
Submitted:
28 October 2024
Posted:
28 October 2024
You are already at the latest version
Abstract
The dietary proteins are one of the most important factors determining health conditions in human. The sufficient digestion and absorption of dietary proteins in the digestive tract have positive effects on performance and recovery in sportspeople or athletes. Improving of protein digestibility is a strategy for maintaining health status and optimal performance in sport or exercise activities. The hypothesis of the present study is that Weizmannia coagulans BC 99 (BC99) can increase muscle mass and strength. This randomized double-blind, controlled trial assigned 72 male college students to receive probiotics (n = 36, 20.25±1.03 years; 1.79±5.94 cm; 73.55±8.73 kg, protein powder with BC99) or placebo (n = 36, 20.19±0.79 years; 179.25±5.16 cm; 73.61±8.24 kg, protein powder) for 12-week. At the baseline and final stages of the study, strength tests and body composition assessment were performed; Blood and stool samples were taken at the end of the 12-week intervention, digestive enzymatic activity of stool samples, biochemical parameters, amino acids and hormone level of plasma were analyzed. BC99 administration significantly improved strength performance, skeletal muscle mass, activity of pepsin and trypsin, the concentrations of branched chain amino acids and essential amino acids, reduced activities of creatine kinase and lactic dehydrogenase and urea nitrogen (BUN) level, and increased testosterone and glucagon-like peptide-1 level in male college students. Therefore, BC99 supplementation can be an important nutritional strategy to improve strength performance, body composition, protein digestion and body metabolism in various groups of individuals.
Keywords:
probiotic
; strength performance
; skeletal muscle
; protein digestion
; inflammation
; oxidative stress
Introduction
Dietary protein is widely regarded as a key macronutrient to maintain a good nutrition and health status, and it could be hydrolyzed into amino acids, dipeptides, or tripeptides by proteases and peptidases in the digestive tract. The amino acids, as an important hydrolysate of protein, are mainly absorbed in small intestine and are transported via the portal blood to peripheral tissues, and are used as raw materials for building tissues or organs, meanwhile, exerting other metabolic regulation function [1,2]. Amino acid availability is often dependent on the content and composition of dietary protein, thus, the content, digestion rates of protein in diet are the determinants of its nutritional value. An adequate intake of proteins in the diet is critical for maintaining a good health in human and mammal animals. Some current evidence has suggested that the insufficient digestion of protein, reduction of enzyme secretion and its activity led to reduction of amino acid uptake [3], and affect the metabolism and function of amino acids in skeletal, liver and other tissue [4]. Therefore, the digestion of food proteins in the gastrointestinal tract and amino acid metabolism are two most important factors determining health conditions and physiologic function.
It is well established that optimal performance of athlete in some sports is associated with skeletal muscle mass and strength. Muscle protein synthesis (MPS) and breakdown (MPB) in skeletal muscle can lead to changes in skeletal muscle mass and muscle strength [5]. Reduction of skeletal muscle mass are associated with metabolic disorders [6] and aging [7].
Dietary supplements are used for improving body composition, endurance, sports performance and metabolic adaptations [8,9]. Protein and amino acids have been used for maintaining strength, recovering muscular mass or preventing nutritional deficiencies [10]. Intake of protein and amino acids can also stimulate MPS [10,11]. However, undigested protein and amino acids can be utilized by bacteria in the intestines and produce harmful metabolites including skatole and sulfide [12], and can cause gastrointestinal discomfort [13]. Thus, intervention strategies have been proposed to improve protein digestibility in gastrointestinal tract and amino acid metabolism levels in body.
Probiotics are living microorganisms that have positive effects on health of the host when ingested in adequate quantities [14]. Many studies have suggested that probiotics can influence the digestion and utilization of proteins in the intestinal tract. Improvement of dietary protein digestion by probiotics can be achieved through regulation of the intestinal microbiota, increase of digestive enzyme activity, improvement of digestibility and absorption. Studies in animals have demonstrated that Lactobacillus plantarum GF103 and Bacillus subtilis B27 improved digestibility of crude protein [15]. Clinical trials have also showed that the probiotic Bacillus coagulans GBI-30 increase the digestion and uptake of plant proteins [16]. These results showed that probiotics can beneficially modulate protein digestion and utilization in the intestinal tract.
Several studies have shown that gut microbiota could influence muscle metabolism and function [17,18]. Therefore, the hypothesis of the ‘gut–muscle axis’ has been raised to study the associations between microbiota and muscle [19], and lead to some new research projects which focus on probiotics to target muscle mass and function [20,21]. Probiotic strains of commercial use mainly include the genera Lactobacillus, Bifidobacterium, and Bacillus [22]. Weizmannia coagulans BC99 (BC99) is a Gram-positive, spore-forming, and lactic-acid-producing probiotic strain isolated from the fecal sample of a healthy infant in Inner Mongolia [23], and are used as probiotics and has exhibited its potential to improve intestinal diseases, such as irritable bowel syndrome, acute diarrhea, colitis and constipation [24], however, the effect of BC99 on physical performance, including muscle mass and strength, has not yet been evaluated. The purpose of this study is to evaluate the effects of BC99 on physical performance and verify whether BC99 would promote protein digestion in college athletes. The hypothesis of the present study is that BC99 can increase muscle mass and strength.
Materials and Method
2.1. Ethics Approval and Consent to Participate
The present study was conducted at the college of food and bioengineering, Henan University of Science and Technology, China. It was conducted in accordance with the Declaration of Helsinki, and the study protocol was approved by the Research Ethics Committee of Henan University of Science and Technology (NCT06307821), all participants signed an approved informed consent form prior to participation.
2.2. Participant Characteristics
Male college students who agreed to take probiotics were recruited for this study. Those who had been exposed to antibiotics within 3 months or had neuromuscular disorder, acute or chronic metabolic, respiratory or cardiovascular disorder and any other signs of ill health were excluded.
A total of 72 participants who met the enrollment criteria were randomly assigned to the placebo group (n = 36) and the probiotic group (n = 36). Participant characteristics are shown in Table 1. Differences between the placebo and probiotics groups were not statistically significant.
Powdered probiotics (3-g dose of milk protein concentrate with 6×109 CFU BC99/pack) and placebo (3-g dose of milk protein concentrate/pack) were obtained from the Wecare Probiotics Co., Ltd (Suzhou, China), and the two were indistinguishable by the naked eye. Participants were instructed to consume 8 pack (either placebo or probiotic) per day for the duration of the twelve-week supplementation period. The participants were periodically monitored to ensure full compliance with the treatment of this study.
