Breton et al. [7] conclude that as far as obesity is concerned, no specific bacterial signature has been identified. Nevertheless, key obesity markers, such as increased adiposity, dyslipidemia and insulin resistance are associated with a relatively poor gut microbiota, meaning generally low bacterial gene pool [254].
The obese enterotypes, including abundances of the gut microbiome phyla/genera/species also depend on the stage of life: Mariat et al. [255]reported Firmicutes/Bacteroidetes ratio 0.4, 10.9 and 0.6 in infants, adults and elderly subjects. Abundances of the gut microbiota can also be rapidly changed as a consequence of dietary modifications [157].
The following factors relating gut microbiome composition to obesity can be suggested: gut microbiota-derived metabolites (SCFAs; secondary BAs), an ability of the gut microbiota to mediate low-grade inflammation [6] and effects of the gut microbiota on the intestinal immunity in obesogenic diets [9].
SCFAs produced during the fermentation of indigestible dietary substances (DF, RS) inhibit fat accumulation in adipose tissue, increase energy expenditure and stimulate production of the satiety-inducing hormones [256], such as GLP-1 and PYY with a consequence of decreasing weight gain [257].
One of the hallmarks of obesity is a state of chronic low-grade inflammation [258]. Bacteroidetes, as the Gram-negative bacteria, contain in their cell walls endotoxin lipopolysaccharide (LPS) that stimulates adipose tissue deposition, insulin resistance and increases an inflammation grade [259]. Trøseid et al. [260] reported elevated plasma LPS levels in obese individuals compared to healthy controls, including significant correlation between the intra-abdominal fat volume and plasma LPS level. On the other hand, higher abundances of Gram-positive bacteria Lactobacillus spp. and Bifidobacterium spp. within the gut microbiome are able to reduce intestinal permeability and improve systemic inflammation [261,262].
Cell-mediated and humoral innate intestinal immunity consists of Goblet cells producing mucus, Paneth cells able to secrete antimicrobial peptides (AMPs), intestinal epithelial cells, innate lymphoid cells and myeloid cells [9,263]. A direct involvement of intestinal microbiota in defensive mechanisms was demonstrated in the case of Akkermansia muciniphila that restored the obesogenic diet-induced down-regulation of AMPs [264]. Toll-like receptors (TLRs) and NOD-like receptors (NLRs), as principal pattern recognition receptors, sense microorganisms-associated patterns (MAMPs: LPS, flagellin, peptidoglycan) and so initiate intestinal immune activation at the cellular level [9].
As far as adaptive immunity is concerned, intestinal microbiota positively modifies intestinal T-helper 17 (Th 17) cells. Th 17 cells secrete interleukin 17 (IL-17) and IL-22 that induce production of AMPs and tight junction proteins and so protect gut barrier integrity [265]. Failure of the host defense as a consequence of reduced IgA (immunoglobulin A)+ cells and decreased IgA levels in a diet-induced obesity was demonstrated by Luck et al. [266]. Obesity was related with impairment of the IgA production also in the study of Petersen et al. [267], where increased amounts of Desulfovibrio spp. and decreased abundance in Clostridium spp. was reported.
Gut microbiota can affect host immune system in obesity also via structural and metabolic mediators [9]. Cani et al. [268] demonstrated involvement of LPS from Gram-negative bacteria in obesity-associated inflammation. On the other hand, flagellin is able to ameliorate diet-induced obesity [269] and muramyl dipeptide (derived from the bacterial peptidoglycan) reduced inflammation and insulin resistance via NOD2 receptor signaling in the study of Cavallari et al. [270]. Mazmanian et al. [271]reported induction of IL-10 in intestinal T-cells by the polysaccharide A (PSA) from the capsule of Bacteroides fragilis, which prevented intestinal inflammation; the immune reaction was suppressed by PSA from Bacteroides fragilis via activation of TLR2 on the CD4+ T-cells. TLR2 was also involved in a signaling pathway of a membrane protein from Akkermansia muciniphila exerting anti-obesity effects [272].
Apart from SCFAs and secondary BAs (see parts 4.1 and 4.2 of this study), at least two more microbiota-derived metabolites affect the host health status in obesity, including insulin resistance [9]. On the one hand, imidazole propionate, a histidine metabolite, belongs to the risk factors for insulin resistance and consequently for T2D, expansion of the imidazole propionate-producing bacteria can be elicited by dietary changes resulting to the shift in the gut environment [273]. On the other hand, an immune homeostasis can be boosted by microbial metabolites produced from tryptophan [274].
The key transcription factor in this context, widely expressed in immune cells, is aryl hydrocarbon receptor (AhR). Indoles, catabolites of tryptophan, are important AhR ligands, and the gut microbiota in subjects with metabolic syndrome were reported to show a reduced ability to metabolize tryptophan into effective AhR ligands [275]. Reduced synthesis of the AhR agonists can contribute to the obesity pathogenesis via decreasing production of IL-22. Gut microbiome of obese individuals shows an increased expression of indoleamine-2,3-dioxigenase, stimulating the kynurenine pathway. This leads to degradation, instead to production, of indole derivatives and decreased concentration of IL-22 with a consequence of chronic inflammation and decreased insulin sensitivity [276].
7. Conclusions
From the recent data concerning associations between gut microbiome, host diet and obesity follows that these relationships are extremely complicated, and despite the often-quoted microbiota-gut-brain axis as a potential target for interventions, deeper characterization of the associations is still missing. Relationships between gut microbiota and obesity are bidirectional and evidence to identify their temporal relations is still insufficient. No specific gut bacterial signature has been identified up to now as far as obesity is concerned. Moreover, gut microbiota may favour obesity via modulation of energy homeostasis, lipopolysaccharide-stimulated inflammation and fat deposition, though it is not clear, which specific bacterial communities contribute to the development of obesity. Findings regarding relationship between secondary bile acids (as products of the gut microbiota metabolism) and obesity are also still mostly inconclusive. It therefore be concluded that further experimental effort is necessary to fulfill the potential of the gut microbiome, including products of its metabolism SCFAs and BAs, in combination with a diet based on a sufficient amount of dietary fibre/resistant starch, to modulate obesity meaningfully.
Author Contributions
“Conceptualization, T.K. and O.P.; methodology, T.K., O.P., G.F.; software, O.P.; investigation, O.P., T.K., G.F.; resources, T.K., O.P., G.F.; data curation, T.K., O.P., G.F.; writing—original draft preparation, T.K., G.F.; writing—review and editing, O.P., T.K.; visualization, O.P.; supervision, T.K.; project administration, T.K.; funding acquisition, O.P. All authors have read and agreed to the published version of the manuscript.”
Funding
“This research received no external funding”
Acknowledgments
The study was supported by the Internal Grant Agency of the Faculty of AgriSciences of the Mendel University in Brno (project No. IGA24-AF-IP-048).
Conflicts of Interest
“The authors declare no conflicts of interest.”
References
- Haslam, D.W; James, W.T.P. Obesity. Lancet 2005, 366, 1197-1209. [CrossRef]
- Schwartz, M.W.; Seeley, R.J.; Zeltser, L.M.; Drewnowski, A.; Ravussin, E.; Redman, L.M.; Leibel, R.L. Obesity Pathogenesis: An Endocrine Society Scientific Statement. Endocr. Rev. 2017, 38, 267-296. [CrossRef]
- Blüher, M. Obesity: Global Epidemiology And Pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288-298. [CrossRef]
- Strohacker, K.; Carpenter, K.C.; McFarlin, B.K. Consequences of Weight Cycling: An Increase in Disease Risk? Int. J. Exerc. Sci. 2009, 2(3), 191-201.
- Dombrowski, S.U.; Knittle, K.; Avenell, A.; Araujo-Soares, V.; Sniehotta, F.F. Long Term Maintenance Of Weight Loss With Non-Surgical Interventions In Obese Adults: Systematic Review And Meta-Analyses Of Randomised Controlled Trials. BMJ 2014, 348, g2646-g2646. [CrossRef]
- Lim, Y.Y.; Lee, Y.S.; Ooi, D.S.Q. Engineering The Gut Microbiome For Treatment Of Obesity: A Review Of Current Understanding And Progress. Biotechnol. J. 2020, 15. [CrossRef]
- Breton, J.; Galmiche, M.; Déchelotte, P. Dysbiotic Gut Bacteria In Obesity: An Overview Of The Metabolic Mechanisms And Therapeutic Perspectives Of Next-Generation Probiotics. Microorganisms 2022, 10. [CrossRef]
- Moser, B.; Milligan, M.A.; Dao, M.C. The Microbiota-Gut-Brain Axis: Clinical Applications In Obesity And Type 2 Diabetes. Rev. Invest. Clin. 2022, 74. [CrossRef]
- Liébana-García, R.; Olivares, M.; Bullich-Vilarrubias, C.; López-Almela, I.; Romaní-Pérez, M.; Sanz, Y. The Gut Microbiota As A Versatile Immunomodulator In Obesity And Associated Metabolic Disorders. Best Pract. Res. Clin. Endoc. Metab. 2021, 35. [CrossRef]
- Pedroza Matute, S.; Iyavoo, S. Exploring The Gut Microbiota: Lifestyle Choices, Disease Associations, And Personal Genomics. Front. Nutr. 2023, 10. [CrossRef]
- Hodgkinson, K.; El Abbar, F.; Dobranowski, P.; Manoogian, J.; Butcher, J.; Figeys, D.; Mack, D.; Stintzi, A. Butyrate’s Role In Human Health And The Current Progress Towards Its Clinical Application To Treat Gastrointestinal Disease. Clin. Nutr. 2023, 42, 61-75. [CrossRef]
- Portincasa, P.; Bonfrate, L.; Vacca, M.; De Angelis, M.; Farella, I.; Lanza, E.; Khalil, M.; Wang, D.Q.-H.; Sperandio, M.; Di Ciaula, A. Gut Microbiota And Short Chain Fatty Acids: Implications In Glucose Homeostasis. Int. J. Mol. Sci. 2022, 23. [CrossRef]
- Fusco, W.; Lorenzo, M.B.; Cintoni, M.; Porcari, S.; Rinninella, E.; Kaitsas, F.; Lener, E.; Mele, M.C.; Gasbarrini, A.; Collado, M.C.; et al. Short-Chain Fatty-Acid-Producing Bacteria: Key Components Of The Human Gut Microbiota. Nutrients 2023, 15. [CrossRef]
- Li, R.; Andreu-Sánchez, S.; Kuipers, F.; Fu, J. Gut Microbiome And Bile Acids In Obesity-Related Diseases. Best Pract. Res. Clin. Endoc. Metab. 2021, 35. [CrossRef]
- Larabi, A.B.; Masson, H.L.P.; Bäumler, A.J. Bile Acids As Modulators Of Gut Microbiota Composition And Function. Gut Microbes 2023, 15. [CrossRef]
- Singh, J.; Metrani, R.; Shivanagoudra, S.R.; Jayaprakasha, G.K.; Patil, B.S. Review On Bile Acids: Effects Of The Gut Microbiome, Interactions With Dietary Fiber, And Alterations In The Bioaccessibility Of Bioactive Compounds. J. Agric. Food Chem. 2019, 67, 9124-9138. [CrossRef]
- Beane, K.E.; Redding, M.C.; Wang, X.; Pan, J.H.; Le, B.; Cicalo, C.; Jeon, S.; Kim, Y.J.; Lee, J.H.; Shin, E.-C.; et al. Effects Of Dietary Fibers, Micronutrients, And Phytonutrients On Gut Microbiome: A Review. Appl. Biol. Chem. 2021, 64. [CrossRef]
- Kok, C.R.; Rose, D.; Hutkins, R. Predicting Personalized Responses To Dietary Fiber Interventions: Opportunities For Modulation Of The Gut Microbiome To Improve Health. Annu. Rev. Food Sci. Technol. 2023, 14, 157-182. [CrossRef]
- Chen, Z.; Liang, N.; Zhang, H.; Li, H.; Guo, J.; Zhang, Y.; Chen, Y.; Wang, Y.; Shi, N. Resistant Starch And The Gut Microbiome: Exploring Beneficial Interactions And Dietary Impacts. Food Chem. X 2024, 21. [CrossRef]
- Dagan, T.; Roettger, M.; Bryant, D.; Martin, W. Genome Networks Root The Tree Of Life Between Prokaryotic Domains. Genome Biol. Evol. 2010, 2, 379-392. [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is The Healthy Gut Microbiota Composition? A Changing Ecosystem Across Age, Environment, Diet, And Diseases. Microorganisms 2019, 7. [CrossRef]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The Human Gut Microbiota: Metabolism And Perspective In Obesity. Gut Microbes 2018, 1-18. [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al. Enterotypes Of The Human Gut Microbiome. Nature 2011, 473, 174-180. [CrossRef]
- Putignani, L.; Del Chierico, F.; Petrucca, A.; Vernocchi, P.; Dallapiccola, B. The Human Gut Microbiota: A Dynamic Interplay With The Host From Birth To Senescence Settled During Childhood. Pediatr. Res. 2014, 76, 2-10. [CrossRef]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.M.; Yang, S.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K.; et al. Association Between Breast Milk Bacterial Communities And Establishment And Development Of The Infant Gut Microbiome. JAMA Pediatr. 2017, 171. [CrossRef]
- Zhang, P. Influence Of Foods And Nutrition On The Gut Microbiome And Implications For Intestinal Health. Int. J. Mol. Sci. 2022, 23. [CrossRef]
- David, L.A.; Materna, A.C.; Friedman, J.; Campos-Baptista, M.I.; Blackburn, M.C.; Perrotta, A.; Erdman, S.E.; Alm, E.J. Host Lifestyle Affects Human Microbiota On Daily Timescales. Genome Biol. 2014, 15. [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human Gut Microbiome Viewed Across Age And Geography. Nature 2012, 486, 222-227. [CrossRef]
- Bajinka, O.; Tan, Y.; Abdelhalim, K.A.; Özdemir, G.; Qiu, X. Extrinsic Factors Influencing Gut Microbes, The Immediate Consequences And Restoring Eubiosis. AMB Express 2020, 10. [CrossRef]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment Dominates Over Host Genetics In Shaping Human Gut Microbiota. Nature 2018, 555, 210-215. [CrossRef]
- Jeffery, I.B.; Claesson, M.J.; O'Toole, P.W.; Shanahan, F. Categorization Of The Gut Microbiota: Enterotypes Or Gradients? Nat. Rev. Microbiol. 2012, 10, 591-592. [CrossRef]
- Knights, D.; Ward, T.L.; McKinlay, C.E.; Miller, H.; Gonzalez, A.; McDonald, D.; Knight, R. Rethinking “Enterotypes.” Cell Host Microbe 2014, 16, 433-437. [CrossRef]
- Thursby, E.; Juge, N. Introduction To The Human Gut Microbiota. Biochem. J. 2017, 474, 1823-1836. [CrossRef]
- Patra, D.; Banerjee, D.; Ramprasad, P.; Roy, S.; Pal, D.; Dasgupta, S. Recent Insights Of Obesity-Induced Gut And Adipose Tissue Dysbiosis In Type 2 Diabetes. Front. Mol. Biosci. 2023, 10. [CrossRef]
- Vijay, A.; Valdes, A.M. Retracted Article Review: Role Of The Gut Microbiome In Chronic Diseases: A Narrative Review. Eur. J. Clin. Nutr. 2022, 76, 489-501. [CrossRef]
- Liu, B.-N.; Liu, X.-T.; Liang, Z.-H.; Wang, J.-H. Gut Microbiota In Obesity. World J. Gastroenterol. 2021, 27, 3837-3850. [CrossRef]
- Stewart, E.J. Growing Unculturable Bacteria. J. Bacteriol. 2012, 194, 4151-4160. [CrossRef]
- Kim, M.; Chun, J. 16S Rrna Gene-Based Identification Of Bacteria And Archaea Using The Eztaxon Server. In New Approaches to Prokaryotic Systematics; Methods in Microbiology; Elsevier, 2014; pp. 61-74 ISBN 9780128001769.
