
Submitted:
25 October 2024
Posted:
28 October 2024
You are already at the latest version
Abstract
Given the pivotal role played by the microbiome in skin health it is important to understand how its composition varies with age, gender and body site, and regular use of topical products. Four studies have been carried out to determine the effects of long term (4 week) use of different dexpanthenol containing topical products on the skin microbiome of a varied population with cosmetically dry skin. Skin microbiome composition was assessed before and after product usage. No significant changes in microbiome richness or diversity were found for the individual test products, however meta-analysis of the combined dataset did show changes in microbiome composition as a function of subject sex, age and body site. The work presented here demonstrates how the use of carefully formulated topical products on skin, when used in a way which is representative of real life usage conditions, can respect the microbial diversity present on skin across a widely varied study population.
Keywords:
Microbiome
; Topical products
; Skin health.
; dry skin
; dexpanthenol
; repair
; xerosis
1. Introduction
The microbiome forms an integral part of skin, and can impact its form and function [1,2]. Far from being a static and unchanging part of the skin, the microbiome varies over time as a result of key life stages such as puberty and menopause [3]. It is important that the use of topical skin care products does not negatively impact the microbiome composition, and respects the natural balance present at this different life stages as microbiome dysbiosis is linked with a range of skin conditions such as Xerosis, Atopic Dermatitis and Psoriasis [4,5,6,7,8]. Recently, four human studies were carried out to examine the skin microbiome composition, and the effects of using topical cosmetic products containing dexpanthenol along with ingredients such as niacinamide and lipids which are included with the aim of improving xerotic skin [9]. Three of these studies examined topical moisturizing products, and one looked at products designed to cleanse the skin. While some preliminary results have been shared with an emphasis on the behavior of the wash products [10], meta-analysis has now been carried out on the results of the studies to enable a deep dive into the microbiome of the study population and the effects of the products on it. This article discusses the results of these studies and this meta-analysis and how the skin microbiome varies with sex, age and body location and usage of a range of topical cosmetic products, as well as the roles that these types of products have in helping to protect the skin.
2. Materials and Methods
Four studies to examine the effects of topical moisturizing products on skin microbiome composition were run between January and June 2023 in Hamburg, Germany. Studies were conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board of proderm GmbH (IRB code 2022/028, 15th January 2023, IRB code 2022/029 15th January 2023, IRB code 2023/001 11th January 2023, IRB code 2023/002 11th January 2023). The subjects gave consent to take part in writing in each of the four studies. They also consented that their (pseudo-anonymized) data can be used for further scientific analyses. Key inclusion criteria were; female or male, aged between 18 to 65 years of age and having cosmetically dry skin (according to Corneometer® CM825 values < 37 a.u. (arbitrary units) on one volar forearm at screening). The population demographics across the four studies are given in Table 1.
Each study was based on microbiome samples collected from the participants to test four different treatments (2 test products in a split-design each vs. untreated). Product identifying codes are given in Table 2. All products contained dexpanthenol and a range of other ingredients, the compositions of which are listed in Supplementary Information, section S.1. Two sites were on the face, and two on the volar forearm. Prior to the start of the study the subjects were instructed not to apply any leave on cosmetics (e.g. creams, lotions) to the volar forearms within the last 3 days, or bring the test area in contact with water (e.g. no bathing and swimming, only careful bathing excluding the test area) within the last 2 hours. Products were applied once daily to each site for 28 days by the subjects after being provided with training on how much product to apply and where. Subjects were instructed to apply 0.5g of the relevant test product to half their face and one volar forearm once daily throughout the study. Subjects were instructed to not apply any leave on cosmetics (e.g. creams, lotions) other than the provided test products to the test areas throughout the entire course of the study. The last application of the test products was performed 24 ± 2 hours prior to the scheduled visits. Only subjects who completed the product usage as outlined in the protocol (the per protocol population) were included in the data analysis.
For microbiome assessment the participants were swabbed at Day 0 (D0) before product usage commenced and Day 29 (D29) after 4 weeks of regular product usage, with comparisons made between treated and untreated samples per test area (face and forearm) per time point (D0 and D29). DNA was extracted from the swab samples using the NucleoSpin 96 Soil (Macherey-Nagel) kit. Bead beating was done on a Vortex-Genie 2 horizontally at 2700 rpm for 2x5 min. Negative controls (sterile water treated as a sample and no-template PCR controls) were included per batch of samples from the DNA extraction and throughout the laboratory process (including sequencing). A ZymoBIOMICS Microbial Community DNA Standard (Zymo Research) was also included in the analysis as a positive (mock) control. Full details on the microbiome assessment are given in Supplementary Information, section S.2.
Microbiome Alpha diversity was calculated as the number of entities detected (‘richness’), or as the Shannon index based on natural logarithm. The Shannon diversity index accounts for both the number of different species and abundance evenness of the species. To reduce bias from differences in sequencing depth, alpha diversity analyses were based on values calculated from rarefied abundance data (rarefied to a sequencing depth of 4620 reads). A sequencing depth-adjusted richness measure was also calculated as rarefaction may not remove all the sequencing depth related bias. The paired Wilcoxon signed rank test was used as the statistical test used to detect significant differences in alpha diversity measures between groups.
Beta diversity was calculated as weighted UniFrac distances and Bray-Curtis dissimilarity for visualization of samples and air swabs. To enable visual interpretation of distances between the samples in a two-dimensional plot, the samples can be projected onto the first two dimensions of a principal co-ordinates analysis (PCoA) of the distances. Each point of a PCoA plot represents a sample. The longer the distance between two samples, the further apart are the corresponding sample points in the PCoA plot, i.e., points near each other represent samples with similar microbiome compositions. Ellipses were added to the PCoA plots that are centered on the group centroids and cover one standard deviation of the mean of the centroid position, i.e., the ellipses indicate how certain the positions of the group centroids are.
When comparing the beta diversity, a PERMANOVA test was performed using the adonis2 function from the vegan R package with 1000 permutations. While PCoA plots reduce the beta diversity information into two dimensions, PERMANOVA significance tests directly regress the complete beta diversity information (i.e. the distances) to the groups to be compared.
The meta-analysis merged data from the cosmetic studies focusing on the merged datasets for the Phylum and Species of the bacteria present. Data were merged using PANDAS library (Python version 3.10.12). Exploratory analyses were conducted using R libraries (SparkR, sparklyr, dyplr, tidyr, ggplot2, ggpubr, vegan) (R version 4.3.1). To assess for pre- and post-treatment changes in the microbiome, Wilcoxon signed rank tests were used (R library stats).
3. Results
Changes in Shannon Diversity Index for the individual test products are given in Table 3. No significant differences were found for any of the products between baseline and post treatment.
Beta diversity analysis showed no differences between D0 and D29 or between the products and the relevant untreated comparisons at the 95% confidence level (individual data not shown here). Paired Wilcoxon signed rank tests was used to compare abundances at the level of individual ASVs, species, genera, families, orders, and phyla to test for differences in abundance between treatment time points along with whether the levels increased or decreased, Table 4.
Following assessment of data from the individual products, meta-analysis was carried out to determine the effects of topical moisturizer usage on the overall study population as well as to further understand the impact of age, sex and body site on microbiome composition. Microbial composition for the entire study population, including the top three phyla, at the beginning of the study and after product usage on the face and forearm is shown in Figure 1. Once daily product usage for 4 weeks did not result in any significant changes to the gross overall microbial population composition.
Microbiome ‘richness’ scores and Shannon Diversity Index at baseline (D0) as a function of age group and sex and body site (face and forearm) are shown in Figure 2. Variation in the richness of the face microbiome by age group was observed but with, with no clear trend, however richness was greatest in the oldest age group (60+ years). Non-parametric Kruskal-Wallis test indicated that there were significant differences in both ‘richness’ (p=0.001) and Shannon Diversity Index (p<0.001) of the face by age and sex. There were no significant differences in the ‘richness’ (p=0.063) of the forearm by age and sex, however there was a significant difference in Shannon Diversity Index on the forearm (p<0.001).
