
Submitted:
23 October 2024
Posted:
25 October 2024
You are already at the latest version
Abstract
Ziziphus tree species offer valuable socio-economic and ecological benefits but experience significant damage from insect pests. In Ethiopia, there is limited knowledge on the insects attacking Ziziphus fruits and a study aimed to identify these pests, assess their impact and understand how different land use types (LUTs), affect them were conducted. Sampling involved collecting fifty fruits and ten leaves from ten randomly chosen Ziziphus trees per LUT within each agroecological zone from August to December in 2022 and 2023. Samples were visually assessed for incidence and infestation levels, and the five morphotypes were identified using molecular techniques through phylogenetic analysis. Fruit pest incidence varied during the season yet a positive correlation (r = 0.84) was observed among the assessment months and years. Most fruits showed low to medium infestation levels (5–50%), while severe infestations (>75%) were predominant in the lowland agroecological zone. The insects attributed to the damage were identified as Carpomya incompleta (Becker), Drosophila hydei Sturtevant, D. simulans Sturtevant and Zaprionus indianus Gupta. Fruits showed higher incidence and infestation levels than leaves, indicating significant yield and income losses. Thus, implementing effective management strategies is vital to minimize these losses and achieve sustainable production in Ethiopia.
Keywords:
Land use
; farmland
; home garden
; roadside
; pest severity
; yield losses
1. Introduction
Ziziphus tree species are tropical resources that offer substantial economic, environmental, and social benefits to rural communities inhabiting arid and semi-arid regions [1,2]. The trees are steadily gaining attention among local communities owing to their resilience to harsh climatic conditions and consistent fruiting even amidst drought and climate change [3,4]. However, despite their manifold benefits and growing community interest, Ziziphus production encounters setbacks attributable to various biotic and abiotic factors. Among these, the most significant biotic hindrances arise from insect pests, which commonly afflict Ziziphus leaves, flowers and fruits [5,6]. The presence of insect pests poses a substantial risk factor in diminishing both the quantity and quality of Ziziphus fruits and leaves [7], resulting in significant losses in production [8,9,10]. The species of insect pests, their incidence, and intensity of these pests are subject to fluctuations in climatic conditions, seasonal changes, agricultural practices, ecological factors and the species of the Ziziphus tree [11].
Fruit flies, fruit borers, termites, stone weevils, bark-eating caterpillars, scale insects and mites are among the common insect pests of Ziziphus, each inflicting varying levels of infestation [12,13,14]. Tephrididae fruit flies have previously been reported as the primary menace among the plethora of insect pests affecting Ziziphus species., posing severe threats to fruit growth, yield, and quality, particularly affecting mature fruits [15,16]. Fruit fly infestations can result in significant reductions in fruit yield, ranging from 13–20% [17], 36–40% [12], and in extreme cases, up to 80–100% [11]. Additionally, Lepidopteran fruit borers cause substantial losses, with potential impacts of up to 70% [18]. Moreover, factors such as fruit maturity stage, seasonal and weather conditions, and human activities including industrial operations and residential encroachments contribute to the variation in severity of insect pest infestations [9,16].
Assessing and evaluating the incidence and infestation levels of insect pests impacting the quality and quantity of Ziziphus fruits and leaves across diverse land uses and seasons is essential for determining variations and pinpointing critical infestation periods throughout the year [19]. Moreover, identifying the insect pests is crucial for determining the economically important pests and devising improved integrated management strategies. Morphological and molecular identification techniques serve as appropriate methods for identifying specific types of pests. Morphological analysis entails investigating the morphological characters of different body parts, which is crucial for easily identifying the taxa [20,21]. While molecular species identification, utilising genetic markers, represents a valuable addition to complement traditional morphological methods [22,23], and an approach, often achieved through DNA barcoding, involves the use of short DNA sequences for species identification [24,25].
The production and utilisation of Ziziphus fruits for food and other purposes is becoming increasingly common in Ethiopia mainly among the rural communities in some parts of Ethiopia. It is more valued as food, feed and medicinal plant in northern parts of Ethiopia than in other parts of the country, particularly in some parts of Tigray and Amhara regions where frequent drought could cause food shortages. It is known that Ziziphus fruits are not only used for domestic consumption by rural communities but also sold in the markets as a supplementary source of cash income for households.
Despite its increasing significance as an important tree species of multiple uses, and given the fact that Ziziphus fruit production is challenged by biotic and abiotic agents, particularly by insect pests, pertinent information regarding the species of insect pests affecting Ziziphus fruits and their incidence and infestation levels in Ethiopia is notably lacking. There is also a dearth of knowledge concerning the correlation between Ziziphus fruit insect pest incidence and other underlying factors such as agroecology and seasonal variations in the country. The current study was therefore conducted to document the species of insect pests associated with Ziziphus fruit damage and to assess their incidence and infestation levels in Ethiopia.
2. Materials and Methods
2.1. Study sites
The study was conducted in three distinct land use types (LUTs), namely, farmland (FL), home garden (HG), and roadside (RS), in lowland and midland agro-ecological zones in Bosset district of East Shewa Zone of Oromia regional state and Bati district of Oromia Special zone of Amhara regional state, respectively, in Ethiopia (Figure 1). These two zones within the Oromia regional state and the Amhara regional state were deliberately chosen due to the presence of Ziziphus species trees. Geographically, Bosset and Bati are situated between 8° 34'59'' N and 39° 28'59'' E and 11° 11’ N and 40° 1’ E, respectively.
Bosset's elevation ranges from 1400 to 2500 meters above sea level, while Bati's elevation varies from 1800 to 2200 meters above sea level. The agro-ecological zones in the two districts also differ slightly in temperature, where temperatures fluctuate between 26°C and 34°C in Bosset and between 23°C and 30°C in Bati. Regarding precipitation, Bosset experiences abundant rainfall from June to August, a dry season from September to February, followed by a short rainy season from March to May. Similarly, Bati receives elevated rainfall between June and August, a dry spell stretching from September to February, and a short rainy season between March and April. Bosset receives a mean minimum and maximum annual rainfall of 600–900 mm, while Bati receives 550–700 mm [26].
2.2. Sampling and data collection
A three-stage stratified purposive sampling technique was employed to identify and select LUTs where Ziziphus species abundantly grow as well as areas where fruit collection and utilisation are actively practiced. As the initial stage, a reconnaissance survey was conducted in the North Omo Zone and Konso Zone of the Southern Nations, Nationalities and Peoples regional state, the East Shewa Zone of the Oromia regional state, and the Oromia Special Administrative Zone and the South Wollo Zone of the Amhara regional state together with the respective zonal natural resource experts to identify suitable field sites for the study. Subsequently, two zones, namely the East Shewa zone of the Oromia regional state and the Oromia Special Zone of the Amhara regional state, were selected based on the availability of Ziziphus trees and the culture of collecting and utilization of Ziziphus fruits. In the second stage, two suitable districts, one in each of the selected zones namely, Bosset district from the East Shewa Zone and Bati district from the Oromia Special Administrative Zone, were selected. In the third stage, three LUTs; FL, HG, and RS, were identified within each of the selected districts.
After the selection of suitable LUTs, a total of 30 trees from each of the districts (10 from each LUT) were randomly selected following FAO guidelines [27] and evaluated for incidence [28] and level of infestation [29]. Accordingly, approximately 50±15 fruits and 10±5 leaves per tree were collected randomly from ten trees from each of the LUTs in Bosset during the period from August to November, and Bati during October–December in 2022 and 2023. Those months were selected for fruit collection because fruit maturation occurs in those months at the two study sites. Fruit collection involved shaking trees and beating the branches with long sticks followed by collecting the fruits that fell to the ground in paper bags that were properly labelled with the district name, land use type, tree number, and date of collection. The fruits of each tree were transported from the field to the base station and separately sorted according to level of infestation into different categories based on criteria established by [29], which categorise infestation levels as very low (≤ 5%), low (6–10%), medium (11-20%), severe (21–50%) and very severe (>50%).
Similarly, leaf samples were collected from each of the ten trees from three randomly selected branches, one each from the lower, middle, and top of the crown at the rate of 10 leaves per branch and a total of 30 leaves per tree in a paper bag labelled as described above in the case of fruit collection. The leaf samples were transported to the base camp for further evaluation. At the base camp, after mixing the 30 leaves obtained from the three levels of the crown of a sampled tree, 10 leaves per tree were randomly selected for evaluation of the incidence and level of infestation by insect pests through the visual scoring method ranging from 0 to 5 described by [28]. According to this method, the different levels of leaf infestation are 0 = nil, no infestation; 1 = low (less than 5% of leaf area affected); 2 = medium (5-20% of leaf area affected); 3 = severe (20-50% of leaf area affected); 4 = very severe (50-80% of leaf area affected); and 5 = extreme damage (80-100% of leaves affected), complete or near-complete defoliation. The incidence of insect pests was calculated as a percentage of the total fruits and leaves inspected.
2.3. Insect rearing and identifications
To document and characterize insect pests affecting Ziziphus fruits in the study area, 15 fruits exhibiting the signs of insect damage were randomly selected per LUT and incubated in insect-rearing cages at the forest entomology and pathology laboratory of the Ethiopian Forestry Development (EFD) in Addis Ababa, Ethiopia for 14-21 days. The insect-rearing cages were daily monitored to follow up on the development of the emerging insects followed by adult counts every seven days. Matured insects were transferred to a killing jar containing a cotton swab immersed in 97% ethanol and transferred, using forceps, to clean vials (20 mL) for preservation. Insect specimens were stored at the EFD laboratory for DNA extraction until all necessary adults were collected from all study sites. Proper labelling comprising the site of collection, collection date, agroecological zone, and LUTs from where fruits were collected was ensured both for the rearing cages and insect preservation vials.