- Sanschagrin, S.; Yergeau, E. Next-Generation Sequencing Of 16S Ribosomal Rna Gene Amplicons. J. Vis. Exp. 2014. [CrossRef]
- Lim, M.Y.; Hong, S.; Kim, B.-M.; Ahn, Y.; Kim, H.-J.; Nam, Y.-D. Changes In Microbiome And Metabolomic Profiles Of Fecal Samples Stored With Stabilizing Solution At Room Temperature: A Pilot Study. Sci Rep 2020, 10. [CrossRef]
- Salipante, S.J.; Kawashima, T.; Rosenthal, C.; Hoogestraat, D.R.; Cummings, L.A.; Sengupta, D.J.; Harkins, T.T.; Cookson, B.T.; Hoffman, N.G.; Löffler, F.E. Performance Comparison Of Illumina And Ion Torrent Next-Generation Sequencing Platforms For 16S Rrna-Based Bacterial Community Profiling. Appl. Environ. Microbiol. 2014, 80, 7583-7591. [CrossRef]
- Byrd, D.A.; Sinha, R.; Hoffman, K.L.; Chen, J.; Hua, X.; Shi, J.; Chia, N.; Petrosino, J.; Vogtmann, E.; Rao, K. Comparison Of Methods To Collect Fecal Samples For Microbiome Studies Using Whole-Genome Shotgun Metagenomic Sequencing. mSphere 2020, 5, e00827-19. [CrossRef]
- Tessler, M.; Neumann, J.S.; Afshinnekoo, E.; Pineda, M.; Hersch, R.; Velho, L.F.M.; Segovia, B.T.; Lansac-Toha, F.A.; Lemke, M.; DeSalle, R.; et al. Large-Scale Differences In Microbial Biodiversity Discovery Between 16S Amplicon And Shotgun Sequencing. Sci Rep 2017, 7. [CrossRef]
- Souche, E.; Beltran, S.; Brosens, E.; Belmont, J.W.; Fossum, M.; Riess, O.; Gilissen, C.; Ardeshirdavani, A.; Houge, G.; van Gijn, M.; et al. Recommendations For Whole Genome Sequencing In Diagnostics For Rare Diseases. Eur. J. Hum. Genet. 2022, 30, 1017-1021. [CrossRef]
- Song, Z.; Cai, Y.; Lao, X.; Wang, X.; Lin, X.; Cui, Y.; Kalavagunta, P.K.; Liao, J.; Jin, L.; Shang, J.; et al. Taxonomic Profiling And Populational Patterns Of Bacterial Bile Salt Hydrolase (Bsh) Genes Based On Worldwide Human Gut Microbiome. Microbiome 2019, 7. [CrossRef]
- Combrink, L.; Humphreys, I.R.; Washburn, Q.; Arnold, H.K.; Stagaman, K.; Kasschau, K.D.; Jolles, A.E.; Beechler, B.R.; Sharpton, T.J. Best Practice For Wildlife Gut Microbiome Research: A Comprehensive Review Of Methodology For 16S Rrna Gene Investigations. Front. Microbiol. 2023, 14. [CrossRef]
- Beghini, F.; McIver, L.J.; Blanco-Míguez, A.; Dubois, L.; Asnicar, F.; Maharjan, S.; Mailyan, A.; Manghi, P.; Scholz, M.; Thomas, A.M.; et al. Integrating Taxonomic, Functional, And Strain-Level Profiling Of Diverse Microbial Communities With Biobakery 3. eLife 2021, 10. [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-Omics Of The Gut Microbial Ecosystem In Inflammatory Bowel Diseases. Nature 2019, 569, 655-662. [CrossRef]
- Karcher, N.; Pasolli, E.; Asnicar, F.; Huang, K.D.; Tett, A.; Manara, S.; Armanini, F.; Bain, D.; Duncan, S.H.; Louis, P.; et al. Analysis Of 1321 Eubacterium Rectale Genomes From Metagenomes Uncovers Complex Phylogeographic Population Structure And Subspecies Functional Adaptations. Genome Biol. 2020, 21. [CrossRef]
- Tett, A.; Huang, K.D.; Asnicar, F.; Fehlner-Peach, H.; Pasolli, E.; Karcher, N.; Armanini, F.; Manghi, P.; Bonham, K.; Zolfo, M.; et al. The Prevotella Copri Complex Comprises Four Distinct Clades Underrepresented In Westernized Populations. Cell Host Microbe 2019, 26, 666-679.e7. [CrossRef]
- Pinart, M.; Dötsch, A.; Schlicht, K.; Laudes, M.; Bouwman, J.; Forslund, S.K.; Pischon, T.; Nimptsch, K. Gut Microbiome Composition In Obese And Non-Obese Persons: A Systematic Review And Meta-Analysis. Nutrients 2022, 14. [CrossRef]
- Knight, R.; Vrbanac, A.; Taylor, B.C.; Aksenov, A.; Callewaert, C.; Debelius, J.; Gonzalez, A.; Kosciolek, T.; McCall, L.-I.; McDonald, D.; et al. Best Practices For Analysing Microbiomes. Nat. Rev. Microbiol. 2018, 16, 410-422. [CrossRef]
- Amir, A.; McDonald, D.; Navas-Molina, J.A.; Debelius, J.; Morton, J.T.; Hyde, E.; Robbins-Pianka, A.; Knight, R.; Arumugam, M. Correcting For Microbial Blooms In Fecal Samples During Room-Temperature Shipping. mSystems 2017, 2, e00199-16. [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A Human Gut Microbial Gene Catalogue Established By Metagenomic Sequencing. Nature 2010, 464, 59-65. [CrossRef]
- Postler, T.S.; Ghosh, S. Understanding The Holobiont: How Microbial Metabolites Affect Human Health And Shape The Immune System. Cell Metab. 2017, 26, 110-130. [CrossRef]
- Zhang, K.; Zhang, Q.; Qiu, H.; Ma, Y.; Hou, N.; Zhang, J.; Kan, C.; Han, F.; Sun, X.; Shi, J. The Complex Link Between The Gut Microbiome And Obesity-Associated Metabolic Disorders: Mechanisms And Therapeutic Opportunities. Heliyon 2024, 10. [CrossRef]
- Wu, J.; Wang, K.; Wang, X.; Pang, Y.; Jiang, C. The Role Of The Gut Microbiome And Its Metabolites In Metabolic Diseases. Protein Cell 2021, 12, 360-373. [CrossRef]
- Frolova, M.S.; Suvorova, I.A.; Iablokov, S.N.; Petrov, S.N.; Rodionov, D.A. Genomic Reconstruction Of Short-Chain Fatty Acid Production By The Human Gut Microbiota. Front. Mol. Biosci. 2022, 9. [CrossRef]
- Sonnenburg, E. D.; Sonnenburg, J. L. Starving Our Microbial Self: The Deleterious Consequences Of A Diet Deficient In Microbiota-Accessible Carbohydrates. Cell Metab. 2014, 20, 779-786. [CrossRef]
- Ayakdaş, G.; Ağagündüz, D. Microbiota-Accessible Carbohydrates (Macs) As Novel Gut Microbiome Modulators In Noncommunicable Diseases. Heliyon 2023, 9. [CrossRef]
- Topping, D.L.; Clifton, P.M. Short-Chain Fatty Acids And Human Colonic Function: Roles Of Resistant Starch And Nonstarch Polysaccharides. Physiol. Rev. 2001, 81, 1031-1064. [CrossRef]
- Topping, D.L. Short-chain fatty acids produced by intestinal bacteria. Asia. Pac. J. Clin. Nutr. 1996, 5, 15-19.
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free Fatty Acid Receptors In Health And Disease. Physiol. Rev. 2020, 100, 171-210. [CrossRef]
- Sivaprakasam, S.; Bhutia, Y.D.; Yang, S.; Ganapathy, V. Short-Chain Fatty Acid Transporters: Role in Colonic Homeostasis. Compr. Physiol. 2017, 8, 299-314. doi: 10.1002/cphy.c170014.
