
Submitted:
25 October 2024
Posted:
29 October 2024
You are already at the latest version
Abstract
Fusarium wilt caused by Fusarium oxysporum f. sp. cubense (Foc) is a major disease affecting banana production around the globe. To study the movement of Foc in planta and investigate potential physical mechanisms of deterrence, the roots of a set of banana cultivars were challenged with a GFP-tagged subtropical race 4 strain of Foc. These plants included susceptible and resistant varieties including the plantain 'FHIA25' (AAB), dessert hybrids 'Lady Finger' (AAB), 'FHIA02' (AAAA), as well as the industry preferred Cavendish line, 'Williams' (AAA), and its somaclone derivative 'GCTCV119' (AAA). GFP localisation in planta confirmed that Foc can infect all four cultivars. Its movement through roots to the rhizome and to the aerial parts of the plants was evident. Tyloses formation in the root vasculature was also detected by GFP auto-fluorescence and further confirmed using scanning electron microscopy. While tyloses formation did not prevent colonisation and infection, their formation restricted the fungus within the xylem vessels, slowing its proliferation. This mechanism may be an important factor in providing plant host tolerance to Foc but it doesn't provide complete immunity. Fluorescently tagged Foc is a useful tool for understanding the infection process of the Fusarium wilt disease of banana and aiding in the integrated management of this important disease.
Keywords:
Fusarium wilt of banana
; green fluorescent protein
; tyloses formation
; Fusarium oxysporum f. sp. cubense
; banana plantains
; banana hybrids
; confocal laser microscopy
; scanning electron microscopy
; reporter gene system
; Fusarium-banana interactions
1. Introduction
Banana (Musa sp.), an herbaceous genus of plants native to tropical Indomalaya and Australia, is an important food crop world-wide and is valued for its flavours and nutritional values. Banana is widely grown in the tropics and is consumed as staple food in many countries, having contributed to a total global production of 135 million tons in 2022, with world exports totaling 24 million tons worth approximately 13 billion USD [1].
Cavendish (genome AAA), including the cultivar known as 'Williams', is the most dominant dessert banana in the market accounting for 47% of the global banana production [1]. The over-reliance on so few cultivars and lack of genetic diversity makes banana crops vulnerable to pests and diseases, leading to major outbreaks that put constrains on production at both commercial and small-scale levels [2]. One such disease is the Fusarium wilt of banana (FWB), caused by the soil-borne fungus Fusarium oxysporum f. sp. cubense (Foc). The Fusarium oxysporum species complex (FOSC) contains many plant pathogens that infect economically important crop species, identified as crop specialist forms (formae speciales) or defined by the hosts on which they cause disease. Within formae speciales, isolates are classified using a race structure based on the cultivars to which they are pathogenic [3,4,5]. However, the relationships of different formae speciales of F. oxysporum is complex with evidence of a polyphyletic history and horizontal gene transfer [5,6].
Within the banana-infecting Fusaria, Foc race 1 and Foc race 4 have had the greatest impact on global banana production. Foc race 1caused the demise of 'Gros Michel' (AAA) banana in the 1950s leading to its replacement by the Foc race 1 resistant Cavendish cultivars [7]. Foc race 1 causes disease on the cultivars ‘Gros Michel’ (AAA), ‘Maqueño’ (AAB), ‘Silk’ (AAB), ‘Pome’ (AAB) and ‘Pisang Awak’ (ABB) [8]. The emergence of Foc race 4 from Southeast Asia in 1960s saw Cavendish cultivars and certain other cultivated forms succumbing once again to Fusarium wilt [9]. Foc race 4 is divided into; strains that infect Cavendish after a period of relatively cooler temperatures in the subtropics known as Subtropical race 4 (STR4) and strains known as Tropical race 4 (TR4) that cause infection under both tropical and subtropical conditions [10,11]. Foc TR4 has now spread across all major continents, decimating banana plantations in its wake and finally reaching the major banana production regions of Latin America in 2019 [12]. Foc TR4 was first recorded in the early 1990s in Southeast Asia and soon after in 1997 in Northern Territory, Australia [13]. However, it was almost another two decades before it was detected in Australia's main banana production area, North Queensland, when in 2015, TR4 was reported in Tully, resulting in an unprecedented increase in regional biosecurity monitoring and strict quarantine control [14].
Fusarium wilt is not a disease that can be easily eradicated, so it is important to minimise the dissemination of infection through quarantine control or the use of clean planting materials [15]. For locations where the disease has spread to, there are no effective chemical control methods against FWB, hence the focus has been on developing cultivars carrying genetic resistance to Foc [16,17,18]. Somaclonal variants with enhanced resistance to Foc TR4 and Foc STR4 have been identified from Cavendish derived plants [9,19]. The resistance of banana genotypes to both Foc STR4 and TR4 have been investigated in field and pot trials [20,21,22,23]. Rhizomes appear to play an important role in determining a plant's response to Foc [20]. Some of these somaclonal variants, particularly the Giant Cavendish tissue culture variants (GCTCV), have shown promising results in TR4-infested field trials in the Philippines [24], Australia [21], Africa [25] and China [22] as well as in pot trials [20,22].
Conventional crossing has also been used to develop Foc resistant cultivars. The Honduran Foundation for Agricultural Research (FHIA) established a banana breeding program to develop resistant banana types, to combat a range of different diseases as alternatives to the predominantly susceptible Cavendish cultivars [26]. These varieties include dwarf desserts banana, plantain and cooking bananas hybrids, some of which were resistant to Foc Race 4 types. Field evaluations suggest that some of these FHIA varieties can be as productive and robust as natural hybrids [27]. Despite their high yields and resistances to Fusarium wilt, the FHIA hybrids have not met the needs and preferences of the market [28]. However, these hybrids were derived in turn from a set of diverse progenitor hybrids (SH hybrids) that have been used extensively in the breeding programs at the International Institute for Tropical Agriculture (IITA) in East and West Africa, the National Agricultural Research Organisation (NARO) in Uganda, as well as the Brazilian Agricultural Research Corporation (Embrapa) in Brazil and The Centre for International Cooperation in Agricultural Research (CIRAD) in France. So far, FHIA lines have been used for dissecting disease responses against Foc TR4 in field and pot trials [20,21].
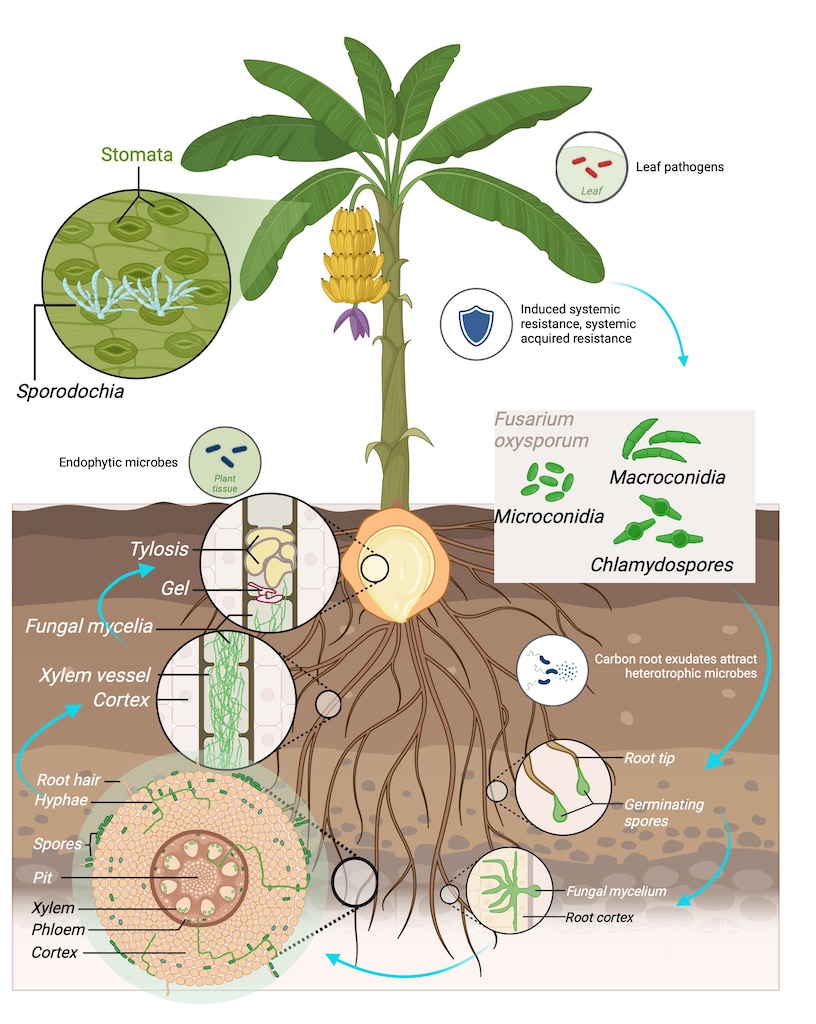
Foc produces three different types of asexual spores; the short-lived microconidia and macroconidia, and the long-lived chlamydospores. All three types of spores have the ability to cause infection on susceptible banana plants [29]. Foc is known to survive in the soil for decades, persisting in the soil on dead host plant tissues in the absence of a suitable host either as chlamydospores or as saprophytes [30]. Foc spores germinate and directly adhere to the roots and gain entry through the root epidermis or open wounds [31,32], followed by the movement into the xylem vessels of the roots and rhizomes. As the fungus colonises the plant, extensive mycelial networks occlude the xylem vessels, obstructing nutrient and water uptake, which results in plant wilting and eventual plant death [33]. External symptoms of FWB include pseudostem splitting, leaf yellowing, and necrosis and stunted growth. The fungus is not transferred to the fruit parts, but the infected plants will produce less fruit [7]. Recently it has been demonstrated that Foc TR4 can colonise the fruit peduncle in Foc TR4-infected banana plants as improvements in agronomic practices to manage the disease have resulted in affected plants producing bunches [34].
Previous studies in many different host species, have demonstrated a series of plant defense reactions against Fusarium pathogens, including papilla formation, production of antimicrobial substances, cell wall lignification, occlusion by gums, gels or tyloses within xylem vessels and vessel crushing [35,36]. The roles of each of these plant reactions in resistance are unclear, but they all contribute to the overall resistance capacity in plants. For instance, tyloses formation is considered a protective response of trees and herbaceous plants to vascular damage whether from mechanical injury or fungal or bacterial infections [37]. Tyloses formation occurs in the vessel lumens of root xylem in infected plants. They are formed from the extension of parenchyma cells through the pit membrane of the inner xylem wall and can fill up the entire xylem lumen. The amount of tyloses accumulation can vary depending on the resistance level of a cultivar. VanderMolen et al. (1987) showed that tyloses formation occurs in both susceptible and resistant cultivars, where rapid occlusion with tyloses occurred in infected root xylem vessels of resistant cultivars, whereas susceptible cultivars showed a similar tyloses initiation with plant growth reduced at later stages [38]. Tyloses formation has been shown to have a significant role in plant defense mechanisms in relation to the susceptibility of banana plants.
