
Submitted:
29 October 2024
Posted:
30 October 2024
You are already at the latest version
Abstract
Cardiovascular diseases (CVDs), which comprise coronary heart disease, hypertension and stroke, collectively represent the number one cause of death globally. Atherosclerosis is the dominant cause of CVDs; and its risk factors are elevated levels of low-density lipoprotein-cholesterol and triglycerides, hypertension, cigarette smoking, obesity and diabetes mellitus. In addition, diverse evidence highlighted the role played by inflammation and clonal haematopoiesis, eventually leading to the immunity involvement. The human microbiota project and subsequent studies using next-generation sequencing technology have highlighted that thousands of different microbial species are present in the human gut. The disturbances in the gut microbiota (GM) composition, i.e., gut dysbiosis, have been associated with diseases ranging from localized gastrointestinal disorders to metabolic and cardiovascular illnesses. Of note, experimental studies suggested that GM, host immune cells and marine-derived ingredients work together to ensure the intestinal wall integrity. This review discusses current evidence concerning the links among GM, marine-derived ingredients and human inflammatory disease. In detail, we summarize the impact of fish-derived proteins and algae components, on CVDs risk factors and gut microbiome. Furthermore, we describe the interplay among these dietary components, probiotics/prebiotics and CVDs.
Keywords:
Cardiovascular diseases
; atherosclerosis risk factors
; microbiota
; fish protein hydrolysates
; seaweeds
; probiotics
1. Introduction
1.1. Cardiovascular Diseases Risk Factors
Cardiovascular diseases (CVDs), which comprise coronary heart disease, hypertension and stroke, collectively represent the number one cause of death globally [1]. Atherosclerosis, the dominant CVDs cause, is the process leading to the accumulation of fatty and/or fibrous material in the innermost layer of arteries, aka the intima. Subsequently, atherosclerotic plaque can acquire more fibrous material and gather calcium mineral. Advanced atherosclerotic plaques can produce a flow-limiting obstruction or disrupt and promote the thrombus development, both these phenomena lead to tissue ischaemia and, eventually, to the clinical manifestations of atherosclerosis, i.e., myocardial infarction, strokes and peripheral artery disease [2]. Risk factors for atherosclerotic lesion’s development and its thrombotic complications are elevated levels of low-density lipoprotein-cholesterol (LDL-C) and triglycerides (TG), hypertension, cigarette smoking, obesity and diabetes mellitus. Diverse evidence also highlighted the role played by inflammation and clonal haematopoiesis, eventually leading to the immunity involvement [3,4].
Of note, a recent study demonstrated that in patients already taking statin therapy, residual inflammatory risk appears to be strongly associated with future CV events than residual cholesterol risk [5,6]. These data strengthen the hypothesis that concomitant targeting of inflammation and atherogenic lipids, will further diminish CVDs’ risk. However, conventional therapeutic approaches, often exhibit limitations, i.e., side effects [7] and inadequate disease control [8,9,10]. Additionally, it is well known that all the above-mentioned CVD’s risk factors are susceptible to lifestyle modifications, such as diet and physical exercise [11]. In line with this evidence, in recent years diverse natural products and their derivates have garnered increasing attention as care standard, along with pharmacotherapy, for the CVDs treatment [12].
1.2. Microbiota-Immune Axis and CVDs: State of the Art
10–100 trillion symbiotic microbial cells reside in the human body and are known as the human microbiota. The totality of these cells, belonging to bacteria, fungi and parasites together with genetic material (e.g. from viruses) make up the human microbiome. Our genetic ancestry is made by a combination of human and microbial species; thus, our metabolism is determined by microbial and human signatures [13].
Recent research has shown that the gut microbiota (GM) acts as an endocrine organ, playing a role in modulating immunity and influencing the development of inflammatory, metabolic and infectious diseases [14]. Dietary fibre, which is the indigestible components of fruits, vegetables, and grains, is recognized as a significant energy source for bacteria that produce short chain fatty acids (SCFAs) [15]. When there is a balanced composition of human microbiome, it leads to a healthy intestinal epithelial barrier and the recruitment and activation of the appropriate immune cells through the secretion of metabolites, particularly SCFAs, and the expression of microbial components [16]. The Western diet, rich in simple carbohydrates and saturated fats, along with reduced physical activity, has contributed to gut and skin dysbiosis, altering immune balance. A shift from living in close contact with natural environments, combined with changes in eating habits, such as an increase in ultra-processed foods, red meats and a reduction in fibre-rich foods, has led to a GM depletion [17,18]. Eventually these conditions can lead to metabolic problems and variations in symbiotic microorganisms, rushing the CVDs development [19]. An increasing body of research indicates that intestinal bacteria and their metabolites are crucial in the development of CVDs. Some studies suggest that hypertension patients often show a lower diversified gut microbiota, along with an increased prevalence of certain bacteria like Clostridiales and Bacteroidales [20,21]. These microbiota changes, observed in both humans and animal models, may play a role in hypertension's development by affecting blood pressure regulation through microbial by-products such as SCFAs [20,21]. Systolic blood pressure instead is correlated with Robinsoniella and Catabacter abundance in a study on Coronary Artery Risk Development in Young Adults (CARDIA) [22].
In addition, cigarette smoke may affect the GM composition and function by upregulating oxidative stress-related enzymes, modifying the gut mucin layer and the expression of intestinal tight junction proteins, as well as by stimulating the spread of non-commensal bacteria [23]. The smokers GM differs from that of non-smokers, with a higher relative abundance of Actinobacteria and Cyanobacteria [24].
Different investigations indicate a correlation between GM and CVDs, with trimethylamine-N-oxide (TMAO) identified as a crucial metabolite [25]. Indeed, TMAO is synthesized from dietary choline and carnitine [26], by gut bacterial TMA-lyase, which produces TMA that is subsequently adsorbed by intestinal cells and transported to the liver. There it is oxidized by the enzyme flavin-containing monooxygenase 3 (FMO3). A meta-analysis of prospective studies revealed that individuals with elevated plasma TMAO levels had a 23% higher risk of CV events [27] and a 62% greater risk of all-cause mortality [28]. Altogether, these data suggest that TMAO has been linked to an elevated CVDs risk [29].
Conversely, the gut microbiota produces metabolites like SCFAs and in detail the butyrate that shows beneficial effects on the host, including promoting growth and reducing inflammation in intestinal epithelial cells. Additionally, butyrate plays a key role in maintaining immune balance in the gut by facilitating communication between the host and microbiota [16].
Nutritional interventions targeting the gut microbiota, such as probiotics, prebiotics, and postbiotics, have shown promise in preventing CVDs, especially when implemented early in life [30].
1.3. Marine-Derived Compounds and CVDs
Marine species, aka mammals, fish, seaweeds, sea anemones, sponges, represent approximately one-half of the global biodiversity. Therefore, the sea offers a wonderful resource for novel compounds, potentially able to improve the health of the worldwide population. Attention has been drawn to the beneficial effects of fish consumption, due to the ability of fish ingredients, mainly omega-3 polyunsaturated fatty acids (n-3 PUFAs) and proteins, to lower CVDs risk factors [31,32,33] and modulate inflammation [34,35,36,37].
Based on these data, the US Food and Drug Administration (FDA) have formally declared that consumption of up to 3 g/d of marine-derived n-3 PUFAs is generally considered as safe. In line with this health claim, fish consumption is still recommended in the 2020-2025 Dietary Guidelines for Americans and by the American Heart Association [38]. Additionally, a recent positional paper strengthened the positive relationship between the replacement of proteins from red meat with proteins from fish and reduced risk of CVDs [39]. In agreement, two meta-analyses reported a significant inverse association between fish consumption and all-cause of mortality, with a nadir at consumption of 60-80 g/d and that this inverse association is marked influenced by regional differences, respectively [40,41]. More in detail, Jayedi et al, found that in Western studies the risk of all-cause and CVD mortality decreased in a dose-dependent manner and then increased with a relatively sharp trend, suggesting a U-shape curve association. While, in Asian studies this dose-response relationship was linear [41]. Recently, Zhou et al in a general Chinese population found a reverse J-shaped association between fish-derived protein and new-onset hypertension [42]. Meaning that, there is a window of consumption (appropriate level) where the risk of new-onset hypertension is lower. These data confirmed previously results that showed a U-shaped curve association between protein intake and health [43]. Additionally, recent data have documented seaweeds as promising reservoirs of bioactive compounds able at targeting multiple aspects of CVDs, including inflammation [44,45]. Specifically, seaweeds contain polysaccharides, proteins, pigments, lipids, sterols, terpenes and phenolic compounds [46].
This review summarizes the impact of marine-derived ingredients, aka fish-derived proteins and algae components, on CVDs risk factors and gut microbiome. Furthermore, we describe the interplay among these dietary components, probiotics/prebiotics and CVDs.
2. Fish-Derived Proteins and CVDs Risk Factors
Fish-derived proteins contain all the essential amino acids, mainly lysine and leucine, some non-essential amino acids (aspartic acid, glutamic acid, and alanine), together with the amino acid-derived organic acid taurine [47,48,49]. Different studies have proved that the enzymatic digestion of fish by-products is an efficient means of producing peptides with enhanced bioactivity [48]. Furthermore, experimental and clinical data have investigated the impact of fish-derived proteins/bioactive peptides on lipid profile, glucose metabolism, inflammation, and blood pressure.
2.1. Experimental Studies
A recent systematic review and meta-analysis described all the pre-clinical data performed in rodents and published before July 15, 2022 [33]. This paper concluded that intake of protein from fish muscles or fish by-products significantly decrease circulating total cholesterol (TC) concentration when compared to their control group. Of note, the authors highlighted that the stronger effect of fish-derived proteins’ intake was observed in the subgroup comprising genetically modified rodent models, which spontaneously develop hypertension after birth, and rodents fed diets enriched with cholesterol alone or in combination with cholate (added to exacerbate hypercholesterolemia [50,51,52]. This data, indicating that the potency for preventing an increase of TC was higher than that for lowering TC plasma levels, may have relevant clinical application, albeit not directly transferable to human. Furthermore, the authors analysed diverse mechanisms of action to justify the hypocholesterolemic effects exerted by the dietary intake of fish or fish proteins (Figure 1).
