
Submitted:
30 October 2024
Posted:
31 October 2024
You are already at the latest version
Abstract
Desmoplasia is a hallmark feature of pancreatic ductal adenocarcinoma (PDAC) that contributes significantly to treatment resistance. Approaches to enhance drug delivery into fibrotic PDAC tumors continues to be an important unmet need. In this study, we have engineered a tumor-colonizing, E.coli-based agent that expresses both collagenase and hyaluronidase as a strategy to reduce desmoplasia and enhance intratumoral perfusion of anticancer agents. Overall, we observed that the tandem expression of both these enzymes by tumor-colonizing E.coli resulted in the reduced presence of intratumoral collagen and hyaluronan, which likely contributed to enhanced chemotherapeutic efficacy observed when used in combination. These results highlight the importance of combination treatments involving the depletion of desmoplastic components in PDAC prior to, or during, treatment.
Keywords:
pancreatic ductal adenocarcinoma
; fibrosis
; desmoplasia
; resistance
; hyaluronan
; collagen
; bacterial vector
; hyaluronidase
; collagenase
1. Introduction
Pancreatic ductal adenocarcinoma (PDAC) is the third leading cause of cancer-related death and is expected to rise by 2030. At time of diagnosis, 80% of patients have advanced disease that is ineligible for surgical resection, and 50% have metastasized tumors, leaving PDAC patients with an overall survival of less than a year [1,2,3]. FOLFIRONOX or gemcitabine plus albumin-bound paclitaxel is the first-line standard of care for PDAC patients, with a majority progressing to second-line treatment due to chemoresistance [3,4]. The extracellular matrix (ECM) in PDAC, which is characterized by desmoplasia due to excess production of collagens and hyaluronan (HA), is considered a major contributor to therapeutic resistance. Increased tissue rigidity, interstitial fluidic pressure, and dampened immune response are outcomes of the highly dense tumor stroma. Previous studies have shown that drug penetration is significantly hindered with increased collagen and HA, and depletion of ECM components leads to better drug perfusion and decreased tumor growth in PDAC models [5,6]. In addition to serving as a biophysical barrier, ECM components act as signaling molecules that influence immune responses within tumors. Excess collagen is linked to low CD8+ T cell infiltration and has been proposed as a biomarker for immunotherapy resistance, while upregulation of HA production in tumor cells correlates with immunosuppressive macrophage phenotypes [7,8].
Collagen I is highly abundant in desmoplastic tumors and is a fibril-forming collagen that is crosslinked by LOXL2, a lysyl oxidase that is upregulated in PDAC. Excessive collagen crosslinking leads to fibrosis and a stiff ECM that enhances migration of tumor cells and promotes stem cell transformation [9]. PDAC cell metabolism and proliferation is reliant on collagen I. For example, proline availability is regulated by collagen biosynthesis, and signaling through the DDR1-NFκB-p62-NRF2 cascade leads to changes in mitochondrial protein expression [10,11,12]. Furthermore, PDAC cells use collagen I in an autocrine signaling loop to promote an immunosuppressive tumor microenvironment (TME) with limited T cell infiltration [13]. Collagen IV is also abundant in PDAC and has been proposed as a serum biomarker, as tumor cells actively secrete collagen IV in both primary and metastatic tumors, forming irregular basement membrane-like structures [14]. Type IV collagen in PDAC has been associated with metastasis and angiogenesis, and production of collagen IV allows tumor cells to secrete and anchor to their own basement membrane through integrin receptors, supporting tumor cell survival [15]. Metastasis is often seen in murine models engrafted with cancer cell lines known to produce more collagen IV through interaction with integrins on tumor cell surfaces [16]. In addition to collagens, HA overexpression is prevalent in a majority of PDAC patients, as well as a plethora of other cancers [17]. Low molecular weight HA expressed by PDAC cells interacts with cell surface signaling molecules to increase proliferation through the TLR4 signaling cascade, and in turn increases lymphatic permeability, cell adhesion and matrix remodeling to facilitate metastasis, local inflammation and fibrosis [18,19]. Collagen I, collagen IV, and HA are abundant in PDAC stroma and are negatively correlated with patient survival and chemotherapeutic response [20,21], making these ECM proteins enticing therapeutic targets that may delay disease progression and increase susceptibility to chemo- and immunotherapy.
Collagenases and hyaluronidases, enzymes that degrade collagen and HA respectively, are being explored in preclinical and clinical studies to sensitize PDAC tumors to the immune system and increase permeability for therapeutic intervention [22]. Bacterial collagenases were first studied and isolated from Clostridium histolyticum due to their ability to degrade human tissue [23,24], while bacterial hyaluronidases were characterized by Karl Meyer in the 1970s for their pathological role [25]. The FDA has approved use of bacterial collagenases and hyaluronidases for various diseases, including the localized treatment of solid tumors. Since collagen and HA play an important role in tissue integrity throughout the body, systemic administration is not a viable option and intratumoral injection for localized treatment remains challenging [26,27,28]. To overcome these challenges, we previously showed that attenuated Salmonella typhimurium engineered to express collagenase or hyaluronidase selectively colonize PDAC tumors in mice and degrade collagen or HA, respectively, to increase sensitivity to immuno- and chemotherapy [29,30].
To attain a synergistic effect with increased ECM degradation and decreased tumor progression, we have employed ColH, a collagenase isolated from Clostridium histolyticum [31], and HylB, a hyaluronidase found in Streptococcus agalactiae [32], in order to effectively degrade the ECM within solid PDAC tumors, and we have engineered E.coli strain BL21 to express both ColH and HylB as a single inducible agent with tumor specificity (BL21-TAN). BL21-TAN efficiently colonizes tumors, expresses both enzymes, and depletes collagen and HA content in PDAC tumor models, leading to an enhanced therapeutic outcome when combined with chemotherapeutic treatment. We predict that BL21-TAN has the potential to serve as a combination treatment to increase sensitivity of PDAC tumors to both chemotherapy and immunotherapy, significantly improving the outcomes of patients diagnosed with PDAC.
2. Materials and Methods
Animals and Cell Lines
C57BL/6 and NOD/SCID gamma (NSG) mice were bred and housed at the City of Hope (COH) Biomedical Research Center (BRC). For all studies, animals were handled according to standard IACUC guidelines under an approved protocol. KPC cells were generously provided by Dr. Laleh Melstrom. The UPN cell line was generated at COH from de-identified patient PDAC resection. BxPC3 was obtained commercially from ATCC (CRL-1687). KPC and UPN cells were maintained in DMEM and BxPC3 in RPMI. Both medias contained 10% FBS, 2mM L-glutamine, 100 units/mL penicillin, and 100 μg/mL streptomycin. Prior to tumor cell engraftment, cells were passaged ≤5 times at ≤80% confluency.
E.coli Strains and Generation of BL21-TAN
BL21 (DE3) chemically competent E. coli was obtained from New England Biolabs and cultured in LB media. The ColH (Genbank Accession no. BAA34542) and HylB (Genbank Accession no. AAA56749.1) amino acid sequences were used to synthesize a codon-optimized cDNA sequences inserted into the NcoI site of the pBAD-His-A bacterial expression vector (Biomatik), with a c-Myc tag fused to ColH and a hexahistadine tag fused to HylB. In-frame insertion of TAN into the pBAD vector allowed for expression of ColH and HylB upon induction with L-arabinose. Plasmids were transformed into BL21 (DE3) and spread onto LB plates containing 100 μg/mL ampicillin and incubated overnight at 37°C. Positive clones were identified by colony PCR.
Bacterial Growth, Viability, and Analysis of TAN Expression
BL21-TAN was cultured in media with (induced, I) or without (uninduced, U) 0.1% L-arabinose at 37°C, 225 rpm for time intervals ranging from 2 to 24 h. ColH was detected in bacterial lysates by Western blot using a primary monoclonal mouse anti-Myc antibody (R&D). HylB was detected in bacterial lysates by Western blot using primary monoclonal goat anti-His antibody (R&D). Growth kinetics were monitored through absorbance readings at 600 nm (Genesys 30, Thermo Fisher Scientific) up to 24 h. For immunofluorescence, uninduced and induced BL21-TAN grown for ~4 h were fixed with 4% paraformaldehyde at room temperature (RT) for 30 minutes, and permeabilized with 0.1% Triton-X 100/PBS pH=7.2 at RT for 30 minutes followed by lysozyme (Sigma, 100 μg/mL final concentration in 5 mM EDTA) treatment at RT for 45 minutes. Fixed/permeabilized bacteria were incubated with primary antibody (1:100) for 30 minutes with shaking in a humidified 37°C incubator followed by incubation with FITC-conjugated anti-mouse secondary (1:200, Abcam) and DAPI for 30 minutes with shaking in a humidified 37°C incubator.
Fluorometric Substrate Assays
BL21-TAN was cultured under uninduced or induced conditions for 4 h at 37°C. To measure collagenase activity, substrates consisting of bovine skin collagen type I, human placenta collagen type IV, and pig skin gelatin conjugated to FITC (Thermo Fisher Scientific) were used. The reaction was started by the addition of BL21-TAN under uninduced or induced conditions. Hyaluronidase activity was measured using hyaluronan conjugated to FITC (Thermo Fisher Scientific). Fluorescence intensity was captured using an iBright FL1500 Imaging System (Thermo Fisher Scientific). Fluorescence intensity was quantified in relation to the BL21-TAN uninduced control condition using ImageJ (NIH).