Green Fluorescent Protein (GFP)-tagged Foc strains have been used to observe the movement of Foc in banana hosts resistant and susceptible to Foc TR4 [39,40] and Foc STR4 [33]. In the study presented here, histological process of Foc infection was investigated using five banana cultivars: 'Williams' Cavendish (AAA), its somaclone 'GCTCV119' (AAA), as well as Pome type cultivar known in Australia as 'Lady Finger' (AAB), 'FHIA02' (AAAA), and 'FHIA25' (AAB) hybrids. We observed that host susceptibility is associated with the active proliferation of the fungus in the xylem vessels of the rhizomes. In comparison, the fungus was restricted in the rhizomes of the Foc resistant cultivars 'FHIA25' and 'GCTCV119', although, the ability of Foc to still enter the roots appeared unhindered in these cultivars. Given that Foc TR4 resistant cultivars have already been deployed in some of the Foc TR4 affected regions around the world, it will be important to understand the level of resistance such cultivars possess. It is equally important to address what constitutes resistance specifically relating to plant host's response to Foc and whether the fungal presence can still be detected in resistant cultivars. Thus, addressing some of these questions allows the suitability of deploying apparent resistant genotypes in Foc TR4 affected regions to be assessed.
2. Materials and Methods
2.1. The GFP-tagged Fusarium Oxysporum f. sp. Cubense Isolate
The GFP-tagged Foc STR4 (GFP-Foc-STR4) strain UQ6817 was derived from the strain BRIP40389 (Queensland Plant Pathology Herbarium) and has been described in previous studies with respect to its pathogenicity on Musa [20,41]. The strain was single-spored and stored in the form of water-agar plugs in sterile water at 4°C.
2.2. The banana Cultivars and Plant Growth Conditions
Tissue-culture banana plantlets of the known Foc STR4 susceptible lines 'FHIA02', 'Lady Finger', 'Williams', and the Foc STR4 tolerant/resistant lines 'GCTCV119' (somaclonal variant of 'Williams') and 'FHIA25' were de-flasked into 30 cell potting trays of 35 cm (length) x 29 cm (width) x 5.5 cm (depth). The trays were placed on a lab bench and were positioned under fluorescent light set at a day/night cycle of 16 h / 8 h and temperature of 20-22°C. The soil mix, UQ23, contained 70% composted pine bark 0-5 mm in size and 30% coco peat and has a pH range of 5.5-6.5. After 4 weeks of hardening-off post tissue culture, plants were repotted into 140 mm diameter pots and to which a teaspoon of a balanced fertiliser (Osmocote) was added. Plants were transferred to a temperature-controlled glasshouse with day and night temperatures set at 22°C/26°C and were grown for 6 weeks under a regime of watering to field capacity once every 2-3 days.
2.3. Preparation of Spore Suspension For Plant Inoculation
The GFP-Foc-STR4 strain was grown on full strength potato dextrose agar (PDA), supplemented with 100 mg/L hygromycin B, and was incubated at 25°C for 4 days. Four 5 mm2 mycelial blocks of GFP-Foc-STR4 were cut from a fully colonised PDA plate and were used to inoculate 500 mL of sterile potato dextrose broth (PDB), supplemented with 50mg/L hygromycin B. The cultures were placed on a shaker and agitated at 28°C, 180 rpm for 4 days. The culture was then filtered through four layers of sterile Miracloth and the spores were collected and then washed with sterile distilled water (SDW). For root dipping, the concentration of the spore suspension was adjusted to 2×106 spores per mL with SDW. Additional spore suspension was used to drench the soil at a concentration of approximately 50,000 spores per gram of soil.
For plants subjected to scanning electron microscopy, the plants were inoculated with 45 g of GFP-Foc-STR4 infested Japanese millet (Echinochloa esculenta) variety 'Shirohie', as previously described [20]. A spore suspension of the same concentration and application method as above was also directly poured onto the root zone of the plants. Non-inoculated plants served as controls.
2.4. Plant Inoculation
After 6 weeks, 30 plants of each 'FHIA25', 'FHIA02', 'Lady Finger', 'Williams' and 'GCTCV119' were inoculated by root dipping, followed by soil-drenching using the GFP-Foc-STR4 spore suspension. Plants with five to six healthy leaves and a stem height of 30 cm were selected and plant roots were washed with SDW and then dipped in the spore suspension for 2 hours. Instead of a spore suspension, non-inoculated control plants were root-dipped in SDW for 2 hours. All plants were then transplanted to 250 mm diameter pots containing the soil mix UQ23.
2.5. Plant Harvest and Visualisation of GFP Colonised Plants
Plants were harvested at weekly time points up to 70 days throughout this experiment. External symptoms were visually assessed on plant leaves and pseudostems at harvest. Internal symptoms including discolouration within the roots, rhizome and stems were assessed on the day when confocal microscopy was performed to detect the localisation of GFP-Foc-STR4 inside the host. For visualisation under a confocal microscope, a double-edged razor blade was used for sectioning plant tissues (transverse and longitudinal), including roots, stem, rhizomes and leaf parts. Sections were mounted in sterile deionised water.
GFP was detected using Zeiss 700 Laser Scanning Microscope (Zeiss, Oberkochen, Germany) and a laser at an excitation wavelength of 488 nm. The Z-stack function was used to capture 3D images consisting of 10-30 optical slices taken at intervals of 1-5 μm. The T-PMT (transmission detector setting) was also used to see the sectioned plant tissues in an overlay of brightfield.
2.6. Reisolation of GFP-Foc-STR4 from Symptomatic Plants
Diseased plant materials were surface sterilised using 0.5% bleach for 30 seconds and then washed twice in SDW before blotting dry on sterile paper towel under laminar flow hood. Tissues including leaf, petiole, stem just above the rhizome, central cylinder of the rhizome and rhizome node connecting roots, were cut into 2-5 mm sections using a sterile scalpel and then embedded into water agar. The plates were incubated at 25°C for 10 days. The presence of Fo-like spores and mycelia were identified using a dissecting microscope. The Fo-like colonies (white mycelia with pink to mauve staining of agar) were then sub-cultured onto half strength PDA, supplemented with 100 mg/L hygromycin B. Plates were scored for the presence or absence of Fo-like growth after 10 days of incubation at 25°C. These colonies were further examined under confocal microscope to confirm GFP fluorescence.
2.7. Sample Preparation and Scanning Electron Microscopy
Primary roots and rhizomes at the base of the banana plants were sectioned for scanning electron microscopy (SEM) at 2- and 6-weeks post inoculation. The sample preparation protocol was adapted from a previous study [42]. Sections of 0.5 to 1 cm in size were fixed in a buffer containing 2.5% glutaraldehyde, 0.1 M monobasic sodium phosphate and 0.1 M dibasic sodium phosphate. The samples were embedded in a specimen tube containing agarose (50%, w/v) and were sliced at a depth of 100 μm using a vibratome (VT1000 S, Leica Biosystems, Wetzlar, Germany). Sectioned samples were stored in a 6% sodium azide buffer.
The primary root and rhizome were washed twice by the same 0.1 M sodium phosphate buffer to replace the glutaraldehyde. Following this, all processing was performed in a Biowave microwave (Ted Pella, Redding, USA). Each step consisted of a one minute on, one minute off, one minute on process using the Biowave operated at 150W. Dehydration in ethanol used a graded series of 60 %, 70 %, 80 %, 90 % and two 100 % steps. This was followed by chemical drying using hexamethyldisilazane (HMDS) added in steps as 1:1 diluted with absolute ethanol, then twice in 100 ethanol. The 100% HMDS was left overnight to evaporate.
Once dried, each section was mounted onto an aluminium stub using double-sided sticky carbon tabs, and was sputter coated with platinum to create electrical conductivity on the sample surface. Approximately 400 sections were imaged by the SEM: HitachiTM4000Plus Bench top (Hitachi High-Tech, Tokyo, Japan) in a high-vacuum mode operating at 15 kV and the working distance of 15 mm to 18 mm. The presence or absence of the tyloses within the tissue due to Foc inoculation were analysed by SEM comparing with previous studies [38,43].
3. Results
Assessment of symptomatic plants at 32-35 dpi revealed varying degree of internal discolouration in the rhizomes of all cultivars except 'GCTCV119' (Figure S1). Extensive discolouration was also observed in the pseudostems of all cultivars. Reisolation of F. oxysporum-like colonies from symptomatic 'FHIA02' and 'FHIA25' plants showed that GFP-Foc-STR4 was present in the rhizome and lower stem of both cultivars at 35 dpi (Table S1).
3.1. The Detection of Foc at the Onset of Infection in the Banana Cultivars
Assessment of the suspension culture of GFP-Foc-STR4 showed strong fluorescence associated with both the microconidia and macroconidia of this strain, consistent with the constitutive expression of GFP (Figure 1A).
At 5 days post inoculation (dpi), highly abundant microconidia and mycelial networks were detected under a confocal microscope in the xylems of lateral and main roots of 'FHIA02' plants (Figure 1B-C). Extensive microconidia and hyphae were detected in the lateral roots of 'Williams' at 5 dpi (Figure 1E) and appeared to occlude the xylem of lateral roots at 9 dpi (Figure 1F). Likewise, the presence of mycelia was abundant in the xylem vessels of the lateral roots of 'Lady Finger' plants at 8 dpi (Figure 1J) and was observed at a similar level in 'FHIA2'. This is consistent with the susceptibility of these two cultivars to Foc. By contrast, the presence of the fungus, in the tolerant/resistant line 'FHIA25' was detected at a lower frequency. This was observed in particular for plants at 5 dpi, where only a single hyphal strand and a few conidia were detected (Figure 1D). In GCTCV119 plants, the hyphae were detected in the lateral roots at 5-7 dpi (Figure 1G, 1I) but to a lesser extent than in 'Williams', 'Lady Finger' or 'FHIA02'.
3.2. Foc STR4 Colonisation of Roots and Rhizome in 'FHIA02'
At 12-70 dpi, it was evident that the fungus had moved uninhibited through the plant vasculature in both the roots and rhizomes of 'FHIA02' (Figure 2). Extensive mycelial networks were detected in the fine roots of 'FHIA02' as early as 12 dpi (Figure 2A). Chlamydospores and a spore germ tube were evident on the epidermis of the lateral roots at 14 dpi (Figure 2B). Intercellular movement of the hyphae was also detected at 14 dpi (Figure 2C). Apparent establishment of the fungus through a wound site in a root hair was visualised at 19 dpi (Figure 2D). Extensive mycelial networks and germinated spores were detected not only in the fine roots but also in the main roots and root nodes connecting to the rhizome at 26-42 dpi (Figure 2E-J).
In the rhizome, intercellular presence of the fungus through the rhizome cortex was observed as early as 14 dpi (Figure 3A-C). An intercellular hypha with an apparent single terminal chlamydospore was also visualised (Figure 3B). Strong vascular fluorescence with microconidia forming in clusters was observed in the rhizome at 14 dpi (Figure 3C). Abundant germinated microconidia were observed in the xylem vessels at 36 dpi (Figure 3D-E). Tyloses were noted by the appearance of fine angular intra-xylem walled networks similar to those published by VanderMolen et al. (1987) [38]. These structures auto-fluoresced as did the plant cell wall structures of the xylem (Figure 3D-E). The tyloses were observed apparently occluding vessels of the rhizome at 68-70 dpi (Figure 3F-G). Microconidia and chlamydospores were co-localised in and around the xylem vessels.
