
Submitted:
30 October 2024
Posted:
31 October 2024
You are already at the latest version
Abstract
Eggplant is a vegetable grown worldwide, and due to quality standards, large amounts of biomass are generated after harvest. Biomass is considered a source of bioactive compounds with antioxidant properties. Therefore, this research aimed to evaluate microencapsulated alkaloids' bioaccessibility (BA) and antioxidant capacity from eggplant fruit biomass. Eggplant biomass was collected, and the total alkaloid content, antioxidant capacity, and alkaloid profile were determined before and after the digestion in vitro for encapsulated and non-encapsulated alkaloids. The bioaccessibility of microencapsulated alkaloids (12 % BA) increased three-fold compared to non-encapsulated (4 %BA). The antioxidant capacity of digested microcapsules measured by ORAC and TEAC assays was increased (30 and 8 µmol TE/g powder, respectively), and their bioaccessibility was higher than non-encapsulate alkaloids. Solamargine and solasonine decreased during in vitro digestion by 17 and 15 %BA, respectively. However, microencapsulation showed the protection of these alkaloids during in vitro digestion. This study demonstrates that microencapsulated alkaloids from eggplant biomass manage to protect bioactive compounds from in vitro digestion, having antioxidant potential mainly through hydrogen atom transfer. Therefore, microencapsulation represents an alternative to protect alkaloids and give added value to eggplant plant biomass.
Keywords:
Eggplant
; alkaloids
; microparticles
; bioaccessibility
; antioxidant capacity
1. Introduction
Eggplant (Solanum melongena L.) is a vegetable that belongs to the genus Solanum, considered the largest of the Solanaceae family, to which the potato (Solanum tuberosum) and tomato (Solanum lycopersicum L.) also belong [1]. In 2021, Mexico produced 125,531 t, while the state of Sinaloa contributed 95.6% of national production [2]. However, due to quality standards, an excess of agricultural biomass is produced in the fields [3,4]. Annually, according to the International Society of Horticultural Sciences (ISHS), 140 billion t of biomass of horticultural products is generated in the world, representing losses of 40% in post-harvest, and the main biomass found includes seeds, peels, leaves, roots, stems and fruits [5,6]. Much of the biomass is not used, and to reduce large amounts of biomass, it is returned to the soil, burned, or used as livestock feed [4,7]. These methods do not always turn out to be profitable and can have a negative effect on climate change; for this reason, it is considered an issue of social, economic, and environmental concern [5,6].
In eggplant cultivation, its biomass represents an important source of secondary metabolites that are present throughout the entire plant; one of the groups of compounds with important biological activity is alkaloids, which were identified in eggplant fruit, stems, leaves, roots, and flowers [8,9,10,11], some studies report solasonine and solamargine as the alkaloids synthesized in the highest proportion in eggplant [12]. Previous studies have found that alkaloids have been associated with various properties such as antioxidants, anticancer, and antiproliferative [13,14,15,16,17], prevent strokes [18], anti-inflammatory, antiepileptic, analgesic, hypolipidemic, hypotensive, and nervous system depressants [14,19,20,21]. Structurally, alkaloids are varied, and their biological activity is subject to their structure and bioaccessibility [22].
Bioaccessibility is defined as the percentage of compounds released from a food matrix that are accessible for absorption by the epithelial cells of the small intestine [23,24]. It has been shown that bioactive compounds have low bioaccessibility, which can hinder their biological activity. In this sense, the evaluation of piperine alkaloid from Piper nigrum mixed in food preparation showed a bioaccessibility of 60%. In terms of content, after digestion, the piperine alkaloid decreased from 6.5 to 3.9 µg/g [25]. On the other hand, Pasli, et al. [26] reported that a simulated digestion decreased the total phenolic and flavonoid content of eggplant extracts and also a reduction in the antioxidant capacity. Therefore, there is a need to protect these compounds from degradation during the digestive process. One of the most used strategies to enhance the bioaccessibility of bioactive compounds such as alkaloids is the microencapsulation process; this technique also allows these to be released at specific sites in a controlled manner and under certain conditions [27]. Spray drying has become a widely used method for encapsulating compounds. The leading encapsulating agent is maltodextrin (a water-soluble biopolymer), which is used to protect different bioactive compounds, just like Srinivasan and Shanmughasundaram [28], microencapsulated the alkaloid vasicine, derived from Adhatoda vasica Nees., by spray-drying, using maltodextrin in various proportions and obtained an encapsulation efficiency in a range of 69 to 84 %, as well as alkaloid retention of 69 %.
There is little or no information about the effect of gastrointestinal digestion on microencapsulated eggplant biomass alkaloids; for this reason, the objective of this study was to evaluate the bioaccessibility and antioxidant capacity of microencapsulated alkaloids from the biomass of the eggplant plant (Solanum melongena L.).
2. Materials and Methods
2.1. Biomass Collection
The eggplant plants (Solanum melongena L.) were collected in June 2022, 15 days after the last harvest (open field), from a farm in Villa Juárez, Navolato, Sinaloa, Mexico. The fruit was washed with water and rinsed in a chlorinated solution of 50 ppm; finally, samples were dried at 20 °C, then were freeze-dried (−49 °C and 0.079 bars), ground, and stored in sealed acetate bags at -20 °C.
