
Submitted:
30 October 2024
Posted:
30 October 2024
You are already at the latest version
Abstract
Lactococcus spp. has emerged as a pathogen that is affecting global aquaculture, with L. garvieae, L. petauri, and L. formosensis causing piscine lactococcosis. While antimicrobials are commonly used to treat diseases in aquaculture, reports of antimicrobial resistance in fish isolates are increasing. However, little is known about the susceptibility patterns of Lactococcus spp. strains isolated from native fish species in Brazil. This study aimed to assess the antimicrobial susceptibility of these strains and establish a provisional epidemiological cut-off value for L. garvieae using the normalized resistance interpretation approach. A total of 47 isolates were tested: 17 L. garvieae, 24 L. petauri, and 6 L. formosensis. The isolates were classified as wild-type or non-wild-type (NWT) based on inhibition zone diameters. Isolates classified as NWT for three or more antimicrobial classes were considered multidrug-resistant, and the multiple antibiotic resistance (MAR) index was calculated. The results revealed heterogeneity in antimicrobial resistance profiles, with higher resistance to trimethoprim/sulfamethoxazole and norfloxacin. Resistance to other antimicrobials, including florfenicol and oxytetracycline (approved for use in Brazil), varied according to the bacterial species. Lactococcus petauri (87.5%) and L. formosensis (66.7%) showed the highest multidrug resistance, compared to L. garvieae (11.7%), along with higher MAR index values. These findings suggest that multidrug-resistant strains could pose future challenges in the production of native species, underscoring the need for ongoing monitoring of antimicrobial resistance and responsible use of antimicrobials in aquaculture.
Keywords:
disk diffusion
; epidemiological cut-off values
; fish
; piscine lactococcosis
1. Introduction
Piscine lactococcosis is considered an emerging bacterial disease for fish farming worldwide [1], and the number of hosts in which Lactococcus garvieae, L. petauri and L. formosensis have been detected has expanded [2,3,4,5,6,7]. The disease is currently a significant health challenge for Oncorhynchus mykiss and Oreochromis niloticus production, causing high mortality rates and significant economic losses [8,9,10].
One of the main methods for controlling outbreaks of bacterial diseases in fish farms is antibiotic therapy [11]. However, there are already reports of Lactococcus spp. strains becoming resistant to the main drugs used in aquaculture [12,13]. The indiscriminate use of antimicrobials has been reported by producers and technicians from different fish farms, which can result in bacterial resistance to specific drug [14]. As a result, a substance already used by a producer may no longer be effective in treating bacteriosis, thereby necessitating the use of another antibiotic. Additionally, it is worth mentioning that the rate of approval for new drugs is slower than the evolution of bacterial resistance, leading producers to use off-label drugs [15].
One way to monitor antimicrobial resistance in lactococcosis-causing bacteria is through the use of laboratory methods such as disk diffusion [10] and broth dilution [16] methods. The former is considered an inexpensive, reliable and simple technique that can be easily applied in a laboratory routine, while in comparison the latter is technically demanding and labor-intensive [17]. However, in Brazil, few studies have evaluated these methodologies for testing Lactococcus spp. strains, whether using isolates from terrestrial mammals or aquatic animals. In Brazil, the disk diffusion assay has been performed for isolates of L. petauri from farmed Oreochromis niloticus, and resistance for some isolates to amoxicillin, erythromycin, florfenicol and norfloxacin was identified. In addition, all the isolates evaluated were considered resistant to trimethoprim/sulfamethoxazole [10]. Bacteria of the genus Lactococcus have also been isolated from native Brazilian fish species [2], and little is known about the use of antimicrobials in these species and their antimicrobial susceptibility profiles.
The main problem in determining the sensitivity of bacterial fish pathogens to antimicrobials is the lack of reference values. Without these values, it is not possible to determine whether an isolate is sensitive or resistant. There are no internationally recognized epidemiological cut-off values for disk diffusion data for Lactococcus spp. strains in the Clinical Laboratory Standards Institute (CLSI) or the European Committee on Antimicrobial Susceptibility Testing (EUCAST) guidelines. Previous studies have generated provisional epidemiological cut-off values for L. petauri from disk diffusion zone data using the normalized resistance interpretation (NRI) method [10] for L. garvieae and L. petauri from minimum inhibitory concentration data using NRI and ECOFFinder approaches [16]. Nevertheless, for disk diffusion zone data, there are no reports of established cut-off values in the literature for L. garvieae and L. formosensis.
Therefore, the aim of this study was to evaluate the susceptibility profile of Lactococcus spp. strains obtained from native Brazilian fish species to different antimicrobials and to calculate the provisional epidemiological cut-off values (pECVs) for Lactococcus garvieae.
2. Materials and Methods
2.1. Bacterial Strains and Identification
A total of 47 Lactococcus spp. strains (n = 6 L. formosensis, n = 17 L. garvieae and n = 24 L. petauri) were used in this study. The isolates were obtained from eleven native fish species (Arapaima gigas, Brycon amazonicus, Cichla sp., Colossoma macropomum, Hoplias macrophtalmus, Hoplias malabaricus, Lophiosilurus alexandri, Phractocephalus hemioliopterus, Pseudoplatystoma corruscans, Pseudoplatystoma fasciatum and a hybrid of Pseudoplatystoma) originating from free-living fish or commercial farms, between 2012 and 2024, in six Brazilian states (Amazonas, Bahia, Mato Grosso do Sul, Minas Gerais, Pará and São Paulo) (Table 1) [2,18,19,20,21,22]. These isolates were obtained through routine laboratory diagnosis of bacterial diseases in fish conducted by the Laboratory of Aquatic Animal Diseases (Veterinary School, Federal University of Minas Gerais, Belo Horizonte, Brazil), Laboratory of Applied Microbiology of Aquatic Organisms (Nilton Lins University, Manaus, Brazil), Laboratory of Microbiology and Parasitology of Aquatic Organisms (Aquaculture Center of São Paulo State University, São Paulo, Brazil), and Fisheries Institute (São Paulo, Brazil). Of these, 11, originating from Arapaima gigas (n = 7), Cichla sp. (n = 1), Hoplias malabaricus (n = 1) and Pseudoplatystoma sp. (n = 2), were recovered through bacterial examination after the recent disease outbreak. Furthermore, all the selected isolates were identified to species level using matrix-assisted laser desorption ionization time-of-flight (MALDI-TOF) mass spectrometry (Bruker Daltonics, Germany) [20] with the Bruker MALDI Biotyper database (v13.0.0.2) followed by gyrB sequencing [23]. The isolates were stored at -70 °C in BHI broth with 15% glycerol until use.
2.2. Susceptibility Testing
Disk diffusion tests against Lactococcus spp. were carried out according to the protocol provided in the CLSI guideline VET03, with adaptations recommended for bacteria of the genus Streptococcus (Group 4) [24]. The disks used contained 10 µg amoxicillin, 15 µg erythromycin, 30 µg florfenicol, 10 µg neomycin, 10 µg norfloxacin, 30 µg oxytetracycline and 1.25/23.75 µg trimethoprim/sulfamethoxazole. The disks were obtained from a commercial company (Oxoid, United Kingdom).
The selected isolates (Table 1) were thawed, inoculated onto Man Rogosa & Sharpe (MRS, Merck, Germany) agar, and incubated at 28 ºC for 48 h. After incubation, colonies were collected and suspended in sterile saline solution until they reached an absorbance of between 0.08 and 0.13 (DO625) nm using a spectrophotometer (Spectrum, China). Muller-Hinton agar enriched with 5% defibrinated sheep blood was inoculated with the bacterial suspension using sterile swabs. Then, antimicrobial disks were placed on the agar and the plates were incubated at 28 ºC for 24 h. Additionally, the quality control reference strains Escherichia coli ATCC 25922 and Aeromonas salmonicida subsp. salmonicida ATCC 33658 were grown on blood agar, incubated at 28 ºC for 24 h and subjected to the same experimental conditions described above as recommended by the CLSI for this method. All the procedures were performed in duplicate. The diameter of the inhibition zone of all the isolates was measured.