3.3. GFP-Foc-STR4 in the Foc Resistant Cultivar 'FHIA25'
In the resistant cultivar 'FHIA25', presence of the fungus in the fine roots was clearly visualised at 12-49 dpi (Figure 4). The extent of colonisation by the fungus in the roots appeared similar to that in 'FHIA02'. At 12 dpi, microconidia produced on false heads in monophialides were detected in the fine roots (Figure 4A). Germinated spores in the xylem vessels were also observed at 14 dpi (Figure 4B). The infection appeared extensive and consistent across the rest of the time points, with the fungus having clearly colonised the xylem vessels at 21 dpi (Figure 4C) and extended through the lateral root nodes connecting to the rhizome at 26 dpi (Figure 4D). This process of continual occurrence of spore germination was evident even at the later time points up to 29 dpi, with germinated microconidia and hyphae clearly visualised in the root cap and the elongation zone (Figure 4E-F). Plants can mount a defense response to a xylem-invading vascular wilt pathogen by depositing a vascular coating made up of lignin and suberin in the colonised vessels [44]. The vascular coating is typically visualised as auto-fluorescence from phenolic deposits. Strong GFP fluorescence was detected in the colonised compartments at 36-49 dpi, suggesting the presence of vascular coatings at these time points (Figure 4G-I). Co-locolisation of the GFP auto-fluorescence with the fungus was apparent in these regions.
When the rhizome of 'FHIA25' was assessed for the presence of Foc STR4, unlike the root tissues, it did appear that the fungus was more restricted when compared with 'FHIA02' (Figure 5). Strong GFP auto-fluorescence was associated with the infected regions in the rhizome and main root nodes connecting to the rhizome (Figure 5A, 5D-E, 5G-I). The presence of GFP-Foc-STR4 also appeared confined in the xylem vessels (Figure 5B-C). At 36-41 dpi, tyloses were observed in the vasculature of the rhizome (Figure 5D-E). The movement of hyphae was restricted between individual tyloses (Figure 5F). Tyloses formation was consistently detected in the rhizome at 42-70 dpi (Figure 5G-I). At the same time, minimal fungal presence was detected in these regions.
3.4. Assessing the Presence of Foc in the Leaves of FHIA02 and FHIA25
Presence of the fugus was consistently detected in the outer leaf sheath and petiole edge of both 'FHIA02' and 'FHIA25' plants (Figure 6). Presence of microconidia and hyphae was detected at time points as early as 14-16 dpi (Figure 6A, 6G). In both cultivars, the fungus was confined to the xylem vessels in the midrib of the leaf sheath at 29 dpi (Figure 6B, 6H). Mycelial networks were also detected on the epidermis of leaf sheaths (Figure 6C, 6I). At 54-62 dpi, macroconidia and chlamydospores were abundantly visualised on the edge of the petiole of a senescing leaf in 'FHIA02' (Figure 6D-E). Chlamydospores and hyphae were also observed in the petiole of 'FHIA25' at 62-70 dpi (Figure 6J-K)
3.5. Analysis of Foc infected 'FHIA02' and 'FHIA25' Plants Using Scanning Electron Microscopy
An analysis using scanning electron microscopy on the primary roots of both cultivars detected the presence of tyloses and fungal structures in the xylem vessels (Figure 7 and Figure 8). Tyloses were clearly identified in the vascular bundles of both cultivars at 14 and 42 dpi (Figure 7C, 7E, 7F, 7H-I, 8C, 8F, 8H-I). Tyloses of various sizes, including those that were taking shape, were observed in the xylem vessels of 'FHIA02' and 'FHIA25' plants inoculated with GFP-Foc-STR4 (Figure 7F, 8F, 8H). The tyloses were in close proximity to the fungus (Figure 7C) and in some cases, were forming in the same vascular bundle as the fungus (Figure 8C). Furthermore, some tyloses were also detected in the vascular bundles of control plants of 'FHIA02' not treated with Foc, which suggests that tyloses can form during normal plant growth (Figure 7A).
Fungal growth was observed in the parenchyma cells and vascular bundles of both cultivars at 14 dpi and 42 dpi, consistent with the detection of the fungus inside the host at these time points in the GFP study. Clear presence of mycelia was detected within individual vascular bundles and the pits of the main roots (Figure 7C, 7E, 8C, 8G). As far as roots are concerned, there were no noticeable differences in the number of tyloses formed or the extent of colonisation by the fungus in the vasculature between 'FHIA02' and 'FHIA25'.
3.6. The movement of GFP-Foc-STR4 in 'Williams', 'GCTCV119' and 'Lady Finger'
The GFP tagged fungus was visualised in the primary roots of 'GCTCV119', 'Williams' and 'Lady Finger' at 15-45 dpi (Figure S2). Chlamydospores and hyphae were detected in the root vasculature of each cultivar. To assess whether the fungus moves through the roots into the rhizome, we assessed its presence in the root nodes connecting to the corms in each cultivar at 32-60 dpi. GFP tagged hyphae and mycelia were present in the rhizome nodes of all three cultivars, suggesting that the fungus moved from the roots into the rhizome through the vasculature (Figure S3). Within the rhizomes, the fungus was not detected in 'GCTCV119' at 26-42 dpi (Figure S4A-C). However, chlamydospores and hyphae were clearly present in the vasculature of the corm in 'Lady Finger' and 'Williams' at 27-40 dpi (Figure S4D-I). Furthermore, the regions where infection was observed also showed a strong GFP fluorescence.
Tyloses were localised within the vascular bundles of primary roots and corm in 'GCTCV119' and 'Williams' plants at 55-59 dpi. In 'GCTCV119', tyloses were also formed in the non-inoculated controls, indicating that tyloses can be naturally formed in plants (Figure 9A) as noted above for the FHIA lines. In both 'Williams' and its somaclonal derivative 'GCTCV119', GFP-Foc-STR4 appeared to be confined in the occluded xylem vessels (Figure 9B, 9E-F) or in between two adjacent vessels (Figure 9D).
Above ground, GFP-Foc-STR4 was detected in the pseudostem of 'Lady Finger' and 'Williams' at 18-20 dpi (Figure S5A-C) but was not detected in that of 'GCTCV119' at 28 dpi (Figure S5E). At 51-59 dpi, mycelia were detected in the pseudostem of all three cultivars (Figure S5B, S5D-F). In the leaves of all three cultivars, mycelial networks and sporodochia were consistently detected at both early (13-40 dpi) and late (60-65 dpi) time points (Figure S5G-L). Sporodochia were also detected in the pseudostem of 'GCTCV119' at 55 dpi (Figure S5F).
4. Discussion
Green fluorescent proteins have been a critical tool in understanding pathogen and host plant interactions in planta [20,45,46]. Being able to visualise the fungus in plantae means that the progress of infection can be analysed in great detail. In F. oxysporum, the production of microconidia and macroconidia are critical for the fungus to proliferate inside the hosts. Understanding the accumulation of spores in specific compartments of plants has relevance for the management and containment of Foc in the field. The plant responses associated with the pathogen attack are not well known in resistant/tolerant cultivars such as 'FHIA25' and 'GCTCV119'.
The movement of Foc through the roots was observed in all five cultivars, including 'Williams', 'Lady Finger', 'FHIA02', 'FHIA25', and 'GCTCV119'. Despite having good levels of resistance against Foc STR4, chlamydospores and microconidia of the inoculated GFP-STR4 isolate were observed to attach and germinate on the surface of the root tip and fine root hairs in 'FHIA25'. This was followed by the penetration of the root surface and the movement of hyphae in the root vascular systems. These observations suggest that Foc can actively enter banana hosts via the root systems regardless of the level of resistance the hosts carry. In both 'FHIA02' and 'FHIA25', the intercellular movement of the fungus was observed on the epidermis and then in the elongation zones of the roots. This is consistent with the characteristic movement of a biotrophic pathogen and has been observed in the infection process of other Fusarium species [47,48]. The presence of the fungus was consistently detected in the xylem vessels of both the roots and root nodes connecting to the rhizome. This suggests that the vascular streams facilitate the movement of the fungus inside the plant host in previously described susceptible and resistant lines.
Our previous study showed that the plant host's response, in the rhizome, plays an important role in inhibiting the fungus from spreading to the rest of the plant [20]. In this study, a differential response was observed in the rhizome of 'FHIA02' and 'FHIA25' plants. In the symptomatic rhizomes of 'FHIA02', proliferation of spores and mycelia was clearly detected in the xylem vessels. Interestingly, only a few hyphae were detected in these regions of 'FHIA25', and they appeared to be confined in the xylem. The formation of tyloses wase detected in these regions of both cultivars. Furthermore, in the susceptible cultivars 'Lady Finger' and 'Williams', the observations were that they were extensively colonised by the fungus in the rhizome, a characteristic of Foc susceptibility, whereas the fungus displayed limited movement and appeared confined in the rhizome of 'GCTCV119'. Taken together, these observations suggest that the rhizome is essential in the interplay between the pathogen and plant host.
A similar pattern of restricted colonisation in Dianthus caryophyllus by F. oxysporum f. sp. dianthi has been reported and further characterisation revealed that the infected regions of the xylem became compartmentalised by cell wall thickening, hyperplasia of parenchyma cells and the built-up of vascular occluded materials [36]. In the current study, vascular occluded xylem vessels were detected in the rhizome of both resistant and susceptible cultivars inoculated with the fungus. Vascular occlusion in the rhizome was detected as early as 36 dpi in 'FHIA25' and was detected for the first time in the 'FHIA2' at 68 dpi. Although SEM showed that vascular occlusion occurred as early as 14 dpi in the main roots of both 'FHIA2' and 'FHIA25' inoculated with the fungus. Occluded vessels were similarly visualised in 'GCTCV119' and 'Williams' at 56-59 dpi. These results collectively suggest that vascular occlusion is an inducible plant mechanism to prevent the spread of an invading pathogen. While it may contribute to deterring the movement of the pathogen inside the host, tyloses formed as extensions of contact parenchyma cells [38] do not fully explain the host resistances observed in 'FHIA25' and 'GCTCV119'. Formation of tyloses is typically triggered by infections [49], wounding [50], heartwood formation [51] and abscission [52]. The ability to form tyloses was an important factor in resistance to F. oxysporum in cotton [53] and has been shown to be upregulated by an exogenous chemical application in banana [54]. Therefore, the presence of tyloses observed in this study is consistent with their roles in growth and development, as well as in the regulation of stresses including pathogen attacks and activation of oxidative stress.
Another aspect of pathogen deterrence is the formation of vascular coatings in or around infected regions observed in this study. These regions have been identified as plant physico-chemical barriers induced against xylem vascular wilt pathogens [44]. Particularly, gel and lignin depositions have been found to be associated with vertical and horizontal restrictions, respectively, of F. oxysporum f. sp. cubense in banana hosts [38,55]. These mechanisms involving the formation of gels, gums, or mucilage in and around the vascular systems have been shown to limit fungal growth in banana [43], tomato [56], carnation [57], pea [35,58], cotton [45,53], and bean plants [59]. These barriers are mostly composed of carbohydrates like pectin, polyphenols, and sometimes phytoalexins, lignin-like compounds or lipoidal substances [35]. It was observed in peas that the production of carbohydrates and polyphenolics by vascular parenchyma cells progressively accumulated in the lumen of xylem cells, highlighting important functions of these compounds in defense against F. oxysporum f. sp. pisi [35].