2.2. Alkaloid Extraction
The extract rich in alkaloids was obtained using the QuEChERS method with some modifications, as reported by Lehotay [29]. For this, 3 g of dried sample was homogenized with 12 mL of distilled water and 15 mL of 1 % acidified acetonitrile, sonicated for 10 min, then 4 g of magnesium sulfate and 1 g of sodium acetate were added, and centrifuged at 4,000 rpm for 5 min at 4 °C. The supernatant was purified with a C18 cartridge, dried in a SyncorePlus, and then stored at -20 °C.
2.3. Totals Alkaloids Content
The method based on the alkaloid reaction with bromocresol green (BCG) was used with some modifications [30]. To quantify the alkaloid content were weighted 63 mg of the dry extract obtained by the QUECHERS method and 0.5 g microcapsule (1 mL of water was added to release the alkaloids), 2.5 mL of chloroform was added to each sample, the mixture was placed in a separation funnel, and 2.5 mL of phosphate buffer pH 7.4 and 2.5 mL of BCG were added. The organic phase was collected and quantified using a spectrophotometer at 470 nm. The results were calculated as mg solasodine equivalent per g dry extract (mg ESS/g DE) and as mg solasodine equivalent per g microencapsulated powder (mg ESS/g powder).
2.4. Antioxidant Capacity Assay
The antioxidant capacity of the alkaloid-rich extract without microencapsulation and microencapsulation was evaluated before and after in vitro gastrointestinal digestion through the Trolox equivalent antioxidant capacity (TEAC) assay described by Karadag, et al. [31], the ion reduction capacity by ferric ion reducing antioxidant power (FRAP) was also determined, following the methodology of Benzie and Strain [32], and the oxygen radical absorbance capacity (ORAC) according to Huang, et al. [33]. The results were expressed as equivalent micromoles of Trolox per g of dry extract (µmol TE/g DE) and µmol TE/g powder.
2.5. Identification and Quantification of Alkaloids by Ultra High-Resolution Liquid Chromatography/Mass Spectrometry (UPLC/MS)
The UPLC chromatographic system coupled to a Waters Xevo TQ-S mass spectrometer was used to identify and quantify the alkaloids and glycoalkaloids in the dry extract. Samples were automatically injected through a Waters Sample Manager–FTN Acquity system to an Acquity H series UPLC equipped with an Acquity UPLC BEH Phenyl 1.7 µm, 2.1 x 100 mm column. The conditions were mobile phase A (5 mM ammonium formate, pH 3.0) and phase B (acetonitrile + 0.1 % formic acid). At time 0, it started with 90 % A and 10 % B with low flow that gradually increased 0.3 mL/min and was maintained in these conditions for 5 min, then 10 % A and 90 % B; At 5.1 min, it was raised to 90 % A and 10 % B and was maintained until 8 min. Likewise, the conditions for the Waters Xevo TQ-S mass spectrometer were established through the MassLynx software. The conditions were as follows: positive electrospray ionization (ESI+), source temperature 150 °C, cone voltages were 60 to 100 V, and capillary voltages were 3.21 kV. The desolvation temperature was 400 °C, the desolvation gas flow was 650 L/h, and the collision gas flow was 0.15 mL/min. The run time was 10 min with an injection volume of 5 µL, and the MRM (multiple reaction monitoring) mode was used for analyte analysis. Retention time and transitions using MRM were used for identification, and calibration curves (solamargine and solasonine) were used for quantification to compare the area under the curve of the obtained peaks.
2.6. Alkaloid Microencapsulation
An aliquot of 50 mL of alkaloid-rich extract (concentration of 84 mg of dry extract) stock was mixed with 8 g maltodextrin (MD) ten dextrose equivalents (DE) as wall material. The mixture was homogenized on a stir plate at 600 rpm until completely dissolved. Subsequently, the mixture was fed to a Spray Dryer Yamato ADL311S. The conditions were inlet temperature 145 °C and outlet 80 °C, the atomization pressure was 0.1 MPa, the feed flow was 5 mL/min, and the airflow was 0.32 m3/min. The recovered powder (alkaloids microencapsulate) was weighed to obtain the yield of the process and stored at 20 °C [34].
2.7. Physical Characterization of the Microcapsules
2.7.1. Morphology
The morphology of the microencapsulate was analyzed using scanning electron microscopy (SEM) model EVO-50, Carl Zeiss brand. The powder was coated with gold in a DESK II model ionizer, Denton Vacuum brand, operating with a voltage of 10 kV and under high vacuum conditions. The size was determined using ImageJ Software.
2.7.2. Moisture
The moisture percentage in the microcapsules was determined using the gravimetric method AOAC 925.09 [35].
2.7.3. Process Yield
The encapsulation yield of the process was calculated using a gravimetric technique, which is the relationship between the weight of powder after drying and the total solids at the beginning of the feed. It was reported as a percentage [36].
2.7.4. Encapsulation Efficiency (EE)
These were calculated by the content of total alkaloids in the encapsulated powder using the value of the encapsulated extract obtained concerning the theoretical encapsulated extract (mg solasodine equivalents) and reported as a percentage.