They found that in almost half of the analysed studies a lower TC concentration was associated with higher faecal excretion of cholesterol and/or bile acids. Furthermore, in two papers [53,54] the above-described effects were also combined with higher mRNA expression levels of cholesterol 7-alpha-hydroxylase (CYP7A1) [55], which is the first and rate-liming enzyme in the cholesterol metabolism, such as the bile acid synthesis. On the contrary, the impact of fish or fish-proteins on the expression of 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase (HMG-CoA reductase), LDL-receptor (LDL-R), and acyl-CoA:cholesterol acyltransferase (ACAT2) was difficult to assess, because it was marked influenced by the rodent model used. However, an elegant and recent work demonstrated that the hypocholesterolemic effect exerted by Alcalase-silver carp hydrolysate (Alcalase-SCH) was associated with an up-regulation of LDL-R expression and a down-regulation of Niemann-Pick C1-like 1 (NPC1L1) and ACAT2 [56]. In addition, these authors identified novel peptides, present in the Alcalase-SCH, as main contributors to the hypocholesterolemic activity of Alcalase-SCH [56]. Of note, in line with data obtained with soya, potatoes and rice proteins, lower methionine/glycine and lysine/arginine ratios were also observed in fish proteins compared with casein, together with a lower TC plasma level. In addition, salmon protamine is a strongly alkaline polycationic low-molecular-weight protein, in which nearly two-thirds of the amino acid composition is arginine [57,58,59]. It is well known that arginine, being a precursor for nitric oxide (NO) synthesis, may positively affect vascular function [60]. Indeed, arginine supplementation has been shown to decrease neointimal formation in animal models [61,62], and to improve flow-mediated vasodilation in humans [63].
These data were subsequently confirmed by diverse experiments. Oral administration of jellyfish collagen hydrolysate (JCH) was able to prevent the increase of serum glucose, TC and TG levels, together with the body gain weight in a mouse model of obesity, aka mice fed a high-fat diet (HFD) [64]. Additionally, JCH administration modulated oxidative stress and inflammatory response, crucial factors implicated in obesity-related pathologies, and helped recover the alteration on microbiota composition induced by high-fat diet, specifically by contrasting the lowering of Romboutsia’s abundance [64]. Similar data were published by Shi et al, in healthy mice fed a chow diet and treated with Half-fin Anchovy hydrolysate (HAHp) or with its Maillard reaction products (HAHp-MRPs) by oral gavage [65]. Significantly, the glycation process or Maillard reaction, aka the chemical process involving proteins and sugars during food processing, can enhance protein and peptide functionalities, including antioxidant and antihypertensive activity. The glycated proteins or peptides may resist digestion and undergo fermentation in the colon, potentially benefiting gut health. Studies have shown that glycated proteins, such as those from pea [66] and milk, can exhibit similar probiotic effects as Galactooligosaccharides (GOS) alone [67]. GOS are a type of prebiotic that support beneficial intestinal bacteria and produce SCFAs that have a variety of biological functions, hence promoting gut health [68]. GOS ferments quickly, producing gas and bloating. This has raised interest in prebiotics that affect the distal colon and are linked to a lower risk of colon cancer [69]. Glycated peptides' effect on the gut microbiota is yet unclear, though [70]. Jin et al. [71] investigated the effects of GOS glycated with fish peptides on GM of rats using the Maillard reaction. The composition of the gut microbiota and colonic fermentation were affected by the new glycoconjugates, offering the first in vivo proof of these prebiotic effects. Additionally, Han et al. [72] explored the chemical characteristics of glycoconjugates of myofibrillar proteins from grass carp that were conjugated with glucose via the Maillard reaction during dry heating. Glycation increased furosine levels, promoted structural changes in the proteins and reduced protein digestibility. The butyrate production during fermentation was influenced by glycation and showed positive correlation with Mitsuokella, Lachnospiraceae_UCG-004, Sutterella, Salinimicrobium, Fodinibius and Nitriliruptor, but anti-correlation with Enterococcus, Dorea, Escherichia-Shigella, and Phascolarctobacterium. These findings demonstrated that the glycation of myofibrillar proteins could have positive outcomes on gut health [72].
Lin et al, found that small-molecule peptides from the bone collagen of Harpadon nehereus (HNCP) exerted antidiabetic effects in Streptozotocin induced diabetes mice [73]. Specifically, HNCP administration significantly decreased the plasma levels of glucose, TC, TG, LDL-C and increased HDL-C concentration and insulin secretion. Moreover, HNCP improved glucose metabolism and showed remarkable antioxidant activity in this type 1 diabetic mouse model by regulating the expression levels of glycosynthesis and gluconeogenesis-related [i.e., glucokinase (GK), phosphoenolpyruvate carboxikinase1 (PEPCK1) and glucose-6-phosphate (G6Pase)] and antioxidant enzymes [i.e., catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GSH-Px) and quinone oxidoreductase 1 (NQO1)], respectively. Additionally, the same authors demonstrated that this latter effect, aka the antioxidant activity, was mediated by the activation of the nuclear factor-erythroid 2-related factor 2 (Nrf2) pathway [73]. It has been established the crucial role played by Nrf2 in redox balance, inflammation, cytotoxicity and cellular metabolism, and its involvement in many oxidative stress-based diseases [74]. Similar results were obtained in Streptozotocin-induced diabetes rats treated with the small peptide (<1kDa) fraction from Takifugu bimaculatus skin hydrolysate (TBP) [75]. Specifically, TBP was chosen because in an in vitro assay exhibited the strongest dipeptidyl peptidase-IV (DPP-IV) inhibitory activity. DPP-IV inhibition hinders the degradation of glucagon-like peptide 1 (GLP-1) and glucose-dependent insulinotropic polypeptide (GIP) that are released post-prandially, increasing their half-life and amplifying the insulin effect on glucose homeostasis [76]. In the in vivo experiment, TBS diminished weight loss, lowered fasting blood glucose concentrations, increased insulin secretion, improved irregular hormonal fluctuations and lipid metabolism, and mitigated histopathological damage in the pancreas and liver. Additionally, the relative abundance of Firmicutes decreased, alongside the increase in Bacteroidetes, significant modifications were observed at the genus level, and two metabolites, hippuric acid and ergosta-5,7,22,24(28)-tetraen-3beta-al were identified following TBP administration [75]. In line with these data, a salmon peptide fraction (SPF), containing low-molecular-weight peptides, was able to prevent the development of obesity and metabolic disorders, dampening inflammation in both hepatic and intestinal tissues, and to modulate thrombosis risk factors in high-fat-high-sugar-fed vitamin D-deficient dyslipidemic mice [77]. Interestingly, Fang et al. applied a multistage strategy, in detail, a molecular docking-based virtual screening within a small library of marine-derived natural products with follow-up in vitro and in vivo phenotypic assays, aiming at discovery new lipid lowering molecules [78].
Hypertension, as above already mentioned, is a major risk factor for CVDs. One pharmacological approach aiming at reducing blood pressure is represented by the angiotensin-I-converting enzyme (ACE) inhibitors. ACE is a key enzyme that catalyzes the conversion of angiotensin I (an inactive decapeptide) to angiotensin II (octapeptide), a potent vasoconstrictor, which stimulates the release of aldosterone, and eventually increases the blood pressure. Fish-derived bioactive peptides have been widely investigated for their anti-hypertensive effects, such as ACE inhibition. All the studies published before 2020, have been collected in reviews [32,79,80]. According to the results obtained, the bio-efficacy and bioavailability of the final peptide products are marked affected by the used extraction processes (the enzymatic hydrolysis as well as isolation/purification techniques). Additionally, size and chain length together with the presence of some amino acids (tyrosine, tryptophan, proline, and phenylalanine) at the C-terminal of the fish-derived peptide’ structures are crucial for ACE inhibition and antihypertensive effects [32,79,80]. In vitro experiments demonstrated that protein hydrolysates from fish by-products exerted competitive, non-competitive and mixed inhibition modes against ACE. In line with these data, in vivo experiments, mainly performed in spontaneously hypertensive rats (SHRs), proved the strong antihypertensive activity of protein hydrolysate from diverse marine organisms. Of note, grass carp peptides, rich in phenylalanine, leucine, aspartic acid, and glycine, significantly reduced the systolic blood pressure compared to the control group treated with captopril, the drug of choice for hypertensive patients [81]. Similar data were obtained by Chen et al, by administrating the Leu-Ser-Gly-Tyr-Gly-Pro peptide [82] from tilapia skin gelatine to SHRs [83]. Moreover, these authors via molecular docking comparison identified four connecting residues of the ACE active site, which may justify the mechanism of inhibition [83]. Recently, an experimental study demonstrated that both intact and hydrolysed blue whiting proteins reduced blood pressure in an obese rat model, inhibiting renin activity but not showing ACE inhibitory effect [84]. Whereas, a peptide composed of 13 amino acid residues, DPALATEPDPMPF, obtained from Nile tilapia (Orechromis niloticus) exhibited potent ACE inhibitory and radical scavenging activities, suggesting a potential it use in functional foods [85]. Indeed, ZBPHs administration to rats fed a high-cholesterol/cholic acid containing diet attenuated cholesterol-caused cardiac injury, testified by biochemical and histological improvement as well as significantly protecting heart genomic DNA’s oxidative damage induced by Fenton’s reagent [86]. Finally, Maneesai et al, investigated the impact of tuna protein hydrolysate (TPH) on CV remodelling and dysfunction in a rat model of metabolic syndrome (MS) [87]. The results of this study demonstrated that TPH supplementation improved all the metabolic parameters, including dyslipidemia, hyperglycemia, obesity, hypertension, cardiac hypertension, endothelial dysfunction, oxidative stress and inflammation, in the in a dose-dependent manner. These effects were related to the TPH ability at modulating angiotensin II receptor type 1 (AT1R)/NADPH oxidase 2 (NOX2), endothelial nitric oxide (eNOS), Nfr2/heme oxygenase 1 (HO-1) and peroxisome proliferator-activated receptor (PPAR)gamma/nuclear factor kappa B (NF-kB) protein expression in heart and aorta [87].
Various experimental studies have investigated the impact of fish protein hydrolysates (FPHs), from salmon or anchovy by products (spine, viscera, collagen), on atherosclerosis development. All the studies were performed on genetically modified mice, aka apoE-deficient mice, fed high-fat [88,89] or high-fat/high cholesterol diet [90,91]. Altogether, the results demonstrated that these FPHs reduced plaque area and lipid accumulation in the aorta as well as in the aortic sinus. Conversely, no differences in extracellular matrix, macrophages and T-lymphocytes were observed in the plaque area of FPHs-fed mice compared to control animals. Of note, these effects were associated with lower levels of pro-inflammatory cytokines in the serum and aorta [88,89,90,91,92]. Interestingly, two studies showed that taurine proved efficacy in reducing the atherosclerosis development in both apoE-deficient mice fed chow diet with or without TMAO [93,94]. Furthermore, the authors demonstrated that dietary taurine exerted its anti-atherosclerotic effects via increasing the hepatic gene expression of conjugated bile acid synthesis and eventually, increasing the conjugated BA to unconjugated BA ratio in the liver as well as serum. Meanwhile, taurine improved the TMAO-induced abnormal bile acid profile in the gallbladder. Moreover, taurine increased bile acid deconjugation, by enhancing the genera Ruminiclostridium level, and excretion of fecal neutral sterols. In line with the data obtained with the FPHs, taurine positively modified the TMAO-induced inflammation in both serum and aorta [93,94].