Wilcoxon-Mann Whitney U tests were used to compare the richness of the treated versus untreated groups after product usage (D29). Separate analyses were run for face and forearm. Results for the face showed no mean difference in the ‘richness’ (p = 0.545) or Shannon Index (p = 0.661). The forearm results also showed no difference in the ‘richness’ (p = 0.240) or Shannon Index (p = 0.193).
Using meta-analysis the effects of age and sex on microbiome diversity were examined. Relative abundance of the main microbial phyla on forearm and face as function of subject age is given in Figure 3.
Relative abundance of the main microbial phyla on forearm and face as function of subject sex is given in Figure 4.
Individual Wilcoxon-signed rank tests were used to compare the pre- and post-treatment counts and relative abundance for face and forearm for the three phyla. There was no significant difference in the pre- and post-treatment, for the treatment groups for Actinobacteriota and Firmicutes (both counts and relative abundance, face and forearm, p>0.05). There were significant differences in the pre- and post-treatment groups for Proteobacteria facial relative abundances (p=0.045) and forearm counts (p=0.015) between the pre- and post-treatment groups.
A total of 34 adverse events (AEs) were reported across the 4 studies. Of the reported AEs, 17 were attributed to the product usage, and 17 occurred in the untreated groups. All product related AEs were minor and all AEs were monitored until resolved.
4. Discussion
Originally considered to be just a dead substrate on the surface of the skin, the stratum corneum actually hosts an extremely diverse ecosystem, comprising a range of environments including dry, moist and oily and with a range of cell types and appendages such as sebaceous and sweat glands and hair follicles [11,12,13]. With such a varied range of environments available, different microbial species colonize the various parts of the body [2,14,15]. Far from being simple colonizers, these microbial communities have been shown to actively contribute to skin homeostasis and health [1], and microbiome dysbiosis has been linked with a variety of skin disorders, such as Atopic Dermatitis, Psoriasis and acne [2,16]. In addition to substances such as sebum which are produced by the body, the skin is also exposed to a wide variety of external chemical agents including topical products and even pollution, all of which can be expected to have an impact on its properties and therefore the microbiome that is present [17,18,19]. It is therefore important to understand how the microbial populations on skin vary throughout life, and the impact of topical skin products on them.
Overall, in the data presented here, topical product usage had a relatively minor impact on the skin microbiome composition after 4 weeks of use. Shannon diversity indices were not significantly changed (Table 3) nor was overall beta diversity. There were however some changes in individual species/genus/phylum levels, for the products (Table 4). It should be noted though that some minor but significant changes at this species/genus/phylum level were seen for the untreated sites as well (individual data for the untreated sites not shown here). As will be mentioned later the impact of the ingredients present in the products also needs to be considered. For instance Xanthomonas species are used in the production of xanthan gum, a common ingredient in hand creams and lotions [20,21]. Xanthan gum is present in a number of the products tested here and the level present on the skin was found to have increased after usage of products DBL and DHC. This has potential skin benefits when used on dry skin as the Xanthomonas genus has a specific enzymatic system for hydrolyzing keratin [22]. The presence of Xanthomonas on skin after usage of topical products containing xanthan gum has been reported before [4]. This emphasizes the need to fully understand the composition of the products being used on skin when interpreting microbiome assessments.
How do the results presented here compare with what has been previously reported with regards to the effects of topical product usage on the skin microbiome? There has been a vast array of studies carried out to look at the effects of cosmetic usage on the skin microbiome, with some studies done using simple cosmetics, while others look to the effects of pre-, pro- or post-biotics ingredients, and with claims of improving bacterial diversity or preserving it [23,24,25,26]. The usage of make up on the face has been reported to increase microbiome diversity, to a greater extent than this is seen with moisturizers and cleansers [27]. Many topical product studies have claimed that their results show an improvement in microbiome structure and composition, however it is important to consider the potential effects of study design in these results. For instance, some studies used the products twice daily [28,29,30,31], others twice daily while continuing to use the subjects own skin routine in addition [32]. Other studies have shown no significant impact on diversity [33]. While the ingredients in some topical products have even been shown to negatively impact microbiome diversity [34], which is a cautionary reminder to those developing products for use on skin.
While research for what topical products need to do to be considered to ‘microbiome friendly’ is still in in its infancy, it does make sense for these types of products to not be drastically impacting the existing microbiome, especially on skin which is already considered healthy [35]. It is also important to ensure that study design should not be overly impacting the results, for instance be making sure that product usage during testing is representative of real-life usage.
Skin microbiome composition and diversity, although typically relatively stable in the short term, can change dramatically with age and sex and with life events such as puberty, pregnancy and menopause all being linked with these variations [12,36,37,38,39].
In the data presented here variation in the richness of the face microbiome by age group was observed but with no clear trend, however, richness was greatest in the oldest age group (60+ years). Overall though, there were significant differences in both ‘richness’ (p=0.001) and Shannon Diversity Index (p<0.001) of the face by age and sex. There were no significant differences in the ‘richness’ (p=0.063) of the forearm by age and sex, however, there was a significant difference in Shannon Diversity Index on the forearm (p<0.001). Generally, microbiome diversity is reported to increase with age which reflects the results reported here [40,41,42]. While skin microbial composition in healthy subjects has been reported to remain relatively stable over time during adulthood [36,41], physiological changes which occur with age, for instance variations in sebum secretion and immune function as well as a decrease in sweat production, may impact the skin microbiome of older individuals. A significant modification in the structure and composition of the skin microbiome has been observed with increased age [43], particularly in women [44]. Increased sebaceous gland activity and a concomitant rise in Cutibacterium acnes levels, has been reported to reduce microbiome diversity in adults vs. infants [45,46] although as the test groups here did not include subjects going through puberty this observation could not be tested for. Increase in microbial diversity was seen with increasing age in a Korean study, however, no difference in alpha diversity was reported by gender [47]. Quan et al. reported that skin microbiome diversity was significantly greater in adults than in adolescents or the elderly [43]. The skin’s ability to retain moisture as well as its sebum production capacities are impacted by both aging and gender [48]. Cutibacterium species prefer an environment with higher moisture and sebum. Male skin microbiome composition varies dramatically across the first 25 years of life, with an increase in diversity being seen with increasing age [49]. Also, males, who have greater sebum secretion that remains stable with ageing [48], were found to have a greater relative abundance of Cutibacterium bacteria, which increased with age. An increase in skin alpha diversity has been reported for post-menopausal women following a decrease in sebaceous gland activity and subsequent reduction in skin sebum levels [44].
It is often assumed by the consumer that increased microbial diversity on skin is better, however it has been reported that drier skin sites are more diverse than those with moist or sebaceous skin [14], and that with subjects who report drier skin the microbiome richness on the face is higher than has been observed for those without dry skin [50]. At other body sites the question of diversity is further complicated. For example, a healthy vagina microbiome typically has low microbiome diversity, while a healthy gut microbiome has high diversity [51,52]. The potential for cross talk between these different microbiomes should also be considered [53,54]. Given the variability of the microbiome across the body and during life, rather than simply aiming for increased diversity with the use of topical moisturizing products, there is a growing emphasis on creating products which respect the microbiome present on the skin especially on skin which is classified as ‘healthy’ as opposed to being in a disease state as the composition of skin flora is reflective of the changes occurring in the body with age [23,35,55,56]. Based on this, is the statement ‘increased microbiome diversity is better for the skin’ too simplistic? Future work should be aimed at addressing this.
Specific bacterial species have been linked with a range of skin conditions such as Xerosis, Atopic Dermatitis (AD) and Psoriasis [4,5,6,7,8]. However, it is not always clear whether these relationships are causal or correlative [57]. Average bacterial counts of the three most prevalent species (Staphylococcus epidermidis, Staphylococcus aureus, Cutibacterium acnes and others) at baseline (D0) for both females and males, grouped by body site are shown in Table 5.
