
Submitted:
28 October 2024
Posted:
28 October 2024
You are already at the latest version
Abstract
Epicauta gorhami is a hypermetamorphic insect that mainly forage soybeans during the adult stage. However, the lack of appropriate references hinders our studying of the gene function in E. gorhami. In this study, refer to five computational tools (Ct value, geNorm, NormFinder, BestKeeper and RefFinder), the stability of 10 housekeeping genes (GAPDH, ACT, RPL4, RPL27, α-TUB, RPS18, EF1α, RPS28, RPL13 and SOD) was assessed by RT-qPCR under three different conditions (adult ages, tissues/organs and temperatures). The findings suggested that SOD and RPS18 were the most ideal references for examine gene transcripts among diverse adult ages and at various temperatures; A pair of RPS18 and RPS28 was the most reliable genes to assess gene expressions in diverse adult tissues. Finally, The relative expression levels of EgUAP were computed after normalization RPS18 and RPS28 with across diverse adult tissues. As expected, EgUAP expression was abundant in the foregut, trachea and antenna, and low in the midgut, hindgut and epidermis. These findings will lay a solid foundation for analysing the gene expression of E. gorhami.
Keywords:
Epicauta gorhami
; Glycine max
; reference genes
; RT-qPCR
; RefFinder
1. Introduction
The bean blister beetle, Epicauta gorhami (Coleoptera: Meloidae) is a hypermetamorphic insect that can forage soybean (Glycine max) during the adult stage [1,2,3,4]. It poses potential threats to soybean production, which is an crucial oilseed, protein and biodiesel crop [5,6]. The adults of this specie mainly feed on leaves of G. max [1,2,3,4]. Previous studies on E. gorhami have traditionally placed a strong emphasis on diapause [1,3], seasonal adaptation [2] and the phylogenetic relationship [4]. However, there is currently no one research on gene functions in E. gorhami. In order to further explore molecular mechanisms of target genes, it is imperative to screening the suitable reference genes.
The reverse tanscriptase-quantitative polymerase chain reaction (qRT-PCR) has amounts of advantages of high sensitivity, specificity and accuracy, which is widely employed to analyse the target gene transcripts [7,8]. For measuring accurately the mRNA levels of target genes by qRT-PCR, selecting the most optimal references to conduct normalization is essential [9,10]. If unstable reference genes are employed, the results of gene transcripts will be inaccurate [8,9]. Hence, selecting appropriate reference genes should be assessed under various experimental conditions [11,12,13].
As usual, housekeeping genes (HKGs), such as actin (ACT), ribosomal protein (RPL and RPS), and elongation factor 1α (EF1α), are widely employed as references in insect species, due to relative stable expression levels. However, the stability of internal genes may not always constant under diverse experimental treatments [13,14]. In the potato ladybird Henosepilachna vigintioctomaculata, RPL6 and RPL13 are the suitable references in different developmental stages, whereas RPS18 and EF1α are more optimal among diverse tissues [13]. In the sugarcane stem borer Chilo sacchariphagus, β-ACT and RPL7 show the best stable expression under different tissues, whereas EF1α and SDHA exhibit stability between sexes [14]. Hence, to evaluating reference genes at different backgrounds is necessary.
Currently, it has made important advances in the study of reference genes selection in numerous insect species, such as Mylabris sibirica (RPL6 and RPL13) [11], Phthorimaea operculella (EF1α and RPL13) [12], H. vigintioctomaculata (RPS18 and RPL13) [13] and C. sacchariphagus (β-ACT and RPL7 across various tissues and at distinct temperatures, EF1α and SDHA between sexes) [14]. To sum up, each experimental condition need at least two reference genes to measure the target gene transcripts in insect species.
In the paper, we selected 10 potential reference genes from the transcriptomes of E. gorhami, which were ACT, EF1α, GAPDH (glyceraldehyde-3-phosphate dehydrogenase), α-TUB (α-tubulin), RPL4, RPL13, RPL27, RPS18, RPS28 and SOD (superoxide dismutase). The stablity of these genes was assessed among diverse adult ages, across different adult tissues and in various temperatures in E. gorhami. In addition, the expression pattern of E. gorhami uridine diphosphate (UDP)-N-acetylglucosamine-pyrophosphorylase (EgUAP) across different adult tissues was used to evaluate the results. These findings will lay an vital basis for further research on gene functions in E. gorhami.
2. Materials and Methods
2.1. Insect Rearing and Treatment Methods
E. gorhami adults used in the study were obtained in August 2024 from G. max plants in Enshi Tujia and Miao Autonomous Prefecture, Hubei Province, China (coordinates:
N30°19'43", E109°38'48"E). The adults were fed under suitable conditions (25±2 °C, 16h:8h photoperiod, and 50 ∼ 60% relative humidity) in the insectary for one week.
Adult ages: Four newly emerged adults (two males and two females) were sampled every 1 day for 3 day, as one biological replicate. Three biological replicates were conducted.
Adult tissues: These tissues (foregut, midgut, hindgut, antenna, trachea and epidermis) were collected from the 10 healthy 5-day-old adults (5 males and 5 females) in three biological replicates.
Temperature: Four newly emerged adults (2 males and 2 females) were treated under three temperature ( 4 ◦C, 25 ◦C and 37 ◦C for 6 h, respectively), as one biological replicate. Three biological replicates were conducted.
2.2. Total RNA Isolation and cDNA Synthesis
Total RNA of each sample was extracted by TRIzol reagent (YiFeiXue Tech, Nanjing, China), refer to the manufacturer's instructions. The RNA purity and concentration were detected by the NanoDrop ND-1000 spectrophotometer Subsequently, cDNA synthesis was manipulated using the HiScript III RT SuperMix (Vazyme Biotech Co.,Ltd, Nanjing, China).
2.3. Selection of Candidate References and Primer Design
According to the transcriptome of E. gorhami, a total of 10 candidate references were selected, including EF1α, ACT, RPL4, α-TUB,RPL13, RPL27, RPS18, GAPDH, RPS28 and SOD, which are commonly utilized as reference genes in other insects [15]. The information of references was displayed in Table S1.
Primer design was executed by the Primer Premier 5.0 software, based on the RT-PCR primer design principles. The resultant sequences were uploaded to GenBank, with the accession numbers of PQ497541-PQ497541 (Table S1).
2.4. Quantitative Real-Time PCR (qRT-PCR)
The primers for qRT-PCR were performed by Primer3web version 4.1.0 (https://primer3.ut.ee/), and were located in Table 1. qRT-PCR experiment was executed by CFX96 Real-Time System (Bio-Rad Laboratories, Hercules, CA, USA) and ChamQ Universal SYBR qPCR Master Mix (Vazyme Biotech co.,ltd). Each reaction consisted of a final volume 20 μL, containing 10 μL of 2× ChamQ Universal SYBR qPCR Master Mix, 0.4 μL of reverse primer (10 μM), 1 μL cDNA template and 8.2 μL of RNase Free water. The thermocycling procedure was an initial step of 95°C for 30 s, followed by 40 cycles of 95°C for 5 s, annealing at 60°C for 34 s. After the reaction, a melting curve analysis was one cycle of 95 ℃ for 15 s, 60 ℃ for 60 s and 95 ℃ for 1 s, which was used to confirm the specificity of the amplified product. The amplification efficiency (E) was computed using a series of 3-fold dilutions of cDNA template refer to the equation: E = (10[−1/slope] − 1) × 100% [16]. All experiments were conducted in triplicate.
2.5. Assessment of Reference Gene Stability
The raw Ct values were required by the BioRadCFXManager software. The stability of 10 reference genes under different backgrounds were assessed using a range of algorithms, including BestKeeper [17], Normfinder [18], geNorm [9] and the ΔCt method [19]. Finally, the RefFinder [20,21] was applied to analyse the comprehensive ranking of each experimental condition.