Identification of insects based on morphological features including wings, legs, head, body colours, antennae, thorax, and abdominal size and associated structures following available catalogues and identification keys [20,21]. Examination of insect structures was carried out using EMZ-5 binocular 7X-45X Zoom Stereo Microscope and photographs were taken using a digital camera and sorted in morphotypes for documentation. Subsequently, based on similarity in morphological features, the insect specimens were categorized into ten different groups assumed to be different species. For the identification of the insects, besides the morphological features, molecular data was also employed. Molecular species identification, utilising genetic markers, represents a valuable addition to complement traditional morphological methods [22,30]. This approach, often achieved through DNA barcoding, involves the use of short DNA sequences for species identification [24,25]. Accordingly, the morphological categorization was followed by the sequencing of a genetic marker Cytochrome c oxidase subunit 1 (COI) for molecular identification [33]. Polymerase chain reaction (PCR) and PCR-based typing methods serve as superior markers, making them ideal tools for species diagnosis [32,33].
2.4. DNA extraction
Following the morphological categorization of the insect specimens as described above, two insect specimens from each of the morphological groups were randomly selected for genomic DNA extraction. DNA was extracted from whole insect bodies [34] using the Quick-Start Protocol, and extraction of genomic DNA was conducted at the Holleta Agricultural Research Centre, National Biotechnology Institute, Holleta, Ethiopia. The DNeasy Blood and Tissue Kit (cat. nos. 69504 and 69506) was employed during the DNA extraction process. The DNA concentration was determined by an ultraviolet fluorescence spectrophotometer (Eppendorf, Germany), and the DNA was stored at −20 °C [35].
2.5. Polymerase Chain Reaction (PCR), DNA sequencing and phylogenetic analyses
The amplification and sequencing of the COI region of DNA was conducted at the molecular biology laboratory of the International Livestock Research Institute (ILRI) in Nairobi, Kenya. Amplification of the mitochondrial COI subunit 1 was carried out using the forward primer, LCO1490 (5'-ggtcaacaaatcataaagatattgg-3') and the reverse primer, HC02198 (5'-taaacttcagggtgaccaaaaaatca-3') using 2 U of Fast Start Taq DNA polymerase in a 20-μl reaction under standard conditions (L and H refer to light and heavy DNA strands and the numbers (1490 and 2198) refer to the position of the 5' nucleotide, while the 3' end of each primer is on a second-position nucleotide) [36]. Four ladders (designated as molecular markers), each approximately 710 base pairs in size, confirmed the successful amplification of the COI target region. PCR was performed (200 µM each dNTP, 200 nM each primer, 2 mM MgCl2) with 2 µl of the DNA extract as a template using the forward and reverse primers. The DNA fragments were amplified through 35 cycles at the following parameters: two minutes at 95°C, one minute at 40°C, and one and a half minutes at 72°C followed by a final extension step at 72°C for seven minutes. PCR products were confirmed by gel electrophoresis using 1.5% w/v Hi-Res standard agarose/1X TAE gel (Cambridge reagents, Thermo Scientific), stained with the dye GelRed Nucleic Acid Gel Stain.
The amplified DNA fragments were sequenced employing the Sanger dideoxy method [37]. Purified PCR product (25 ng/1μL), primer (5 pmol/μL), and Big Dye Terminator v3.1 (0.5μL) were brought to 10 μL in molecular H2O and sequenced using the protocol: 2 min at 96°C (10 s at 96°C, 10 s at 50°C, and 4 min at 60°C) × 30, and final hold at 72°C for 4 min in an ABI 9700 thermocycler. Sequencing reactions were analysed on an Applied Biosystems 3730 DNA Analyzer. All sequences obtained from the genetic analyser were edited using BioEdit v. 7.7.1 bioinformatics software and consensus sequences were obtained for each of the sequenced specimens [39]. After the consensus sequences were obtained, the DNA sequences were saved in FASTA format and were sent as query sequences to the National Centre for Biotechnology Information (NCBI) database to be verified through the Basic Local Alignment Search Tool (BLAST) [39]. Accordingly, after the BLAST search was performed in the NCBI database, matching sequences from the database providing possible species names with varying percentages of similarity with the query sequence along with additional descriptive information on the insect it originated from were obtained. Subsequently, the best matching sequences were saved and used as reference taxa for phylogenetic analysis and thus to confirm the identity of the insect species obtained from Ziziphus fruits collected during the current study.
For subsequent phylogenetic analysis, explorations of identical sequence arrangements were made using BLASTN, and the identities of the closest relatives of the sequences (similarity ≥ 99%) were retrieved from the GenBank. The evolutionary history and relationships of taxa were inferred using the neighbor-joining method [40]. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) is shown next to the branches [41]. The tree was drawn to scale, with branch lengths (next to the branches) in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the p-distance method [42] and presented in the units of the number of base differences per site. The analysis encompassed 8, 12, 7, 9, and 13 nucleotide sequences for C. incompleta, D. hydei, Z. indianus, D. simulans, and P. concolor, respectively. All positions containing gaps and missing data were eliminated using the complete deletion method. The final dataset comprised a total of 524, 450, 223, 597, and 532 positions for C. incompleta, D. hydei, Z. indianus, D. simulans, and P. concolor, respectively. Evolutionary analyses were conducted using Molecular Evolution Genetic Analysis (MEGA) MEGA11 software [43].
2.6. Statistical analysis
A one-way analysis of variance (ANOVA) was employed to examine variations in the percentage of incidence and infestation levels across the different LUTs, agroecological zones and fruit production years. The normal distribution of the data set was tested using the Shapiro-Wilk test and considered significant at p ≥ 0.05. Leven's test was used to calculate the homogeneity of variance of the data. The data exhibited normal distribution and the significance of mean differences was assessed through Tukey’s Honest Significant Difference (Tukey’s HSD) post hoc test, with a significance level set at P ≤ 0.05. The Pearson correlation coefficient (r) was calculated to assess the relationships between insect pest incidences, and agroecological zones, LUTs, assessment months and fruit production years. Furthermore, linear regression analysis was employed to identify the factors contributing to the high incidence of insect pests on Ziziphus trees. All statistical analyses were performed using R free software version 4.3.2 [44].
3. Results
3.1. Factors influencing Ziziphus fruit and leaves insect pest incidences
The visual examination revealed insect pest infestations in both Ziziphus fruits and leaves and the damage was prevalent across all the study sites. Various symptoms of damage were observed on both fruits and leaves caused by various insect pests. Insect damage was evident across all agroecological zones, land uses, and throughout the two-year assessment period. The timing of fruit collection (month) and production years were significant factors influencing the incidence of Ziziphus fruit insects, where variations were observed within months of each year (Table 1). Only the production year was found to influence leaf insect pest incidence percentage. Agroecological zone and land use did not influence leaves insect pest incidence (Table 1).
The incidence and infestation levels on Ziziphus fruits varied across assessment months in each fruit production year. The highest incidence occurred in November 2022, while it was in November and December 2023. The lowest percentage of incidence was recorded in September and December 2022 and in September 2023 (Figure 2).
Insect pest incidence exhibited minimal correlation with the factors examined in this study. A significant positive correlation (P < 0.05) was observed between land use types and AEZ, as well as assessment months and production years over the two-year study period for both fruits and leaves. However, no correlation was identified between the assessment months and the two AEZ (Table 2).
3.2. Ziziphus tree fruits and leaves insect pests infestation levels
The average level of non-infested fruits per tree ranged from 6% to 13%. Most fruits had low to medium infestation levels, consistently observed over the study years. Significant variations emerged in severe- and very severe level infestation between the two assessment years, peaking in the 2023 fruiting season (Table 3). In contrast, insect pest occurrence on Ziziphus leaves was notably low, with the majority showing no signs of infestation, indicating overall leaf health. No significant differences were found among the three LUT regarding leaf infestation levels (Table 3). However, despite the low infestation level, there was notable variation between the two fruiting years. Additionally, variations were observed between the two AEZ, with the lowland AEZ experiencing the highest leaf infestation levels. However, no instances of very severe pest infestation levels of leaves were recorded in either AEZ (Table 3).
Means followed by a common letter in the same column for each factor (LUT, AEZ and year) are not significantly different at P < 0.05, Tukey’s HSD test; Means are followed by the Standard Error (SE) of the mean; LUT: land use types; HG: home garden; FL: farmland and RS: roadside; AEZ= Agroecological zones.
3.3. Morphological characterization and molecular identification
In the present study, mature adults of insects that were captured from Ziziphus fruits incubated in insect-rearing cages in the laboratory were categorized into ten different groups based on similarities and differences in morphological characters of wings, legs, head, body colour, antennae, thorax, and abdomen.
The BLAST search results with the DNA sequences obtained after sequencing the COI region of 20 insect specimens randomly selected from the ten morphologically separated groups revealed that, the insect groups collected from Ziziphus fruits in Ethiopia were related to six different insect species. Accordingly, based on the highest per cent similarity (≥ 99) of the query sequences with those of the GenBank sequences (Table 4), five of the 20 query sequences matched with C. incompleta and C. vesuviana, six with D. hydei, four with Z. indianus, three with Simulium spp., one with D. simulans and two with P. concolor. Phylogenetic analyses using the sequences obtained from our study and those of the highly matching GenBank sequences for each of the six taxa revealed that there seems to be more diversity of insects associated with Ziziphus fruits in Ethiopia than the results from the BLAST search mentioned above.