2.8. Bioaccessibility Assay
The percentage of bioaccessibility (% BA) of the alkaloids in the extract and the microparticles was determined following the INFOGEST method by Brodkorb, et al. [37] with some modifications. This in vitro model simulates the digestion of products when they pass through the mouth, stomach, and small intestine, imitating the chemical composition, pH of the digestive fluids, temperatures, and transit times. To start the process, 8 mg of dry extract rich in alkaloids and 0.5 g of microencapsulated extract were used, and 2 mL of the simulated oral phase was added and incubated for 2 min at 37 °C. After that, 2 mL of the gastric phase was added, the pH was adjusted to 3, and the mixture was incubated at 37 °C for 2 h. Finally, 4 mL of the intestinal phase was added proportionately to the total liquid until the gastric phase. The pH was adjusted to 7 and incubated for 2 h at 37 °C. After this process, absolute methanol was added in a 1:1 ratio. The samples were centrifuged at 10,000 rpm for 15 min at a temperature of 4 °C. The supernatant was recovered and stored at −20 °C. The in vitro bioaccessibility of extract and microencapsulated was calculated with the following equation:
The final concentration is the result obtained through some assay used to evaluate the end of the intestinal phase, and the initial concentration is the result obtained from undigested samples.
2.9. Statistical Analysis
It was a completely randomized experimental design with one factor and three replications. The results obtained were analyzed using an analysis of variance (ANOVA), using Tukey's mean comparison test (p≤0.05), with a significance value of 5 % in case of significant differences. The statistical package used to analyze the results was Minitab version 2019.
3. Results
3.1. Total Alkaloid Content of Eggplant Biomass Extract
The content of total alkaloids in the fruit biomass extract was quantified before and after the in vitro gastrointestinal simulation. The total content of alkaloids is higher in the undigested extract (173 mg ESS/g), but after the digestion process, a significant reduction was shown (7 mg ESS/g); these changes caused a bioaccessibility of 4% (Table 1).
3.2. Antioxidant Capacity of Eggplant Alkaloids Fruit Extract
The antioxidant capacity values of the alkaloid-rich extract from the fruit, measured by ORAC, TEAC, and FRAP, before and after the in vitro digestive simulation ranged from 800 to 1778 µmol TE/g DE (Table 1). Our results showed a higher antioxidant capacity in the undigested extract measured by ORAC and TEAC assay. In vitro digestion significantly reduced the antioxidant capacity of the alkaloids in each method evaluated. In addition, the percentage of bioaccessibility (%BA), determined by TEAC, FRAP, and ORAC, ranged from 6 to 31%, with the highest bioaccessibility by the ORAC method at 31%.
3.3. Identification and Quantification of Alkaloids by Ultra High-Resolution Liquid Chromatography Coupled to Mass Spectrometry (UPLC/MS) in Extract
The alkaloids and glycoalkaloids of eggplant fruit were extracted using the QuEChERS method. The alkaloid extract was quantitatively evaluated before and after the in vitro digestive simulation using commercial standards (solamargine and solasonine) previously reported in the genus Solanum. The extract from the eggplant biomass showed a higher concentration of solamargine with values of 2485 ng/g, while solasonine was 1724 ng/g before the in vitro digestion process. After digestion, the concentration of these glycoalkaloids was reduced by up to 80% (Table 2).
3.4. Physical Characterization of Microcapsules
3.4.1. Morphology
One of the essential characteristics of the encapsulation process is its size and shape. In this sense, the particles obtained had a diameter that varied from 1 to 14 µm; the shapes were spherical and irregular, with depressions on the surface (Figure 1).
3.4.2. Moisture and Yield
The moisture of the encapsulated products was low (3.4 %); according to NOM-183-SCFI-2012, powdered dairy products must have a maximum of 4 % moisture to maintain their stability. The yield in this research was below (45 %), which was considered favorable (>50 %) [38].
3.4.3. Encapsulation Efficiency
This variable is defined as the amount of alkaloids that managed to be trapped within the wall material (maltodextrin)[39]. In this research, the encapsulation efficiency was 80 %.
3.5. Total Alkaloid Content of Encapsulated Eggplant Alkaloids Fruit
The results of the total alkaloid content of the digested and undigested microcapsule are shown in Table 3, where a decrease from 1.59 to 0.185 mg ESS/g of powder was observed. The alkaloid content decreased after in vitro digestive simulation; however, regarding bioaccessibility percentage, it was higher (12%) than the non-encapsulated extract (4%).
3.6. Antioxidant Capacity of Encapsulated Eggplant Alkaloids Fruit
The antioxidant capacity for the ORAC assay presented the highest value in undigested microcapsules (Table 3). Meanwhile, in TEAC and FRAP, the values were lower. After the in vitro digestive simulation, the antioxidant capacity of the microcapsules increased almost twice by the ORAC (30 µmol TE/g) and TEAC (8 µmol TE/g) assays; FRAP, on the contrary, decreased. The bioaccessibility of the antioxidant capacity of the microcapsules loaded with alkaloid-rich extract evaluated by the three methods ranged from 70% to values greater than 100%, which were higher than those presented by the unencapsulated extract (6 to 31% BA).
3.7. Identification and Quantification of Alkaloids by Ultra High-Resolution Liquid Chromatography Coupled to Mass Spectrometry (UPLC/MS) of Encapsulated Eggplant Alkaloids Fruit
The compounds that were quantified in undigested and digested encapsulated eggplant alkaloid extract were solamargine and solasonine. Undigested microcapsules had a higher solamargine content than solasonine (Table 4). After the in vitro digestive simulation, an increase in the content of digested microencapsulated glycoalkaloids was observed compared to the undigested microcapsule; the values ranged between 9.743 and 12.74 ng/g.