Although the overall levels of S. aureus are relatively low compared with other bacterial species, women appear to have more of it present than men. Elevated S. aureus levels are a potential issue for skin as this can result in increased α-Toxin and proteases which can damage the skin barrier and lead to inflammation [58]. S. aureus in particular has been widely reported to be elevated in subjects with AD, but also to be an initial colonizer of dry skin regions [5,59,60,61,62] and is a strong driver of the ‘itch-scratch cycle’ by its ability to directly activate pruriceptor sensory neurons to drive itch [63]. Available study data of differences in skin hydration between men and women widely varies [64] However, despite strong clinical or objective data on the subject women do report experiencing dry skin more frequently than men [65]. Elevated baseline levels of S. aureus may indicate a higher propensity of women to develop issues with dry, sensitive skin, which would be in keeping with existing data on sex related differences in skin sensitivity [66,67]. This emphasizes the important ongoing need to adequately hydrate and moisturize the skin, especially for women.
It is important discuss the limitations of the retrospective analysis. There was a sample size discrepancy between genders, with a reduced representation of males (n=28) compared to females (n=63). Additionally, there was an uneven distribution of subjects across various age groups, and overall base size was low. Although attempts were made to address this imbalance through age stratification, it resulted in the creation of relatively small groups with unequal age ranges, deviating from standard age stratification by decades. Further work is required focusing on specific age groups to further explore the initial results presented here. These clinical studies and the retrospective analysis solely focuses on assessing the evolution of the skin microbiome, without considering other skin biomarkers such as hydration levels and housing habits (urban vs. rural) which have been reported to impact microbial diversity [43,68]. The study population was predominantly Caucasian (described as White in Table 1), so it was not possible to examine the effects of ethnicity on microbiome composition. Amongst the genetic factors which shape the skin microbiome, ethnicity appears to be a secondary, although not insignificant, one [69]. The microbiome is assumed here to be bacterial in nature, however, bacteria form only part of the living ecosystem present on the stratum corneum. For example, fungi can also be considered as part of the overall microbiota of the skin, and they can have a dramatic impact on its health and function [70,71,72,73]. Incorporating multiple skin biomarkers could lead to a more comprehensive understanding of the dynamics of the skin microbiome. In addition to parameters such as bacterial diversity, more research is required to assess the metabolite pathways at the molecular level and their impact on the microbiome. Given their potential to act as a food source for bacteria the interaction of ingredients in the topical formulation with the microbiome will be the subject of future work.
5. Conclusions
Microbiome composition is important to skin health, form and function. However, the skin is a complex structure and subject to changes as a function of age, gender and body site, and the use of topical products. It is therefore important to understand how these intrinsic and extrinsic factors impact microbiome composition. Four studies have been carried out to determine the effects of long term (4 week) use of different dexpanthenol containing topical skin care products on the skin microbiome of a varied population with cosmetically dry skin. Skin microbiome composition was assessed before and after products usage. No significant changes in microbiome richness or diversity were found for the individual test products, however, meta-analysis of the combined dataset did show changes in microbiome composition as a function of subject sex, age and body site. While low microbial diversity is linked with certain skin conditions such as Acne Vulgaris, it is not sufficient to state that increased diversity is representative of better quality skin. It is important to demonstrate that the use of carefully formulated topical products on skin, when used in a way which is representative of real-life usage conditions, can respect the microbial diversity present on skin across a widely varied study population without significantly impacting the natural diversity which is present.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. S.1: Test product ingredients; S.2: Microbiome assessment.
Author Contributions
Conceptualization, Z.Q., G.B., R.d.S., S.M., K.U. and E.P.; methodology, Z.Q., G.B., R.d.S., S.M. K.U. and E.P.; software, G.B., S.M. and K.U.; validation, Z.Q., R.d.S, and E.P.; formal analysis, Z.Q., S.M., K.U. and E.P.; investigation, K.U. and S.M.; resources, R.d.S.; data curation, K.U.; writing—original draft preparation, Z.Q. and E.P.; writing—review and editing, Z.Q., G.B., R.d.S. S.M., K.U. and E.P.; visualization, Z.Q., G.B., S.M., K.U. and E.P.; supervision, R.d.S. and E.P.; project administration, R.d.S. and K.U. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by Bepanthen, Bayer Consumer Care A.G Basel Switzerland. ZQ, EP, GB and RdS are employees of Bayer Consumer Care A.G.
Institutional Review Board Statement
The studies were conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board of proderm GmbH (IRB code 2022/028, 15th January 2023, IRB code 2022/029 15th January 2023, IRB code 2023/001 11th January 2023, IRB code 2023/002 11th January 2023).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Acknowledgments
Authors gratefully acknowledge that microbiome analysis was performed by Clinical Microbiomics Denmark and medical writing of the manuscript carried out by Dr Jonathan Crowther (JMC Scientific Consulting Ltd, UK).
Conflicts of Interest
E.P, Z.Q, RdS, G.B are employees of Bayer Consumer Care A.G.. The authors declare no conflicts of interest.
References
- Sfriso, R.; Egert, M.; Gempeler, M.; Voegeli, R.; Campiche, R. Revealing the secret life of skin - with the microbiome you never walk alone. Int. J. Cosmet. Sci. 2020, 42(2), 116–126. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2020, 16(3), 143–155. [Google Scholar] [CrossRef] [PubMed]
- Townsend, E.C.; Kalan, L.R. The dynamic balance of the skin microbiome across the lifespan. Biochem. Soc. Trans. 2023, 51(1), 71–86. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Murphy, B.; Grimshaw, S.; Hoptroff, M.; Paterson, S.; Arnold, D.; Cawley, A.; Adams, S.E.; Falciani, F.; Dadd, T.; Eccles, R.; Mitchell, A.; Lathrop, W.F.; Marrero, D.; Yarova, G.; Villa, A.; Bajor, J.S.; Feng, L.; Mihalov, D.; Mayes, A.E. Alteration of barrier properties, stratum corneum ceramides and microbiome composition in response to lotion application on cosmetic dry skin. Sci. Rep. 2022, 12, 5223. [Google Scholar] [CrossRef] [PubMed]
- Ogonowska, P.; Szymczak. K.; Empel, J.; Urbaś, M.; Woźniak-Pawlikowska, A.; Barańska-Rybak, W.; Świetlik, D.; Nakonieczna, J. Staphylococcus aureus from Atopic Dermatitis Patients: Its Genetic Structure and Susceptibility to Phototreatment. Microbiol. Spectr. 2023, 11, e04598–22. [Google Scholar] [CrossRef]
- Wrześniewska, M.; Wołoszczak, J.; Świrkosz, G.; Szyller, H.; Gomułka, K. The Role of the Microbiota in the Pathogenesis and Treatment of Atopic Dermatitis-A Literature Review. Int. J. Mol. Sci. 2024, 25(12), 6539. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Korneev, A.; Peshkova, M.; Koteneva, P.; Gundogdu, A.; Timashev, P. Modulation of the skin and gut microbiome by psoriasis treatment: a comprehensive systematic review. Arch. Dermatol. Res. 2024, 316(7), 374. [Google Scholar] [CrossRef] [PubMed]
- Thio, H.B. The Microbiome in Psoriasis and Psoriatic Arthritis: The Skin Perspective. J. Rheumatol. Suppl. 2018, 94, 30–31. [Google Scholar] [CrossRef] [PubMed]