2.3. Experimental Design
This study was conducted using a randomized, double-blind design. The study procedures are shown in Figure 1.
During 12-week intervention period, all participants have the identical exercise protocol because they were from same major.
2.4. Strength Performance and Body Composition
At the baseline and final stages of the study, strength tests were performed, the one repetition maximum (1RM) of the barbell bench press and squat was assessed, and the number of 80%RM repetitions was scored. body composition was assessed through InBody270 (Biospace Co., Ltd, Seoul, Korea) at the baseline and final stages of the study.
2.5. Enzyme Assays
At the final stages of the study, stool samples were collected using sterilized EP tubes, and stored at -80 ℃ until assayed. Frozen 100 mg stool samples were placed into 2 mL centrifuge tube with 1 mL PBS, and centrifuged at 3500g for 10 min at 4 °C after being homogenized. Then, the supernatant was harvested for analysis of digestive enzyme. The activities of digestive enzyme including pepsin, trypsin and lipase were determined according to assay kit instructions (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The enzyme activities were expressed as units per milligram or gram of protein, and the amount of protein of samples was determined using a commercially available bicinchoninic acid kit (Nanjing Jiancheng Bioengineering Institute).
2.6. Venous Blood Collection and Processing
At the final stages of the study, venous blood was collected after 12 h fasting, and plasma samples were obtained by centrifuging at 3000 rpm for 5 min at 4 ℃, then all samples were stored at -80 ℃ for further analysis. Blood were also taken at 3h post-ingestion, the plasma samples were obtained for amino acid analyses according to the above method.
2.7. Amino Acid Determination
The Agilent 1290 Infinity ultra-high performance liquid chromatography and QTRAP® 6500+ tandem triple quadrupole mass spectrometer (UHPLC-MS/MS, SCIEX Pte. Ltd., USA) were used for free amino acids analysis in plasma sample, and quantified by isotope internal standard. A test solution was prepared according to the following steps: 50 μL of plasma sample and 50 μL of mixed solution of isotope internal standards with 16 amino acid (50 μM) were added to 400 μL of methanol /acetonitrile (v/v, 1:1) solution, separately. Then the above solution was mixed on a vortex mixer for 1 min and left to stand for 1 h at 4 ℃ to allow protein complete precipitation. Subsequently, a resulting test solution was obtained via centrifugation at 12000 rpm for 20 min at 4°C.
Chromatographic separation was carried out on Waters ACQUITY UPLC BEH Amide column (2.1 mm ×150 mm, 1.7 µm). Column temperature was set as 35 °C. Mobile phase A consisted of MQ water containing 0.1% (v/v) formic acid and mobile phase B was a mixture of 0.1% formic acid-acetonitrile. The flow rate and injection volume were 300 µL/min and 2 µL, respectively. The gradient profile is as follows: Initial, 85% B; 1 min, 85% B; 3 min, 50% B; 11.5 min, 40% B; 12 min, 75% B.
The MS conditions were as follows: The electro-spray ionization source (ESI) was operated using the positive mode with capillary voltage of 4.5 kV and the ion source temperature of 500°C. The flow rates of ion source gas1, ion source gas2, and curtain gas were set at 55 L/h, 50 L/h and 30 L/h, respectively.
2.8. Biochemistry Analysis
The biochemical parameters in plasma including lactate dehydrogenase (LDH) and creatine kinase (CK) were measured using the commercial kit according to the manufacturer’s protocol (Nanjing Jiancheng Bioengineering Institute).
Myoglobin (Mb) and blood urea nitrogen (BUN) were measured using automatic biochemical analyzer at the Health Management Center, the Hospital of Henan University of Science and Technology,
2.9. Hormone Assays
The levels of testosterone and glucagon-like peptide-1 (GLP-1) in plasma were determined using commercially available kit (Nanjing Jiancheng Bioengineering Institute).
2.10. Statistical Analysis
Student’s t-test and the Mann Whitney test were used to compare quantitative variables between the two groups at baseline. All clinical and laboratory parameters were compared between the treatment groups (placebo and probiotic) and over time (baseline, and 12 week) using generalized linear models. p-values < 0.05 were considered statistically significant. All calculations were performed using IBM SPSS Statistics version 22.0.
3. Results
3.1. Effect of W. coagulans BC99 on strength performance
As shown in Table 2, strength performance was compared prior to and following the intervention. There were no significant differences in the bench press and squat between the two groups at the baseline. After a 12-week intervention, the number of 80%RM repetitions in bench press and squat in the probiotics group was significantly higher than that of the placebo group, no significant differences were observed between the two group after the 1RM test of bench press and squat. Moreover, compared with that in the baseline stage of study, the 1RM test of bench press and squat in both placebo and probiotics group in the final stage of study was higher (p < 0.05).
3.2. Effect of W. coagulans BC99 on Body Composition
Body composition changes were also compared prior to and following the intervention. After a 12-week intervention, compared with placebo group, probiotics groups showed a significant increase in muscle mass and fat free mass (p < 0.05), a significant decrease in fat mass (p < 0.05), and decreasing trend in body weight, however, the difference were not statistically significant (p = 0.07). Compared with that in the baseline stage, body composition in the final stage was not significant differences in the placebo group. However, compared with that in the baseline stage, muscle mass, fat free mass showed a significant increase (p < 0.05), and reduces in fat mass (p < 0.01) in the probiotics group. The results are presented in Table 3.
3.3. Effect of W. coagulans BC99 on Enzymatic Activity of Stool Samples
After a 12-week intervention, compared with those in the placebo group, the activities of pepsin, trypsin and lipase were significantly increase (p < 0.01) in the probiotics group. The results are presented in Figure 2.
3.4. Effect of W. coagulans BC99 on Plasma Concentration of Amino Acids
To investigate whether BC99 intervention increased the plasma concentration of amino acids by enhancing digestibility, 22 amino acids were analyzed (Table 4). The concentrations of alanine, arginine, Gamma- aminobutyric acid (GABA), glutamine, isoleucine, leucine, taurine, and tyrosine were significantly higher in the probiotic group than those in the placebo group (Figure 3). Compared with those in the placebo group, the concentrations of proline and valine tended to increase in the probiotic group, although the increase was nonsignificant. In particular, the concentrations of branched chain amino acids (BCAA) including isoleucine and leucine, and essential amino acid (EAA), were significantly higher in probiotic group than those in the placebo group. Compared to the placebo group, the concentrations of BCAA and EAA were 1.61 and 1.27 times higher in the probiotic group, respectively.
