
Submitted:
29 October 2024
Posted:
30 October 2024
You are already at the latest version
Abstract
Atherosclerosis, a major risk factor for cardiovascular diseases is influenced by modifiable factors such as adiposity and blood cholesterol. Diet is crucial in these areas, particularly regarding antioxidant, inflammatory, and obesity effects. Coffee, a globally popular stimulant beverage has garnered significant attention for its potential impact on cardiovascular diseases. Recent insights reinforce the need to re-examine the relationship between coffee consumption and atherosclerosis progression. Coffee’s complex composition includes polyphenols, renowned for their antioxidant and anti-inflammatory properties, as well as potential weight-reducing effects. In addition, studies have demonstrated that certain coffee compounds such as chlorogenic acid, caffeic, p-coumaric, and ferulic acid can prevent atherogenesis by preventing oxidation of low-density lipoproteins. Conversely, diterpenes, found in some coffee brews, can elevate cholesterol levels posing risk to coronary health. Notably, coffee intake has been shown to influence gut microbiota diversity, potentially contributing to anti-obesity effects. This review explores the insights from preclinical and clinical studies investigating the potential mechanisms through which coffee consumption may reduce the risk of atherosclerosis. Highlighting the potential benefits of moderate filtered-coffee consumption, as well as the potential risks associated with excessive coffee consumption. Understanding this relationship is crucial for informing public health recommendations and guiding future research.
Keywords:
Cardiovascular disease
; Atherosclerosis
; Obesity
; coffee consumption
; Cholesterol
1. Introduction
Coffee is one of the most popular beverages consumed globally. According to the International Coffee Organization [1], annual per capita consumption in the top coffee consuming countries ranges from 15.4 to 22.3 kilograms. Coffee contains a complex mixture of compounds with a myriad of bioactive molecules. In addition, coffee brewing method also influences the beverage composition having effects mainly in lipid profile [2]. Coffee contains phytochemicals with well-recognized antioxidant properties, as evidenced in both animal models [3] and humans [4]. Extensive research has explored the antioxidant properties of coffee [5]. Additionally, coffee intake has not been linked to the development of coronary or carotid atherosclerosis [6,7]. This review delves into the potential mechanisms through which coffee compounds may influence atherosclerosis progression, considering associated factors such as obesity and lipid profile. By examining the diverse bioactive molecules in coffee and their potential effects on cardiovascular health, this review aims to contribute to a better understanding of the role of coffee consumption in preventing atherosclerosis and related risk factors.
2. Coffee Composition
The Coffee has a complex composition containing a diverse array of biologically active compounds. The specific composition can vary depending on the type of coffee bean, such as Coffea arabica, Coffea canephora, Coffea liberica. Key components include phenolic acids (chlorogenic acids, cafestol, and kahweol), alkaloids (caffeine and trigonelline), methylxanthines (caffeine, theobromine, and theophylline), and nicotinic acid (vitamin B3) [8]. Chlorogenic acid stands out as a particularly potent antioxidant and is also found in other foods, for instance wine, tea, apples, mushrooms etc. [9]. The biological properties of coffee are primarily attributed to key compounds such as caffeine, cafestol, kahweol, ferulic acid, chlorogenic acid, and trigonelline.
3. Bioactive Compounds in Coffee and Its Potential to Prevent Atherosclerosis
3.1. The Impact of Coffee Consumption on Obesity
Obesity is a well-established risk factor for cardiovascular disease. Adipose tissue expansion during obesity contribute to adipocyte dysfunction, chronic low-grade inflammation, raise of cholesterol levels, oxidative stress, and endothelial dysfunction [10]. A few studies have investigated the potential effects of coffee intake on adiposity. Peroxisome proliferator-activated receptors (PPARs) are transcription factors activated by lipid ligands and are critical in energy homeostasis and metabolism [11]. A recent study demonstrated that caffeine and chlorogenic acid (CGA), when loaded into solid lipid nanoparticles, can decrease the expression of PPAR-γ and CCAAT/enhancer binding protein alpha (c/EBP-α), adipogenic biomarkers, in a 3T3-F422A preadipocyte cell line. This suggests a potential reversible effect on adipogenesis by coffee bioactives [12]. Additionally, mice fed with a combination of CGA and caffeine (0.2%/0.03%) for 24 weeks exhibited decreased intraperitoneal adipose tissue, body weight, serum/hepatic of total cholesterol, triacylglycerol (TAG), and leptin levels. This response was associated with increased AMP-activated protein kinase (AMPK) expression and decreased PPAR-γ2 expression in the liver [13]. AMPK may promote lipolysis since and PPAR-γ2 is a physiological sensor of lipid levels. In this respect, the effect of coffee in the regulation of lipid metabolism reducing the size and number of adipocytes has been widely described in vitro [14,15,16]. Furthermore, a randomization control trial found that 500 mg/day of green coffee bean extract supplementation reduced body weight and improved lipid profiles in patients [17]. Moreover, a clinical trial in overweight/obese patients with type 2 diabetes demonstrated the protective effects of 800 mg/day of green coffee extract for 10 weeks, leading to reductions in body weight, body mass index, systolic blood pressure, C-reactive protein (CRP), triglycerides, and higher HDL levels [18]. Coffee intake can influence adiposity, in part by stimulating the sympathetic nervous system, which can increase energy expenditure and brown adipose tissue activity, leading to weight loss [19,20]. While moderate coffee consumption can contribute to weight management, excessive intake may have negative consequences, as demonstrated by a 2-year follow up study, that found a correlation between high coffee consumption and increased body adiposity in individuals with kidney transplant [21]. A three-year follow-up study in elderly participants with metabolic syndrome demonstrated that moderate, but not high, coffee consumption was associated with a reduction in total fat tissue, trunk fat, and visceral adipose tissue. However, heavy coffee intake was linked to unhealthier lifestyle habits among consumers, which could potentially counteract the beneficial effects of coffee on weight loss [22]. These findings suggest that the dosage of coffee consumption can determine its impact on weight loss.
3.1.1. Coffee Consumption and Gut Microbiota in Weigh Loss
Coffee metabolism occurs in three phases: early absorption within the first 1-2 hours in the stomach and small intestine, intermediate absorption between 4-8 hours, and late absorption phase in the large intestine after 8 hours. This late absorption phase is particularly influenced by gut microbiota [23]. In the colon, coffee is fermented by gut microbiota, which can regulate the bioavailability and biological activity of coffee polyphenols [24]. Coffee compounds, including CGA, and caffeine, can exert beneficial effects on the intestinal microbiota, potentially leading to weight loss. A study showed that coffee consumption can have to have positive effects in high-fat fed rats, including decreased body weight, adiposity, and improved gut microbiota diversity [24]. In another study using C57BL/6 male mice fed a high-fat diet, a 150 mg/Kg CGA solution for 20 weeks led to a loss of body weight and prevented subcutaneous and visceral weight gain, through its regulation of gut microbiota [25]. Of note, gut microbiota diversity has been well-established as a significant factor in atherosclerosis progression [26]. While green coffee extract intake did not directly decrease atherosclerotic lesions or serum lipids in ApoE-/- mice fed a high-fat diet for 14 weeks, it did result in reduced adiposity, weight gain, and improved gut miocrobiota diversity [27]. Furthermore, in an animal model of high-fat diet using Wistar rats, a freeze-dried coffee solution was found to increase the population of Bifidobacterium and improve HDL-C reverse cholesterol transport. However, despite these positive effects, it did not prevent weight gain in the experimental rats [28]. These findings suggest that coffee consumption, particularly through the action of chlorogenic acid and caffeine, can be beneficial for weight loss, potentially due to its impact on gut microbiota diversity. This highlights the importance of gut microbiota in mediating the cardiovascular effects of dietary compounds like coffee.