- Tabat, M.W.; Marques, T.M.; Markgren, M.; Löfvendahl, L.; Brummer, R.J.; Wall, R. Acute Effects Of Butyrate On Induced Hyperpermeability And Tight Junction Protein Expression In Human Colonic Tissues. Biomolecules 2020, 10. [CrossRef]
- Liang, L.; Liu, L.; Zhou, W.; Yang, C.; Mai, G.; Li, H.; Chen, Y. Gut Microbiota-Derived Butyrate Regulates Gut Mucus Barrier Repair By Activating The Macrophage/Wnt/Erk Signaling Pathway. Clin. Sci. 2022, 136, 291-307. [CrossRef]
- Blottiere, H.M.; Buecher, B.; Galmiche, J.-P.; Cherbut, C. Molecular Analysis Of The Effect Of Short-Chain Fatty Acids On Intestinal Cell Proliferation. Proc. Nutr. Soc. 2003, 62, 101-106. [CrossRef]
- Donohoe, D. R.; Garge, N.; Zhang, X.; Sun, W.; O'Connell, T. M.; Bunger, M. K.; Bultman, S. J. The Microbiome And Butyrate Regulate Energy Metabolism And Autophagy In The Mammalian Colon. Cell Metab. 2011, 13, 517-526. [CrossRef]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The Short-Chain Fatty Acid Acetate Reduces Appetite Via A Central Homeostatic Mechanism. Nat. Commun. 2014, 5. [CrossRef]
- González Hernández, M.A.; Canfora, E.E.; Jocken, J.W.E.; Blaak, E.E. The Short-Chain Fatty Acid Acetate In Body Weight Control And Insulin Sensitivity. Nutrients 2019, 11. [CrossRef]
- Yoshida, H.; Ishii, M.; Akagawa, M. Propionate Suppresses Hepatic Gluconeogenesis Via Gpr43/Ampk Signaling Pathway. Arch. Biochem. Biophys. 2019, 672. [CrossRef]
- den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.-J.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet–Induced Obesity Via A Pparγ-Dependent Switch From Lipogenesis To Fat Oxidation. Diabetes 2015, 64, 2398-2408. [CrossRef]
- Xiong, Y.; Miyamoto, N.; Shibata, K.; Valasek, M.A.; Motoike, T.; Kedzierski, R.M.; Yanagisawa, M. Short-Chain Fatty Acids Stimulate Leptin Production In Adipocytes Through The G Protein-Coupled Receptor Gpr41. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 1045-1050. [CrossRef]
- Slavin, J. Fiber And Prebiotics: Mechanisms And Health Benefits. Nutrients 2013, 5, 1417-1435. [CrossRef]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Di Yu; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation Of Inflammatory Responses By Gut Microbiota And Chemoattractant Receptor Gpr43. Nature 2009, 461, 1282-1286. [CrossRef]
- Zapolska-Downar, D.; Naruszewicz, M. Propionate reduces the cytokine-induced VCAM-1 and ICAM-1 expression by inhibiting nuclear factor-kappa B (NF-kappaB) activation. J. Physiol. Pharmacol. 2009, 60, 123-31.
- Cox, M.A.; Jackson, J.; Stanton, M.; Rojas-Triana, A.; Bober, L.; Laverty, M.; Yang, X.; Zhu, F.; Liu, J.; Wang, S.; et al. Short-Chain Fatty Acids Act As Antiinflammatory Mediatorsby Regulating Prostaglandin E2 And Cytokines. World J. of Gastroenterol. 2009, 15. [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role Of Short-Chain Fatty Acids From Gut Microbiota In Gut-Brain Communication. Front. Endocrinol. 2020, 11. [CrossRef]
- Yang, G.; Chen, S.; Deng, B.; Tan, C.; Deng, J.; Zhu, G.; Yin, Y.; Ren, W. Implication Of G Protein-Coupled Receptor 43 In Intestinal Inflammation: A Mini-Review. Front. Immunol. 2018, 9. [CrossRef]
- Tsukuda, N.; Yahagi, K.; Hara, T.; Watanabe, Y.; Matsumoto, H.; Mori, H.; Higashi, K.; Tsuji, H.; Matsumoto, S.; Kurokawa, K.; et al. Key Bacterial Taxa And Metabolic Pathways Affecting Gut Short-Chain Fatty Acid Profiles In Early Life. ISME J. 2021, 15, 2574-2590. [CrossRef]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.-zhong; Abe, F.; Osawa, R. Age-Related Changes In Gut Microbiota Composition From Newborn To Centenarian: A Cross-Sectional Study. BMC Microbiol. 2016, 16. [CrossRef]
- Ghosh, T.S.; Shanahan, F.; O’Toole, P.W. The Gut Microbiome As A Modulator Of Healthy Ageing. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 565-584. [CrossRef]
- Herrmann, E.; Young, W.; Reichert-Grimm, V.; Weis, S.; Riedel, C.; Rosendale, D.; Stoklosinski, H.; Hunt, M.; Egert, M. In Vivo Assessment Of Resistant Starch Degradation By The Caecal Microbiota Of Mice Using Rna-Based Stable Isotope Probing—A Proof-Of-Principle Study. Nutrients 2018, 10. [CrossRef]
- Sela, D.A.; Mills, D.A. Nursing Our Microbiota: Molecular Linkages Between Bifidobacteria And Milk Oligosaccharides. Trends Microbiol. 2010, 18, 298-307. [CrossRef]
- Conlon, M.; Bird, A. The Impact Of Diet And Lifestyle On Gut Microbiota And Human Health. Nutrients 2015, 7, 17-44. [CrossRef]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic Distribution Of Three Pathways For Propionate Production Within The Human Gut Microbiota. ISME J. 2014, 8, 1323-1335. [CrossRef]
- Louis, P.; Flint, H.J. Formation Of Propionate And Butyrate By The Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29-41. [CrossRef]
- Anand, S.; Kaur, H.; Mande, S.S. Comparative In Silico Analysis Of Butyrate Production Pathways In Gut Commensals And Pathogens. Front. Microbiol. 2016, 7. [CrossRef]
- Singh, V.; Lee, G.D.; Son, H.W.; Koh, H.; Kim, E.S.; Unno, T.; Shin, J.-H. Butyrate Producers, “The Sentinel Of Gut”: Their Intestinal Significance With And Beyond Butyrate, And Prospective Use As Microbial Therapeutics. Front. Microbiol. 2023, 13. [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role Of Human Gut Lachnospiraceae. Microorganisms 2020, 8. [CrossRef]
- Martín, R.; Bermúdez-Humarán, L.G.; Langella, P. Searching For The Bacterial Effector: The Example Of The Multi-Skilled Commensal Bacterium Faecalibacterium Prausnitzii. Front. Microbiol. 2018, 9. [CrossRef]
- Miquel, S.; Martín, R.; Rossi, O.; Bermúdez-Humarán, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium Prausnitzii And Human Intestinal Health. Curr. Opin. Microbiol. 2013, 16, 255-261. [CrossRef]
- Litvak, Y.; Byndloss, M.X.; Bäumler, A.J. Colonocyte Metabolism Shapes The Gut Microbiota. Science 2018, 362. [CrossRef]
- Rivera-Chávez, F.; Zhang, L. F.; Faber, F.; Lopez, C. A.; Byndloss, M. X.; Olsan, E. E.; Xu, G.; Velazquez, E. M.; Lebrilla, C. B.; Winter, S. E.; et al. Depletion Of Butyrate-Producing Clostridia From The Gut Microbiota Drives An Aerobic Luminal Expansion Of Salmonella. Cell Host Microbe 2016, 19, 443-454. [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-Chain Fatty Acids In Control Of Body Weight And Insulin Sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577-591. [CrossRef]
- HAMER, H.M.; JONKERS, D.; VENEMA, K.; VANHOUTVIN, S.; TROOST, F.J.; BRUMMER, R.-J. Review Article: The Role Of Butyrate On Colonic Function. Aliment. Pharmacol. Ther. 2008, 27, 104-119. [CrossRef]
- Kim, K.N.; Yao, Y.; Ju, S.Y. Short Chain Fatty Acids And Fecal Microbiota Abundance In Humans With Obesity: A Systematic Review And Meta-Analysis. Nutrients 2019, 11. [CrossRef]
- Lange, O.; Proczko-Stepaniak, M.; Mika, A. Short-Chain Fatty Acids—A Product Of The Microbiome And Its Participation In Two-Way Communication On The Microbiome-Host Mammal Line. Curr. Obes. Rep. 2023, 12, 108-126. [CrossRef]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota And Scfa In Lean And Overweight Healthy Subjects. Obesity 2010, 18, 190-195. [CrossRef]
- De la Cuesta-Zuluaga, J.; Mueller, N.; Álvarez-Quintero, R.; Velásquez-Mejía, E.; Sierra, J.; Corrales-Agudelo, V.; Carmona, J.; Abad, J.; Escobar, J. Higher Fecal Short-Chain Fatty Acid Levels Are Associated With Gut Microbiome Dysbiosis, Obesity, Hypertension And Cardiometabolic Disease Risk Factors. Nutrients 2019, 11. [CrossRef]
- Wang, Y.; Wang, H.; Howard, A.G.; Meyer, K.A.; Tsilimigras, M.C.B.; Avery, C.L.; Sha, W.; Sun, S.; Zhang, J.; Su, C.; et al. Circulating Short-Chain Fatty Acids Are Positively Associated With Adiposity Measures In Chinese Adults. Nutrients 2020, 12. [CrossRef]
- Monte, M.J.; Marin, J.J.G.; Antelo, A.; Vazquez-Tato, J. Bile Acids: Chemistry, Physiology, And Pathophysiology. World J. of Gastroenterol. 2009, 15. [CrossRef]
- Guzior, D.V.; Quinn, R.A. Review: Microbial Transformations Of Human Bile Acids. Microbiome 2021, 9. [CrossRef]
- Quinn, R.A.; Melnik, A.V.; Vrbanac, A.; Fu, T.; Patras, K.A.; Christy, M.P.; Bodai, Z.; Belda-Ferre, P.; Tripathi, A.; Chung, L.K.; et al. Global Chemical Effects Of The Microbiome Include New Bile-Acid Conjugations. Nature 2020, 579, 123-129. [CrossRef]
- Di Ciaula, A.; Garruti, G.; Lunardi Baccetto, R.; Molina-Molina, E.; Bonfrate, L.; Wang, D.Q.-H.; Portincasa, P. Bile Acid Physiology. Ann. Hepatol. 2017, 16, S4-S14. [CrossRef]
- Grüner, N.; Mattner, J. Bile Acids And Microbiota: Multifaceted And Versatile Regulators Of The Liver–Gut Axis. Int. J. Mol. Sci. 2021, 22. [CrossRef]
- Hofmann, A., F. The Enterohepatic Circulation Of Bile Acids In Mammals: Form And Functions. Front. Biosci. 2009, Volume. [CrossRef]
- Dawson, P.A.; Karpen, S.J. Intestinal Transport And Metabolism Of Bile Acids. J. Lipid Res. 2015, 56, 1085-1099. [CrossRef]
- Kliewer, S.A.; Mangelsdorf, D.J. Bile Acids As Hormones: The Fxr-Fgf15/19 Pathway. Dig. Dis. 2015, 33, 327-331. [CrossRef]
- Fiorucci, S.; Carino, A.; Baldoni, M.; Santucci, L.; Costanzi, E.; Graziosi, L.; Distrutti, E.; Biagioli, M. Bile Acid Signaling In Inflammatory Bowel Diseases. Dig. Dis. Sci. 2021, 66, 674-693. [CrossRef]
- Friedman, E.S.; Li, Y.; Shen, T.-C.D.; Jiang, J.; Chau, L.; Adorini, L.; Babakhani, F.; Edwards, J.; Shapiro, D.; Zhao, C.; et al. Fxr-Dependent Modulation Of The Human Small Intestinal Microbiome By The Bile Acid Derivative Obeticholic Acid. Gastroenterology 2018, 155, 1741-1752.e5. [CrossRef]