4. Discussion
In plants, alkaloids act as a defense mechanism against biotic and abiotic stress, such as attacks by pests, herbivores, pathogens, UV radiation, drought, etc. Different types can be found, such as tropane alkaloids, pyrrolizidine alkaloids, indolic alkaloids, and steroidal alkaloids. In this sense, research has indicated that the solanum species has many biologically active alkaloids[40].
Authors various reported total alkaloids, however, are expressed as alkaloids different, such as Păltinean, et al. [41], who found 8.6 mg of chelidonine equivalents per gram in an alkaloid-rich extract of Fumaria species in eggplant biomass was reported 26 mg of atropine equivalents per gram [8]. Our results were expressed as mg of solasodine per gram DE with a value of 173. These differences could be attributed mainly to how the results are described and the species, type of extract, and extraction method. On the other hand, after the in vitro digestion process in extracts obtained from the eggplant fruit biomass, the total alkaloid content is almost entirely reduced, with a bioaccessibility of 4%; this behavior was previously reported regarding the content of piperine mixed in food after the in vitro digestive simulation indicated a decrease due to pH variations [25]. This effect could be due to the conditions of the different digestion phases, mainly pH [43], which could be causing physicochemical transformations by oxidation or interactions with other groups of compounds [44].
The TEAC assay measures the antioxidant capacity of hydrophilic and lipophilic compounds, offering a perspective on the group of compounds able to interact with the radical [45]. In this research, a decrease in the digested extract was shown. However, it has greater antioxidant capacity than what was reported in eggplant fruits in the commercial stage (7 µmol TE/g) by Elizalde-Romero, et al. [46]; these differences are attributed to the stage of maturity and the type of compounds evaluated because the extraction method that was carried out removes the majority of secondary metabolites such as phenols, saponins, flavonoids, and some alkaloids. This allows us to show that the alkaloid-rich extract of eggplant biomass has antioxidant properties, as do the alkaloids of other species of the Solanum genus, such as Solanum macrocarpon L. and Solanum nigrum L.[47].
Regarding the effect of in vitro digestion, our results showed a lower antioxidant capacity in the digested extract compared to the undigested, which indicates that it is less bioaccessible (12%); this behavior was similar to Solanum nigrum evaluated by Moyo, et al. [48] reported 650 µmol TE/g of undigested extract and 379 µmol ET/g of digested extract. Similarly, the fruit of Solanum lycopersicum, the undigested extract, showed greater antioxidant capacity than the digested extract, 713 µmol TE/g, and 430 µmol TE/g, respectively [49]. These results were superior to those reported in this research; however, the behavior of the extracts during the assay was similar, demonstrating the negative impact of in vitro digestion caused by the gastrointestinal environment.
Regarding the results by FRAP, which consist of the reduction of the ferric ion, it was observed a greater antioxidant capacity obtained in the undigested extract of the biomass of the eggplant fruit; these results were higher than those of the eggplant fruit collected after harvest with values of 107 µmol TE/g [8]; likewise, in different varieties of eggplant, values of 0.82 to 8.11 µmol TE/g were reported [50]. These differences are attributed to the group of compounds that were extracted and evaluated, as well as to the eggplant's variety, region, and stage of maturity. About in vitro digestive simulation, it was found that the digested fruit extract decreased its antioxidant capacity just like extract from the fruit of Solanum lycopersicum reported 0.477 µmol TE/g of undigested extract to 0.276 µmol TE/g of digested [49]. These behaviors are attributable to the physicochemical and structural characteristics of the compounds affected by the digestive simulation. On the other hand, the antioxidant capacity was higher in this research and is related to the ability of alkaloids to reduce metal ions [51].
The ORAC method uses a radical generator to analyze the antioxidant capacity of compounds based on the transfer of hydrogen atoms [52,53]. The inhibition of the peroxyl radical in this study was more significant in the undigested extract (1778 µmol TE/g). Compared to that reported in eggplant fruit biomass, its antioxidant capacity is almost 3 times higher (547 µmol TE/g) [8]. The differences could be due to the type of extract used for the assay. Similarly, our data were higher than the benzylisoquinoline alkaloids in Plumula nelumbinis with a value of 0.00553 µmol TE/g [54], the same as the alkaloids present in Catharanthus roseus with values of 185 µmol TE/g [55] and 56 µmol TE/g [56]. The previously reported data were lower than this research's, possibly due to the biosynthetic origin and the type of alkaloids specific to each species and genus [57]. After in vitro digestive simulation, a lower capacity to transfer hydrogen atoms was observed; however, our data were superior to the hydrophilic compounds of Solanum lycopersicum with values of 310 µmol TE/g of undigested extract and 270 µmol TE/g of digested extract [58]. Solanum nigrum leaves obtained values of 299 µmol TE/g of undigested extract and 620 µmol TE/g of digested extract [48]. The authors maintain that these differences could be related to the availability of hydroxyl groups of the compounds in the extracts and their physicochemical properties.
In general, a reduction in the antioxidant capacity of the alkaloid extract from eggplant biomass was observed in the three assays after in vitro digestion. In addition, a low bioaccessibility was obtained, which varied from 6 to 31% depending on the assay.