The presence of chlamydospores and hyphae on the petiole and outer leaf sheath was detected in both 'FHIA02' and 'FHIA25'. Despite 'FHIA25' being resistant to Foc STR4, the fungus was able to move through the outer leaf sheaths and reach the aerial parts of the plant. Furthermore, sporodochia were detected in the pseudostem of 'GCTCV119' as well as around the stomata in the leaves of 'William' and 'Lady Finger'. These results are consistent with a previous study which showed the movement of Foc through the leaf sheaths [40] and capability of reaching the outside of the leaf sheath via stomata [33]. However, the transmission of Foc through the aerial part of a Foc resistant plant has not been previously reported.
To date, conventional breeding efforts have not yielded varieties resistant to Foc TR4 that are also agronomically viable [12]. 'GCTCV119', is one of the Cavendish tissue culture derived somaclones that have been shown to carry improved tolerance to Foc TR4 [9]. Although it should be noted that 'GCTCV119' and the improved variant 'GCTCV218', also known as 'Formosana', are not truly resistant to Foc TR4 [12]. 'GCTCV218' did seem to be moderately resistant to Foc TR4 in one study [25] and intermediately resistant only in the first cropping cycle in another study [21]. 'GCTCV218' has replaced Cavendish cultivars in TR4-affected regions in the Philippines and Mozambique [25]. Similarly, 'FHIA25' has previously shown to be highly resistant to both Foc STR4 and TR4 [20,21]. Given the evidence in this study to suggest that Foc can colonise resistant/tolerant cultivars such as 'GCTCV119' and 'FHIA25', it is important then to understand the inoculum threshold levels these and other resistant cultivars can withstand before they succumb to the disease. Further stress testing of these cultivars under different conditions will need to be performed to guide the deployment of these cultivars in Foc TR4-affected regions and to maintain and improve appropriate management strategies to control Foc TR4.
5. Conclusions
In this study, we have assessed the in plantae movement of GFP-tagged Foc STR4 in five important banana cultivars and we found that the pathogen is restricted in the rhizome of Foc resistant cultivars, but not in the susceptible cultivars. This suggests that the rhizome is a key factor in determining the magnitude and the spread of the fungus to other parts of the plant. The presence of the fungus was detected in the leaves and the outer leaf sheaths of the resistant cultivars. This has important implications regarding the potential spread of this pathogen amongst the Foc race 4 resistant cultivars that are currently being used in Foc TR4 affected regions around the world. Results from this study can be used to improve existing monitoring and containment protocols currently placed in regions afflicted by the Fusarium wilt of banana, specifically in relation to the deployment of resistant cultivars in the field. It should also be noted that bioseurity concerns prohibited the use of Foc TR4 in this study.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Assessment of discolouration in the rhizomes of the banana cultivars used in this study.; Figure S2: GFP-Foc-STR4 visualised in the main roots of 'GCTCV119' (G), 'Williams' (W) and 'Lady Finger' (LF) at 15—45 dpi; Figure S3: GFP-Foc-STR4 visualised in the corm nodes of 'Williams' (W), 'GCTCV119' (G) and 'Lady Finger' (LF) at 26—60 dpi; Figure S4: GFP-Foc-STR4 visualised in the corms of 'Williams' (W), 'GCTCV119' (G) and 'Lady Finger' (LF) at 26—51 dpi; Figure S5: GFP-Foc-STR4 visualised in the pseudostems and leaves of 'Lady Finger' (LF), 'Williams' (W) and 'GCTCV119' (G) at 18—65 dpi; Table S1: Reisolation of Fusarium oxysporum-like colonies from the leaves, petioles, lower sems and rhizomes of 'FHIA02' and 'FHIA25' plants inoculated with GFP-Foc-STR4.
Author Contributions
Conceptualization, A.C. and E.A.B.A.; methodology, A.C., T.Y.C., Y.C. and S.M.A.F.; software, A.C., T.Y.C., Y.C. and S.M.A.F.; validation, A.C., J.A., J.S., H.C., S.G., B.F., A.S., D.M.G. and E.A.B.A.; formal analysis, A.C., T.Y.C., Y.C. and S.M.A.F.; investigation, T.Y.C., Y.C. and S.M.A.F.; resources, A.C. and E.A.B.A.; data curation, A.C.; writing—original draft preparation, A.C.; writing—review and editing, A.C., T.Y.C., Y.C., S.M.A.F., J.A., J.S., H.C., S.G., B.F., A.S., D.M.G. and E.A.B.A.; visualization, A.C.; supervision, A.C. and E.A.B.A.; project administration, A.C. and E.A.B.A.; funding acquisition, A.C. and E.A.B.A. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by the Bill and Melinda Gates Foundation through its grant to the International Institute of Tropical Agriculture (IITA) under the project Accelerated Breeding of Better Bananas, grant number IITA 20600.15/0008-8—Phase II. Research funding support for this project was also provided by Hort Innovation Australia under grant BA17006. Jay Anderson was supported by Hort Innovation Australia grant BA14014. Jiaman Sun was supported by Guangxi Natural Science Foundation grant 2021GXNSFAA196014.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Acknowledgments
We would like to kindly thank Sharon Hamill for the supply of tissue culture derived plants of the banana cultivars used in this study. We would also like to thank Ning Chen for the in vitro multiplication and maintenance of these plantlets in tissue culture. We also graciously thank Richard I. Webb for providing technical support in scanning electron microscopy.
Plant material declaration
Plants used in this study were generated from the Australian in vitro banana cultivar collection that is maintained in the Quality Banana Approved Nursery (QBAN) scheme accredited Plant Biotechnology Laboratory located at the Maroochy Research Facility, Department of Agriculture and Fisheries, Nambour, Queensland, Australia. All cultivars were sourced under agreements allowing them to be used for research purposes. No ownership is claimed for those cultivars sourced from owners or other institutions. The plants were destructively sampled for the purposes of this study and were not further propagated.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- FAO. https://www.fao.org/statistics/en. 2024.
- Ortiz, R.; Ferris, R.; Vuylsteke, D. Banana and plantain breeding. In Bananas and plantains; Springer: 1995; pp. 110-146.
- Maryani, N.; Lombard, L.; Poerba, Y.; Subandiyah, S.; Crous, P.; Kema, G. Phylogeny and genetic diversity of the banana Fusarium wilt pathogen Fusarium oxysporum f. sp. cubense in the Indonesian centre of origin. Stud mycol 2019, 92, 155-194. [CrossRef]
- Lombard, L.; Sandoval-Denis, M.; Lamprecht, S.; Crous, P.W. Epitypification of Fusarium oxysporum–clearing the taxonomic chaos. Pers: Mol Phylogeny Evol Fungi 2019, 43, 1-47. [CrossRef]
- Czislowski, E.; Fraser-Smith, S.; Zander, M.; O'Neill, W.T.; Meldrum, R.A.; Tran-Nguyen, L.T.T.; Batley, J.; Aitken, E.A.B. Investigation of the diversity of effector genes in the banana pathogen, Fusarium oxysporum f. sp. cubense, reveals evidence of horizontal gene transfer. Mol Plant Pathol 2018, 19, 1155-1171. [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: concordant evidence from nuclear and mitochondrial gene genealogies. Proc Nat Acad Sci 1998, 95, 2044-2049. [CrossRef]
- Pegg, K.G.; Coates, L.M.; O’Neill, W.T.; Turner, D.W. The Epidemiology of Fusarium Wilt of Banana. Front Plant Sci 2019, 10. [CrossRef]
- Ploetz, R.C. Management of Fusarium wilt of banana: A review with special reference to tropical race 4. Crop Prot 2015, 73, 7-15. [CrossRef]
- Hwang, S.C.; Ko, W.H. Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan. Plant dis 2004, 88, 580-588. [CrossRef]
- Buddenhagen, I. (2009). Understanding strain diversity in Fusarium oxysporum f. sp. cubense and history of introduction of 'Tropical Race 4' to better manage banana production. Acta Hortic 828, 193-204. [CrossRef]
- Mostert, D.; Molina, A.B.; Daniells, J.; Fourie, G.; Hermanto, C.; Chao, C.P.; Fabregar, E.; Sinohin, V.G.; Masdek, N.; Thangavelu, R. The distribution and host range of the banana Fusarium wilt fungus, Fusarium oxysporum f. sp. cubense, in Asia. PLoS One 2017, 12, e0181630. [CrossRef]
- Munhoz, T.; Vargas, J.; Teixeira, L.; Staver, C.; Dita, M. Fusarium Tropical Race 4 in Latin America and the Caribbean: status and global research advances towards disease management. Front Plant Sci 2024, 15. [CrossRef]
- Ploetz, R.C. Fusarium wilt of banana. Phytopathology 2015, 105, 1512-1521. [CrossRef]
- O’Neill, W.T.; Henderson, J.; Pattemore, J.A.; O’Dwyer, C.; Perry, S.; Beasley, D.R.; Tan, Y.P.; Smyth, A.; Goosem, C.; Thomson, K. Detection of Fusarium oxysporum f. sp. cubense tropical race 4 strain in northern Queensland. Australas Plant Dis Notes 2016, 11, 1-3. [CrossRef]
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti, E.S.G.; Staver, C.P. Fusarium wilt of banana: current knowledge on epidemiology and research needs toward sustainable disease management. Front Plant Sci 2018, 9. [CrossRef]
- Dale, J.; James, A.; Paul, J.Y.; Khanna, H.; Smith, M.; Peraza-Echeverria, S.; Garcia-Bastidas, F.; Kema, G.; Waterhouse, P.; Mengersen, K. Transgenic Cavendish bananas with resistance to Fusarium wilt tropical race 4. Nat Commun 2017, 8, 1496. [CrossRef]
- Chen, A.; Sun, J.; Martin, G.; Gray, L.-A.; Hřibová, E.; Christelová, P.; Yahiaoui, N.; Rounsley, S.; Lyons, R.; Batley, J. et al. Identification of a major QTL-controlling resistance to the Subtropical Race 4 of Fusarium oxysporum f. sp. cubense in Musa acuminata ssp. malaccensis. Pathogens 2023, 12, 289. [CrossRef]
- Chen, A.; Sun, J.; Viljoen, A.; Mostert, D.; Xie, Y.; Mangila, L.; Bothma, S.; Lyons, R.; Hřibová, E.; Christelová, P. et al. Genetic mapping, candidate gene identification and marker validation for host plant resistance to the race 4 of Fusarium oxysporum f. sp. cubense using Musa acuminata ssp. malaccensis. Pathogens 2023, 12, 820. [CrossRef]
- Rebouças, T.A.; de Jesus Rocha, A.; Cerqueira, T.S.; Adorno, P.R.; Barreto, R.Q.; dos Santos Ferreira, M.; Lino, L.S.M.; de Oliveira Amorim, V.B.; dos Santos-Serejo, J.A.; Haddad, F. Pre-selection of banana somaclones resistant to Fusarium oxysporum f. sp. cubense, subtropical race 4. Crop Prot 2021, 147, 105692. [CrossRef]
- Chen, A.; Sun, J.; Matthews, A.; Armas-Egas, L.; Chen, N.; Hamill, S.; Mintoff, S.; Tran-Nguyen, L.T.; Batley, J.; Aitken, E.A. Assessing variations in host resistance to Fusarium oxysporum f. sp. cubense race 4 in Musa species, with a focus on the subtropical race 4. Front Microbiol 2019, 10, 1062. [CrossRef]
- Mintoff, S.J.; Nguyen, T.V.; Kelly, C.; Cullen, S.; Hearnden, M.; Williams, R.; Daniells, J.W.; Tran-Nguyen, L.T. Banana cultivar field screening for resistance to Fusarium oxysporum f. sp. cubense tropical race 4 in the Northern Territory. J Fungus 2021, 7, 627. [CrossRef]
- Zuo, C.; Deng, G.; Li, B.; Huo, H.; Li, C.; Hu, C.; Kuang, R.; Yang, Q.; Dong, T.; Sheng, O. Germplasm screening of Musa spp. for resistance to Fusarium oxysporum f. sp. cubense tropical race 4 (Foc TR4). Eur J Plant Pathol 2018, 151, 723-734. [CrossRef]
- Zhan, N.; Kuang, M.; He, W.; Deng, G.; Liu, S.; Li, C.; Roux, N.; Dita, M.; Yi, G.; Sheng, O. Evaluation of resistance of banana genotypes with AAB genome to Fusarium wilt Tropical Race 4 in China. J Fungus 2022, 8, 1274. [CrossRef]
- Molina, A.; Razo, S.; Sinohin, V.G.; Herradura, L. DA-BAR Terminal Report: Mitigating banana Fusarium wilt Tropical Race 4 through a farmer-participatory approach of developing disease management strategies; Bioversity International, 2018. [CrossRef]
- Viljoen, A.; Mostert, D.; Chiconela, T.; Beukes, I.; Fraser, C.; Dwyer, J.; Murray, H.; Amisse, J.; Matabuana, E.L.; Tazan, G. Occurrence and spread of the banana fungus Fusarium oxysporum f. sp. cubense TR4 in Mozambique. S Afr J Sci 2020, 116, 1-11. [CrossRef]
- Daly, A.; Walduck, G. Fusarium wilt of bananas (Panama disease):(Fusarium oxysporum f. sp. cubense). Agnote - Northern Territory of Australia, No.I51, pp. 1-5, Northern Territory, Australia, 2006.