2.2. Clinical Studies
Diverse clinical study investigating the impact of fish-derived peptides on CVD’ risk factors have been performed thus far. Three reviews summarized the clinical trials published before 2020 [31,48,79]. Additionally, a recent systematic review and meta-analysis of randomized controlled trials (RCTs) investigated the impact of lean fish and fish-derived proteins consumption on lipid profile [95]. Even though some of the studies reported an overall positive metabolic effect of consuming different fish protein hydrolysate, such as effect on body weight [96,97], TG and TC concentration [98,99,100], glucose metabolism [101,102,103,104] and hypertension [105], the majority of RCTs show highly inconsistent results. Indeed, Tou et al. concluded that additional better-designed, longer, and larger RCTs are mandatory to achieve a final statement of the impact of lean fish and fish proteins on serum lipid levels. Moreover, these new clinical studies are needed to appropriately inform the public about nutritional differences among fish species, eventually helping consumers to make more evidence-based dietary choices [95]. In line with the above-citated experimental data, more clinical studies documented the beneficial activity of taurine on CVD’ risk factors [31]. Notably, taurine seems to reduce blood pressure by acting as an antagonist of angiotensin II [106].
3. Seaweed Components and CVDs Risk Factors
Different studies have shown beneficial effects of seaweed polyphenols, especially florotannins, on inflammation, oxidative stress, hyperglycemia, and hyperlipidaemia [107]. Additionally, fucoxanthin and brown seaweed-derived florotannins have significant anti-inflammatory and antioxidant activities, which may contribute to CV protection [108]. Moreover, marine microalgae provide vital nutrients and metabolites, including carotenoids and polysaccharides, which may help prevent heart disease [109]. By including these substances into functional foods, the worldwide burden of CVDs may be reduced and CV health may be improved. Overall, despite ongoing clinical application challenges the range of pharmacological activity of marine compounds offer a unique possibility for novel CVD treatments [110].
3.1. Experimental Studies
Algal polysaccharides, particularly fucoidan and laminarin, have attracted attention for their therapeutic potential in atherosclerosis. In vitro studies have shown that algal polysaccharides can inhibit LDL oxidation, a critical step in the atherosclerosis development. This antioxidant effect is essential for preventing the formation of foam cells, which contribute to plaque formation in the arteries [111]. In addition, the observed anti-inflammatory properties of these polysaccharides, may help at reducing vascular inflammation, eventually supporting CV health. Moreover, these compounds have been shown to inhibit atherosclerotic plaque formation and improve endothelial function [111].
Studies in genetically modified mice demonstrated the role of fucoidan (a polysaccharide composed of L-fucose extracted from brown seaweed) in the atherosclerosis management [112,113]. Specifically, these studies shown that intragastric gavage or intraperitoneal administration of fucoidan proved efficacy in decreasing arterial plaque formation as well as macrophage plaque accumulation and smooth muscle cell proliferation (Table 1).
In addition, the authors reported the fucoidan ability to improve the lipid profile, such as a reduction of TC, HDL-C and TG and a rise of HDL-C. By looking for the potential mechanisms of action, they found that in fucoidan-treated mice the hepatic gene expression of sterol regulatory element-binding protein 1 (SREBP1), acetyl-CoA carboxylase (ACC), fatty acid synthetase (FAS), SREBP2 and HMG-CoA reductase was down-regulated, whereas LDL-R gene expression was up-regulated compared with control animals [112,113]. In conclusion, these data suggest that fucoidan is able to impair atherosclerotic plaque development by increasing the lipid metabolism/uptake and decreasing the lipid synthesis. Finally, they also reported a reduction of reactive oxidative species (ROS) as well as of pro-inflammatory mediators. In other words, these compounds could be used as dietary supplements for the atherosclerosis prevention and management.
Eisenia bicyclis (Kjellman) Setchell (EEB) 30% ethanol extract's anti-obesity action has been tested on 3T3-L1 preadipocytes and C57BL/6 mice fed (HFD) [114]. The 3T3-L1 cells' differentiation, proliferation, and mitotic clonal expansion (MCE) were all lowered by EEB treatment. In the subcutaneous and liver tissues of HFD-fed mice, oral treatment of EEB inhibited lipogenesis and adipogenesis. While, EEB increased thermogenesis in brown adipose tissue (BAT) [114]. Mice given oral doses of Monostroma nitidum's rhamnan sulphate (RS), showed a considerable rise in body weight and food intake, accompanied by a decrease in plasma TC, glucose and insulin levels. This latter effect testifies that RS improves insulin resistance. RS feeding modified GM by activating pathways linked to glycolysis and the tricarboxylic acid cycle (TCA) and reducing the Firmicutes/Bacteroidetes (F/B) ratio, thus exerting an-anti-obesity action [115].
In mice fed high-fat/high cholesterol diet, the serum levels of TC, LDL-C, and TG were significantly lowered, and the expression of NPC1L1, a crucial transporter for intestinal cholesterol absorption was downregulated, by laminarin treatment [116].
Oligosaccharide extracted from Enteromorpha prolifera oligosaccharide (EPO) manifested antioxidative, anti-inflammatory, and anti-diabetic effects, when administrated to mice [117]. EPO regulates the crotonylation of XPO1 (Exportin for nuclear export of NES-containing proteins and RNAs) and HSPA8 HSPA8 (heat shock protein family A member 8) proteins, modulating the expression of key genes involved in cell cycle and aging. EPO is also involved in glucose metabolism by inhibiting the crotonylation of HSPA8-K126 and activating the AKT pathway. Finally, the crotonylation of histones in intestinal cells, increasing the abundance of butyric acid-producing bacteria Ruminococcaceae, are promoted by EPO as well [117]. In a diabetic mouse model, Enteromorpha prolifera's sulfated polysaccharide (EP) enhanced glucose tolerance, decreased blood glucose levels, and boosted liver glycogen content. EP's ability to boost AKT phosphorylation was found to be responsible for its antidiabetic benefits. This, in turn, led to the reduction of Glycogen synthase kinase-3β (GSK-3β) and Forkhead box protein O1 (FOXO1) activity, which in turn encouraged glycogen production and reduced gluconeogenesis [117]. In addition, EP inhibited the elevated expression of O-GlcNAc transferase (OGT) induced by diabetes, suggesting that it acted similarly to an OGT inhibitor and contributed to mitigate hyperglycaemia [118].
The three Undaria pinnatifida extracts, UPLW, Undaria pinnatifida low-temperature water extract; UPHW, Undaria pinnatifida high-temperature water extract; UPE, Undaria pinnatifida ethanol extract (UPLW, UPHW, and UPE) had different chemical profiles, suggesting that the extraction approach had an impact on the extracts' composition [119]. UPLW exhibited the highest inhibitory impact on sucrose, but UPHW extract was more effective in inhibiting α-glucosidase activity toward maltose. After oral delivery of glucose, maltose, or sucrose to mice, the UPLW extract was the most successful in lowering postprandial blood glucose levels in those animals [119].
A recent review by Fernando et al. [120] describes the methodologies to obtain protein and protein-hydrolysates from microalgae, and the studies demonstrated their biological properties. The nutritional relevance of the microalgae is due to their high protein content, ranging from 50 to 70%, based on species, growth phase and light quality [120]. Similar to what mentioned earlier for FPHs, these microalgae-derived proteins, mainly their bioactive peptides obtained through hydrolysis by proteolytic enzymes or microorganisms, display antioxidant, anti-inflammatory, antihypertensive, and immunomodulatory effects, which are crucial in reducing CVD risk factors. Antioxidant peptides have been produced by the hydrolysis of Chlorella sp. [121], Navicula sp. [122] and Spiurulina sp [123] and extensively investigated in in vitro experiments. However, more studies are need to well identify the amino acid sequences with antioxidant capacity and then to verify this data in vivo models as well as clinical trials. Antihypertensive peptides derived from Chlorella vulgaris, Chlorella ellipsiodea, Spirulina platensis and Nannochloropsis oculate have been tested using both in vitro and in vivo (mainly using SHRs) experimental settings [124,125]. The most potent antihypertensive peptides seem to contain a high percentage of hydrophobic amino acid residues, such as proline, and the basic mechanisms are related to ACE and renin inhibitions [126]. Finally, two in vitro studies demonstrated the capacity of peptides isolated from Chlorella pyrenoidosa and Spirulina maxima to down-regulated the gene expression levels of E- and P-selectins, intercellular cell adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1), monocyte chemoattractant protein (MCP-1) and endothelin-1 (ET-1) [127,128].
Although the potential CVD efficacy of proteins produced from algae appear promising, more in vivo experimental studies are required to confirm these results. Finally, clinical trials are needed to validate the pre-clinical data and to investigate the potential applications of these proteins in functional foods and nutraceuticals [120,129].
3.2. Clinical Studies
According to a review, polyphenol-rich marine extracts can successfully can significantly reduce plasma TC, LDL-C, and glucose levels, but didn’t find relevant effects of seaweed polyphenols on other biomarkers, such as postprandial blood glucose, fasting insulin, related to CVDs, indicating the need for further research to clarify these relationships [130]. The contrasting results could be explained by variations in the types of polyphenols used, population characteristics, and study design.A form of green microalgae called Chlorella has also been the subject of several clinical studies examining its impact on CV risk factors [131,132,133]. A systematic review found that supplementation with Chlorella can reduce fasting blood glucose, TC and LDL-C concentrations and systolic and diastolic blood pressure, but no effects were observed on TG and HDL-C levels [134]. Chlorella's rich nutritional profile, which includes antioxidants and phytochemicals that may act in concert to support CV health, is believed to be the source of its health benefits [133].
Gdue is an algal extract made from the combination of Ascophyllum nodosum and Fucus vesiculosus, together with chromium picolinate [135]. The two algae that make up Gdue have a 95:5 ratio (Ascophyllum nodosum 95, Fucus vesiculosus 5), and can help at maintaining body weight and promoting metabolism, especially for fats and carbohydrates [135]. Specifically, overweight or obese patients with elevated fasting LDL-C were treated with Gdue for six-months. Gdue-treated patients showed metabolic syndrome metrics improvements, i.e., reduction of body weight and waist circumference, blood pressure, fasting blood TG, LDL-C and glucose concentrations, associated with a drop in HbA1c, and a rise of HDL-C levels. Overall, the study showed a 27.7% relative CVD risk reduction [135].
Finally, ProAlgaZyme is a novel and proprietary infusion of freshwater algae in purified water. The infusion effects on the CV risk factors associated with metabolic syndrome are assessed in a randomized double-blind placebo-controlled study with 60 overweight and obese individuals aged 25 to 60 [136]. Over the course of ten weeks, ProAlgaZyme consumption significantly improved serum lipid profiles, reduced inflammatory markers, and significantly decreased weight and glucose levels in overweight and obese subjects [136]. According to a recent review [137], marine polyphenols found in algae, fish and crustaceans possess anti-inflammatory properties that may help at reducing inflammatory responses, eventually improving to CVD profile.