In eggplant fruits, the main glycoalkaloids reported in S. melongena are solasonine and solamargine, both glycosides of solasodine [57]. In previous research, the content of solasonine and solamargine in eggplant fruit was 0.062 ng/g and 0.373 ng/g, respectively [12]; these values were lower than what was found in this study, which may be caused by the collection time, type of species, and crop conditions [9]. Currently, there are few reports on the bioaccessibility of alkaloids of the Solanum genus, so this research is one of the few to report the effect of simulated gastrointestinal digestion on solamargine and solasonine content. In previous studies, it was reported that the alkaloid piperine (Piper nigrum) decreased by 60% after the digestion process [25]. Bioactive compounds tend to undergo structural changes such as isomerization, attributed to digestion conditions, the action of intestinal enzymes, the presence of chemical elements such as transition metals, and the presence of oxygen [43]. Regarding the decrease in glycoalkaloids, the possible hydrolysis of glycosylated molecules is considered, caused by the enzymes and the pH of the different digestive phases [59]. Besides, the differences observed between the assays could be explained by the ability of the compounds to transfer electrons [51], reduce metal ions [41], and transfer hydrogen atoms [53].
In the next stage of the study, the alkaloid-rich extract of the fruit of the eggplant biomass was microencapsulated with maltodextrin. The size of the microcapsules obtained was considered relatively homogeneous and uniform and not very narrow, which is regarded as favorable to maintaining the consistency of the microencapsulation [60]. The observed roughness is a common characteristic of microcapsules made with maltodextrin and spray drying due to rapid evaporation of moisture and cooling [61]. Maltodextrin microcapsules with anthocyanins from the peel of Solanum melongena were made with an inlet temperature of 180 °C, forming smooth, dented, and irregular microcapsules due to the rapid loss of moisture due to high inlet temperature [62]. On the other hand, agglomerations were reported in maltodextrin microcapsules with an extract of the alkaloid vasicine from Adhatoda vasica Nees prepared with an inlet temperature of 110 °C [28]. Under the same conditions as our study, sizes of 12 µm and a spherical morphology with depressions were obtained in maltodextrin microcapsules loaded with oregano phenolic compounds [34]. The differences between our study and what was found in the literature are due to the variations in the inlet temperature [38].
Moisture is an indicator of quality and stability. Low moisture percentages prevent powder hardening, guarantee a long shelf life, and protect it from microbiological contamination during storage [63]. Our data is similar to that of Arrazola, Herazo and Alvis [62], who reported 3.4% humidity in anthocyanin microparticles from eggplant peel using maltodextrin as an encapsulating agent. Besides, maltodextrin microcapsules with alkaloids from the Adhatoda vasica plant with an inlet temperature of 110 °C had a moisture of 5.1% [28].
The yield of the powder obtained indicates the efficiency of the spray drying process. In this research, the yield was lower than that obtained in maltodextrin microcapsules (20 DE) with purple potato compounds (Solanum tuberosum L.); the process was made at an inlet temperature of 130 °C with a feed of 100 g, generating a yield of 58% [64]. Our data were slightly lower than those reported by Sarabandi, Jafari, Mahoonak, and Mohammadi [61], who worked with eggplant peel metabolites microencapsulated with maltodextrin (18-20 DE) with an air inlet of 140 °C and a feed of 300 mL, resulting in a yield of 52%. This result could be due to the quantity and low viscosity of the solution fed, as it is related to greater water elimination and less adhesion of the encapsulated microparticles on the walls of the dryer. In this sense, the value considered favorable is greater than 50%, and the main factors that influence this percentage are the viscosity of the wall material and the content of fed solids, so the differences found are attributed to the content of the feeding solution.
The EE is considered an essential characteristic during the encapsulation process, as it is defined as the amount of material to be encapsulated that was encapsulated within a wall material [39]. This research was similar to microcapsules of alkaloid-rich extract of Adhatoda vasica Nees leaves prepared with maltodextrin at an inlet temperature of 80 °C, obtaining 84 % EE [28]. Contrary to this, phenolic compounds from the fruit of Malphigia emarginata DC fruit were microencapsulated with maltodextrin, using an inlet temperature of 170 °C, obtaining 69 % EE [65]. These differences may be due to high temperatures during the spray drying process, which can cause the loss of volatile active compounds, resulting in low encapsulation efficiency [66].
In this same stage of encapsulated alkaloids, the total content of undigested and digested alkaloids was evaluated. After in vitro digestion simulation, the total content of microencapsulated alkaloids decreased; however, bioaccessibility was higher in the encapsulated extract than in the non-encapsulated extract. These results agree with those previously reported, which claim that the microencapsulation of bioactive compounds increases the percentage of bioaccessibility concerning the non-encapsulated sample when they pass through the three phases of the digestive system because the encapsulating agent manages to protect them from the conditions of the gastrointestinal simulation phases, enzymatic and pH variation [44,67,68].
Antioxidant compounds are sensitive to high temperatures, light, and pH; for that reason, encapsulation techniques have been carried out to protect them and improve their functionality[69]. In this sense, the microcapsules were evaluated using the TEAC test, where a greater antioxidant capacity was found in the digested microencapsulate (8 µmol TE/g powder), coinciding with what was reported in MD microcapsules from the fruit of Eugenia stipitata with values of 136 µmol TE/g and 253 µmol TE/g powder undigested and digested, respectively [70]. This may be due to the deprotonation of the hydroxyl groups of the bioactive compounds at high pH [26].