2.6. Validation of Selected Reference Genes Under Diverse Adult Tissues
The transcription level of uridine diphosphate (UDP)-N-acetylglucosamine-pyrophosphorylase (UAP, GenBank: PQ497551) in E. gorhami was utilized to assess the stability of selected reference genes across various adult tissues. The primer sequence of UAP was as follows:Forward: CGCTACAACGTAACGCCATC, Reverse: CCCCACAATCGCTACGTTTC. The relative transcription level of EgUAP were computed under adult tissues by the 2-∆∆Ct method, based on RPS18 and RPS28. The SPSS Statistics 29 software package was then used to evaluate the variance in expression levels of EgUAP among different adult tissues through one-way ANOVA analysis followed by Tukey’s test for multiple comparisons.
3. Results
3.1. Selection of Candidate Reference Genes
Ten commonly used reference genes (ACT, GAPDH, RPL4, EF1α,RPL13, RPL27, RPS18, RPS28, α-TUB and SOD) in E. gorhami were choosed. The resultant sequences were uploaded to GenBank, with the accession numbers of PQ497541-PQ497541 (Table S1).
The products of qRT-PCR were verified by sequencing. The melting curve analysis confirmed the specificities of each primer pair for qRT-PCR. As expected, the efficiency (E) of 10 primer pairs were between 95.50% (EF1α) and 102.73% (α-TUB), with the correlation coefficients (R2) varying from 0.993 to 0.999 (Table 1). These results demonstrated that efficiency of primers reached the standards of traditional qRT-PCR [22].
3.2. Expression Levels of Reference Genes
The threshold-cycle (Ct) of 10 reference genes under different tested conditions were shown (Figure 1). Among various adult ages, SOD and RPS18 had smaller gene expression variations, whereas EF1α and GAPDH had higher (Figure 1A). Across different adult tissues and under temperature treatments, the expression fluctuations were lower in RPS18 and RPS28 and higher in ACT and GAPDH (Figure 1B,C). A combination of above results indicated that the expression difference was small in all reference genes except for ACT and GAPDH ( Figure 1D).
3.3. Stability of the Ten HKGs Among Adult Ages
Based on average expression stability (M-values) and pairwise variation (V-values), the geNorm algorithm evaluate the gene stability. These results exhibited that ACT, RPS18 and SOD were the most suitable, with M-values below 0.20 (Figure 2A, Table 2). Moreover, V-values suggested that V2/3 to V9/10 values were below 0.15, showing that two diverse references were equal for assessing the gene transcript among adult ages (Figure 2B).
The NormFinder algorithm indicated that the stable ranking of ten reference genes from the most to the least stable were as follows: SOD, RPS18, ACT, RPL13, RPL4, RPS28, RPL27, α-TUB, EF1α and GAPDH, with the p value of 0.091, 0.143, 0.172, 0.243, 0.268, 0.329, 0.458, 0.651, 0.849 and 0.954, respectively (Figure 2C, Table 2). The BestKeeper analysis showed that RPL4, RPS28 and SOD were the top three stable genes (Figure 2D, Table 2).
Refer to the results of RefFinder, the ranking of 10 reference genes at adult ages were as follows: SOD>RPS18>ACT>RPL4>RPS28>RPL13>EF1α>RPL27>α-TUB>GAPDH (Figure 5A). Therefore, the combination of SOD and RPS18 are the most appropriate for gene transcript analysis by qRT-PCR among various adult ages (Table 3).
3.4. Stability of the Ten HKGs Across Various Adult Tissues
The geNorm results displayed that RPS28, EF1α, RPL4 and RPS18 were the top four stable genes (Figure 3A, Table 2). V-values data demonstrated that all values of from V2/3 to V9/10 were below 0.15, showing that two reference genes in various adult tissues were suitable (Figure 3B).
According to the NormFinder data, the rankings of reference genes were as follows:RPS18>RPS28>RPL13>RPL4>EF1α>RPL27>α-TUB>SOD>ACT>GAPDH (Figure 3C, Table 2). Moreover, the p values of all reference genes were less than 1.0 (Figure 3C, Table 2). The BestKeeper results indicated that RPS18 and RPS28 were the most stable, with the SD values of 0.700 and 0.914, respectively (Figure 3D, Table 2). Furthermore, the SD values of ACT, GADPH, α-TUB and RPL4 were above 1.0 (Figure 3D, Table 2).
3.5. Stability of the Ten HKGs at Diverse Temperature Conditions
The geNorm results indicated that RPS18, SOD and RPL13 were the most steady references in diverse temperatures, with M-values of 0.123, 0.123 and 0.188, respectively (Figure 4A, Table 2). Moreover, V-values data displayed that all values were below 0.15, showing that two reference genes in various temperature treaments were suitable (Figure 4B).
The NormFinder results manifested that the steady rankings were RPS28, RPS18, SOD, EF1α, α-TUB, RPL13, RPL4, RPL27, GAPDH and ACT (Figure 4C, Table 2). BestKeeper data uncovered that the SD values of these genes were less than 1.0, except for ACT( Figure 4D, Table 2).
Refer to the results of RefFinder, the stability orders were as follows: RPS18>SOD>RPS28>RPL13>RPL27>RPL4>EF1α>α-TUB>GAPDH>ACT (Figure 5C). When combinating the three diverse treaments together, the RefFinder results showed that the stability ranking in sequence were RPS18, RPL13, RPS28, RPL4, RPL27, EF1α, SOD, α-TUB, GAPDH and ACT (Figure 5D). To sum up, RPS18 and SOD are the best reference gene pair to evaluate the gene transcript in diverse temperature conditions (Table 3).
3.6. Validation of the Selected Reference Genes
To estimate the performance of the recommended reference genes, the relative mRNA level of EgUAP was measured using the most stable references (RPS18 and RPS28) under various adult tissues. Our results indicated that the transcription level of EgUAP was high in the foregut, trachea and antenna, and low in the midgut, hindgut and epidermis in E. gorhami (Figure 6).
4. Discussion
In the study, the expression stabilities of ten selected reference genes in E. gorhami were estimated to perform RT-PCR analysis normalization, laying an vital basis for future study of gene functions. Notably, we evaluated the stability of these genes under three different conditions (adult ages, adult tissues/organs and temperatures) for the first time in E. gorhami. These findings suggested that SOD and RPS18 were the most ideal reference combination to measure gene transcription levels among different adult ages (Figure 1, Figure 2 and Figure 5, Table 2 and Table 3) and at various temperatures (Figure 1, Figure 4 and Figure 5, Table 2 and Table 3); RPS18 and RPS28 was the most reliable genes to assess gene expressions under diverse adult tissues (Figure 1, Figure 3 and Figure 5, Table 2 and Table 3).
Ribosomal proteins play an vital function on ribosome assembly, which bind to four ribosomal RNAs (rRNAs) to constitute the ribosomes [23]. Similar to our findings, ribosomal proteins are selected as the most optimal reference genes in insect species, such as Coleopterans M. sibirica (RPL6 and RPL13) [11], Leptinotarsa decemlineata (RP18 and RP4) [24], Phaedon brassicae (RPL32 and RPL19) [25], Henosepilachna vigintioctopunctata (RPL13 and RPS18) [15], Tribolium castaneum (RPS6, RPL13a, RPS3 and RPL18) [26] ,Ips sexdentatus (RPS3) [27] and H. vigintioctomaculata (RPS18 and RPL13) [13], Hemipterans Psammotettix striatus (RPLP2) [28], Rhopalosiphum padi (RPL13, RPS6 and RPS18) [29], Aphis glycines (RPS9) [30], Ferrisia gilli (RPS8, RPL40 and RPL7) [31], Diaphorina citri (RPL7) [32] and Dichelops melacanthus (RPL9 and RPS23) [33], Hymenopterans Anastatus japonicus (RPL13 and RPS6) [34], Lepidopterans Mythimna loreyi (RPL10, RPL27 and RPS3) [35], Plutella xylostella (RPS13 and RPS23) [36], Spodoptera litura (RPS13 and RPLP0) [37], P. operculella (RPL13) [12], Bombyx mori (RPS7) [38] and Helicoverpa armigera (RPS15 and RPL27) [39], Dipterans Aphidoletes aphidimyza (RPL8 and RPS3) [40], Exorista sorbillans (RP49) [41] and Chlorops oryzae (RPS15) [42], and Orthopteran Locusta migratoria (RPL32) [43], and Thysanopterans Megalurothrips usitatus (RPL30) [44] and Frankliniella occidentalis (RPL32) [45].