4. Beneficial Effects of Probiotic/Prebiotics and Marine Derived Compounds
Due to potential health benefits, especially regarding GM and CVDs, marine-derived ingredients have stimulated a bunch of investigations. Additionally, marine sources are a rich source of prebiotics, which are non-digestible food components that specifically promote the growth and activity of healthy gut microbiota.
Probiotics are able to reduce TMAO concentrations, with Lactobacillus rhamnosus GG showing efficacy in both human and animal studies. However, the probiotics’ effects on TMAO reduction appear to be strain-specific [138]. Furthermore probiotics, prebiotics, and synbiotics have demonstrated efficacy in lowering cholesterol concentrations, mitigating inflammation, and exhibiting antioxidative and antiplatelet effects [139]. Both probiotics and prebiotics can modify the GM composition, promoting the proliferation of beneficial bacterial strains and rectifying dysbiosis linked to CVD risk factors [140].
Additionally, faecal microbiota transplantation has emerged as a potential therapeutic approach for CVDs [141]. Altogether, these data underscore the relevance of gut microbiota-targeted strategies in CVD prevention and treatment. Nevertheless, despite encouraging findings from in vitro studies, animal research, and select human clinical trials, additional rigorously designed clinical investigations are imperative to comprehensively elucidate the long-term effects and underlying mechanisms of these dietary interventions in the context of CVD prevention and treatment [139].
Recent studies have placed a great deal of efforts in studying the function of fish-derived proteins and how they affect the human microbiome. In detail one study documents that dietary protein sources, including fish hydrolysates, can alter GM composition and enhance beneficial bacteria [25]. Importantly, FPHs possess excellent digestibility, absorption, water-holding capacity, texture, gelling, whipping, and emulsification properties when introduced into the food matrix [142].
Gabolysat®, a fish protein hydrolysate extracted from cod and mackerel, known for having anxiolytic [143] and gastric protective effects [42], may exert beneficial actions on the colonic mucosal barrier integrity, especially by increasing the mRNA levels of the anti-inflammatory cytokine, interleukin 10 (IL-10) [145]. Fish protein hydrolysates, such as those from salmon and mackerel, have been shown to increase beneficial bacteria while reducing harmful strains in mice GM, suggesting a protective role against metabolic diseases [146]. In addition, Sivixay et al demonstrated that the combination of fish proteins and the prebiotic raffinose positively affect the GM composition and its metabolic functions. Specifically, the data evidenced that this dietary combination influenced the GM diversity by increasing the Akkermansia muciniphila abundance. This bacterium is the most abundant species in the human intestinal microbiota and it has been inversely associated with body weight, inflammatory index, insulin resistance, glucose tolerance, and development of atherosclerosis in several experimental studies [147,148].
Marine Algae Polysaccharides (MAPs) have the potential to modify the gut microbiota in a way that improves heart failure treatment and CV health. MAPs can stimulate GM to produce healthier SCFAs. By influencing gut microbiome's regulation of bile acid metabolism MAPs can also improve CV health [149]. According to De Brito Alves et al. [150], supplementation of Spirulina platensis increases production of beneficial microbial metabolites like SCFAs, improve gut barrier function, enhancing the GM diversity and composition, which, as previously reported, has been linked to a number of health advantages, including anti-inflammatory, anti-obesity, and anti-diabetic effects. Anyways, to evaluate Spirulina platensis's impact directly on the gut microbiome, more clinical trials are still needed.
5. Conclusions
CVDs represent the number one cause of death globally and atherosclerosis is the dominant cause of CVDs. Risk factors for atherosclerotic lesion’s development are dyslipidaemia, hypertension, cigarette smoking, obesity and diabetes mellitus. Diverse evidence also highlighted the role played by inflammation and clonal haematopoiesis, eventually leading to the immunity involvement. Of note, it has been observed that in patients already taking statin therapy, residual inflammatory risk appears to be strongly associated with future CV events than residual cholesterol risk. The GM acts as an endocrine organ, playing a role in modulating immunity and influencing the development of inflammatory, metabolic and infectious diseases. In fact, gut dysbiosis have been associated with inflammatory based-diseases ranging from localized gastrointestinal disorders to metabolic and CV illnesses. There is growing evidence that FPHs and marine-derived compounds, especially those generated by algae, have positive effects on the CV system. In addition, including marine-derived compounds in dietary choices is a viable way to improve the GM health and lower the risk of CVDs. Prebiotics and marine probiotics work synergistically to promote gut health, which is essential for preserving CV health. However, additional and specific in vivo experimental and clinical studies are needed to draw an adequate conclusion. Finally, tailored clinical trials including patients in primary and secondary CV preventions, are mandatory to design new therapeutic protocols, where the “standard of care” therapies will be implemented with these marine-derived ingredients, aiming at reducing the side effects of the traditional drugs [109].
Author Contributions
Font investigation and literature review: I.L., and C.P. Conceptualization, I.L., and C.P.; writing—original draft preparation, I.L., and C.P.; writing—review and editing, I.L., A.A., and C.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MUR Progetto Eccellenza, by BANDO DI ATENEO PROGETTI “PROBLEM-DRIVEN”- PROGETTO FONZIE (D.R.1116/2022 prot.202389) and by European Union - Next Generation EU, in the context of The National Recovery and Resilience Plan, Investment 1.5 Ecosystems of Innovation, Project Tuscany Health Ecosystem (THE), CUP: B83C22003920001.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Benjamin EJ, Blaha MJ, Chiuve SE, Cushman M, Das SR, Deo R, et al. Heart Disease and Stroke Statistics-2017 Update: A Report From the American Heart Association. Circulation. 2017 Mar 7;135(10):e146–603. [CrossRef]
- Libby P, Ridker PM, Hansson GK. Progress and challenges in translating the biology of atherosclerosis. Nature. 2011 May 19;473(7347):317–25. [CrossRef]
- Libby P, Buring JE, Badimon L, Hansson GK, Deanfield J, Bittencourt MS, et al. Atherosclerosis. Nat Rev Dis Primers. 2019 Aug 16;5(1):56.
- Liberale L, Badimon L, Montecucco F, Lüscher TF, Libby P, Camici GG. Inflammation, Aging, and Cardiovascular Disease: JACC Review Topic of the Week. J Am Coll Cardiol. 2022 Mar 1;79(8):837–47.
- Marchesi M, Parolini C, Caligari S, Gilio D, Manzini S, Busnelli M, et al. Rosuvastatin does not affect human apolipoprotein A-I expression in genetically modified mice: a clue to the disputed effect of statins on HDL. British Journal of Pharmacology. 2011 Nov;164(5):1460. [CrossRef]
- Ridker PM, Bhatt DL, Pradhan AD, Glynn RJ, MacFadyen JG, Nissen SE, et al. Inflammation and cholesterol as predictors of cardiovascular events among patients receiving statin therapy: a collaborative analysis of three randomised trials. Lancet. 2023 Apr 15;401(10384):1293–301. [CrossRef]
- Parolini C. Marine n-3 polyunsaturated fatty acids: Efficacy on inflammatory-based disorders. Life Sci. 2020:263:118591. [CrossRef]
- Parolini C, Marchesi M, Chiesa G. HDL therapy for the treatment of cardiovascular diseases. Curr Vasc Pharmacol. 2009;7:550–556. [CrossRef]
- Parolini C, Adorni MP, Busnelli M, Manzini S, Cipollari E, Favari E, et al. Infusions of Large Synthetic HDL Containing Trimeric apoA-I Stabilize Atherosclerotic Plaques in Hypercholesterolemic Rabbits. Can J Cardiol. 2019 Oct;35(10):1400–8.
- Jamadade P, Nupur N, Maharana KC, Singh S. Therapeutic Monoclonal Antibodies for Metabolic Disorders: Major Advancements and Future Perspectives. Curr Atheroscler Rep. 2024 Jul 15; [CrossRef]
- Onvani S, Haghighatdoost F, Surkan PJ, Larijani B, Azadbakht L. Adherence to the Healthy Eating Index and Alternative Healthy Eating Index dietary patterns and mortality from all causes, cardiovascular disease and cancer: a meta-analysis of observational studies. J Hum Nutr Diet. 2017 Apr;30(2):216–26. [CrossRef]
- Pereira L, Valado A. Algae-Derived Natural Products in Diabetes and Its Complications-Current Advances and Future Prospects. Life (Basel). 2023 Aug 29;13(9):1831. [CrossRef]
- Curini L, Amedei A. Cardiovascular Diseases and Pharmacomicrobiomics: A Perspective on Possible Treatment Relevance. Biomedicines. 2021 Sep 28;9(10):1338. [CrossRef]
- Soldati L, Di Renzo L, Jirillo E, Ascierto PA, Marincola FM, De Lorenzo A. The influence of diet on anti-cancer immune responsiveness. Journal of Translational Medicine. 2018 Mar 20;16(1):75.
- Dübüş EN, Lamminpää I, Nannini G, Niccolai E. Nourishing Immunity and Combatting Neuroinflammation: The Power of Immunonutrition and The Microbiome. FBL. 2023 Aug 24;28(8):178. [CrossRef]
- Lamminpää I, Niccolai E, Amedei A. Probiotics as adjuvants to mitigate adverse reactions and enhance effectiveness in Food Allergy Immunotherapy. Scandinavian Journal of Immunology. n/a(n/a):e13405. [CrossRef]
- Rinninella E, Tohumcu E, Raoul P, Fiorani M, Cintoni M, Mele MC, et al. The role of diet in shaping human gut microbiota. Best Practice & Research Clinical Gastroenterology. 2023 Feb 1;62–63:101828.