The reduction of metal ions was more significant in the undigested microcapsules (3.4 µmol TE/g powder), similar to the MD microcapsules of bioactive compounds from the fruit of Eugenia stipitate; the authors reported that after in vitro digestion simulation, the antioxidant capacity decreases slightly[70]. This is attributed to the ability to chelate metals due to the sample's pH and the method's optimal pH and structural modifications due to enzymatic hydrolysis, which causes the breakdown of glycosidic bonds [44].
The inhibition of peroxyl radicals by microencapsulated alkaloids was more significant after in vitro digestion simulation, consistent with Tomé-Sánchez, et al. [71], who reported that after digestion process a two-fold increase in antioxidant capacity, which has been attributed to possible degradation of polymer during its passage through the different digestive phases, achieving the total release of the compounds in the intestinal phase, causing chemical transformations in the structures of the metabolites due to the effect of digestive enzymes that cause deprotonation [44].
The antioxidant capacity of the alkaloids microencapsulated by the different assays after digestion in vitro was increased, obtaining a bioaccessibility above 50 %.
Previous studies have indicated that solamargine and solasonine are the main chemical compounds in Solanum species that also have beneficial health properties [72]. In this sense, we can observe the identification and quantification of these compounds in both the undigested and digested microcapsules. In the digested microcapsules, the content of solamargine and solasonine increased by more than 100%. The increase observed after in vitro digestive simulation is consistent with previous investigations on ergot alkaloids, where some compounds' increases are attributed to bidirectional epimerization caused by intestinal enzymes [73]. Likewise, other compounds' isomerization has been described, and they speculate that it is due to temperature and prolonged exposure in the small intestine [74]. In the same way, Vronen [75] described the chemical hydrolysis of glycoalkaloids in potatoes. They mentioned that it is caused by time, temperature, and acid concentration, allowing the formation of compounds β and γ as new hydrolysis products.
5. Conclusions
The microencapsulation process with maltodextrin using the spray drying method managed to trap the alkaloids from the eggplant biomass, which allowed them to be protected during in vitro gastrointestinal digestion, increasing their bioaccessibility three-fold, while antioxidant capacity increased more than 50 %BA; likewise, solamargine and solasonine glycoalkaloids in microcapsule increase during digestion in vitro. However, it is necessary to continue exploring different wall materials and encapsulation methods to obtain encapsulates with more resistance to digestion and for the alkaloids to be more bioaccessible.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, B.G.G.-S., L.A.C.-A, J.B.H, N.L.-L and E.P.G.-G; methodology, B.G.G.-S., M.G.-C., P.B.-B and L.A.C.-A.; software, M.G.C. and P.B.-B; formal analysis, B.G.G.-S., L.A.C.-A, J.B.H, N.L.-L and E.P.G.-G.; investigation, B.G.G.-S, and L.A.C.-A.; writing—original draft preparation, B.G.G.-S.; writing—review and editing, B.G.G.-S., L.A.C.-A, J.B.H, and N.L.-L.; visualization, B.G.G.-S., L.A.C.-A.; supervision, L.A.C.-A.; project administration, L.A.C.-A, J.B.H, N.L.-L and E.P.G.-G.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author/s.
Acknowledgments
B.G.G.-S. Thanks, CONAHCYT, for the scholarship granted for its graduate studies.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Agoreyo, B.O.; Obansa, E.S.; Obanor, E.O. Comparative nutritional and phytochemical analyses of two varieties of Solanum melongena. Sci. World J. 2012, 7, 5-8.
- SIAP. Panorama agroalimentario 2022. Servicio de información agroalimentaria y pesquera 2022, 42-43.
- Mauro, R.P.; Agnello, M.; Rizzo, V.; Graziani, G.; Fogliano, V.; Leonardi, C.; Giuffrida, F. Valorización de residuos de campo de berenjena como fuente de fitoquímicos. Sci Horticulturae 2020. [CrossRef]
- Maurya, R.; Bharti, C.; Singh, T.D.; Pratap, V. Crop Residue Management for Sustainable Agriculture. Int J Curr Microbiol Appl Sci 2020, 9, 2020. [CrossRef]
- Lobo, M.G.; Dorta, E. Utilization and management of horticultural waste. In Postharvest Technology of Perishable Horticultural Commodities; Elsevier: 2019; pp. 639-666.