Under different adult ages and temperatures, SOD was verified to be the most reliable gene (Figure 1, Figure 2, Figure 4 and Figure 5, Table 2 and Table 3). Superoxide dismutase (SOD) is a crucial antioxidant enzyme, catalyzing the conversion of reactive oxygen species into oxygen and hydrogen peroxide [46]. Consistent with our data, SOD is recommended as the most appropriate reference gene in Spodoptera frugiperda [47], Thrips tabaci [48], Riptortus pedestris [49] and Spodoptera exigua [50].
In addition, The BestKeeper analysis data indicated that the SD values of α-TUB, EF1α, GAPDH and ACT were greater than 1.0 (Table 2), showing that these genes were unaccommodated as reference genes to perform RT-qPCR normalization. Similar results have been verified in other insects, such as P. operculella [12], Ophraella communa [51], A. aphidimyza [40], Hippodamia convergens [52], H. vigintioctomaculata [13], Colaphellus bowringi [53], H. vigintioctopunctata [15] and M. sibirica [11].
To further validate the accuracy of RPS18 and RPS28 in RT-qPCR normalization in E. gorhami, we examined the relative transcript level of EgUAP under diverse adult tissues. Our results exhibited that EgUAP expression was high in the foregut, trachea and antenna, and low in the midgut, hindgut and epidermis (Figure 6).
Overall, it is essential to screen and verify the most suitable references to guarantee the accuracy of gene expression. The study would offer a solid basis for further molecular functions of target genes in E. gorhami.
5. Conclusions
Ten potential reference genes in E. gorhami were assessed for accurate RT-qPCR analysis in E. gorhami under three different treaments. These results demonstrated that the most steady reference genes were as follows: SOD and RPS18 for different adult ages and various temperatures, RPS18 and RPS28 for adult tissues. This study is the first time to establish the RT-qPCR normalization analyses in E. gorhami, facilitating further research on gene functions of E. gorhami.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: A list of primers used for RT-PCR of the genes.
Author Contributions
Experiments were designed by G.-F.Y., X.-T.Y., Y.Z., G.W., X.-H.Y., C.-H.S. Figures designed and created by G.-F.Y., X.-T.Y., Y.Z., J.-H.L., G.W., X.-H.Y., C.-H.S. Statistical analyses were implemented by G.-F.Y., X.-T.Y., Y.Z., J.-H.L., X.-F.L., L.Z., H.-H.Z., L.J.,G.W., X.-H.Y., C.-H.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Major Special Project of Breeding for Genetically Modified Organisms in China (No. 2016ZX08012-005) and the Innovation Project of Chinese Academy of Agricultural Sciences (No. 2060302-049-091).
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Terao, M.; Hirose, Y.; Shintani, Y. Effects of temperature and photoperiod on termination of pseudopupal diapause in the bean blister beetle, Epicauta gorhami. J. Insect Physiol. 2012, 58, 737–742. [CrossRef]
- Shintani, Y.; Terao, M.; Tanaka, S. Adaptive significance of precocious pupation in the bean blister beetle, Epicauta gorhami (Coleoptera: Meloidae), a hypermetamorphic insect. J. Insect Physiol. 2017, 99, 107–112. [CrossRef]
- Terao, M.; Tokuda, M.; Shintani, Y. Geographic Variation in Photoperiodic Response for Induction of Pseudopupal Diapause in Epicauta gorhami (Coleoptera: Meloidae). Environ. Èntomol. 2021, 50, 1145–1150. [CrossRef]
- Zhou, Z.; Liu, Y.; Chen, X. Structural Features and Phylogenetic Implications of Three New Mitochondrial Genomes of Blister Beetles (Coleoptera: Meloidae). J. Insect Sci. 2021, 21. [CrossRef]
- Hina, A.; Razzaq, M.K.; Abbasi, A.; Shehzad, M.B.; Arshad, M.; Sanaullah, T.; Arshad, K.; Raza, G.; Ali, H.M.; Hayat, F.; et al. Genomic blueprints of soybean (Glycine max) pathogen resistance: revealing the key genes for sustainable agriculture. Funct. Plant Biol. 2024, 51. [CrossRef]
- Vianna, G.; Cunha, N.; Rech, E. Soybean seed protein storage vacuoles for expression of recombinant molecules. Curr. Opin. Plant Biol. 2023, 71, 102331. [CrossRef]
- ozefczuk, J.; Adjaye, J. Quantitative real-time PCR-based analysis of gene expression. Methods Enzymol. 2011, 500, 99-109.
- Derveaux, S.; Vandesompele, J.; Hellemans, J. How to do successful gene expression analysis using real-time PCR. Methods 2010, 50, 227–230. [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, Research0034.
- Valasek, M.A.; Repa, J.J. The power of real-time PCR. Adv. Physiol. Educ. 2005, 29, 151–159. [CrossRef]
- Shen, C.-H.; Tang, M.; Li, X.-F.; Zhu, L.; Li, W.; Deng, P.; Zhai, Q.; Wu, G.; Yan, X.-H. Evaluation of reference genes for quantitative expression analysis in Mylabris sibirica (Coleoptera, Meloidae). Front. Physiol. 2024, 15, 1345836. [CrossRef]
- Shen, C.H.; Peng, L.J.; Zhang, Y.X.; Zeng, H.R.; Yu, H.F.; Jin, L.; Li, G.Q. Reference genes for expression analyses by qRT-PCR in Phthorimaea operculella (Lepidoptera: Gelechiidae). Insects. 2022, 13, 140.
- Zhang, Y.-X.; Tan, Q.; Shen, C.-H.; Wu, J.-J.; Wu, Y.-K.; Li, W.-Z.; Jin, L.; Li, G.-Q. Reference gene selection for transcriptional profiling by RT-qPCR in the 28-spotted larger potato ladybird. J. Asia Pac. Entomol. 2022, 25, 101900.
- Wang, Z.; Shang, X.; Wei, J.; Tian, X.; Liu, Y.; Zhang, G. Evaluation and validation of reference genes for gene expression analysis using qRT-PCR in the sugarcane stem borer Chilo sacchariphagus (Lepidoptera: Pyralidae). Insects. 2024, 15, 594.
- Lü, J.; Yang, C.; Zhang, Y.; Pan, H. Selection of reference genes for the normalization of RT-qPCR data in gene expression studies in insects: A systematic review. Front. Physiol. 2018, 9, 1560.
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper--Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509-515.