The molecular identification unveiled the presence of four fruit fly species, comprising one tephritidae species and three Drosophilidae species, as pests affecting Ziziphus fruits. The tephritid C. incompleta (Becker 1903) (Diptera: Tephritidae) was observed, alongside several drosophilids: D. hydei Sturtevant 1921; Z. indianus Gupta 1970; and D. simulans Sturtevant 1919 (all Diptera: Drosophilidae). In addition, the Braconidae species, P. concolor Szepligeti 1910 (Opiinae; Psyttalia), a parasitoid of C. incompleta (Becker 1903) and Simulium spp., (Diptera: Simuliidae) was identified during the current study. Given that Simulium spp. was found to be a non-insect pest of Ziziphus, it was excluded from further analysis. All insect pests were detected across the two AEZ and the three LUTs. Carpomya incompleta exhibited the highest occurrence across all three LUT, throughout the assessment months, and in both AEZ (Table 5). The count of emergence holes, indicative of adult insect activity, ranged from one to four per fruit, averaging about three holes per fruit. Interestingly, during the rearing phase of the adults, two distinct types of insect pests were observed emerging from a single fruit in certain samples. The extent of damage inflicted by each species was not determined, but the cumulative damage caused by the aforementioned species was assessed.
3.4. Phylogenetic characterization
Phylogenetic analyses using the neighbour-joining method for each taxon revealed further insights on whether the insects from our study were closely related to their respective matches from the GenBank. Accordingly, five sequences (ZFBO T01 022, ZFBO T04 022, ZFBA T06 022, ZFBO T17 022 and ZFBA T18 022) that closely matched with C. incompleta and C. vesuviana formed a distinct and strongly supported clade of their own with a bootstrap value of 100%. This clade was further divided into four branches that are also strongly supported by bootstrap values but negligible genetic distances. Similarly, those sequences from the GenBank claimed to be from C. incompleta and C. vesuviana formed a clade of their own which was further divided into distinct C. incompleta and C. vesuviana (Figure 3a).
In the phylogenetic analysis of six sequences (ZFBO T03 022, ZFBO T14 022, ZFBO T02 022, ZFBA T12 022, ZFBA T08 022 and ZFBO T09 022) with the highest sequence similarity with those of D. hydei, revealed that two of them (ZFBO T03 022, ZFBO T14 022) formed a strongly supported clade with those of D. hydei while the remaining four (ZFBO T02 022, ZFBA T12 022, ZFBA T08 022 and ZFBO T09 022) formed a separate and strongly supported clade on their own suggesting that those insects represented by these sequences were not exactly identical to D. hydei (Figure 3b). Likewise, the phylogenetic analysis of the sequence (ZFBO T13 022) that matched with the D. simulans revealed that it formed a strongly supported but small clade with only one of the D. simulans sequences while the rest of the D. simulans sequences formed a separate clade (Figure 3c). This indicates that the specimen from Ziziphus from Ethiopia is actually a D. simulans specimen.
Phylogenetic analysis of the sequences (ZFBO T05 022, ZFBO T10 022, ZFBO T11 022 and ZFBA T19 022) that matched with Zaprionus indianus in the BLAST search result showed that they formed a clade with sequences of Z. indianus although the statistical support for the branches was not strong. However, two of the sequences (ZFBO T05 022, ZFBO T10 022) formed a strongly supported sub-branch within the clade, while the other two sequences (ZFBO T11 022 and ZFBA T19 022) formed another sub-branch with a relatively weaker bootstrap support (Figure 3d).
The phylogenetic analysis of the fifth taxon represented by sequences (ZFBA T15 022 and ZFBO T16 022) that showed the highest sequence similarity match with those of P. concolor based on the BLAST search revealed that only one of the two (ZFBO T16 022) formed a strongly supported clade with P. concolor sequences while the second one formed a separate branch alone. Furthermore, the P. concolor sequences were also separated into two strongly supported clades (Figure 3e).
4. Discussion
The incidence of insect pests on Ziziphus fruits varied among the assessment months over two years. The percentage incidence of insect pests on Ziziphus fruits was above 77% across the AEZ and throughout the two fruit production years. The farmlands exhibited the highest percentage incidence during the 2022 fruiting year (89%), while the lowest incidence was observed in roadside areas (77%) during the 2023 fruit production year. Very severe infestations of >80%% were observed in the lowland AEZ, likely attributable to the availability of a suitable environment such as suitable temperature and humidity for insect pest reproduction [12,45]. These variations in incidences and levels of infestations across different agro-ecological zones can also be associated with variations in temperature, humidity, and rainfall, critical parameters for insect pest reproduction [46,47,48,49].
Insect pests contribute to yield reduction and post-harvest losses, consequently diminishing the quality and quantity of the fruit [50,51,52]. Such reductions in fruit quality can have adverse economic impacts on communities reliant on these fruits for their livelihood income [53]. Ensuring high fruit quality, such as size, shape, colour, and freedom from defects and decay, is crucial for fetching higher prices [54,55]. Additionally, desirable qualities like sweetness, softness, juiciness, thin skin, and well-developed flavour render fruits more susceptible to insect pest attacks [51]. In the present study, the majority of fruits were classified under low, medium, and severe infestation levels, with consistent patterns observed across production years and no significant variation among LUT. The incidence and infestation levels on Ziziphus fruits exhibited variability across assessment months. In both assessment years, the highest percentage of incidence occurred during November and December, while the lowest was recorded in September. The various assessment months were positively correlated with the fruit production years, while no correlation was observed between the different LUT, months of assessment, and AEZ.
In contrast, the occurrence of insect pests on Ziziphus leaves was low, under 50%. No instances of very severe pest infestation levels were recorded in the AEZ, LUT, production years, or assessment months. Despite the low percentage of incidence, significant variations were observed among the different LUTs, particularly on the roadside, which exhibited the highest percentage of infestation on Ziziphus leaves. Severe insect pest infestations on leaves could lead to complete defoliation, affecting growth and fruit production due to the cessation of photosynthesis [56-57. Additionally, infestation levels were generally low over the two years, with the majority of leaves showing no signs of infestation, indicating their overall health. The percentage incidence of insect pests on Ziziphus leaves remained below 50% throughout all assessment months over the two production years. The highest percentage incidence was observed in September of the 2022 production year, while the lowest occurred in October of the 2023 production year, which could be associated with climate variability between the two production years [10]. The low percentage incidence and infestation levels might be due to the resistance of the leaves to insect pests [58,59]. Molecular identification unveiled the presence of four fruit fly species, comprising one Tephritidae species and three Drosophilidae species, as pests affecting Ziziphus fruits.
The formation of a separate and strongly supported clade by the sequence from specimens collected from Ziziphus fruits in Ethiopia suggests that the insects may not be C. incompleta or C. vesuviana but may be a species that belongs to the same genus but not the same species. Furthermore, there seems to be some degree of genetic variation among the specimens collected from Ethiopia which could only be verified with further analysis using additional molecular markers. In the present study, the tephritid C. incompleta (Becker, 1903) (Diptera: Tephritidae) was found to be the most dominant insect pest. Carpomya incompleta is a monophagous pest of Ziziphus spp., causing significant negative impacts and contributing to producing low-yield and poor-quality fruits [60]. It has been recorded in several countries in Africa, Asia, the Middle East, and Europe [61,62]. Additionally, another tephritid fruit fly C. vesuviana Costa (Diptera: Tephritidae), which is also a monophagous pest of Zizyphus species, is a destructive pest contributing to low yield and poor quality of fruits [46,63,64]. Other tephritid fruit fly species, Bactrocera zonata and B. dorsalis (Diptera: Tephritidae), are common insects that frequently attack and severely damage Ziziphus fruits [57].
Alongside three drosophilids: D. hydei Sturtevant, 1921; Zaprionus indianus Gupta, 1970; and Drosophila simulans Sturtevant, 1919 (all Diptera: Drosophilidae), were also found to be impacting insect pests of Ziziphus fruits in Ethiopia. The phylogenetic analysis of the sequences identified as D. hydei from BLAST search indicated that the specimen from Ziziphus fruits in Ethiopia was probably not a single species but two different species, one of them being D. hydei since it formed a strongly supported branch along with D. hydei sequences while the other group appears to be distinct from D. hydei and maybe another Drosophila sp. In this study, the Ethiopia isolate ZFBO T13 022 was which demonstrated intraspecific similarity with D. simulans isolate KX161438.1:1-673 retrieved from GenBank, supported by a robust bootstrap value of 100%, thereby indicating a complete resemblance of the Ethiopian isolate to D. simulans. Drosophila hydei and D. simulans were reported as impacting insect pests on many fruits such as bananas, apples, melons, and berries (cherry, raspberry, blackberry, strawberry, and blueberry) with wide distribution ranges caused due to the transportation of contaminated fruits and the changing climates [65,66]. Several Drosopiliade lay their eggs in decaying fruits and might be secondary pests, yet some species also lay eggs on fresh fruits and their larvae feed on healthy fruit. Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) mainly infests healthy, unwounded thin-skinned fruits such as soft and stone fruits, cherries and apricots [67,68] and larval feeding and development on fruit flesh make fruits unmarketable [69].
The BLAST search result indicated that four sequences from the current study matched with those of Z. indianus from the GenBank database. Nevertheless, from the phylogenetic tree of the sequences from Ethiopia and those from the GenBank, it could be said that the specimens from Ethiopia do not appear to be Z. indianus but may be a related species since the bootstrap support for the branches was low. Given the existence of several species under the Genus Zaprionus and the fact that Z. indianus also known as fig fly, is a generalist that lives on a large number of host plants using their fruits for oviposition [70], the specimens from Ziziphus fruits in Ethiopia could be considered to belong to the Genus Zaprionus. Zaprionus indianus was reported first from Tunisian olives [71], and thereafter, was introduced to various countries across the globe and became pests of multiple fruits such as olive, berries, peach, apple and Ziziphus [72]. Zaprionus indianus mostly lays eggs in decaying fruit or fruit with injuries or mechanical damage yet it can also oviposit undamaged healthy fruit such as figs, strawberries and guavas [73].