3.5. Effect of W. coagulans BC99 on Plasma BIOCHEMICAL parameters
After a 12-week intervention, compared with those of the placebo group, a statistically significant decrease in the activities of LDH (p < 0.05), CK (p < 0.01) and the concentration of BUN (p < 0.05) of the probiotics group was observed. There is no significant difference in the concentration of Mb between two groups. The results are presented in Table 5.
3.6. Effect of W. coagulans BC99 on plasma hormone level
After a 12-week intervention, compared with those in the placebo group, plasma testosterone and GLP-1 levels were significantly increased in probiotics group (Figure 4).
4. Discussion
In this study, we evaluate if BC99 may obtain optimal athletic performance by improving protein digestion, amino acid metabolism in male students of sport majors. As we expected, BC99 significantly increased strength performance in male college students. Notably, we observed that protein supplementation (placebo group) had also significant the improvement effect for strength performance from pretest to posttest. Several studies have showed that protein supplementation contribute to strength recovery and improvement of run performance and muscle mass [25,26]. In contrast, some studies found that protein supplementation had no efficacy for strength and performance [27-29]. These discrepancies can be associated with the digestibility of the proteins in the gastrointestinal tract besides protein types, protein dose and metabolic status of host. Overall, this study provide evidence that BC99 exhibited good efficacy in improvement of strength performance in the male students.
Skeletal muscle accounting for about 40% of body weight in human. Skeletal muscle mass affect muscle strength [30] and superior athletic performance [31]. Skeletal muscle is a highly plastic tissue, being able to respond to numerous environmental cues, such as exercise and nutrition [32]. Such response to environmental and physiological changing conditions, can lead to changes of skeletal muscle mass, function and composition. In health, muscle mass is maintained by precisely regulated muscle MPS and MPB [5]. In this study, the significant differences between the placebo and probiotics groups in muscle mass, fat mass and fat free mass were observed (Table 3), suggesting that increase in muscle quality may have contributed to improvement of strength performance after BC99 intervention.
It has been suggested that free essential amino acid (EAA) compositions stimulated muscle protein synthesis more than the same profile and amount of EAAs in dietary protein [33] and the higher protein digestion and amino acid absorption were more effective for stimulating MPS [33-35], indicating that enhancing of digestion rate of dietary protein can facilitate release of free amino acids into blood circulation system, increase the bioavailability of dietary protein, and stimulate MPS.
We subsequently evaluated activity of protease including pepsin and trypsin in stool samples and concentration of amino acid in plasma, the results showed that BC99 increased activity of pepsin and trypsin and concentration of amino acid, specifically, levels of BCAA and EAA were significantly increased. The increasing of plasma amino acid concentration seems to be, at least partly, attributed to the higher protein digestion after ingestion of BC 99 compared with single protein supplement. Clinic studies have shown that BCAA can promote MPS [36], and EAA can also effectively stimulate MPS [37]. Muscle protein synthesis is the metabolic basis for increased muscle mass and physical function [38]. Together with the above results, we concluded that probiotic BC99 could enhance muscle protein anabolism by facilitating digestion and absorption of dietary protein. In addition, we also found the ingestion of BC 99 increased activity of lipase, these results provided the evidence that BC 99 could also improve digestion of other nutrients beside protein in dietary, and BC99 supplementation may be an important strategy to improve digestive function for patient with indigestion.
BUN is a fatigue-related biomarker. BC99 reduced BUN level. LDH is an enzyme which catalyzes conversion of pyruvate to lactic acid [39], and creatine kinase can convert creatine into the high-energy phosphocreatine molecule [40]. The excessive production of LDH and creatine kinase leads to muscle damage [41], thus both them are biomarker of muscle damage. In this study, we observed that BC99 intervention attenuated CK and LDH levels, and decreased BUN, these results indicated that BC99 could improve endurance and reduced fatigue, and decreased muscle damage. Despite Mb is important in maintaining oxygen consumption and tension generation in skeletal muscle, the Mb content between the two group had no significant difference in this study.
A continuous turnover of protein (synthesis and breakdown) maintains the functional integrity and quality of skeletal muscle. Hormones are important regulators of this remodeling process. Anabolic hormones stimulate human muscle growth mainly by increasing protein synthesis or by decreasing protein breakdown. Both testosterone and glucagon-like peptide-one (GLP-1) are critical hormone relating to skeletal muscle protein metabolism. Subnormal testosterone levels led to unfavorable changes in body composition such as reducing of muscle mass and strength, as well as increasing of fat mass [42]. testosterone supplementation increased muscle mass by inhibiting protein degradation in older men [43]. Some evidence suggests that testosterone supplementation attenuates decreasing of muscle mass and grip strength [43,44], increased lean body mass and leg and arm muscle strength [45]. GLP-1 is a peptide hormone with 30-amino-acid, and affect muscle mass and its function [46,47]. GLP-1 can augment MPS in skeletal muscle of older humans [48].
Previous studies have suggested gut microbiota affect hormone level of host [49]. Probiotics possesses health promoting effect by increasing intestinal bacterial diversity and producing numerous metabolites [50]. Recent study showed that probiotics supplementary changed hormone levels and metabolic parameters of host [51]. In our study, administration of BC 99 for twelve weeks significantly increased testosterone and GLP-1 levels compared to that in the placebo group. It is possible that BC99 increase MPS by regulating hormone changes.
The study contributes to building the scientific foundation for using probiotics BC 99 as a potential tool for improving muscle protein synthesis and enhancing exercise capacity.
5. Conclusions
The present study demonstrated that BC99 intervention can significantly improve strength performance, body composition, digestion and absorption of amino acids, and regulate metabolism of skeletal muscle-related biochemical parameters and hormone levels. Therefore, probiotic BC99 has significant market value in various groups of individuals-from athletes to patients with obesity and even sarcopenia. However, further studies are required to understand dose response and the underlying mechanisms, and best practices to advance the application of BC99 as functional products.
Author Contributions
Designed study: SG and LC. Data collection: LC, YW, MG, YZ, JZ, ST, XL, PT and SG. Analyzed data: LC and YW. Prepared initial draft: LC and SG. Approved final version: all authors.
Funding
This study was supported by Major Science and Technology Special Projects in Henan Province (231100310200), Key Research and Development Projects in Henan Province (241111314200) and Key Research and Development Projects in Henan Province (221111111400).