3.2. Anti-Atherogenic Effects of Bioactive Compounds of Coffee
Oxidation of low-density lipoprotein (ox-LDL) is a pivotal step in atherogenesis, as outlined by the oxidation theory of atherosclerosis. While ox-LDL is a key factor, other components, such as lysophosphatidylcholine (lysoPC) and oxisterols, also contribute to atherosclerosis progression. LysoPC, a breakdown product of phosphatidylcholine, is a component of ox-LDL. It plays a significant role in endothelial dysfunction and cardiovascular disease [29,30]. Oxylipins, derived from oxidized polyunsaturated fatty acids, and oxysterols, such as 7-ketocholesterol (7-KC), are also implicated in atherosclerosis due to their roles in inflammation and vascular function [31,32]. As previously mentioned, coffee is a rich source of antioxidants, including alkaloids, flavonoids, and phenolic compounds. CGA and its metabolites, such as ferulic, isoferulic, and vanillic acids, have demonstrated potent antioxidant activity [33,34]. Given that oxidized LDL (ox-LDL) is a contributing factor for the development of atherosclerotic plaques, the antioxidant properties of coffee compounds, including phenolic acids, have been shown to protect LDL from oxidation both in vitro and in human studies.
An in vitro study demonstrated that coffee compounds, including caffeic acid, 1-methyluric acid, and 1,3,7-trimethyluric acid, can prevent LDL oxidation [33]. Additionally, in vitro and ex vivo data showed that acute coffee consumption containing 420 mg of CGA was able to increase antioxidant capacity in plasma samples from healthy volunteers and delay LDL oxidation [23]. A study of healthy male students aged 20 to 31 that consuming coffee for 7 days led to a significant decrease in total cholesterol, LDL-C levels, lipid peroxidation markers, and a significant reduction in LDL susceptibility to oxidation [35]. These findings were further supported by another study on samples from 10 healthy volunteers, which showed that consuming 200 mL of filtered coffee increased the resistance of LDL to oxidation, likely due to the incorporation of coffee’s phenolic compounds, such as caffeic, p-coumaric, and ferulic acids, into LDL [36]. A larger study of 169 individuals found that coffee consumption can decrease lysoPC levels in plasma, primarily mediated by coffee polyphenols [37]. Moreover, consuming 4 to 8 cups of paper-filtered coffee per day was found to reduce lysoPC levels in 47 habitual coffee drinkers [38]. Furthermore, it has been suggested that coffee polyphenols can be transported in the plasma bound to LDL, potentially carrying antioxidants to the arterial intima and endothelial cells to protect against cardiovascular diseases [39]. Another study found that consuming 482 ± 61 mL/day of medium light roast or medium roast paper-filtered for 4 weeks increased antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx), however did not significantly reduce ox-LDL levels in healthy volunteers, suggesting that individual responses to coffee consumption may vary [40]. Additionally, coffee polyphenols, such as CGA, are metabolized by gut microbiota into smaller phenolic acids like dihydroferulic acid (DHFA). These metabolites can be detected in plasma at concentrations up to 1μM and have been shown to exhibit anti-inflammatory and antioxidant properties, potentially mitigating atherogenic progression [41]. A study on 74 subjects consuming two types of filtered coffee (high and low CGA content) for 8 weeks found that coffee with high CGA content decreased oxylipins levels, lipid peroxidation markers, and inflammatory markers in plasma. Moreover, in vitro experiments using THP-1 monocyte-derived macrophages exposed to 25 μg/mL ox-LDL further demonstrated that treatment with 1μM DHFA led to a decline in ox-LDL uptake, ox-LDL receptors cluster of differentiation 36 (CD36), scavenger receptor class A (SR-A), lectin-like oxidized LDL receptor-1 (LOX-1) expression, ROS production, and oxylipin levels [41]. In a more recent study, DHFA demonstrated its ability to reduce atherogenic progression in cultured human macrophages exposed to inflammatory and oxidative stress conditions (50 μg/mL ox-LDL, 10 ng/mL LPS, or 20 μM 7KC). DHFA decreased reactive oxygen species (ROS), 8-isoprostane (8-IsoP), ox-LDL uptake, CD36 expression, and inflammatory mediators, while simultaneously increasing IL-10, and prostaglandin PGE1, a potent vasodilator [42]. These findings further support the promising role of coffee’s bioactive compounds in preventing oxidation of LDL, inflammation, lysoPC, mitigating the progression of atherosclerosis. Moreover, the effects of coffee-related compounds on ox-LDL may vary depending on factors such as intervention time, coffee concentration, the content of CGA. A detailed overview of the effects of coffee and its bioactive compounds is presented in Table 1.
3.3. Coffee Consumption on Lipid Metabolism and Inflammation
A clinical trial carried out in habitual coffee drinkers demonstrated that increasing coffee doses had favorable effects on certain markers of subclinical inflammation (IL-18 and 8-isoprostane) without adversely affecting proatherogenic lipids [43]. Another clinical study found that moderate coffee consumption (3 cups per day for 8 weeks) of a soluble mixture of green and roasted coffee provided benefits for both healthy and hypercholesterolemic subjects. Coffee intake decreased total cholesterol, LDL-cholesterol, VLDL-cholesterol, and triglycerides, along with improved antioxidant capacity, decreased MDA, carbonylated groups, and decreasing C-reactive protein (CRP), which is associated with chronic inflammatory conditions [4]. Experimental data in INS-1 cells exposed to 3mM streptozotocin and preincubated with kahweol (2.5 and 5 μM) showed increase of antioxidant enzymes and anti-inflammatory effects by downregulating NF-κB [44]. Indeed, kahweol has been shown to activate Nrf2 signaling, which is associated with antioxidant and anti-inflammatory responses [45]. In vitro studies have demonstrated anti-inflammatory properties of caffeine, including its ability to suppress NF-κB activation in RAW264.7 cells stimulated with LPS [46]. Additionally, caffeine, at various doses (0.019 mM, 0.102 mM, and 1.16 mM), has been shown to exert anti-inflammatory effects in peripheral blood mononuclear cells by downregulating the expression of pro-inflammatory genes such as signal transducer and activator of transcription 1 (STAT1), tumor necrosis factor (TNF), interferon gamma (IFNG), and PPARG, as well as cytokines including IL-8, IL-4, IL-10, and TNF-α [47]. Treatment of RAW 264.7 cells with coffee pulp extract, CGA, and caffeine, in the presence of LPS, reduced NFκB activation and the expression of inflammatory markers such as TNF-α, IL-6, iNOS, COX-2, and PGE2 [48]. In a mouse model of high-fat diet-induced obesity, coffee intake (2% freeze-dried coffee or 2% green coffee extract) for 9 weeks, decreased body weight gain and reduced the expression of inflammatory markers, including activating transcription factor 3 (Atf3), Fos, and suppressor of cytokine signaling (Socs3) [49]. Furthermore, a study in C57BL/6 mice fed a high-fat diet found that consuming instant organic coffee (instant organic coffee 0.1% v/v) for 4 weeks prevented glucose intolerance, hypertrophy, and reduced macrophage infiltration, IL-6, and TNF-α levels, suggesting a decrease in adipose tissue inflammation [20]. Coffee and its constituents have been shown to exhibit anti-inflammatory effects also in human subjects. A study in subjects with and without coronary artery disease showed that acute caffeine ingestion (200 mg), decreased CRP in plasma and improved brachial endothelial function [50]. Two large cohorts of health professionals, with a follow up between 9 to 14 years, demonstrated that regular coffee consumption, both caffeinated and decaffeinated was associated with lower levels of inflammatory mediators such as CRP, leptin, and IL-6. These findings suggest that coffee compounds beyond caffeine may be responsible for anti-inflammatory effects [51]. Long-term filtered caffeinated and decaffeinated coffee consumption in healthy and type 2 diabetes women, has been demonstrated to diminish inflammation, as indicated by lower levels of CRP, and prevent endothelial dysfunction, as assessed by a decreased of E-selectin [52]. In a placebo-controlled trial in cyclists, high-CGA coffee consumption (prepared using the Turkish method) for 2 weeks increased antioxidant capacity but did not decrease post-exercise inflammation [53]. This may be due to CGA’s primary role as an antioxidant rather than anti-inflammatory agent.