Psyttalia concolor is known to be a parasitoid of the tephritid pests and therefore has been extensively used in biological control programmes against these pests. It is also known to be a member of a complex of closely related species from Africa that are difficult to separate apart based on morphological features alone [74]. The grouping of one of our sequences with one group of P. concolor sequences might indicate that the species represented by the sequence is actually P. concolor, while the separation of the second sequence into a separate branch on its own might indicate that the specimen represents one of the closely related members of the P.concolor species complex which we could not name based only on the data we currently have. Furthermore, the finding of the insect specimens that are already in use for biological control against fruit fly pests is interesting for further research on biological control of the tephritid pests affecting Zizyphus fruits in Ethiopia. Psyttalia concolor native to the Mediterranean are used as biological control of arthropod pests, and parasitize C. incompleta and the olive fruit fly (Bactrocera oleae) insects. They contribute a potential for agricultural pest controls [75].
Among the four fruit flies in the current study, Zaprionus indianus showed the lowest record compared to D. hydei, D. simulans and C. incompleta. The variation in the distribution of insect pests across various LUTs, agro-ecological zones, and different fruit production years might be related to various abiotic factors such as temperature, rainfall, and relative humidity, and biotic factors such as varietal resistances [13,68,76]. Drosophila hydei and D. simulans showed similar distributions among the three LUTs and the two AEZ. Zaprionus indianus showed the lowest record compared to D. hydei, D. simulans and C. incompleta. The variation might be related to various abiotic factors such as temperature, rainfall, and relative humidity [13,76]. Various insect pests have been reported from different countries for their detrimental impacts on Ziziphus fruit production, attacking the fruits at various stages of maturity [57,58,66].
5. Conclusions
Ziziphus tree species, which serve various purposes for communities including consumption, market sale, firewood, construction material, farm utensils, and fencing, face challenges in growth and production in Ethiopia due to both biotic and abiotic factors. Among biotic constraints, insect pests present significant challenges to Ziziphus trees, affecting flower, fruit and foliage production. The present study provides valuable insights into the frequency and severity of insects infesting Ziziphus tree fruits and leaves, which have significantly hampered fruit production, resulting in considerable yield reductions. Across the study sites, various damage symptoms were observed on both fruits and leaves over the span of two years. These distinct symptoms observed consistently throughout the assessment period, were attributed to different species of fruit fly insects. Higher incidences of pests and more severe infestations were observed on fruits harvested from farmlands, whereas comparatively lower occurrences were recorded on fruits obtained from roadsides over the two fruit production years. Conversely, leaves exhibited notably lower percentages of incidence and infestation levels compared to fruits. In lowland AEZ, higher percentages of incidence and infestation levels were observed. Analysis across different assessment months revealed peak percentages of incidence and infestation levels in November, with the lowest levels occurring in September across both LUT and AEZ. These assessment months emerged as significant influencers regarding the incidence of Ziziphus fruit insect pests. Moreover, this study identified the types of insect pests responsible for significant yield losses on Ziziphus fruits in Ethiopia. Both morphological (based on phenotypic characterizations) and molecular (based on DNA barcoding) techniques were employed to confirm the identification of specific insect pests. The study revealed that C. incomplete, D. hyde, D. simulans, and Z. indianus were the fruit fly insect pests impacting Ziziphus fruits in Ethiopia. The isolates from Ethiopia formed separate clades among themselves and between the sequences retrieved from the GenBank except for the Ethiopia isolate ZFBO T13 022 which demonstrated a complete resemblance and formed the same clade with D. simulans.
Overall, fruits exhibited significantly higher percentages of incidence and infestation levels compared to leaves, indicating substantial yield losses due to fruit fly insect pests within the study areas. The major fruit fly insect pests impacting Ziziphus fruits were C. incomplete, while the lowest distribution was observed in Z. indianus. Therefore, it is imperative to implement appropriate management strategies to mitigate significant yield losses and promote sustainable Ziziphus fruit production.
6. Patents
Supplementary Materials
The following supporting information can be downloaded at: www. https://submit.ncbi.nlm.nih.gov/subs/sra/SUB14806558/metadata_SUB14806558.
Author Contributions
“Conceptualization, methodology, formal analysis, data curation and writing—original draft preparation, Tigabu Redae; writing—review and editing, visualization, supervision, Samora M. Andrew, Miriam F. Karlsson and Abdella Gure. All authors have read and agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.”.
Funding
“This research received no external funding” or “This research was funded by the Swedish International Development Agency (SIDA) under the REFOREST (Regional Research School in Forest Sciences) program, Grant Number 13394”, which is gratefully acknowledged and “no funding was received for the APC”.
Data Availability Statement
Data will be available up on request.
Acknowledgments
We appreciate the Ethiopian Forestry Development Office, Addis Ababa, Ethiopia, for allowing us to use the laboratory to rear and preserve adults and prepare insect pests for DNA extraction. The International Livestock Research Institute in Nairobi, Kenya through the molecular biology laboratory assisted with conducting PCR amplification and DNA sequencing of the whole specimens during the molecular identification. Dr Wendu Admasu, Mr. Weldesenbet Beze and Mr. Dejen for their invaluable assistance with sample preparations, data recording, adult rearing and preservation, and morphological identifications as well as DNA extractions.
Conflicts of Interest
Declare conflicts of interest or state “The authors declare no conflicts of interest.” “The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Mongalo, N.I.; Mashele, S.S.; Makhafola, T.J. Ziziphus mucronata Willd. (Rhamnaceae): its botany, toxicity, phytochemistry and pharmacological activities. Heliyon, 2020, 6, e03708. [Google Scholar] [CrossRef] [PubMed]
- Leakey, R.R.B.; Tientcheu, A.M.L.; Awazi, N.P.; Assogbadjo, A.E.; Mabhaudhi, T.; Hendre, P.S.; Degrande, A.; Hlahla, S.; Manda, L. The future of food: domestication and commercialization of indigenous food crops in Africa over the third decade (2012-2021). Sustainability, 2022, 14, 2355. [Google Scholar] [CrossRef]
- Leakey, R.R.B.; Weber, J.C.; Page, T.; Cornelius, J.P.; Akinnifesi, F.K.; Roshetko, J.M.; Tchoundjeu, Z.; Jamnadass, R. Tree Domestication in Agroforestry: Progress in the Second Decade (2003–2012). In Advances in Agroforestry—The Future of Global Land Use; Springer: New York, NY, USA, 2012; pp. 145–173. [Google Scholar]
- Kalaba, F.K.; Chirwa, P.W.; Prozesky, H. The contribution of indigenous fruit trees in sustaining rural livelihoods and conservation of natural resources. African Journal of Wood Science and Forestry, 2019, 7, 1–6. [Google Scholar]
- Karuppaiah, V. Biology and management of ber fruit fly, Carpomyia vesuviana Costa (Diptera : Tephritidae): A review. African Journal of Agricultural Research, 2014, 9, 1310–1317. [Google Scholar]
- Yadav, J.; Gaur, R.K.; Kumar, Y. Evaluation of antixenotic and allelochemical traits of ber (Ziziphus mauritiana Lamk) fruits as a source of host plant resistance against fruit fly (Carpomyia vesuviana Costa) (Diptera : Tephritidae) in a semi-arid region of India. Phytoparasitica, 2020, 48, 607–620. [Google Scholar] [CrossRef]
- Dhaliwal, G.S.; Singh, R. Host Plant Resistance to Insects: Concepts and applications. New Delhi, Panama. Publication, 2014, pp:578.
- Ibrahim, A.S.; Samira, A.M.; Mahmoud, M.E.E.; Salih, A.I.S.; Saqib, A.; Ali, A. Monitoring of Tephritidae of Fruit Trees and Their Level of Infestation in South Kordofan State, Sudan. International Journal of Agriculture Institute. Canadian Journal of Plant Protection, 2014, 2, 1–7. [Google Scholar]
- Haldhar, S.M.; Deshwal, H.L.; Jat, G.C.; Berwal, M.K. Pest scenario of ber (Ziziphus mauritiana Lam. ) in arid regions of Rajasthan: a review. Journal of Agriculture and Ecology, 2016, 1, 10–21. [Google Scholar]
- Meghwal, P.R.; Kumar, P.; Singh, D. Climate variability during flowering and fruiting reduces fruit yield of ber (Ziziphus mauritiana ) in Western Rajasthan. Journal of Agriculture and Ecology, 2018, 6, 31–38. [Google Scholar] [CrossRef]
- Karuppaiah, V. Seasonality and management of stone weevil, Aubeus himalayanus Voss (Curculionidae : Coleoptera): An emerging pest in Indian Jujube (Ziziphus mauritiana L. ). Africa Journal of Agricultural Research, 2015, 10, 871–876. [Google Scholar]
- Sharma, V.P.; Lal, O.P.; Rohidas, S.B.; Pramanick, P.K. Varietal resistance in ber (Ziziphus mauritiana Lamk. ) against the fruit fly, Carpomyia vesuviana Costa (Diptera : Tephritidae) under the field conditions. Journal of Entomological Research, 1998, 22, 61–67. [Google Scholar]
- Karuppaiah, V.; More, T.A. , Sivalingam, P. N.; Hanif, K.; Bagle, B.G. Prevailing insect pests of ber (Zizipus mauritiana Lamk) and their natural enemies in a hot arid ecosystem. Haryana Journal of Horticultural Sciences, 2010, 39, 214–216. [Google Scholar]
- Singh, M.P. Managing menace of insect pests on ber. Indian horticulture, 2018, 53, 31–32. [Google Scholar]
- Gaur, R.K.; Kumar, M.; Sharma, S.; Singh, B. Survey studies on insects and non-insect pests associated with ber crop in South West Haryana. Journal of Entomology and Zoology Studies, 2020, 8, 856–863. [Google Scholar]
- Girma N, Zinabu N, Mubarek E. Assessment of Diseases and Insect Pests on Ziziphus Species Fruit in Waghimra Administrative Zone, Amhara National Regional State, Ethiopia. In Editors, Menale Wondie, Abraham Abiyu and Mulugeta Alemayehu (2017) Proceedings of the 10th and 11th Annual Regional Conference on Completed Research Activities of Forestry, March 6-13, Amhara Agricultural Research Institute (ARARI), Bahir Dar, Ethiopia, 2017.