Immunohistochemistry/Immunofluorescence (IHC/IF)
FFPE tumors from C57BL/6 or NSG mice were sectioned (5 μm), transferred to glass slides, deparaffinized, and rehydrated. To determine in vitro enzymatic function, slides were treated with BL21-TAN uninduced or induced for 4 h. To examine in vivo colonization, expression, and function, mice were treated with BL21-TAN or BL21-eGFP control prior to tumor collection. Slides were stained with trichrome and cover slipped. Hematoxylin and methyl blue channels were separated using color deconvolution, and thresholds were set to cover positive staining area for hematoxylin (red) and the raw integrated density was used to measure the density of collagen. To determine HA density, slides were stained with HA-binding protein (HABP). (Millipore Sigma) BL21 colonization was detected using a primary anti-BL21 lysate antibody (R&D), ColH was detected using a primary monoclonal mouse anti-Myc antibody (R&D), and HylB was detected using primary monoclonal goat anti-His antibody (R&D). Brightfield and fluorescent images were acquired with a 63x 1.4NA Plan-Apochromat objective (630x total magnification). Stitching was performed using ZEN 2.3 Pro (Blue) software (Carl Zeiss Inc.). Quantification of brightfield images was performed using ImageJ (NIH) and quantification of fluorescence intensity was performed using Quantitative Pathology & Bioimage Analysis (QuPath) software (v0.2.1, University of Edinburgh, Edinburgh, UK) [33].
Tumor Implantation, Administration, and Induction of BL21-TAN
For all experiments, 6-8 week-old mice were used. 2x106 BxPC3 cells or 5x105 KPC cells were subcutaneously injected into the right flank of NSG mice or C57Bl/6 mice, respectively, in a volume of 100 μL PBS using a 27-gauge needle. Mice with palpable tumors (>150mm3) were intravenously injected with 5x107 colony-forming units (CFUs) BL21-TAN or BL21-eGFP control per day for three consecutive days. Forty-eight hours following final BL21 administration, protein expression was induced by administration of 40 mg L-arabinose through intraperitoneal route. Forty-eight hours following induction, tumors were collected and analyzed via IHC/IF staining for colonization, enzymatic expression, and enzymatic function.
Combination Treatment Studies with Gemcitabine
Following BL21 induction treatment as described above, mice were subsequently administered gemcitabine (15 mg/kg) or PBS 48 hours post-induction. Maintenance doses of gemcitabine or PBS control were administered every 3 days thereafter.
Statistics
All statistical analyses were performed using Prism software by GraphPad (v9). Data was analyzed by performing Student’s t-test and 2-way ANOVA. Unless otherwise indicated, all error bars represent the standard error of the mean.
3. Results
3.1. Construction and Inducible Expression of BL21-TAN
To obtain inducible expression of ColH and HylB, we utilized the pBAD expression system, which employs the araBAD operon that is tightly regulated by L-arabinose availability [34]. The use of this system allows for controlled expression of ColH and HylB within the tumor, limiting off-target toxicities that are expected with systemic collagenase and hyaluronidase expression [35,36,37]. We designed a construct that encodes for ColH with a C-terminal Myc tag and HylB with a C-terminal His tag under the regulation of the pBAD promoter. Each gene is preceded by an independent N-terminal ribosomal binding site to ensure expression of two individual proteins upon induction (Figure 1A). The constructed plasmid was then transformed into BL21 E. coli, which was selected for its ability to colonize solid tumors and its efficiency in the production of recombinant proteins [38,39,40]. Colonies that integrated the plasmid were grown in media to the exponential phase (OD600, ~0.5) and placed under uninduced (U; 0% L-arabinose) or induced (I; 0.02% L-arabinose) conditions for 4 additional hours. Western blot analysis of cell lysates and conditioned culture media (Figure 1B) confirmed the expression of both ColH and HylB under induced conditions, detected by their C-terminal tags. Minimal to no expression was observed in uninduced samples, demonstrating tight regulation of protein expression. The presence of both ColH and HylB in the culture media further suggests that these proteins are secreted by BL21 E. coli post induction. To determine the effect of TAN expression on bacterial growth kinetics, BL21-TAN was grown in uninduced or induced conditions, alongside BL21-eGFP as an additional control and the optical density (OD600) was measured at multiple timepoints over 24 hours (Figure 1C). There was an observable initial increase in OD600 that stabilized by 16 hours at an approximate OD600 of 5.0 in all three conditions, suggesting that ColH and HylB expression, overall, has no detrimental effect on BL21 growth. While we have demonstrated tight regulation of protein expression and sufficient growth of bacterial cells under induced conditions, it is important that both ColH and HylB are expressed simultaneously in a single bacterium for further research. To observe the proteins, we performed immunofluorescent staining of the C-terminal Myc-tag and His-tag fused to ColH and HylB, respectively (Figure 1D). Staining of both proteins in single bacterium was observed in cultures grown under induced conditions and was observed outside of the genomic DNA staining (DAPI). This confirms that both ColH and HylB are expressed simultaneously in the same bacterium and, in this case, are likely localized to the periplasm on their way to being secreted.
3.2. BL21-TAN Degrades Collagen I, Collagen IV, and Hyaluronic acid In Vitro
To assess the enzymatic activity of ColH and HylB produced by BL21-TAN, we grew the engineered bacterial cultures under uninduced and induced conditions and performed in vitro functional assays utilizing FITC-labeled pig skin gelatin (Figure 2A), bovine skin collagen type I (Figure 2B), human placenta collagen type IV (Figure 2C), or HA (Figure 2D) as substrates. Following co-incubation of FITC-labeled substrate with BL21-TAN, the extent of substrate degradation by enzymatic hydrolysis was measured by an increase in fluorescent molecules. Collagen type I and IV, two prevalent collagen types in PDAC, were degraded when co-incubated with induced BL21-TAN, consistent with previous studies on the functional range of ColH [14,31]. Additionally, an increase in fluorescence intensity was observed in FITC-HA substrate solutions following co-incubation with induced BL21-TAN, indicating the degradation of hyaluronic acid. These results confirm that ColH and HylB expressed by BL21-TAN are enzymatically active. Significant increases in fluorescence intensity between uninduced and induced conditions were observed in assays with FITC-collagen I (p<0.0001, t-test), collagen IV (p<0.0001, t-test), and HA (p<0.0001, t-test), demonstrating that BL21-TAN is tightly regulated by the pBAD inducible expression system in vitro, with induction resulting in increased functional enzyme production. In contrast, no difference was observed between uninduced and induced cultures co-incubated with FITC-gelatin, suggesting no effect on baseline gelatin degradation. This finding is in agreement with previous studies confirming that ColH does not exhibit gelatinase activity [31].
3.3. BL21-TAN Effectively Depletes PDAC Tumor-Derived Collagen and Hyaluronic Acid
Collagen and HA derived from healthy tissue differ in structure and function in comparison to tumor-derived collagen and HA. Collagen in healthy tissue is flexible with varying diameter between different tissue types, while tumor-derived collagen is thick, stiff, and aligned, leading to dense, fibrotic tissue that promotes tumor cell growth and metastasis [41,42]. HA metabolism is aberrantly regulated in tumor tissue compared to healthy tissue, resulting in an increase in the abundance of low molecular weight HA in tumor tissue, which promotes cancer cell proliferation, angiogenesis, and metastasis [43]. To determine the efficacy of BL21-TAN in degrading tumor-derived collagen and HA, we obtained serial sections of tumors grown from the murine-derived KPC cell line and human-derived BxPC3 cell line, as well as a de-identified patient (UPN) derived PDAC tumor and treated them with uninduced or induced cultures of BL21-TAN in vitro overnight. Tumor sections were then subjected to staining with trichrome to detect collagen (Figure 3A) or biotin-labeled HA binding protein (HABP; Figure 3B). Collagen content in the trichrome-stained sections was quantified by deconvolution using ImageJ and the raw integrated density of collagen was measured (Figures 3C and S1), while QuPath was used to quantify HA density, determined by the relative fluorescence intensity of HABP (Figure 3D). A significant decrease in both collagen density (p= 0.0014 for KPC, p= 0.005 for BXPC3, p=0.0002 for UPN, t-tests) and HA content (p= 0.0065 for KPC, p<0.0001 for BXPC3, p=0.0029 for UPN, t-tests) was observed in sections treated with induced cultures of BL21-TAN when compared to sections incubated with uninduced cultures in all three tumors. These results suggest that ColH and HylB produced by BL21-TAN effectively deplete tumor-derived collagen and HA and further support the ability to regulate enzymatic function through induced expression of TAN.
3.4. BL21-TAN Expresses Both ColH and HylB in PDAC Murine Models
We evaluated the ability of intravenously delivered BL21-TAN to colonize tumors in C57BL/6 mice with subcutaneous (s.c.) KPC tumors and NSG mice with s.c. BxPC3 tumors through immunofluorescence analysis. BL21-eGFP was used as a vector control to display any observed effects caused by bacterial colonization alone. Following intravenous administration of BL21-eGFP or BL21-TAN and subsequent induction with intraperitoneal (i.p.) L-arabinose, BL21 staining was observed in both tumor models treated with BL21-eGFP and BL21-TAN, as expected (Figure 4). These results demonstrate effective tumor colonization by BL21, independent of the expressed genes. To ensure that BL21 colonized tumors while effectively clearing from systemic circulation, we examined the spleens, where bacteria from the bloodstream typically accumulate during infection [44]. Spleens from tumor-bearing mice treated with ampicillin-resistant BL21 were homogenized and subjected to microbial enumeration by serial plating on LB agar containing ampicillin (data not shown). No bacterial colonies were detected, indicating that BL21 colonization was limited to the tumors. Together these data strongly suggest that BL21 colonization is tumor specific. Next, we assessed in vivo enzyme expression following systemic treatment of tumor-bearing mice with BL21-TAN. Antibodies against the Myc-tag on ColH and the His-tag on HylB were used to visualize the expression of bacterially produced enzymes after induction of the pBAD promoter. KPC and BxPC3 tumors treated with BL21-TAN dual expression of ColH and HylB (Figure 4), indicating successful in vivo induction of the TAN construct. ColH and HylB expression was localized to regions with high BL21 density, further confirming that BL21-TAN expresses both ECM-degrading enzymes within the tumor microenvironment in two PDAC tumor models.