- Ginos, B.N.R.; Navarro, S.L.; Schwarz, Y.; Gu, H.; Wang, D.; Randolph, T.W.; Shojaie, A.; Hullar, M.A.J.; Lampe, P.D.; Kratz, M.; et al. Circulating Bile Acids In Healthy Adults Respond Differently To A Dietary Pattern Characterized By Whole Grains, Legumes And Fruits And Vegetables Compared To A Diet High In Refined Grains And Added Sugars: A Randomized, Controlled, Crossover Feeding Study. Metabolism 2018, 83, 197-204. [CrossRef]
- Vaz, F.M.; Ferdinandusse, S. Bile Acid Analysis In Human Disorders Of Bile Acid Biosynthesis. Mol. Asp. Med. 2017, 56, 10-24. [CrossRef]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The Impact Of Dietary Fiber On Gut Microbiota In Host Health And Disease. Cell Host Microbe 2018, 23, 705-715. [CrossRef]
- Jones, B.V.; Begley, M.; Hill, C.; Gahan, C.G.M.; Marchesi, J.R. Functional And Comparative Metagenomic Analysis Of Bile Salt Hydrolase Activity In The Human Gut Microbiome. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 13580-13585. [CrossRef]
- Joyce, S.A.; Gahan, C.G.M. Disease-Associated Changes In Bile Acid Profiles And Links To Altered Gut Microbiota. Dig. Dis. 2017, 35, 169-177. [CrossRef]
- Ridlon, J.M.; Kang, D.-J.; Hylemon, P.B. Bile Salt Biotransformations By Human Intestinal Bacteria. J. Lipid Res. 2006, 47, 241-259. [CrossRef]
- Doden, H.L.; Ridlon, J.M. Microbial Hydroxysteroid Dehydrogenases: From Alpha To Omega. Microorganisms 2021, 9. [CrossRef]
- Funabashi, M.; Grove, T.L.; Wang, M.; Varma, Y.; McFadden, M.E.; Brown, L.C.; Guo, C.; Higginbottom, S.; Almo, S.C.; Fischbach, M.A. A Metabolic Pathway For Bile Acid Dehydroxylation By The Gut Microbiome. Nature 2020, 582, 566-570. [CrossRef]
- Hofmann, A.F.; Sjövall, J.; Kurz, G.; Radominska, A.; Schteingart C.D.; Tint, G.S.; Vlachevic, Z.R.; Setchell, K.D.R. A proposed nomenclature for bile acids. J. Lipid Res. 1992, 33, 599-604. [CrossRef]
- Jia, B.; Park, D.; Chun, B.H.; Hahn, Y.; Jeon, C.O. Diet-Related Alterations Of Gut Bile Salt Hydrolases Determined Using A Metagenomic Analysis Of The Human Microbiome. Int. J. Mol. Sci. 2021, 22. [CrossRef]
- Ridlon, J.M.; Harris, S.C.; Bhowmik, S.; Kang, D.-J.; Hylemon, P.B. Consequences Of Bile Salt Biotransformations By Intestinal Bacteria. Gut Microbes 2016, 7, 22-39. [CrossRef]
- Gérard, P. Metabolism Of Cholesterol And Bile Acids By The Gut Microbiota. Pathogens 2014, 3, 14-24. [CrossRef]
- Urdaneta, V.; Casadesús, J. Interactions Between Bacteria And Bile Salts In The Gastrointestinal And Hepatobiliary Tracts. Front. Med. 2017, 4. [CrossRef]
- Sannasiddappa, T.H.; Lund, P.A.; Clarke, S.R. In Vitro Antibacterial Activity Of Unconjugated And Conjugated Bile Salts On Staphylococcus Aureus. Front. Microbiol. 2017, 8. [CrossRef]
- Kurdi, P.; Kawanishi, K.; Mizutani, K.; Yokota, A. Mechanism Of Growth Inhibition By Free Bile Acids In Lactobacilli And Bifidobacteria. J. Bacteriol. 2006, 188, 1979-1986. [CrossRef]
- Bachmann, V.; Kostiuk, B.; Unterweger, D.; Diaz-Satizabal, L.; Ogg, S.; Pukatzki, S.; Picardeau, M. Bile Salts Modulate The Mucin-Activated Type Vi Secretion System Of Pandemic Vibrio Cholerae. Plos Neglect. Trop. Dis. 2015, 9. [CrossRef]
- Abt, M.C.; McKenney, P.T.; Pamer, E.G. Clostridium Difficile Colitis: Pathogenesis And Host Defence. Nat. Rev. Microbiol. 2016, 14, 609-620. [CrossRef]
- Guinan, J.; Villa, P.; Thangamani, S. Secondary Bile Acids Inhibit Candida Albicans Growth And Morphogenesis. Pathog. Dis. 2018, 76. [CrossRef]
- Bustos, A.Y.; Font de Valdez, G.; Fadda, S.; Taranto, M.P. New Insights Into Bacterial Bile Resistance Mechanisms: The Role Of Bile Salt Hydrolase And Its Impact On Human Health. Food Res. Int. 2018, 112, 250-262. [CrossRef]
- Levy, M.; Blacher, E.; Elinav, E. Microbiome, Metabolites And Host Immunity. Curr. Opin. Microbiol. 2017, 35, 8-15. [CrossRef]
- de Diego-Cabero, N.; Mereu, A.; Menoyo, D.; Holst, J.J.; Ipharraguerre, I.R. Bile Acid Mediated Effects On Gut Integrity And Performance Of Early-Weaned Piglets. BMC Vet. Res. 2015, 11. [CrossRef]
- Verbeke, L.; Farre, R.; Verbinnen, B.; Covens, K.; Vanuytsel, T.; Verhaegen, J.; Komuta, M.; Roskams, T.; Chatterjee, S.; Annaert, P.; et al. The Fxr Agonist Obeticholic Acid Prevents Gut Barrier Dysfunction And Bacterial Translocation In Cholestatic Rats. Am. J. Pathol. 2015, 185, 409-419. [CrossRef]
- Xu, M.; Cen, M.; Shen, Y.; Zhu, Y.; Cheng, F.; Tang, L.; Hu, W.; Dai, N. Deoxycholic Acid-Induced Gut Dysbiosis Disrupts Bile Acid Enterohepatic Circulation And Promotes Intestinal Inflammation. Dig. Dis. Sci. 2021, 66, 568-576. [CrossRef]
- Lajczak, N.K.; Saint-Criq, V.; O’Dwyer, A.M.; Perino, A.; Adorini, L.; Schoonjans, K.; Keely, S.J. Bile Acids Deoxycholic Acid And Ursodeoxycholic Acid Differentially Regulate Human Β-Defensin-1 And -2 Secretion By Colonic Epithelial Cells. Faseb J. 2017, 31, 3848-3857. [CrossRef]
- Cipriani, S.; Mencarelli, A.; Chini, M.G.; Distrutti, E.; Renga, B.; Bifulco, G.; Baldelli, F.; Donini, A.; Fiorucci, S.; Ryffel, B. The Bile Acid Receptor Gpbar-1 (Tgr5) Modulates Integrity Of Intestinal Barrier And Immune Response To Experimental Colitis. PLoS One 2011, 6. [CrossRef]
- Ichikawa, R.; Takayama, T.; Yoneno, K.; Kamada, N.; Kitazume, M.T.; Higuchi, H.; Matsuoka, K.; Watanabe, M.; Itoh, H.; Kanai, T.; et al. Bile Acids Induce Monocyte Differentiation Toward Interleukin-12 Hypo-Producing Dendritic Cells Via A Tgr5-Dependent Pathway. Immunology 2012, 136, 153-162. [CrossRef]
- Vavassori, P.; Mencarelli, A.; Renga, B.; Distrutti, E.; Fiorucci, S. The Bile Acid Receptor Fxr Is A Modulator Of Intestinal Innate Immunity. J. Immunol. 2009, 183, 6251-6261. [CrossRef]
- Biagioli, M.; Carino, A.; Cipriani, S.; Francisci, D.; Marchianò, S.; Scarpelli, P.; Sorcini, D.; Zampella, A.; Fiorucci, S. The Bile Acid Receptor Gpbar1 Regulates The M1/M2 Phenotype Of Intestinal Macrophages And Activation Of Gpbar1 Rescues Mice From Murine Colitis. J. Immunol. 2017, 199, 718-733. [CrossRef]
- Haselow, K.; Bode, J.G.; Wammers, M.; Ehlting, C.; Keitel, V.; Kleinebrecht, L.; Schupp, A.-K.; Häussinger, D.; Graf, D. Bile Acids Pka-Dependently Induce A Switch Of The Il-10/Il-12 Ratio And Reduce Proinflammatory Capability Of Human Macrophages. J. Leukoc. Biol. 2013, 94, 1253-1264. [CrossRef]
- Chávez-Talavera, O.; Haas, J.; Grzych, G.; Tailleux, A.; Staels, B. Bile Acid Alterations In Nonalcoholic Fatty Liver Disease, Obesity, Insulin Resistance And Type 2 Diabetes: What Do The Human Studies Tell? Curr. Opin. Lipidology 2019, 30, 244-254. [CrossRef]
- Vincent, R.P.; Omar, S.; Ghozlan, S.; Taylor, D.R.; Cross, G.; Sherwood, R.A.; Fandriks, L.; Olbers, T.; Werling, M.; Alaghband-Zadeh, J.; et al. Higher Circulating Bile Acid Concentrations In Obese Patients With Type 2 Diabetes. Ann. Clin. Biochem. 2013, 50, 360-364. [CrossRef]
- Prinz, P.; Hofmann, T.; Ahnis, A.; Elbelt, U.; Goebel-Stengel, M.; Klapp, B.F.; Rose, M.; Stengel, A. Plasma Bile Acids Show A Positive Correlation With Body Mass Index And Are Negatively Associated With Cognitive Restraint Of Eating In Obese Patients. Front. Neurosci. 2015, 9. [CrossRef]
- Chen, L.; van den Munckhof, I.C.L.; Schraa, K.; ter Horst, R.; Koehorst, M.; van Faassen, M.; van der Ley, C.; Doestzada, M.; Zhernakova, D.V.; Kurilshikov, A.; et al. Genetic And Microbial Associations To Plasma And Fecal Bile Acids In Obesity Relate To Plasma Lipids And Liver Fat Content. Cell Reports 2020, 33. [CrossRef]
- Montagnana, M.; Danese, E.; Giontella, A.; Bonafini, S.; Benati, M.; Tagetti, A.; Dalbeni, A.; Cavarzere, P.; Gaudino, R.; Pucci, M.; et al. Circulating Bile Acids Profiles In Obese Children And Adolescents: A Possible Role Of Sex, Puberty And Liver Steatosis. Diagnostics 2020, 10. [CrossRef]
- Haeusler, R.A.; Astiarraga, B.; Camastra, S.; Accili, D.; Ferrannini, E. Human Insulin Resistance Is Associated With Increased Plasma Levels Of 12A-Hydroxylated Bile Acids. Diabetes 2013, 62, 4184-4191. [CrossRef]
- Chiang, J.Y.L.; Ferrell, J.M. Bile Acids As Metabolic Regulators And Nutrient Sensors. Annu. Rev. Nutr. 2019, 39, 175-200. [CrossRef]
- Chávez-Talavera, O.; Tailleux, A.; Lefebvre, P.; Staels, B. Bile Acid Control Of Metabolism And Inflammation In Obesity, Type 2 Diabetes, Dyslipidemia, And Nonalcoholic Fatty Liver Disease. Gastroenterology 2017, 152, 1679-1694.e3. [CrossRef]
- Preidis, G.A.; Kim, K.H.; Moore, D.D. Nutrient-Sensing Nuclear Receptors Pparα And Fxr Control Liver Energy Balance. J. Clin. Invest. 2017, 127, 1193-1201. [CrossRef]
- Sagar, N.M.; McFarlane, M.; Nwokolo, C.; Bardhan, K.D.; Arasaradnam, R.P. Mechanisms Of Triglyceride Metabolism In Patients With Bile Acid Diarrhea. World J. Gastroenterol. 2016, 22. [CrossRef]
- Ma, K. Farnesoid X Receptor Is Essential For Normal Glucose Homeostasis. J. Clin. Invest. 2006, 116, 1102-1109. [CrossRef]
- Kir, S.; Beddow, S.A.; Samuel, V.T.; Miller, P.; Previs, S.F.; Suino-Powell, K.; Xu, H.E.; Shulman, G.I.; Kliewer, S.A.; Mangelsdorf, D.J. Fgf19 As A Postprandial, Insulin-Independent Activator Of Hepatic Protein And Glycogen Synthesis. Science 2011, 331, 1621-1624. [CrossRef]
- Gowda, G.A.N.; Ijare, O.B.; Somashekar, B.S.; Sharma, A.; Kapoor, V.K.; Khetrapal, C.L. Single-Step Analysis Of Individual Conjugated Bile Acids In Human Bile Using 1 H Nmr Spectroscopy. Lipids 2006, 41, 591-603. [CrossRef]
- Yang, Y.; Griffiths, W.J.; Nazer, H.; Sjövall, J. Analysis of bile acids and bile alcohols in urine by capillary column liquid chromatography-mass spectrometry using fast atom bombardment or electrospray ionization and collision-induced dissociation. Biomed. Chromatogr. 1997, 11, 240-55.