- Stettler, H.; Crowther, J.M.; Brandt, M.; Boxshall, A.; Lu, B.; de Salvo, R.; Laing, S.; Hennighausen, N.; Bielfeldt, S.; Blenkiron, P. Multi parametric biophysical assessment of treatment effects on xerotic skin. Skin Health Dis. 2021, 1(2), e21. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Qaizar, Z.; Sun, C.; Unbereit, K.; Stettler, H. Skin microbiome effects of long term dexpanthenol based gentle wash and emollient products formulated for dry skin. European Academy of Dermatology and Venereology conference, Berlin, Germany, 11-14 October 2023. [Google Scholar]
- Kligman, A.M. A.M. A brief history of how the dead stratum corneum became alive. In Skin Barrier; Elias, P.M., Feingold, K.R., Eds.; Informa Healthcare Inc.: New York, 2010; pp. 15–24. [Google Scholar]
- Pagac, M.P.; Stalder, M.; Campiche, R. Menopause and facial skin microbiomes: a pilot study revealing novel insights into their relationship. Front. Aging. 2024, 5, 1353082. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Crowther, J.M. Stratum corneum or stratum ecologica? Int. J. Cosmet. Sci. 2019, 41(2), 200–201. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Losada, M.; Crandall, K.A. Spatial diversity of the skin bacteriome. Front. Microbiol. 2023, 14, 1257276. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Brandwein, M.; Fuks, G.; Israel, A.; Hodak, E.; Sabbah, F.; Steinberg, D.; Bentwich, Z.; Shental, N.; Meshner, S. Biogeographical Landscape of the Human Face Skin Microbiome Viewed in High Definition. Acta Derm. Venereol. 2021, 101(11), adv00603. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- 16. Kim, J.; Park, T.; Kim, H.J.; An, S.; Sul, W.J. Inferences in microbial structural signatures of acne microbiome and mycobiome. J. Microbiol. 2021, 59(4), 369–375. [Google Scholar] [CrossRef] [PubMed]
- McCall, L.I.; Callewaert, C.; Zhu, Q.; Song, S.J.; Bouslimani, A.; Minich, J.J.; Ernst, M.; Ruiz-Calderon, J.F.; Cavallin, H.; Pereira, H.S.; Novoselac, A.; Hernandez, J.; Rios, R.; Branch, O.H.; Blaser, M.J.; Paulino, L.C.; Dorrestein, P.C.; Knight, R.; Dominguez-Bello, M.G. Home chemical and microbial transitions across urbanization. Nat. Microbiol. 2020, 5(1), 108–115. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gu, X.; Li, Z.; Su, J. Air pollution and skin diseases: A comprehensive evaluation of the associated mechanism. Ecotoxicol. Environ. Saf. 2024, 278, 116429. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Q.; Li, X.; Zhang, R.Y.; Yuan, C.; Yan, B.; Humbert, P.; Quan, Z.X. Effects of Investigational Moisturizers on the Skin Barrier and Microbiome following Exposure to Environmental Aggressors: A Randomized Clinical Trial and Ex Vivo Analysis. J. Clin. Med. 2023, 12(18), 6078. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Palaniraj, A.; Jayaraman, V. Production, recovery and applications of xanthan gum by Xanthomonas campestris. J. Food Eng. 2011, 106(1), 1–12. [Google Scholar] [CrossRef]
- Dai, X.; Gao, G.; Wu, M.; Wei, W.; Qu, J.; Li, G.; Ma, T. Construction and application of a Xanthomonas campestris CGMCC15155 strain that produces white xanthan gum. Microbiologyopen 2019, 8, e631. [Google Scholar] [CrossRef]
- De Toni, C.H.; Richter, M.F.; Chagas, J.R.; Henriques, J.A.; Termignoni, C. Purification and characterization of an alkaline serine endopeptidase from a feather-degrading Xanthomonas maltophilia strain. Can. J. Microbiol. 2002, 48(4), 342–348. [Google Scholar] [CrossRef] [PubMed]
- Han, J.H.; Kim, H.S. Skin Deep: The Potential of Microbiome Cosmetics. J. Microbiol. 2024, 62(3), 181–199. [Google Scholar] [CrossRef] [PubMed]
- Woolery-Lloyd, H.; Andriessen, A.; Day, D.; Gonzalez, N.; Green, L.; Grice, E.; Henry, M. Review of the microbiome in skin aging and the effect of a topical prebiotic containing thermal spring water. J. Cosmet. Dermatol. 2023, 22(1), 96–102. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Mao, J.; Diaz, I.; Kopylova, E.; Melnik, A.V.; Aksenov, A.A.; Tipton, C.D.; Soliman, N.; Morgan, A.M.; Boyd, T. Multi-omic approach to decipher the impact of skincare products with pre/postbiotics on skin microbiome and metabolome. Front. Med. (Lausanne) 2023, 10, 1165980. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- De Almeida, C.V.; Antiga, E.; Lulli, M. Oral and Topical Probiotics and Postbiotics in Skincare and Dermatological Therapy: A Concise Review. Microorganisms 2023, 11(6), 1420. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Staudinger, T.; Pipal, A.; Redl, B. Molecular analysis of the prevalent microbiota of human male and female forehead skin compared to forearm skin and the influence of make-up. J. Appl. Microbiol. 2011, 110(6), 1381–1389. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, E.; Åkerström, U.; Berger-Picard, N.; Lataste, S.; Gillbro, J.M. Randomized comparative double-blind study assessing the difference between topically applied microbiome supporting skincare versus conventional skincare on the facial microbiome in correlation to biophysical skin parameters. Int. J. Cosmet. Sci. 2023, 45(1), 83–94. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jeong, S.E.; Lee, S.; Kim, S.; Han, H.; Jeon, C.O. Effects of cosmetics on the skin microbiome of facial cheeks with different hydration levels. Microbiologyopen 2018, 7(2), e00557. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cui, S.; Pan, M.; Tang, X.; Liu, G.; Mao, B.; Zhao, J.; Yang, K. Metagenomic insights into the effects of cosmetics containing complex polysaccharides on the composition of skin microbiota in females. Front. Cell. Infect. Microbiol. 2023, 13, 1210724. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Iglesia, S.; Kononov, T.; Zahr, A.S. A multi-functional anti-aging moisturizer maintains a diverse and balanced facial skin microbiome. J. Appl. Microbiol. 2022, 133(3), 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.K.; Lee, S.; Myoung, J.; Hwang, S.J.; Lim, J.M.; Jeong, E.T.; Park, S.G.; Youn, S.H. Effect of the skincare product on facial skin microbial structure and biophysical parameters: A pilot study. Microbiologyopen 2021, 10(5), e1236. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Callejon, S.; Giraud, F.; Larue, F.; Buisson, A.; Mateos, L.; Grare, L.; Guyoux, A.; Perrier, E.; Ardiet, N.; Trompezinski, S. Impact of Leave-on Skin Care Products on the Preservation of Skin Microbiome: An Exploration of Ecobiological Approach. Clin. Cosmet. Investig. Dermatol. 2023, 16, 2727–2735. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhao, H.; Yu, F.; Wang, C.; Han, Z.; Liu, S.; Chen, D.; Liu, D.; Meng, X.; He, X.; Huang, Z. The impacts of sodium lauroyl sarcosinate in facial cleanser on facial skin microbiome and lipidome. J. Cosmet. Dermatol. 2024, 23(4), 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- van Belkum, A.; Lisotto, P.; Pirovano, W.; Mongiat, S.; Zorgani, A.; Gempeler, M.; Bongoni, R.; Klaassens, E. Being friendly to the skin microbiome: Experimental assessment. Front. Microbiomes 2023, 1, 1077151. [Google Scholar] [CrossRef]
- Oh, J.; Byrd, A.L.; Park, M. NISC Comparative Sequencing Program; Kong HH, Segre JA. Temporal Stability of the Human Skin Microbiome. Cell 2016, 165(4), 854–866. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Park, J.; Schwardt, N.H.; Jo, J.H.; Zhang, Z.; Pillai, V.; Phang, S.; Brady, S.M.; Portillo, J.A.; MacGibeny, M.A.; Liang, H.; Pensler, M.; Soldin, S.J.; Yanovski, J.A.; Segre, J.A.; Kong, H.H. Shifts in the Skin Bacterial and Fungal Communities of Healthy Children Transitioning through Puberty. J. Invest. Dermatol. 2022, 142(1), 212–219. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Leem, S.; Keum, H.L.; Song, H.J.; Gu, K.N.; Kim, Y.; Seo, J.Y.; Shin, J.G.; Lee, S.G.; Lee, S.M.; Sul, W.J.; Kang, N.G. Skin aging-related microbial types separated by Cutibacterium and α-diversity. J. Cosmet. Dermatol. 2024, 23(3), 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Radocchia, G.; Brunetti, F.; Marazzato, M.; Totino, V.; Neroni, B.; Bonfiglio, G.; Conte, A.L.; Pantanella, F.; Ciolli, P.; Schippa, S. Women Skin Microbiota Modifications during Pregnancy. Microorganisms 2024, 12(4), 808. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Somerville, D.A. The normal flora of the skin in different age groups. Br. J. Dermatol. 1969, 81(4), 248–258. [Google Scholar] [CrossRef] [PubMed]