2.3. Calculation of Provisional Lactococcus garvieae Epidemiological Cut-Off Values
As there is no established zone diameter cut-off for Lactococcus garvieae generated by a standard method, this study calculated the pECV for each antimicrobial agent tested using the automatic normalized resistance interpretation (NRI) method (www.bioscand.se/nri) from the inhibition zone data that was generated [25,26]. The isolates were then classified as wild-type (WT) or non-wild-type (NWT) [27]. To meet the minimum requirements of the NRI method [28], the disk diffusion data of the Lactococcus garvieae strains isolated from Oreochromis niloticus (n = 3), Trichogaster lalius (n = 1), and Xiphophorus maculatus (n = 1) from the Laboratory of Aquatic Animal Diseases culture collection were included in the calculation of the pECVs (Supplementary Table 2).
2.4. Data Analysis
Lactococcus petauri strains were classified as WT or NWT according to the previously established pECV (Egger et al. 2023). However, as Lactococcus formosensis has no number of suggested observations to set a reliable pECV calculation, the inhibition zone data were shown as maximum, minimum, mean and standard deviation values. Regardless of the bacterial species, bacteria that did not present an inhibition zone and did not have a defined ECV were considered NWT for the antimicrobials. R software v.4.3.1 [29] and RAWGraphs v2.0 [30] were used for data visualization. Isolates classified as NWT for at least three classes of antimicrobials were considered to be multidrug-resistant bacteria [31]. The multiple antibiotic resistance (MAR) index was also calculated [32].
3. Results
3.1. Bacterial Identification
From the sequencing of the gyrB gene, Lactococcus formosensis (n = 1, Arapaima gigas), L. garvieae (n = 1, Cichla sp.; n = 1, Hoplias malabaricus; n = 1, Pseudoplatystoma sp.) and L. petauri (n = 6, Arapaima gigas; n = 1, Pseudoplatystoma sp.) were detected from the current disease outbreaks in native fish species (Table 1). The gyrB gene sequences of these isolates were included in the NCBI database.
The remaining isolates used in this study were identified at the species level in a previous study [2].
3.2. Quality Control
The reference strains E. coli ATCC 25922 and Aeromonas salmonicida subsp. salmonicida ATCC 33658 presented inhibition zone diameters within the acceptable ranges established by the CLSI (Table 2).
3.3. Antimicrobial Susceptibility for Lactococcus formosensis
The disk diffusion assay for L. formosensis strains exhibited zones ranging between 6 mm and 30 mm (Table 2 and Supplementary Table 1). All the isolates were categorized as NWT (no observation of inhibition zone) for trimethoprim/sulfamethoxazole and norfloxacin (Figure 1). A total of one and four strains were categorized as NWT for florfenicol and oxytetracycline, respectively (Figure 1). For these antimicrobials, all the isolates from Arapaima gigas were categorized as NWT for oxytetracycline, and the LG91-23 strain (from Pseudoplatystoma sp.) was NWT for both drugs (Figure 2). Since there is no pECV for Lactococcus formosensis, it is not possible to determine whether the other isolates are resistant to other antimicrobials. In addition, four isolates were classified as multidrug-resistant. The MAR index of the isolates varied between 0.285 and 0.571 (Figure 3, Supplementary Table 1).
3.4. Antimicrobial Susceptibility for Lactococcus garvieae
The disk diffusion assay for L. garvieae strains exhibited zones raging between 6 mm and 31 mm (Table 2 and Supplementary Table 1). The distribution of the inhibition zones obtained from all the evaluated isolates are shown in Supplementary Figure 1. The calculated pECVs for the antimicrobials are presented in Table 2. A total of eight and fifteen isolates (Figure 1) presented a zone of complete inhibition of 6 mm for norfloxacin and trimethoprim/sulfametoxazole, respectively, preventing the establishment of the ECV for these antimicrobials. However, these isolates were categorized as NWT. Based on the calculated pECVs, all the isolates were classified as WT for amoxicillin, erythromycin and neomycin. One and five isolates were classified as NWT for florfenicol and oxytetracycline, respectively, especially those strains isolated from Pseudoplatystoma sp. (Figure 2). In addition, two isolates were classified as multidrug-resistant. The MAR index of the isolates varied between 0.00 and 0.428 (Figure 3, Supplementary Table 1).
3.5. Antimicrobial Susceptibility for Lactococcus petauri
The disk diffusion assay for the L. petauri strains exhibited zones ranging between 6 mm and 31 mm (Table 2 and Supplementary Table 1). All the isolates were classified as WT for neomycin. A total of 1, 8, 11, 22, 22 and 23 isolates were classified as NWT for amoxicillin, erythromycin, florfenicol, oxytetracycline, norfloxacin and trimethoprim/sulfametoxazole, respectively (Supplementary Table 1). A resistance phenotype for florfenicol and oxytetracycline was observed in all the native Brazilian fish species in which L. petauri was isolated (Figure 2). Twenty-one isolates were classified as multidrug-resistant. The MAR index varied between 0.285 and 0.857 (Figure 3, Supplementary Table 1).
4. Discussion
Currently, antimicrobial resistance is one of the biggest threats to public health [33], especially with the emergence of multidrug-resistant strains [34]. The antimicrobial susceptibility profile in lactococcosis-causing bacteria strains has been studied by several different institutions and researchers using various techniques, such as disk diffusion [35,36], broth dilution [12,37] and the Etest [38]. Studies have shown that most of the isolates evaluated are resistant to ampicillin [36,39], florfenicol [40], flumequine [35], nalidixic acid [39,41,42], norfloxacin [40], tetracycline [13] and trimethoprim/sulfamethoxazole [35,39,41,43]. Although detected at lower percentages, there are also records of resistance to amoxicillin (16-23%), bacitracin (42%), ciprofloxacin (4%), chloramphenicol (18%), enrofloxacin (33-67%), erythromycin (16-52%), kanamycin (33%), oxytetracycline (4-44%) and streptomycin (33%) [13,35,36,39,42,43].
In the scientific literature, it is possible to observe heterogeneity in the antimicrobial resistance profiles for Lactococcus spp. strains, which may be related to the different species within the genus. This is because most of the articles, including some recent ones, did not perform the correct taxonomic classification of the isolates, which is currently recommended [44]. Only three studies assessed the antimicrobial resistance profile after correct species identification, using the disk diffusion [10,45] and broth dilution [16]. Furthermore, Öztürk et al. [16] suggest that this heterogeneity is related to the overuse or misuse of antimicrobials at the farm level and the lack of established susceptibility cut-off values for each of the three species that cause piscine lactococcosis. Here, we evaluated the antimicrobial resistance profiles of L. formosensis, L. garvieae and L. petauri strains isolated from native fish species in Brazil using disk diffusion susceptibility testing and established pECVs for five out of seven antimicrobials for L. garvieae strains.
Regardless of the bacterial species evaluated in our study, the trimethoprim/sulfametoxazole resistance phenotype stood out (L. formosensis = 100%, L. garvieae = 88.2%, L. petauri = 95.8%) (Figure 1). Resistance to this drug has previously been reported in the literature for Lactococcus spp. strains isolated from Oncorhynchus mykiss, Dicentrarchus labrax and Oreochromis niloticus [6,10,35,39,40,41,43]. For the other drugs, interspecies variation has been observed.
Unfortunately, due to the limited number of isolates identified as L. formosensis in our study, it was not possible to establish a pECV and, therefore, classification as WT or NWT could not be performed. However, we considered those isolates for which no inhibition zones were observed for the antimicrobials tested to be NWT. Thus, in addition to trimethoprim/sulfametoxazole, all the isolates were considered NWT for norfloxacin. This result is in disagreement with the study conducted by Lin et al. [46], in which all the L. formosensis strains isolated from milk samples of a cow with clinical mastitis were susceptible to quinolones via broth dilution testing. We also did not observe the formation of inhibition zones in four isolates (three from Arapaima gigas and one from Pseudoplatystoma sp.) for oxytetracycline, nor in one Pseudoplatystoma sp. isolate (LG91-23) for florfenicol (Figure 2a). Chan et al. [45] evaluated susceptibility using the disk diffusion method for an L. formosensis strain obtained from a human with bacteremia and found that the isolate was susceptible to tetracycline. There is no mention in the literature regarding resistance profiles to florfenicol, regardless of the host evaluated; however, a previous study demonstrated resistance to another amphenicol, chloramphenicol, for all the isolates evaluated [46]. Although we cannot determine susceptibility for other drug classes, the literature mentions L. formosensis resistance to aminoglycosides and macrolides, and susceptibility to β-lactams [45,46]. If we consider the pECV of L. garvieae and L. petauri from this study, all the isolates would be classified as WT for amoxicillin, erythromycin, florfenicol and neomycin, which would corroborate the previous information. Additionally, the AM-LG05 strain would be classified as NWT for oxytetracycline, increasing the number of multidrug-resistant isolates. It was possible to observe that the antimicrobial resistance profile was similar among the Arapaima gigas isolates, as all the isolates share the same geographic origin.