- Lamminpää I, Boem F, Amedei A. Health-promoting worms? Prospects and pitfalls of helminth therapy. BioEssays. n/a(n/a):2400080. [CrossRef]
- Harris EV, de Roode JC, Gerardo NM. Diet-microbiome-disease: Investigating diet’s influence on infectious disease resistance through alteration of the gut microbiome. PLoS Pathog. 2019 Oct;15(10):e1007891. [CrossRef]
- Nesci A, Carnuccio C, Ruggieri V, D’Alessandro A, Di Giorgio A, Santoro L, et al. Gut Microbiota and Cardiovascular Disease: Evidence on the Metabolic and Inflammatory Background of a Complex Relationship. International Journal of Molecular Sciences. 2023 Jan;24(10):9087. [CrossRef]
- Guo Y, Li X, Wang Z, Yu B. Gut Microbiota Dysbiosis in Human Hypertension: A Systematic Review of Observational Studies. Front Cardiovasc Med [Internet]. 2021 May 14 [cited 2024 Oct 21];8. Available from: https://www.frontiersin.org/journals/cardiovascular-medicine/articles/10.3389/fcvm.2021.650227/full. [CrossRef]
- Sun S, Lulla A, Sioda M, Winglee K, Wu MC, Jacobs DR, et al. Gut Microbiota Composition and Blood Pressure. Hypertension. 2019 May;73(5):998–1006. [CrossRef]
- Savin Z, Kivity S, Yonath H, Yehuda S. Smoking and the intestinal microbiome. Arch Microbiol. 2018 Jul;200(5):677–84. [CrossRef]
- Sublette MG, Cross TWL, Korcarz CE, Hansen KM, Murga-Garrido SM, Hazen SL, et al. Effects of Smoking and Smoking Cessation on the Intestinal Microbiota. J Clin Med. 2020 Sep 14;9(9):2963. [CrossRef]
- Illiano P, Brambilla R, Parolini C. The mutual interplay of gut microbiota, diet and human disease. FEBS J. 2020 Mar;287(5):833–55. [CrossRef]
- Koeth RA, Levison BS, Culley MK, Buffa JA, Wang Z, Gregory JC, et al. γ-Butyrobetaine is a proatherogenic intermediate in gut microbial metabolism of L-carnitine to TMAO. Cell Metab. 2014 Nov 4;20(5):799–812. [CrossRef]
- Qi J, You T, Li J, Pan T, Xiang L, Han Y, et al. Circulating trimethylamine N-oxide and the risk of cardiovascular diseases: a systematic review and meta-analysis of 11 prospective cohort studies. J Cell Mol Med. 2018 Jan;22(1):185–94. [CrossRef]
- Heianza Y, Ma W, Manson JE, Rexrode KM, Qi L. Gut Microbiota Metabolites and Risk of Major Adverse Cardiovascular Disease Events and Death: A Systematic Review and Meta-Analysis of Prospective Studies. J Am Heart Assoc. 2017 Jun 29;6(7):e004947. [CrossRef]
- Astudillo AA, Mayrovitz HN. The Gut Microbiome and Cardiovascular Disease. Cureus [Internet]. 2021 Apr 16 [cited 2024 Sep 13]; Available from: https://www.cureus.com/articles/55496-the-gut-microbiome-and-cardiovascular-disease.
- Hsu CN, Hou CY, Hsu WH, Tain YL. Cardiovascular Diseases of Developmental Origins: Preventive Aspects of Gut Microbiota-Targeted Therapy. Nutrients. 2021 Jul;13(7):2290. [CrossRef]
- Tørris C, Småstuen MC, Molin M. Nutrients in Fish and Possible Associations with Cardiovascular Disease Risk Factors in Metabolic Syndrome. Nutrients. 2018 Jul 23;10(7):952. [CrossRef]
- Abachi S, Bazinet L, Beaulieu L. Antihypertensive and Angiotensin-I-Converting Enzyme (ACE)-Inhibitory Peptides from Fish as Potential Cardioprotective Compounds. Mar Drugs. 2019 Oct 29;17(11):613. [CrossRef]
- O’Keeffe M, Gudbrandsen OA. Effects of diets containing proteins from fish muscles or fish by-products on the circulating cholesterol concentration in rodents: a systematic review and meta-analysis. Br J Nutr. 2023 Aug 14;130(3):389–410. [CrossRef]
- Jensen IJ, Mæhre HK. Preclinical and Clinical Studies on Antioxidative, Antihypertensive and Cardioprotective Effect of Marine Proteins and Peptides-A Review. Mar Drugs. 2016 Nov 18;14(11):211.
- Serhan CN, Levy BD. Resolvins in inflammation: emergence of the pro-resolving superfamily of mediators. J Clin Invest. 2018 Jul 2;128(7):2657–69. [CrossRef]
- Bäck M, Hansson GK. Omega-3 fatty acids, cardiovascular risk, and the resolution of inflammation. FASEB J. 2019 Feb;33(2):1536–9.
- Parolini C. The Role of Marine n-3 Polyunsaturated Fatty Acids in Inflammatory-Based Disease: The Case of Rheumatoid Arthritis. Mar Drugs. 2023 Dec 27;22(1):17. [CrossRef]
- Snetselaar LG, de Jesus JM, DeSilva DM, Stoody EE. Dietary Guidelines for Americans, 2020–2025. Nutr Today. 2021;56(6):287–95.
- Zhubi-Bakija F, Bajraktari G, Bytyçi I, Mikhailidis DP, Henein MY, Latkovskis G, et al. The impact of type of dietary protein, animal versus vegetable, in modifying cardiometabolic risk factors: A position paper from the International Lipid Expert Panel (ILEP). Clin Nutr. 2021 Jan;40(1):255–76. [CrossRef]
- Zhao LG, Sun JW, Yang Y, Ma X, Wang YY, Xiang YB. Fish consumption and all-cause mortality: a meta-analysis of cohort studies. Eur J Clin Nutr. 2016 Feb;70(2):155–61. [CrossRef]
- Jayedi A, Shab-Bidar S, Eimeri S, Djafarian K. Fish consumption and risk of all-cause and cardiovascular mortality: a dose-response meta-analysis of prospective observational studies. Public Health Nutr. 2018 May;21(7):1297–306. [CrossRef]
- Zhou C, Wu Q, Ye Z, Liu M, Zhang Z, Zhang Y, et al. Inverse Association Between Variety of Proteins With Appropriate Quantity From Different Food Sources and New-Onset Hypertension. Hypertension. 2022 May;79(5):1017–27. [CrossRef]
- Drummen M, Tischmann L, Gatta-Cherifi B, Adam T, Westerterp-Plantenga M. Dietary Protein and Energy Balance in Relation to Obesity and Co-morbidities. Front Endocrinol (Lausanne). 2018;9:443. [CrossRef]
- Pradhan B, Nayak R, Patra S, Jit BP, Ragusa A, Jena M. Bioactive Metabolites from Marine Algae as Potent Pharmacophores against Oxidative Stress-Associated Human Diseases: A Comprehensive Review. Molecules. 2020 Dec 23;26(1):37. [CrossRef]
- Bocanegra A, Macho-González A, Garcimartín A, Benedí J, Sánchez-Muniz FJ. Whole Alga, Algal Extracts, and Compounds as Ingredients of Functional Foods: Composition and Action Mechanism Relationships in the Prevention and Treatment of Type-2 Diabetes Mellitus. Int J Mol Sci. 2021 Apr 7;22(8):3816. [CrossRef]
- Suleria HAR, Osborne S, Masci P, Gobe G. Marine-Based Nutraceuticals: An Innovative Trend in the Food and Supplement Industries. Mar Drugs. 2015 Oct 14;13(10):6336–51. [CrossRef]
- Marchesi M, Parolini C, Valetti C, Mangione P, Obici L, Giorgetti S, et al. The intracellular quality control system down-regulates the secretion of amyloidogenic apolipoprotein A-I variants: a possible impact on the natural history of the disease. Biochim Biophys Acta. 2011 Jan;1812(1):87–93. [CrossRef]
- Dale HF, Madsen L, Lied GA. Fish-derived proteins and their potential to improve human health. Nutr Rev. 2019 Aug 1;77(8):572–83. [CrossRef]
- Yamori Y, Taguchi T, Hamada A, Kunimasa K, Mori H, Mori M. Taurine in health and diseases: consistent evidence from experimental and epidemiological studies. J Biomed Sci. 2010 Aug 24;17 Suppl 1(Suppl 1):S6. [CrossRef]
- Caligari S, Chiesa G, Johnson SK, Camisassi D, Gilio D, Marchesi M, et al. Lupin (Lupinus albus) protein isolate (L-ISO) has adequate nutritional value and reduces large intestinal weight in rats after restricted and ad libitum feeding.Ann Nutr Metab. 2006;50:528-537. [CrossRef]
- Parolini C, Caligari S, Gilio D, Manzini S, Busnelli M, Montagnani M, et al. Reduced biliary sterol output with no change in total faecal excretion in mice expressing a human apolipoprotein A-I variant. Liver Int. 2012 Oct;32(9):1363–71. [CrossRef]
- Parolini C, Busnelli M, Ganzetti GS, Dellera F, Manzini S, Scanziani E, et al. Magnetic resonance imaging visualization of vulnerable atherosclerotic plaques at the brachiocephalic artery of apolipoprotein E knockout mice by the blood-pool contrast agent B22956/1. Mol Imaging. 2014;13.