- Pattnaik, M.; Pandey, P.; Martin, G.J.O.; Mishra, H.N.; Ashokkumar, M. Innovative technologies for extraction and microencapsulation of bioactives from plant-based food waste and their applications in functional food development. Foods 2021, 10, 279. [CrossRef]
- Kazemi, M.; Khodaiyan, F.; Hosseini, S.S.; Najari, Z. An integrated valorization of industrial waste of eggplant: Simultaneous recovery of pectin, phenolics and sequential production of pullulan. J. Waste Manag. 2019, 100, 101-111. [CrossRef]
- Contreras-Angulo, L.A.; Moreno-Ulloa, A.; Carballo-Castañeda, R.A.; León-Felix, J.; Romero-Quintana, J.G.; Aguilar-Medina, M.; Ramos-Payán, R.; Heredia, J.B. Metabolomic analysis of phytochemical compounds from agricultural residues of eggplant (Solanum melongena L.). Molecules 2022, 27, 7013. [CrossRef]
- Distl, M.; Wink, M. Identification and quantification of steroidal alkaloids from wild tuber-bearing Solanum species by HPLC and LC-ESI-MS. Potato Res. 2009, 52, 79-104. [CrossRef]
- Matias, L.J.; Mercadante-Simões, M.O.; Royo, V.A.; Ribeiro, L.M.; Santos, A.C.; Fonseca, J. Structure and histochemistry of medicinal species of Solanum. Rev. bras. farmacogn. 2016, 26, 147-160. [CrossRef]
- Morillo, M.; Rojas, J.; Lequart, V.; Lamarti, A.; Martin, P. Natural and synthetic derivatives of the steroidal glycoalkaloids of Solanum genus and biological activity. Nat Prod Res 2020. [CrossRef]
- Lelario, F.; De Maria, S.; Rivelli, A.R.; Russo, D.; Milella, L.; Bufo, S.A.; Scrano, L. A complete survey of glycoalkaloids using LC-FTICR-MS and IRMPD in a commercial variety and a local landrace of eggplant (Solanum melongena L.) and their anticholinesterase and antioxidant activities. Toxins (Basel) 2019, 11, 230. [CrossRef]
- Chang, L.C.; Tsai, T.R.; Wang, J.J.; Lin, C.N.; Kuo, K.W. The rhamnose moiety of solamargine plays a crucial role in triggering cell death by apoptosis. Biochem. Biophys. Res. Commun. 1998, 242, 21-25. [CrossRef]
- Friedman, M. Chemistry and anticarcinogenic mechanisms of glycoalkaloids produced by eggplants, potatoes, and tomatoes. J. Agric. Food Chem. 2015, 63, 3323-3337. [CrossRef]
- Milner, S.E.; Brunton, N.P.; Jones, P.W.; O'Brien, N.M.; Collins, S.G.; Maguire, A.R. Bioactividades de glicoalcaloides y sus agliconas de especies de Solanum. J. Agric. Food Chem. 2011, 59 (8), 3454-3484. [CrossRef]
- Xie, X.; Zhu, H.; Yang, H.; Huang, W.; Wu, Y.; Wang, Y.; Luo, Y.; Wang, D.; Shao, G. Solamargine triggers hepatoma cell death through apoptosis. Oncol. Lett. 2015, 10, 168-174. [CrossRef]
- Zhao, D.K.; Zhao, Y.; Chen, S.Y.; Kennelly, E. Solanum steroidal glycoalkaloids: Structural diversity, biological activities, and biosynthesis. Natural Product Reports 2021, 38, 1423-1444. [CrossRef]
- Sharma, T.; Airao, V.; Panara, N.; Vaishnav, D.; Ranpariya, V.; Sheth, N.; Parmar, S. Solasodine protects rat brain against ischemia/reperfusion injury through its antioxidant activity. Eur. J. Pharmacol. 2014, 7. [CrossRef]
- Chauhan, K.; Sheth, N.; Ranpariya, V.; Parmar, S. Anticonvulsant activity of solasodine isolated from Solanum sisymbriifolium fruits in rodents. Pharm. Biol. 2011, 49, 194-199. [CrossRef]
- Das, M.; Barua, N. Pharmacological activities of Solanum melongena Linn.(Brinjal plant). Int. J. Green Pharm. 2013, 7. [CrossRef]
- Vohora, S.B.; Kumar, I.; Khan, M.S.Y. Effect of alkaloids of Solanum melongena on the central nervous system. J. Ethnopharmacol. 1984, 11, 331-336. [CrossRef]
- Shahidi, F.; Peng, H. Bioaccessibility and bioavailability of phenolic compounds. JFB 2018, 4, 11–68-11–68. [CrossRef]
- Shahidi, F.; Pan, Y. Influence of food matrix and food processing on the chemical interaction and bioaccessibility of dietary phytochemicals: A review. Crit. Rev. Food Sci. Nutr 2022, 62, 6421-6445. [CrossRef]
- Thakur, N.; Raigond, P.; Singh, Y.; Mishra, T.; Singh, B.; Lal, M.K.; Dutt, S. Recent updates on bioaccessibility of phytonutrients. Trends Food Sci. Technol. 2020, 97, 366-380. [CrossRef]
- Azevedo, M.; Leite, I.B.; Queiroz, C.; Fialho, E. Spiced risotto: Cooking processing and simulated in vitro digestion on curcuminoids, capsaicin and piperine. J CULIN. 2019, 17, 256-267. [CrossRef]
- Pasli, A.A.; Yavuz-Düzgün, M.; Altuntas, U.; Altin, G.; Özçelik, B.; Firatligil, E. In vitro bioaccessibility of phenolics and flavonoids in various dried vegetables, and the determination of their antioxidant capacity via different spectrophotometric assays. Food Res Int 2019, 26, 793-800.