- Griffiths, W.J.; Sjövall, J. Bile Acids: Analysis In Biological Fluids And Tissues. J. Lipid Res. 2009, 51, 23-41. [CrossRef]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect Of Diet On The Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11. [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly And Reproducibly Alters The Human Gut Microbiome. Nature 2014, 505, 559-563. [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns With Gut Microbial Enterotypes. Science 2011, 334, 105-108. [CrossRef]
- Xu, Z.; Knight, R. Dietary Effects On Human Gut Microbiome Diversity. Br. J. Nutr. 2015, 113, S1-S5. [CrossRef]
- Walker, A.W.; Ince, J.; Duncan, S.H.; Webster, L.M.; Holtrop, G.; Ze, X.; Brown, D.; Stares, M.D.; Scott, P.; Bergerat, A.; et al. Dominant And Diet-Responsive Groups Of Bacteria Within The Human Colonic Microbiota. ISME J. 2011, 5, 220-230. [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact Of Diet In Shaping Gut Microbiota Revealed By A Comparative Study In Children From Europe And Rural Africa. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 14691-14696. [CrossRef]
- Zimmer, J.; Lange, B.; Frick, J.-S.; Sauer, H.; Zimmermann, K.; Schwiertz, A.; Rusch, K.; Klosterhalfen, S.; Enck, P. A Vegan Or Vegetarian Diet Substantially Alters The Human Colonic Faecal Microbiota. Eur. J. Clin. Nutr. 2012, 66, 53-60. [CrossRef]
- Sidhu, S.R.K.; Kok, C.W.; Kunasegaran, T.; Ramadas, A. Effect Of Plant-Based Diets On Gut Microbiota: A Systematic Review Of Interventional Studies. Nutrients 2023, 15. [CrossRef]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence Of Diet On The Gut Microbiome And Implications For Human Health. J. Transl. Med. 2017, 15. [CrossRef]
- Kase, B.E.; Liese, A.D.; Zhang, J.; Murphy, E.A.; Zhao, L.; Steck, S.E. The Development And Evaluation Of A Literature-Based Dietary Index For Gut Microbiota. Nutrients 2024, 16. [CrossRef]
- Schoonakker, M.P.; van Peet, P.G.; van den Burg, E.L.; Numans, M.E.; Ducarmon, Q.R.; Pijl, H.; Wiese, M. Impact Of Dietary Carbohydrate, Fat Or Protein Restriction On The Human Gut Microbiome: A Systematic Review. Nutr. Res. Rev. 2024, 1-18. [CrossRef]
- Dai, Z.-L. Amino Acid Metabolism In Intestinal Bacteria: Links Between Gut Ecology And Host Health. Front. Biosci. 2011, 16. [CrossRef]
- Bartlett, A.; Kleiner, M. Dietary Protein And The Intestinal Microbiota: An Understudied Relationship. iScience 2022, 25. [CrossRef]
- Wan, Y.; Wang, F.; Yuan, J.; Li, J.; Jiang, D.; Zhang, J.; Li, H.; Wang, R.; Tang, J.; Huang, T.; et al. Effects Of Dietary Fat On Gut Microbiota And Faecal Metabolites, And Their Relationship With Cardiometabolic Risk Factors: A 6-Month Randomised Controlled-Feeding Trial. Gut 2019, 68, 1417-1429. [CrossRef]
- Clemente-Suárez, V.J.; Beltrán-Velasco, A.I.; Redondo-Flórez, L.; Martín-Rodríguez, A.; Tornero-Aguilera, J.F. Global Impacts Of Western Diet And Its Effects On Metabolism And Health: A Narrative Review. Nutrients 2023, 15. [CrossRef]
- Piernas, C.; Gao, M.; Jebb, S.A. Dietary Patterns Derived By Reduced Rank Regression And Non-Communicable Disease Risk. Proc. Nutr. Soc. 2022, 1-8. [CrossRef]
- Beam, A.; Clinger, E.; Hao, L. Effect Of Diet And Dietary Components On The Composition Of The Gut Microbiota. Nutrients 2021, 13. [CrossRef]
- Muscogiuri, G.; Cantone, E.; Cassarano, S.; Tuccinardi, D.; Barrea, L.; Savastano, S.; Colao, A. Gut Microbiota: A New Path To Treat Obesity. Int. J. Obes. Suppl. 2019, 9, 10-19. [CrossRef]
- Borrego-Ruiz, A.; Borrego, J.J. Human Gut Microbiome, Diet, And Mental Disorders. International Microbiology 2024. [CrossRef]
- Nagpal, R.; Neth, B.J.; Wang, S.; Craft, S.; Yadav, H. Modified Mediterranean-Ketogenic Diet Modulates Gut Microbiome And Short-Chain Fatty Acids In Association With Alzheimer's Disease Markers In Subjects With Mild Cognitive Impairment. EBioMedicine 2019, 47, 529-542. [CrossRef]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention Of Cardiovascular Disease With A Mediterranean Diet Supplemented With Extra-Virgin Olive Oil Or Nuts. N. Engl. J. Med. 2018, 378. [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-Level Adherence To A Mediterranean Diet Beneficially Impacts The Gut Microbiota And Associated Metabolome. Gut 2016, 65, 1812-1821. [CrossRef]
- Garcia-Mantrana, I.; Selma-Royo, M.; Alcantara, C.; Collado, M.C. Shifts On Gut Microbiota Associated To Mediterranean Diet Adherence And Specific Dietary Intakes On General Adult Population. Front. Microbiol. 2018, 9. [CrossRef]
- Pagliai, G.; Russo, E.; Niccolai, E.; Dinu, M.; Di Pilato, V.; Magrini, A.; Bartolucci, G.; Baldi, S.; Menicatti, M.; Giusti, B.; et al. Influence Of A 3-Month Low-Calorie Mediterranean Diet Compared To The Vegetarian Diet On Human Gut Microbiota And Scfa: The Cardiveg Study. Eur. J. Nutr. 2020, 59, 2011-2024. [CrossRef]
- Wang, Y.; Wymond, B.; Tandon, H.; Belobrajdic, D.P. Swapping White For High-Fibre Bread Increases Faecal Abundance Of Short-Chain Fatty Acid-Producing Bacteria And Microbiome Diversity: A Randomized, Controlled, Decentralized Trial. Nutrients 2024, 16. [CrossRef]
- Kaoutari, A.E.; Armougom, F.; Gordon, J.I.; Raoult, D.; Henrissat, B. The Abundance And Variety Of Carbohydrate-Active Enzymes In The Human Gut Microbiota. Nat. Rev. Microbiol. 2013, 11, 497-504. [CrossRef]
- Holscher, H.D. Dietary Fiber And Prebiotics And The Gastrointestinal Microbiota. Gut Microbes 2017, 8, 172-184. [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation Of The Human Colonic Microbiota: Introducing The Concept Of Prebiotics. J. Nutr. 1995, 125, 1401-1412. [CrossRef]
- Roberfroid, M. Prebiotics: The Concept Revisited1. J. Nutr. 2007, 137, 830S-837S. [CrossRef]
- Deehan, E.C.; Duar, R.M.; Armet, A.M.; Perez-Muñoz, M.E.; Jin, M.; Walter, J.; Britton, R.A.; Cani, P.D. Modulation Of The Gastrointestinal Microbiome With Nondigestible Fermentable Carbohydrates To Improve Human Health. Microbiol. Spectr. 2017, 5. [CrossRef]
- Delcour, J.A.; Aman, P.; Courtin, C.M.; Hamaker, B.R.; Verbeke, K. Prebiotics, Fermentable Dietary Fiber, And Health Claims. Adv. Nutr. 2016, 7, 1-4. [CrossRef]
- Guzman, J.R.; Conlin, V.S.; Jobin, C. Diet, Microbiome, And The Intestinal Epithelium: An Essential Triumvirate? Biomed Res. Int. 2013, 2013, 1-12. [CrossRef]
- Mogensen, T.H. Pathogen Recognition And Inflammatory Signaling In Innate Immune Defenses. Clin. Microbiol. Rev. 2009, 22, 240-273. [CrossRef]
- Lozupone, C.A.; Knight, R. Species Divergence And The Measurement Of Microbial Diversity. Fems Microbiol. Rev. 2008, 32, 557-578. [CrossRef]
- Cadotte, M.W.; Jonathan Davies, T.; Regetz, J.; Kembel, S.W.; Cleland, E.; Oakley, T.H. Phylogenetic Diversity Metrics For Ecological Communities: Integrating Species Richness, Abundance And Evolutionary History. Ecol. Lett. 2010, 13, 96-105. [CrossRef]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links Between Diet, Gut Microbiota Composition And Gut Metabolism. Proc. Nutr. Soc. 2015, 74, 13-22. [CrossRef]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary Fiber Intervention On Gut Microbiota Composition In Healthy Adults: A Systematic Review And Meta-Analysis. Am. J. Clin. Nutr. 2018, 107, 965-983. [CrossRef]
- Zhang, C.; Yin, A.; Li, H.; Wang, R.; Wu, G.; Shen, J.; Zhang, M.; Wang, L.; Hou, Y.; Ouyang, H.; et al. Dietary Modulation Of Gut Microbiota Contributes To Alleviation Of Both Genetic And Simple Obesity In Children. EBioMedicine 2015, 2, 968-984. [CrossRef]
- Simpson, H.L.; Campbell, B.J. Review Article: Dietary Fibre-Microbiota Interactions. Aliment. Pharmacol. Ther. 2015, 42, 158-179. [CrossRef]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Wang, J.; Sailer, M.; Theis, S.; Verbeke, K.; Raes, J. Prebiotic Inulin-Type Fructans Induce Specific Changes In The Human Gut Microbiota. Gut 2017, 66, 1968-1974. [CrossRef]
- Liu, F.; Li, P.; Chen, M.; Luo, Y.; Prabhakar, M.; Zheng, H.; He, Y.; Qi, Q.; Long, H.; Zhang, Y.; et al. Fructooligosaccharide (Fos) And Galactooligosaccharide (Gos) Increase Bifidobacterium But Reduce Butyrate Producing Bacteria With Adverse Glycemic Metabolism In Healthy Young Population. Sci Rep 2017, 7. [CrossRef]
- Hamaker, B.R.; Tuncil, Y.E. A Perspective On The Complexity Of Dietary Fiber Structures And Their Potential Effect On The Gut Microbiota. J. Mol. Biol. 2014, 426, 3838-3850. [CrossRef]
- Tuncil, Y.E.; Thakkar, R.D.; Arioglu-Tuncil, S.; Hamaker, B.R.; Lindemann, S.R.; Young, V.B. Subtle Variations In Dietary-Fiber Fine Structure Differentially Influence The Composition And Metabolic Function Of Gut Microbiota. mSphere 2020, 5, e00180-20. [CrossRef]
- Cantu-Jungles, T.M.; Hamaker, B.R. Tuning Expectations To Reality: Don’t Expect Increased Gut Microbiota Diversity With Dietary Fiber. The J. Nutr. 2023, 153, 3156-3163. [CrossRef]
- Bai, J.; Li, Y.; Li, T.; Zhang, W.; Fan, M.; Zhang, K.; Qian, H.; Zhang, H.; Qi, X.; Wang, L. Comparison Of Different Soluble Dietary Fibers During The In Vitro Fermentation Process. J. Agric. Food Chem. 2021, 69, 7446-7457. [CrossRef]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial Degradation Of Complex Carbohydrates In The Gut. Gut Microbes 2014, 3, 289-306. [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The Carbohydrate-Active Enzymes Database (Cazy) In 2013. Nucleic Acids Res. 2013, 42, D490-D495. [CrossRef]