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: a miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [CrossRef]
- Xie, F.; Wang, J.; Zhang, B. RefFinder: a web-based tool for comprehensively analyzing and identifying reference genes. Funct. Integr. Genom. 2023, 23, 1–5. [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [CrossRef]
- Landry-Voyer, A.-M.; Hassani, Z.M.; Avino, M.; Bachand, F. Ribosomal Protein uS5 and Friends: Protein–Protein Interactions Involved in Ribosome Assembly and Beyond. Biomolecules 2023, 13, 853. [CrossRef]
- Shi, X.-Q.; Guo, W.-C.; Wan, P.-J.; Zhou, L.-T.; Ren, X.-L.; Ahmat, T.; Fu, K.-Y.; Li, G.-Q. Validation of reference genes for expression analysis by quantitative real-time PCR in Leptinotarsa decemlineata (Say). BMC Res. Notes 2013, 6, 93–10. [CrossRef]
- Ma, L.; Jiang, T.; Liu, X.; Xiao, H.; Peng, Y.; Zhang, W. Evaluation of candidate reference genes for gene expression analysis in the brassica leaf beetle, Phaedon brassicae (Coleoptera: Chrysomelidae). PLOS ONE 2021, 16, e0251920. [CrossRef]
- Toutges, M.J.; Hartzer, K.; Lord, J.; Oppert, B. Evaluation of Reference Genes for Quantitative Polymerase Chain Reaction across Life Cycle Stages and Tissue Types of Tribolium castaneum. J. Agric. Food Chem. 2010, 58, 8948–8951. [CrossRef]
- Sellamuthu, G.; Amin, S.; Bílý, J.; Synek, J.; Modlinger, R.; Sen, M.K.; Chakraborty, A.; Roy, A. Reference Gene Selection for Normalizing Gene Expression in Ips Sexdentatus (Coleoptera: Curculionidae: Scolytinae) Under Different Experimental Conditions. Front. Physiol. 2021, 12. [CrossRef]
- Yuan, F.; Xie, Z.; Li, Z.; Lian, P.; Wei, C. Screening of reference genes for gene expression study in different tissues from the transcriptome data of the vector leafhopper Psammotettix striatus. Gene 2024, 927, 148696. [CrossRef]
- Li, M.; Li, X.; Wang, C.; Li, Q.; Zhu, S.; Zhang, Y.; Li, X.; Yang, F.; Zhu, X. Selection and validation of reference genes for qRT-PCR analysis of Rhopalosiphum padi (Hemiptera: Aphididae). Front. Physiol. 2021, 12, 663338.
- Bansal, R.; Mamidala, P.; Mian, M.A.R.; Mittapalli, O.; Michel, A.P. Validation of Reference Genes for Gene Expression Studies in Aphis glycines (Hemiptera: Aphididae). J. Econ. Èntomol. 2012, 105, 1432–1438. [CrossRef]
- Bansal, R.; Haviland, D.R.; Hunter, W.B. Selection and validation of reference genes for quantifying gene expression in the Gill's mealybug. J. Econ. Entomol. 2023, 116, 2166-2172.
- Bassan, M.M.; Angelotti-Mendonça, J.; Alves, G.R.; Yamamoto, P.T.; Filho, F.d.A.A.M. Selection of Reference Genes for Expression Studies in Diaphorina citri (Hemiptera: Liviidae). J. Econ. Èntomol. 2017, 110, 2623–2629. [CrossRef]
- Pinheiro, D.H.; Moreira, R.O.; Leite, N.A.; Redoan, A.C.; Xavier, A.D.S.; Barros, B.A.; Carneiro, N.P. Suitable reference genes for RT-qPCR analysis in Dichelops melacanthus (Hemiptera: Pentatomidae). Mol. Biol. Rep. 2020, 47, 4989-5000.
- Liu, Z.; Xiao, J.; Xia, Y.; Wu, Q.; Zhao, C.; Li, D. Selection and validation of reference genes for RT-qPCR-based analyses of Anastatus japonicus Ashmead (Hymenoptera: Helicopteridae). Front. Physiol. 2022, 13, 1046204.
- Wang, L.; Yang, C.; Liu, Q.; Zhang, X.; Mei, X.; Zhang, T.; Ning, J. Validation and Evaluation of Reference Genes for Quantitative Real-Time PCR Analysis in Mythimna loreyi (Lepidoptera: Noctuidae). Insects 2024, 15, 185. [CrossRef]
- Fu, W.; Xie, W.; Zhang, Z.; Wang, S.; Wu, Q.; Liu, Y.; Zhou, X.; Zhou, X.; Zhang, Y. Exploring Valid Reference Genes for Quantitative Real-time PCR Analysis in Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Biol. Sci. 2013, 9, 792–802. [CrossRef]
- Wu, S.; Luo, Y.; Zeng, Z.; Yu, Y.; Zhang, S.; Hu, Y.; Chen, L. Determination of internal controls for quantitative gene expression of Spodoptera litura under microbial pesticide stress. Sci. Rep. 2024, 14, 1–13. [CrossRef]
- Dalai, M.; Jagota, A. Identification of specific reference gene for normalization of RT-qPCR data in rhythmic gene expression studies of the effect of developmental hormone antagonist in postembryonic development in Bombyx mori. Front. Insect Sci. 2024, 4, 1362473.
- Zhang, S.; An, S.; Li, Z.; Wu, F.; Yang, Q.; Liu, Y.; Cao, J.; Zhang, H.; Zhang, Q.; Liu, X. Identification and validation of reference genes for normalization of gene expression analysis using qRT-PCR in Helicoverpa armigera (Lepidoptera: Noctuidae). Gene. 2015, 555, 393-402.
- Shen, X.X.; Zhang, G.Q.; Zhao, Y.X.; Zhu, X.X.; Yu, X.F.; Yang, M.F.; Zhang, F. Selection and validation of optimal reference genes for RT-qPCR analyses in Aphidoletes aphidimyza Rondani (Diptera: Cecidomyiidae). Front. Physiol. 2023, 14, 1277942.
- Yang, J.; Jiang, Z.; Xu, Q.; Liu, X.; Dai, M.; Li, B.; Wei, J. Evaluation of suitable reference genes for expression analysis using quantitative real-time polymerase chain reaction in the parasitoid Exorista sorbillans (Diptera: Tachinidae). Arch. Insect Biochem. Physiol. 2023, 113, e22009. [CrossRef]
- Tian, P.; Qiu, L.; Zhou, A.; Chen, G.; He, H.; Ding, W.; Li, Y. Evaluation of Appropriate Reference Genes For Investigating Gene Expression in Chlorops oryzae (Diptera: Chloropidae). J. Econ. Èntomol. 2019, 112, 2207–2214. [CrossRef]
- Yang, Q.; Li, Z.; Cao, J.; Zhang, S.; Zhang, H.; Wu, X.; Zhang, Q.; Liu, X. Selection and Assessment of Reference Genes for Quantitative PCR Normalization in Migratory Locust Locusta migratoria (Orthoptera: Acrididae). PLOS ONE 2014, 9, e98164. [CrossRef]
- Hou, Q.; Yuan, L.; Jin, H.; Yan, H.; Li, F.; Wu, S. Identification and validation of reference genes for normalization of gene expression analysis using qRT-PCR in Megalurothrips usitatus (thysanoptera: thripidae). Front. Physiol. 2023, 14, 1161680.
- Zheng, Y.T.; Li, H.B.; Lu, M.X.; Du, Y.Z. Evaluation and validation of reference genes for qRT-PCR normalization in Frankliniella occidentalis (Thysanoptera: Thripidae). PLoS One. 2014, 9, e111369.
- Yamamoto, K.; Yamaguchi, M. Characterization of a novel superoxide dismutase in Nilaparvata lugens. Arch. Insect Biochem. Physiol. 2021, 109, e21862. [CrossRef]
- Han, S.; Qin, Q.; Wang, D.; Zhou, Y.; He, Y. Selection and evaluation of reference genes for qRT-PCR in Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects. 2021, 12, 902.
- Singh, S.; Gupta, M.; Pandher, S.; Kaur, G.; Goel, N.; Rathore, P.; Palli, S.R. RNA sequencing, selection of reference genes and demonstration of feeding RNAi in Thrips tabaci (Lind.) (Thysanoptera: Thripidae). BMC Mol. Biol. 2019, 20, 1–21. [CrossRef]
- Wang, L.; Liu, Q.; Guo, P.; Gao, Z.; Chen, D.; Zhang, T.; Ning, J. Evaluation of Reference Genes for Quantitative Real-Time PCR Analysis in the Bean Bug, Riptortus pedestris (Hemiptera: Alydidae). Insects 2023, 14, 960. [CrossRef]
- Zhu, X.; Yuan, M.; Shakeel, M.; Zhang, Y.; Wang, S.; Wang, X.; Zhan, S.; Kang, T.; Li, J. Selection and evaluation of reference genes for expression analysis using qRT-PCR in the beet armyworm Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae). PLoS One. 2014, 9, e84730.