Acknowledgments
The authors would like to thank all the volunteers participated in this study. Thanks also to Xiaojing Lei and Huili Liu for assistance with data collection and technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fuchs, C.J.; Hermans, W.J.H.; Holwerda, A.M.; Smeets, J.S.J.; Senden, J.M.; Kranenburg, J.; Gijsen, A.P.; Wodzig, W.K.H.W.; Schierbeek, H.; Verdijk, L.B.; et al. Branched-chain amino acid and branched-chain ketoacid ingestion increases muscle protein synthesis rates in vivo in older adults: a double-blind, randomized trial. Am. J. Clin. Nutr. 2019, 110, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Bröer, S.; Bröer, A. Amino acid homeostasis and signalling in mammalian cells and organisms. Biochem. J. 2017, 474, 1935–1963. [Google Scholar] [CrossRef]
- Lee, S.; Jo, K.; Jeong, H.G.; Yong, HI.; Choi, Y.S.; Kim, D.J.; Jung, S. ; Freezing-then-aging treatment improved the protein digestibility of beef in an in vitro infant digestion model. Food. Chem. 2021, 350, 129224. [Google Scholar] [CrossRef]
- Paulusma, C.C.; Lamers, W.H.; Broer, S.; Lamers, W.H.; Broer, S.; Graaf, S.F.J. Amino acid metabolism, transport and signalling in the liver revisited. Biochem. Pharmacol. 2022, 201, 115074. [Google Scholar] [CrossRef] [PubMed]
- Jang, L.G.; Choi, G.; Kim, S.W.; Kim, B.Y.; Lee, S.; Park, H. The combination of sport and sport-specific diet is associated with characteristics of gut microbiota: an observational study. J. Int. Soc. Sport. Nutr. 2019, 16, 21. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.E.; Goodpaster, B.; Wing, R.R.; Simoneau, J.A. Skeletal muscle fatty acid metabolism in association with insulin resistance, obesity, and weight loss. Am. J. Physiol. 1999, 277, E1130–E1141. [Google Scholar] [CrossRef] [PubMed]
- Ebert, S.M.; Dyle, M.C.; Bullard, S.A.; Dierdorff, J.M.; Murry, D.J.; Fox, D.K.; Bongers, K.S.; Lira, V.A.; Meyerholz, D.K.; Talley, J.J.; et al. Identification and Small Molecule Inhibition of an Activating Transcription Factor 4 (ATF4)-dependent Pathway to Age-related Skeletal Muscle Weakness and Atrophy. J. Biol. Chem. 2015, 290, 25497–25511. [Google Scholar] [CrossRef]
- Mohr, A.E.; Jäger, R.; Carpenter, K.C.; Kerksick, C.M.; Purpura, M.; Townsend, J.R.; West, N.P.; Black, K.; Gleeson, M.; Pyne, D.B.; et al. The athletic gut microbiota. J. Int. Soc. Sport. Nutr. 2020, 17, 24. [Google Scholar] [CrossRef]
- Maughan, R.J. Nutritional ergogenic aids and exercise performance. Nutr. Res. Rev. 1999, 12, 255–80. [Google Scholar] [CrossRef]
- Jäger, R.; Kerksick, C.M.; Campbell, B.I.; Cribb, P.J.; Wells, S.D.; Skwiat, TM.; Purpura, M.; Ziegenfuss, T.N.; Ferrando, A.A.; Arent, SM.; et al. International society of sports nutrition position stand: protein and exercise. J. Int. Soc. Sport. Nutr. 2017, 14, 20. [Google Scholar] [CrossRef]
- Holwerda, A.M.; Paulussen, K.J.M.; Overkamp, M.; Goessens, J.P.B.; Kramer, I.F.; Wodzig, W.K.W.H.; Verdijk, L.B.; Loon, L.J.C. Dose-dependent increases in whole-body net protein balance and dietary protein-derived amino acid incorporation into myofibrillar protein during recovery from resistance exercise in older men. J. Nutr. 2019, 149, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Blachier, F. Amino acid-derived bacterial metabolites in the colorectal luminal fluid: effects on microbial communication, metabolism, physiology, and growth. Microorganisms 2023, 11, 1317. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.Y. Dietary protein intake and human health. Food. Funct. 2016, 7, 1251–1265. [Google Scholar] [CrossRef]
- Sanders, ME.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: from biology to the clinic. Nat. Rev. Gastro. Hepat. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Zhang, R.; Zhou, M.; Tu, Y.; Zhang, N.F.; Deng, K.D.; Ma, T.; Diao, Q.Y. ; Effect of oral administration of probiotics on growth performance, apparent nutrient digestibility and stress-related indicators in Holstein calves. J. Anim. Physiol. Anim. Nutr. (Berl). 2016, 100, 33–38. [Google Scholar] [CrossRef]
- Keller, D.; Van Dinter, R.; Cash, H.; Farmer, S.; Venema, K. Bacillus coagulans GBI-30, 6086 increases plant protein digestion in a dynamic, computer-controlled in vitro model of the small intestine (TIM-1). Benef. Microbes. 2017, 8, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Grosicki, G.J.; Fielding, R.A.; Lustgarten, M.S. Gut microbiota contribute to age-related changes in skeletal muscle size, composition, and function: biological basis for a gut-muscle axis. Calcified. Tissue. Int. 2018, 102, 433–442. [Google Scholar] [CrossRef]