- Maeda H, Hosomi R, Yokoyama T, Ikeda Y, Nishimoto A, Tanaka G, et al. Dietary Alaska pollock protein attenuates liver steatosis and alters gut microbiota in leptin-deficient ob/ob mice. Journal of Functional Foods. 2020 Dec 1;75:104266. [CrossRef]
- Hosomi R, Fukunaga K, Arai H, Nishiyama T, Yoshida M. Effects of dietary fish protein on serum and liver lipid concentrations in rats and the expression of hepatic genes involved in lipid metabolism. J Agric Food Chem. 2009 Oct 14;57(19):9256–62. [CrossRef]
- Parolini C, Rigamonti E, Marchesi M, Busnelli M, Cinquanta P, Manzini S, et al. Cholesterol-lowering effect of dietary Lupinus angustifolius proteins in adult rats through regulation of genes involved in cholesterol homeostasis. Food Chem. 2012 Jun 1;132(3):1475–9. [CrossRef]
- Wang K, Han L, Tan Y, Hong H, Fan H, Luo Y. Novel Hypocholesterolemic Peptides Derived from Silver Carp Muscle: The Modulatory Effects on Enterohepatic Cholesterol Metabolism In Vitro and In Vivo. J Agric Food Chem. 2023 Apr 12;71(14):5565–75. [CrossRef]
- Mijiti M, Mori R, Huang B, Tsukamoto K, Kiriyama K, Sutoh K, et al. Anti-Obesity and Hypocholesterolemic Actions of Protamine-Derived Peptide RPR (Arg-Pro-Arg) and Protamine in High-Fat Diet-Induced C57BL/6J Mice. Nutrients. 2021 Jul 22;13(8):2501. [CrossRef]
- Hosomi R, Fukunaga K, Arai H, Kanda S, Nishiyama T, Yoshida M. Effect of dietary protamine on lipid metabolism in rats. Nutr Res Pract. 2010 Dec;4(6):462–9. [CrossRef]
- Hosomi R, Miyauchi K, Yamamoto D, Arai H, Nishiyama T, Yoshida M, et al. Salmon Protamine Decreases Serum and Liver Lipid Contents by Inhibiting Lipid Absorption in an In Vitro Gastrointestinal Digestion Model and in Rats. J Food Sci. 2015 Oct;80(10):H2346-2353. [CrossRef]
- Luiking YC, Ten Have GAM, Wolfe RR, Deutz NEP. Arginine de novo and nitric oxide production in disease states. Am J Physiol Endocrinol Metab. 2012 Nov 15;303(10):E1177-1189. [CrossRef]
- Ohwada T, Ishibashi T, Yaoita H, Shindo J, Noji H, Ohkawara H, et al. Different contribution of apoptosis to the antiproliferative effects of L-arginine, enalapril and losartan on neointimal growth inhibition after balloon arterial injury. Circ J. 2002 Oct;66(10):965–71. [CrossRef]
- Dellera F, Ganzetti GS, Froio A, Manzini S, Busnelli M, Meinitzer A, et al. L-homoarginine administration reduces neointimal hyperplasia in balloon-injured rat carotids. Thromb Haemost. 2016 Aug 1;116(2):400–2. [CrossRef]
- Bai Y, Sun L, Yang T, Sun K, Chen J, Hui R. Increase in fasting vascular endothelial function after short-term oral L-arginine is effective when baseline flow-mediated dilation is low: a meta-analysis of randomized controlled trials. Am J Clin Nutr. 2009 Jan;89(1):77–84. [CrossRef]
- Lv Z, Zhang C, Song W, Chen Q, Wang Y. Jellyfish Collagen Hydrolysate Alleviates Inflammation and Oxidative Stress and Improves Gut Microbe Composition in High-Fat Diet-Fed Mice. Mediators Inflamm. 2022;2022:5628702. [CrossRef]
- Shi M, Song R, Gu L. Different Regulatory Effects of Heated Products and Maillard Reaction Products of Half-Fin Anchovy Hydrolysates on Intestinal Antioxidant Defense in Healthy Animals. Int J Mol Sci. 2023 Jan 25;24(3):2355. [CrossRef]
- Parolini C, Manzini S, Busnelli M, Rigamonti E, Marchesi M, Diani E, et al. Effect of the combinations between pea proteins and soluble fibres on cholesterolaemia and cholesterol metabolism in rats. Br J Nutr. 2013 Oct;110(8):1394–401. [CrossRef]
- Hernandez-Hernandez O, Sanz ML, Kolida S, Rastall RA, Moreno FJ. In vitro fermentation by human gut bacteria of proteolytically digested caseinomacropeptide nonenzymatically glycosylated with prebiotic carbohydrates. J Agric Food Chem. 2011 Nov 23;59(22):11949–55. [CrossRef]
- Monteagudo-Mera A, Arthur JC, Jobin C, Keku T, Bruno-Barcena JM, Azcarate-Peril MA. High purity galacto-oligosaccharides enhance specific Bifidobacterium species and their metabolic activity in the mouse gut microbiome. Benef Microbes. 2016;7(2):247–64. [CrossRef]
- Terpend K, Possemiers S, Daguet D, Marzorati M. Arabinogalactan and fructo-oligosaccharides have a different fermentation profile in the Simulator of the Human Intestinal Microbial Ecosystem (SHIME ®). Environ Microbiol Rep. 2013 Aug;5(4):595–603. [CrossRef]
- Joubran Y, Moscovici A, Portmann R, Lesmes U. Implications of the Maillard reaction on bovine alpha-lactalbumin and its proteolysis during in vitro infant digestion. Food Funct. 2017 Jun 21;8(6):2295–308. [CrossRef]
- Jin W, Han K, Dong S, Yang Y, Mao Z, Su M, et al. Modifications in gut microbiota and fermentation metabolites in the hindgut of rats after the consumption of galactooligosaccharide glycated with a fish peptide. Food Funct. 2018 May 1;9(5):2853–64. [CrossRef]
- Han K, Yao Y, Dong S, Jin S, Xiao H, Wu H, et al. Chemical characterization of the glycated myofibrillar proteins from grass carp (Ctenopharyngodon idella) and their impacts on the human gut microbiota in vitro fermentation. Food Funct. 2017 Mar 22;8(3):1184–94. [CrossRef]
- Lin Q, Guo Y, Li J, He S, Chen Y, Jin H. Antidiabetic Effect of Collagen Peptides from Harpadon nehereus Bones in Streptozotocin-Induced Diabetes Mice by Regulating Oxidative Stress and Glucose Metabolism. Mar Drugs. 2023 Sep 29;21(10):518. [CrossRef]
- G Bardallo R, Panisello-Roselló A, Sanchez-Nuno S, Alva N, Roselló-Catafau J, Carbonell T. Nrf2 and oxidative stress in liver ischemia/reperfusion injury. FEBS J. 2022 Sep;289(18):5463–79. [CrossRef]
- Xu M, Chen B, Qiao K, Liu S, Su Y, Cai S, et al. Mechanism of Takifugu bimaculatus Skin Peptides in Alleviating Hyperglycemia in Rats with Type 2 Diabetic Mellitus Based on Microbiome and Metabolome Analyses. Marine Drugs. 2024 Aug;22(8):377. [CrossRef]
- Bodnaruc AM, Prud’homme D, Blanchet R, Giroux I. Nutritional modulation of endogenous glucagon-like peptide-1 secretion: a review. Nutr Metab (Lond). 2016;13:92. [CrossRef]
- Valle M, Mitchell PL, Pilon G, Varin T, Hénault L, Rolin J, et al. Salmon peptides limit obesity-associated metabolic disorders by modulating a gut-liver axis in vitamin D-deficient mice. Obesity. 2021;29(10):1635–49. [CrossRef]
- Fang Y, She J, Zhang X, Gu T, Xie D, Luo X, et al. Discovery of Anti-Hypercholesterolemia Agents Targeting LXRα from Marine Microorganism-Derived Natural Products. J Nat Prod. 2024 Feb 23;87(2):322–31. [CrossRef]
- Wijesekara I, Kim SK. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: prospects in the pharmaceutical industry. Mar Drugs. 2010 Mar 31;8(4):1080–93. [CrossRef]
- Busnelli M, Manzini S, Sirtori CR, Chiesa G, Parolini C. Effects of Vegetable Proteins on Hypercholesterolemia and Gut Microbiota Modulation. Nutrients. 2018:10:1249. [CrossRef]
- Wang S, Lin L mei, Wu Y ning, Fang M, Yu Y qin, Zhou J, et al. Angiotensin I Converting Enzyme (ACE) inhibitory activity and antihypertensive effects of grass carp peptides. Food Sci Biotechnol. 2014 Oct 1;23(5):1661–6.
- Sun L, Zhang Y, Zhuang Y. Antiphotoaging effect and purification of an antioxidant peptide from tilapia (Oreochromis niloticus) gelatin peptides. Journal of Functional Foods. 2013;1(5):154–62. [CrossRef]
- Chen J, Ryu B, Zhang Y, Liang P, Li C, Zhou C, et al. Comparison of an angiotensin-I-converting enzyme inhibitory peptide from tilapia (Oreochromis niloticus) with captopril: inhibition kinetics, in vivo effect, simulated gastrointestinal digestion and a molecular docking study. J Sci Food Agric. 2020 Jan 15;100(1):315–24.
- Drotningsvik A, Oterhals Å, Mjøs SA, Vikøren LA, Flesland O, Gudbrandsen OA. Effects of intact and hydrolysed blue whiting proteins on blood pressure and markers of kidney function in obese Zucker fa/fa rats. Eur J Nutr. 2021 Feb;60(1):529–44. [CrossRef]
- Vo TS, Ngo DH, Kim JA, Ryu B, Kim SK. An antihypertensive peptide from tilapia gelatin diminishes free radical formation in murine microglial cells. J Agric Food Chem. 2011 Nov 23;59(22):12193–7. [CrossRef]
- Ktari N, Bkhairia I, Nasri R, Ben Abdallah Kolsi R, Ben Slama-Ben Salem R, Ben Amara I, et al. Zebra blenny protein hydrolysates as a source of bioactive peptides with prevention effect against oxidative dysfunctions and DNA damage in heart tissues of rats fed a cholesterol-rich diet. Food Res Int. 2017 Oct;100(Pt 1):423–32. [CrossRef]
- Maneesai P, Wattanathorn J, Potue P, Khamseekaew J, Rattanakanokchai S, Thukham-Mee W, et al. Cardiovascular complications are resolved by tuna protein hydrolysate supplementation in rats fed with a high-fat diet. Sci. Rep. 2023;13:12880. [CrossRef]
- Parolini C, Vik R, Busnelli M, , Bjørndal B, Holm S, Brattelid T, et al. A Salmon Protein Hydrolysate Exerts Lipid-Independent Anti-Atherosclerotic Activity in ApoE-Deficient Mice. PLoS One. 2014;9:e97598.
- Abbate JM, Macrì F, Arfuso F, Iaria C, Capparucci F, Anfuso C, et al. Anti-Atherogenic Effect of 10% Supplementation of Anchovy (Engraulis encrasicolus) Waste Protein Hydrolysates in ApoE-Deficient Mice. Nutrients. 2021 Jul;13(7):2137. [CrossRef]
- Liu H, Yang Y, Liu Y, Cui L, Fu L, Li B. Various bioactive peptides in collagen hydrolysate from salmo salar skin and the combined inhibitory effects on atherosclerosis in vitro and in vivo. Food Res Int. 2022 Jul;157:111281. [CrossRef]
- Yang Y, Liu H, Cui L, Liu Y, Fu L, Li B. A Collagen-Derived Oligopeptide from Salmo salar Collagen Hydrolysates Restrains Atherogenesis in ApoE–/– Mice via Targeting P2Y12 Receptor. Molecular Nutrition & Food Research. 2022;66(13):2200166.
- Yang Y, Liu H, Cui L, Liu Y, Fu L, Li B. A Collagen-Derived Oligopeptide from Salmo salar Collagen Hydrolysates Restrains Atherogenesis in ApoE–/– Mice via Targeting P2Y12 Receptor. Mol. Nutr. Food Res. 2022;66:e2200166.
- Kondo Y, Toda Y, Kitajima H, Oda H, Nagate T, Kameo K, et al. Taurine Inhibits Development Of Atherosclerotic Lesions In apolipoprotein E-Deficient Mice. Clin. Exp. Pharmacol. Physiol. 2001;28:809-815.
- Yang JY, Zhang TT, Yu ZL, Wang CC, Zhao YC, Wang YM, et al. Taurine Alleviates Trimethylamine N-Oxide-Induced Atherosclerosis by Regulating Bile Acid Metabolism in ApoE–/– Mice. J Agric Food Chem. 2022 May 11;70(18):5738–47. [CrossRef]
- Tou JC, Gucciardi E, Young I. Lipid-modifying effects of lean fish and fish-derived protein consumption in humans: a systematic review and meta-analysis of randomized controlled trials. Nutr Rev. 2021 Dec 8;80(1):91–112. [CrossRef]
- Ramel A, Jonsdottir MT, Thorsdottir I. Consumption of cod and weight loss in young overweight and obese adults on an energy reduced diet for 8-weeks. Nutr Metab Cardiovasc Dis. 2009 Dec;19(10):690–6. [CrossRef]
- Nobile V, Duclos E, Michelotti A, Bizzaro G, Negro M, Soisson F. Full article: Supplementation with a fish protein hydrolysate (Micromesistius poutassou): effects on body weight, body composition, and CCK/GLP-1 secretion. Food Nutr. Res. 2016;60:29857.
- Telle-Hansen VH, Larsen LN, Høstmark AT, Molin M, Dahl L, Almendingen K, et al. Daily intake of cod or salmon for 2 weeks decreases the 18:1n-9/18:0 ratio and serum triacylglycerols in healthy subjects. Lipids. 2012 Feb;47(2):151–60.