- Luna, P.C. Skin Microbiome as Years Go By. Am. J. Clin. Dermatol. 2020, 21 (Suppl 1), 12–17. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Myers, T.; Bouslimani, A.; Huang, S.; Hansen, S.T.; Clavaud, C.; Azouaoui, A.; Ott, A.; Gueniche, A.; Bouez, C.; Zheng, Q.; Aguilar, L.; Knight, R.; Moreau, M.; Song, S.J. A multi-study analysis enables identification of potential microbial features associated with skin aging signs. Front. Aging. 2024, 4, 1304705. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ying, S.; Zeng, D.N.; Chi, L.; Tan, Y.; Galzote, C.; Cardona, C.; Lax, S.; Gilbert, J.; Quan, Z.X. The Influence of Age and Gender on Skin-Associated Microbial Communities in Urban and Rural Human Populations. PLoS One 2015, 10(10), e0141842. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shibagaki, N.; Suda, W.; Clavaud, C.; Bastien, P.; Takayasu, L.; Iioka, E.; Kurokawa, R.; Yamashita, N.; Hattori, Y.; Shindo, C.; Breton, L.; Hattori, M. Aging-related changes in the diversity of women's skin microbiomes associated with oral bacteria. Sci. Rep. 2017, 7(1), 10567. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Murphy, B.; Hoptroff, M.; Arnold, D.; Cawley, A.; Smith, E.; Adams, S.E.; Mitchell, A.; Horsburgh, M.J.; Hunt, J.; Dasgupta, B.; Ghatlia, N.; Samaras, S.; MacGuire-Flanagan, A.; Sharma, K. Compositional Variations between Adult and Infant Skin Microbiome: An Update. Microorganisms 2023, 11(6), 1484. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Leyden, J.J.; McGinley, K.J.; Mills, O.H.; Kligman, A.M. Age-related changes in the resident bacterial flora of the human face. J. Invest. Dermatol. 1975, 65(4), 379–381. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Son, S.M.; Park, H.; Kim, B.K.; Choi, I.S.; Kim, H.; Huh, C.S. Taxonomic profiling of skin microbiome and correlation with clinical skin parameters in healthy Koreans. Sci. Rep. 2021, 11(1), 16269. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Luebberding, S.; Krueger, N.; Kerscher, M. Skin physiology in men and women: in vivo evaluation of 300 people including TEWL, SC hydration, sebum content and skin surface pH. Int. J. Cosmet. Sci. 2013, 35(5), 477–483. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Tang, M.; Dong, K.; Zhou, J.; Wang, D.; Song, L. Changes in the skin microbiome during male maturation from 0 to 25 years of age. Skin Res. Technol. 2023, 29(9), e13432. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Russo, E.; Di Gloria, L.; Cerboneschi, M.; Smeazzetto, S.; Baruzzi, G.P.; Romano, F.; Ramazzotti, M.; Amedei, A. Facial Skin Microbiome: Aging-Related Changes and Exploratory Functional Associations with Host Genetic Factors, a Pilot Study. Biomedicines 2023, 11(3), 684. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ottinger, S.; Robertson, C.M.; Branthoover, H.; Patras, K.A. The human vaginal microbiota: from clinical medicine to models to mechanisms. Curr. Opin. Microbiol. 2024, 77, 102422. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489(7415), 220–230. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chaudhary, P.P.; Kaur, M.; Myles, I.A. Does "all disease begin in the gut"? The gut-organ cross talk in the microbiome. Appl. Microbiol. Biotechnol. 2024, 108(1), 339. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhou, Z.; Feng, Y.; Xie, L.; Ma, S.; Cai, Z.; Ma, Y. Alterations in gut and genital microbiota associated with gynecological diseases: a systematic review and meta-analysis. Reprod. Biol. Endocrinol. 2024, 22(1), 13. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wallen-Russell, C.; Wallen-Russell, S. ; Meta Analysis of Skin Microbiome: New Link between Skin Microbiota Diversity and Skin Health with Proposal to Use This as a Future Mechanism to Determine Whether Cosmetic Products Damage the Skin. Cosmetics 2017, 4(2), 14. [Google Scholar] [CrossRef]
- Carvalho, M.J.; S Oliveira, A.L.; Santos Pedrosa, S.; Pintado, M.; Pinto-Ribeiro, I.; Madureira, A.R. Skin Microbiota and the Cosmetic Industry. Microb. Ecol. 2023, 86(1), 86–96. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Thompson, A.; Jaros, J.; Blackcloud, P.; Hsiao, J.; Shi, V.Y. Updated understanding of Staphylococcus aureus in atopic dermatitis: From virulence factors to commensals and clonal complexes. Exp. Dermatol. 2021, 30(10), 1532–1545. [Google Scholar] [CrossRef] [PubMed]
- Blicharz, L.; Rudnicka, L.; Samochocki, Z. Staphylococcus aureus: an underestimated factor in the pathogenesis of atopic dermatitis? Postepy. Dermatol. Alergol. 2019, 36(1), 11–17. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tauber, M.; Balica, S.; Hsu, C.Y.; Jean-Decoster, C.; Lauze, C.; Redoules, D.; Viodé, C.; Schmitt, A.M.; Serre, G.; Simon, M.; Paul, C.F. Staphylococcus aureus density on lesional and nonlesional skin is strongly associated with disease severity in atopic dermatitis. J. Allergy. Clin. Immunol. 2016, 137, 1272–1274.e3. [Google Scholar] [CrossRef] [PubMed]
- Cleary, J.M.; Lipsky, Z.W.; Kim, M.; Marques, C.N.H.; German, G.K. Heterogeneous ceramide distributions alter spatially resolved growth of Staphylococcus aureus on human stratum corneum. J. R. Soc. Interface. 2018, 15(141), 20170848. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Imokawa, G.; Abe, A.; Jin, K.; Higaki, Y.; Kawashima, M.; Hidano, A. Decreased level of ceramides in stratum corneum of atopic dermatitis: an etiologic factor in atopic dry skin? J. Investig. Dermatol. 1991, 96(4), 523–526. [Google Scholar] [CrossRef]
- Watanabe, M.; Tagami, H.; Horii, I.; Takahashi, M.; Kligman, A.M. Functional analyses of the superficial stratum corneum in atopic xerosis. Arch. Dermatol. 1991, 127(11), 1689–1692. [Google Scholar] [CrossRef]
- Deng, L.; Costa, F.; Blake, K.J.; Choi, S.; Chandrabalan, A.; Yousuf, M.S.; Shiers, S.; Dubreuil, D.; Vega-Mendoza, D.; Rolland, C.; et al. S. aureus drives itch and scratch-induced skin damage through a V8 protease-PAR1 axis. Cell 2023, 186, 5375–5393.e25. [Google Scholar] [CrossRef] [PubMed]
- Rahrovan, S.; Fanian, F.; Mehryan, P.; Humbert, P.; Firooz, A. Male versus female skin: What dermatologists and cosmeticians should know. Int. J. Womens Dermatol. 2018, 4(3), 122–130. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jemec, G.B.; Serup, J. Scaling, dry skin and gender. A bioengineering study of dry skin. Acta Derm. Venereol. Suppl. (Stockh.) 1992, 177, 26–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Su, Y.; Zheng, B.; Wen, S.; Liu, D.; Ye, L.; Yan, Y.; Elias, P.M.; Yang, B.; Man, M.Q. Gender-related characterization of sensitive skin in normal young Chinese. J. Cosmet. Dermatol. 2020, 19(5), 1137–1142. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Farage, M.A. The Prevalence of Sensitive Skin. Front. Med. 2019, 6, 98. [Google Scholar] [CrossRef]
- Callewaert, C.; Ravard Helffer, K.; Lebaron, P. Skin Microbiome and its Interplay with the Environment. Am. J. Clin. Dermatol. 2020, 21 (Suppl 1), 4–11. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Skowron, K.; Bauza-Kaszewska, J.; Kraszewska, Z.; Wiktorczyk-Kapischke, N.; Grudlewska-Buda, K.; Kwiecińska-Piróg, J.; Wałecka-Zacharska, E.; Radtke, L.; Gospodarek-Komkowska, E. Human Skin Microbiome: Impact of Intrinsic and Extrinsic Factors on Skin Microbiota. Microorganisms 2021, 9(3), 543. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nguyen, U.T.; Kalan, L.R. Forgotten fungi: the importance of the skin mycobiome. Curr. Opin. Microbiol. 2022, 70, 102235. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jo, J.H.; Kennedy, E.A.; Kong, H.H. Topographical and physiological differences of the skin mycobiome in health and disease. Virulence 2017, 8(3), 324–333. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Meray, Y.; Gençalp, D.; Güran, M. Putting It All Together to Understand the Role of Malassezia spp. in Dandruff Etiology. Mycopathologia 2018, 183(6), 893–903. [Google Scholar] [CrossRef] [PubMed]
- Leyden, J.J.; Kligman, A.M. Interdigital athlete's foot: new concepts in pathogenesis. Postgrad. Med. 1977, 61(6), 113–116. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Baseline (D0) and post product usage (D29) average relative abundance of the top 3 phyla (Actinobacteriota, Firmicutes, Proteobacteria) and other species by body site (face, forearm).