Coffee has shown to have beneficial effects on inflammation and cardiovascular health; however, the influence of inter-individual difference should be considered, including genetic variations and lifestyle habits. A study showed that coffee consumption triggered an anti-inflammatory response in individuals with the adenosine A2A receptor (ADORA2A) TT genotype after resistance exercise or in physically active subjects [54]. Another study examining peripheral blood mononuclear cells from healthy subjects before and after coffee consumption found that inflammatory markers exhibited a differential response among individuals, concluding that caffeine may be pro-inflammatory in some individuals [55]. Indicating that the optimal dosing regimen influences the coffee-associated effects, yet the individual response pattern is also affected by genetic variations. Genetic polymorphisms in metabolic enzymes, such as cytochrome P4501A2 [56], adenosine receptors [57], and transcription factor Nrf2 [58], can account for individual variations in caffeine metabolism and response. Additionally, lifestyle factors play a significant role. A cross-sectional study found that while habitual caffeine intake in healthy subjects could decrease CRP in plasma, also the physical activity and sedentarism impact the inflammatory status [59]. These factors can interfere with the anti-inflammatory effects associated with coffee consumption.
3.4. Coffee Extraction Method Influence the Lipid Profile
Coffee consumption has been linked to various health benefits effects in the context of cardiovascular health. Some studies have shown that coffee may contribute to negative effects on serum lipids, mediated mainly by the lipid-soluble diterpenes, cafestol and kahweol [60]. However, the specific outcomes can vary depending on the coffee’s composition and the extraction method used [61,62]. Although diterpenes are intrinsic compounds of coffee lipid fraction, it is important to highlight that coffee extraction has a certain complexity and not only determines the coffee flavor profile, but also physicochemical characteristics related to biologically active substances aside of caffeine, such as lipid-soluble compounds [61]. While coffee polyphenols offer protective effects against cardiovascular diseases and associated risk factors, coffee oils can have detrimental effects by raising serum lipid levels. This raises concerns about the potential impact of coffee intake on cardiovascular health. For instance, coffee brewed using a French press typically has a low content of polyphenols and high content of kahweol and cafestol, both found in the lipid fraction, while filtered coffee has the opposite profile [62]. Regular consumption of Turkish coffee, which is high in cafestol and kahweol, has been shown to increase cholesterol, triglycerides, and LDL-C levels, suggesting a higher susceptibility for cardiovascular diseases [63]. This is primarily due to a lower bile acid excretion and neutral sterols [64]. Specifically, boiled and unfiltered coffee brew contains higher levels of cafestol and kahweol, which can contribute to elevated cholesterol levels in serum [63,65]. A study demonstrated that consumption of unfiltered coffee (Turkish method) increased total cholesterol to HDL ratio in women with vitamin D deficiency, potentially increasing their risk of hyperlipidemia [66]. This supports a susceptibility of hyperlipidemic subjects to enhance serum lipids by drinking boiled and unfiltered coffee. Conversely, paper-filtering coffee can effectively remove most of coffee oils, preventing the cholesterol raising effect, enabling the anti-atherogenic effects mediated mainly by coffee phenolic compounds [65]. For instance, in traditionally prepared coffee in Singapore and India the content of diterpenes is negligible (0.09 mg/cup), which did not increase serum cholesterol and triglycerides levels, whereas higher content of diterpenes (around 4.43 mg/cup) were found in unfiltered coffee from Indonesia, Scandinavian boiled, Turkish, and French press, associated with an increase in serum cholesterol and triglycerides [67]. A study, without specifying the preparation method, demonstrated that consuming at least 5 cups of plain black coffee per week, increased HDL-C levels, which is crucial for cardiometabolic health by its antiatherogenic potential [68]. Not only the diterpene content in coffee brew could be a concern, but the dosage consumed is also a crucial factor. A study examining the effects of filtered-coffee consumption on lipid profiles found that heavy consumption increased total cholesterol, triglycerides, and VLDL-C. In contrast, moderate consumption did not significantly alter lipid profiles [69]. Moreover, a study by Svatun et al [70] found that consuming 3 to 5 cups of espresso per day was associated with higher serum cholesterol levels in both men and women. Similarly, consuming more than 6 cups of boiled/plunger coffee per day also led to increased cholesterol levels in both genders. However, high intake of filtered coffee (more than 6 cups per day) only elevated serum cholesterol in women [70]. This suggests that diterpene content by the brewing method, the amount of coffee consumed, and sex differences determine the effects of coffee consumption on cardiometabolic health (Figure 1).
Figure 1.
Coffee extraction methods significantly influence the physicochemical characteristics of the final brew, particularly the content of cafestol and kahweol. These compounds can directly affect lipid profiles in coffee consumers, which is crucial for cardiovascular health.
Figure 1.
Coffee extraction methods significantly influence the physicochemical characteristics of the final brew, particularly the content of cafestol and kahweol. These compounds can directly affect lipid profiles in coffee consumers, which is crucial for cardiovascular health.
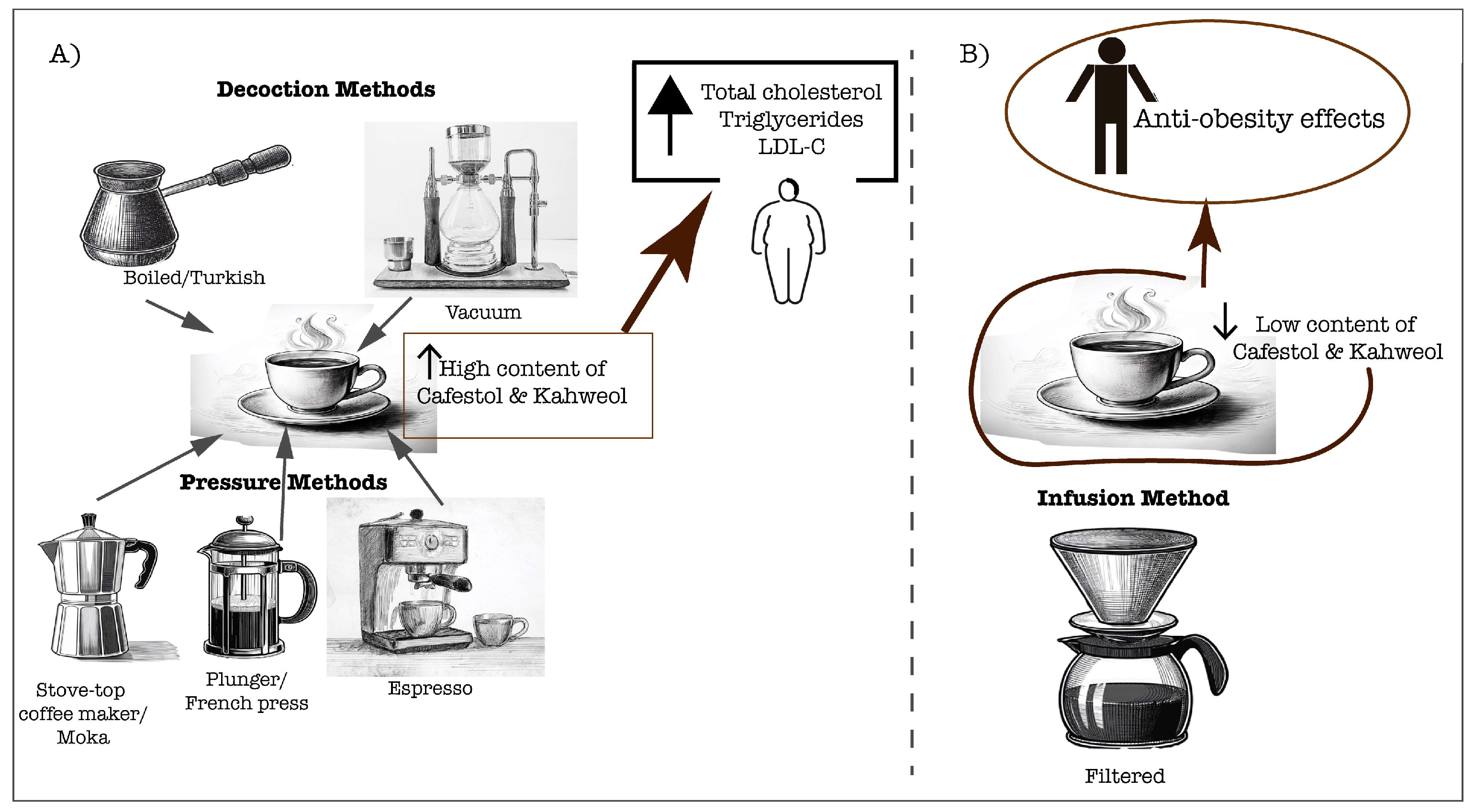

Table 1.