- Cecchini, D.A.; Laville, E.; Laguerre, S.; Robe, P.; Leclerc, M.; Doré, J.; Henrissat, B.; Remaud-Siméon, M.; Monsan, P.; Potocki-Véronèse, G.; et al. Functional Metagenomics Reveals Novel Pathways Of Prebiotic Breakdown By Human Gut Bacteria. PLoS One 2013, 8. [CrossRef]
- Saito, Y.; Shigehisa, A.; Watanabe, Y.; Tsukuda, N.; Moriyama-Ohara, K.; Hara, T.; Matsumoto, S.; Tsuji, H.; Matsuki, T.; Zhou, N.-Y. Multiple Transporters And Glycoside Hydrolases Are Involved In Arabinoxylan-Derived Oligosaccharide Utilization In Bifidobacterium Pseudocatenulatum. Appl. Environ. Microbiol. 2020, 86, e01782-20. [CrossRef]
- Kim, C.C.; Healey, G.R.; Kelly, W.J.; Patchett, M.L.; Jordens, Z.; Tannock, G.W.; Sims, I.M.; Bell, T.J.; Hedderley, D.; Henrissat, B.; et al. Genomic Insights From Monoglobus Pectinilyticus: A Pectin-Degrading Specialist Bacterium In The Human Colon. ISME J. 2019, 13, 1437-1456. [CrossRef]
- Boger, M.C.L.; Lammerts van Bueren, A.; Dijkhuizen, L.; McBain, A.J. Cross-Feeding Among Probiotic Bacterial Strains On Prebiotic Inulin Involves The Extracellular Exo -Inulinase Of Lactobacillus Paracasei Strain W20. Appl. Environ. Microbiol. 2018, 84, e01539-18. [CrossRef]
- Delannoy-Bruno, O.; Desai, C.; Raman, A.S.; Chen, R.Y.; Hibberd, M.C.; Cheng, J.; Han, N.; Castillo, J.J.; Couture, G.; Lebrilla, C.B.; et al. Evaluating Microbiome-Directed Fibre Snacks In Gnotobiotic Mice And Humans. Nature 2021, 595, 91-95. [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria And Butyrate-Producing Colon Bacteria: Importance And Strategies For Their Stimulation In The Human Gut. Front. Microbiol. 2016, 7. [CrossRef]
- Koropatkin, N.M.; Cameron, E.A.; Martens, E.C. How Glycan Metabolism Shapes The Human Gut Microbiota. Nat. Rev. Microbiol. 2012, 10, 323-335. [CrossRef]
- Sheridan, P.; Martin, J.C.; Lawley, T.D.; Browne, H.P.; Harris, H.M.B.; Bernalier-Donadille, A.; Duncan, S.H.; O'Toole, P.W.; P. Scott, K.; J. Flint, H. Polysaccharide Utilization Loci And Nutritional Specialization In A Dominant Group Of Butyrate-Producing Human Colonic Firmicutes. Microb. Genomics 2016, 2. [CrossRef]
- Rodriguez, J.; Hiel, S.; Neyrinck, A.M.; Le Roy, T.; Pötgens, S.A.; Leyrolle, Q.; Pachikian, B.D.; Gianfrancesco, M.A.; Cani, P.D.; Paquot, N.; et al. Discovery Of The Gut Microbial Signature Driving The Efficacy Of Prebiotic Intervention In Obese Patients. Gut 2020, 69, 1975-1987. [CrossRef]
- Chen, T.; Long, W.; Zhang, C.; Liu, S.; Zhao, L.; Hamaker, B.R. Fiber-Utilizing Capacity Varies In Prevotella- Versus Bacteroides-Dominated Gut Microbiota. Sci Rep 2017, 7. [CrossRef]
- Christensen, L.; Roager, H.M.; Astrup, A.; Hjorth, M.F. Microbial Enterotypes In Personalized Nutrition And Obesity Management. Am. J. Clin. Nutr. 2018, 108, 645-651. [CrossRef]
- Van den Abbeele, P.; Duysburgh, C.; Ghyselinck, J.; Goltz, S.; Berezhnaya, Y.; Boileau, T.; De Blaiser, A.; Marzorati, M. Fructans With Varying Degree Of Polymerization Enhance The Selective Growth Of Bifidobacterium Animalis Subsp. Lactis Bb-12 In The Human Gut Microbiome In Vitro. Appl. Sci. 2021, 11. [CrossRef]
- Klimenko, N.S.; Tyakht, A.V.; Popenko, A.S.; Vasiliev, A.S.; Altukhov, I.A.; Ischenko, D.S.; Shashkova, T.I.; Efimova, D.A.; Nikogosov, D.A.; Osipenko, D.A.; et al. Microbiome Responses To An Uncontrolled Short-Term Diet Intervention In The Frame Of The Citizen Science Project. Nutrients 2018, 10. [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker Of Gut Dysbiosis In Obese Patients? Nutrients 2020, 12. [CrossRef]
- Mobeen, F.; Sharma, V.; Prakash, T. Enterotype Variations Of The Healthy Human Gut Microbiome In Different Geographical Regions. Bioinformation 2018, 14, 560-573. [CrossRef]
- Healey, G.; Murphy, R.; Butts, C.; Brough, L.; Whelan, K.; Coad, J. Habitual Dietary Fibre Intake Influences Gut Microbiota Response To An Inulin-Type Fructan Prebiotic: A Randomised, Double-Blind, Placebo-Controlled, Cross-Over, Human Intervention Study. Br. J. Nutr. 2018, 119, 176-189. [CrossRef]
- Smith, A.M. The Biosynthesis Of Starch Granules. Biomacromolecules 2001, 2, 335-341. [CrossRef]
- Sim, L.; Willemsma, C.; Mohan, S.; Naim, H.Y.; Pinto, B.M.; Rose, D.R. Structural Basis For Substrate Selectivity In Human Maltase-Glucoamylase And Sucrase-Isomaltase N-Terminal Domains. J. Biol. Chem. 2010, 285, 17763-17770. [CrossRef]
- Birt, D.F.; Boylston, T.; Hendrich, S.; Jane, J.-L.; Hollis, J.; Li, L.; McClelland, J.; Moore, S.; Phillips, G.J.; Rowling, M.; et al. Resistant Starch: Promise For Improving Human Health. Adv. Nutr. 2013, 4, 587-601. [CrossRef]
- Jane, J.L Structural Features Of Starch Granules II. In Starch; Elsevier, 2009; pp. 193-236 ISBN 9780127462752. doi: 10.1016/B978-0-12-746275-2.00006-9.
- Sajilata, M.G.; Singhal, R.S.; Kulkarni, P.R. Resistant Starch–A Review. Compr. Rev. Food. Sci. Food Saf. 2006, 5, 1-17. [CrossRef]
- McCleary, B.V.; Monaghan, D.A. Measurement Of Resistant Starch. J. AOAC Int. 2002, 85, 665-675. [CrossRef]
- Englyst, H.N.; Hudson, G.J. The classification and measurement of dietary carbohydrates. Food Chem. 1996, 57, 15-21. doi.org/10.1016/0308-8146(96)00056-8.
- Raigond, P.; Ezekiel, R.; Raigond, B. Resistant Starch In Food: A Review. J. Sci. Food Agric. 2015, 95, 1968-1978. [CrossRef]
- Macfarlane, S.; Macfarlane, G.T. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 2003, 62, 67-72. doi:10.1079/PNS2002207.
- Morrison, D.J.; Preston, T. Formation Of Short Chain Fatty Acids By The Gut Microbiota And Their Impact On Human Metabolism. Gut Microbes 2016, 7, 189-200. [CrossRef]
- Turroni, F.; Peano, C.; Pass, D.A.; Foroni, E.; Severgnini, M.; Claesson, M.J.; Kerr, C.; Hourihane, J.; Murray, D.; Fuligni, F.; et al. Diversity Of Bifidobacteria Within The Infant Gut Microbiota. PLoS One 2012, 7. [CrossRef]
- Suzuki, T. Regulation Of Intestinal Epithelial Permeability By Tight Junctions. Cell. Mol. Life Sci. 2013, 70, 631-659. [CrossRef]
- Segain, J.-P. Butyrate Inhibits Inflammatory Responses Through Nfkappa B Inhibition: Implications For Crohn's Disease. Gut 47, 397-403. [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal Microbe-Derived Butyrate Induces The Differentiation Of Colonic Regulatory T Cells. Nature 2013, 504, 446-450. [CrossRef]
- Krishnan, V.; Awana, M.; Samota, M.K.; Warwate, S.I.; Kulshreshtha, A.; Ray, M.; Bollinedi, H.; Singh, A.K.; Thandapilly, S.J.; Praveen, S.; et al. Pullulanase Activity: A Novel Indicator Of Inherent Resistant Starch In Rice (Oryza Sativa. L). Int. J. Biol. Macromol. 2020, 152, 1213-1223. [CrossRef]
- Barros, F.; Awika, J.; Rooney, L.W. Effect Of Molecular Weight Profile Of Sorghum Proanthocyanidins On Resistant Starch Formation. J. Sci. Food Agric. 2014, 94, 1212-1217. [CrossRef]
- Cao, R.; Liu, X.; Liu, Y.; Zhai, X.; Cao, T.; Wang, A.; Qiu, J. Applications Of Nuclear Magnetic Resonance Spectroscopy To The Evaluation Of Complex Food Constituents. Food Chem. 2021, 342. [CrossRef]
- Cuevas-Sierra, A.; Ramos-Lopez, O.; Riezu-Boj, J.I.; Milagro, F.I.; Martinez, J.A. Diet, Gut Microbiota, And Obesity: Links With Host Genetics And Epigenetics And Potential Applications. Adv. Nutr. 2019, 10, S17-S30. [CrossRef]
- Bien, J.; Palagani, V.; Bozko, P. The Intestinal Microbiota Dysbiosis And Clostridium Difficile Infection: Is There A Relationship With Inflammatory Bowel Disease? Ther. Adv. Gastroenterol. 2013, 6, 53-68. [CrossRef]
- Shanahan, F. The Colonic Microbiota In Health And Disease. Curr. Opin. Gastroenterol. 2013, 29, 49-54. [CrossRef]
- Peterson, C.T.; Sharma, V.; Elmén, L.; Peterson, S.N. Immune Homeostasis, Dysbiosis And Therapeutic Modulation Of The Gut Microbiota. Clin. Exp. Immunol. 2015, 179, 363-377. [CrossRef]
- DeGruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current Understanding Of Dysbiosis In Disease In Human And Animal Models. Inflamm. Bowel Dis. 2016, 22, 1137-1150. [CrossRef]
- Ciobârcă, D.; Cătoi, A.F.; Copăescu, C.; Miere, D.; Crișan, G. Bariatric Surgery In Obesity: Effects On Gut Microbiota And Micronutrient Status. Nutrients 2020, 12. [CrossRef]
- Heiss, C.N.; Olofsson, L.E. Gut Microbiota-Dependent Modulation Of Energy Metabolism. J. Innate Immun. 2018, 10, 163-171. [CrossRef]
- Verdam, F.J.; Fuentes, S.; de Jonge, C.; Zoetendal, E.G.; Erbil, R.; Greve, J.W.; Buurman, W.A.; de Vos, W.M.; Rensen, S.S. Human Intestinal Microbiota Composition Is Associated With Local And Systemic Inflammation In Obesity. Obesity 2013, 21. [CrossRef]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Tameda, M.; Shiraki, K.; Ito, M.; Takei, Y.; et al. Comparison Of The Gut Microbiota Composition Between Obese And Non-Obese Individuals In A Japanese Population, As Analyzed By Terminal Restriction Fragment Length Polymorphism And Next-Generation Sequencing. BMC Gastroenterol. 2015, 15. [CrossRef]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; et al. Association Between Body Mass Index And Firmicutes/Bacteroidetes Ratio In An Adult Ukrainian Population. BMC Microbiol. 2017, 17. [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity Alters Gut Microbial Ecology. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 11070-11075. [CrossRef]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence Of Probiotics On The Firmicutes/Bacteroidetes Ratio In The Treatment Of Obesity And Inflammatory Bowel Disease. Microorganisms 2020, 8. [CrossRef]