For the L. petauri strains, we compared the results using the previously established pECV. All the isolates evaluated were classified as WT for neomycin, thereby corroborating with Egger et al. [10]. For amoxicillin and erythromycin, our isolates demonstrated a low frequency of NWT detection, 4.1% and 33.3%, respectively, when compared to other antimicrobials. A previous study demonstrated resistance of 6% and 25% for L. petauri strains isolated from Oncorhynchus mykiss for erythromycin and amoxicillin, respectively [16]. For isolates obtained from Oreochromis niloticus, the NWT percentages were lower, around 3% for both antimicrobials [10]. A high percentage of isolates classified as NWT for norfloxacin was observed in our study (91.7%) compared to the results obtained in Oreochromis niloticus (16.75%) [10]. Regarding florfenicol and oxytetracycline, 11.4% and 91.6% of the isolates were classified as NWT, respectively. In Oncorhynchus mykiss and Oreochromis niloticus isolates, NWT values of 0% and 12.5% for oxytetracycline and 3.4% to 9.4% for florfenicol have been observed [10,16]. The resistance profile was similar among the Pseudoplatystoma sp. isolates from the states of Mato Grosso and Minas Gerais, as well as most of the Arapaima gigas isolates from Bahia (Supplementary Table 1). However, for isolates obtained from this latter fish species in the northern region of Brazil, individual variation was detected, as were the cases with Colossoma macropomum and Brycon amazonicus isolates (Figure 2c).
There are no studies that have standardized cut-off values, even provisional ones, for L. garvieae strains following its correct taxonomic identification. Therefore, our study is the first to do so. However, we emphasize that the ECVs presented here are provisional. To generate ECVs that are relevant to disk diffusion data for L. garvieae, a larger number of isolates (over 100 observations) from at least five different laboratories would be required [28]. As previously mentioned, all the isolates evaluated were classified as being WT for amoxicillin, erythromycin and neomycin. In contrast, isolates from Oncorhynchus mykiss exhibited varying susceptibility to these antimicrobials. Approximately 2.6%, 24.4% and 6.4-11.5% of the isolates were resistant to amoxicillin, erythromycin and aminoglycosides, respectively [16]. A total of 47% of the isolates from native fish species in Brazil were considered to be NWT for norfloxacin, in contrast to previous studies that reported low (7.7%) or no resistance to quinolones [16,45]. A total of 5.8% and 29.4% of the isolates were classified as NWT for florfenicol and oxytetracycline, respectively. However, the literature reports resistance rates of 26.9% for florfenicol and 17.9% for oxytetracycline [16]. The antimicrobial resistance profile observed in our study for the L. garvieae strains was not consistent among the aquatic host species analyzed or with the origin of the isolates, demonstrating a heterogeneous profile (Figure 2b).
Our study showed that the L. garvieae strains tend to be more sensitive to antimicrobials when compared to the L. formosensis and L. petauri strains. Furthermore, the proportion of L. garvieae isolates with a MAR index greater than 0.3 (11.7%) was lower than that found in L. formosensis (66.7%) and L. petauri (87.5%) (Figure 3). The most efficient measure to control bacterial diseases is the use of antimicrobials [11]. However, since the isolates evaluated in this study were classified as multi-resistant to several antimicrobials (Supplementary Table 1), treating piscine lactococcosis in native Brazilian fish species becomes challenging. Unfortunately, little is known about the use of antimicrobials in native fish species in Brazil. However, prophylactic and metaphylactic use of antimicrobials, especially oxytetracycline, in larviculture of native species and during the feeding training of carnivorous species like Pseudoplatystoma sp. and Arapaima gigas has been reported by producers and technicians in the country. Additionally, it is known that commercial fish farming in Brazil involves off-label use of amoxicillin, enrofloxacin and norfloxacin [15,47]. In the central-western region of Brazil, fluoroquinolones, especially norfloxacin and enrofloxacin, intended for the treatment of cattle, have also been used off-label in Pseudoplatystoma sp.. Therefore, the widespread use of these antimicrobials may have contributed to the increase in resistance among Lactococcus spp. strains. It is also worth mentioning that when MAR index values exceed 0.2 (in our case, over 0.3 due to the number of antibiotics tested), a high environmental risk of spreading antimicrobial resistance is predicted [32]. In this context, the shared production of native fish species and Oreochromis niloticus could pose a risk of transmitting antimicrobial-resistant Lactococcus spp. strains, or it could enable L. petauri isolates from Oreochromis niloticus to acquire resistance genes in this production environment, resulting in an unsatisfactory therapeutic approach during disease outbreaks. Therefore, monitoring of antimicrobial resistance in Lactococcus spp. strains becomes essential.
Sun et al. [48] report that acquired resistance in microorganisms occurs for two reasons: the natural resistance of bacteria to certain antimicrobials and acquired resistance due to continuous exposure to antimicrobials. Once the bacteria becomes resistant, this resistance can be transferred to the other bacterial species through horizontal gene transfer [49]. Furthermore, some L. garvieae strains carry these antimicrobial resistance genes on transferable R plasmids [35]. The acquisition and transfer of antimicrobial resistance genes have been considered to be responsible for the spread and distribution of antimicrobial resistance [16]. Previous studies indicate a high prevalence of antimicrobial resistance genes in Lactococcus spp. isolates from Oncorhynchus mykiss [13,16,35]. There is no description of the detection of resistance genes in Lactococcus spp. strains from Brazilian fishes. However, given the higher percentage of multi-resistant isolates, future studies should be conducted to identify resistance genes, particularly those encoding antimicrobial resistance, using genomic tools.
In Brazil, only florfenicol and oxytetracycline are approved antimicrobial agents for use in aquaculture [50]. Both antimicrobials act against Gram-negative and Gram-positive bacteria; they are bacteriostatic drugs that work by binding to bacterial ribosomal subunits and inhibiting protein synthesis [49]. However, neither of these antimicrobials has been evaluated for their therapeutic efficacy in fish either naturally or experimentally infected with Lactococcus spp. in Brazil. Nevertheless, the administration of oxytetracycline in Oncorhynchus mykiss in Greece was reported to be unsatisfactory in both prophylactic and therapeutic treatments [51]. The circulation of florfenicol- and oxytetracycline-resistant strains in Brazilian fish farms could become a significant health issue when producing native species. The oral administration of amoxicillin, erythromycin and flumequine did not yield significant results in the treatment of Oncorhynchus mykiss and Dicentrarchus labrax with lactococcosis [6,51]. However, based on the antimicrobial susceptibility tests from our study, amoxicillin and neomycin could be tested for their therapeutic efficacy against piscine lactococcosis in Brazil.
5. Conclusions
When the pECVs of L. garvieae strains and the antimicrobial resistance profiles of L. garvieae, L. formosensis and L. petauri were assessed, a higher percentage of resistance to various antimicrobials was observed among the evaluated isolates, especially for L. petauri, including multidrug-resistant strains. This is quite different from what has been observed in Oreochromis niloticus farms in Brazil, thus making it essential to monitor the susceptibility of the isolates and raise awareness among producers about the correct use of antibiotics.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Histograms of the inhibition zone for Lactococcus garvieae strains against amoxicillin (AMO), erythromycin (ERY), florfenicol (FLO), neomycin (NEO), norfloxacin (NOR), oxytetracycline (OXY) and trimethoprim/sulfamethoxazole (SXT); Table S1: Inhibition zones diameters (mm) of antimicrobial agents against Lactococcus spp. strains determined using disk diffusion susceptibility assay and MAR index calculated per isolate; Table S2: Inhibition zones diameters (mm) of antimicrobial agents against Lactococcus garvieae strains used to satisfy the minimum requirements of the NRI method.