Evidence of the impact of coffee and coffee bioactive on adiposity, gut microbiota, antioxidant enzymes, inflammation, and lipid profile.
Table 1.
Evidence of the impact of coffee and coffee bioactive on adiposity, gut microbiota, antioxidant enzymes, inflammation, and lipid profile.
| Coffee/Coffee Bioactive | Study Model | Main findings | Study details | References |
|---|---|---|---|---|
|
Caffeine and CGA⇧ |
3T3-F422A preadipocyte cell line |
⇩ PPAR-γ expression ⇩c/EBP-α |
Caffeine 1 mM + CGA 0.5 mM loaded into solid lipid nanoparticles |
Uner and Celebi, 2023 [12] |
|
Caffeine and CGA |
Female ICR mice |
⇩ Adipose tissue ⇩ Body weight ⇩Total cholesterol (serum and hepatic) ⇩Triglycerides ⇩ Leptin levels ⇧ AMPK activation ⇩ PPAR-γ2 liver expression |
Diet containing: 0.2% CGA 0.03% caffeine For 24 weeks |
Zheng et al., 2014 [13] |
|
Green coffee bean extract |
Individuals, over the age of 18, (n=103) |
⇩ Body weight Lipid profile improvement |
500 mg/day green coffee extract Supplementation for at least 1 week to 8 weeks |
Kanchanasurakit et al., 2023 [17] |
|
Green coffee extract |
Overweight/ obese patients with type 2 diabetes (n=44) |
⇩ Body weight ⇩ Body mass index ⇩ Systolic blood pressure ⇩ C-reactive protein ⇩ Triglycerides ⇧ HDL levels |
800 mg/day green coffee extracct supplementation for 10 weeks |
Khalili-Moghadam et al., 2023 [18] |
|
Coffee |
Kidney transplant recipients aged 49.5 years (n=170) |
⇧ Body adiposity (central adiposity) Lower muscle quality |
Median coffee consumption 200 mL/day 2 years-follow up |
Costa et al., 2023 [21] |
|
Coffee |
Individuals with metabolic syndrome (n= 1,483) |
⇩ Total fat tissue ⇩ Trunk fat ⇩ Visceral adipose tissue |
Moderate coffee consumption (1-7 cups/week) 3 years-follow up |
Henn et al., 2023 [22] |
|
Instant coffee |
High-fat fed rats (Male Sprague Dawley) |
⇩ Body weight ⇩ Adiposity ⇧ Insulin resistance ⇩Firmicutes (F)-to-Bacteroidetes ratio and Clostridium Cluster XI ⇧ Enterobacteria |
Instant caffeinated coffee (20 g/L) for 10 weeks |
Cowan et al., 2014 |
|
CGA |
C57BL/6 male mice fed a high fat diet |
⇩ Body weight ⇩ Subcutaneous and visceral weight ⇧ short chain fatty acid producers (Dubosiella, Romboutsia, Mucispirillum, and Faecalibaculum) ⇧ Akkermansia |
150 mg/Kg CGA solution for 20 weeks |
Ye et al., 2021 [25] |
|
Green coffee extract |
Apo-/- mice fed antiatherogenic diet |
⇩ Adiposity ⇩ Weigh gain ⇩ Inflammatory infiltrate in adipose tissue Improved microbiota diversity ⇧ Desulfovibrio ⇧ Mogibacteriaceae |
Green coffee extract 220 mg/Kg for 14 weeks |
Caro-Gómez et al., 2019 [27] |
|
Freeze-dried coffee solution |
Wistar rats fed high-fat diet |
⇧ Bifidobacterium spp. ⇧ HDL-C reverse cholesterol transport ⇩ II1b mRNA Did not improved weight gain |
Freeze-dried coffee solution at a dose of 0.39 g/100 g for 8 weeks |
Cavalcanti et al., 2022 [28] |
| Caffeic acid, 1-methyluric acid and 1,3,7-trimethyluric acid |
In vitro and ex vivo study on plasma from healthy indivi-duals |
Prevention of LDL oxidation by cooper |
0.5 μM caffeic acid, 3 μM 1,3,7-trimethyluric acid, 30 μM 1-methyluric acid, caffeic acid |
Gómez-Ruiz et al., 2007 [33] |
|
Acute coffee consumption (400 mg CGA) |
In vitro and ex vivo experiments on plasma from healthy volunteers (n=20) after drinking coffee |
⇧ Antioxidant capacity of plasma Prevention of LDL oxidation |
Acute coffee consumption containing 420 mg of CGA (400 mL of coffee |
Lara-Guzmán et al., 2016 [23] |
|
Coffee |
Healthy male volunteers aged 20 to 31 (n=11) | ⇩ Total cholesterol ⇩ LDL-C ⇩ MDA ⇩ LDL oxidation |
Coffee intake, 24 g total per day for 1 week |
Yukawa et al., 2004 [35] |
|
Filtered Coffee /caffeic acid |
Ex vivo and in vitro experiments in plasma from healthy volunteers (n=10) |
⇩ LDL oxidation Incorporation of caffeic, p-coumaric, and ferulic acids into LDL |
Coffee consumption (200 mL) In vitro: 1, 10, 100 nmol/L caffeic acid incubated with isolated LDL from healthy subjects |
Natella et al., 2007 [36] |
|
Coffee (high content of polyphenols) |
Healthy subjects aged 20 years or older (n= 169) |
⇩ plasma LysoPC levels |
Low coffee consumption (≤100 mL/day), high coffee consumption >100 mL/day) |
Miranda et al., 2017 [37] |
|
Filtered coffee |
Habitual coffee drinkers (n=47) |
⇩ plasma LysoPC levels |
First month: no coffee consumption Second month: 4 cups of paper-filtered coffee/day Third month: 8 cups of paper-filtered coffee/day |
Kuang et al., 2018 [38] |
|
Filtered coffee |
Healthy volunteers (n=20) |
⇧ SOD ⇧ Catalase ⇧ GPx Did not reduce ox-LDL levels |
482 ± 61 mL/day medium light roast or medium roast paper-filtered for 4 weeks |
Corrêa et al., 2012 [40] |
|
Filtered coffee with high content of CGA and low content of kahweol and cafestol/DHFA in in vitro experiments |
Subjects (n=74) aged between 20 and 60 years. In vitro experiments in THP-1 monocyte-derived macrophages |
⇩ Oxylipins levels in plasma ⇩ Lipid peroxidation markers ⇩ Inflammatory markers No significant differences on ox-LDL levels in plasma In vitro data: ⇩ Ox-LDL uptake ⇩ CD36 expression ⇩ SR-A expression ⇩ LOX-1 expression ⇩ ROS production ⇩ oxylipins profile |
Consumption of coffee A containing 787 mg CGA (n=24), coffee B containing 407 mg CGA (n=25), 400 mL/day for 8 weeks. In vitro experiment: 25 μg/mL ox-LDL, 1μM DHFA, and 1μM phenolic acid |
Lara-Guzmán et al., 2020 [41] |
|
DHFA |
Culture human macrophages |
⇩ ROS production ⇩ 8-Isoprostane ⇩ Ox-LDL uptake ⇩ CD36 expression ⇩ inflammatory mediators (TNF-α, IL-6, and IL-17) ⇧ IL-10 ⇧ PGE1 |
THP-1 monocyte-derived macrophages were exposed to 50 μg/mL oxLDL, 10 ng/mL LPS or 20 μM 7KC treated with 1 μM DHFA |
Lara-Guzmán et al., 2024 |
|
Filtered coffee |
Habitual coffee drinkers (n=47) younger than 65 years with elevated risk of type 2 diabetes |
⇩ IL-18 ⇩ 8-Isoprostane ⇧ adiponectin ⇧ Caffeine in serum ⇧ CGA in serum ⇧ Caffeic acid metabolites in serum ⇧HDL ⇩LDL/HDL ratio |
First month: no coffee Second month: 4 cups/day Third month: 8 cups/day |
Kempf et al., 2010 [43] |
|
Green and roasted coffee |