- Zhang, Y.; Chen, J.; Chen, G.; Ma, C.; Chen, H.; Gao, X.; Tian, Z.; Cui, S.; Tian, Z.; Guo, J.; et al. Identification and Validation of Reference Genes for Quantitative Gene Expression Analysis in Ophraella communa. Front. Physiol. 2020, 11, 355. [CrossRef]
- Yang, C.; Preisser, E.L.; Zhang, H.; Liu, Y.; Dai, L.; Pan, H.; Zhou, X. Selection of reference genes for RT-qPCR analysis in Coccinella septempunctata to assess un-intended effects of RNAi transgenic plants. Front. Plant. Sci. 2016, 7, 1672.
- Tan, Q.-Q.; Zhu, L.; Li, Y.; Liu, W.; Ma, W.-H.; Lei, C.-L.; Wang, X.-P. A De Novo Transcriptome and Valid Reference Genes for Quantitative Real-Time PCR in Colaphellus bowringi. PLOS ONE 2015, 10, e0118693–e0118693. [CrossRef]
Figure 1.
Expression levels of ten house-keeping genes in Epicauta gorhami. The mean Ct values for 10 candidate reference genes are shown in four different experiments: (A) adult ages, (B) adult tissues, (C) temperature, (D) All samples. Mean Ct values for the ten candidate reference genes are presented in violin plot. Abbreviation: ACT, actin; SOD, Superoxide dismutase; α-TUB, α-tubulin; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; EF1α, elongation factor 1α; RPL4, RPL13 ,RPL27, RPS18 and RPS28, ribosomal protein. The abbreviations are exactly the same as Figure 2, Figure 3, Figure 4 and Figure 5.
Figure 1.
Expression levels of ten house-keeping genes in Epicauta gorhami. The mean Ct values for 10 candidate reference genes are shown in four different experiments: (A) adult ages, (B) adult tissues, (C) temperature, (D) All samples. Mean Ct values for the ten candidate reference genes are presented in violin plot. Abbreviation: ACT, actin; SOD, Superoxide dismutase; α-TUB, α-tubulin; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; EF1α, elongation factor 1α; RPL4, RPL13 ,RPL27, RPS18 and RPS28, ribosomal protein. The abbreviations are exactly the same as Figure 2, Figure 3, Figure 4 and Figure 5.
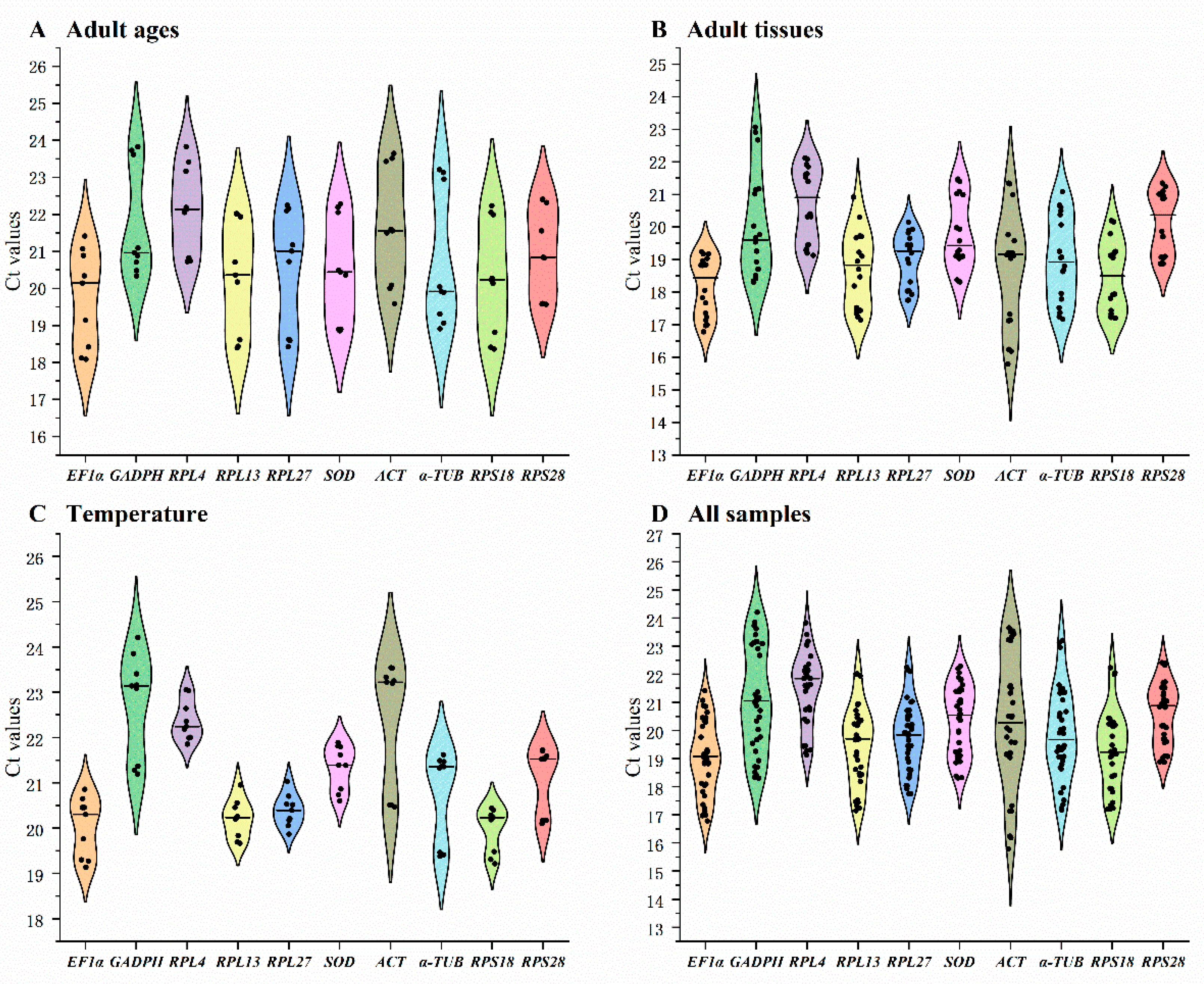
Figure 2.
Stability of the ten house-keeping genes in Epicauta gorhami during various adult ages. Diverse ages of Epicauta gorhami adults were sampled (collected on the first to third days of the newly-emerged adults). The expression stability rankings are determined by geNorm, NormFinder and BestKeeper.
Figure 2.
Stability of the ten house-keeping genes in Epicauta gorhami during various adult ages. Diverse ages of Epicauta gorhami adults were sampled (collected on the first to third days of the newly-emerged adults). The expression stability rankings are determined by geNorm, NormFinder and BestKeeper.