Author Contributions
Conceptualization: AECR, ACCB, HCM, HCPF and GCT; Methodology: AECR, ACCB, HCM, CRMSM, JMT, HCPF and GCT; Formal analysis: AECR, ACCB and HCM; Investigation: AECR, ACCB, HCM, CRMSM, LFFN, FP, SUG, CAGL, HCPF and GCT; Resources: FP, SUG, CAGL, HCPF and GCT; Data curation: LFFN and GCT; Writing—original draft preparation, AECR and GCT; Writing—review and editing, AECR, ACCB, HCM, CRMSM, JMT, LFFN, FP, SUG, CAGL, HCPF and GCT; Visualization: GCT; Supervision: SUG, HCPF and GCT; Project administration: GCT; Funding acquisition: HCPF and GCT. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) through the PROCAD/Amazônia (grant number 88881.200614/2018-01) and PDPG-CAPES, Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG, grant numbers APQ-01227-22, APQ-04309-22 and PPM-00779-18), and Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM, grant number 01.02.016301.03071/2022-11).
Ethical Approval
No ethical approval was required as the research in this article is related to microorganisms.
Data Availability Statement
The gyrB gene sequences of the Lactococcus spp. strains isolated from the native Brazilian fish species were included in the NCBI database as follows: L. formosensis – 167/23-09: PQ529765; L. garvieae – LG114-23: PQ529769, LG116-23: PQ529771, LG119-24: PQ529773; L. petauri – 167/23-03: PQ529760, 167/23-04: PQ529761, 167/23-05: PQ529762, 167/23-07: PQ529763, 167/23-08: PQ529764, 167/23-10: PQ529766, LG117-23: PQ529772.
Acknowledgments
The authors gratefully acknowledge the support provided by Maria José Tavares Ranzani-Paiva for conducting this study.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
References
- Altinok, I.; Ozturk, R.C.; Ture, M. NGS Analysis revealed that Lactococcus garvieae Lg-Per was Lactococcus petauri in Türkiye. J. Fish Dis. 2022, 45, 1839–1843. [Google Scholar] [CrossRef] [PubMed]
- Barbanti, A.C.C.; do Rosário, A.E.C.; da Silva Maia, C.R.M.; Rocha, V.P.; Costa, H.L.; Trindade, J.M.; Nogueira, L.F.F.; Rosa, J.C.C.; Ranzani-Paiva, M.J.T.; Pilarski, F.; et al. Genetic characterization of lactococcosis-causing bacteria isolated from Brazilian native fish species. Aquaculture 2024, 593, 741305. [Google Scholar] [CrossRef]
- Bondavalli, F.; Colussi, S.; Pastorino, P.; Zanoli, A.; Bezzo Llufrio, T.; Fernández-Garayzábal, J.F.; Acutis, P.L.; Prearo, M. First report of Lactococcus petauri in the pumpkinseed (Lepomis gibbosus) from Candia Lake (Northwestern Italy). Fishes 2024, 9. [Google Scholar] [CrossRef]
- Khoo, L.H.; Austin, F.W.; Quiniou, S.M.A.; Gaunt, P.S.; Riecke, D.K.; Jacobs, A.M.; Meals, K.O.; Dunn, A.W.; Griffin, M.J. Lactococcosis in silver carp. J. Aquat. Anim. Health 2014, 26, 1–8. [Google Scholar] [CrossRef]
- Abraham, T.; Yazdi, Z.; Littman, E.; Shahin, K.; Heckman, T.I.; Quijano Cardé, E.M.; Nguyen, D.T.; Hu, R.; Adkison, M.; Veek, T.; et al. Detection and virulence of Lactococcus garvieae and L. petauri from four lakes in Southern California. J. Aquat. Anim. Health 2023, 35, 187–198. [Google Scholar] [CrossRef]
- Salogni, C.; Bertasio, C.; Accini, A.; Gibelli, L.R.; Pigoli, C.; Susini, F.; Podavini, E.; Scali, F.; Varisco, G.; Alborali, G.L. The characterisation of Lactococcus garvieae isolated in an outbreak of septicaemic disease in farmed sea bass (Dicentrarchus labrax, Linnaues 1758) in Italy. Pathogens 2024, 13. [Google Scholar] [CrossRef]
- Neupane, S.; Rao, S.; Yan, W.-X.; Wang, P.-C.; Chen, S.-C. First identification, molecular characterization, and pathogenicity assessment of Lactococcus garvieae isolated from cultured pompano in Taiwan. J. Fish Dis. 2023, 46, 1295–1309. [Google Scholar] [CrossRef]
- Shahin, K.; Veek, T.; Heckman, T.I.; Littman, E.; Mukkatira, K.; Adkison, M.; Welch, T.J.; Imai, D.M.; Pastenkos, G.; Camus, A.; et al. Isolation and characterization of Lactococcus garvieae from rainbow trout, Onchorhyncus mykiss, from California, USA. Transbound. Emerg. Dis. 2022, 69, 2326–2343. [Google Scholar] [CrossRef]
- Ortega, C.; Irgang, R.; Valladares-Carranza, B.; Collarte, C.; Avendaño-Herrera, R. First identification and characterization of Lactococcus garvieae isolated from rainbow trout (Oncorhynchus mykiss) cultured in Mexico. Animals 2020, 10. [Google Scholar] [CrossRef]
- Egger, R.C.; Rosa, J.C.C.; Resende, L.F.L.; de Pádua, S.B.; de Oliveira Barbosa, F.; Zerbini, M.T.; Tavares, G.C.; Figueiredo, H.C.P. Emerging fish pathogens Lactococcus petauri and L. garvieae in Nile tilapia (Oreochromis niloticus) farmed in Brazil. Aquaculture 2023, 565, 739093. [Google Scholar] [CrossRef]
- Gazal, L.E. de S.; Brito, K.C.T. de; Kobayashi, R.K.T.; Nakazato, G.; Cavalli, L.S.; Otutumi, L.K.; Brito, B.G. de Antimicrobials and resistant bacteria in global fish farming and the possible risk for public health. Arq. Inst. Biol. (Sao. Paulo). 2020, 87. [Google Scholar]
- Duman, M.; Buyukekiz, A.G.; Saticioglu, I.B.; Cengiz, M.; Sahinturk, P.; Altun, S. Epidemiology, genotypic diversity, and antimicrobial resistance of Lactococcus garvieae in farmed rainbow trout (Oncorhynchus mykiss). IFRO 2020, 19, 1–18. [Google Scholar]
- Raissy, M.; Moumeni, M. Detection of antibiotic resistance genes in some Lactococcus garvieae strains isolated from infected rainbow trout. Iran. J. Fish. Sci. 2016, 15, 221–229. [Google Scholar]
- Monteiro, S.H.; Francisco, J.G.; Andrade, G.C.R.M.; Botelho, R.G.; Figueiredo, L.A.; Tornisielo, V.L. Study of spatial and temporal distribution of antimicrobial in water and sediments from caging fish farms by on-line SPE-LC-MS/MS. J. Environ. Sci. Heal. Part B 2016, 51, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Leal, C.A.G. Resistência a antimicrobianos na piscicultura. Panor. da Aquicultura 2022, 31, 38–47. [Google Scholar]
- Öztürk, R.Ç.; Ustaoglu, D.; Ture, M.; Bondavalli, F.; Colussi, S.; Pastorino, P.; Vela, A.I.; Kotzamanidis, C.; Fernandez-Garayzábal, J.F.; Bitchava, K.; et al. Epidemiological cutoff values and genetic antimicrobial resistance of Lactococcus garvieae and L. petauri. Aquaculture 2024, 593, 741340. [Google Scholar] [CrossRef]
- Carvalho, G.M.; Silva, B.A.; Xavier, R.G.C.; Zanon, I.P.; Vilela, E.G.; Nicolino, R.R.; Tavares, G.C.; Silva, R.O.S. Evaluation of disk diffusion method for testing the rifampicin, erythromycin, and tetracycline susceptibility of Clostridioides (Prev. Clostridium) difficile. Anaerobe 2023, 80, 102720. [Google Scholar] [CrossRef]
- Sebastião, F.A.; Furlan, L.R.; Hashimoto, D.T.; Pilarski, F. Identification of bacterial fish pathogens in Brazil by direct colony PCR and 16S rRNA gene sequencing. Adv. Microbiol. 2015, 05, 409–424. [Google Scholar] [CrossRef]
- Fukushima, H.C.S.; Leal, C.A.G.; Cavalcante, R.B.; Figueiredo, H.C.P.; Arijo, S.; Moriñigo, M.A.; Ishikawa, M.; Borra, R.C.; Ranzani-Paiva, M.J.T. Lactococcus garvieae outbreaks in Brazilian farms: Lactococcosis in Pseudoplatystoma sp. – Development of an autogenous vaccine as a control strategy. J. Fish Dis. 2017, 40, 263–272. [Google Scholar] [CrossRef]
- Assis, G.B.N.; Pereira, F.L.; Zegarra, A.U.; Tavares, G.C.; Leal, C.A.; Figueiredo, H.C.P. Use of MALDI-TOF mass spectrometry for the fast identification of Gram-positive fish pathogens. Front. Microbiol. 2017, 8, 1492. [Google Scholar] [CrossRef]
- Tavares, G.C.; de Queiroz, G.A.; Assis, G.B.N.; Leibowitz, M.P.; Teixeira, J.P.; Figueiredo, H.C.P.; Leal, C.A.G. Disease outbreaks in farmed Amazon catfish (Leiarius marmoratus x Pseudoplatystoma corruscans) caused by Streptococcus agalactiae, S. iniae, and S. dysgalactiae. Aquaculture 2018, 495, 384–392. [Google Scholar] [CrossRef]
- Rosário, A.E.C. do; Henrique, M.C.; Costa, É.J.C. da; Barbanti, A.C.C.; Tavares, G.C. Surtos de bacteriose em juvenis de pirarucu (Arapaima gigas) provevientes de pisciculturas amazônicas e avaliação da sensibilidade de Aeromonas jandaei a antimicrobianos. In Enfermidades parasitárias e bacterianas na piscicultura brasileira: insights e perspectivas; Cruz, M.G. da, Castro, J. da S., Jerônimo, G.T., Eds.; i-EDUCAM, 2023; pp. 33–46. [Google Scholar]
- Ferrario, C.; Ricci, G.; Milani, C.; Lugli, G.A.; Ventura, M.; Eraclio, G.; Borgo, F.; Fortina, M.G. Lactococcus garvieae: where is it from? A first approach to explore the evolutionary history of this emerging pathogen. PLoS One 2013, 8, e84796. [Google Scholar] [CrossRef] [PubMed]
- CLSI VET03: Methods for Antimicrobial Broth Dilution and Disk Diffusion Susceptibility Testing of Bacteria Isolated from Aquatic Animals; Clinical and Laboratory Standards Institute: Wayne, USA, 2020.