- Bagle, B. Incidence and control of fruitfly (Carpomyia vesuviana Costa) of ber (Ziziphus mauritiana Lamk. ). Indian Journal of Plant Protection, 1992, 20, 205–207. [Google Scholar]
- Sonawane, B.R. Study of pest of tropical ber with special reference to fruit borer Meridarchis scyrodes Meyr. MSc Thesis, Poona University, Poona.
- Mayhew, P.J. Explaining global insect species richness: lessons from a decade of macro-evolutionary entomology. Entomologia experimentalists et applicate, 2018, 166, 225–250. [Google Scholar] [CrossRef]
- Whiting, D. Key to insect orders. CMG GardenNotes. 2017, pp:314–316.
- Tahir, H.; Noor, M.; Mehmood, A.; Sherawat, S.M.; Qazi, M.A. Evaluating the accuracy of morphological identification of insect pests of rice crops using DNA barcoding. Mitochondrial DNA Part B, 2018, 3, 1220–1224. [Google Scholar] [CrossRef]
- Smith, M.A. , Eveleigh, E. S.; Mccann, K.S. Barcoding a quantified food web: crypsis, concepts, ecology and hypotheses. PLoS One, 2011, 6, e14424. [Google Scholar]
- Wallace, L.J.; Boilard, S.M.; Eagle, S.H. DNA barcodes for everyday life: routine authentication of natural health products, Food Research International, 2012, 49, 446–452.
- Bihon, W.; Burgess, T.; Slippers, B.; Wingfield, M.J.; Wingfield, B.D. Distribution of Diplodia pinea and its genotypic diversity within asymptomatic Pinus patula trees. Australasian Plant Pathology, 2011, 40, 540–548. [Google Scholar] [CrossRef]
- Chacón, I.A.; Janzen, D.H.; Hallwachs, W. Cryptic species within cryptic moths: new species of Dunama Schaus (Notodontidae, Nystaleinae) in Costa Rica. ZooKeys, 2013, 264, 11–45. [Google Scholar] [CrossRef]
- NMSA. Addis Ababa National Meteorology Service Agency (NMSA),2020, Addis Ababa, Ethiopia.
- FAO (Food and Agriculture Organization of the United Nations), success stories. climate-smart agriculture (CSA) on the ground understanding, 2015.
- Madden, L.V.; Hughes, G.; van den Bosch, F. The Study of Plant Disease Epidemics APS Press, St. Paul, MN, 2007.
- Nagrare, V.S.; Kranthi, S.; Biradar, V.K.; Zade, N.N.; Sangode, V.; Kakde, G.; Shukla, R.M.; Shivare, D.; Khadi, B.M.; Kranthi, K.R. Widespread infestation of the exotic mealybug species Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) on cotton in India. Bull. Entomology Research, 2011, 99, 537–541. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Analysis of phylogenetic relationships by amplification and direct sequencing of ribosomal genes. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, Editors. PCR Protocols; Academic Press; New York, 1990.
- Costa, F.O.; Dewaard, J.R.; Boutillier, J. Biological identifications through DNA barcodes: the case of the Crustacea. Canadian Journal of Fishery and Aquatic Sciences, 2007, 62, 272–295.
- Saiki, R.K.; Gelfand, D.H.; Stoffel, S.; Scharf, S.J.; Higuchi, R.; Horn, G.T.; Mullis, K.B.; Erlich, H.A. Primer directed enzymatic ampliÞcation of DNA with a thermostable DNA polymerase. Science, 1988, 239, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Marion, C.; Jade, L.; Laury-Ann, D.; Ariane, T.; Grégoire, C.; Cloé, L.; Eric, P.; Geneviève, J.; Parent. Optimized QIAGEN DNeasy Blood & Tissue kit Protocol for Environmental DNA Extraction. 2023, protocols.io. [CrossRef]
- Borghuis, A.; Pinto, J.D.; Platner, G.R.; Stouthamer, R. Partial cytochrome oxidase II sequences distinguish the sibling species Trichograma minutum Riley and T. platneri Nagarkatti. Biol. Con.. 2004, 30, 90–94. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular cloning: a laboratory manual (2nd Ed). Cold Spring Harbor, NY. Cold Spring Harbor Laboratory Press, Selection of the Formosan subterranean termite. J. Beh.Eco., 1989, 19, 764–773. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Marine Bio. and Biot., 1994, 3, 294–299. [Google Scholar]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proceedings of the National Academy of Sciences, 1977, 74, 5413–5467. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: a user-friendly biological sequence alignment [ed. ], and analysis program for Windows 95/98/NT. Nucleic Acids Symposiums Serous, 1999, 41, 95–98. [Google Scholar]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: architecture and applications. BMC Bioinformatics, 2009, 10, 421. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics. Oxford University Press, New York.
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Molecular Biology and Evolution, 2011. [CrossRef]
- Team, R.C. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2023, URL https://www.R-project.org/.
- Kavitha Z, Savithri P (2002) New record of some natural enemies on ber pests in Tirupati Region. South Indian Horti., 2002, 50(4-6):513-514.
- Lakra, R.K.; Singh, Z. Seasonal fluctuations in the incidence of ber fruitfly Carpomyia vesuviana Costa (Diptera: Tephritidae) under agroclimatic conditions of Hisar. Haryana Agricultural University J. Res., 1985, 15, 42–50. [Google Scholar]
- Coissac, E.; Hollingsworth, P.M.; Lavergne, S. From barcodes to genomes: extending the concept of DNA barcoding, Molecular Ecology, 2016, 25, 1423–1428.
- Nandihalli, B.S.; Patil, D.R.; Jagginavar, S.B.; Biradar, A.P.; Guled, M.B.; Surkod, V.S. Incidence of fruit borer (Meridarchis scyrodes Meyr.), and fruit fly (Carpomyia vesuviana Costa) on different varieties of ber. Advances in Agri. Res. in India, 1996, 6, 13–18. [Google Scholar]
- Gopali, J.B.; Sharanabasappa Yelshetty, S. Incidence of ber fruit borer, Meridarchi scyrodes Meyrick (Lepidoptera: Carposinidae) in relation to weather parameters. Insect Envi., 2003, 9, 165–166. [Google Scholar]
- Grewal, J.S.; Kapoor. Relative incidence of infestation by two species of fruit flies Carpomyia vesuviana and Dacuszonatus (Diptera: Tephritidae) on ber in the Punjab. Indian J. of Eco., 1986, 8, 123–125. [Google Scholar]
- Farrar, N.; Golestaneh, R.; Askari, H.; Assareh, M.H. Studies on parasitism of Fopius carpomyie (Silvestri) (Hymenoptera: Braconidae), an egg-pupal parasitoid of ber fruit fly, Carpomyia vesuviana Costa (Diptera: Tephritidae). Bushehr-Iranian Acta Horti 2009, 840, 431–438. [Google Scholar] [CrossRef]
- Sander, N.L.; da Silva, C.J.; Duarte, A.V.M.; Zago, B.W.; Galbiati, C. The Influence of Environmental Features on the Morphological Variation in Mauritia flexuosa L.f. Fruits and Seeds. Plants, 2020, 9, 1–10. [Google Scholar] [CrossRef]
- Arora, P.K.; Kaur, N.; Thind, S.K.; Aulakh, P.S. Screening of some ber varieties for resistance against fruit fly. Horti. J., 2001, 14, 117–120. [Google Scholar]
- Mitra, S.K.; Dhaliwal, S.S. Effect of potassium on fruit quality and their storage life. Proceedings IPI-OUAT-IPNI International symposium, 2009.
- Karar, H.; Bashir, M.A.; Khan, K.A.; Farooq, A.B.; Aziz, I.; Ali, H.; Ghramh, H.A.; Abbas, G.; Alghanem, S.M. Response of leading ber (Zizyphus jujuba) varieties against fruit flies (Tephritidae : Diptera) and estimation of their losses. Fre. Envi. Bulletin, 2020, 29, 10311–10319. [Google Scholar]
- Muhammad S (2006) Incidence of Insect Pests on Ber (Zizyphus jujube) Tree. Journal of Zoology, 38, 261-263.
- Sarwar, M. Incidence of insect pests on Ber (Zizyphus jujube) tree. Pakistan J. of Zoo., 2006, 38, 261–263. [Google Scholar]
- Tembo, L.; Chiteka, Z.; Kadzere, I.; Akinnifesi, F.K.; Tagwira, F. Ripening stage and drying method affecting colour and quality attributes of Ziziphus mauritiana fruits in Zimbabwe. Afri.. J. of Biotec., 2008, 7, 2509–2513. [Google Scholar]
- Nizamani IA, Rustamani MA, Nizamani SM, Nizamani SA, Khaskheli MI (2015) Population Density of Foliage Insect Pest on Jujube, Ziziphus mauritiana Lam. Ecosystem. Journal of Basic and Applied Sciences, 11, 304–313.
- Al-Masudey, A.D.; Al-Yousuf, A.A. Effect of jujube fruit cultivars on chemical control of jujabe fruit fly Carpomyia incompleta. Kufa J. of Agri. Sci., 2013, 5, 111–124. [Google Scholar]
- Hoddle, M.S.; Mound, L.A. The genus Scirtothrips in Australia (Thysanoptera, Thripidae). Zootaxa, 2003, 268, 1–40. [Google Scholar] [CrossRef]
- Korneyev, V.A.; Mishustin, R.I.; Korneyev, S.V. The Carpomyini fruit flies (Diptera: Tephritidae) of Europe, Caucasus, and Middle East: new records of pests, with improved keys. Vestn Zoo., 2017, 51, 453–470. [Google Scholar] [CrossRef]
- CABI (2022) Invasive species compendium. Available online: https://www. cabi.org/isc/datasheet/11408.