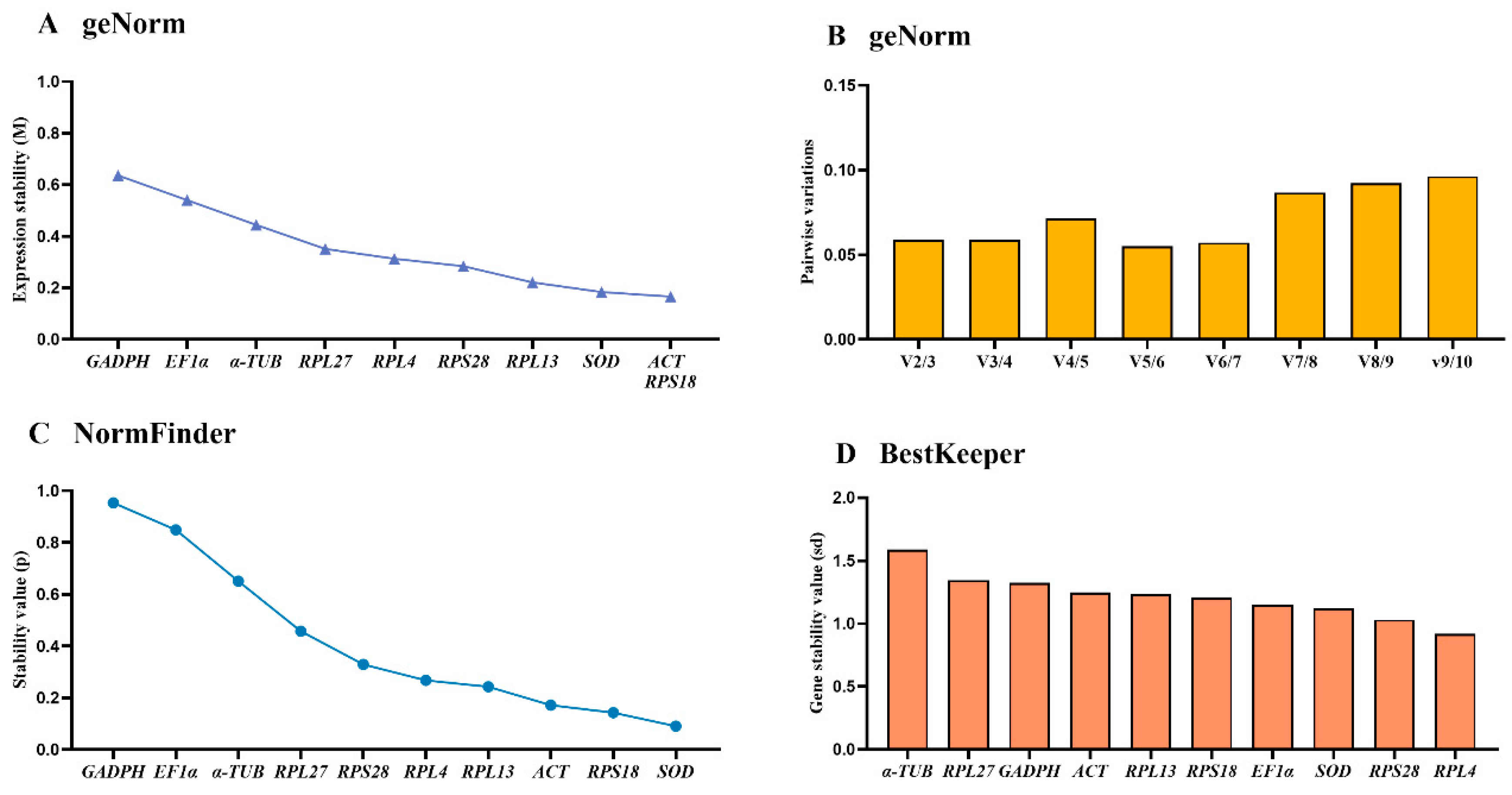
Figure 3.
Stabilities of the ten house-keeping genes in Epicauta gorhami among diverse adult tissues. Foregut, midgut, hindgut, epidermis, trachea and antenna were dissected from the 5-day-old adults. The expression stability rankings are determined by geNorm, NormFinder and BestKeeper.
Figure 3.
Stabilities of the ten house-keeping genes in Epicauta gorhami among diverse adult tissues. Foregut, midgut, hindgut, epidermis, trachea and antenna were dissected from the 5-day-old adults. The expression stability rankings are determined by geNorm, NormFinder and BestKeeper.
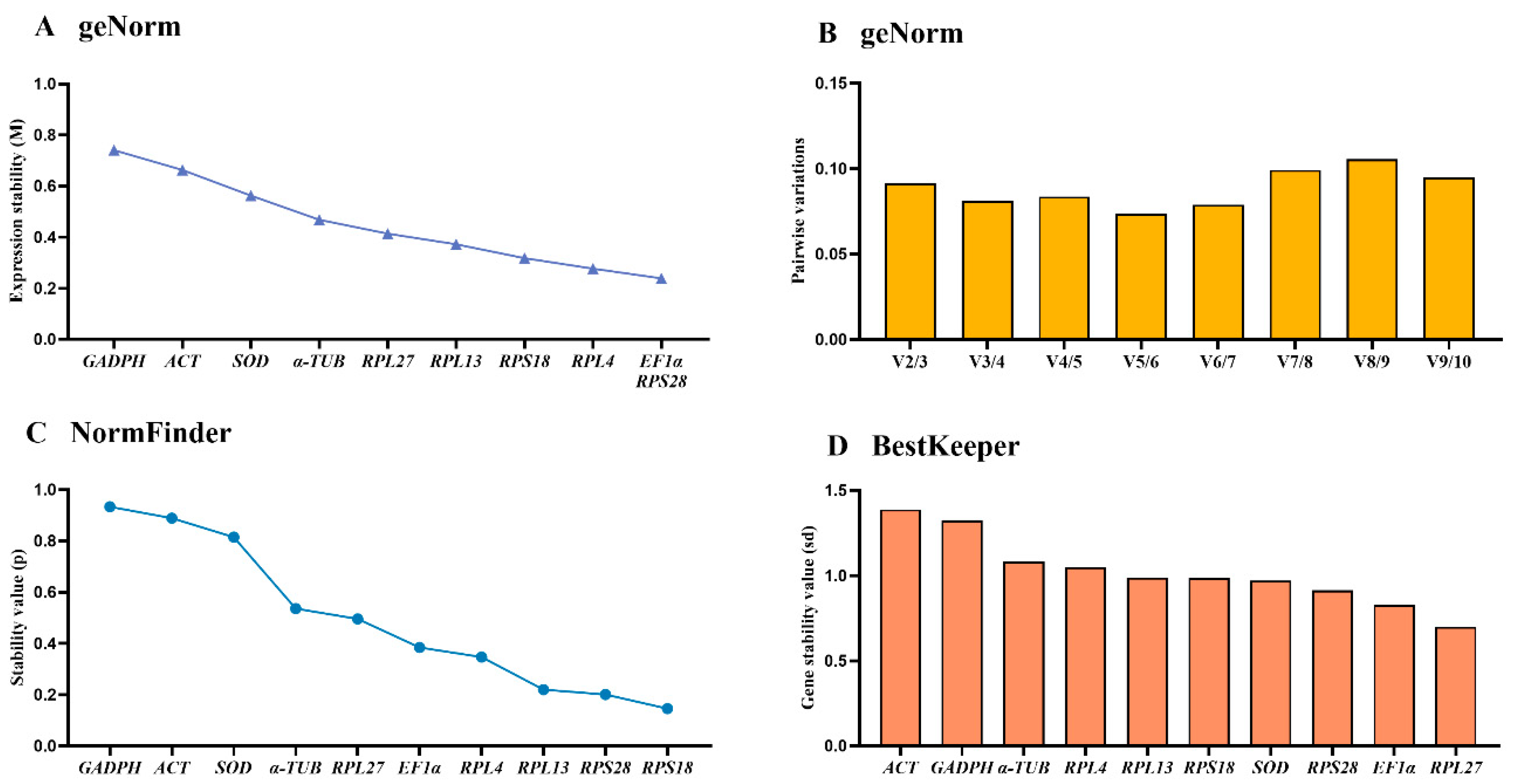
Figure 4.
Stability of the ten house-keeping genes in Epicauta gorhami under different temperatures. The newly-emerged adults reared under three temperatures (4°C, 25 °C and 37 °C) were collected. The expression stability rankings are determined by geNorm, NormFinder and BestKeeper.
Figure 4.
Stability of the ten house-keeping genes in Epicauta gorhami under different temperatures. The newly-emerged adults reared under three temperatures (4°C, 25 °C and 37 °C) were collected. The expression stability rankings are determined by geNorm, NormFinder and BestKeeper.