- Aadland EK, Lavigne C, Graff IE, Eng Ø, Paquette M, Holthe A, et al. Lean-seafood intake reduces cardiovascular lipid risk factors in healthy subjects: results from a randomized controlled trial with a crossover design. Am J Clin Nutr. 2015 Sep;102(3):582–92. [CrossRef]
- Aadland EK, Graff IE, Lavigne C, Eng Ø, Paquette M, Holthe A, et al. Lean Seafood Intake Reduces Postprandial C-peptide and Lactate Concentrations in Healthy Adults in a Randomized Controlled Trial with a Crossover Design. J Nutr. 2016 May;146(5):1027–34. [CrossRef]
- Vikøren LA, Nygård OK, Lied E, Rostrup E, Gudbrandsen OA. A randomised study on the effects of fish protein supplement on glucose tolerance, lipids and body composition in overweight adults. Br J Nutr. 2013 Feb 28;109(4):648–57. [CrossRef]
- Dale HF, Jensen C, Hausken T, Lied E, Hatlebakk JG, Brønstad I, et al. Effect of a cod protein hydrolysate on postprandial glucose metabolism in healthy subjects: a double-blind cross-over tria. J Nutr. Sci. 2018;7:e33.l.
- Ouellet V, Marois J, Weisnagel SJ, Jacques H. Dietary cod protein improves insulin sensitivity in insulin-resistant men and women: a randomized controlled trial. Diabetes Care. 2007;30:2816-2821.
- Ouellet V, Weisnagel SJ, Marois J, Bergeron J, Julien P, Gougeon R, et al. Dietary cod protein reduces plasma C-reactive protein in insulin-resistant men and women. J Nutr. 2008;138:2386-2391. [CrossRef]
- Kawasaki T, Seki E, Osajima K, Yoshida M, Asada K, Matsui T, et al. Antihypertensive effect of valyl-tyrosine, a short chain peptide derived from sardine muscle hydrolyzate, on mild hypertensive subjects. J Hum Hypertens. 2000 Aug;14(8):519–23. [CrossRef]
- Sun Q, Wang B, Li Y, Sun F, Li P, Xia W, et al. Taurine Supplementation Lowers Blood Pressure and Improves Vascular Function in Prehypertension: Randomized, Double-Blind, Placebo-Controlled Study. Hypertension. 2016 Mar;67(3):541–9.
- Murray M, Dordevic AL, Ryan L, Bonham MP. An emerging trend in functional foods for the prevention of cardiovascular disease and diabetes: Marine algal polyphenols. Critical Reviews in Food Science and Nutrition. 2018 May 24;58(8):1342–58. [CrossRef]
- Akram W, Rihan M, Ahmed S, Arora S, Ahmad S, Vashishth R. Marine-Derived Compounds Applied in Cardiovascular Diseases: Submerged Medicinal Industry. Marine Drugs. 2023 Mar;21(3):193. [CrossRef]
- Yurika N, Montuori E, Lauritano C. Marine Microalgal Products with Activities against Age-Related Cardiovascular Diseases. Mar. Drugs. 2024;22:229. [CrossRef]
- Giuliani ME, Bigossi G, Lai G, Marcozzi S, Brunetti D, Malavolta M. Marine Compounds and Age-Related Diseases: The Path from Pre-Clinical Research to Approved Drugs for the Treatment of Cardiovascular Diseases and Diabetes. Marine Drugs. 2024 May;22(5):210. [CrossRef]
- Patil NP, Le V, Sligar AD, Mei L, Chavarria D, Yang EY, et al. Algal Polysaccharides as Therapeutic Agents for Atherosclerosis. Front Cardiovasc Med. 2018;5:153.
- Yokota T, Nomura K, Nagashima M, Kamimura N. Fucoidan alleviates high-fat diet-induced dyslipidemia and atherosclerosis in ApoE(shl) mice deficient in apolipoprotein E expression. J Nutr Biochem. 2016 Jun;32:46–54.
- Park J, Yeom M, Hahm DH. Fucoidan improves serum lipid levels and atherosclerosis through hepatic SREBP-2-mediated regulation. J Pharmacol Sci. 2016 Jun;131(2):84–92. [CrossRef]
- Ys Y, Ks C, Sy L, Sw H, Yr K, Jk L, et al. Anti-obesity effects of a standardized ethanol extract of Eisenia bicyclis by regulating the AMPK signaling pathway in 3T3-L1 cells and HFD-induced mice. Food & function [Internet]. 2024 Jun 17 [cited 2024 Sep 18];15(12). Available from: https://pubmed.ncbi.nlm.nih.gov/38771619/.
- Y S, L Z, T I, K N, K M, R U, et al. Lipid- and glucose-lowering effects of Rhamnan sulphate from Monostroma nitidum with altered gut microbiota in mice. Food science & nutrition [Internet]. 2024 Mar 25 [cited 2024 Sep 18];12(6). Available from: https://pubmed.ncbi.nlm.nih.gov/38873438/.
- He Z, Zhang Z, Xu P, Dirsch VM, Wang L, Wang K. Laminarin Reduces Cholesterol Uptake and NPC1L1 Protein Expression in High-Fat Diet (HFD)-Fed Mice. Mar Drugs. 2023 Nov 29;21(12):624. [CrossRef]
- Shan S, Zhang Z, Nie J, Wen Y, Wu W, Liu Y, et al. Marine algae-derived oligosaccharide via protein crotonylation of key targeting for management of type 2 diabetes mellitus in the elderly.Pharmacol Res. 2024;205:107257. [CrossRef]
- Lin D, Zhang N, Wu S, Wang S, Huang F, Lin Y, et al. Structural Analysis and Novel Mechanism of Enteromorpha prolifera Sulfated Polysaccharide in Preventing Type 2 Diabetes Mellitus. Plant Foods Hum Nutr [Internet]. 2023 Dec 12 [cited 2024 Sep 18]; Available from: https://link.springer.com/10.1007/s11130-023-01129-8. [CrossRef]
- Lee SM, Park S-Y, Kim JY. Comparative evaluation of the antihyperglycemic effects of three extracts of sea mustard (Undaria pinnatifida): In vitro and in vivo studies. Food Res Int. 2024;190:114623. [CrossRef]
- Fernando R, Sun X, Rupasinghe HPV. Production of Bioactive Peptides from Microalgae and Their Biological Properties Related to Cardiovascular Disease. Macromol. 2024 Sep;4(3):582–96. [CrossRef]
- Olena Z, Yang Y, TingTing Y, XiaoTao Y, HaiLian R, Xun X, et al. Simultaneous preparation of antioxidant peptides and lipids from microalgae by pretreatment with bacterial proteases. Bioresour Technol. 2022 Mar;348:126759. [CrossRef]
- Kang KH, Qian ZJ, Ryu B, Karadeniz F, Kim D, Kim SK. Antioxidant peptides from protein hydrolysate of microalgae Navicula incerta and their protective effects in HepG2/CYP2E1 cells induced by ethanol. Phytother Res. 2012 Oct;26(10):1555–63.
- Abo-Shady AM, Gheda SF, Ismail GA, Cotas J, Pereira L, Abdel-Karim OH. Antioxidant and Antidiabetic Activity of Algae. Life (Basel). 2023 Feb 7;13(2):460. [CrossRef]
- Suetsuna K, Chen JR. Identification of antihypertensive peptides from peptic digest of two microalgae, Chlorella vulgaris and Spirulina platensis. Mar Biotechnol (NY). 2001 Jul;3(4):305–9. [CrossRef]
- Sheih IC, Wu TK, Fang TJ. Antioxidant properties of a new antioxidative peptide from algae protein waste hydrolysate in different oxidation systems. Bioresour Technol. 2009 Jul;100(13):3419–25. [CrossRef]
- Fitzgerald C, Gallagher E, Tasdemir D, Hayes M. Heart Health Peptides from Macroalgae and Their Potential Use in Functional Foods. J Agric Food Chem. 2011 Jul 13;59(13):6829–36. [CrossRef]
- Shih MF, Chen LC, Cherng JY. Chlorella 11-peptide inhibits the production of macrophage-induced adhesion molecules and reduces endothelin-1 expression and endothelial permeability. Mar Drugs. 2013 Oct 14;11(10):3861–74. [CrossRef]
- Vo TS, Kim SK. Down-regulation of histamine-induced endothelial cell activation as potential anti-atherosclerotic activity of peptides from Spirulina maxima. Eur J Pharm Sci. 2013 Oct 9;50(2):198–207. [CrossRef]
- Ejike CECC, Collins SA, Balasuriya N, Swanson AK, Mason B, Udenigwe CC. Prospects of microalgae proteins in producing peptide-based functional foods for promoting cardiovascular health. Trends in Food Science & Technology. 2017 Jan 1;59:30–6. [CrossRef]
- Cherry P, O’Hara C, Magee PJ, McSorley EM, Allsopp PJ. Risks and benefits of consuming edible seaweeds. Nutr Rev. 2019 May 1;77(5):307–29. [CrossRef]
- Otsuki T, Shimizu K, Maeda S. Changes in arterial stiffness and nitric oxide production with Chlorella-derived multicomponent supplementation in middle-aged and older individuals. J Clin Biochem Nutr. 2015;57(3):228–32. [CrossRef]
- Ryu NH, Lim Y, Park JE, Kim J, Kim JY, Kwon SW, et al. Impact of daily Chlorella consumption on serum lipid and carotenoid profiles in mildly hypercholesterolemic adults: a double-blinded, randomized, placebo-controlled study. Nutrition Journal. 2014 Jun 11;13(1):57. [CrossRef]
- Bito T, Okumura E, Fujishima M, Watanabe F. Potential of Chlorella as a Dietary Supplement to Promote Human Health. Nutrients. 2020 Sep;12(9):2524. [CrossRef]
- Potential of Chlorella as a Dietary Supplement to Promote Human Health - PMC [Internet]. [cited 2024 Sep 17]. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7551956/.
- Nicolucci A, Rossi MC, Petrelli M. Effectiveness of Ascophyllum nodosum and Fucus vesiculosus on Metabolic Syndrome Components: A Real-World, Observational Study. J Diabetes Res. 2021;2021:3389316. [CrossRef]
- Oben J, Enonchong E, Kuate D, Mbanya D, Thomas TC, Hildreth DJ, et al. The effects of ProAlgaZyme novel algae infusion on metabolic syndrome and markers of cardiovascular health. Lipids Health Dis. 2007 Sep 5;6:20. [CrossRef]
- Pereira L, Cotas J. Therapeutic Potential of Polyphenols and Other Micronutrients of Marine Origin. Marine Drugs. 2023 Jun;21(6):323. [CrossRef]
- Cantero MA, Guedes MRA, Fernandes R, Lollo PCB. Trimethylamine N-oxide reduction is related to probiotic strain specificity: A systematic review. Nutrition Research. 2022 Aug 1;104:29–35. [CrossRef]
- Olas B. Probiotics, Prebiotics and Synbiotics—A Promising Strategy in Prevention and Treatment of Cardiovascular Diseases? International Journal of Molecular Sciences. 2020 Jan;21(24):9737.