3.5. BL21-TAN Depletes Collagen and Hyaluronan in Murine Model of PDAC
We demonstrated that BL21-TAN preferentially colonizes tumors and displays dual expression of ColH and HylB, therefore the next important question is whether these enzymes are functionally active under physiological conditions. To determine the functional capacity of ECM-degrading enzymes produced by BL21-TAN, BxPC3 tumors in NSG mice treated with either BL21-eGFP or BL21-TAN were analyzed for collagen and HA abundance using trichrome (Figure 5A) or HABP (Figure 5B). BxPC3 tumors from mice treated with BL21-TAN exhibited significantly lower collagen levels compared to tumor sections from mice treated with BL21-eGFP (p<0.0001, t-test ; Figure 5C). Similarly, IHC staining of hyaluronan revealed significantly less hyaluronan in tumors from mice treated with BL21-TAN compared to tumors treated with BL21-eGFP(p<0.0001, t-test ; Figure 5C). These data suggest that BL21-TAN effectively degrades intratumoral collagen and HA under physiological conditions.
3.6. Pre-Treatment with BL21-TAN Increases Gemcitabine Efficacy in Murine Model of PDAC
Desmoplasia in PDAC tumors has been shown to be a pathological barrier, hindering drug penetration by affecting vasculature, increasing interstitial pressure, and increasing organ stiffness [45,46]. Collagen and hyaluronan promote therapeutic resistance in solid tumors through signaling pathways and receptors, such as TGF-β, LAIR-1, and CD44 [7,47,48]. Therefore, we hypothesized that PDAC tumors treated with ECM-degrading BL21-TAN would lead to increased efficacy of chemotherapeutic interventions. NSG mice bearing BxPC3 tumors were treated with BL21-eGFP or BL21-TAN for three consecutive days and induced with 40mg L-arabinose delivered via i.p. injection twenty-four hours after the final BL21 dose. Control groups receiving PBS only or gemcitabine only were administered PBS. Gemcitabine was administered by i.p. injection 48 hours post-induction, and then every three days for a total of 23 days, until mice in the PBS-control group reached humane endpoints (Figure 6A). Tumor volume was measured every 3-5 days, and mouse weights were monitored to assess potential toxicity. Tumors in mice receiving control treatments began growing exponentially around day 14, while the growth rate of tumors in mice that received BL21-TAN and gemcitabine combination treatment was inhibited. By day 23, the tumor sizes in the BL21-TAN plus gemcitabine combination treatment group was significantly smaller (Figure 6B) than those treated with PBS (p=0.0002, mixed-effects), gemcitabine only (p<0.0001, mixed-effects) and BL21-TAN only (p<0.0001, mixed-effects). This data indicates that BL21-TAN pre-treatment enhanced the efficacy of gemcitabine in PDAC tumors. Furthermore, the tumor sizes were significantly lower in the BL21-TAN plus gemcitabine group compared to the BL21-eGFP plus gemcitabine group (p=0.0006, mixed-effects), suggesting that the enhanced efficacy of combination treatment is specifically due to the degradation of collagen and HA within the tumor by BL21-TAN. BL21-TAN monotherapy also appeared to decelerate tumor growth, albeit not as well as combination with chemotherapy. Furthermore, mouse weights were comparable across all treatments, indicating that treatment with BL21-TAN was not a source of increased toxicity (Figure 6C). Taken together with our previous results, our study showed that BL21-TAN pretreatment depletes HA and collagen in the PDAC tumors, leading to an increased efficacy of chemotherapeutic treatment.
4. Discussion
Desmoplasia is a hallmark of PDAC which leads to obstructed drug penetration and immunosuppression, making effective therapeutic treatment difficult in PDAC patients [5,6]. Tumor metabolism, survival and migration is promoted by abundant, aberrant collagen and HA in PDAC tumors [9,10,11,12,18,19]. For these reasons, depletion of collagen and HA are anticipated to serve as an effective therapeutic intervention for desmoplastic tumors [35,49]. Various different avenues for targeting the fibrotic ECM are being explored, such as inducing the apoptosis of cells responsible for ECM deposition, inhibiting collagen synthesis and collagen cross-linking, and HA degradation to decrease interstitial fluid pressure. However, these therapies have had limited success in clinical trials due to difficulties with targeted delivery of molecules, insufficient tumor regression and increased disease progression [50,51]. Therefore, development of tumor-targeting, ECM depleting treatments are necessary.
Collagen and HA are integral components of the ECM present in most tissues. Therefore, to achieve targeted treatment while limiting systemic toxicity, we, along with other groups, previously employed tumor-colonizing bacteria engineered to deliver ECM-degrading enzymes directly to the tumor [29,52,53]. Here, we have engineered gram-negative bacteria to deliver both collagenase and hyaluronidase, as a single agent therapeutic, to PDAC tumor models. We show that the engineered bacteria successfully colonize murine models of PDAC tumors, express ECM-degrading enzymes, and deplete tumor-derived collagen and HA.
Degradation of ECM-proteins increased biomaterial diffusion and distribution in tumor-like spheroids in vitro and intratumoral distribution in vivo [54,55,56,57]. Therefore, we theorized that BL21-TAN pre-treatment would increase penetration of gemcitabine in PDAC tumors, following degradation of tumor ECM. We showed that tumor-bearing mice pre-treated with BL21-TAN followed by gemcitabine had increased survival and decreased tumor burden compared to mice treated with gemcitabine alone. This indicates that BL21-TAN pre-treatment increases efficacy of chemotherapy in PDAC models, most likely by creating a less fibrotic microenvironment which facilitated diffusion of gemcitabine.
Tumor-derived collagen and HA differ in function and structure compared to healthy tissue and contribute to the immunosuppressive TME in solid tumors. Leukocyte-associated immunoglobulin-like receptor-1 (LAIR-1), an inhibitory receptor, binds to collagen in the TME and leads to decreased immune activation and decreased immune cell infiltration [7,58]. HA promotes the differentiation of tumor-associated macrophages, resulting in increased immunosuppression within the TME through PD-1/PD-L1 signaling. Dense ECM also leads to interstitial fluid flow toward tumor margins, leading to the expression of TGFβ in cancer-associated fibroblasts, which polarizes regulatory T cell and immunosuppressive macrophage phenotypes [59,60]. Furthermore, increased organ stiffness, due to desmoplasia, is correlated to a poor immunotherapeutic response and has been suggested as a prognostic marker for immunotherapeutic intervention [61,62]. Future studies might consider looking into the infiltration and activation status of immune cells following treatment with BL21-TAN, as there is evidence that fibrotic, tumor-associated ECM contributes to the immunosuppressive TME.
5. Conclusions
This work, in addition to our previous work utilizing bacteria expressing only a single ECM-degrading enzyme, emphasizes the importance of eliminating desmoplasia in PDAC to maximize the effectiveness of therapy.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Representative analysis showing deconvolution of trichrome staining for collagen density within tumor sections.
Author Contributions
LCA and ERM conceived and planned the experiments. LCA, SL, NDE, AFS, IRT and ERM carried out the experiments. LCA and ERM contributed to the interpretation of the results. LCA and ERM generated figures. LCA wrote the manuscript. All authors provided critical feedback to significantly improve experimental design, analysis and composition of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Cancer Institute (NCI) of the National Institutes of Health (NIH) under grant numbers R01CA266472, R01CA272732, R21CA256593, and R21CA293969. Research reported in this publication included work performed in the Molecular Pathology, Animal Resource Center, and Light Microscopy Digital Imaging cores supported by the NCI of the NIH under grant number P30CA033572. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Institutional Review Board Statement