Figure 1.
Baseline (D0) and post product usage (D29) average relative abundance of the top 3 phyla (Actinobacteriota, Firmicutes, Proteobacteria) and other species by body site (face, forearm).
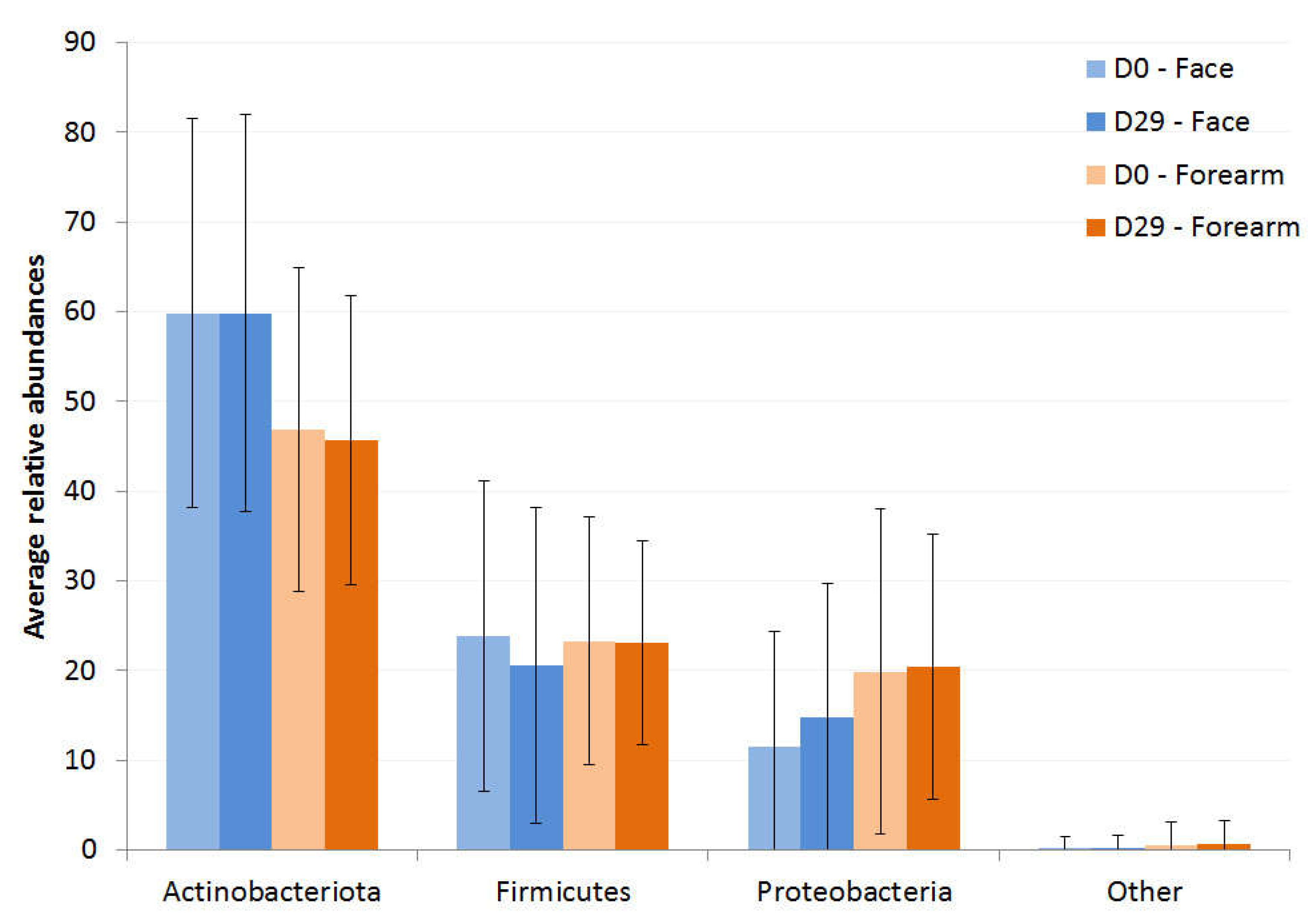
Figure 2.
Alpha diversity at baseline (D0) by age group and sex shown as mean and standard deviation. a). variation in the richness on face, b). Shannon diversity of facial microbiome, c). variation in the richness of forearm, d). Shannon diversity of forearm microbiome.
Figure 2.
Alpha diversity at baseline (D0) by age group and sex shown as mean and standard deviation. a). variation in the richness on face, b). Shannon diversity of facial microbiome, c). variation in the richness of forearm, d). Shannon diversity of forearm microbiome.
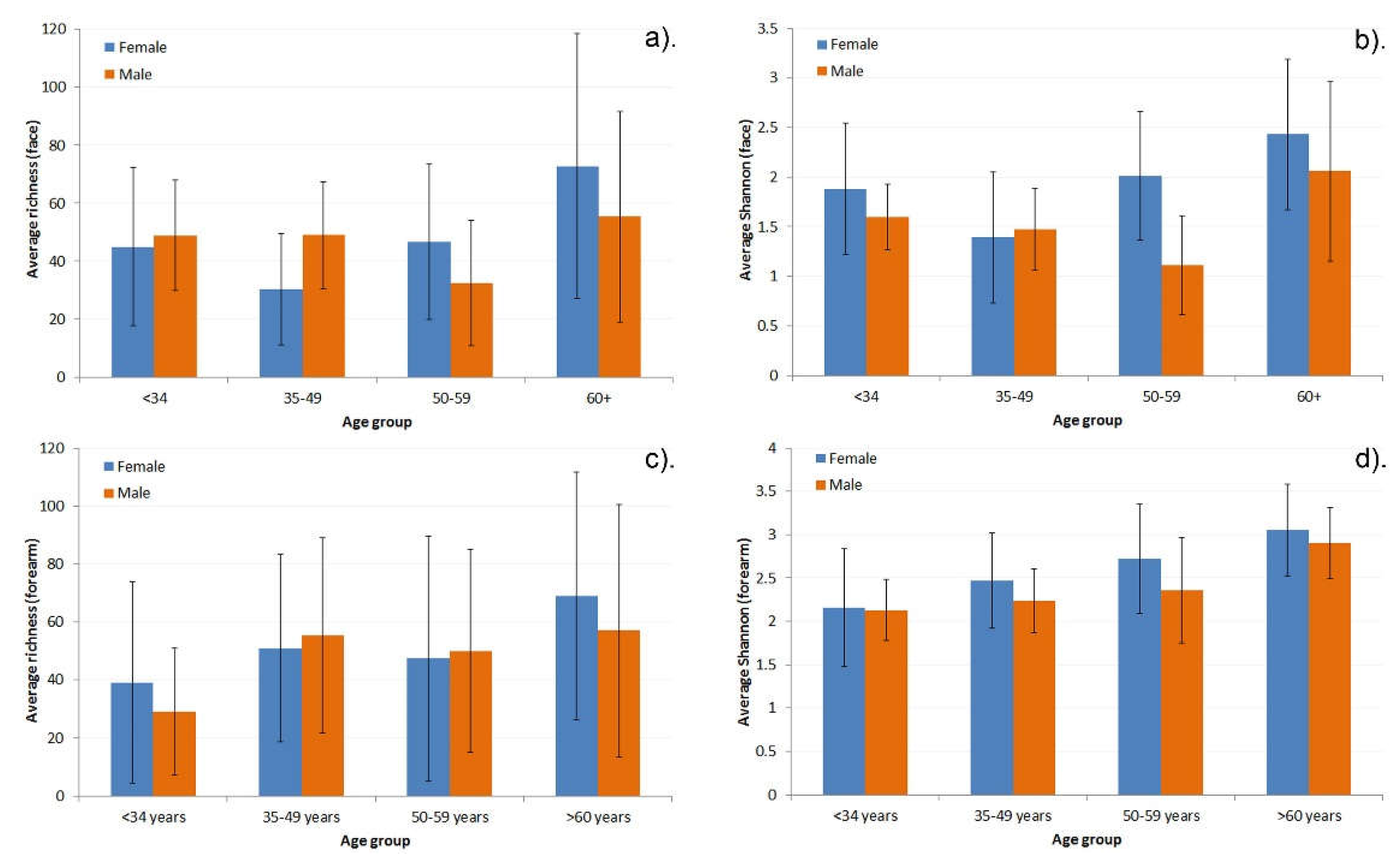
Figure 3.