- Ghanbari F, Hasani S, Aghili ZS, Asgary S. The potential preventive effect of probiotics, prebiotics, and synbiotics on cardiovascular risk factors through modulation of gut microbiota: A review. Food Science & Nutrition. 2024;12(7):4569–80. [CrossRef]
- Murphy K, O’Donovan AN, Caplice NM, Ross RP, Stanton C. Exploring the Gut Microbiota and Cardiovascular Disease. Metabolites. 2021 Jul 29;11(8):493. [CrossRef]
- Cunha SA, Pintado ME. Bioactive peptides derived from marine sources: Biological and functional properties. Trends in Food Science & Technology. 2022 Jan 1;119:348–70. [CrossRef]
- Freret T, Largilliere S, Nee G, Coolzaet M, Corvaisier S, Boulouard M. Fast Anxiolytic-Like Effect Observed in the Rat Conditioned Defensive Burying Test, after a Single Oral Dose of Natural Protein Extract Products. Nutrients. 2021 Jul 17;13(7):2445. [CrossRef]
- Fitzgerald AJ, Rai PS, Marchbank T, Taylor GW, Ghosh S, Ritz BW, et al. Reparative properties of a commercial fish protein hydrolysate preparation. Gut. 2005 Jun;54(6):775–81. [CrossRef]
- Langlois LD, Oddoux S, Aublé K, Violette P, Déchelotte P, Noël A, et al. Effects of Glutamine, Curcumin and Fish Bioactive Peptides Alone or in Combination on Intestinal Permeability in a Chronic-Restraint Stress Model. Int J Mol Sci. 2023 Apr 13;24(8):7220.
- Daskalaki MG, Axarlis K, Tsoureki A, Michailidou S, Efraimoglou C, Lapi I, et al. Fish-Derived Protein Hydrolysates Increase Insulin Sensitivity and Alter Intestinal Microbiome in High-Fat-Induced Obese Mice. Mar Drugs. 2023 Jun 2;21(6):343. [CrossRef]
- Schneeberger M, Everard A, Gómez-Valadés AG, Matamoros S, Ramírez S, Delzenne NM, et al. Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci Rep. 2015 Nov 13;5:16643. [CrossRef]
- Li J, Lin S, Vanhoutte PM, Woo CW, Xu A. Akkermansia Muciniphila Protects Against Atherosclerosis by Preventing Metabolic Endotoxemia-Induced Inflammation in Apoe-/- Mice. Circulation. 2016 Jun 14;133(24):2434–46.
- Cheong KL, Yu B, Chen J, Zhong S. A Comprehensive Review of the Cardioprotective Effect of Marine Algae Polysaccharide on the Gut Microbiota. Foods. 2022 Nov 8;11(22):3550. [CrossRef]
- Alves JL de B, Costa PCT da, Sales LCS de, Silva Luis CC, Bezerra TPT, Souza MLA, et al. Shedding light on the impacts of Spirulina platensis on gut microbiota and related health benefits. Crit Rev Food Sci Nutr. 2024 Feb 29;1–14.
Figure 1.
Schematic representation of the fish proteins hydrolysates (FPHs) mechanism of actions. HMGCoA red: 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase; LDL-R: low-density lipoprotein-receptor, ACAT2: acyl-CoA:cholesterol acyltransferase; NPC1L1: Niemann-Pick C1-like 1; CYP7A1: cholesterol 7-alpha-hydroxylase; GK: glucokinase; PEPCK1; phosphoenolpyruvate carboxikinase1; G6Pase: glucose-6-phosphate; DPP-IV: dipeptidyl peptidase-IV; CAT: catalase; SOD: superoxide dismutase; GSH-Px: glutathione peroxidase; NQO1: quinone oxidoreductase 1; Nrf2: nuclear factor-erythroid 2-related factor 2; IL: interleukin; TNF: tumour necrosis factor; ACE: angiotensin-I-converting enzyme.
Figure 1.
Schematic representation of the fish proteins hydrolysates (FPHs) mechanism of actions. HMGCoA red: 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase; LDL-R: low-density lipoprotein-receptor, ACAT2: acyl-CoA:cholesterol acyltransferase; NPC1L1: Niemann-Pick C1-like 1; CYP7A1: cholesterol 7-alpha-hydroxylase; GK: glucokinase; PEPCK1; phosphoenolpyruvate carboxikinase1; G6Pase: glucose-6-phosphate; DPP-IV: dipeptidyl peptidase-IV; CAT: catalase; SOD: superoxide dismutase; GSH-Px: glutathione peroxidase; NQO1: quinone oxidoreductase 1; Nrf2: nuclear factor-erythroid 2-related factor 2; IL: interleukin; TNF: tumour necrosis factor; ACE: angiotensin-I-converting enzyme.
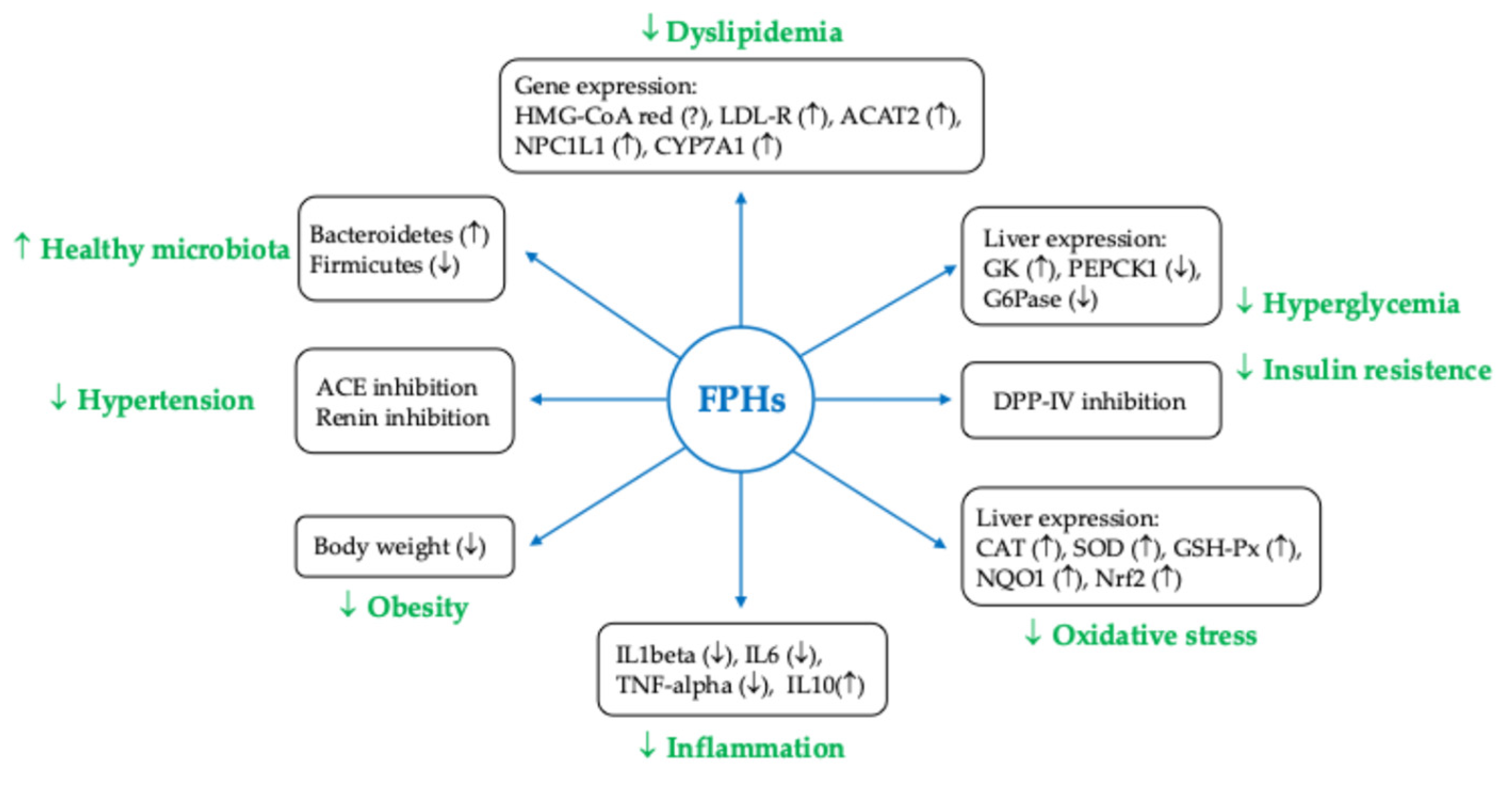

Table 1.
The main mechanisms of action of seaweed compounds in promoting cardiovascular health involve their antioxidant, anti-inflammatory, and metabolic modulation effects.
Table 1.
The main mechanisms of action of seaweed compounds in promoting cardiovascular health involve their antioxidant, anti-inflammatory, and metabolic modulation effects.
| Antioxidant Activity | Seaweed polyphenols, especially florotannins and fucoxanthin, exhibit significant antioxidant properties that help inhibit LDL oxidation—a crucial step in the development of atherosclerosis. This activity reduces the formation of foam cells, which contribute to arterial plaque formation [107,111]. |
| Anti-inflammatory Effects | Algal polysaccharides like fucoidan and laminarin demonstrate anti-inflammatory properties that can help lower vascular inflammation. This reduction in inflammation is essential for maintaining CV health and preventing atherosclerosis [111,112,113] |
| Lipid Metabolism Regulation | Fucoidan has been shown to improve lipid profiles by down-regulating genes involved in lipid synthesis (like SREBP1, ACC, and FAS) and up-regulating genes involved in lipid uptake (such as LDL-R). This modulation leads to reduced TC, LDL-C, and TG [112,113] |
| Improvement of Endothelial Function | Seaweed extracts have been linked to improved endothelial function, which is critical for vascular health [111]. |
| Impact on Glucose Metabolism | Compounds like oligosaccharides from Enteromorpha prolifera have demonstrated anti-diabetic effects, by enhancing glucose tolerance and reducing blood glucose levels through mechanisms involving the AKT pathway and the inhibition of gluconeogenesis [117,118]. |
| Gut Microbiota Modulation | Some seaweed compounds can alter gut microbiota composition, promoting beneficial bacteria and pathways associated with metabolic health, which may contribute to their anti-obesity effects [115]. |
| Clinical Evidence | Clinical studies, including those on Chlorella and Gdue (a blend of Ascophyllum nodosum and Fucus vesiculosus), support the efficacy of these compounds in reducing CVDs risk factors such as fasting blood glucose, LDL-C, and overall metabolic syndrome markers [134,135,136]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