Normocholesterolemic (n=25) and hypercholesterolemic (n=27) subjects aged 18 to 45 years |
⇩ Total cholesterol ⇩ LDL-C ⇩ VLDL-C ⇩ Triglycerides ⇧ Plasma antioxidant capacity ⇩ MDA levels ⇩ Carbonylation ⇩ CRP |
Moderate coffee consumption (3 cups per day) for 8 weeks |
Martínez-López et al., 2019 [4] |
|
Kahweol |
INS-1 cells |
⇩ NF-κB ⇧ Antioxidant enzymes (Hemeoxygenase-1) ⇧ p-AKT ⇧ BCL-2 |
Cells were exposed to 3mM streptozotocin and pre-incubated with 2.5 and 5 μM Kahweol |
El-Huneidi et al., 2021 [44] |
|
Kahweol |
AREc32 cells |
⇧Nrf2 |
0.02 and 30 μM Kahweol |
Wu et al., 2014 [45] |
|
Caffeine |
RAW264.7 cells |
⇩ NF-κB ⇩ pho-p38MAPK |
Cells were exposed to 1 μg/mL LPS and treated with caffeine (0, 100, 400, 800, 1000, and 1200 μM) |
Hwang et al., 2016 [46] |
|
Caffeine |
Peripheral Blood Mononuclear Cells isolated from 3 healthy individuals |
⇩ STAT1 expression ⇩ TNF expression ⇩ IFNG expression ⇩ PPARG expression ⇩ IL-8, IL-4, IL10, and TNF-α levels |
Caffeine (0.019 mM, 0.102 mM, and 1.16 mM) |
Iris et al., 2018 [47] |
|
Coffee pulp extract/CGA/caffeine |
Raw 264.7 cells |
⇩ TNF-α, IL-6, iNOS, COX-2, and PGE2 expression ⇩NFκB activation ⇩ MAPK signaling |
Cells were stimulated with 1μg/mL LPS and treated with 1000 μg/mL coffee pulp extract, 13.38 μg/mL CGA, and 3.82 μg/mL caffeine |
Ontawong et al., 2023 [48] |
|
Coffee/Green coffee |
C57BL6 male mice |
⇩ Body weight ⇩ Mesenteric fat weight ⇩ Atf3, Fos, and Socs3 ⇩ Hsp70 |
High fat diet 2% freeze-dried caffeinated coffee, decaffeinated coffee, or green coffee for 9 weeks |
Jia et al., 2014 [49] |
|
Instant organic coffee |
C57BL6 male mice |
Improved glucose metabolism ⇩ Adipose tissue inflammation ⇩ Hypertrophy ⇩ Macrophage infiltration ⇩IL-6, TNF-α ⇧ Adapative thermogenesis ⇧ Mitochondrial biogenesis |
High-fat diet +consumption of instant organic coffee (0.1% v/v) for 4 weeks |
Martins et al., 2023 [20] |
|
Caffeine |
Subjects with (n=40) and without coronary artery disease (n=40) |
⇩ CRP in plasma Improvement of brachial endothelial function. |
200 mg Acute C caffeine ingestion |
Shechter et al., 2011 [50] |
|
Caffeinated and decaffeinated coffee |
N= 15,551 women (Nurse’s Health Study) and n= 7,397 men (Health Professionals) | ⇩ CRP ⇩Leptin ⇩ IL-6 ⇩C-peptide ⇩ Estrone, total estradiol, free estradiol ⇧Adiponectin |
Regular coffee consumption; Follow-up between 9 to 14 years |
Hang et al., 2019 [51] |
|
Filtered coffee |
Healthy women (n=730) and women with type 2 diabetes (n=663) aged 43-70 years |
⇩ CRP Prevent endothelial dysfunction ⇩ E-selectin |
Regular caffeinated and decaffeinated coffee consumption. Follow-up of 14 to 15 years |
Lopez-Garcia et al., 2006 [52] |
|
High-CGA coffee |
Cyclists subjects Men (n=10), women (n=5) aged 19 to 51 years |
⇧ Antioxidant capacity in plasma It did not decrease post-exercise inflammation |
High-CGA coffee consumption (300 ml/day) for 2 weeks. Coffee was prepared using the Turkish method. Participation in a 50-Km cycling time trial |
Nieman et al., 2018 [53] |
|
Caffeine/Coffee |
Resistance-trained Iranian men (n=15) around 21 years old. Russian healthy physically active subjects (n=134) aged 28 to 31 years. | ⇩Myeloperoxi-dase ⇩Acetylcholines-terase Association of ADORA2A gene polymorphism with anti-inflammatory effects of caffeine |
6 mg/Kg Acute caffeine consumption before resistance exercise. Regular coffee intake in the physically active subjects. |
Rahimi et al., 2023 [54] |
|
Coffee/caffeine |
Peripheral blood mononuclear cells isolated from 8 healthy individuals |
⇩ Inflammatory markers in some individuals ⇧ inflammatory markers in some individual |
Cells were isolated before and after coffee consumption (3 capsules of coffee containing 165 mg caffeine). Exposed to 1 μg/mL LPS and 5 μg/mL phytohaemagglutinin. Cells were treated with 200 ng/mL caffein in vitro |
Muqaku et al., 2016 [55] |
|
Caffeine |
Healthy subjects: men (n=112) and women (n=132) aged 18 to 55 years | ⇩ CRP in plasma ⇩ Body fatt total and visceral ⇧ Adiponectin ⇧ Il-10 ⇩ IL-6, TNF-α |
Habitual caffeine intake |
Rodas et al., 2020 [59] |
|
Coffee |
Individuals (n=109) aged 22 to 70 years |
⇧ Total cholesterol ⇧ Triglycerides ⇧ LDL-C ⇧VLDL-C |
Regular coffee consumption (Turkish method and instant coffee) |
Saad Al-Fawaeir et al., 2023 [63] |
|
Coffee |
Women with vitamin D deficiency (n=270) aged 18 to 65 years |
⇧ Total cholesterol/HDL ratio |
Turkish coffee consumption during 3 previous months. Moderate consumption (1-2 cups/day). High consumption (⩾ 3 cups/day). 150 mg caffeine per cup |
Habash et al., 2022 [66] |
|
Coffee |
Healthy volunteers (n=3000) |
Filtered coffee: ⇩ Serum cholesterol ⇩ Triglyceride Unfiltered coffee: ⇧ Serum cholesterol ⇧ Triglycerides |
Filtered and unfiltered coffee consumption (1-5 cups/day) |
Naidoo et al., 2011 [67] |
|
Coffee |
Healthy volunteers (n=1272) over the age of 30. |
⇧ HDL-C levels |
Regular plain black coffee consumption (5 cups per week). Fo-llow-up of 13 years |
Chang et al., 2010 [68] |
|
Filtered coffee |
ELSA-Brasil cohort (n=4732) |
⇧ Total cholesterol ⇧ Triglycerides ⇧ VLDL-C ⇧ Triglyceride-rich lipoprotein particles |
Regular high-consumption of filtered coffee (more than 3 cups/day) |
Miranda et al., 2022 [69] |
|
Coffee |
Tromø Study in Norther Norway (n=21083) aged 40 years |
⇧ Total cholesterol levels |
Espresso coffee 3 to 5 cups per day. Boiled/plunger coffee more than 6 cups per day |
Svatun et al., 2020 [70] |
4. Conclusions
In conclusion, habitual and moderate coffee consumption can offer potential benefits in preventing atherosclerosis progression. However, it is essential to consider the brewing method, as diterpenes in boiled and unfiltered coffee may elevate lipid levels, posing risks for cardiovascular diseases. Furthermore, individual responses to coffee consumption can vary based on factors such as dosage and genetic variations. While existing research provides valuable insights, further studies are needed to fully comprehend the complex interplay between coffee components and human health.