- Lustgarten, M.S. The role of the gut microbiome on skeletal muscle mass and physical function: 2019 update. Front. Physiol. 2019, 10, 1435. [Google Scholar] [CrossRef]
- Bindels, L.B.; Delzenne, N.M. Muscle wasting: The gut microbiota as a new therapeutic target? Int. J. Biochem. Cell. 2013, 45, 2186–2190. [Google Scholar]
- Rezaee, N.; Rahmani-Nia, F.; Delfan, M.; Ghahremani, R. Exercise training and probiotic supplementation effects on skeletal muscle apoptosis prevention in type-I diabetic rats. Life. Sci. 2021, 285, 119973. [Google Scholar] [CrossRef]
- Prokopidis, K.; Giannos, P.; Kirwan, R.; Prokopidis, K.; Giannos, P.; Kirwan, R.; Ispoglou, T.; Galli, F.; Witard, OC.; Triantafyllidis, KK.; et al. Impact of probiotics on muscle mass, muscle strength and lean mass: a systematic review and meta-analysis of randomized controlled trials. J. Cachexia. Sarcopeni. 2023, 14, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Soares, MB.; Martinez, R.C.R.; Pereira, E.P.R.; Balthazar, C.F.; Cruz, A.G.; Ranadheera, C.S.; Sant'Ana, A.S. The resistance of and strains with claimed probiotic properties in different food matrices exposed to simulated gastrointestinal tract conditions. Food. Res. Int. 2019, 125, 108542. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zhu, J.; Fang, S.; Zhao, B. Complete genome sequence of Heyndrickxia (Bacillus) coagulans BC99 isolated from a fecal sample of a healthy infant. Microbiol. Resour. Ann. 2024, 13, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, K.; Mohan, M.; Saudagar, P.; Sable, C.; Shinde, S.; Bedade, D. In vitro and in vivo evaluation of probiotic potential and safety assessment of Bacillus coagulans SKB LAB-19 (MCC 0554) in humans and animal healthcare. Regul. Toxicol. Pharmacol. 2022, 133, 105218. [Google Scholar] [CrossRef] [PubMed]
- Cintineo, H.P.; Arent, M.A.; Antonio, J.; Arent, S.M. Effects of protein supplementation on performance andrecovery in resistance and endurance training. Front. Nutr. 2018, 5, 83. [Google Scholar] [CrossRef]
- Huang, W.C.; Chang, Y.C.; Chen, Y.M.; Hsu, Y.J.; Huang, C.C.; Kan, N.W.; Chen, S.S. Whey protein improves marathon-induced injury and exercise performance in elite track runners. Int. J. Med. Sci. 2017, 14, 648–654. [Google Scholar] [CrossRef]
- Forbes, S.C.; Bell, G.J. Whey protein isolate supplementation while endurance training does not alter cycling performance or immune responses at rest or after exercise. Front. Nutr. 2019, 6, 19. [Google Scholar] [CrossRef]
- Williamson, E.; Kato, H.; Volterman, K.A.; Suzuki, K.; Moore, D.R. The effect of dietary protein on protein metabolism and performance in endurance-trained males. Med. Sci. Sport. Exer. 2019, 51, 352–360. [Google Scholar] [CrossRef]
- D'Lugos, A.C.; Luden, N.D.; Faller, J.M.; Akers, J.D.; McKenzie, A.I.; Saunders, M.J. Supplemental protein during heavy cycling training and recovery impacts skeletal muscle and heart rate responses but not performance. Nutrients 2016, 8, 550. [Google Scholar] [CrossRef]
- Janssen, I.; Baumgartner, R.N.; Ross, R.; Rosenberg, I.H.; Roubenoff, R. Skeletal muscle cutpoints associated with elevated physical disability risk in older men and women. Am. J. Epidemiol. 2004, 159, 413–421. [Google Scholar] [CrossRef]
- Tsekoura, M.; Kastrinis, A.; Katsoulaki, M.; Billis, E.; Gliatis, J. Sarcopenia and its impact on quality of life. Adv. Exp. Med. Biol. 2017, 987, 213–218. [Google Scholar] [PubMed]
- Stewart, C.; Rittweger, J. Adaptive processes in skeletal muscle: molecular regulators and genetic influences. J. Musculoskelet. Neuronal. Interact. 2006, 6, 73–86. [Google Scholar] [PubMed]
- Paddon-Jones, D.; Sheffield-Moore, M.; Katsanos, C.S.; Zhang, X.J.; Wolfe, R.R. Differential stimulation of muscle protein synthesis in elderly humans following isocaloric ingestion of amino acids or whey protein. Exp. Gerontol. 2005, 4, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Devries, M.C.; Phillips, S.M. Supplemental protein in support of muscle mass and health: advantage whey. J. Food. Sci. 2015, 80, A8–A15. [Google Scholar] [CrossRef] [PubMed]
- Dangin, M.; Boirie, Y.; Garcia-Rodenas, C.; Gachon, P.; Fauquant, J.; Callier, P.; Ballevre, O.; Beaufrere, B. The digestion rate of protein is an independent regulating factor of postprandial protein retention. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E340–E348. [Google Scholar] [CrossRef]
- Sharples, A.P.; Hughes, D.C.; Deane, C.S.; Saini, A.; Selman, C.; Stewart, C.E. Longevity and skeletal muscle mass: the role of IGF signalling, the sirtuins, dietary restriction and protein intake. Aging. Cell. 2015, 14, 511e. [Google Scholar] [CrossRef]