For all studies, animals were handled according to standard Institutional Animal Care and Use Committee (IACUC) guidelines under an approved protocol.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA Cancer J Clin. 2024;74(1):12-49. Epub 2024/01/17. [CrossRef] [PubMed]
- Duan X, Zhu Q, Zhang X, Shen Z, Huang Y. Expression, biochemical and structural characterization of high-specific-activity beta-amylase from Bacillus aryabhattai GEL-09 for application in starch hydrolysis. Microb Cell Fact. 2021;20(1):182. Epub 2021/09/20. [CrossRef] [PubMed] [PubMed Central]
- Wood LD, Canto MI, Jaffee EM, Simeone DM. Pancreatic Cancer: Pathogenesis, Screening, Diagnosis, and Treatment. Gastroenterology. 2022;163(2):386-402 e1. Epub 2022/04/11. [CrossRef] [PubMed] [PubMed Central]
- Kang J, Hwang I, Yoo C, Kim KP, Jeong JH, Chang HM, Lee SS, Park DH, Song TJ, Seo DW, Lee SK, Kim MH, Hong SM, Shin SH, Hwang DW, Song KB, Lee JH, Kim SC, Ryoo BY. Nab-paclitaxel plus gemcitabine versus FOLFIRINOX as the first-line chemotherapy for patients with metastatic pancreatic cancer: retrospective analysis. Invest New Drugs. 2018;36(4):732-41. Epub 2018/04/05. [CrossRef] [PubMed]
- Hwang HJ, Oh MS, Lee DW, Kuh HJ. Multiplex quantitative analysis of stroma-mediated cancer cell invasion, matrix remodeling, and drug response in a 3D co-culture model of pancreatic tumor spheroids and stellate cells. J Exp Clin Cancer Res. 2019;38(1):258. Epub 2019/06/16. [CrossRef] [PubMed] [PubMed Central]
- Jacobetz MA, Chan DS, Neesse A, Bapiro TE, Cook N, Frese KK, Feig C, Nakagawa T, Caldwell ME, Zecchini HI, Lolkema MP, Jiang P, Kultti A, Thompson CB, Maneval DC, Jodrell DI, Frost GI, Shepard HM, Skepper JN, Tuveson DA. Hyaluronan impairs vascular function and drug delivery in a mouse model of pancreatic cancer. Gut. 2013;62(1):112-20. Epub 2012/04/03. [CrossRef] [PubMed] [PubMed Central]
- Peng DH, Rodriguez BL, Diao L, Chen L, Wang J, Byers LA, Wei Y, Chapman HA, Yamauchi M, Behrens C, Raso G, Soto LMS, Cuentes ERP, Wistuba, II, Kurie JM, Gibbons DL. Collagen promotes anti-PD-1/PD-L1 resistance in cancer through LAIR1-dependent CD8(+) T cell exhaustion. Nat Commun. 2020;11(1):4520. Epub 2020/09/11. GlaxoSmithKline, Astellas, Ribon Therapeutics and Sanofi. D.L.G. receives research grant funding from AstraZeneca, Janssen, Ribon Therapeutics, Astellas and Takeda. L.A.B. declares consulting work for AstraZeneca, AbbVie, GenMab, BergenBio, Pharma Mar, SA. L.A.B. receives research grant funding from AbbVie, AstraZeneca, GenMab, Tolero Pharmaceuticals. All other authors declare that they have no conflict of interests. [CrossRef] [PubMed] [PubMed Central]
- Kuang DM, Wu Y, Chen N, Cheng J, Zhuang SM, Zheng L. Tumor-derived hyaluronan induces formation of immunosuppressive macrophages through transient early activation of monocytes. Blood. 2007;110(2):587-95. Epub 2007/03/31. [CrossRef] [PubMed]
- Karsdal MA, Nielsen SH, Leeming DJ, Langholm LL, Nielsen MJ, Manon-Jensen T, Siebuhr A, Gudmann NS, Ronnow S, Sand JM, Daniels SJ, Mortensen JH, Schuppan D. The good and the bad collagens of fibrosis - Their role in signaling and organ function. Adv Drug Deliv Rev. 2017;121:43-56. Epub 2017/07/25. [CrossRef] [PubMed]
- Palka J, Oscilowska I, Szoka L. Collagen metabolism as a regulator of proline dehydrogenase/proline oxidase-dependent apoptosis/autophagy. Amino Acids. 2021;53(12):1917-25. Epub 2021/04/06. [CrossRef] [PubMed] [PubMed Central]
- Olivares O, Mayers JR, Gouirand V, Torrence ME, Gicquel T, Borge L, Lac S, Roques J, Lavaut MN, Berthezene P, Rubis M, Secq V, Garcia S, Moutardier V, Lombardo D, Iovanna JL, Tomasini R, Guillaumond F, Vander Heiden MG, Vasseur S. Collagen-derived proline promotes pancreatic ductal adenocarcinoma cell survival under nutrient limited conditions. Nat Commun. 2017;8:16031. Epub 2017/07/08. [CrossRef] [PubMed] [PubMed Central]
- Su H, Yang F, Fu R, Trinh B, Sun N, Liu J, Kumar A, Baglieri J, Siruno J, Le M, Li Y, Dozier S, Nair A, Filliol A, Sinchai N, Rosenthal SB, Santini J, Metallo CM, Molina A, Schwabe RF, Lowy AM, Brenner D, Sun B, Karin M. Collagenolysis-dependent DDR1 signalling dictates pancreatic cancer outcome. Nature. 2022;610(7931):366-72. Epub 2022/10/06. [CrossRef] [PubMed] [PubMed Central]
- Chen Y, Yang S, Tavormina J, Tampe D, Zeisberg M, Wang H, Mahadevan KK, Wu CJ, Sugimoto H, Chang CC, Jenq RR, McAndrews KM, Kalluri R. Oncogenic collagen I homotrimers from cancer cells bind to alpha3beta1 integrin and impact tumor microbiome and immunity to promote pancreatic cancer. Cancer Cell. 2022;40(8):818-34 e9. Epub 2022/07/23. [CrossRef] [PubMed] [PubMed Central]
- Ohlund D, Lundin C, Ardnor B, Oman M, Naredi P, Sund M. Type IV collagen is a tumour stroma-derived biomarker for pancreas cancer. Br J Cancer. 2009;101(1):91-7. Epub 2009/06/06. [CrossRef] [PubMed] [PubMed Central]
- Ohlund D, Franklin O, Lundberg E, Lundin C, Sund M. Type IV collagen stimulates pancreatic cancer cell proliferation, migration, and inhibits apoptosis through an autocrine loop. BMC Cancer. 2013;13:154. Epub 2013/03/28. [CrossRef] [PubMed] [PubMed Central]
- Burnier JV, Wang N, Michel RP, Hassanain M, Li S, Lu Y, Metrakos P, Antecka E, Burnier MN, Ponton A, Gallinger S, Brodt P. Type IV collagen-initiated signals provide survival and growth cues required for liver metastasis. Oncogene. 2011;30(35):3766-83. Epub 2011/04/12. [CrossRef] [PubMed]
- Markowska A, Antoszczak M, Markowska J, Huczynski A. Role of Hyaluronic Acid in Selected Malignant Neoplasms in Women. Biomedicines. 2023;11(2). Epub 2023/02/26. [CrossRef] [PubMed] [PubMed Central]
- Michalczyk M, Humeniuk E, Adamczuk G, Korga-Plewko A. Hyaluronic Acid as a Modern Approach in Anticancer Therapy-Review. Int J Mol Sci. 2022;24(1). Epub 2023/01/09. [CrossRef] [PubMed] [PubMed Central]
- Inokoshi Y, Tanino Y, Wang X, Sato S, Fukuhara N, Nikaido T, Fukuhara A, Saito J, Frevert CW, Munakata M. Clinical significance of serum hyaluronan in chronic fibrotic interstitial pneumonia. Respirology. 2013;18(8):1236-43. Epub 2013/06/26. [CrossRef] [PubMed]
- Magzoub M, Jin S, Verkman AS. Enhanced macromolecule diffusion deep in tumors after enzymatic digestion of extracellular matrix collagen and its associated proteoglycan decorin. FASEB J. 2008;22(1):276-84. Epub 2007/09/01. [CrossRef] [PubMed]
- Whatcott CJ, Diep CH, Jiang P, Watanabe A, LoBello J, Sima C, Hostetter G, Shepard HM, Von Hoff DD, Han H. Desmoplasia in Primary Tumors and Metastatic Lesions of Pancreatic Cancer. Clin Cancer Res. 2015;21(15):3561-8. Epub 2015/02/20. [CrossRef] [PubMed] [PubMed Central]
- Park W, Chawla A, O'Reilly EM. Pancreatic Cancer: A Review. JAMA. 2021;326(9):851-62. Epub 2021/09/22. [CrossRef] [PubMed] [PubMed Central]
- Maclennan JD, Mandl I, Howes EL. Bacterial digestion of collagen. J Clin Invest. 1953;32(12):1317-22. Epub 1953/12/01. [CrossRef] [PubMed] [PubMed Central]
- Harrington DJ. Bacterial collagenases and collagen-degrading enzymes and their potential role in human disease. Infect Immun. 1996;64(6):1885-91. Epub 1996/06/01. [CrossRef] [PubMed] [PubMed Central]
- Jedrzejas MJ. Structural and functional comparison of polysaccharide-degrading enzymes. Crit Rev Biochem Mol Biol. 2000;35(3):221-51. Epub 2000/07/25. [CrossRef] [PubMed]
- Burgeson RE, Nimni ME. Collagen types. Molecular structure and tissue distribution. Clin Orthop Relat Res. 1992(282):250-72. Epub 1992/09/01. [PubMed]
- Fraser JR, Laurent TC, Laurent UB. Hyaluronan: its nature, distribution, functions and turnover. J Intern Med. 1997;242(1):27-33. Epub 1997/07/01. [CrossRef] [PubMed]
- Lian C, Wang X, Qiu X, Wu Z, Gao B, Liu L, Liang G, Zhou H, Yang X, Peng Y, Liang A, Xu C, Huang D, Su P. Collagen type II suppresses articular chondrocyte hypertrophy and osteoarthritis progression by promoting integrin beta1-SMAD1 interaction. Bone Res. 2019;7:8. Epub 2019/03/12. [CrossRef] [PubMed] [PubMed Central]
- Ebelt ND, Zamloot V, Zuniga E, Passi KB, Sobocinski LJ, Young CA, Blazar BR, Manuel ER. Collagenase-Expressing Salmonella Targets Major Collagens in Pancreatic Cancer Leading to Reductions in Immunosuppressive Subsets and Tumor Growth. Cancers (Basel). 2021;13(14). Epub 2021/07/25. [CrossRef] [PubMed] [PubMed Central]