Author Contributions
Conceptualization, M.B.C.; writing—original draft preparation, M.B.C.; writing—review and editing, M.B.C. All authors have read and agreed to the published version of the manuscript.
Funding
Open access funding for this article was supported by Instituto Nacional de Cardiología Ignacio Chávez.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- International Coffee Organization [ICO]. Coffee Report and Outlook, December 2023 [PDF]. 2023. Retrieved from https://icocoffee.org/documents/cy2023-24/Coffee_Report_and_Outlook_December_2023_ICO.pdf.
- Farag, M.A.; Zayed, A.; Sallam, I.E.; Abdelwareth, A; Wessjohann, L.A. Metabolomics-Based Approach for Coffee Beverage Improvement in the Context of Processing, Brewing Methods, and Quality Attributes. Foods 2022, 11(6):864. [CrossRef]
- Higgins, L.G.; Cavin, C.; Itoh, K.; Yamamoto, M.; Hayes, J.D. Induction of cancer chemopreventive enzymes by coffee is mediated by transcription factor Nrf2. Evidence that the coffee-specific diterpenes cafestol and kahweol confer protection against acrolein. Toxicol Appl Pharmacol 2008, 226:328–337. [CrossRef]
- Martínez-López, S.; Sarriá, B.; Mateos, R.; Bravo-Clemente, L. Moderate consumption of a soluble green/roasted coffee rich in caffeoylquinic acids reduces cardiovascular risk markers: results from a randomized, cross-over, controlled trial in healthy and hypercholesterolemic subjects. Eur J Nutr 2019, 58(2):865-878. [CrossRef]
- Shanmugam, G. Polyphenols: potent protectors against chronic diseases. Nat Prod Res 2024:1-3. [CrossRef]
- Reis, J.P.; Loria, C.M.; Steffen, L.M.; Zhou, X.; van Horn, L.; Siscovick, D.S.; Jacobs, D.R. Jr.; Carr, J.J. Coffee, decaffeinated coffee, caffeine, and tea consumption in young adulthood and atherosclerosis later in life: the CARDIA study. Arterioscler Thromb Vasc Biol 2010, 30(10):2059-66. [CrossRef]
- Khalili-Moghadam, S.; Hedayati, M.; Golzarand, M.; Mirmiran, P. Effects of green coffee aqueous extract supplementation on glycemic indices, lipid profile, CRP, and malondialdehyde in patients with type 2 diabetes: a randomized, double-blind, placebo-controlled trial. Front Nutr 2023, 10:1241844. [CrossRef]
- de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Magalhães Júnior, A.I.; do Prado, F.G.; Pagnoncelli, M.G.B.; Karp, S.G.; Soccol, C.R. Chemical composition and health properties of coffee and coffee by-products. Adv Food Nutr Res 2020, 91:65-96. [CrossRef]
- Purushothaman, A.; Babu, S.S.; Naroth, S.; Janardanan, D. Antioxidant activity of caffeic acid: thermodynamic and kinetic aspects on the oxidative degradation pathway. Free Radic Res 2022, 56(9-10):617-630. [CrossRef]
- Engin, A. Endothelial Dysfunction in Obesity. Adv Exp Med Biol 2017, 960:345-379. [CrossRef]
- Wagner, N.; Wagner, K.D. The Role of PPARs in Disease. Cells 2020, 9(11):2367. [CrossRef]
- Uner, B.; Macit Celebi, M.S. Anti-obesity effects of chlorogenic acid and caffeine- lipid nanoparticles through PPAR-γ/C/EBP-ɑ pathways. Int J Obes (Lond) 2023, 47(11):1108-1119. [CrossRef]
- Zheng, G.; Qiu, Y.; Zhang, Q.F.; Li, D. Chlorogenic acid and caffeine in combination inhibit fat accumulation by regulating hepatic lipid metabolism-related enzymes in mice. Br J Nutr 2014, 112(6):1034-40. [CrossRef]
- Wang, Q.; Hu, G.; Lu, Q.; Hong, D.; Al-Romaima, A.; Qiu, M.; Xiong, W. Identification and screening of novel diterpenoids from roasted arabica coffee in the regulation of lipid content in white adipocytes. Food Funct 2023, 14(11):5138-5150. [CrossRef]
- Quarta, S.; Scoditti, E.; Carluccio, M.A.; Calabriso, N.; Santarpino, G.; Damiano, F.; Siculella, L.; Wabitsch, M.; Verri, T.; Favari, C.; Del Rio, D.; Mena, P.; De Caterina, R.; Massaro, M. Coffee Bioactive N-Methylpyridinium Attenuates Tumor Necrosis Factor (TNF)-α-Mediated Insulin Resistance and Inflammation in Human Adipocytes. Biomolecules 2021, 11(10):1545. [CrossRef]
- Duangjai, A.; Nuengchamnong, N.; Suphrom, N.; Trisat, K.; Limpeanchob, N.; Saokaewm S. Potential of Coffee Fruit Extract and Quinic Acid on Adipogenesis and Lipolysis in 3T3-L1 Adipocytes. Kobe J Med Sci 2018, 64(3):E84-E92.
- Kanchanasurakit, S.; Saokaew, S.; Phisalprapa, P.; Duangjai, A. Chlorogenic acid in green bean coffee on body weight: a systematic review and meta-analysis of randomized controlled trials. Syst Rev 2023, 12(1):163. [CrossRef]
- Khalili-Moghadam, S.; Hedayati, M.; Golzarand, M.; Mirmiran, P. Effects of green coffee aqueous extract supplementation on glycemic indices, lipid profile, CRP, and malondialdehyde in patients with type 2 diabetes: a randomized, double-blind, placebo-controlled trial. Front Nutr 2023, 10:1241844. [CrossRef]
- Van Schaik, L.; Kettle, C.; Green, R.; Irving, H.R.; Rathnerm J.A. Effects of Caffeine on Brown Adipose Tissue Thermogenesis and Metabolic Homeostasis: A Review. Front Neurosci 2021, 15:621356. [CrossRef]
- Martins, B.C.; Soares, A.C.; Martinsm F.F.; Resende, A.C.; Inada, K.O.P., Souza-Mello, V.; Nunes, N.M.; Daleprane, J.B. Coffee consumption prevents obesity-related comorbidities and attenuates brown adipose tissue whitening in high-fat diet-fed mice. J Nutr Biochem 2023, 117:109336. [CrossRef]
- Costa, M.S.D.; Pontes, K.S.D.S.; Guedes, M.R.; Barreto Silva, M.I.; Klein, M.R.S.T. Association of habitual coffee consumption with obesity, sarcopenia, bone mineral density and cardiovascular risk factors: A two-year follow-up study in kidney transplant recipients. Clin Nutr 2023, 42(10):1889-1900. [CrossRef]
- Henn, M.; Babio, N.; Romaguera, D.; Vázquez-Ruiz, Z.; Konieczna, J.; Vioque, J.; Torres-Collado, L.; Razquin, C.; Buil-Cosiales, P.; Fitó, M.; Schröder, H.; Hu, F.B.; Abete, I.; Zulet, M.Á.; Fernández-Villa, T.; Martín, V.; Estruch, R.; Vidal, J.; Paz-Graniel, I.; Martínez, J.A.; Salas-Salvadó, J.; Martínez-González, M.A.; Ruiz-Canela, M. Increase from low to moderate, but not high, caffeinated coffee consumption is associated with favorable changes in body fat. Clin Nutr 2023, 42(4):477-485. [CrossRef]
- Lara-Guzmán, O.J.; Álvarez-Quintero, R.; Osorio, E.; Naranjo-Cano, M, Muñoz-Durango K. GC/MS method to quantify bioavailable phenolic compounds and antioxidant capacity determination of plasma after acute coffee consumption in human volunteers. Food Res Int 2016, 89(Pt 1):219-226. [CrossRef]