- Zavitha Z, Savithri P, Vijayaragavan C (2002) Insect pests of ber, Ziziphusjujuba in Tirupati Region. Ins. Envi., 2002, 7, 157–158.
- Balikai, R.A.; Kotikal, Y.K.; Prasanna, P.M. Global scenario of insect and non-insect pests of jujube and their management options. Acta Horti., 2013, 993, 253–277. [Google Scholar] [CrossRef]
- Inmaculada, G.J.; Enrique, Q.M.; Meelad, Y.Y. Zizyphus fruit fly (Carpomya incompleta (Becker), Diptera: Tephritidae) is expanding its range in Europe. Spa. J. of Agri. Res., 2022, 20 (4): e10SC02.
- Cini, A.; Ioriatti, C.; Anfora, A. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bulletin of Insectology, 2012, 65, 149–160. [Google Scholar]
- Sasaki, M.; Sato, R. Bionomics of the cherry drosophila, Drosophila suzukii Matsumura (Diptera: Drosophilidae) in Futeushima prefecture. Annual Report of the Society of Plant Protection of North Japan, 1995, 46, 164–172. [Google Scholar]
- Rota-Stabelli O, Blaxter M, Anfora G (2013) Drosophila suzukii. Current Biology, 23, R8–R9.
- Commar, L.S.; Conceição, L.G.; Carlos, C.R.; Claudia, M.A.C. Taxonomic and evolutionary analysis of Zaprionus indianus and its colonization of Palearctic and Neotropical regions, Gen. and Mol. Bio., 2012, 35, 395–406. [Google Scholar]
- Szépligeti, G. In: Marchal P. (Ed.), Sur un Braconide [Hym.] nouveau, parasite du Dacus oleae. Bulletin de la Société entomologique de France, 1910, 13, 243–244.
- Daniell, R.R.F.; Elton, L.A. Occurrence of Zaprionus indianus Gupta (Diptera: Drosophilidae) In “Juazeiro” Fruits Ziziphus Joazeiro Mart. (Rhamnaceae). In The State of Rio Grande Do Norte, Brazil. Revista Brasileira de Fruticultura, Jaboticabal-SP, 2011, 33, 1356–1358. [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health), Bragard, C.; Baptista, P.; Chatzivassiliou, E.; Di Serio, F.; Gonthier, P.; Jaques, M.J.A.; Justesen, A.F.; Magnusson, C.S.; Milonas, P.; Navas-Cortes, J.A.; Parnell, S.; Potting, R.; Reignault, P.L.; Stefani, E.; Thulke, H.H.; Van der Werf, W.; Vicent Civera, A.; Yuen, J.; Zappala, L.; Gregoire, J.C.; Malumphy, C.; Kertesz, V.; Maiorano, A.; MacLeod, A. Pest categorisation of Zaprionus indianus. EFSA, 2022, 20, 7144. [CrossRef]
- Brunner, P.C.; Flemming, C.; Frey, J.E. A molecular identification key for economically important thrips species (Thysanoptera: Thripidae) using direct sequencing and a PCR-RFLP-based approach. Agri. and For.Ent., 2002, 4, 127–136. [Google Scholar] [CrossRef]
- Rugman-Jones, P.F.; Robert, W.; Tom van, N.; Richard, S. Molecular differentiation of the Psyttalia concolor (Szépligeti) species complex (Hymenoptera: Braconidae) associated with olive fly, Bactrocera oleae (Rossi) (Diptera: Tephritidae), in Africa. Bi. Con., 2009, 49, 17–26. [Google Scholar] [CrossRef]
- Amiri, A.; Talebi, A.A.; Zamani, A.A.; Kamali, K. Effect of temperature on demographic parameters of the hawthorn red midget moth, Phyllonorycter corylifoliella, on apple. J. Ins. Sci., 2010, 10, 134. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Incidence of insect pests on Ziziphus fruits across the assessment months in Ethiopia.
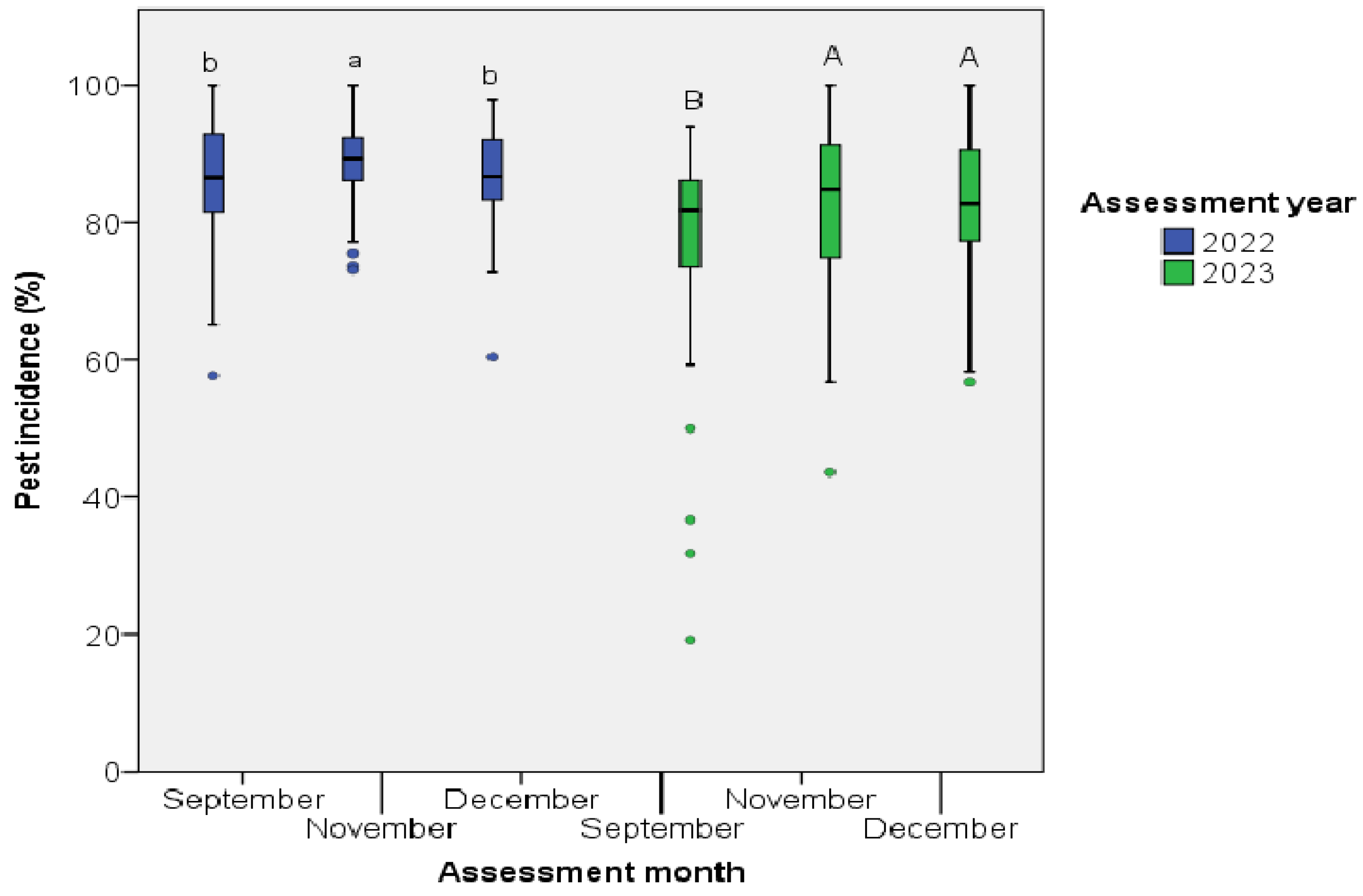
Figure 3.
Phylogenetic tree drawn according to the neighbor-joining method based on barcode sequences from specimens; (a) C. incompleta, (b) D. hydei, (c) D. simulans, (d) Z. indianus and (e) P. concolor in Ethiopia.
Figure 3.
Phylogenetic tree drawn according to the neighbor-joining method based on barcode sequences from specimens; (a) C. incompleta, (b) D. hydei, (c) D. simulans, (d) Z. indianus and (e) P. concolor in Ethiopia.