- Ebelt ND, Zuniga E, Passi KB, Sobocinski LJ, Manuel ER. Hyaluronidase-Expressing Salmonella Effectively Targets Tumor-Associated Hyaluronic Acid in Pancreatic Ductal Adenocarcinoma. Mol Cancer Ther. 2020;19(2):706-16. Epub 2019/11/07. [CrossRef] [PubMed] [PubMed Central]
- Zhang YZ, Ran LY, Li CY, Chen XL. Diversity, Structures, and Collagen-Degrading Mechanisms of Bacterial Collagenolytic Proteases. Appl Environ Microbiol. 2015;81(18):6098-107. Epub 2015/07/08. [CrossRef] [PubMed] [PubMed Central]
- Wang Z, Guo C, Xu Y, Liu G, Lu C, Liu Y. Two novel functions of hyaluronidase from Streptococcus agalactiae are enhanced intracellular survival and inhibition of proinflammatory cytokine expression. Infect Immun. 2014;82(6):2615-25. Epub 2014/04/09. [CrossRef] [PubMed] [PubMed Central]
- Bankhead P, Loughrey MB, Fernandez JA, Dombrowski Y, McArt DG, Dunne PD, McQuaid S, Gray RT, Murray LJ, Coleman HG, James JA, Salto-Tellez M, Hamilton PW. QuPath: Open source software for digital pathology image analysis. Sci Rep. 2017;7(1):16878. Epub 2017/12/06. [CrossRef] [PubMed] [PubMed Central]
- Guzman LM, Belin D, Carson MJ, Beckwith J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J Bacteriol. 1995;177(14):4121-30. Epub 1995/07/01. [CrossRef] [PubMed] [PubMed Central]
- Whatcott CJ, Han H, Posner RG, Hostetter G, Von Hoff DD. Targeting the tumor microenvironment in cancer: why hyaluronidase deserves a second look. Cancer Discov. 2011;1(4):291-6. Epub 2011/11/05. [CrossRef] [PubMed] [PubMed Central]
- Ferreira AM, Gentile P, Chiono V, Ciardelli G. Collagen for bone tissue regeneration. Acta Biomater. 2012;8(9):3191-200. Epub 2012/06/19. [CrossRef] [PubMed]
- Ouyang Z, Dong L, Yao F, Wang K, Chen Y, Li S, Zhou R, Zhao Y, Hu W. Cartilage-Related Collagens in Osteoarthritis and Rheumatoid Arthritis: From Pathogenesis to Therapeutics. Int J Mol Sci. 2023;24(12). Epub 2023/06/28. [CrossRef] [PubMed] [PubMed Central]
- Jiang SN, Phan TX, Nam TK, Nguyen VH, Kim HS, Bom HS, Choy HE, Hong Y, Min JJ. Inhibition of tumor growth and metastasis by a combination of Escherichia coli-mediated cytolytic therapy and radiotherapy. Mol Ther. 2010;18(3):635-42. Epub 2010/01/07. [CrossRef] [PubMed] [PubMed Central]
- Gentschev I, Petrov I, Ye M, Kafuri Cifuentes L, Toews R, Cecil A, Oelschaeger TA, Szalay AA. Tumor Colonization and Therapy by Escherichia coli Nissle 1917 Strain in Syngeneic Tumor-Bearing Mice Is Strongly Affected by the Gut Microbiome. Cancers (Basel). 2022;14(24). Epub 2022/12/24. [CrossRef] [PubMed] [PubMed Central]
- Sun XM, Zhang ZX, Wang LR, Wang JG, Liang Y, Yang HF, Tao RS, Jiang Y, Yang JJ, Yang S. Downregulation of T7 RNA polymerase transcription enhances pET-based recombinant protein production in Escherichia coli BL21 (DE3) by suppressing autolysis. Biotechnol Bioeng. 2021;118(1):153-63. Epub 2020/09/09. [CrossRef] [PubMed]
- Popova NV, Jucker M. The Functional Role of Extracellular Matrix Proteins in Cancer. Cancers (Basel). 2022;14(1). Epub 2022/01/12. [CrossRef] [PubMed] [PubMed Central]
- Song K, Yu Z, Zu X, Li G, Hu Z, Xue Y. Collagen Remodeling along Cancer Progression Providing a Novel Opportunity for Cancer Diagnosis and Treatment. Int J Mol Sci. 2022;23(18). Epub 2022/09/24. [CrossRef] [PubMed] [PubMed Central]
- Donelan W, Dominguez-Gutierrez PR, Kusmartsev S. Deregulated hyaluronan metabolism in the tumor microenvironment drives cancer inflammation and tumor-associated immune suppression. Front Immunol. 2022;13:971278. Epub 2022/10/15. [CrossRef] [PubMed] [PubMed Central]
- Siggins MK, Sriskandan S. Bacterial Lymphatic Metastasis in Infection and Immunity. Cells. 2021;11(1). Epub 2022/01/12. [CrossRef] [PubMed] [PubMed Central]
- DuFort CC, DelGiorno KE, Carlson MA, Osgood RJ, Zhao C, Huang Z, Thompson CB, Connor RJ, Thanos CD, Scott Brockenbrough J, Provenzano PP, Frost GI, Michael Shepard H, Hingorani SR. Interstitial Pressure in Pancreatic Ductal Adenocarcinoma Is Dominated by a Gel-Fluid Phase. Biophys J. 2016;110(9):2106-19. Epub 2016/05/12. [CrossRef] [PubMed] [PubMed Central]
- Netti PA, Berk DA, Swartz MA, Grodzinsky AJ, Jain RK. Role of extracellular matrix assembly in interstitial transport in solid tumors. Cancer Res. 2000;60(9):2497-503. Epub 2000/05/16. [PubMed]
- Chaudhry GE, Akim A, Naveed Zafar M, Safdar N, Sung YY, Muhammad TST. Understanding Hyaluronan Receptor (CD44) Interaction, HA-CD44 Activated Potential Targets in Cancer Therapeutics. Adv Pharm Bull. 2021;11(3):426-38. Epub 2021/09/14. [CrossRef] [PubMed] [PubMed Central]
- Chakravarthy A, Khan L, Bensler NP, Bose P, De Carvalho DD. TGF-beta-associated extracellular matrix genes link cancer-associated fibroblasts to immune evasion and immunotherapy failure. Nat Commun. 2018;9(1):4692. Epub 2018/11/10. [CrossRef] [PubMed] [PubMed Central]
- Xu S, Xu H, Wang W, Li S, Li H, Li T, Zhang W, Yu X, Liu L. The role of collagen in cancer: from bench to bedside. J Transl Med. 2019;17(1):309. Epub 2019/09/16. [CrossRef] [PubMed] [PubMed Central]
- Ji T, Feng W, Zhang X, Zang K, Zhu X, Shang F. HDAC inhibitors promote pancreatic stellate cell apoptosis and relieve pancreatic fibrosis by upregulating miR-15/16 in chronic pancreatitis. Hum Cell. 2020;33(4):1006-16. Epub 2020/06/12. [CrossRef] [PubMed] [PubMed Central]
- Ferrara B, Pignatelli C, Cossutta M, Citro A, Courty J, Piemonti L. The Extracellular Matrix in Pancreatic Cancer: Description of a Complex Network and Promising Therapeutic Options. Cancers (Basel). 2021;13(17). Epub 2021/09/11. [CrossRef] [PubMed] [PubMed Central]
- Zamloot V, Ebelt ND, Soo C, Jinka S, Manuel ER. Targeted Depletion of Hyaluronic Acid Mitigates Murine Breast Cancer Growth. Cancers (Basel). 2022;14(19). Epub 2022/10/15. [CrossRef] [PubMed] [PubMed Central]
- Kim JS, Park JE, Choi SH, Kang SW, Lee JH, Lee JS, Shin M, Park SH. ECM-targeting bacteria enhance chemotherapeutic drug efficacy by lowering IFP in tumor mouse models. J Control Release. 2023;355:199-210. Epub 2023/02/08. [CrossRef] [PubMed]
- McKee TD, Grandi P, Mok W, Alexandrakis G, Insin N, Zimmer JP, Bawendi MG, Boucher Y, Breakefield XO, Jain RK. Degradation of fibrillar collagen in a human melanoma xenograft improves the efficacy of an oncolytic herpes simplex virus vector. Cancer Res. 2006;66(5):2509-13. Epub 2006/03/03. [CrossRef] [PubMed]
- Eikenes L, Tufto I, Schnell EA, Bjorkoy A, De Lange Davies C. Effect of collagenase and hyaluronidase on free and anomalous diffusion in multicellular spheroids and xenografts. Anticancer Res. 2010;30(2):359-68. Epub 2010/03/25. [PubMed]
- Goodman TT, Olive PL, Pun SH. Increased nanoparticle penetration in collagenase-treated multicellular spheroids. Int J Nanomedicine. 2007;2(2):265-74. Epub 2007/08/29. [PubMed] [PubMed Central]
- Kohli AG, Kivimae S, Tiffany MR, Szoka FC. Improving the distribution of Doxil(R) in the tumor matrix by depletion of tumor hyaluronan. J Control Release. 2014;191:105-14. Epub 2014/05/24. [CrossRef] [PubMed] [PubMed Central]
- Larsen AMH, Kuczek DE, Kalvisa A, Siersbaek MS, Thorseth ML, Johansen AZ, Carretta M, Grontved L, Vang O, Madsen DH. Collagen Density Modulates the Immunosuppressive Functions of Macrophages. J Immunol. 2020;205(5):1461-72. Epub 2020/08/26. [CrossRef] [PubMed]
- Gong D, Shi W, Yi SJ, Chen H, Groffen J, Heisterkamp N. TGFbeta signaling plays a critical role in promoting alternative macrophage activation. BMC Immunol. 2012;13:31. Epub 2012/06/19. [CrossRef] [PubMed] [PubMed Central]
- Jacob SS, Shastry P, Sudhakaran PR. Monocyte-macrophage differentiation in vitro: modulation by extracellular matrix protein substratum. Mol Cell Biochem. 2002;233(1-2):9-17. Epub 2002/06/27. [CrossRef] [PubMed]
- Huang Y, Lei X, Sun L, Liu Y, Yang J. Leveraging various extracellular matrix levels to assess prognosis and sensitivity to immunotherapy in patients with ovarian cancer. Front Oncol. 2023;13:1163695. Epub 2023/05/25. [CrossRef] [PubMed] [PubMed Central]
- Yuan G, Xie F, Song Y, Li Q, Li R, Hu X, Zang M, Cheng X, Lu G, Huang J, Fan W, Rong X, Sun J, Chen J. Hepatic Tumor Stiffness Measured by Shear Wave Elastography Is Prognostic for HCC Progression Following Treatment With Anti-PD-1 Antibodies Plus Lenvatinib: A Retrospective Analysis of Two Independent Cohorts. Front Immunol. 2022;13:868809. Epub 2022/06/28. [CrossRef] [PubMed] [PubMed Central]
Figure 1.