- Smith, P.; Finnegan, W.; Ngo, T.; Kronvall, G. Influence of incubation temperature and time on the precision of MIC and disc diffusion antimicrobial susceptibility test data. Aquaculture 2018, 490, 19–24. [Google Scholar] [CrossRef]
- Kronvall, G.; Kahlmeter, G.; Myhre, E.; Galas, M.F. A new method for normalized interpretation of antimicrobial resistance from disk test results for comparative purposes. Clin. Microbiol. Infect. 2003, 9, 120–132. [Google Scholar] [CrossRef]
- Silley, P. Susceptibility testing methods, resistance and breakpoints: what do these terms really mean? Rev. Sci. Tech. - Off. Int. des Épizooties 2012, 31, 33–41. [Google Scholar] [CrossRef]
- Smith, P. Eight rules for improving the quality of papers on the antimicrobial susceptibility of bacteria isolated from aquatic animals. Dis. Aquat. Organ. 2020, 139, 87–92. [Google Scholar] [CrossRef]
- R Core Team R: A language and environment for statistical computing 2024.
- Mauri, M.; Elli, T.; Caviglia, G.; Uboldi, G.; Azzi, M. RAWGraphs: A visualisation platform to create open outputs. In Proceedings of the Proceedings of the 12th Biannual Conference on Italian SIGCHI Chapter; Association for Computing Machinery: New York, NY, USA, 2017. [Google Scholar]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Assessing the antimicrobial susceptibility of bacteria obtained from animals. Vet. Microbiol. 2010, 141, 1–4. [Google Scholar] [CrossRef]
- Krumperman, P.H. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef]
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- McEwen, S.A.; Collignon, P.J. Antimicrobial resistance: a One Health perspective. Microbiol. Spectr. 2018, 6, 10–1128. [Google Scholar] [CrossRef]
- Torres-Corral, Y.; Santos, Y. Predicting antimicrobial resistance of Lactococcus garvieae: PCR detection of resistance genes versus MALDI-TOF protein profiling. Aquaculture 2022, 553, 738098. [Google Scholar] [CrossRef]
- Altan, E.; Korun, J. Lactococcus garvieae isolates from rainbow trout (Oncorhynchus mykiss, W.) compared by PLG and SA1B10 PCR primer pairs. J. Ilm. Platax 2021, 9, 18–28. [Google Scholar] [CrossRef]
- Kawanishi, M.; Kojima, A.; Ishihara, K.; Esaki, H.; Kijima, M.; Takahashi, T.; Suzuki, S.; Tamura, Y. Drug resistance and pulsed-field gel electrophoresis patterns of Lactococcus garvieae isolates from cultured Seriola (Yellowtail, Amberjack and Kingfish) in Japan. Lett. Appl. Microbiol. 2005, 40, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Lim, F.H.; Jenkins, D.R. Native valve endocarditis caused by Lactococcus garvieae: an emerging human pathogen. BMJ Case Rep. 2017, 2017, bcr–2017. [Google Scholar] [CrossRef] [PubMed]
- Korun, J.; Altan, E.; Teker, S.; Ulutaş, A. A study on the antimicrobial resistance of Lactococcus garvieae. Acta Aquat. 2021, 8, 98–102. [Google Scholar] [CrossRef]
- Sezgin, S.S.; Yılmaz, M.; Arslan, T.; Kubilay, A. Current antibiotic sensitivity of Lactococcus garvieae in rainbow trout (Oncorhynchus mykiss) farms from Southwestern Turkey. J. Agric. Sci. 2023, 29, 630–642. [Google Scholar] [CrossRef]
- Chang, P.H.; Lin, C.W.; Lee, Y.C. Lactococcus garvieae infection of cultured rainbow trout, Oncorhynchus mykiss, in Taiwan and associated biophysical characteristics and histopathology. Bull. Eur. Assoc. Fish Pathol. 2002, 22, 319–327. [Google Scholar]
- Majeed, S.; De Silva, L.A.D.S.; Kumarage, P.M.; Heo, G.J. Characterization of pathogenic Lactococcus garvieae isolated from farmed mullet (Mugil cephalus). Vet. Integr. Sci. 2025, 23, 1–17. [Google Scholar]
- Ture, M.; Boran, H. Phenotypic and genotypic antimicrobial resistance of Lactococcus sp. strains isolated from rainbow trout (Oncorhynchus mykiss). Bull. Vet. Inst. Pulawy 2015, 59, 37–42. [Google Scholar] [CrossRef]
- Vela, A.I.; del Mar Blanco, M.; Colussi, S.; Kotzamanidis, C.; Prearo, M.; Altinok, I.; Acutis, P.L.; Volpatti, D.; Alba, P.; Feltrin, F.; et al. The association of Lactococcus petauri with lactococcosis is older than expected. Aquaculture 2024, 578, 740057. [Google Scholar] [CrossRef]
- Chan, Y.-X.; Cao, H.; Jiang, S.; Li, X.; Fung, K.-K.; Lee, C.-H.; Sridhar, S.; Chen, J.H.-K.; Ho, P.-L. Genomic investigation of Lactococcus formosensis, Lactococcus garvieae, and Lactococcus petauri reveals differences in species distribution by human and animal sources. Microbiol. Spectr. 2024, 12, e00541–24. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Han, J.; Barkema, H.W.; Wang, Y.; Gao, J.; Kastelic, J.P.; Han, B.; Qin, S.; Deng, Z. Comparative genomic analyses of Lactococcus garvieae isolated from bovine mastitis in China. Microbiol. Spectr. 2023, 11, e02995–22. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, T.F.; Leibowitz, M.P.; Leal, C.A.G. Local epidemiological cutoff values and antimicrobial susceptibility profile for Brazilian Francisella orientalis isolates. Aquaculture 2022, 553, 738054. [Google Scholar] [CrossRef]
- Sun, K.; Wang, H.; Zhang, M.; Xiao, Z.; Sun, L. Genetic mechanisms of multi-antimicrobial resistance in a pathogenic Edwardsiella tarda strain. Aquaculture 2009, 289, 134–139. [Google Scholar] [CrossRef]
- Bondad-Reantaso, M.G.; MacKinnon, B.; Karunasagar, I.; Fridman, S.; Alday-Sanz, V.; Brun, E.; Le Groumellec, M.; Li, A.; Surachetpong, W.; Karunasagar, I.; et al. Review of alternatives to antibiotic use in aquaculture. Rev. Aquac. 2023, 15, 1421–1451. [Google Scholar] [CrossRef]
- Leal, C.A.G.; Silva, B.A.; Colombo, S.A. Susceptibility profile and epidemiological cut-off values are influenced by serotype in fish pathogenic Streptococcus agalactiae. Antibiotics 2023, 12. [Google Scholar] [CrossRef]
- Savvidis, G.K.; Anatoliotis, C.; Kanaki, Z.; Vafeas, G. Epizootic outbreaks of lactococcosis disease in rainbow trout, Oncorhynchus mykiss (Walbaum), culture in Greece. Bull. Eur. Assoc. Fish Pathol. 2007, 27, 223–228. [Google Scholar]
Figure 1.