- Cani, P.D.; Moens de Hase, E.; Van Hul, M. Gut Microbiota And Host Metabolism: From Proof Of Concept To Therapeutic Intervention. Microorganisms 2021, 9. [CrossRef]
- Peters, B.A.; Shapiro, J.A.; Church, T.R.; Miller, G.; Trinh-Shevrin, C.; Yuen, E.; Friedlander, C.; Hayes, R.B.; Ahn, J. A Taxonomic Signature Of Obesity In A Large Study Of American Adults. Sci Rep 2018, 8. [CrossRef]
- Hu, H.-J.; Park, S.-G.; Jang, H.B.; Choi, M.-G.; Park, K.-H.; Kang, J.H.; Park, S.I.; Lee, H.-J.; Cho, S.-H.; Zoetendal, E.G. Obesity Alters The Microbial Community Profile In Korean Adolescents. PLoS One 2015, 10. [CrossRef]
- Duncan, S.H.; Lobley, G.E.; Holtrop, G.; Ince, J.; Johnstone, A.M.; Louis, P.; Flint, H.J. Human Colonic Microbiota Associated With Diet, Obesity And Weight Loss. Int. J. Obes. 2008, 32, 1720-1724. [CrossRef]
- Méndez-Salazar, E.O.; Ortiz-López, M.G.; Granados-Silvestre, M. de los Á.; Palacios-González, B.; Menjivar, M. Altered Gut Microbiota And Compositional Changes In Firmicutes And Proteobacteria In Mexican Undernourished And Obese Children. Front. Microbiol. 2018, 9. [CrossRef]
- Million, M.; Maraninchi, M.; Henry, M.; Armougom, F.; Richet, H.; Carrieri, P.; Valero, R.; Raccah, D.; Vialettes, B.; Raoult, D. Retracted Article: Obesity-Associated Gut Microbiota Is Enriched In Lactobacillus Reuteri And Depleted In Bifidobacterium Animalis And Methanobrevibacter Smithii. Int. J. Obes. 2012, 36, 817-825. [CrossRef]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S.; et al. Richness Of Human Gut Microbiome Correlates With Metabolic Markers. Nature 2013, 500, 541-546. [CrossRef]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimarăes, V.D.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J.-P. The Firmicutes/Bacteroidetes Ratio Of The Human Microbiota Changes With Age. BMC Microbiol. 2009, 9. [CrossRef]
- Larraufie, P.; Martin-Gallausiaux, C.; Lapaque, N.; Dore, J.; Gribble, F.M.; Reimann, F.; Blottiere, H.M. Scfas Strongly Stimulate Pyy Production In Human Enteroendocrine Cells. Sci Rep 2018, 8. [CrossRef]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The Short Chain Fatty Acid Propionate Stimulates Glp-1 And Pyy Secretion Via Free Fatty Acid Receptor 2 In Rodents. Int. J. Obes. 2015, 39, 424-429. [CrossRef]
- Mraz, M.; Haluzik, M. The Role Of Adipose Tissue Immune Cells In Obesity And Low-Grade Inflammation. J. Endocrinol. 2014, 222, R113-R127. [CrossRef]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes In Gut Microbiota Control Metabolic Endotoxemia-Induced Inflammation In High-Fat Diet–Induced Obesity And Diabetes In Mice. Diabetes 2008, 57, 1470-1481. [CrossRef]
- Trøseid, M.; Nestvold, T.K.; Rudi, K.; Thoresen, H.; Nielsen, E.W.; Lappegård, K.T. Plasma Lipopolysaccharide Is Closely Associated With Glycemic Control And Abdominal Obesity. Diabetes Care 2013, 36, 3627-3632. [CrossRef]
- Cani, P.D.; Neyrinck, A.M.; Fava, F.; Knauf, C.; Burcelin, R.G.; Tuohy, K.M.; Gibson, G.R.; Delzenne, N.M. Selective Increases Of Bifidobacteria In Gut Microflora Improve High-Fat-Diet-Induced Diabetes In Mice Through A Mechanism Associated With Endotoxaemia. Diabetologia 2007, 50, 2374-2383. [CrossRef]
- Cani, P.D.; Possemiers, S.; Van de Wiele, T.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M.; et al. Changes In Gut Microbiota Control Inflammation In Obese Mice Through A Mechanism Involving Glp-2-Driven Improvement Of Gut Permeability. Gut 2009, 58, 1091-1103. [CrossRef]
- Bevins, C.L.; Salzman, N.H. Paneth Cells, Antimicrobial Peptides And Maintenance Of Intestinal Homeostasis. Nat. Rev. Microbiol. 2011, 9, 356-368. [CrossRef]
- Everard, A.; Lazarevic, V.; Gaïa, N.; Johansson, M.; Ståhlman, M.; Backhed, F.; Delzenne, N.M.; Schrenzel, J.; François, P.; Cani, P.D. Microbiome Of Prebiotic-Treated Mice Reveals Novel Targets Involved In Host Response During Obesity. ISME J. 2014, 8, 2116-2130. [CrossRef]
- Tan, T.G.; Sefik, E.; Geva-Zatorsky, N.; Kua, L.; Naskar, D.; Teng, F.; Pasman, L.; Ortiz-Lopez, A.; Jupp, R.; Wu, H.-J.J.; et al. Identifying Species Of Symbiont Bacteria From The Human Gut That, Alone, Can Induce Intestinal Th17 Cells In Mice. Proc. Natl. Acad. Sci. U. S. A. 2016, 113. [CrossRef]
- Luck, H.; Khan, S.; Kim, J.H.; Copeland, J.K.; Revelo, X.S.; Tsai, S.; Chakraborty, M.; Cheng, K.; Tao Chan, Y.; Nøhr, M.K.; et al. Gut-Associated Iga+ Immune Cells Regulate Obesity-Related Insulin Resistance. Nat. Commun. 2019, 10. [CrossRef]
- Petersen, C.; Bell, R.; Klag, K.A.; Lee, S.-H.; Soto, R.; Ghazaryan, A.; Buhrke, K.; Ekiz, H.A.; Ost, K.S.; Boudina, S.; et al. T Cell–Mediated Regulation Of The Microbiota Protects Against Obesity. Science 2019, 365. [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic Endotoxemia Initiates Obesity And Insulin Resistance. Diabetes 2007, 56, 1761-1772. [CrossRef]
- Tran, H.Q.; Ley, R.E.; Gewirtz, A.T.; Chassaing, B. Flagellin-Elicited Adaptive Immunity Suppresses Flagellated Microbiota And Vaccinates Against Chronic Inflammatory Diseases. Nat. Commun. 2019, 10. [CrossRef]
- Cavallari, J.F.; Fullerton, M.D.; Duggan, B.M.; Foley, K.P.; Denou, E.; Smith, B.K.; Desjardins, E.M.; Henriksbo, B.D.; Kim, K.J.; Tuinema, B.R.; et al. Muramyl Dipeptide-Based Postbiotics Mitigate Obesity-Induced Insulin Resistance Via Irf4. Cell Metab. 2017, 25, 1063-1074.e3. [CrossRef]
- Mazmanian, S.K.; Round, J.L.; Kasper, D.L. A Microbial Symbiosis Factor Prevents Intestinal Inflammatory Disease. Nature 2008, 453, 620-625. [CrossRef]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A Purified Membrane Protein From Akkermansia Muciniphila Or The Pasteurized Bacterium Improves Metabolism In Obese And Diabetic Mice. Nat. Med. 2017, 23, 107-113. [CrossRef]
- Koh, A.; Molinaro, A.; Ståhlman, M.; Khan, M.T.; Schmidt, C.; Mannerås-Holm, L.; Wu, H.; Carreras, A.; Jeong, H.; Olofsson, L.E.; et al. Microbially Produced Imidazole Propionate Impairs Insulin Signaling Through Mtorc1. Cell 2018, 175, 947-961.e17. [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial Tryptophan Catabolites In Health And Disease. Nat. Commun. 2018, 9. [CrossRef]
- Natividad, J.M.; Agus, A.; Planchais, J.; Lamas, B.; Jarry, A.C.; Martin, R.; Michel, M.-L.; Chong-Nguyen, C.; Roussel, R.; Straube, M.; et al. Impaired Aryl Hydrocarbon Receptor Ligand Production By The Gut Microbiota Is A Key Factor In Metabolic Syndrome. Cell Metab. 2018, 28, 737-749.e4. [CrossRef]
- Laurans, L.; Venteclef, N.; Haddad, Y.; Chajadine, M.; Alzaid, F.; Metghalchi, S.; Sovran, B.; Denis, R.G.P.; Dairou, J.; Cardellini, M.; et al. Genetic Deficiency Of Indoleamine 2,3-Dioxygenase Promotes Gut Microbiota-Mediated Metabolic Health. Nat. Med. 2018, 24, 1113-1120. [CrossRef]
Figure 1.
Formation of short-chain fatty acids and their effects on the human organism. DF, dietary fiber; RS, resistant starch; MACs, microbiota-accessible carbohydrates; SCFAs, short-chain fatty acids; A, acetate; P, propionate; B, butyrate; CAZymes, carbohydrate-active enzymes; GHs, glycoside hydrolases; GTs, glycosyltransferases; PLs, polysaccharide lyases; CEs, carbohydrate esterases; GPR41, G protein-coupled receptor 41; ROS, reactive oxygen species; MCP-1, monocyte chemoattractant protein 1; VCAM-1, vascular cell adhesion molecule 1; CLDN-1, claudin-1; OCLN, occludin; ZO-1, zonula occludens-1; ATP, adenosine triphosphate; AhR, aryl hydrocarbon receptor; PPARγ, peroxisome proliferator-activated receptor gamma; HIF-1, hypoxia-inducible factor 1; NF-κB, nuclear factor kappa B; GLP-1, glucagon-like peptide 1; PYY, peptide YY; HDAC, histone deacetylase; p21Cip1, cyclin-dependent kinase inhibitor 1; D1 and D3, cyclins 1 and 3. Created with Biorender.com
Figure 1.
Formation of short-chain fatty acids and their effects on the human organism. DF, dietary fiber; RS, resistant starch; MACs, microbiota-accessible carbohydrates; SCFAs, short-chain fatty acids; A, acetate; P, propionate; B, butyrate; CAZymes, carbohydrate-active enzymes; GHs, glycoside hydrolases; GTs, glycosyltransferases; PLs, polysaccharide lyases; CEs, carbohydrate esterases; GPR41, G protein-coupled receptor 41; ROS, reactive oxygen species; MCP-1, monocyte chemoattractant protein 1; VCAM-1, vascular cell adhesion molecule 1; CLDN-1, claudin-1; OCLN, occludin; ZO-1, zonula occludens-1; ATP, adenosine triphosphate; AhR, aryl hydrocarbon receptor; PPARγ, peroxisome proliferator-activated receptor gamma; HIF-1, hypoxia-inducible factor 1; NF-κB, nuclear factor kappa B; GLP-1, glucagon-like peptide 1; PYY, peptide YY; HDAC, histone deacetylase; p21Cip1, cyclin-dependent kinase inhibitor 1; D1 and D3, cyclins 1 and 3. Created with Biorender.com
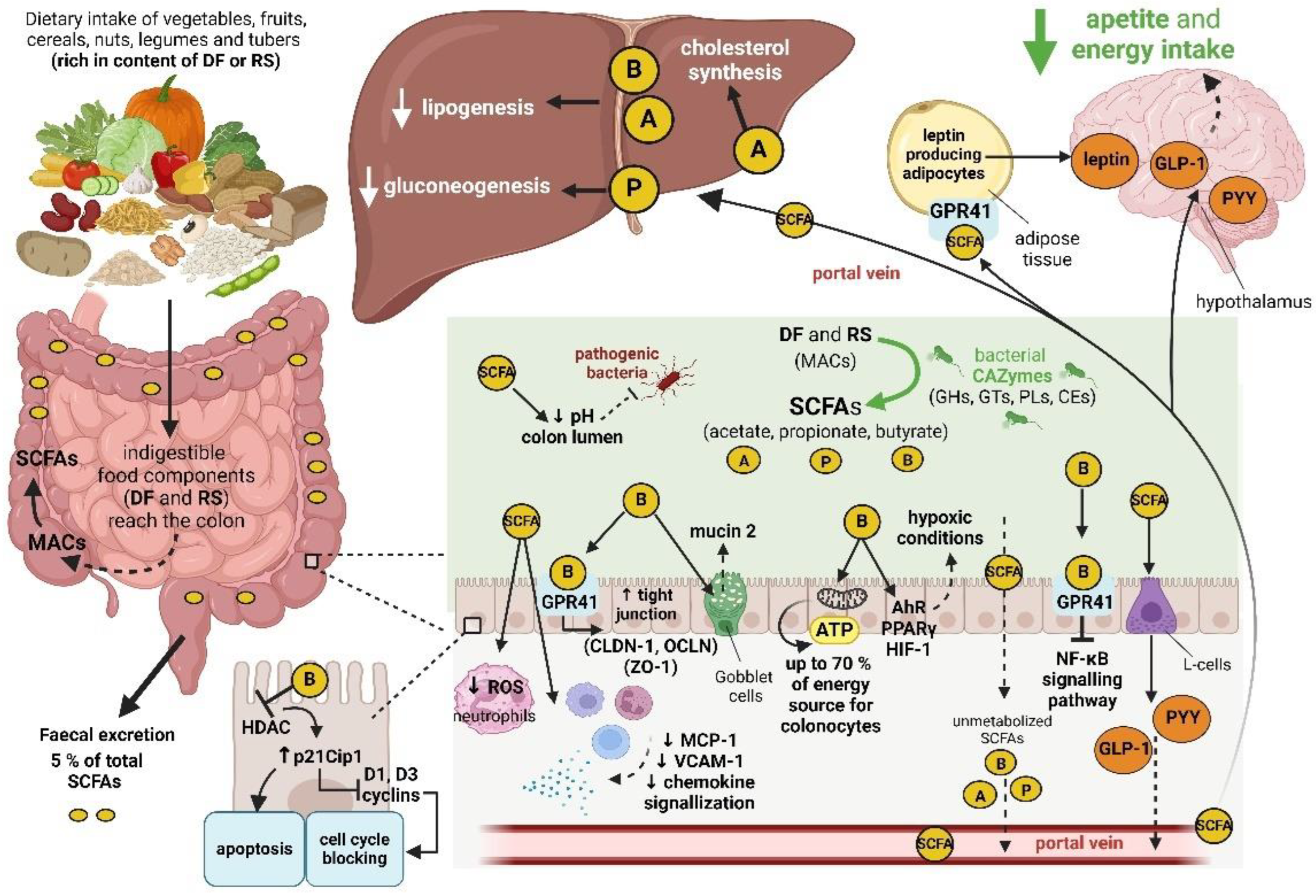
Figure 2.