Tandem Construct Design and Expression Analysis in BL21 E. Coli Transformants. (A) The synthesized sequence encoding for both bacterial collagenase (ColH) and hyaluronidase (HylB) were engineered with independent 5’ ribosomal binding sites (RBS) and fused to a myc or his tag, respectively. The sequence was cloned downstream of the inducible pBAD promoter in the pBAD/HIS A plasmid and then transformed into BL21 to generate BL21-TAN. (B) BL21-TAN cultures were grown to exponential phase (shaking at 37°C) and then left alone (uninduced, U) or induced (I) at a final concentration of 0.02% L-arabinose for 4 hours. Bacterial pellets and culture media (CM) were then subjected to western blot analysis to detect for expression of ColH (anti-Myc) and HylB (anti-His). (C) BL21 transformed with control pBAD-eGFP plasmid (BL21-eGFP) and BL21-TAN were cultured to an optical density (OD600) of ~1. Cultures were either left uninduced (BL21-eGFP, BL21-TAN) or induced at 0.02% L-arabinose (BL21-TAN) and OD600 was measured over time. (D) Uninduced or induced BL21-eGFP (0.02% L-arabinose, 4 hours) were fixed in 4% paraformaldehyde and then stained simultaneously with anti-myc and anti-his to detect ColH and HylB expression, respectively. A representative, single bacterium for each condition is shown.
Figure 1.
Tandem Construct Design and Expression Analysis in BL21 E. Coli Transformants. (A) The synthesized sequence encoding for both bacterial collagenase (ColH) and hyaluronidase (HylB) were engineered with independent 5’ ribosomal binding sites (RBS) and fused to a myc or his tag, respectively. The sequence was cloned downstream of the inducible pBAD promoter in the pBAD/HIS A plasmid and then transformed into BL21 to generate BL21-TAN. (B) BL21-TAN cultures were grown to exponential phase (shaking at 37°C) and then left alone (uninduced, U) or induced (I) at a final concentration of 0.02% L-arabinose for 4 hours. Bacterial pellets and culture media (CM) were then subjected to western blot analysis to detect for expression of ColH (anti-Myc) and HylB (anti-His). (C) BL21 transformed with control pBAD-eGFP plasmid (BL21-eGFP) and BL21-TAN were cultured to an optical density (OD600) of ~1. Cultures were either left uninduced (BL21-eGFP, BL21-TAN) or induced at 0.02% L-arabinose (BL21-TAN) and OD600 was measured over time. (D) Uninduced or induced BL21-eGFP (0.02% L-arabinose, 4 hours) were fixed in 4% paraformaldehyde and then stained simultaneously with anti-myc and anti-his to detect ColH and HylB expression, respectively. A representative, single bacterium for each condition is shown.
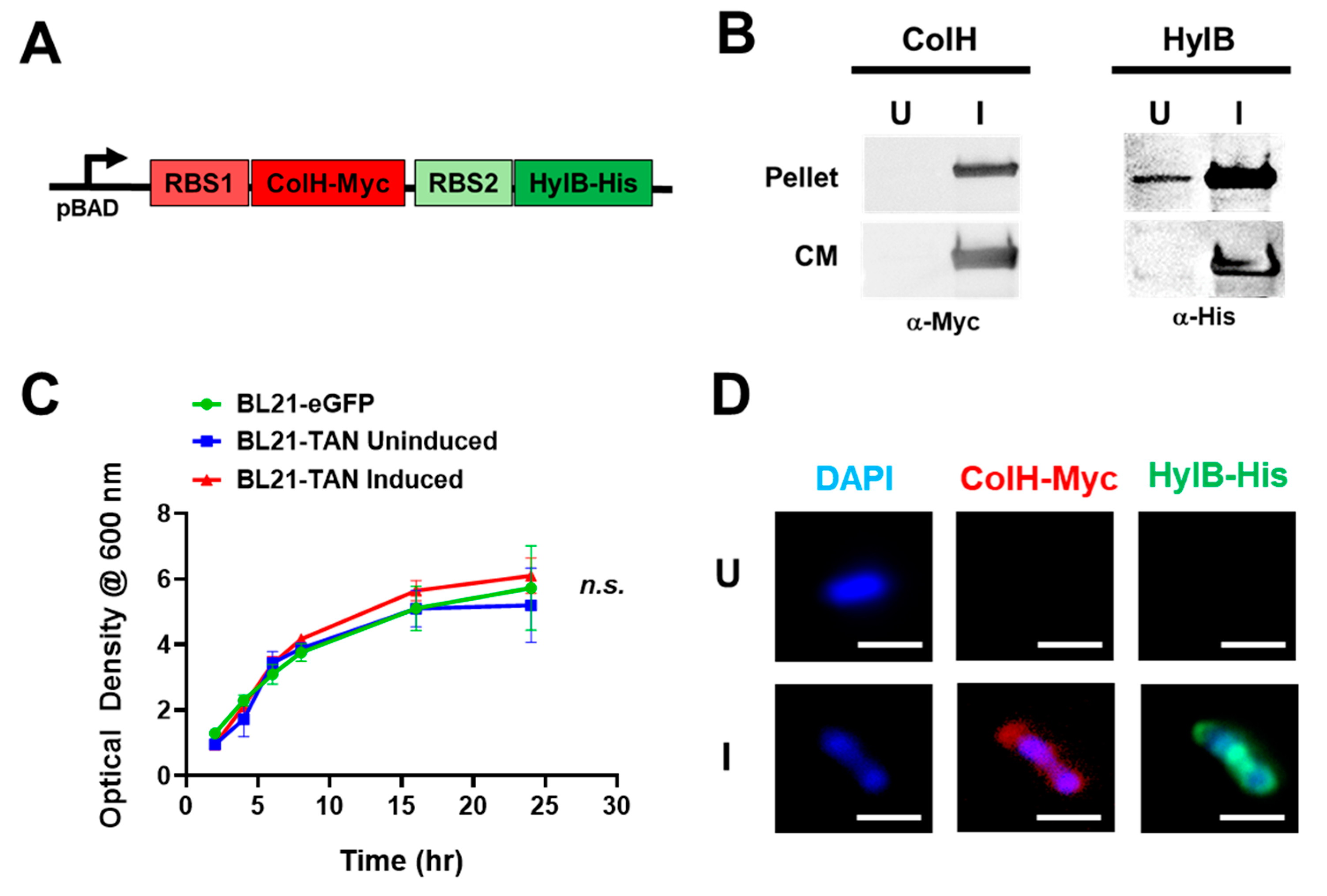
Figure 2.
In Vitro Degradation of Collagens and Hyaluronic Acid by BL21-TAN. Hydrolysis reactions were performed using uninduced (U) or induced (I) BL21-TAN co-incubated with FITC-conjugated pig skin gelatin (A), bovine skin collagen type I (B), human placenta collagen type IV (C) or purified hyaluronic acid (HA) (D) in 50 mM Tris-HCl (pH 8.0) containing 10 mM CaCl2 at 37⁰C. Negative control includes culture media (LB). Increases in fluorescence intensity signifies degradation of FITC-conjugated target. Enzyme activity was measured by monitoring fluorescence (FITC) (ex: 495 nm, em: 519 nm). Data are expressed as mean ± error of mean of three independent experiments. ****p<0.0001, t-test.
Figure 2.
In Vitro Degradation of Collagens and Hyaluronic Acid by BL21-TAN. Hydrolysis reactions were performed using uninduced (U) or induced (I) BL21-TAN co-incubated with FITC-conjugated pig skin gelatin (A), bovine skin collagen type I (B), human placenta collagen type IV (C) or purified hyaluronic acid (HA) (D) in 50 mM Tris-HCl (pH 8.0) containing 10 mM CaCl2 at 37⁰C. Negative control includes culture media (LB). Increases in fluorescence intensity signifies degradation of FITC-conjugated target. Enzyme activity was measured by monitoring fluorescence (FITC) (ex: 495 nm, em: 519 nm). Data are expressed as mean ± error of mean of three independent experiments. ****p<0.0001, t-test.
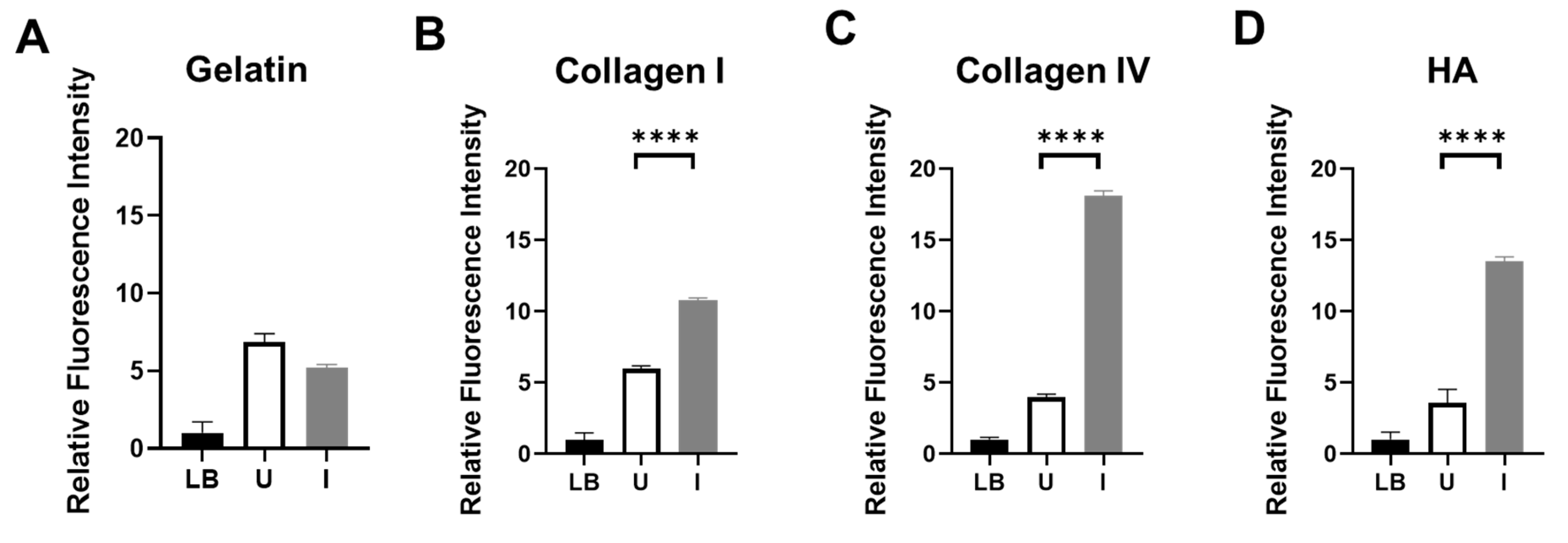
Figure 3.