Disk diffusion scatter plots for antimicrobials versus diameters of inhibition zones for the six Lactococcus formosensis, 17 Lactococcus garvieae and 24 Lactococcus petauri strains evaluated.
Figure 1.
Disk diffusion scatter plots for antimicrobials versus diameters of inhibition zones for the six Lactococcus formosensis, 17 Lactococcus garvieae and 24 Lactococcus petauri strains evaluated.
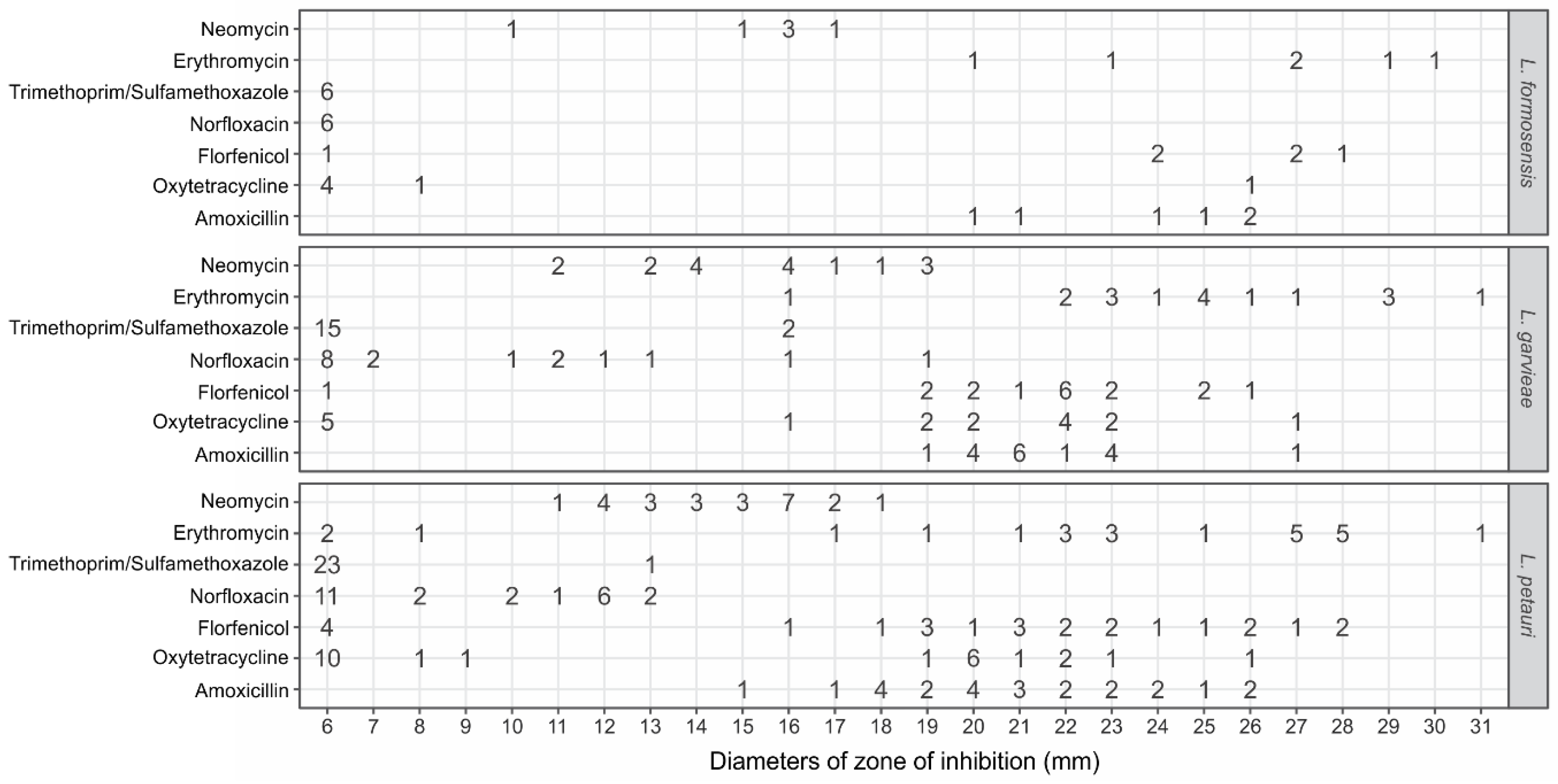
Figure 2.
Alluvial plot demonstrating the association of the host with antimicrobial susceptibility or resistance to florfenicol (FLO) and oxytetracycline (OXY) in Lactococcus formosensis (a), Lactococcus garvieae (b) and Lactococcus petauri (c) strains.
Figure 2.
Alluvial plot demonstrating the association of the host with antimicrobial susceptibility or resistance to florfenicol (FLO) and oxytetracycline (OXY) in Lactococcus formosensis (a), Lactococcus garvieae (b) and Lactococcus petauri (c) strains.
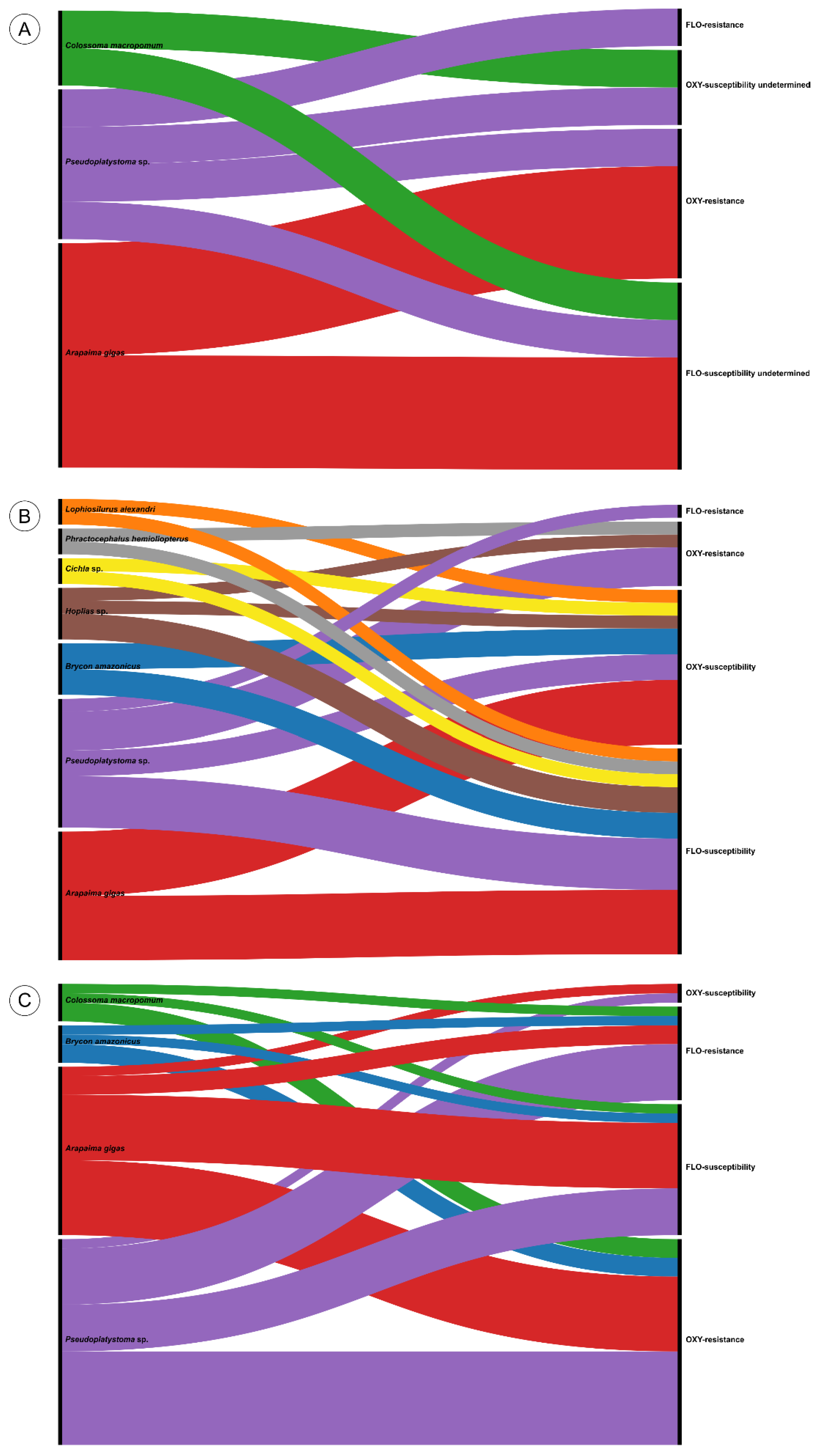
Figure 3.