- Church, D.D.; Hirsch, K.R.; Park, S.; Kim, I-Y.; Gwin, J.S.; Pasiakos, S.M.; Wolfe, R.R.; Ferrando, A.A. Essential amino acids and protein synthesis: insights to maximizing the muscle and whole-body response to feeding. Nutrients 2020, 12, 3717. [Google Scholar] [CrossRef]
- Wolfe, R.R. The underappreciated role of muscle in health and disease. Am. J. Clin. Nutr. 2006, 84, 475–82. [Google Scholar] [CrossRef]
- Cho, H.D.; Lee, J.H.; Jeong, J.H.; Kim, J.Y.; Yee, S.T.; Park, S.K.; Lee, M.K.; Seo, KI. Production of novel vinegar having antioxidant and anti-fatigue activities from Salicornia herbacea L. J. Sci. Food. Agr. 2016, 96, 1085–1092. [Google Scholar] [CrossRef]
- Xu, C.; Lv, J.L.; Lo, Y.M.; Cui, S.W.; Hu, X.; Fan, M. Effects of oat β-glucan on endurance exercise and its anti-fatigue properties in trained rats. Carbohyd. Polym. 2013, 92, 1159–1165. [Google Scholar] [CrossRef]
- Oh, H.A.; Kim, D.E.; Choi, H.J.; Kim, N.J.; Kim, D.H. Anti-fatigue effects of 20(S)-protopanaxadiol and 20(S)-protopanaxatriol in mice. Biol. Pharm. Bull. 2015, 38, 1415–1419. [Google Scholar] [CrossRef] [PubMed]
- Kruse, R.; Petersson, S.J.; Christensen, L.L.; Kristensen, J.M.; Sabaratnam, R.; Ørtenblad, N.; Andersen, M.; Højlund, K. Effect of long-term testosterone therapy on molecular regulators of skeletal muscle mass and fibre-type distribution in aging men with subnormal testosterone. Metabolism 2020, 112, 154347. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, A.A.; Sheffield-Moore, M.; Yeckel, C.W.; Gilkison, C.; Jiang, J.; Achacosa, A.; Lieberman, S.A.; Tipton, K.; Wolfe, R.R.; Urban, R.J. Testosterone administration to older men improves muscle function: molecular and physiological mechanisms. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E601–E607. [Google Scholar] [CrossRef] [PubMed]
- Sinha-Hikim, I.; Cornford, M.; Gaytan, H.; Lee, M.L.; Bhasin, S. Effects of testosterone supplementation on skeletal muscle fiber hypertrophy and satellite cells in community-dwelling older men. J. Clin. Endocrinol. Metab. 2006, 91, 3024–3033. [Google Scholar] [CrossRef]
- Caminiti, G.; Volterrani, M.; Iellamo, F.; Marazzi, G.; Massaro, R.; Miceli, M.; Mammi, C.; Piepoli, M.; Fini, M.; Rosano, G.M. Effect of long-acting testosterone treatment on functional exercise capacity, skeletal muscle performance, insulin resistance, and baroreflex sensitivity in elderly patients with chronic heart failure a double-blind, placebo-controlled, randomized study. J. Am. Coll. Cardiol. 2009, 54, 919–927. [Google Scholar] [CrossRef]
- Katayama, T.; Takada, S.; Masaki, Y.; Kinugawa, S.; Matsumoto, J.; Furihata, T.; Fukushima, A.; Yokota, T.; Okita, K.; Tsutsui, H. The activation of glucagon-like peptide-1 improves the mitochondrial abnormalities in skeletal muscle and exercise intolerance in heart failure mice. J. Card. Fail. 2016, 22, S162–S162. [Google Scholar] [CrossRef]
- Hong, Y.; Lee, J.H.; Jeong, K.W.; Choi, C.S.; Jun, H.S. Amelioration of muscle wasting by glucagon-like peptide-1 receptor agonist in muscle atrophy. J. Cachexia. Sarcopenia. Muscle. 2019, 10, 903–918. [Google Scholar] [CrossRef]
- Tuddenham, S.; Sears, C.L. The intestinal microbiome and health. Curr. Opin. Infect. Dis. 2015, 28, 464–470. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Li, H.; Yu, Z.H.; Zhang, F.; Liang, S.; Liu, H.; Chen, H.; Lü, M.H. The Gut microbiome and sex hormone-related diseases. Front. Microbiol. 2021, 12, 711137. [Google Scholar] [CrossRef]
- Falcinelli, S.; Rodiles, A.; Hatef, A.; Picchietti, S.; Cossignani, L.; Merrifield, D.L.; Unniappan, S.; Carnevali, O. Influence of probiotics administration on gut microbiota core. J. Clin. Gastroenterol. 2018, 52, S50–S56. [Google Scholar] [CrossRef]
- Okuka, N.; Milinkovic, N.; Velickovic, K.; Polovina, S.; Sumarac-Dumanovic, M.; Minic, R.; Korčok, D.; Djordjevic, B.; Ivanovic, N.D. Beneficial effects of a new probiotic formulation on adipocytokines, appetite-regulating hormones, and metabolic parameters in obese women. Food. Funct. 2024, 15, 7658–7668. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The schedule of procedure of the present study.
Figure 2.
Changes of enzymatic activity of stool samples. A: Changes of pepsin activity. B: Changes of trypsin activity. C: Changes of lipase activity. Data are expressed as mean ± SD. ** means the difference is significant at the 0.01 level.
Figure 2.
Changes of enzymatic activity of stool samples. A: Changes of pepsin activity. B: Changes of trypsin activity. C: Changes of lipase activity. Data are expressed as mean ± SD. ** means the difference is significant at the 0.01 level.
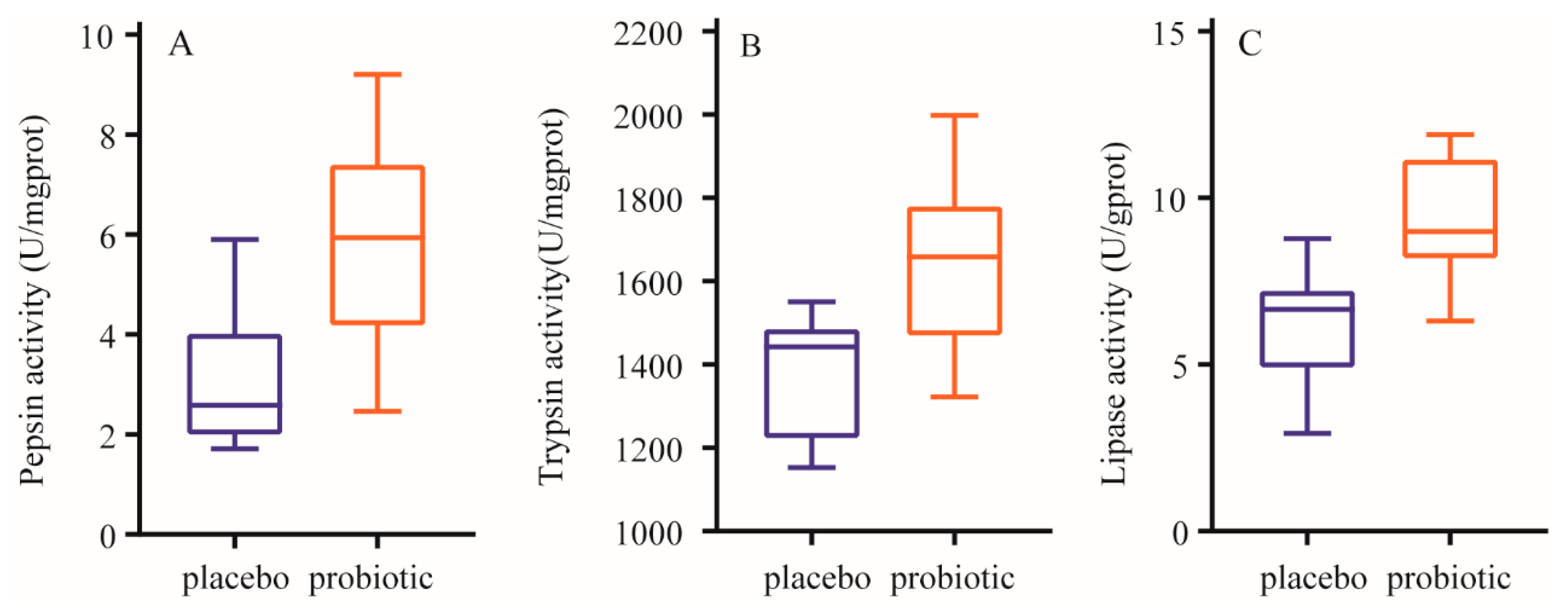
Figure 3.
Comparison of plasma concentrations of significantly changed amino acids.