BL21-TAN Depletes PDAC-Derived Collagen and HA in Serial Tumor Sections. Serial sections of KPC, BxPC3, and de-identified patient (UPN) PDAC tumors were treated overnight with BL21-TAN under uninduced or induced conditions at 37°C. Sections were then stained by trichrome to detect collagen (A) or using biotin-labeled hyaluronic acid binding protein (HABP) followed by streptavidin-FITC. (B). Trichome images were deconvoluted using ImageJ to quantify collagen content (blue staining) in randomly selected fields (10) of each tumor section (C). Fluorescence intensity was used to quantify HA content (FITC/488 channel) was measured using ImageJ and normalized to uninduced treatment (D). Data are expressed as mean ± error of mean. **p<0.01; ***p<0.001; ****p<0.0001, t-test. Scale bar = 20 um.
Figure 3.
BL21-TAN Depletes PDAC-Derived Collagen and HA in Serial Tumor Sections. Serial sections of KPC, BxPC3, and de-identified patient (UPN) PDAC tumors were treated overnight with BL21-TAN under uninduced or induced conditions at 37°C. Sections were then stained by trichrome to detect collagen (A) or using biotin-labeled hyaluronic acid binding protein (HABP) followed by streptavidin-FITC. (B). Trichome images were deconvoluted using ImageJ to quantify collagen content (blue staining) in randomly selected fields (10) of each tumor section (C). Fluorescence intensity was used to quantify HA content (FITC/488 channel) was measured using ImageJ and normalized to uninduced treatment (D). Data are expressed as mean ± error of mean. **p<0.01; ***p<0.001; ****p<0.0001, t-test. Scale bar = 20 um.
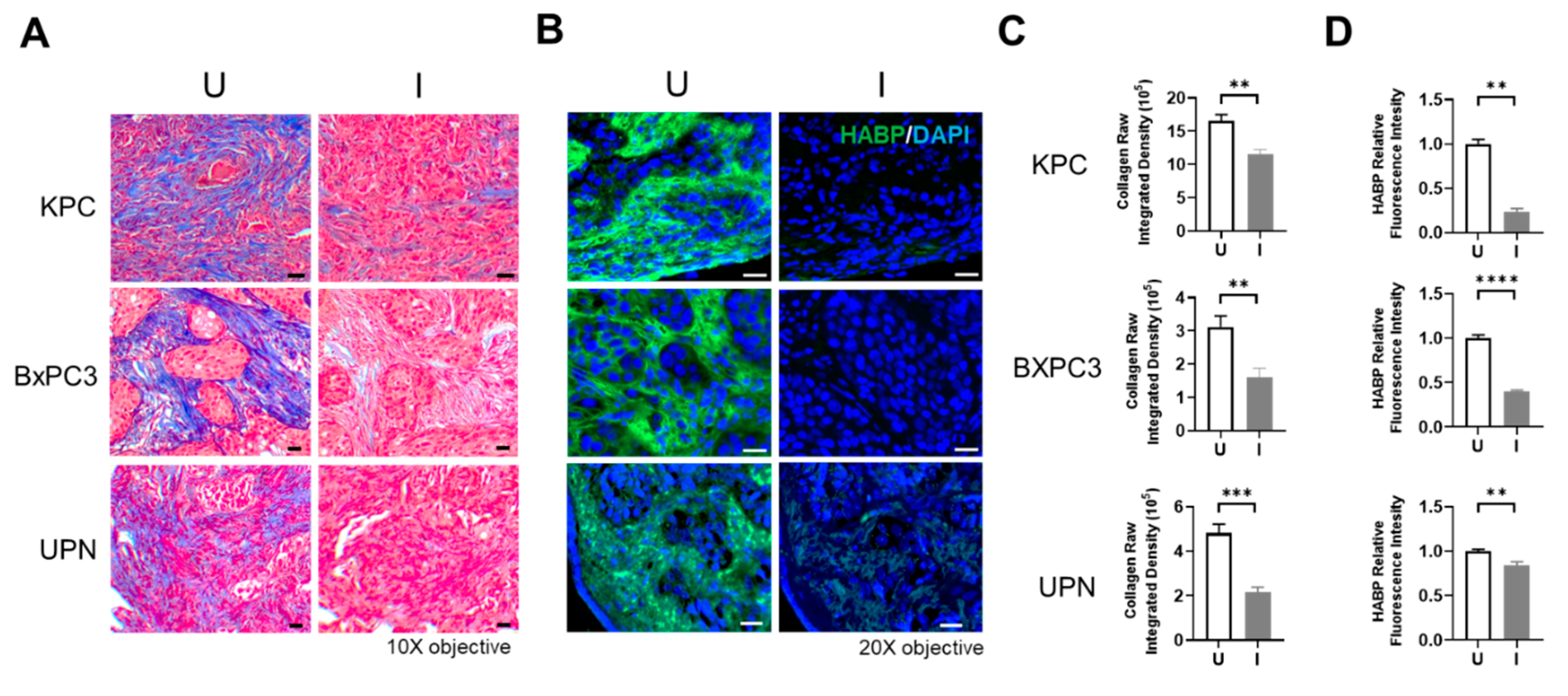
Figure 4.
Intravenously administered BL21-TAN colonizes PDAC tumors and expresses both ColH and HylB. Mice bearing subcutaneous KPC or BxPC3 tumors (6-8mm diameter) were intravenously injected with 5x107 colony-forming units (CFUs) of BL21-TAN for three consecutive days. Twenty-four hours following the final injection, mice were either administered PBS (uninduced, U) or 40 mg L-arabinose (induced, I) intraperitoneally. Tumors were collected 48h post-induction and sections were evaluated for BL21-TAN colonization and enzyme expression by immunofluorescence using antibodies specific to BL21 E.coli, Myc-tag (ColH) and His-tag (HylB). Objective:100X oil. Scale bars = 10 um.
Figure 4.
Intravenously administered BL21-TAN colonizes PDAC tumors and expresses both ColH and HylB. Mice bearing subcutaneous KPC or BxPC3 tumors (6-8mm diameter) were intravenously injected with 5x107 colony-forming units (CFUs) of BL21-TAN for three consecutive days. Twenty-four hours following the final injection, mice were either administered PBS (uninduced, U) or 40 mg L-arabinose (induced, I) intraperitoneally. Tumors were collected 48h post-induction and sections were evaluated for BL21-TAN colonization and enzyme expression by immunofluorescence using antibodies specific to BL21 E.coli, Myc-tag (ColH) and His-tag (HylB). Objective:100X oil. Scale bars = 10 um.
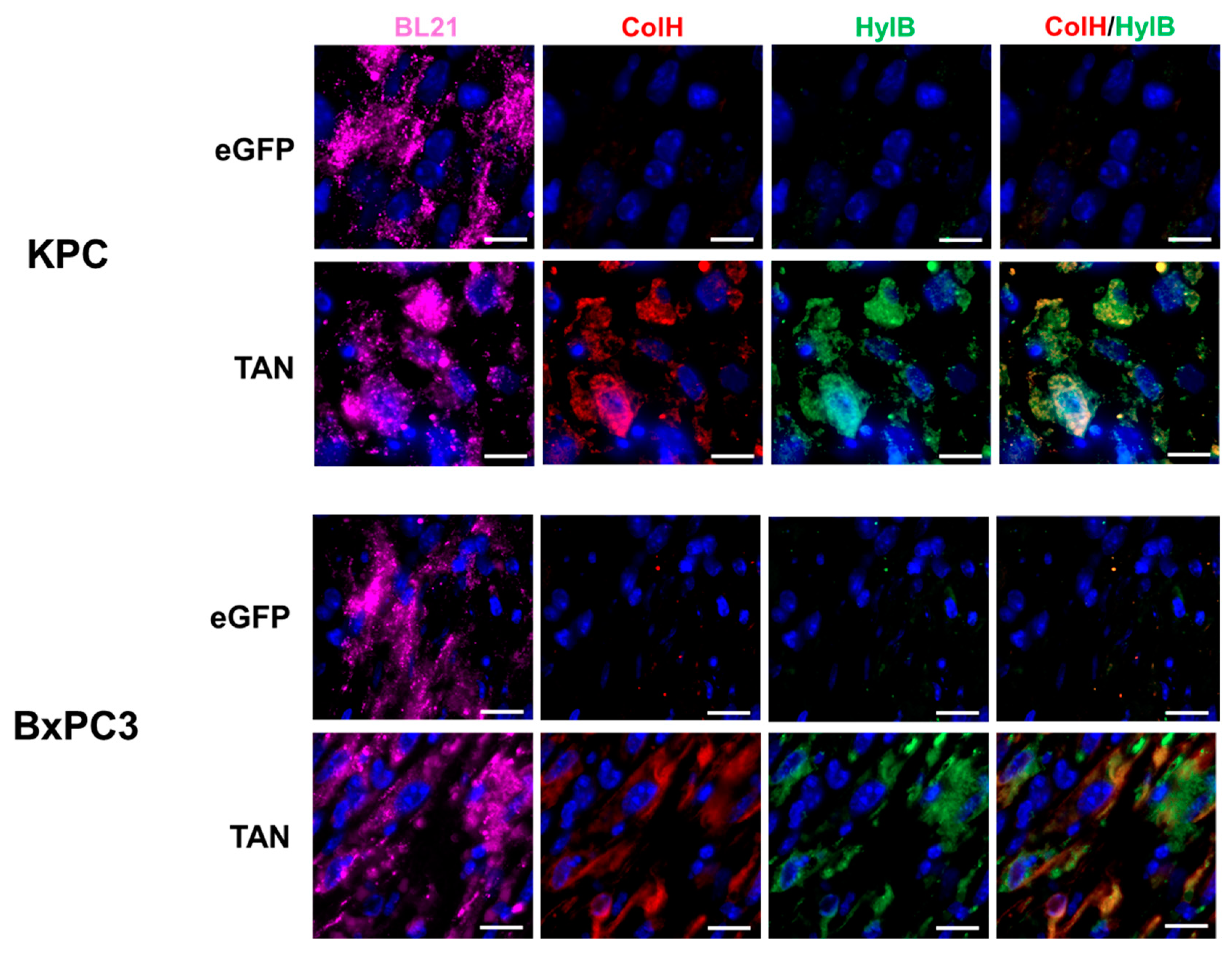
Figure 5.