Multiple antibiotic resistance (MAR) index box plot of Lactococcus spp. strains isolated from native Brazilian fish species.
Figure 3.
Multiple antibiotic resistance (MAR) index box plot of Lactococcus spp. strains isolated from native Brazilian fish species.
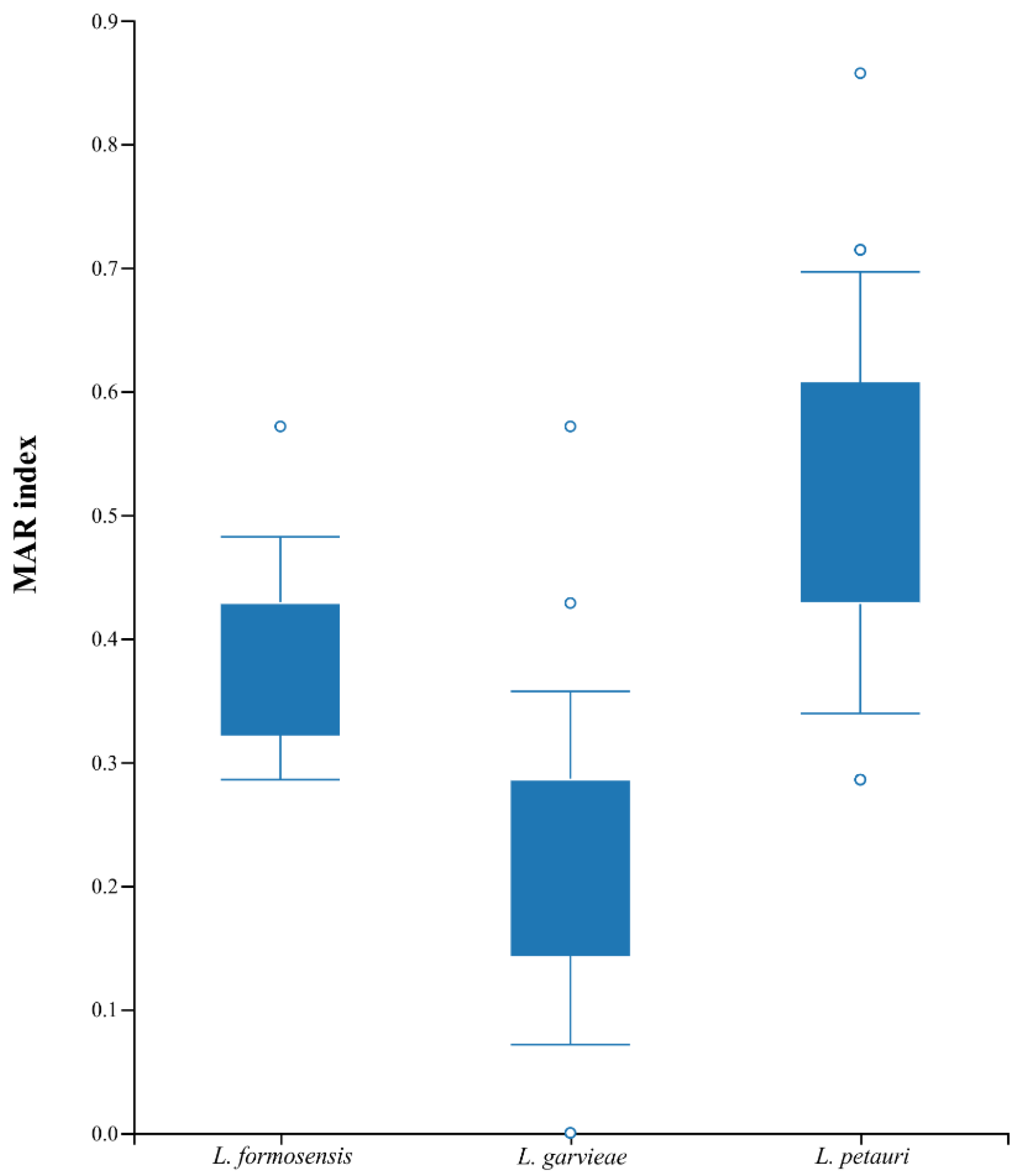

Table 1.
Metadata of the 47 strains of lactococcosis-causing bacteria isolated from the native Brazilian fish species.
Table 1.
Metadata of the 47 strains of lactococcosis-causing bacteria isolated from the native Brazilian fish species.