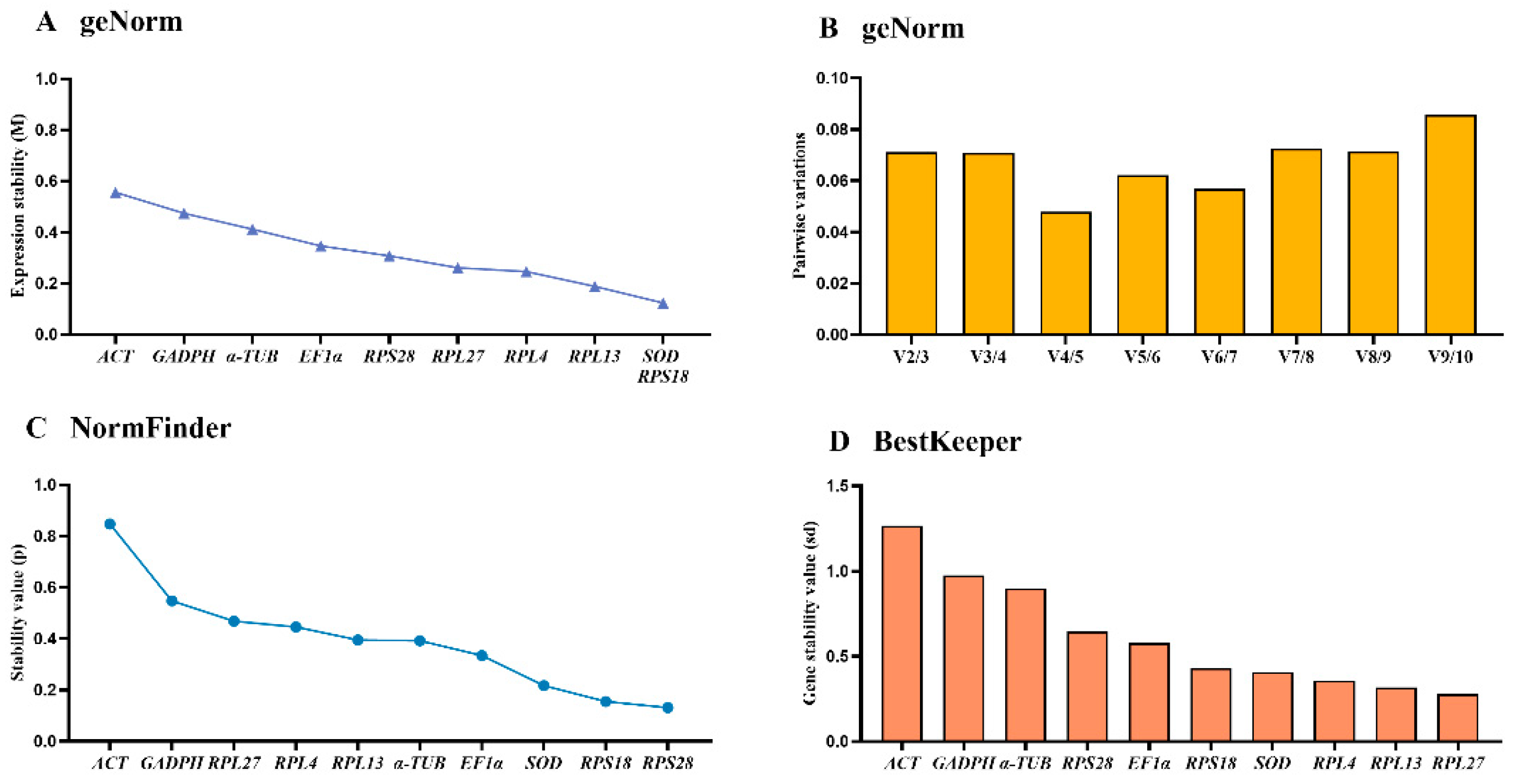
Figure 5.
Stabilities of the ten house-keeping genes in Epicauta gorhami in different samples. The stability of the reference genes as calculated by the Geomean method of RefFinder. A lower Geomean of ranking value denotes more stable expression. (A) adult ages, (B) adult tissues, (C) temperature, (D) all samples.
Figure 5.
Stabilities of the ten house-keeping genes in Epicauta gorhami in different samples. The stability of the reference genes as calculated by the Geomean method of RefFinder. A lower Geomean of ranking value denotes more stable expression. (A) adult ages, (B) adult tissues, (C) temperature, (D) all samples.
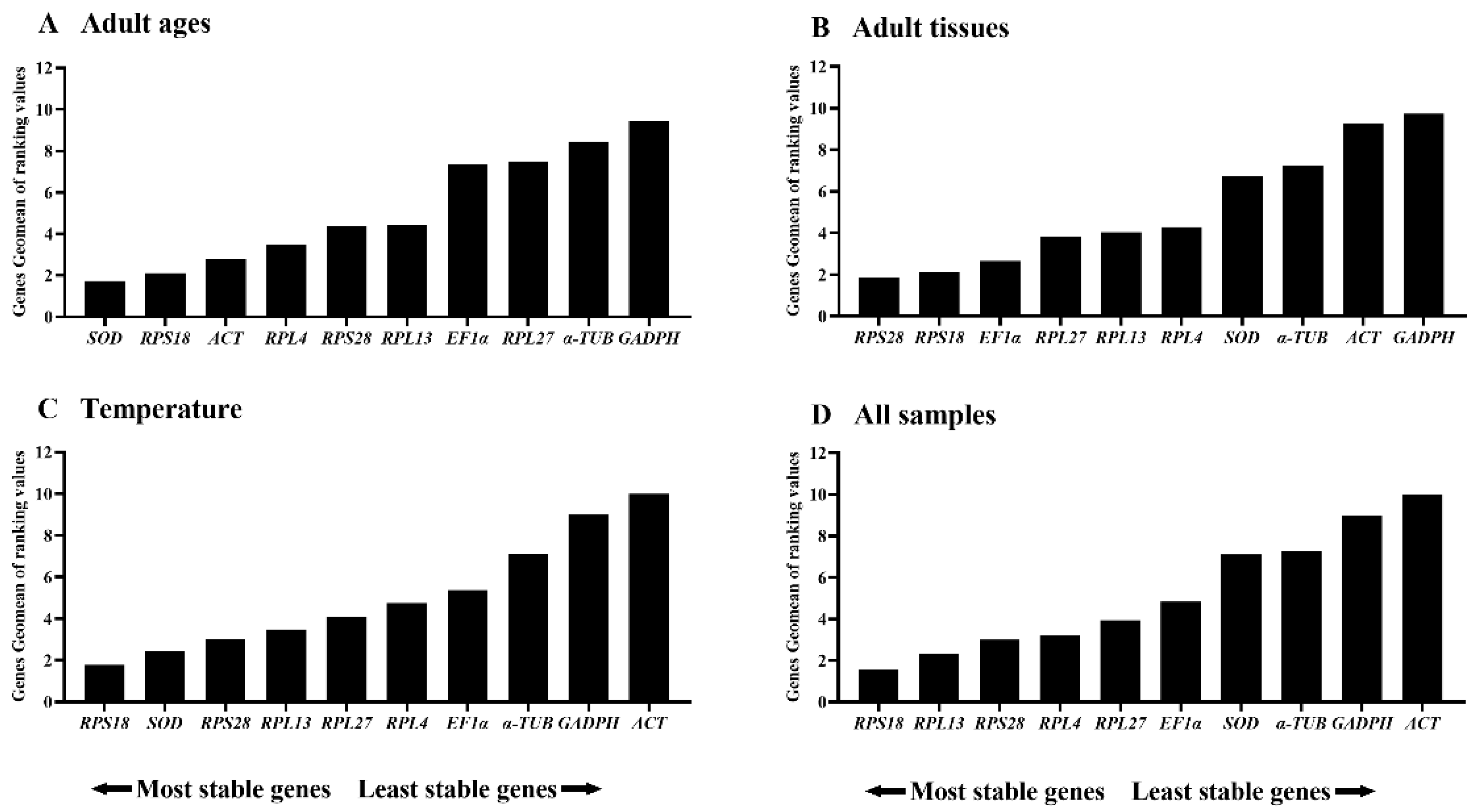
Figure 6.