- Smith, M.; Langdon, P.; Pegg, K.; Daniells, J. Growth, yield and Fusarium wilt resistance of six FHIA tetraploid bananas (Musa spp.) grown in the Australian subtropics. Sci Hortic 2014, 170, 176-181. [CrossRef]
- Sanya, L.N.; Sseguya, H.; Kyazze, F.B.; Diiro, G.M.; Nakazi, F. The role of variety attributes in the uptake of new hybrid bananas among smallholder rural farmers in central Uganda. Agriculture & Food Security 2020, 9, 1. [CrossRef]
- Ploetz, R.C. Fusarium wilt of banana is caused by several pathogens referred to as Fusarium oxysporum f. sp. cubense. Phytopathology 2006, 96, 653-656. [CrossRef]
- Gordon, T.R. Fusarium oxysporum and the Fusarium Wilt Syndrome. Annu Rev Phytopathol 2017, 55, 23-39. [CrossRef]
- Stover, R.H. Fusarial wilt (Panama Disease) of bananas and other Musa species. Commonwealth Mycological Institute, UK. pp. 117. 1962.
- Zhang, L.; Yuan, T.; Wang, Y.; Zhang, D.; Bai, T.; Xu, S.; Wang, Y.; Tang, W.; Zheng, S.J. Identification and evaluation of resistance to Fusarium oxysporum f. sp. cubense tropical race 4 in Musa acuminata Pahang. Euphytica 2018, 214, 106. [CrossRef]
- Warman, N.M.; Aitken, E.A.B. The Movement of Fusarium oxysporum f.sp. cubense (Sub-Tropical Race 4) in susceptible cultivars of banana. Front Plant Sci 2018, 9. [CrossRef]
- Bai, T.; Qin, M.; Li, X.; Fan, H.; Xu, S.; Zeng, L.; Zheng, S.J. An additional threat to ‘Cavendish’banana growers and traders: The infection of banana peduncles by Fusarium oxysporum f. sp. cubense Tropical Race 4 (Foc TR4). Plant Health Progress 2020, 21, 312-316. [CrossRef]
- Bani, M.; Pérez-De-Luque, A.; Rubiales, D.; Rispail, N. Physical and chemical barriers in root tissues contribute to quantitative resistance to Fusarium oxysporum f. sp. pisi in pea. Front Plant Sci 2018, 9. [CrossRef]
- Ouellette, G.; Baayen, R.; Simard, M.; Rioux, D. Ultrastructural and cytochemical study of colonization of xylem vessel elements of susceptible and resistant Dianthus caryophyllus by Fusarium oxysporum f. sp. dianthi. Canad J Bot 1999, 77, 644-663. [CrossRef]
- De Micco, V.; Balzano, A.; Wheeler, E.A.; Baas, P. Tyloses and gums: a review of structure, function and occurrence of vessel occlusions. IAWA J 2016, 37, 186-205.
- VanderMolen, G.E.; Beckman, C.H.; Rodehorst, E. The ultrastructure of tylose formation in resistant banana following inoculation with Fusarium oxysporum f.sp. cubense. Physiol Mol Plant Pathol 1987, 31, 185-200. [CrossRef]
- Li, C.; Chen, S.; Zuo, C.; Sun, Q.; Ye, Q.; Yi, G.; Huang, B. The use of GFP-transformed isolates to study infection of banana with Fusarium oxysporum f. sp. cubense race 4. Eur J Plant Pathol 2011, 131, 327-340. [CrossRef]
- Xiao, R.F.; Zhu, Y.J.; Li, Y.D.; Liu, B. Studies on vascular infection of Fusarium oxysporum f. sp. cubense race 4 in banana by field survey and green fluorescent protein reporter. Int J Plant Pathol 2013, 2, 44-51. [CrossRef]
- Anderson, J.; Aitken, E. Effect of in planta treatment of ‘Cavendish’ Banana with herbicides and fungicides on the colonisation and sporulation by Fusarium oxysporum f. sp. cubense Subtropical Race 4. J Fungus 2021, 7, 184.
- Ratnayake, K.; Joyce, D.C.; Webb, R.I. A convenient sample preparation protocol for scanning electron microscope examination of xylem-occluding bacterial biofilm on cut flowers and foliage. Sci Hortic 2012, 140, 12-18. [CrossRef]
- Rocha, A.d.J.; Soares, J.M.d.S.; Nascimento, F.d.S.; Rocha, A.d.S.; Amorim, V.B.O.d.; Ramos, A.P.d.S.; Ferreira, C.F.; Haddad, F.; Amorim, E.P. Molecular, histological and histochemical responses of banana cultivars challenged with Fusarium oxysporum f. sp. cubense with different levels of virulence. Plants 2022, 11, 2339. [CrossRef]
- Kashyap, A.; Planas-Marquès, M.; Capellades, M.; Valls, M.; Coll, N.S. Blocking intruders: inducible physico-chemical barriers against plant vascular wilt pathogens. J Exp Bot 2020, 72, 184-198. [CrossRef]
- Chen, A.; Morrison, S.; Gregson, A.; Le, D.P.; Urquhart, A.S.; Smith, L.J.; Aitken, E.A.; Gardiner, D.M. Fluorescently Tagged Verticillium dahliae to understand the infection process on cotton (Gossypium hirsutum) and weed plant species. Pathogens 2024, 13, 442. [CrossRef]
- Vallad, G.; Subbarao, K. Colonization of resistant and susceptible lettuce cultivars by a green fluorescent protein-tagged isolate of Verticillium dahliae. Phytopathology 2008, 98, 871-885. [CrossRef]
- Lagopodi, A.L.; Ram, A.F.; Lamers, G.E.; Punt, P.J.; Van den Hondel, C.A.; Lugtenberg, B.J.; Bloemberg, G.V. Novel aspects of tomato root colonization and infection by Fusarium oxysporum f. sp. radicis-lycopersici revealed by confocal laser scanning microscopic analysis using the green fluorescent protein as a marker. Mol Plant Microbe Interact 2002, 15, 172-179. [CrossRef]
- Yuan, H.; Ling, X.; Liu, T.; Chen, T.; Yang, Y.; Yao, S.; Zhang, B. Microscopic observations of strawberry plant colonization by a GFP-labelled strain of Fusarium oxysporum f. sp. fragariae. Canad J Plant Pathol 2014, 36, 501-508. [CrossRef]
- Pérez-Donoso, A.G.; Greve, L.C.; Walton, J.H.; Shackel, K.A.; Labavitch, J.M. Xylella fastidiosa infection and ethylene exposure result in xylem and water movement disruption in grapevine shoots. Plant Physiol 2007, 143, 1024-1036. [CrossRef]
- Sun, Q.; Rost, T.L.; Matthews, M.A. Pruning-induced tylose development in stems of current-year shoots of Vitis vinifera (Vitaceae). Am J Bot 2006, 93, 1567-1576. [CrossRef]
- Taylor, A.M.; Gartner, B.L.; Morrell, J.J. Heartwood formation and natural durability-a review. Wood Fiber Sci 2002, 34, 587-611.
- Dute, R.R.; Duncan, K.M.; Duke, B. Tyloses in abscission scars of loblolly pine. IAWA J 1999, 20, 67-74.
- Shi, J.; Mueller, W.; Beckman, C. Vessel occlusion and secretory activities of vessel contact cells in resistant or susceptible cotton plants infected with Fusarium oxysporum f. sp. vasinfectum. Physiol Mol Plant Pathol 1992, 40, 133-147. [CrossRef]
- Zhou, G.-D.; He, P.; Tian, L.; Xu, S.; Yang, B.; Liu, L.; Wang, Y.; Bai, T.; Li, X.; Li, S. Disentangling the resistant mechanism of Fusarium wilt TR4 interactions with different cultivars and its elicitor application. Front in Plant Sci 2023, 14, 1145837. [CrossRef]
- De Ascensao, A.R.; Dubery, I.A. Panama disease: cell wall reinforcement in banana roots in response to elicitors from Fusarium oxysporum f. sp. cubense Race Four. Phytopathology 2000, 90, 1173-1180. [CrossRef]
- Beckman, C.; Elgersma, D.; MacHardy, W. The localization of fusarial infections in the vascular tissue of single-dominant-gene resistant tomatoes. Phytopathology 1972, 62, 1256–60.
- Baayen, R.; Van Eijk, C.; Elgersma, D. Histology of roots of resistant and susceptible carnation cultivars from soil infested with Fusarium oxysporum f. sp. dianthi. Neth J Plant Pathol 1989, 95, 3-13. [CrossRef]
- Tessier, B.; Mueller, W.; Morgham, A. Histopathology and ultrastructure of vascular responses in peas resistant or susceptible to Fusarium oxysporum f. sp. pisi. Phytopathology 1990, 80, 756-764.
- Pereira, A.C.; Cruz, M.F.A.; Paula Júnior, T.J.; Rodrigues, F.A.; Carneiro, J.E.S.; Vieira, R.F.; Carneiro, P.C.S. Infection process of Fusarium oxysporum f. sp. phaseoli on resistant, intermediate and susceptible bean cultivars. Trop Plant Pathol 2013, 38, 323-328. [CrossRef]
Figure 1.