- Cowan, T.E.; Palmnäs, M.S.; Yang, J.; Bomhof, M.R.; Ardell, K.L.; Reimer, R.A.; Vogel, H.J.; Shearer, J. Chronic coffee consumption in the diet-induced obese rat: impact on gut microbiota and serum metabolomics. J Nutr Biochem 2014, 25(4):489-95. [CrossRef]
- Ye, X.; Liu, Y.; Hu, J.; Gao, Y.; Ma, Y.; Wen, D. Chlorogenic Acid-Induced Gut Microbiota Improves Metabolic Endotoxemia. Front Endocrinol (Lausanne) 2021, 12:762691. [CrossRef]
- Jonsson, A.L.; Bäckhed, F. Role of gut microbiota in atherosclerosis. Nat Rev Cardiol 2017, 14(2):79-87. [CrossRef]
- Caro-Gómez, E.; Sierra, J.A.; Escobar, J.S.; Álvarez-Quintero, R.; Naranjo, M.; Medina, S.; Velásquez-Mejía, E.P.; Tabares-Guevara, J.H.; Jaramillo, J.C.; León-Varela, Y.M.; Muñoz-Durango, K.; Ramírez-Pineda, J.R. Green Coffee Extract Improves Cardiometabolic Parameters and Modulates Gut Microbiota in High-Fat-Diet-Fed ApoE-/- Mice. Nutrients 2019, 11(3):497. [CrossRef]
- Cavalcanti, M.H.; Roseira, J.P.S.; Leandro, E.D.S.; Arruda, S.F. Effect of a freeze-dried coffee solution in a high-fat diet-induced obesity model in rats: Impact on inflammatory response, lipid profile, and gut microbiota. PLoS One 2022, 17(1):e0262270. [CrossRef]
- Paapstel, K.; Kals, J.; Eha, J.; Tootsi, K.; Ottas, A.; Piir, A.; Jakobson, M.; Lieberg, J.; Zilmer, M. Inverse relations of serum phosphatidylcholines and lysophosphatidylcholines with vascular damage and heart rate in patients with atherosclerosis. Nutr Metab Cardiovasc Dis 2018, 28(1):44-52. [CrossRef]
- Stegemann, C.; Pechlaner, R.; Willeit. P.; Langley, S.R.; Mangino, M.; Mayr, U.; Menni, C.; Moayyeri, A.; Santer, P.; Rungger, G.; Spector, T.D.; Willeit, J.; Kiechl, S.; Mayr, M. Lipidomics profiling and risk of cardiovascular disease in the prospective population-based Bruneck study. Circulation 2014, 129(18):1821-31. [CrossRef]
- Bojic, L.A.; McLaren, D.G.; Harms, A.C.; Hankemeier, T.; Dane, A.; Wang, S.P.; Rosa, R.; Previs, S.F.; Johns, D.G.; Castro-Perez, J.M. Quantitative profiling of oxylipins in plasma and atherosclerotic plaques of hypercholesterolemic rabbits. Anal Bioanal Chem 2016, 408(1):97-105. [CrossRef]
- Zmysłowski, A.; Szterk, A. Current knowledge on the mechanism of atherosclerosis and pro-atherosclerotic properties of oxysterols. Lipids Health Dis 2017, 16(1):188. [CrossRef]
- Gómez-Ruiz, J.A.; Leake, D.S.; Ames, J.M. In vitro antioxidant activity of coffee compounds and their metabolites. J Agric Food Chem 2007, 55(17):6962-9. Erratum in: J Agric Food Chem. 2007;55(20):8284. [CrossRef]
- Alnsour, L.; Issa, R.; Awwad, S.; Albals, D.; Al-Momani, I. Quantification of Total Phenols and Antioxidants in Coffee Samples of Different Origins and Evaluation of the Effect of Degree of Roasting on Their Levels. Molecules 2022, 27(5):1591. [CrossRef]
- Yukawa, G.S.; Mune, M.; Otani, H.; Tone, Y.; Liang, X.M.; Iwahashi, H.; Sakamoto, W. Effects of coffee consumption on oxidative susceptibility of low-density lipoproteins and serum lipid levels in humans. Biochemistry (Mosc) 2004, 69(1):70-4. [CrossRef]
- Natella, F.; Nardini, M.; Belelli, F.; Scaccini, C. Coffee drinking induces incorporation of phenolic acids into LDL and increases the resistance of LDL to ex vivo oxidation in humans. Am J Clin Nutr 2007, 86(3):604-9. [CrossRef]
- Miranda, A.M.; Carioca, A.A.F.; Steluti, J.; da Silva, I.D.C.G.; Fisberg, R.M.; Marchioni, D.M. The effect of coffee intake on lysophosphatidylcholines: A targeted metabolomic approach. Clin Nutr 2017, 36(6):1635-1641. [CrossRef]
- Kuang, A.; Erlund, I.; Herder, C.; Westerhuis, J.A.; Tuomilehto, J. Cornelis, M.C. Lipidomic Response to Coffee Consumption. Nutrients. 2018;10(12):1851. [CrossRef]
- Tung, W.C.; Rizzo, B.; Dabbagh, Y.; Saraswat, S.; Romanczyk, M.; Codorniu-Hernández, E.; Rebollido-Rios, R.; Needs, P.W.; Kroon, P.A.; Rakotomanomana, N.; Dangles, O.; Weikel, K.; Vinson, J. Polyphenols bind to low density lipoprotein at biologically relevant concentrations that are protective for heart disease. Arch Biochem Biophys 2020, 694:108589. [CrossRef]
- Corrêa, T.A.; Monteiro, M.P.; Mendes, T.M.; Oliveira, D.M.; Rogero, M.M.; Benites, C.I.; Vinagre, C.G.; Mioto, B.M.; Tarasoutchi, D.; Tuda, V.L.; César, L.A.; Torres, E.A. Medium light and medium roast paper-filtered coffee increased antioxidant capacity in healthy volunteers: results of a randomized trial. Plant Foods Hum Nutr 2012, 67(3):277-82. [CrossRef]
- Lara-Guzmán, O.J.; Medina, S.; Álvarez, R.; Oger, C.; Durand, T.; Galano, J.M.; Zuluaga, N.; Gil-Izquierdo, Á.; Muñoz-Durango, K. Oxylipin regulation by phenolic compounds from coffee beverage: Positive outcomes from a randomized controlled trial in healthy adults and macrophage derived foam cells. Free Radic Biol Med 2020, 160:604-617. [CrossRef]
- Lara-Guzmán, O.J.; Arango-González, Á.; Rivera, D.A.; Muñoz-Durango, K.; Sierra, J.A. The colonic polyphenol catabolite dihydroferulic acid (DHFA) regulates macrophages activated by oxidized LDL, 7-ketocholesterol, and LPS switching from pro- to anti-inflammatory mediators. Food Funct 2024, 15(20):10399-10413. [CrossRef]
- Kempf, K.; Herder, C.; Erlund, I.; Kolb, H.; Martin, S.; Carstensen, M.; Koenig, W.; Sundvall, J.; Bidel, S.; Kuha, S.; Tuomilehto, J. Effects of coffee consumption on subclinical inflammation and other risk factors for type 2 diabetes: a clinical trial. Am J Clin Nutr 2010, 91(4):950-7. [CrossRef]
- El-Huneidi, W.; Anjum, S.; Bajbouj, K.; Abu-Gharbieh, E.; Taneera, J. The Coffee Diterpene, Kahweol, Ameliorates Pancreatic β-Cell Function in Streptozotocin (STZ)-Treated Rat INS-1 Cells through NF-kB and p-AKT/Bcl-2 Pathways. Molecules 2021, 26(17):5167. [CrossRef]
- Wu, K.C.; McDonald, P.R.; Liu, J.; Klaassen, C.D. Screening of natural compounds as activators of the keap1-nrf2 pathway. Planta Med 2014, 80(1):97-104. [CrossRef]
- Hwang, J.H.; Kim, K.J.; Ryu, S.J.; Lee, B.Y. Caffeine prevents LPS-induced inflammatory responses in RAW264.7 cells and zebrafish. Chem Biol Interact 2016, 248:1-7. [CrossRef]