Relative abundance of the top 3 phyla (Actinobacteriota, Firmicutes, Proteobacteria) for a). the face, and b). the forearm, at baseline (D0) and post treatment (D29), split by age group.
Figure 3.
Relative abundance of the top 3 phyla (Actinobacteriota, Firmicutes, Proteobacteria) for a). the face, and b). the forearm, at baseline (D0) and post treatment (D29), split by age group.
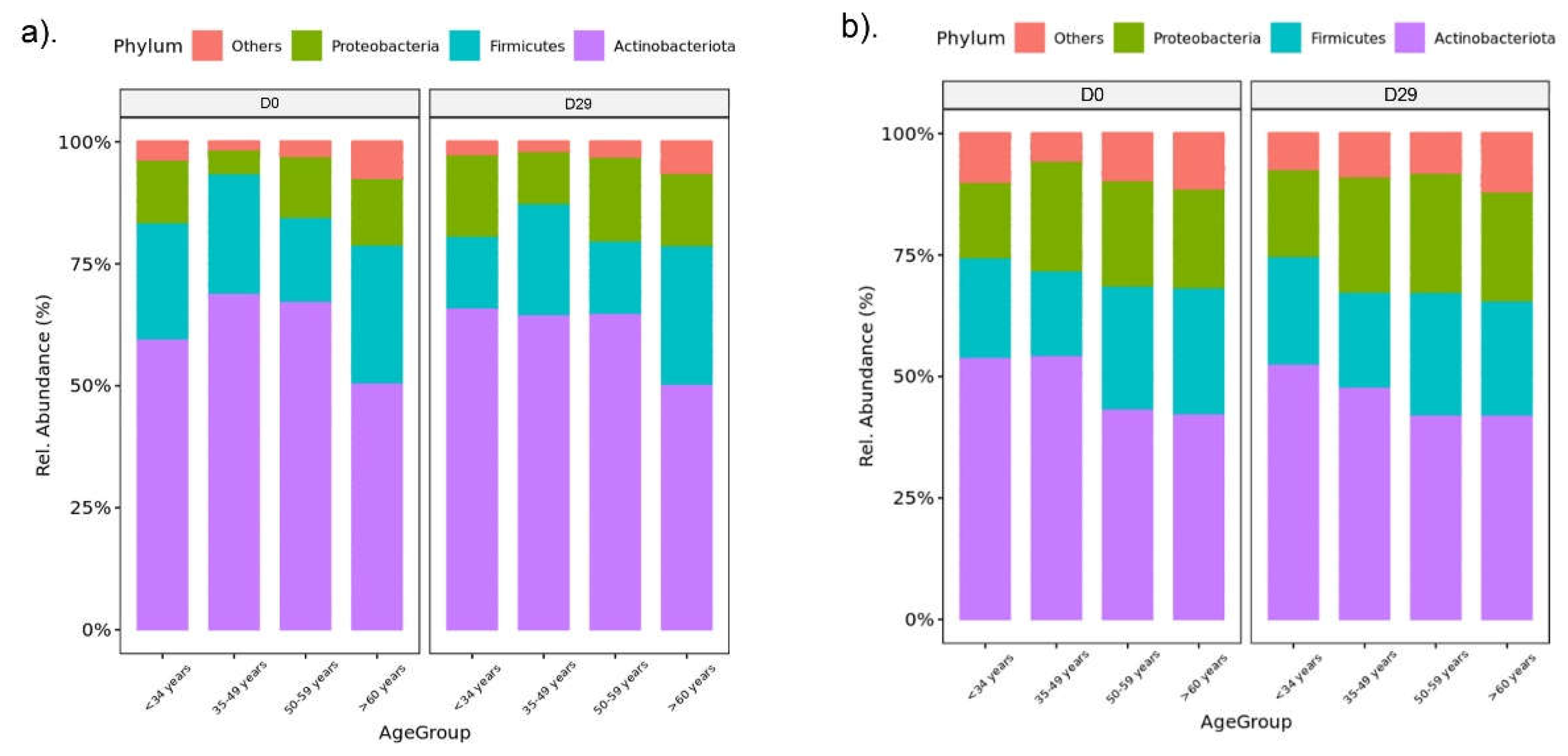
Figure 4.
Relative abundance of the 3 top phyla (Actinobacteriota, Firmicutes, Proteobacteria) for a). the face, and b) the forearm, at baseline (D0) and post treatment (D29), split by sex.
Figure 4.
Relative abundance of the 3 top phyla (Actinobacteriota, Firmicutes, Proteobacteria) for a). the face, and b) the forearm, at baseline (D0) and post treatment (D29), split by sex.