Localisation of GFP-Foc-STR4 in the root systems of 'FHIA02', 'FHIA25', 'Williams', 'GCTCV-119' and 'Lady Finger' at the onset of infection, 5 to 9 dpi. (A) Macroconidia and microconidia of GFP-Foc-STR4 observed under a confocal microscope. (B) 'FHIA02' at 5 dpi, showing mycelia moving in the xylem of a lateral root, in the presence of microconidia. (C) 'FHIA02' at 5dpi, and with GFP tagged spores and mycelia clearly visualised between cortical cells of the elongation zone and the xylem vessels of the primary root. (D) 'FHIA25' at 5 dpi, strong fluorescence was detected in the xylem parenchyma cells and tracheid of the elongation zone of the fine root. A single mycelium confined to the xylem and multiple microconidia observed in the adjoining xylem vessel. Strong discoloration of parenchyma cells and vascular cylinder was also observed. (E) 'Williams' at 5 dpi, Mycelia networks visualised along the surface of the epidermis of a lateral root and in the cortex region adjacent to a xylem vessel. (F) 'Williams' at 9 dpi, an abundance of microconidia was visualised proliferating in the xylem vessels of a lateral root. (G) 'GCTCV119' at 5 dpi, mycelial networks were detected in the xylem vessels of a lateral root. (H) 'GCTCV119' at 7 dpi, a germ tube clearly visible from spores attempting to penetrate the epidermis of a lateral root. (I) 'GCTCV119' at 7 dpi, movement of GFP tagged mycelia detected within the vasculature of a lateral root. (J) 'Lady Finger' at 8 dpi, GFP mycelia networks were detected in the xylem vessels and the cortex region of a lateral root. (K) A longitudinal section of a lateral root of a non-inoculated 'FHIA02' plant at 5 dpi. (L) A longitudinal section of a lateral root of a non-inoculated 'FHIA25' plant at 6 dpi. Abbreviations are annotated as F2 = 'FHIA02'; F25 = 'FHIA25'; G = 'GCTCV119'; LF = 'Lady Finger'; C = non-inoculated control; ma = macroconidia; c = conidia; h = hyphae; m = mycelium; gt = germ tube. Horizontal bars indicate the scale used to capture the confocal images.
Figure 1.
Localisation of GFP-Foc-STR4 in the root systems of 'FHIA02', 'FHIA25', 'Williams', 'GCTCV-119' and 'Lady Finger' at the onset of infection, 5 to 9 dpi. (A) Macroconidia and microconidia of GFP-Foc-STR4 observed under a confocal microscope. (B) 'FHIA02' at 5 dpi, showing mycelia moving in the xylem of a lateral root, in the presence of microconidia. (C) 'FHIA02' at 5dpi, and with GFP tagged spores and mycelia clearly visualised between cortical cells of the elongation zone and the xylem vessels of the primary root. (D) 'FHIA25' at 5 dpi, strong fluorescence was detected in the xylem parenchyma cells and tracheid of the elongation zone of the fine root. A single mycelium confined to the xylem and multiple microconidia observed in the adjoining xylem vessel. Strong discoloration of parenchyma cells and vascular cylinder was also observed. (E) 'Williams' at 5 dpi, Mycelia networks visualised along the surface of the epidermis of a lateral root and in the cortex region adjacent to a xylem vessel. (F) 'Williams' at 9 dpi, an abundance of microconidia was visualised proliferating in the xylem vessels of a lateral root. (G) 'GCTCV119' at 5 dpi, mycelial networks were detected in the xylem vessels of a lateral root. (H) 'GCTCV119' at 7 dpi, a germ tube clearly visible from spores attempting to penetrate the epidermis of a lateral root. (I) 'GCTCV119' at 7 dpi, movement of GFP tagged mycelia detected within the vasculature of a lateral root. (J) 'Lady Finger' at 8 dpi, GFP mycelia networks were detected in the xylem vessels and the cortex region of a lateral root. (K) A longitudinal section of a lateral root of a non-inoculated 'FHIA02' plant at 5 dpi. (L) A longitudinal section of a lateral root of a non-inoculated 'FHIA25' plant at 6 dpi. Abbreviations are annotated as F2 = 'FHIA02'; F25 = 'FHIA25'; G = 'GCTCV119'; LF = 'Lady Finger'; C = non-inoculated control; ma = macroconidia; c = conidia; h = hyphae; m = mycelium; gt = germ tube. Horizontal bars indicate the scale used to capture the confocal images.
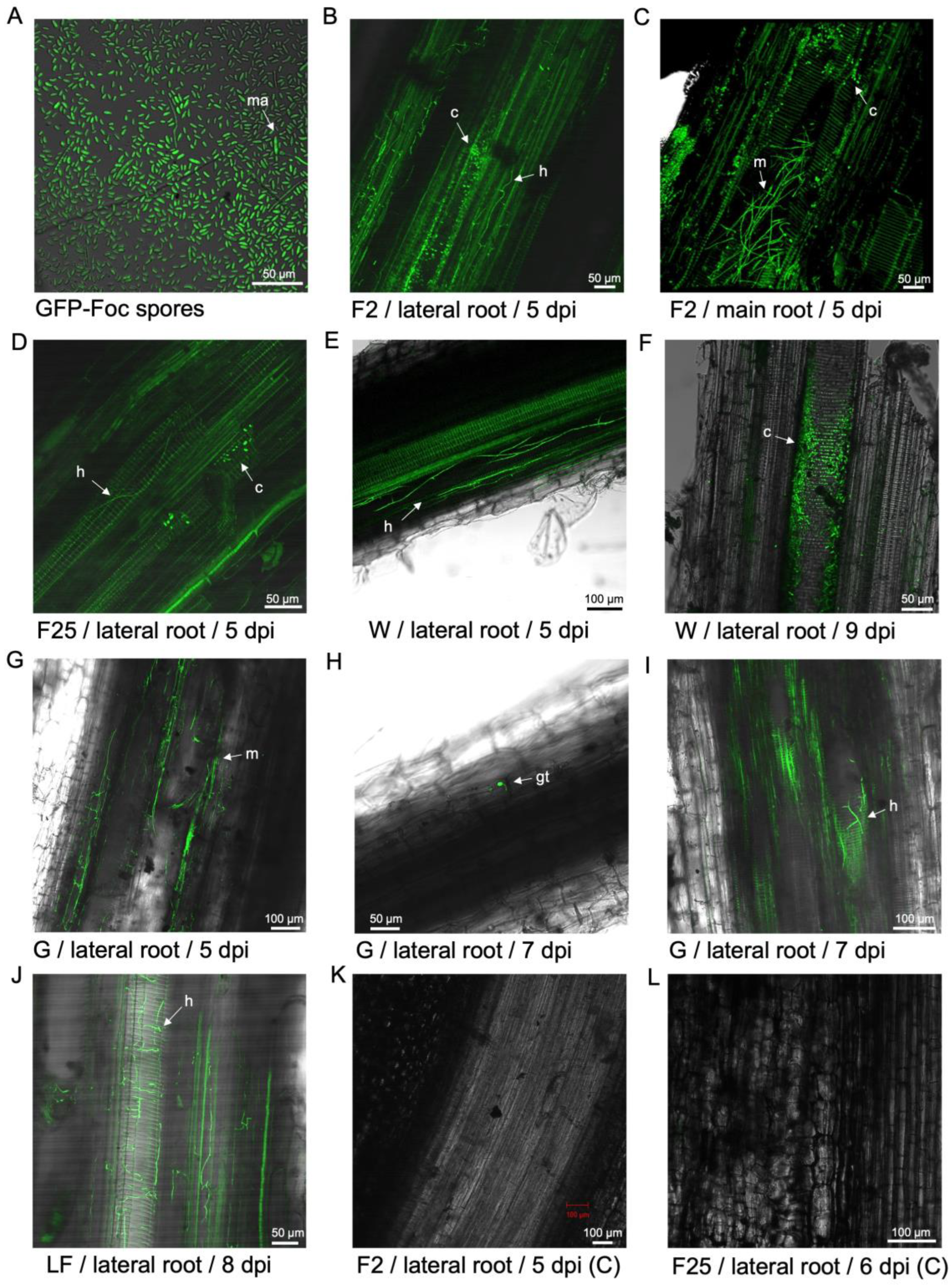
Figure 2.
Localisation of GFP-Foc-STR4 in the susceptible cultivar 'FHIA02'. (A) Microconidia and mycelia moving inter- and intracellularly in the cortex near the xylem vessels of a fine root at 14 dpi. (B) Germinating microconidia, chlamydospores and mycelia present in the epidermis of a lateral root at 14 dpi. (C) Mycelia moving intercellularly in the cortex region of a lateral root. Microconidia produced in false heads on a long monophialide were observed. (D) A wound site was penetrated by the fungus on a lateral root and its movement into the adjacent xylem vessels at 19 dpi. (E) Spore gemination and mycelia colonising the xylem vessels of a fine root at 26 dpi. (F) Spore germination and mycelia movement in the xylem vessels of a primary root at 29 dpi. (G) Spore germination in the xylem of a primary root near a rhizome node, in a region with red discolouration. (H) The movement of mycelia was detected in the cortex region of a main root at 41 dpi. (I) Spore germination and localised mycelia movement in the xylem vessels of the corm joining the main root. (J) Xylem vessels completely colonised by the fungus at 42 dpi. False heads on monophialides were observed. (K) Non-inoculated control of a fine root at 18 dpi. (L) Non-inoculated control of a fine root at 62 dpi. Abbreviations are annotated as F2 = 'FHIA02'; C = non-inoculated control; ma = macroconidia; ch = chlamydospores; c = conidia; h = hyphae; m = mycelium; gt = germ tube. Horizontal bars indicate the scale used to capture the confocal images.
Figure 2.
Localisation of GFP-Foc-STR4 in the susceptible cultivar 'FHIA02'. (A) Microconidia and mycelia moving inter- and intracellularly in the cortex near the xylem vessels of a fine root at 14 dpi. (B) Germinating microconidia, chlamydospores and mycelia present in the epidermis of a lateral root at 14 dpi. (C) Mycelia moving intercellularly in the cortex region of a lateral root. Microconidia produced in false heads on a long monophialide were observed. (D) A wound site was penetrated by the fungus on a lateral root and its movement into the adjacent xylem vessels at 19 dpi. (E) Spore gemination and mycelia colonising the xylem vessels of a fine root at 26 dpi. (F) Spore germination and mycelia movement in the xylem vessels of a primary root at 29 dpi. (G) Spore germination in the xylem of a primary root near a rhizome node, in a region with red discolouration. (H) The movement of mycelia was detected in the cortex region of a main root at 41 dpi. (I) Spore germination and localised mycelia movement in the xylem vessels of the corm joining the main root. (J) Xylem vessels completely colonised by the fungus at 42 dpi. False heads on monophialides were observed. (K) Non-inoculated control of a fine root at 18 dpi. (L) Non-inoculated control of a fine root at 62 dpi. Abbreviations are annotated as F2 = 'FHIA02'; C = non-inoculated control; ma = macroconidia; ch = chlamydospores; c = conidia; h = hyphae; m = mycelium; gt = germ tube. Horizontal bars indicate the scale used to capture the confocal images.