Relative gene expression of EgUAP in various adult tissues of Epicauta gorhami. The relative gene expression level of EgUAP in the foregut, midgut, hindgut, epidermis, trachea and antenna was normalized to the best stable reference genes (RPS18 and RPS28), respectively. The values are means + SE. Different letters indicate significant differences in gene expression among different tissues (P < 0.05).
Figure 6.
Relative gene expression of EgUAP in various adult tissues of Epicauta gorhami. The relative gene expression level of EgUAP in the foregut, midgut, hindgut, epidermis, trachea and antenna was normalized to the best stable reference genes (RPS18 and RPS28), respectively. The values are means + SE. Different letters indicate significant differences in gene expression among different tissues (P < 0.05).

Table 1.
Primers of 10 candidate house-keeping genes used in qRT-PCR.
| Gene | Primer sequences (5'to 3´) | Length (bp) | Slope | R2 | Efficiency (%) |
|---|---|---|---|---|---|
| EF1α | F- ATCATTGACGCACCTGGACA | 99 | -3.435 | 0.999 | 95.50 |
| R- ACCAGTACCAGCAGCAACAA | |||||
| GAPDH | F-ACAGTACATGCCACCACAGC | 180 | -3.351 | 0.999 | 98.82 |
| R-TGGTACACGGAAGGCCATAC | |||||
| RPL4 | F- TCGATGAACCACCGTCAACC | 80 | -3.308 | 0.993 | 100.60 |
| R- CGACCGGTACCCCATGATTC | |||||
| RPL13 | F-AAGCCGCCGGTATTAACAGC | 172 | -3.318 | 0.998 | 100.16 |
| R-TCACCAGGACGCAACTTCTT | |||||
| RPL27 | F-TCGTATTGGTCTTGGCAGGC | 112 | -3.373 | 0.999 | 98.03 |
| R-TCAATGCCGGCAACAATAGC | |||||
| SOD | F-AGTTGTCCATGCTGATCCGG | 95 | -3.32 | 0.998 | 100.08 |
| R-TAACACCACAGGCCAAACGT | |||||
| ACT | F- TACGTGTGGCACCTGAAGAA | 169 | -3.284 | 0.997 | 101.60 |
| R- CCAGTTGTACGACCAGAAGCA | |||||
| α-TUB | F-ATGGGCACGTCTTGATCACA | 174 | -3.258 | 0.999 | 102.73 |
| R-TCATTTTCACCTTCGCCCGA | |||||
| RPS18 | F-AGGTGTTGGTCGTCGTTACTC | 210 | -3.323 | 0.999 | 99.96 |
| R-TCTAAGGTAGCCGATGTGAGC | |||||
| RPS28 | F-GGTGAGCAAAACCGTCAGATC | 90 | -3.347 | 0.997 | 98.97 |
| R-TGCTTCACGTTCAGACTCCA |
Note: ACT, actin; SOD, Superoxide dismutase; α-TUB, α-tubulin; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; EF1α, elongation factor 1α; RPL4, RPL13 ,RPL27, RPS18 and RPS28, ribosomal protein.
Table 2.
Expression stability of the candidate reference genes under different experimental conditions.
Table 2.
Expression stability of the candidate reference genes under different experimental conditions.
| Conditions | CRGs* | geNorm | Normfinder | Normfinder | ΔCt | ||||
|---|---|---|---|---|---|---|---|---|---|
| Stability | Rank | Stability | Rank | Stability | Rank | Stability | Rank | ||
| Developmental ages | EF1α | 0.540 | 8 | 0.849 | 9 | 1.149 | 4 | 0.931 | 9 |
| GADPH | 0.636 | 9 | 0.954 | 10 | 1.325 | 8 | 1.024 | 10 | |
| RPL4 | 0.313 | 5 | 0.268 | 5 | 0.920 | 1 | 0.543 | 5 | |
| RPL13 | 0.221 | 3 | 0.243 | 4 | 1.236 | 6 | 0.500 | 4 | |
| RPL27 | 0.351 | 6 | 0.458 | 7 | 1.344 | 9 | 0.619 | 7 | |
| SOD | 0.184 | 2 | 0.091 | 1 | 1.121 | 3 | 0.443 | 1 | |
| ACT | 0.166 | 1 | 0.172 | 3 | 1.249 | 7 | 0.469 | 3 | |
| α-TUB | 0.444 | 7 | 0.651 | 8 | 1.589 | 10 | 0.782 | 8 | |
| RPS18 | 0.166 | 1 | 0.143 | 2 | 1.209 | 5 | 0.476 | 2 | |
| RPS28 | 0.284 | 4 | 0.329 | 6 | 1.033 | 2 | 0.557 | 6 | |
| Adult tissues | EF1α | 0.239 | 1 | 0.384 | 5 | 0.828 | 2 | 0.621 | 5 |
| GADPH | 0.741 | 9 | 0.933 | 10 | 1.326 | 9 | 1.055 | 10 | |
| RPL4 | 0.277 | 2 | 0.347 | 4 | 1.051 | 7 | 0.614 | 4 | |
| RPL13 | 0.372 | 4 | 0.220 | 3 | 0.989 | 6 | 0.608 | 3 | |
| RPL27 | 0.414 | 5 | 0.496 | 6 | 0.700 | 1 | 0.710 | 6 | |
| SOD | 0.563 | 7 | 0.815 | 8 | 0.971 | 4 | 0.944 | 8 | |
| ACT | 0.663 | 8 | 0.889 | 9 | 1.389 | 10 | 1.012 | 9 | |
| α-TUB | 0.468 | 6 | 0.536 | 7 | 1.085 | 8 | 0.744 | 7 | |
| RPS18 | 0.318 | 3 | 0.146 | 1 | 0.987 | 5 | 0.549 | 1 | |
| RPS28 | 0.239 | 1 | 0.201 | 2 | 0.914 | 3 | 0.552 | 2 | |
| Temparature treatment | EF1α | 0.346 | 6 | 0.334 | 4 | 0.58 | 6 | 0.532 | 5 |
| GADPH | 0.474 | 8 | 0.547 | 9 | 0.976 | 9 | 0.675 | 9 | |
| RPL4 | 0.246 | 3 | 0.446 | 7 | 0.357 | 3 | 0.549 | 6 | |
| RPL13 | 0.188 | 2 | 0.395 | 6 | 0.317 | 2 | 0.522 | 4 | |
| RPL27 | 0.261 | 4 | 0.468 | 8 | 0.278 | 1 | 0.555 | 7 | |
| SOD | 0.123 | 1 | 0.217 | 3 | 0.407 | 4 | 0.431 | 3 | |
| ACT | 0.556 | 9 | 0.847 | 10 | 1.265 | 10 | 0.886 | 10 | |
| α-TUB | 0.412 | 7 | 0.392 | 5 | 0.899 | 8 | 0.570 | 8 | |
| RPS18 | 0.123 | 1 | 0.155 | 2 | 0.429 | 5 | 0.415 | 1 | |
| RPS28 | 0.307 | 5 | 0.131 | 1 | 0.646 | 7 | 0.430 | 2 | |
Table 3.
A list of the recommended reference genes in E. gorhami for different experimental conditions.
Table 3.
A list of the recommended reference genes in E. gorhami for different experimental conditions.
| Experimental conditions | The recommended reference genes | |
|---|---|---|
| Adult ages | SOD | RPS18 |
| Adult tissues | RPS28 | RPS18 |
| Temperature | SOD | RPS18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