In Vivo Collagen and HA Depletion by BL21-TAN. NSG mice with subcutaneous BxPC3 xenografts (6-8mm diameter) were intravenously injected with 5x107 colony-forming units (CFUs) of BL21-eGFP control or BL21-TAN for three consecutive days by intravenous route. Twenty-four hours following the final injection, mice given BL21-TAN were administered 40 mg L-arabinose intraperitoneally. Tumors (n=8) were collected 48h post-induction and serial sections were evaluated for collagen using trichrome staining (A) and HA using HABP staining (B). Regions (box) from each image were magnified for greater resolution of collagen and HA (inset). Random fields (n=15, 10X objective) from each treatment group were used for deconvolution analysis to quantify collagen and HA content in multiple tumors (C). Data are expressed as mean ± error of mean. ****p<0.0001, t-test. Scale bars = 1 mm. .
Figure 5.
In Vivo Collagen and HA Depletion by BL21-TAN. NSG mice with subcutaneous BxPC3 xenografts (6-8mm diameter) were intravenously injected with 5x107 colony-forming units (CFUs) of BL21-eGFP control or BL21-TAN for three consecutive days by intravenous route. Twenty-four hours following the final injection, mice given BL21-TAN were administered 40 mg L-arabinose intraperitoneally. Tumors (n=8) were collected 48h post-induction and serial sections were evaluated for collagen using trichrome staining (A) and HA using HABP staining (B). Regions (box) from each image were magnified for greater resolution of collagen and HA (inset). Random fields (n=15, 10X objective) from each treatment group were used for deconvolution analysis to quantify collagen and HA content in multiple tumors (C). Data are expressed as mean ± error of mean. ****p<0.0001, t-test. Scale bars = 1 mm. .
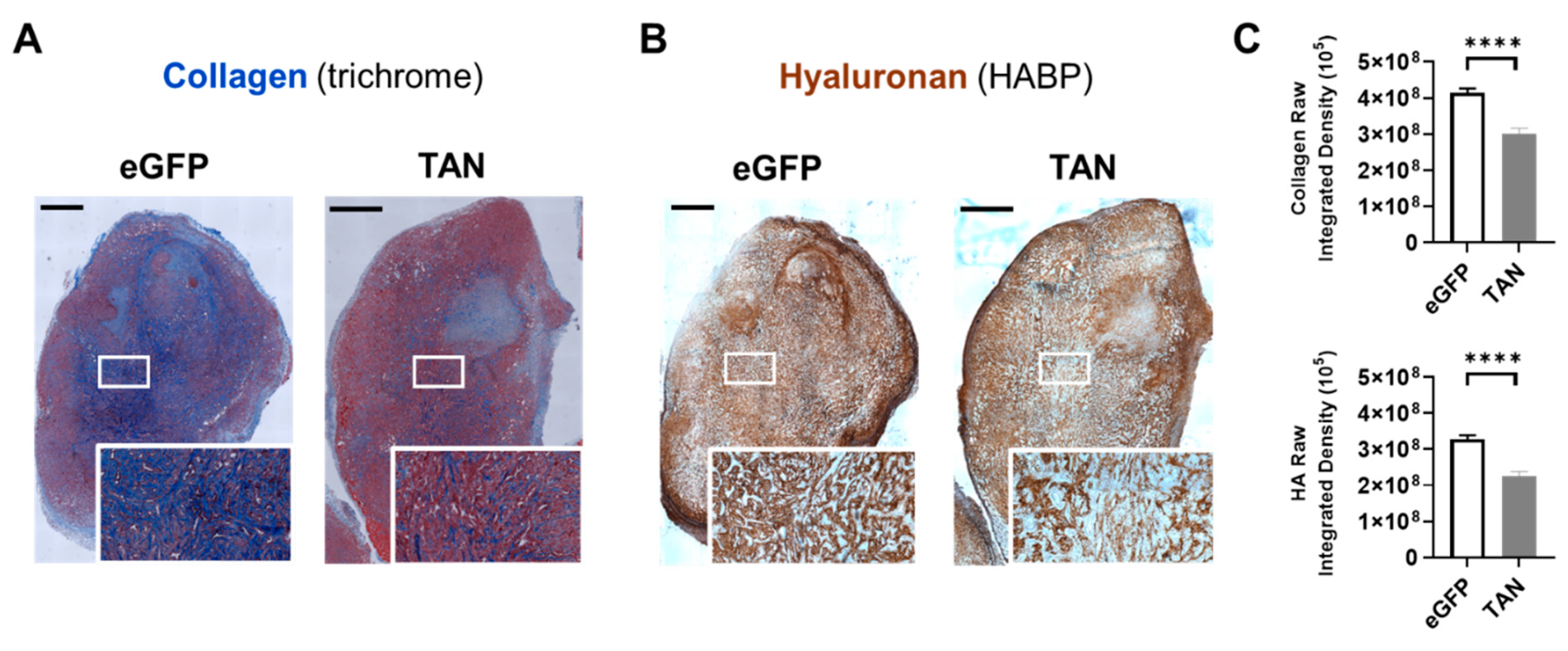
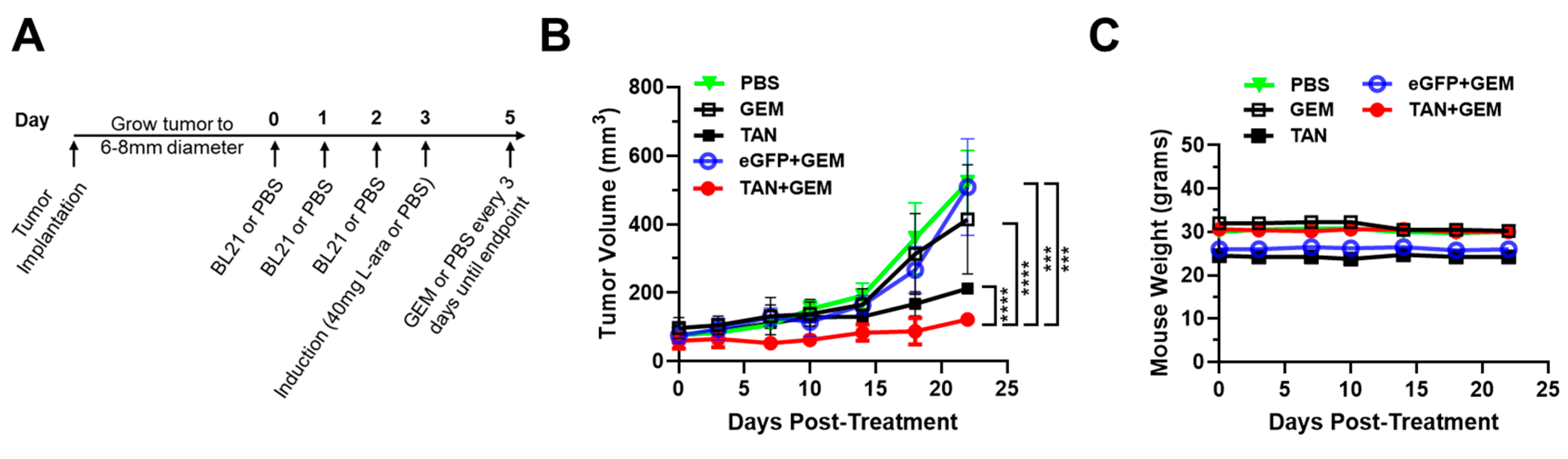
Figure 6.
BL21-TAN Pre-Treatment of PDAC Tumors Enhances Efficacy of Gemcitabine. (A) NSG mice with subcutaneous BxPC3 xenografts (6-8mm diameter) were injected with 5x107 colony-forming units (CFUs) of BL21-eGFP control or BL21-TAN for three consecutive days by intravenous route. 24 hours following the final injection, mice were administered 40 mg L-arabinose (induction) or PBS control intraperitoneally. Gemcitabine (GEM) or vehicle (PBS) were injected intraperitoneally 48 hours after induction and maintenance doses were given every 3 days thereafter. Tumor growth (B) and mouse weights (C) were measured over time until control groups required euthanization. Data are expressed as mean ± error of mean. ***p<0.001; ****p<0.0001, 2way ANOVA.
Figure 6.
BL21-TAN Pre-Treatment of PDAC Tumors Enhances Efficacy of Gemcitabine. (A) NSG mice with subcutaneous BxPC3 xenografts (6-8mm diameter) were injected with 5x107 colony-forming units (CFUs) of BL21-eGFP control or BL21-TAN for three consecutive days by intravenous route. 24 hours following the final injection, mice were administered 40 mg L-arabinose (induction) or PBS control intraperitoneally. Gemcitabine (GEM) or vehicle (PBS) were injected intraperitoneally 48 hours after induction and maintenance doses were given every 3 days thereafter. Tumor growth (B) and mouse weights (C) were measured over time until control groups required euthanization. Data are expressed as mean ± error of mean. ***p<0.001; ****p<0.0001, 2way ANOVA.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