Figure 3.
Localisation of GFP-Foc-STR4 observed in the rhizome of 'FHIA02' during the infection process. (A) Germ tubes of chlamydospores and hyphae moving through cortical cells of a discoloured rhizome at 14 dpi. (B) A discoloured region of a rhizome was associated with a light fluorescence, with chlamydospores and hyphae moving through cortical cells at 14 dpi. (C) Strong fluorescence signals, along with mycelia and microconidia detected in the cortex and the vascular bundles at 14 dpi. (D) Substantial presence of microconidia, germ tubes, hyphae and chlamydospores detected in a vascular bundle at 36 dpi. (E) Extensive mycelia networks established in the xylem vessels at 36 dpi. (F) Tyloses were detected occluding the xylem vessel lumen of a rhizome node, with microconidia and hyphae clearly present in this region at 68 dpi. (G) Two xylem vessels fully occluded by mature tyloses, while microconidia, chlamydospores and mycelia colonised the vessel lumen and along the boundaries of occluding tyloses. (H) Non-inoculated rhizome at 14 dpi. (I) Non-inoculated rhizome showing weak autofluorescence in the vascular regions at 68 dpi. Abbreviations are annotated as F2 = 'FHIA02'; C = non-inoculated control; ch = chlamydospores; c = conidia; h = hyphae; m = mycelium; gt = germ tube; ty = tyloses. Horizontal bars indicate the scale used to capture the confocal images.
Figure 3.
Localisation of GFP-Foc-STR4 observed in the rhizome of 'FHIA02' during the infection process. (A) Germ tubes of chlamydospores and hyphae moving through cortical cells of a discoloured rhizome at 14 dpi. (B) A discoloured region of a rhizome was associated with a light fluorescence, with chlamydospores and hyphae moving through cortical cells at 14 dpi. (C) Strong fluorescence signals, along with mycelia and microconidia detected in the cortex and the vascular bundles at 14 dpi. (D) Substantial presence of microconidia, germ tubes, hyphae and chlamydospores detected in a vascular bundle at 36 dpi. (E) Extensive mycelia networks established in the xylem vessels at 36 dpi. (F) Tyloses were detected occluding the xylem vessel lumen of a rhizome node, with microconidia and hyphae clearly present in this region at 68 dpi. (G) Two xylem vessels fully occluded by mature tyloses, while microconidia, chlamydospores and mycelia colonised the vessel lumen and along the boundaries of occluding tyloses. (H) Non-inoculated rhizome at 14 dpi. (I) Non-inoculated rhizome showing weak autofluorescence in the vascular regions at 68 dpi. Abbreviations are annotated as F2 = 'FHIA02'; C = non-inoculated control; ch = chlamydospores; c = conidia; h = hyphae; m = mycelium; gt = germ tube; ty = tyloses. Horizontal bars indicate the scale used to capture the confocal images.
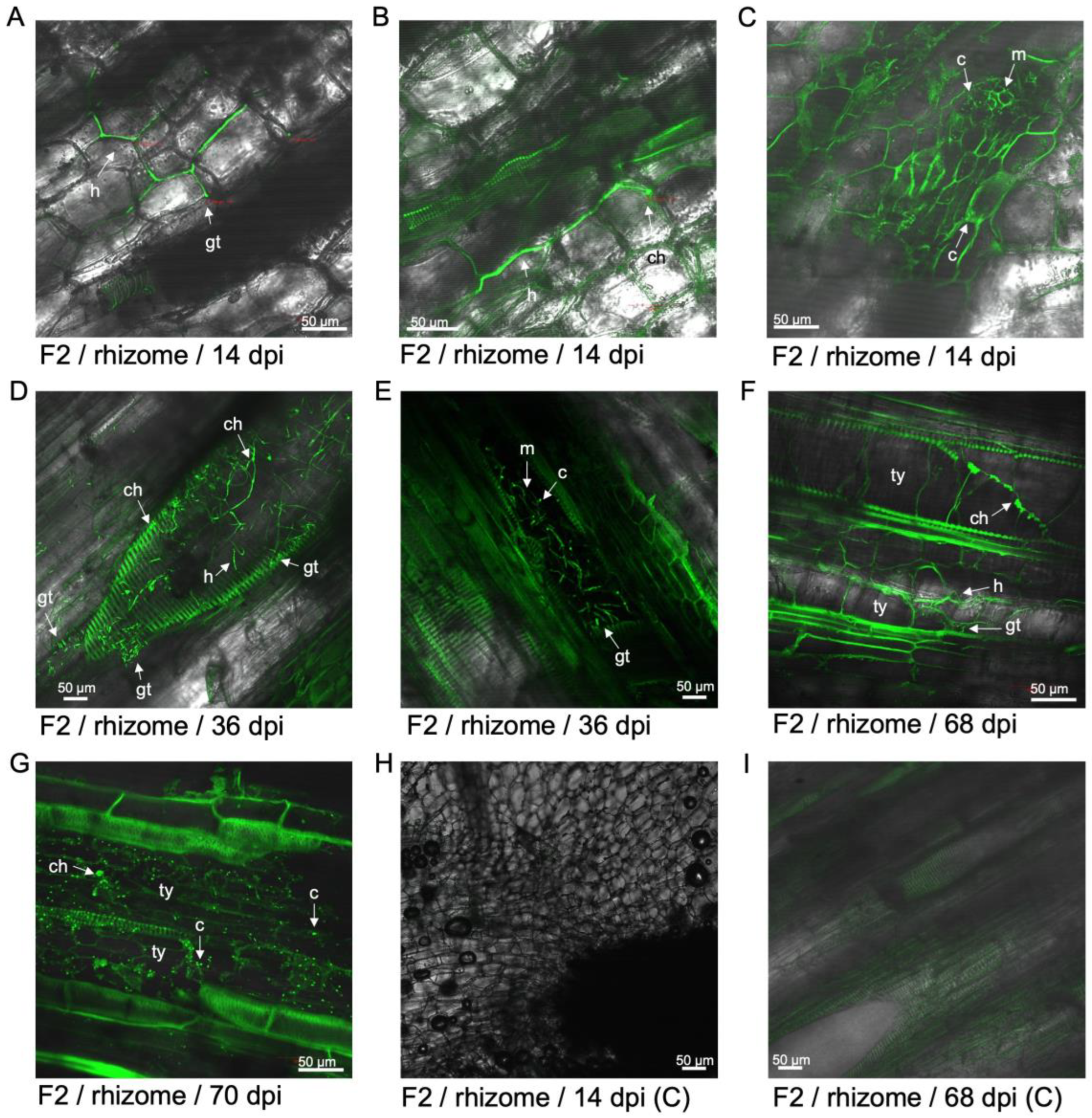
Figure 4.
Localisation of GFP-Foc-STR4 observed in the roots of 'FHIA25' during the infection process. (A) Microconidia and hyphae moving through the epidermis of fine roots in an intercellular manner at 12 dpi. (B) Spore germination and hyphae movement in the tracheid and xylem vessel of fine roots at 12 dpi. (C) Germinating spores and hyphae moving through the xylem vessels of fine roots at 21 dpi. (D) The presence of spores and hyphae detected in the xylem vessels in a lateral root to corm joint at 26 dpi. (E) GFP-Foc-STR4 completely colonised the epidermis of a primary root tip at 29 dpi. (F) Hyphae and germinating spores moving through the xylem vessels of fine roots at 29 dpi. (G) Hyphae and a single terminal chlamydospore detected in the vasculature of a fine root at 36 dpi. (H) GFP fluorescence localised in the vasculature with individual hyphae moving through the cortex. (I) GFP fluorescence associated with hyphae in both the cortex and the vasculature of a root to corm joint at 49 dpi. (J) Non-inoculated lateral roots at 21 dpi. (K) Non-inoculated lateral roots at 36 dpi. (L) Non-inoculated lateral root to corm joint at 51 dpi. Abbreviations are annotated as F25 = 'FHIA25'; C = non-inoculated control; ch = chlamydospores; c = conidia; h = hyphae; m = mycelium; gt = germ tube. Horizontal bars indicate the scale used to capture the confocal images.
Figure 4.
Localisation of GFP-Foc-STR4 observed in the roots of 'FHIA25' during the infection process. (A) Microconidia and hyphae moving through the epidermis of fine roots in an intercellular manner at 12 dpi. (B) Spore germination and hyphae movement in the tracheid and xylem vessel of fine roots at 12 dpi. (C) Germinating spores and hyphae moving through the xylem vessels of fine roots at 21 dpi. (D) The presence of spores and hyphae detected in the xylem vessels in a lateral root to corm joint at 26 dpi. (E) GFP-Foc-STR4 completely colonised the epidermis of a primary root tip at 29 dpi. (F) Hyphae and germinating spores moving through the xylem vessels of fine roots at 29 dpi. (G) Hyphae and a single terminal chlamydospore detected in the vasculature of a fine root at 36 dpi. (H) GFP fluorescence localised in the vasculature with individual hyphae moving through the cortex. (I) GFP fluorescence associated with hyphae in both the cortex and the vasculature of a root to corm joint at 49 dpi. (J) Non-inoculated lateral roots at 21 dpi. (K) Non-inoculated lateral roots at 36 dpi. (L) Non-inoculated lateral root to corm joint at 51 dpi. Abbreviations are annotated as F25 = 'FHIA25'; C = non-inoculated control; ch = chlamydospores; c = conidia; h = hyphae; m = mycelium; gt = germ tube. Horizontal bars indicate the scale used to capture the confocal images.

Figure 5.
Examination of GFP-Foc-STR4 in the rhizome of 'FHIA25' during the infection process. (A) Strong fluorescence associated with the rhizome cortex and vascular bundles at 14 dpi. (B) Hyphae was detected in the rhizome cortex at 14 dpi. (C) Individual hyphae were visualised in the vasculature of a rhizome node, with a strong GFP fluorescence signal detected in this region at 36 dpi. (D) Tyloses were visualised in the vascular bundles at 36 dpi. Microconidia were also present. (E) Tyloses were visualised in the xylem vessels of a corm node at 41 dpi. Strong fluorescence in the vasculature and the presence of hyphae were detected. (F) Mycelia networks were visualised in the rhizome node to a primary root at 41 dpi. (G) Tyloses were visualised to occlude a xylem vessel in the rhizome node joining a primary root at 42 dpi. (H) Tyloses were visualised to occlude a xylem vessel in the rhizome node joining a primary root at 62 dpi. (I) Tyloses were visualised to occlude multiple xylem vessels in the rhizome node joining a primary root at 70 dpi. (J) Non-inoculated rhizome at 16 dpi. Weak autofluorescence in the vasculature was detected. (K) Non-inoculated rhizome at 36 dpi. (L) Non-inoculated rhizome at 70 dpi. Abbreviations are annotated as F25 = 'FHIA25'; C = non-inoculated control; c = conidia; h = hyphae; m = mycelium; gt = germ tube; ty = tyloses. Horizontal bars indicate the scale used to capture the confocal images.
Figure 5.