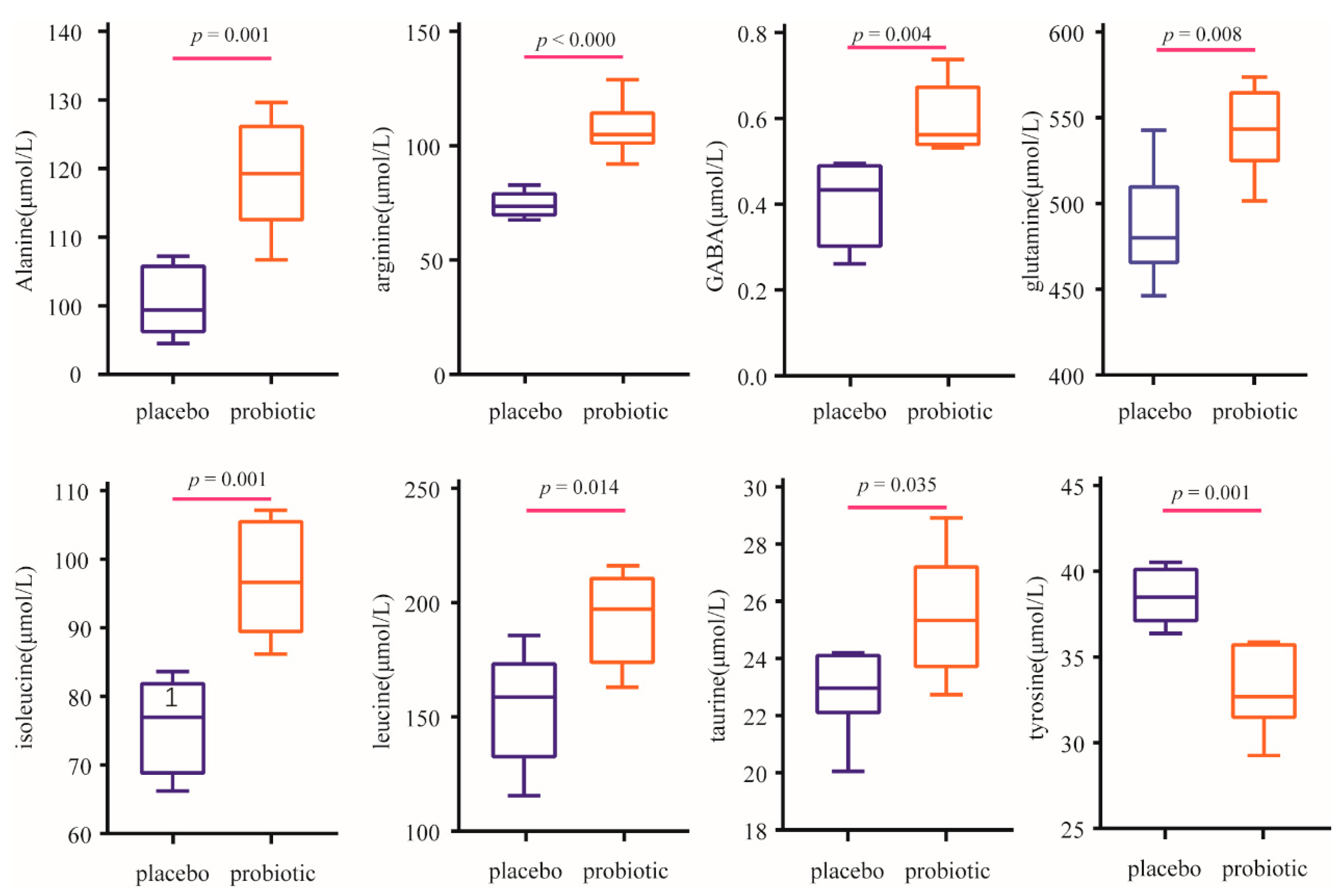
Figure 4.
Changes of plasma hormone level. A: Changes of plasma testosterone level. B: Changes of t glucagon-like peptide-1 level. Data are expressed as mean ± SD. * means the difference is significant at the 0.01 level.
Figure 4.
Changes of plasma hormone level. A: Changes of plasma testosterone level. B: Changes of t glucagon-like peptide-1 level. Data are expressed as mean ± SD. * means the difference is significant at the 0.01 level.
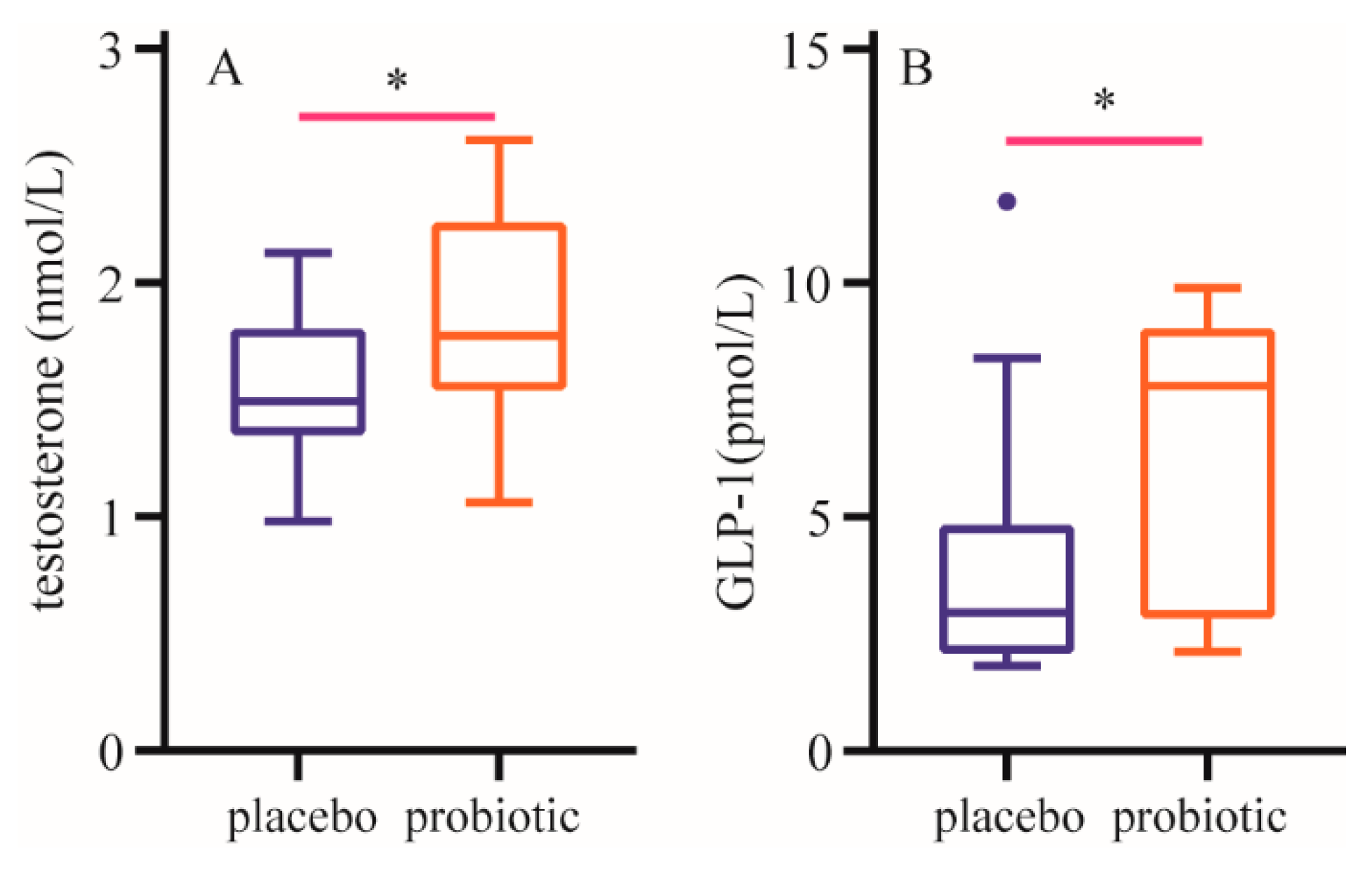

Table 1.