- Iris, M.; Tsou, P.S.; Sawalha, A.H. Caffeine inhibits STAT1 signaling and downregulates inflammatory pathways involved in autoimmunity. Clin Immunol 2018, 192:68-77. [CrossRef]
- Ontawong, A.; Duangjai, A.; Vaddhanaphuti, C.S.; Amornlerdpison, D.; Pengnet, S.; Kamkaew, N. Chlorogenic acid rich in coffee pulp extract suppresses inflammatory status by inhibiting the p38, MAPK, and NF-κB pathways. Heliyon 2023, 9(3):e13917. [CrossRef]
- Jia, H.; Aw, W.; Egashira, K.; Takahashi, S.; Aoyama, S.; Saito, K.; Kishimoto, Y.; Kato, H. Coffee intake mitigated inflammation and obesity-induced insulin resistance in skeletal muscle of high-fat diet-induced obese mice. Genes Nutr 2014, 9(3):389. [CrossRef]
- Shechter, M.; Shalmon, G.; Scheinowitz, M.; Koren-Morag, N.; Feinberg, M.S.; Harats, D.; Sela, B.A.; Sharabi, Y.; Chouraqui, P. Impact of acute caffeine ingestion on endothelial function in subjects with and without coronary artery disease. Am J Cardiol 2011, 107(9):1255-61. [CrossRef]
- Hang, D.; Kværner, A.S.; Ma, W.; Hu, Y.; Tabung, F.K.; Nan, H.; Hu, Z.; Shen, H.; Mucci, L.A.; Chan, A.T.; Giovannucci, E.L.; Song, M. Coffee consumption and plasma biomarkers of metabolic and inflammatory pathways in US health professionals. Am J Clin Nutr 2019,109(3):635-647. [CrossRef]
- Lopez-Garcia, E.; van Dam, R.M.; Qi, L.; Hu, F.B. Coffee consumption and markers of inflammation and endothelial dysfunction in healthy and diabetic women. Am J Clin Nutr 2006, 84(4):888-93. [CrossRef]
- Nieman, D.C.; Goodman, C.L.; Capps, C.R.; Shue, Z.L.; Arnot, R. Influence of 2-Weeks Ingestion of High Chlorogenic Acid Coffee on Mood State, Performance, and Postexercise Inflammation and Oxidative Stress: A Randomized, Placebo-Controlled Trial. Int J Sport Nutr Exerc Metab 2018, 28(1):55-65. [CrossRef]
- Rahimi, M.R.; Semenova, E.A.; Larin, A.K.; Kulemin, N.A.; Generozov, E.V.; Łubkowska, B.; Ahmetov, I.I.; Golpasandi, H. The ADORA2A TT Genotype Is Associated with Anti-Inflammatory Effects of Caffeine in Response to Resistance Exercise and Habitual Coffee Intake. Nutrients 2023, 15:1634. [CrossRef]
- Muqaku, B.; Tahir, A.; Klepeisz, P.; Bileck, A.; Kreutz, D.; Mayer, R.L.; Meier, S.M.; Gerner, M.; Schmetterer, K.; Gerner, C. Coffee consumption modulates inflammatory processes in an individual fashion. Mol Nutr Food Res 2016, 60(12):2529-2541. [CrossRef]
- Yang, A.; Palmer, A.A.; de Wit, H. Genetics of caffeine consumption and responses to caffeine. Psychopharmacology (Berl) 2010, 211(3):245-57. [CrossRef]
- Chen, J.F.; Eltzschig, H.K.; Fredholm, B.B. Adenosine receptors as drug targets--what are the challenges? Nat Rev Drug Discov 2013, 12(4):265-86. [CrossRef]
- Volz, N.; Boettler, U.; Winkler, S.; Teller, N.; Schwarz, C.; Bakuradze, T.; Eisenbrand, G.; Haupt, L.; Griffiths, L.R.; Stiebitz, H.; Bytof, G.; Lantz, I.; Lang, R.; Hofmann, T.; Somoza, V.; Marko, D. Effect of coffee combining green coffee bean constituents with typical roasting products on the Nrf2/ARE pathway in vitro and in vivo. J Agric Food Chem 2012, 60(38):9631-41. [CrossRef]
- Rodas, L.; Riera-Sampol, A.; Aguilo, A.; Martínez, S.; Tauler, P. Effects of Habitual Caffeine Intake, Physical Activity Levels, and Sedentary Behavior on the Inflammatory Status in a Healthy Population. Nutrients 2020, 12(8):2325. [CrossRef]
- Guercia, E.; Berti, F.; De Zorzi, R.; Navarini, L.; Geremia, S.; Medagli, B.; De Conto, M.; Cassetta, A.; Forzato, C. On the Cholesterol Raising Effect of Coffee Diterpenes Cafestol and 16-O-Methylcafestol: Interaction with Farnesoid X Receptor. Int J Mol Sci 2024, 25(11):6096. [CrossRef]
- Cordoba, N.; Fernandez-Alduenda, M.; Moreno, L.F.; Ruiz, Y. Coffee extraction: A review of parameters and their influence on the physicochemical characteristics and flavour of coffee brews. Trends in Food Science and Technology 2020, 96:45-60. [CrossRef]
- Janda, K.; Jakubczyk, K.; Baranowska-Bosiacka, I.; Kapczuk, P.; Kochman, J.; Rębacz-Maron, E.; Gutowska, I. Mineral Composition and Antioxidant Potential of Coffee Beverages Depending on the Brewing Method. Foods 2020, 9(2):121. [CrossRef]
- Saad, Al-Fawaeir.; Jafar, M.; Alawneh, Ibrahim, Al-Odat. Influence of coffee consumption on serum lipid profile parameters: Can coffee consumption lead to health consequences in humans? J Agr Food Res 2023, 14:100904.
- Post, S.M.; de Wit, E.C.; Princen, H.M. Cafestol, the cholesterol-raising factor in boiled coffee, suppresses bile acid synthesis by downregulation of cholesterol 7 alpha-hydroxylase and sterol 27-hydroxylase in rat hepatocytes. Arterioscler Thromb Vasc Biol 1997, 17(11):3064-70. [CrossRef]
- Cai, L.; Ma, D.; Zhang, Y.; Liu, Z.; Wang, P. The effect of coffee consumption on serum lipids: a meta-analysis of randomized controlled trials. Eur J Clin Nutr 2012, 66(8):872-7. [CrossRef]
- Habash, M.; Al-Shakhshir, S.; Abusamak, M.; Mohammad, M.Y.; AbuSamak, M. The association of coffee consumption rate with serum 25-hydroxyvitamin D, non-HDL levels, and TC/HDL ratio in females with vitamin D deficiency. Womens Health (Lond) 2022, 18:17455057221112268. [CrossRef]
- Naidoo, N.; Chen, C.; Rebello, S.A.; Speer, K.; Tai, E.S.; Lee, J.; Buchmann, S.; Koelling-Speer, I.; van Dam, R.M. Cholesterol-raising diterpenes in types of coffee commonly consumed in Singapore, Indonesia and India and associations with blood lipids: a survey and cross sectional study. Nutr J 2011, 10:48. [CrossRef]
- Chang, H.C.; Nfor, O.N.; Ho, C.C.; Chen, P.H.; Kung, Y.Y.; Hsu, S.Y.; Tantoh, D.M.; Liaw, Y.C.; Hsieh, C.F.; Liaw, Y.P. Changes in High-Density Lipoprotein Cholesterol Levels in Relation to Coffee Consumption Among Taiwanese Adults. J Multidiscip Healthc 2020, 13:1427-1432. [CrossRef]
- Miranda, A.M.; Goulart, A.C.; Generoso, G.; Bittencourt, M.S.; Santos, R.D.; Toth, P.P.; Jones, S.R.; Benseñor, I.M.; Lotufo, P.A.; Marchioni, D.M. Association between coffee consumption with serum lipid profile in ELSA-Brasil study: a metabolomic approach. Eur J Nutr 2022, 61(8):4205-4214. [CrossRef]
- Svatun, Å.L.; Løchen, M.L.; Thelle, D.S.; Wilsgaard, T. Association between espresso coffee and serum total cholesterol: the Tromsø Study 2015-2016. Open Heart 2022, 9(1):e001946. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