| Isolate | Species | Host | Origin | Tissue | Year | State | Reference |
|---|---|---|---|---|---|---|---|
| 167/23-02 | L. formosensis | Arapaima gigas | Farmed | Brain | 2023 | BA | [2] |
| 167/23-06 | L. formosensis | Arapaima gigas | Farmed | Brain | 2023 | BA | [2] |
| 167/23-09 | L. formosensis | Arapaima gigas | Farmed | Kidney | 2023 | BA | This study |
| AM-LG05 | L. formosensis | Colossoma macropomum | Farmed | Intestine | 2022 | AM | [2] |
| 52MS | L. formosensis | Pseudoplatystoma fasciatum | Farmed | Brain | 2012 | MS | [18] |
| LG91-23 | L. formosensis | Pseudoplatystoma sp. | Farmed | Brain | 2023 | MG | [2] |
| CRBP53 | L. garvieae | Arapaima gigas | Farmed | Intestine | 2023 | AM | [2] |
| CRBP54 | L. garvieae | Arapaima gigas | Farmed | Intestine | 2023 | AM | [2] |
| CRBP138 | L. garvieae | Arapaima gigas | Farmed | Intestine | 2023 | AM | [2] |
| CRBP144 | L. garvieae | Arapaima gigas | Farmed | Intestine | 2023 | AM | [2] |
| PA-LG01 | L. garvieae | Arapaima gigas | Farmed | Brain | 2018 | PA | [22] |
| LG88-23 | L. garvieae | Brycon amazonicus | Farmed | Brain | 2023 | MG | [2] |
| LG89-23 | L. garvieae | Brycon amazonicus | Farmed | Kidney | 2023 | MG | [2] |
| LG116-23 | L. garvieae | Cichla sp. | Wild | Brain | 2023 | MG | This study |
| LG63-21 | L. garvieae | Hoplias macrophtalmus | Farmed | Kidney | 2021 | MG | [2] |
| LG114-23 | L. garvieae | Hoplias malabaricus | Wild | Brain | 2023 | AM | This study |
| LG10-14 | L. garvieae | Lophiosilurus alexandri | Farmed | Brain | 2014 | MG | [20] |
| LG66-22 | L. garvieae | Phractocephalus hemioliopterus | Farmed | Kidney | 2022 | MG | [2] |
| LG09-14 | L. garvieae | Pseudoplatystoma corruscans | Farmed | Kidney | 2014 | SP | [20] |
| LG23-16 | L. garvieae | Pseudoplatystoma corruscans | Farmed | Brain | 2016 | SP | [21] |
| 177 | L. garvieae | Pseudoplatystoma fasciatum | Farmed | Brain | 2012 | MS | [19] |
| 31MS | L. garvieae | Pseudoplatystoma fasciatum | Farmed | Kidney | 2012 | MS | [18] |
| LG119-24 | L. garvieae | Pseudoplatystoma sp. | Farmed | Brain | 2024 | MG | This study |
| 167/23-03 | L. petauri | Arapaima gigas | Farmed | Kidney | 2023 | BA | This study |
| 167/23-04 | L. petauri | Arapaima gigas | Farmed | Kidney | 2023 | BA | This study |
| 167/23-05 | L. petauri | Arapaima gigas | Farmed | Kidney | 2023 | BA | This study |
| 167/23-07 | L. petauri | Arapaima gigas | Farmed | Kidney | 2023 | BA | This study |
| 167/23-08 | L. petauri | Arapaima gigas | Farmed | Kidney | 2023 | BA | This study |
| 167/23-10 | L. petauri | Arapaima gigas | Farmed | Spleen | 2023 | BA | This study |
| CRBT89 | L. petauri | Arapaima gigas | Farmed | Intestine | 2023 | AM | [2] |
| CRBT98 | L. petauri | Arapaima gigas | Farmed | Intestine | 2023 | AM | [2] |
| CRBP146 | L. petauri | Arapaima gigas | Farmed | Intestine | 2023 | AM | [2] |
| AM-LG07 | L. petauri | Brycon amazonicus | Farmed | Brain | 2022 | AM | [2] |
| AM-LG08 | L. petauri | Brycon amazonicus | Farmed | Brain | 2022 | AM | [2] |
| AM-LG02 | L. petauri | Colossoma macropomum | Farmed | Intestine | 2020 | AM | [2] |
| AM-LG03 | L. petauri | Colossoma macropomum | Farmed | Intestine | 2022 | AM | [2] |
| LG03-18 | L. petauri | Pseudoplatystoma corruscans | Farmed | Brain | 2018 | MG | [2] |
| 14MS | L. petauri | Pseudoplatystoma fasciatum | Farmed | Kidney | 2012 | MS | [18] |
| 176 | L. petauri | Pseudoplatystoma fasciatum | Farmed | Brain | 2012 | MS | [19] |
| 86 | L. petauri | Pseudoplatystoma sp. | Farmed | Brain | 2012 | MS | [19] |
| 89/2 | L. petauri | Pseudoplatystoma sp. | Farmed | Brain | 2012 | MS | [19] |
| 93 | L. petauri | Pseudoplatystoma sp. | Farmed | Brain | 2012 | MS | [19] |
| LG86-23 | L. petauri | Pseudoplatystoma sp. | Farmed | Kidney | 2023 | MG | [2] |
| LG94-23 | L. petauri | Pseudoplatystoma sp. | Farmed | Brain | 2023 | MG | [2] |
| LG104-23 | L. petauri | Pseudoplatystoma sp. | Farmed | Brain | 2023 | MG | [2] |
| LG106-23 | L. petauri | Pseudoplatystoma sp. | Farmed | Kidney | 2023 | MG | [2] |
| LG117-23 | L. petauri | Pseudoplatystoma sp. | Farmed | Kidney | 2023 | MG | This study |
AM: Amazonas; BA: Bahia; MS: Mato Grosso do Sul; MG: Minas Gerais; PA: Pará; SP: São Paulo.
Table 2.
Minimum and maximum values, mean and standard deviation of the inhibition zone diameters, epidemiological cut-off values, and wild type/non-wild type (WT/NWT) percentual for Lactococcus spp. and quality control strains in the antimicrobial susceptibility analysis.
Table 2.
Minimum and maximum values, mean and standard deviation of the inhibition zone diameters, epidemiological cut-off values, and wild type/non-wild type (WT/NWT) percentual for Lactococcus spp. and quality control strains in the antimicrobial susceptibility analysis.
| Antimicrobials | Minimum Value | Maximum Value | Mean ± SD | ECV (mm) | WT (%) | NWT* (%) |
|---|---|---|---|---|---|---|
| Lactococcus formosensisa | ||||||
| Amoxicillin | 19 | 27 | 23.2 ± 2.7 | - | - | - |
| Erythromycin | 20 | 30 | 25.7 ± 3.7 | - | - | - |
| Florfenicol | 6 | 28 | 22.3 ± 7.8 | - | - | 16.7 |
| Neomycin | 10 | 17 | 14.9 ± 2.4 | - | - | - |
| Norfloxacin | 6 | 6 | 6.0 ± 0.0 | - | - | 100 |
| Oxytetracycline | 6 | 27 | 9.5 ± 7.5 | - | - | 66.7 |
| Trimethoprim-sulfametoxazole | 6 | 6 | 6.0 ± 0.0 | - | - | 100 |
| Lactococcus garvieaeb | ||||||
| Amoxicillin | 18 | 28 | 21.4 ± 2.2 | ≥ 11 | 100 | 0 |
| Erythromycin | 16 | 31 | 24.7 ± 3.7 | ≥ 16 | 100 | 0 |
| Florfenicol | 6 | 29 | 20.9 ± 4.4 | ≥ 12 | 94.4 | 5.6 |
| Neomycin | 10 | 19 | 15.1 ± 2.7 | ≥ 7 | 100 | 0 |
| Norfloxacin | 6 | 19 | 9.0 ± 4.0 | - | - | 47 |
| Oxytetracycline | 6 | 27 | 16.5 ± 7.3 | ≥ 10 | 72.2 | 27.8 |
| Trimethoprim-sulfametoxazole | 6 | 19 | 7.2 ± 3.4 | - | - | 88.2 |
| Lactococcus petauric | ||||||
| Amoxicillin | 15 | 26 | 20.5 ± 3.0 | ≥ 16 | 95.8 | 4.2 |
| Erythromycin | 6 | 31 | 22.4 ± 7.1 | ≥ 23 | 66.7 | 33.3 |
| Florfenicol | 6 | 29 | 19.5 ± 6.9 | ≥ 21 | 62.5 | 37.5 |
| Neomycin | 10 | 19 | 14.2 ± 2.2 | ≥ 9 | 100 | 0 |
| Norfloxacin | 6 | 14 | 8.6 ± 2.9 | ≥ 13 | 16.7 | 83.3 |
| Oxytetracycline | 6 | 26 | 13.6 ± 7.5 | ≥ 23 | 16.7 | 83.3 |
| Trimethoprim-sulfametoxazole | 6 | 14 | 6.3 ± 1.4 | - | - | 95.8 |
| Escherichia coliATCC 25922d | ||||||
| Amoxicillin | 14 | 19 | 15.8 ± 2.4 | - | - | - |
| Erythromycin | 12 | 18 | 14.6 ± 2.8 | - | - | - |
| Florfenicol | 19 | 28 | 23.5 ± 4.4 | - | - | - |
| Neomycin | 16 | 20 | 18.0 ± 2.0 | - | - | - |
| Norfloxacin | 24 | 34 | 30.6 ± 5.7 | - | - | - |
| Oxytetracycline | 19 | 27 | 23.2 ± 3.3 | - | - | - |
| Trimethoprim-sulfametoxazole | 25 | 26 | 25.5 ± 0.7 | - | - | - |
| Aeromonas salmonicida subsp. salmonicidaATCC 33658d | ||||||
| Amoxicillin | 24 | 30 | 27.4 ± 3.1 | - | - | - |
| Erythromycin | 19 | 22 | 20.7 ± 1.5 | - | - | - |
| Florfenicol | 32 | 36 | 34.2 ± 1.7 | - | - | - |
| Neomycin | 12 | 20 | 17.3 ± 4.6 | - | - | - |
| Norfloxacin | 21 | 37 | 29.6 ± 8.0 | - | - | - |
| Oxytetracycline | 29 | 32 | 29.7 ± 1.5 | - | - | - |
| Trimethoprim-sulfametoxazole | 24 | 26 | 25.0 ± 1.4 | - | - | - |
a ECV undetermined b pECV determined in this study c pECV determined in Egger et al. [10] d Quality control strains * Regardless of the bacterial species, bacteria that did not present an inhibition zone and did not have a defined epidemiological cutoff value (ECV) were considered NWT for the antimicrobials.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