Structure of quantitatively and qualitatively important primary and secondary bile acids.
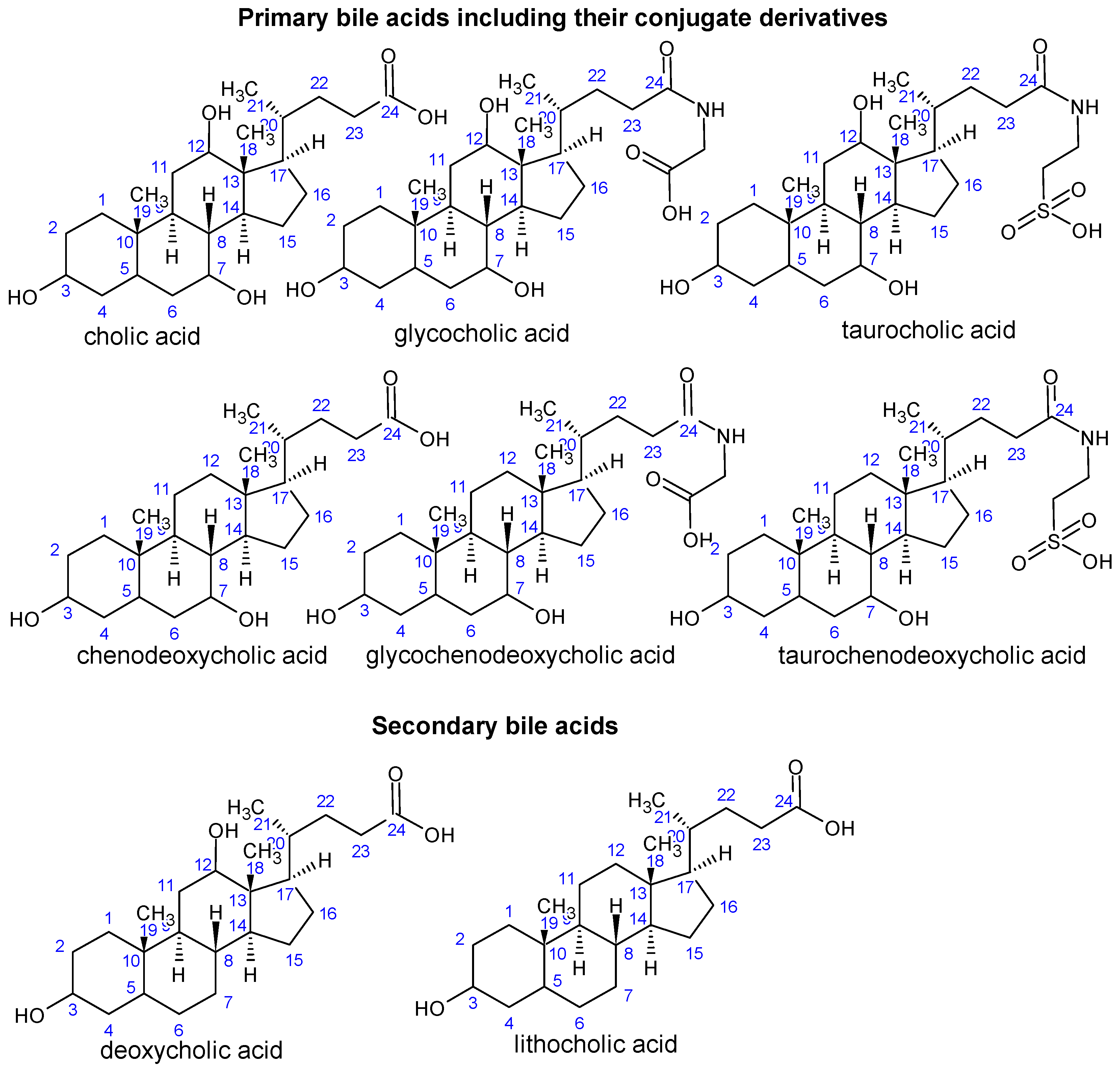
Figure 3.
Summary of the synthesis, reabsorption, biotransformation and physiological effects of bile acids in the human body. Abbreviations: CYP7A1, cholesterol 7α-hydroxylase; CYP8B1, sterol 12α-hydroxylase; CYP27A1, sterol 27α-hydroxylase; CYP7B1, 25-hydroxycholesterol 7α-hydroxylase; BAAT, bile acid-CoA:amino acid N-acyltransferase; TCA, taurocholic acid; GCA, glycocholic acid; TCDCA, taurochenodeoxycholic acid, GCDCA, glycochenodeoxycholic acid; CA, cholic acid; CDCA, chenodeoxycholic acid; DCA, deoxycholic acid; LCA, lithocholic acid; BAs, bile acids; BSHs, bile salt hydrolases; HSDHs, hydroxysteroid dehydrogenases; bai, BA-inducible bai operon; ASBT, apical sodium-dependent bile acid transporter; FXR, farnesoid X receptor; ZO-1, zonula occludens-1; TNF-α, tumor necrosis factor alpha; NF-κB, nuclear factor kappa B; AMPs, antimicrobial peptides; TGR5, Takeda G protein-coupled receptor 5; Mp; macrophages; Mc, monocytes; tDCs, tolerogenic dendritic cells; TGR5-cAMP-PKA, TGR5-cyclic adenosine monophosphate-protein kinase A; IL-1β, IL-6, IL-10, IL-18, interleukin 1 beta, 6, 10, 18; RXR, retinoid X receptor; FGF15/19, fibroblast growth factor 15 and 19; P, phosphorylation; ERKs, extracellular signal-regulated kinases; GNG, gluconeogenesis; GS, glycogen synthesis; SHP; small heterodimer partner; LPL, lipoprotein lipase; SREBP1, sterol regulatory element-binding protein 1; PPARα, peroxisome proliferator-activated receptor alpha; FA, fatty acids; TG, triglycerides; S1PR2, sphingosine-1-phosphate receptor 2; inflamm reg, inflammation regulation. Created with Biorender.com
Figure 3.
Summary of the synthesis, reabsorption, biotransformation and physiological effects of bile acids in the human body. Abbreviations: CYP7A1, cholesterol 7α-hydroxylase; CYP8B1, sterol 12α-hydroxylase; CYP27A1, sterol 27α-hydroxylase; CYP7B1, 25-hydroxycholesterol 7α-hydroxylase; BAAT, bile acid-CoA:amino acid N-acyltransferase; TCA, taurocholic acid; GCA, glycocholic acid; TCDCA, taurochenodeoxycholic acid, GCDCA, glycochenodeoxycholic acid; CA, cholic acid; CDCA, chenodeoxycholic acid; DCA, deoxycholic acid; LCA, lithocholic acid; BAs, bile acids; BSHs, bile salt hydrolases; HSDHs, hydroxysteroid dehydrogenases; bai, BA-inducible bai operon; ASBT, apical sodium-dependent bile acid transporter; FXR, farnesoid X receptor; ZO-1, zonula occludens-1; TNF-α, tumor necrosis factor alpha; NF-κB, nuclear factor kappa B; AMPs, antimicrobial peptides; TGR5, Takeda G protein-coupled receptor 5; Mp; macrophages; Mc, monocytes; tDCs, tolerogenic dendritic cells; TGR5-cAMP-PKA, TGR5-cyclic adenosine monophosphate-protein kinase A; IL-1β, IL-6, IL-10, IL-18, interleukin 1 beta, 6, 10, 18; RXR, retinoid X receptor; FGF15/19, fibroblast growth factor 15 and 19; P, phosphorylation; ERKs, extracellular signal-regulated kinases; GNG, gluconeogenesis; GS, glycogen synthesis; SHP; small heterodimer partner; LPL, lipoprotein lipase; SREBP1, sterol regulatory element-binding protein 1; PPARα, peroxisome proliferator-activated receptor alpha; FA, fatty acids; TG, triglycerides; S1PR2, sphingosine-1-phosphate receptor 2; inflamm reg, inflammation regulation. Created with Biorender.com
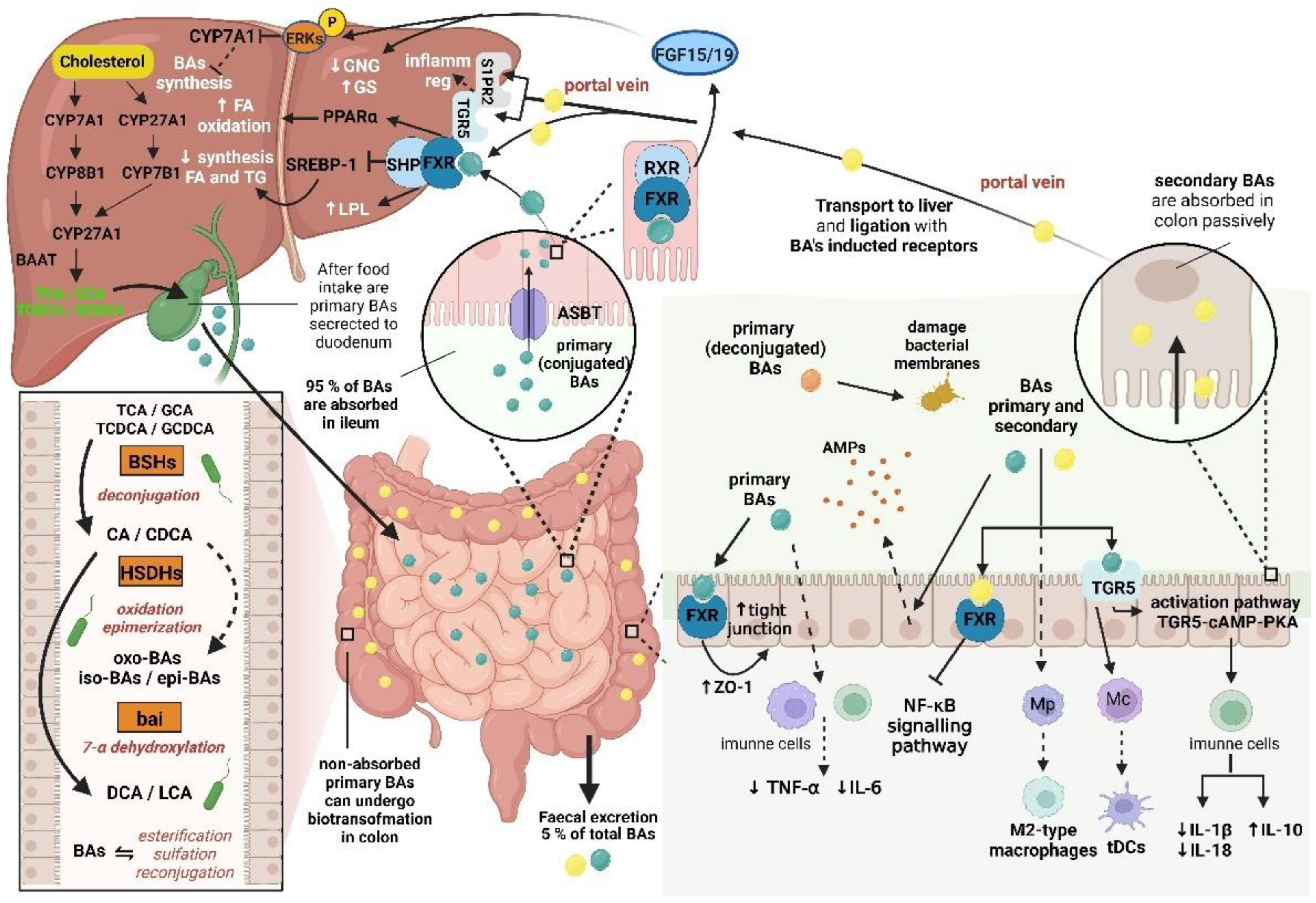
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