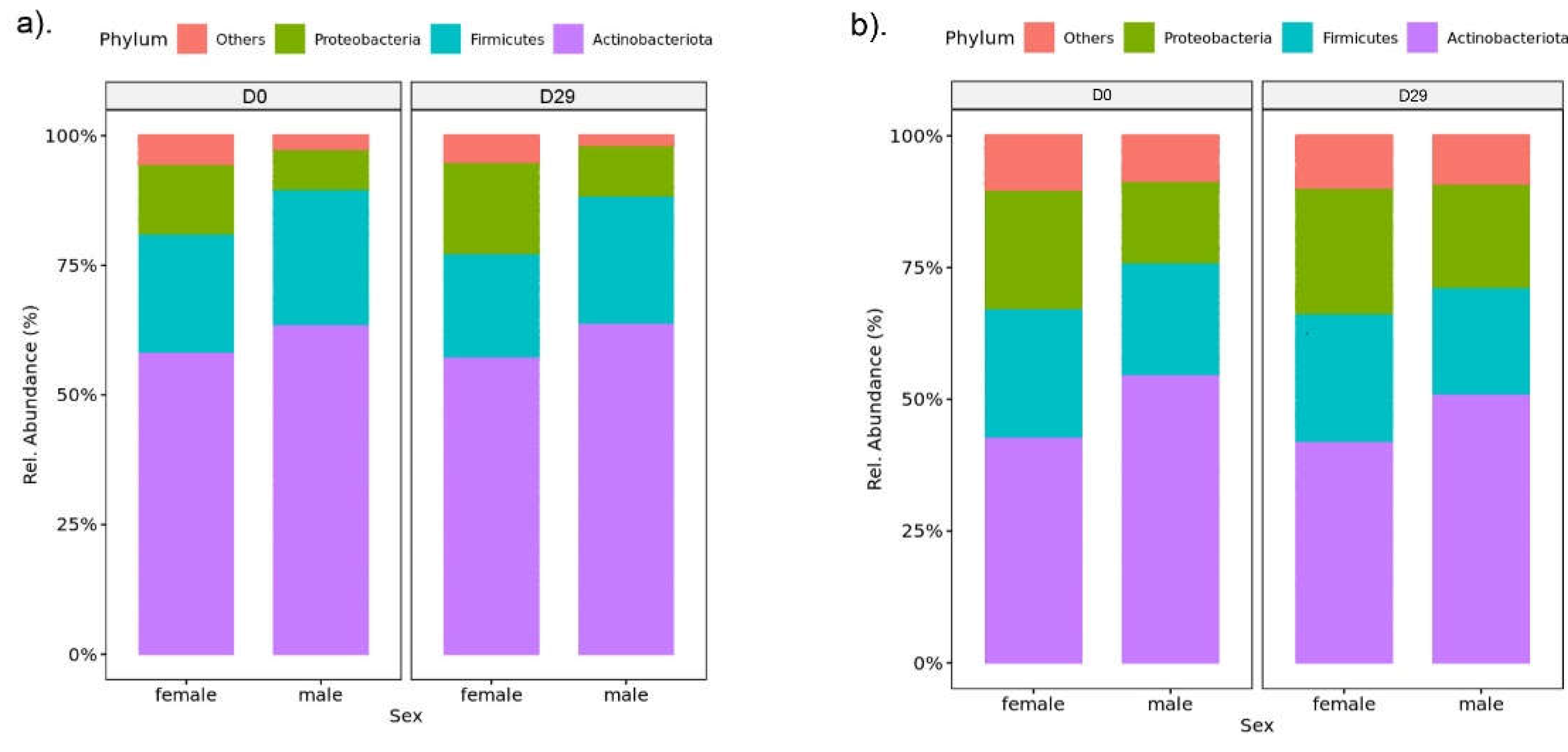

Table 1.
Test population demographics.
| Variable | Male | Female | ||||
| N | % | Mean (SD) | N | % | Mean (SD) | |
| Age (range 22-65) | 52.1 (12.4) | 51.3 (13.1) | ||||
| Fitzpatrick skin type I II III IV V |
0 15 10 2 1 |
0 53.6 35.7 7.1 3.6 |
8 34 19 1 1 |
12.6 54.0 30.2 1.6 1.6 |
||
| Ethnicity White Black Asian |
23 3 2 |
82.1 10.7 7.2 |
61 2 0 |
96.8 3.2 0 |
Table 2.
Product codes and descriptions.
| Product Code | Product Name | |
| DFCD | Dexpanthenol repair complex (DRC) Face cream day | |
| DFCN | DRC Face cream night | |
| DFC-SPF | DRC Face cream (SPF 25) | |
| DFW | Dexpanthenol Face wash | |
| DBL | DRC Body lotion (low lipid level) | |
| DBH | DRC Body balm (high lipid level) | |
| DHC | DRC Hand cream | |
| DBW | Dexpanthenol Body wash | |
Table 3.
Individual product data for median Shannon Diversity Index (with Q1 and Q3, the 1st and 3rd quartile) at baseline (D0) and post treatment (D29). NS – not significant at the 95% confidence level.
Table 3.
Individual product data for median Shannon Diversity Index (with Q1 and Q3, the 1st and 3rd quartile) at baseline (D0) and post treatment (D29). NS – not significant at the 95% confidence level.
| Product | Body site | D0 (Shannon Index) | D29 (Shannon Index) | Difference | |
| DFCD | Face | 1.297 (1.120, 1.598) | 1.263 (1.083, 1.559) | -0.034 (NS) | |
| DFCN | Face | 2.352 (1.525, 2.224) | 2.149 (1.529, 2.135) | -0.203 (NS) | |
| DFC-SPF | Face | 1.601 (1.293, 1.792) | 1.658 (1.326, 1,872) | +0.057 (NS) | |
| DFW | Face | 1.831 (1.476, 2.027) | 1.741 (1.287, 1.902) | -0.090 (NS) | |
| DBL | Forearm | 2.808 (2.250, 2.637) | 2.900 (2.439, 2.864) | +0.092 (NS) | |
| DBH | Forearm | 2.738 (2.442, 2,714) | 3.130 (2.853, 2.986) | +0.392 (NS) | |
| DHC | Forearm | 2.444 (2.194, 2.551) | 2.940 (2.449, 2.846) | +0.496 (NS) | |
| DBW | Forearm | 2.735 (2.544, 2.859) | 2.894 (2.495, 2.784) | +0.159 (NS) | |
Table 4.
Taxonomic microbiome changes after 4 weeks of product usage on the face and forearm. NSD – no significant differences (at the 95% confidence level), ASV - amplicon sequence variant.
Table 4.
Taxonomic microbiome changes after 4 weeks of product usage on the face and forearm. NSD – no significant differences (at the 95% confidence level), ASV - amplicon sequence variant.
| Product | Body site | Taxonomic changes (D29 vs D0) | Category | p value |
| DFCD | Face | NSD | - | - |
| DFCN | Face | NSD | - | - |
| DFC-SPF | Face | NSD | - | - |
| WPF | Face |
Corynebacterium tuberculostearicum ↓ Frankia nodulisporulans ↑ |
Species Species |
0.003 0.002 |
| DBL | Forearm | ASV_0009 – Xanthomonas campestris ↑ Xanthomonas campestris ↑ Veillonella ↑ Xanthomonas ↑ Veillonellaceae ↑ Xanthomonadaceae ↑ Firmicutes_C ↑ |
ASV Species Genus Genus Family Family Phylum |
<0.001 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 |
| DBH | Forearm | Haemophilus_D parainfluenzae ↑ | Species | 0.001 |
| DHC | Forearm | Epilithonimonas ↑ Rothia ↑ Xanthomonas ↑ |
Genus Genus Genus |
0.010 0.007 0.008 |
| WPB | Forearm | NSD | - | - |
Table 5.
Average bacterial counts of the three most prevalent species (S. epidermidis, S. aureus, C. acnes and others) for females and males, grouped by body site. All subjects at D0 are included. SD – standard deviation, CI – confidence interval, N – number of subjects.
Table 5.
Average bacterial counts of the three most prevalent species (S. epidermidis, S. aureus, C. acnes and others) for females and males, grouped by body site. All subjects at D0 are included. SD – standard deviation, CI – confidence interval, N – number of subjects.
| Species | Body site | Sex | Average Count (SD) | Lower CI | Upper CI | N |
| S. epidermidis | Face | Female | 4250.56 (5083.19) | 3352.22 | 5148.90 | 62 |
| Male | 7150.76 (9486.93) | 4643.50 | 9658.03 | 28 | ||
| Forearm | Female | 1607.17 (2122.55) | 1232.06 | 1982.28 | 63 | |
| Male | 2441.20 (2658.55) | 1744.85 | 3137.54 | 28 | ||
| S. aureus | Face | Female | 87.57 (317.14) | 14.31 | 160.83 | 36 |
| Male | 9.38 (38.85) | -6.81 | 25.57 | 11 | ||
| Forearm | Female | 91.43 (370.03) | 4.12 | 178.75 | 36 | |
| Male | 0.00 (0.00) | 0.00 | 0.00 | 11 | ||
| C. acnes | Face | Female | 21448.19 (18560.41) | 18168.06 | 24728.32 | 62 |
| Male | 31771.31 (19974.18) | 26492.41 | 37050.21 | 28 | ||
| Forearm | Female | 6461.51 (9054.49) | 4861.34 | 8061.69 | 63 | |
| Male | 8494.68 (9603.19) | 5979.45 | 11009.91 | 28 | ||
| Other | Face | Female | 32.28 (391.06) | 29.51 | 35.06 | 62 |
| Male | 36.86 (535.64) | 31.08 | 42.63 | 28 | ||
| Forearm | Female | 39.48 (440.08) | 36.34 | 42.62 | 63 | |
| Male | 46.26 (480.83) | 41.13 | 51.39 | 28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