Examination of GFP-Foc-STR4 in the rhizome of 'FHIA25' during the infection process. (A) Strong fluorescence associated with the rhizome cortex and vascular bundles at 14 dpi. (B) Hyphae was detected in the rhizome cortex at 14 dpi. (C) Individual hyphae were visualised in the vasculature of a rhizome node, with a strong GFP fluorescence signal detected in this region at 36 dpi. (D) Tyloses were visualised in the vascular bundles at 36 dpi. Microconidia were also present. (E) Tyloses were visualised in the xylem vessels of a corm node at 41 dpi. Strong fluorescence in the vasculature and the presence of hyphae were detected. (F) Mycelia networks were visualised in the rhizome node to a primary root at 41 dpi. (G) Tyloses were visualised to occlude a xylem vessel in the rhizome node joining a primary root at 42 dpi. (H) Tyloses were visualised to occlude a xylem vessel in the rhizome node joining a primary root at 62 dpi. (I) Tyloses were visualised to occlude multiple xylem vessels in the rhizome node joining a primary root at 70 dpi. (J) Non-inoculated rhizome at 16 dpi. Weak autofluorescence in the vasculature was detected. (K) Non-inoculated rhizome at 36 dpi. (L) Non-inoculated rhizome at 70 dpi. Abbreviations are annotated as F25 = 'FHIA25'; C = non-inoculated control; c = conidia; h = hyphae; m = mycelium; gt = germ tube; ty = tyloses. Horizontal bars indicate the scale used to capture the confocal images.

Figure 6.
Localisation of GFP-Foc-STR4 through the outer leaf sheaths of 'FHIA02' and 'FHIA25'. (A) Microconidia colonising the outer leaf sheaths of 'FHIA02' at 14 dpi. (B) Hyphae moving through xylem vessels of the outer leaf sheaths of 'FHIA02' at 26 dpi. (C) Hyphae moving through the epidermis of outer leaf sheaths of 'FHIA02' at 29 dpi. (D) Edge of a petiole of a senescing showing proliferation of microconidia, hyphae and chlamydospores at 54 dpi. (E) Chlamydospores visualised at the edge of a petiole of a senescing leaf in 'FHIA02' at 62 dpi. (F) Non-inoculated edge of a petiole at 63 dpi. (G) Hyphae and chlamydospore confined to the vasculature of the xylem vessels in the outer leaf sheath of 'FHIA25' at 16 dpi. (H) Hyphae confined to the vasculature in the outer leaf sheath of 'FHIA25' at 21 dpi. (I) Mycelial networks visualised near the vasculatures of a leaf of 'FHIA25' at 29 dpi. (J) Chlamydospores visualised in a discoloured region at the edge of a petiole at the 62 dpi. (K) Chlamydospores and hyphae visualised in the vasculature and surrounding mesophyll cells at the edge of a petiole in 'FHIA25' at 70 dpi. (L) Non-inoculated midrib of a 'FHIA25' leaf at 70 dpi. Abbreviations are annotated as F2 = 'FHIA02'; F25 = 'FHIA25'; C = non-inoculated control; ma = macroconidia; ch = chlamydospores; c = conidia; h = hyphae; m = mycelium; gt = germ tube. Horizontal bars indicate the scale used to capture the confocal images.
Figure 6.
Localisation of GFP-Foc-STR4 through the outer leaf sheaths of 'FHIA02' and 'FHIA25'. (A) Microconidia colonising the outer leaf sheaths of 'FHIA02' at 14 dpi. (B) Hyphae moving through xylem vessels of the outer leaf sheaths of 'FHIA02' at 26 dpi. (C) Hyphae moving through the epidermis of outer leaf sheaths of 'FHIA02' at 29 dpi. (D) Edge of a petiole of a senescing showing proliferation of microconidia, hyphae and chlamydospores at 54 dpi. (E) Chlamydospores visualised at the edge of a petiole of a senescing leaf in 'FHIA02' at 62 dpi. (F) Non-inoculated edge of a petiole at 63 dpi. (G) Hyphae and chlamydospore confined to the vasculature of the xylem vessels in the outer leaf sheath of 'FHIA25' at 16 dpi. (H) Hyphae confined to the vasculature in the outer leaf sheath of 'FHIA25' at 21 dpi. (I) Mycelial networks visualised near the vasculatures of a leaf of 'FHIA25' at 29 dpi. (J) Chlamydospores visualised in a discoloured region at the edge of a petiole at the 62 dpi. (K) Chlamydospores and hyphae visualised in the vasculature and surrounding mesophyll cells at the edge of a petiole in 'FHIA25' at 70 dpi. (L) Non-inoculated midrib of a 'FHIA25' leaf at 70 dpi. Abbreviations are annotated as F2 = 'FHIA02'; F25 = 'FHIA25'; C = non-inoculated control; ma = macroconidia; ch = chlamydospores; c = conidia; h = hyphae; m = mycelium; gt = germ tube. Horizontal bars indicate the scale used to capture the confocal images.

Figure 7.
Scanning electron micrographs of tyloses formation within transverse sectioned primary roots of 'FHIA02'. (A) A transverse section of non-inoculated 'FHIA02' roots at 14 dpi. (B) A magnified view of transverse sectioned roots of non-inoculated 'FHIA02' at 14 dpi. (C) Mycelia and vascular occlusion visualised in the vascular cavities of 'FHIA02' at 14 dpi. (D) Developing tyloses visualised in the vascular cavities of 'FHIA02' at 14 dpi. (E) Extensive mycelial networks visualised in the vascular cavities of 'FHIA02' at 14 dpi. (F) Mature and developing tyloses visualised in the vascular cavities of 'FHIA02' at 14 dpi. (G) Individual occluded xylem cavities, developing tyloses and the presence of hyphae were visualised in the main roots of 'FHIA02' at 42 dpi. (H) Tyloses occluding xylem cavities were visualised in the main roots of 'FHIA02' at 42 dpi. (I) Mature tyloses occluding multiple xylem cavities were visualised in the main roots of 'FHIA02' at 42 dpi. Abbreviations are annotated as F2 = 'FHIA02'; C = non-inoculated control; m = mycelium; ty = tyloses; o = vascular occlusion. Horizontal bars indicate the scale used to capture the confocal images.
Figure 7.
Scanning electron micrographs of tyloses formation within transverse sectioned primary roots of 'FHIA02'. (A) A transverse section of non-inoculated 'FHIA02' roots at 14 dpi. (B) A magnified view of transverse sectioned roots of non-inoculated 'FHIA02' at 14 dpi. (C) Mycelia and vascular occlusion visualised in the vascular cavities of 'FHIA02' at 14 dpi. (D) Developing tyloses visualised in the vascular cavities of 'FHIA02' at 14 dpi. (E) Extensive mycelial networks visualised in the vascular cavities of 'FHIA02' at 14 dpi. (F) Mature and developing tyloses visualised in the vascular cavities of 'FHIA02' at 14 dpi. (G) Individual occluded xylem cavities, developing tyloses and the presence of hyphae were visualised in the main roots of 'FHIA02' at 42 dpi. (H) Tyloses occluding xylem cavities were visualised in the main roots of 'FHIA02' at 42 dpi. (I) Mature tyloses occluding multiple xylem cavities were visualised in the main roots of 'FHIA02' at 42 dpi. Abbreviations are annotated as F2 = 'FHIA02'; C = non-inoculated control; m = mycelium; ty = tyloses; o = vascular occlusion. Horizontal bars indicate the scale used to capture the confocal images.

Figure 8.
Scanning electron micrographs of tyloses formation within transverse sectioned primary roots of 'FHIA25'. (A) Vascular bundles of a non-inoculated plant at 14 dpi. (B) A magnified view of the xylem vessels in the primary vascular bundle at 14 dpi. (C) Tyloses formation and mycelia confined to the xylem cavities in the vascular bundles at 14 dpi. (D) Movement of hyphae visualised in the xylem cavities of the vascular bundles at 14 dpi. (E) Tyloses formation and vascular occlusion visualised in the xylem cavities of the vascular bundles at 14 dpi. (F) A developing tylosis and mycelial networks visualised in the xylem cavities at 14 dpi. (G) Extensive mycelia networks visualised in the xylem vessels at 42 dpi. (H) Mature and developing tyloses visualised occluding the xylem cavities at 42 dpi. (I) Mature tyloses occluding multiple xylem cavities at 42 dpi. F25 = 'FHIA25'; C = non-inoculated control; m = mycelium; ty = tyloses; o = vascular occlusion. Horizontal bars indicate the scale used to capture the confocal images.
Figure 8.
Scanning electron micrographs of tyloses formation within transverse sectioned primary roots of 'FHIA25'. (A) Vascular bundles of a non-inoculated plant at 14 dpi. (B) A magnified view of the xylem vessels in the primary vascular bundle at 14 dpi. (C) Tyloses formation and mycelia confined to the xylem cavities in the vascular bundles at 14 dpi. (D) Movement of hyphae visualised in the xylem cavities of the vascular bundles at 14 dpi. (E) Tyloses formation and vascular occlusion visualised in the xylem cavities of the vascular bundles at 14 dpi. (F) A developing tylosis and mycelial networks visualised in the xylem cavities at 14 dpi. (G) Extensive mycelia networks visualised in the xylem vessels at 42 dpi. (H) Mature and developing tyloses visualised occluding the xylem cavities at 42 dpi. (I) Mature tyloses occluding multiple xylem cavities at 42 dpi. F25 = 'FHIA25'; C = non-inoculated control; m = mycelium; ty = tyloses; o = vascular occlusion. Horizontal bars indicate the scale used to capture the confocal images.

Figure 9.
Tyloses formation and GFP-Foc-STR4 movement in the roots and corm of 'GCTCV119' and 'Williams'. (A) Tyloses visualised in the xylem vessels of a non-inoculated 'GCTCV119' plant at 56 dpi. (B) Movement of hyphae through tyloses was visualised at 56 dpi. (C) Tyloses visualised in the xylem vessels with hyphae present in the same space at 59 dpi. (D) Hyphae confined to the spaces between two xylem vessels in the primary root of 'Williams' at 56 dpi. (E) Hyphae moving in between tyloses in the vasculature of the main root of 'Williams' at 56 dpi. (F) Hyphae moving through the spaces between individual tyloses in the vasculature of 'Williams' at 55 dpi. Abbreviations are annotated as G = 'GCTCV119'; W = 'Williams'; C = non-inoculated control; h = hyphae; m = mycelium; ty = tyloses. Horizontal bars indicate the scale used to capture the confocal images.
Figure 9.
Tyloses formation and GFP-Foc-STR4 movement in the roots and corm of 'GCTCV119' and 'Williams'. (A) Tyloses visualised in the xylem vessels of a non-inoculated 'GCTCV119' plant at 56 dpi. (B) Movement of hyphae through tyloses was visualised at 56 dpi. (C) Tyloses visualised in the xylem vessels with hyphae present in the same space at 59 dpi. (D) Hyphae confined to the spaces between two xylem vessels in the primary root of 'Williams' at 56 dpi. (E) Hyphae moving in between tyloses in the vasculature of the main root of 'Williams' at 56 dpi. (F) Hyphae moving through the spaces between individual tyloses in the vasculature of 'Williams' at 55 dpi. Abbreviations are annotated as G = 'GCTCV119'; W = 'Williams'; C = non-inoculated control; h = hyphae; m = mycelium; ty = tyloses. Horizontal bars indicate the scale used to capture the confocal images.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