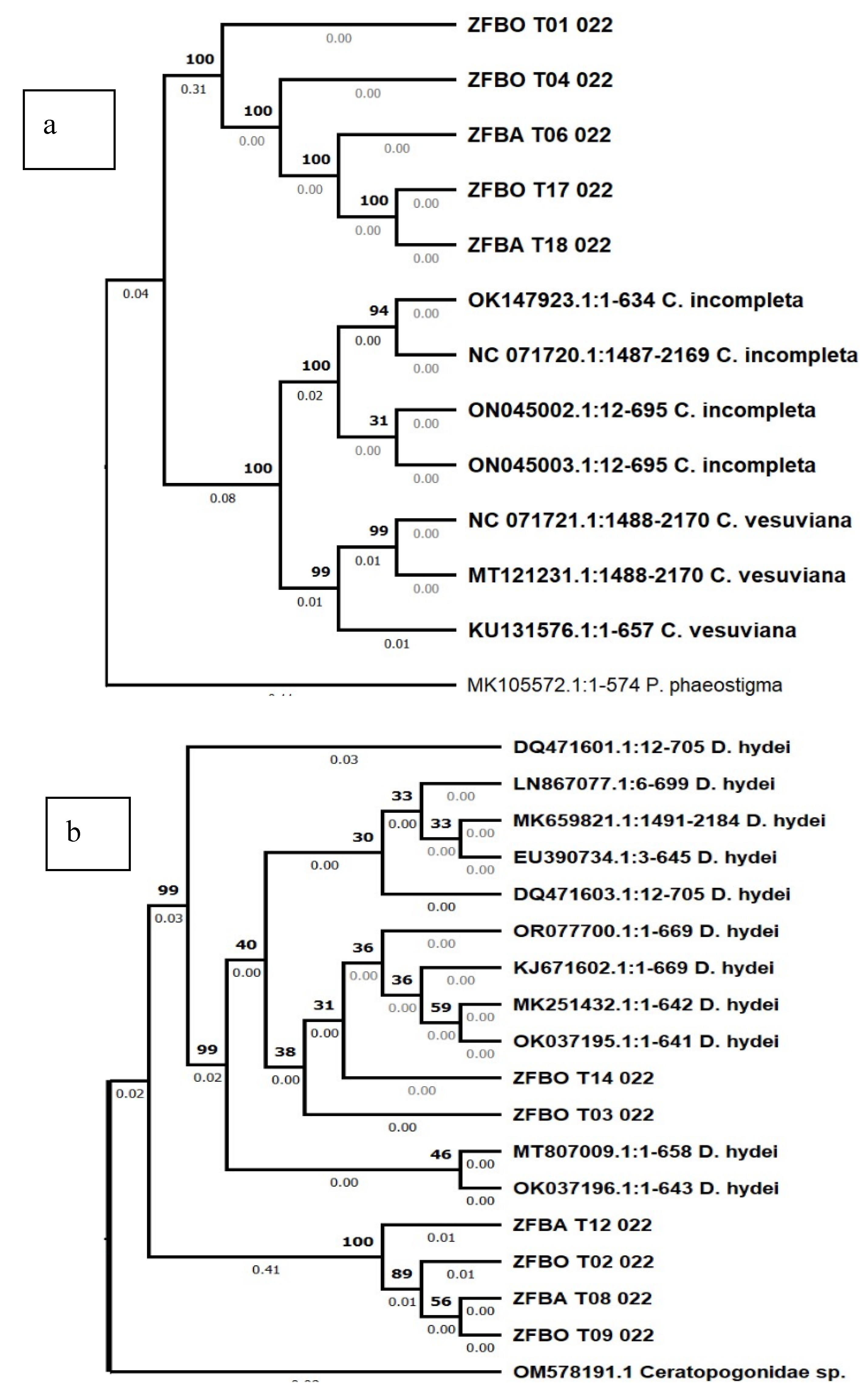
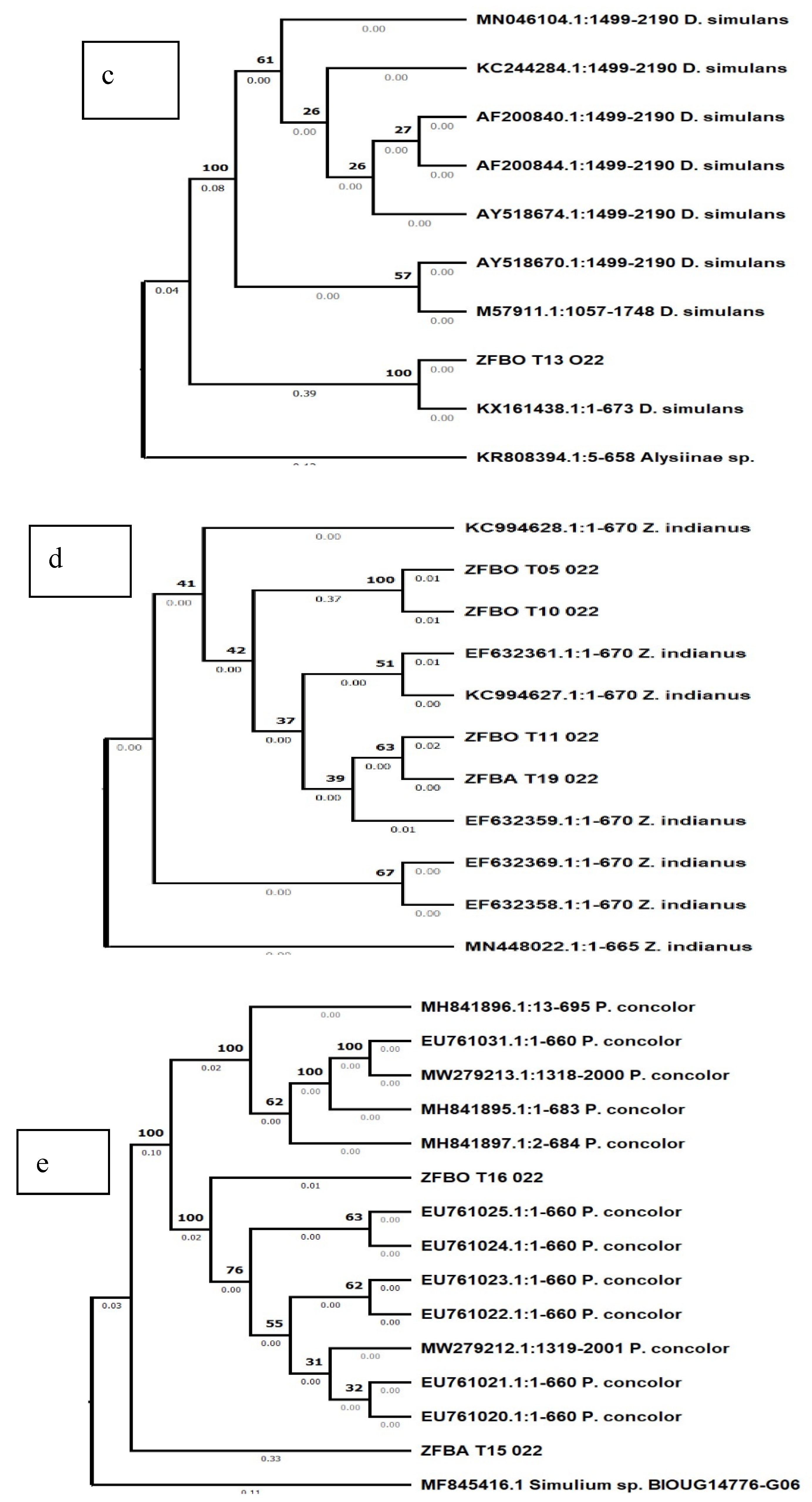

Table 1.
Factors influencing the insect pest incidences of Ziziphus fruits in Ethiopia.
| Fruits | |||||
|---|---|---|---|---|---|
| Variables | Coefficients (B) | S.E | Exponential of B | t-value | P-value |
| Agro-ecological zone | -2.3 | 2.23 | -0.11 | -1.03 | 0.303 |
| Land use types | -0.26 | 0.65 | 0.04 | 0.40 | 0.687 |
| Assessment year | -10.51 | 1.98 | -0.50 | -5.31 | 0.001*** |
| Adjusted R square | 0.096 | ||||
| Standard error | 10.11 | ||||
| p > F | 10.55 | ||||
| Leaves | |||||
| Agro-ecological zone | -2.54 | 2.78 | -0.09 | -0.92 | 0.36 |
| Land use types | 0.60 | 0.81 | 0.07 | 0.74 | 0.46 |
| Assessment months | 0.17 | 0.11 | 0.15 | 1.57 | 0.12 |
| Adjusted R square | 0.069 | ||||
| Standard error | 13.45 | ||||
| P > F | 7..64 | ||||
*- Significant at P < 0.05; CI = confidence interval; B= slope coefficient; SE = Standard Error.
Table 2.
Pearson correlation coefficients between determinant factors of insect pest incidence on Ziziphus fruits in Ethiopia.
Table 2.
Pearson correlation coefficients between determinant factors of insect pest incidence on Ziziphus fruits in Ethiopia.
| Fruits | ||||
|---|---|---|---|---|
| Determinant factors | Insect pest incidence | AEZ | Land use types | Assessment year |
| AEZ | 0.071 | |||
| Land use types | 0.053 | 0.88* | ||
| Assessment year | 0.289 | 0.001 | 0.00 | |
| Assessment months | 0.173 | 0.001 | 0.00 | 0.84* |
| Leaves | ||||
| AEZ | 0.02 | |||
| Land use types | 0.01 | 0.86* | ||
| Assessment year | 0.27 | -0.02 | ||
| Assessment months | 0.18 | 0.004 | 0.004 | 0.85* |
Table 3.
Incidence and infestation level of insect pests on Ziziphus fruits in three land use types in Ethiopia.
Table 3.
Incidence and infestation level of insect pests on Ziziphus fruits in three land use types in Ethiopia.
| Fruits | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| LUT | Mean (± SE) fruits examined/ tree | Mean infestation level per tree | ||||||||
| Very low | Low | Medium | Severe | Very severe | ||||||
| Farmland | 58±1.4 | 6±0.4c | 20.8±0.8 | 21±0.9 | 9 ±0.7 | 5.3±0.5 | ||||
| home garden | 56.9±1.1 | 7.1±0.5b | 19.9±0.7 | 20.7±0.9 | 8.6±0.5 | 5±0.5 | ||||
| Roadside | 59.4±1.2 | 8±0.5a | 19.7±0.7 | 21.7±0.7 | 9±0.6 | 5.1±0.6 | ||||
| AEZ | ||||||||||
| Lowland | 59±0.9 | 9.8±0.5 | 20.4±0.5 | 21.6±0.6 | 8.2±0.4a | 5.9±0.4b | ||||
| Midland | 57.1±0.8 | 9.0±0.5 | 19.8±0.5 | 20.6±0.5 | 9.3±0.4b | 4.2±0.2a | ||||
| Year | ||||||||||
| 2022 | 55.6±0.6 | 7.1±0.3b | 20.1±0.4 | 20.8±0.4 | 7.9±0.3b | 4.9±0.3 | ||||
| 2023 | 60.6±1 | 11.7±0.6a | 20.2±0.6 | 21.4±0.6 | 9.8±0.5a | 5±0.3 | ||||
| Leaves | ||||||||||
| LUT | Mean (± SE) number of leaves examined/ tree | Mean infestation level per tree | ||||||||
| Nil | Low | Medium | Severe | |||||||
| Farmland | 52.6±1.1 | 34.5±1.1 | 13.1±0.5 | 6.6±0.4 | 4.5±0.4 | |||||
| home garden | 49.2±1.1 | 32.7±1.1 | 12.5±0.4 | 6.5±0.4 | 3.7±0.4 | |||||
| Roadside | 53±1.2 | 33.8±1.2 | 13.4±0.5 | 7.3±0.5 | 4.4±0.2 | |||||
| AEZ | ||||||||||
| Lowland | 51.1±0.8 | 32.9±0.8a | 12.8±0.4 | 7.3±0.3b | 4.2±0.3 | |||||
| Midland | 52.7±1.2 | 35.2±1.1b | 13.6±0.4 | 6±0.4a | 4.1±0.3 | |||||
| Year | ||||||||||
| 2022 | 50±0.8 | 30.5±0.6a | 14.2±0.4b | 7.7±0.4b | 4.1±0.3 | |||||
| 2023 | 53.2±1.1 | 36.9±1.1b | 11.8±0.4a | 6.0±0.4a | 4.2±0.2 | |||||
Table 4.