Participant characteristics.
| Placebo (n = 36) | probiotic (n = 36) | |
|---|---|---|
| Age (years) | 20.25±1.03 | 20.19±0.79 |
| Height (cm) | 1.79±5.94 | 179.25±5.16 |
| Weigh (kg) | 73.55±8.73 | 73.61±8.24 |
Values are presented as mean ± SD. There were not statistically significant differences between the placebo and probiotics groups.
Table 2.
Changes of strength performance in the placebo and probiotics groups at baseline and at the end of the 12-week intervention.
Table 2.
Changes of strength performance in the placebo and probiotics groups at baseline and at the end of the 12-week intervention.
| placebo | probiotics | ||||
|---|---|---|---|---|---|
| baseline | end | baseline | end | ||
| Bench press | 1-RM (kg) | 68.06±12.32 | 76.14±9.04* | 70.14±9.74 | 78.27±10.29* |
| 80%RM (times) | 11.42±3.54 | 10.07±2.36 | 11.31±3.09 | 11.41±1.84▲ | |
| Squat | 1-RM (kg) | 114.86±21.51 | 131.90±23.20* | 121.57±17.65 | 133.65±18.84* |
| 80%RM (times) | 10.33±5.39 | 10.91±2.96 | 10.54±4.14 | 13.04±4.01▲ | |
Values are presented as mean ± SD. * means the difference between baseline and study end is significant at the 0.05 level. means the difference between placebo and probiotics groups is significant at the 0.05 level.
Table 3.
Body composition variables in the placebo and probiotics groups at baseline and at the end of the 12-week intervention.
Table 3.
Body composition variables in the placebo and probiotics groups at baseline and at the end of the 12-week intervention.
| placebo | probiotics | |||
|---|---|---|---|---|
| baseline | end | baseline | end | |
| Muscle mass | 34.44±3.78 | 34.15±2.93 | 34.47±3.34 | 36.16±3.12*▲ |
| Fat mass | 11.01±3.41 | 9.90±3.55 | 10.91±3.07 | 8.31±2.18**▲ |
| Fat free mass | 61.28±4.78 | 59.89±4.05 | 61.25±5.69 | 64.77±4.59*▲ |
| Body weight | 73.55±8.73 | 69.94±6.95 | 73.61±8.24 | 70.08±7.18 |
| BMI | 22.26±2.14 | 21.67±2.12 | 22.55±2.13 | 21.95±1.94 |
Values are presented as mean ± SD. *means the difference between baseline and study end is significant at the 0.05 level. ** means the difference between baseline and study end is significant at the 0.01 level. means the difference between placebo and probiotics groups is significant at the 0.05 level.
Table 4.
Plasma concentrations of 22 amino acids (μmol/L).
| Amino acids | Placebo (n = 36) | probiotic (n = 36) | p value |
|---|---|---|---|
| Alanine | 100.45±5.02 | 119.32±8.39 | 0.001 |
| Arginine | 74.30±5.75 | 107.43±12.13 | 0.000 |
| Asparagine | 36.16±3.34 | 40.53±2.84 | 0.210 |
| Aspartic acid | 7.18±1.15 | 7.76±0.39 | 0.273 |
| Cysteine | 7.19±1.16 | 7.78±0.40 | 0.261 |
| GABA | 0.41±0.09 | 0.59±0.08 | 0.004 |
| Glutamate | 1589.43±251.86 | 1704.56±431.67 | 0.008 |
| Glutamine | 486.67±32.71 | 542.75±25.63 | 0.585 |
| Glycine | 107.29±10.67 | 105.34±11.19 | 0.764 |
| Histidine | 87.89±8.82 | 87.60±7.49 | 0.951 |
| Isoleucine | 75.89±7.26 | 90.68±14.11 | 0.001 |
| Leucine | 156.03±22.01 | 192.79±19.41 | 0.014 |
| Lysine | 229.17±48.41 | 240.14±25.52 | 0.108 |
| Methionine | 29.45±2.74 | 32.64±3.89 | 0.132 |
| Phenylalanine | 55.28±2.05 | 53.47±5.23 | 0.449 |
| Proline | 118.92±9.57 | 132.49±12.34 | 0.059 |
| Serine | 83.33±11.38 | 83.15±15.97 | 0.983 |
| Taurine | 22.85±1.51 | 25.51±2.19 | 0.035 |
| Threonine | 190.11±13.40 | 205.05±28.42 | 0.271 |
| Tryptophan | 38.75±5.11 | 37.54±4.69 | 0.678 |
| Tyrosine | 38.54±1.67 | 33.07±2.51 | 0.001 |
| Valine | 120.32±9.28 | 131.25±8.96 | 0.065 |
| BCAA | 350.49±39.83 | 564.18±29.06 | 0.000 |
| EAA | 963.65±65.47 | 1222.31±65.83 | 0.000 |
| Total AA | 3643.83±254.01 | 4092.35±495.51 | 0.077 |
Data are expressed as mean ± SD.
Table 5.
Changes of plasma biochemical parameters in the placebo and probiotics groups.
| Placebo (n = 36) | probiotic (n = 36) | |
|---|---|---|
| LDH(U/L) | 2036.90±273 | 1327.18±368* |
| CK(U/mL) | 0.19±0.04 | 0.09±0.03** |
| BUN(mmol/L) | 5.44±1.05 | 4.96±0.89* |
| Mb(ng/mL) | 1.74±0.33 | 1.60±0.28 |
Data are presented as mean ± SD. *means the difference between baseline and study end is significant at the 0.05 level. ** means the difference between baseline and study end is significant at the 0.01 level.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