Fruit fly isolates used in the insect identification study in Ethiopia.
| Species Name | Isolate number | host | Origin | Collector | Per cent identity | Accession |
|---|---|---|---|---|---|---|
| C. incompleta | AHL2 | Jujube | Iraq | Tahir,H.M. | 99.85 | ON045003 |
| C. incompleta | AHL1 | Jujube | Iraq | Tahir,H.M. | 99.85 | ON045002 |
| C. incompleta | Italy 01 | Jujube | Italy | Zhang Y. | 99.71 | NC_071720 |
| C. vesuviana | I1 | Jujube | Spain | Garrido JI. | 99.68 | OK147923 |
| C. vesuviana | China, Xinjiang 01 | Jujube | China | Zhang,Y. | 95.31 | MT121231 |
| C. vesuviana | Iran 01 | Jujube | Iran | Zhang,Y. | 95.31 | NC071721 |
| C. vesuviana | FUN12 | Jujube | China | Jing,L. | 95.43 | KU131576 |
| C. vesuviana | ZFBO T01 022 | Ziziphus | Ethiopia | Tigabu R. | QU5908887 | |
| C. vesuviana | ZFBO T04 022 | Ziziphus | Ethiopia | Tigabu R. | QU8343087 | |
| C. vesuviana | ZFBO T17 022 | Ziziphus | Ethiopia | Tigabu R. | QU1834357 | |
| C. vesuviana | ZFBA T06 022 | Ziziphus | Ethiopia | Tigabu R. | QU6288145 | |
| C. vesuviana | ZFBA T18 022 | Ziziphus | Ethiopia | Tigabu R. | QU1912175 | |
| D. hydei | CRX36794.1 | Melon | Italy | Patrizia T. | 99.43 | LN867077 |
| D. hydei | DHYDE20161106 | Berry | China | Qian Z.Q. | 99.14 | MK659821 |
| D. hydei | Africa | Berry | China | Wang B.C | 98.85 | DQ471603 |
| D. hydei | CH55 | Melon | Iran | Oshaghi,MA | 99.7 | OR077700 |
| D. hydei | DQ37 | Melon | New Zealand | Simon Hodge | 99.55 | KJ671602 |
| D. hydei | Duke.Bio203L | Berry | USA | Eric Spana | 99.39 | MT807009 |
| D. hydei | TEN104-102 | Melon | Spain | Vilchez R.I. | 99.84 | OK037195 |
| D. hydei | QDE57910.1 | Melon | South Africa | Liana I. A. | 99.53 | MK251432 |
| D. hydei | ABH5 | Melon | Spain | Vilchez R.I. | 99.38 | OK037196 |
| D. hydei | 15085-1641.58 | Melon | Spain | Evans A.L. | 99.38 | EU390734 |
| D. hydei | AQ49 | Berry | China | Wang B.C. | 93.97 | DQ471601 |
| D. hydei | ZFBO T03 022 | Ziziphus | Ethiopia | Tigabu R. | QU3000047 | |
| D. hydei | ZFBO T02 022 | Ziziphus | Ethiopia | Tigabu R. | QU2682351 | |
| D. hydei | ZFBO T09 022 | Ziziphus | Ethiopia | Tigabu R. | QU3295235 | |
| D. hydei | ZFBO T14 022 | Ziziphus | Ethiopia | Tigabu R. | QU7962979 | |
| D. hydei | ZFBA T08 022 | Ziziphus | Ethiopia | Tigabu R. | QU7664841 | |
| D. hydei | ZFBA T12 022 | Ziziphus | Ethiopia | Tigabu R. | QU7906397 | |
| D. simulans | UKG21278.1 | Melon | China | Li,T. | 99.86 | MN046104 |
| D. simulans | sm21 | peach | Brazil | Montooth KL. | 99.86 | KC244283 |
| D. simulans | AU023 | Ziziphus | Kenya | Ballard J.W. | 99.86 | AY518674 |
| D. simulans | Sc00 | melon | Seychelles | Ballard J.W. | 99.86 | AF200844 |
| D. simulans | DSR | Apple | Madagascar | Ballard J.W. | 99.86 | AF200841 |
| D. simulans | DSW | Apple | USA | Ballard J.W. | 99.86 | AF200840 |
| D. simulans | C167 | Banana | Kenya | Ballard J.W. | 99.86 | AF200839 |
| D. simulans | KY215 | Banana | Kenya | Ballard J.W. | 99.71 | AY518672 |
| D. simulans | KY007 | Apple | USA | Ballard J.W. | 99.71 | AY518670 |
| D. simulans | SL3 | Melon | Spain | Satta,Y. | 99.71 | M57911.1 |
| D. simulans | simw501 | Apple | Brazil | Montooth KL. | KC244284 | |
| D. simulans | ZFBO T13 022 | Ziziphus | Ethiopia | Tigabu R. | QU2055377 | |
| Z. indianus | haplotype 6 | Fig, | Brazil | Mendonca MP | 98.96 | KC994628 |
| Z. indianus | Duke.Bio203L | Fig | USA | Mohamed N. | 99.1 | MN448022 |
| Z. indianus | haplotype 5 | Fig | DRC | Mendonca MP | 98.81 | KC994627 |
| Z. indianus | ABR08559.1 | Fig | Brazil | Amir,Y. | 98.81 | EF632369 |
| Z. indianus | ABR08548.1 | Fig | Brazil | Amir,Y. | 98.66 | EF632358 |
| Z. indianus | ABR08551.1 | Fig | Madagascar | Amir,Y. | 98.51 | EF632361 |
| Z. indianus | ABR08549.1 | Fig | Madeira | Amir,Y. | 98.51 | EF632359 |
| Z. indianus | ZFBO T05 022 | Ziziphus | Ethiopia | Tigabu R. | QU2155377 | |
| Z. indianus | ZFBO T10 022 | Ziziphus | Ethiopia | Tigabu R. | QU2212817 | |
| Z. indianus | ZFBO T11 022 | Ziziphus | Ethiopia | Tigabu R. | QU2280917 | |
| Z. indianus | ZFBA T19 022 | Ziziphus | Ethiopia | Tigabu R. | QU2348287 | |
| P. concolor | PRJ076 | Ziziphus | Morocco | Rugman-JP.F. | 99.09 | EU761024 |
| P. concolor | TN0216 | Ziziphus | Italy | Rugman-JP.F. | 99.09 | EU761022 |
| P. concolor | TN0222 | Ziziphus | USA | Rugman-JP.F. | 99.09 | EU761021 |
| P. concolor | TN0227 | Ziziphus | Morocco | Rugman JP.F. | 98.94 | EU761025 |
| P. concolor | TN0223 | Ziziphus | Italy | Rugman-JP.F. | 98.94 | EU761023 |
| P. humilis | Ps29 | Ziziphus | South Africa | Barbara VA. | 95.61 | MH841897 |
| P. humilis | Ps24 | Ziziphus | South Africa | Barbara VA. | 95.61 | MH841896 |
| P. humilis | Ps25 | Ziziphus | South Africa | Barbara VA. | 95.61 | MH841895 |
| P. humilis | QTC30726.1 | Ziziphus | Portugal | Powell,C. | 95.61 | MW279213 |
| P. humilis | TN0220 | Ziziphus | South Africa | Rugman-JP.F. | 95.45 | EU761031 |
| P. humilis | TN0223 | Ziziphus | Namibia | Rugman-JP.F. | 95 | EU761030 |
| P. humilis | ZFBA T15 022 | Ziziphus | Ethiopia | Tigabu R. | QU2497311 | |
| P. humilis | ZFBO T16 022 | Ziziphus | Ethiopia | Tigabu R. | QU2608411 |
Table 5.
Species of insect pests recorded across LUT, assessment months and AEZ in Ethiopia.
| Type of insect pest | Percentage of insect pests recorded across the different | |||||||
| Land use types | Assessment Months | AEZ | ||||||
| Farm land |
Home garden | Roadsides | September | October | November | Low land |
Mid land |
|
| C. incompleta | 51.4 | 47.8 | 39.0 | 43.6 | 41.2 | 36.3 | 48.3 | 42.8 |
| D. hydei | 18.6 | 23.9 | 22.9 | 15.7 | 15.9 | 17.0 | 22.9 | 20.2 |
| D. simulans | 19.9 | 17.6 | 31.1 | 31.6 | 33.0 | 31.0 | 21.8 | 27.4 |
| Z. indianus | 6.3 | 8.8 | 4.6 | 6.7 | 7.4 | 10.8 | 5.6 | 6.0 |
| P. concolor | 3.7 | 1.9 | 2.5 | 2.4 | 2.5 | 5.0 | 1.3 | 3.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