- Marcillo-Parra, V.; Tupuna-Yerovi, D.S.; González, Z.; Ruales, J. Encapsulation of bioactive compounds from fruit and vegetable by-products for food application–A review. Trends Food Sci Tech 2021, 116, 11-23. [CrossRef]
- Srinivasan, K.; Shanmughasundaram, S. Microencapsulation of vasicine alkaloids through spray drying. IAJPR 2016. [CrossRef]
- Lehotay, S. AOAC official method 2007.01 pesticide residues in foods by acetonitrile extraction and partitioning with Magnesium Sulfate. J. AOAC Int. 2007, 90, 485-520. [CrossRef]
- Shamsa, F.; Monsef, H.; Ghamooshi, R.; Verdian-rizi, M. Spectrophotometric determination of total alkaloids in some Iranian medicinal plants. TJPS 2008, 32, 17-20. [CrossRef]
- Karadag, A.; Ozcelik, B.; Saner, S. Review of methods to determine antioxidant capacities. Food Anal. Methods 2009, 2, 41-60. [CrossRef]
- Benzie, I.F.F.; Strain, J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal. Biochem. 1996, 239, 70-76. [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437-4444. [CrossRef]
- Bernal-Millán, M.J.; Gutiérrez-Grijalva, E.P.; Contreras-Angulo, L.; Muy-Rangel, M.D.; López-Martínez, L.X.; Heredia, J.B. Spray-dried microencapsulation of oregano (Lippia graveolens) polyphenols with maltodextrin enhances their stability during in vitro digestion. J. Chem. 2022, 2022. [CrossRef]
- AOAC. Association of official analytical chemists. Available online: https://www.aoac.org/ (accessed on.
- Al-Mubarak, A.; Hamid, N.; Kam, R.; Chan, H. The effects of spray drying conditions on the physical and bioactive properties of New Zealand Tamarillo (Solanum betaceum) powder. Act. Sci. Nutr. Health. 2019, 3, 121-131. [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991-1014. [CrossRef]
- Can-Karaca, A.; Guzel, O.; Ak, M.M. Effects of processing conditions and formulation on spray drying of sour cherry juice concentrate. J Sci Food Agric. 2016, 96, 449-455. [CrossRef]
- Choudhury, N.; Meghwal, M.; Das, K. Microencapsulation: An overview on concepts, methods, properties and applications in foods. Food front 2021, 2, 426-442. [CrossRef]
- Manoharan, R.; Nair, C.S.; Eissa, N.; Cheng, H.; Ge, P.; Ren, M.; Jaleel, A. Therapeutic Potential of Solanum Alkaloids with Special Emphasis on Cancer: A Comprehensive Review. Drug Des Devel Ther. 2024, 3063-3074. [CrossRef]
- Păltinean, R.; Ielciu, I.; Hanganu, D.; Niculae, M.; Pall, E.; Angenot, L.; Tits, M.; Mocan, A.; Babotă, M.; Frumuzachi, O. Biological activities of some isoquinoline alkaloids from Fumaria schleicheri Soy. Will. Plants 2022, 11, 1202. [CrossRef]
- Martini, S.; Conte, A.; Cattivelli, A.; Tagliazucchi, D. Domestic cooking methods affect the stability and bioaccessibility of dark purple eggplant (Solanum melongena) phenolic compounds. Food Chem 2021, 341, 128298. [CrossRef]
- Wojtunik-Kulesza, K.; Oniszczuk, A.; Oniszczuk, T.; Combrzyński, M.; Nowakowska, D.; Matwijczuk, A. Influence of in vitro digestion on composition, bioaccessibility and antioxidant activity of food polyphenols—A non-systematic review. J. Nutr. 2020, 12, 1401. [CrossRef]
- Ydjedd, S.; Bouriche, S.; López-Nicolás, R.; Sánchez-Moya, T.; Frontela-Saseta, C.; Ros-Berruezo, G.; Rezgui, F.; Louaileche, H.; Kati, D.-E. Effect of in vitro gastrointestinal digestion on encapsulated and nonencapsulated phenolic compounds of carob (Ceratonia siliqua L.) pulp extracts and their antioxidant capacity. J. Agric. Food Chem. 2017, 65, 827-835. [CrossRef]
- Nenadis, N.; Tsimidou, M.Z. Assessing the activity of natural food antioxidants. In Oxidation in Foods and Beverages and Antioxidant Applications; Elsevier: 2010; pp. 332-367.
- Elizalde-Romero, C.A.; Basilio-Heredia, J.; Contreras-Angulo, L.A.; Gutiérrez-Grijalva, E.P.; Leyva-López, N. Bioaccesibilidad de extractos hidrofílicos de berenjena (Solanum melongena L.) y su potencial como inhibidor de lipasa Centro de Investigación en Alimentación y Desarrollo A.C., 2020.
- Ogunsuyi, O.B.; Ademiluyi, A.O.; Oboh, G. Solanum leaves extracts exhibit antioxidant properties and inhibit monoamine oxidase and acetylcholinesterase activities (in vitro) in Drosophila melanogaster. JBCPP 2020, 31. [CrossRef]
- Moyo, S.M.; Serem, J.C.; Bester, M.J.; Mavumengwana, V.; Kayitesi, E. The impact of boiling and in vitro human digestion of Solanum nigrum complex (Black nightshade) on phenolic compounds bioactivity and bioaccessibility. Food Res Int 2020, 137, 109720. [CrossRef]
- Izzo, L.; Castaldo, L.; Lombardi, S.; Gaspari, A.; Grosso, M.; Ritieni, A. Bioaccessibility and antioxidant capacity of bioactive compounds from various typologies of canned tomatoes. Front. Nutr. 2022, 9, 849163. [CrossRef]
- Kaur, C.; Nagal, S.; Nishad, J.; Kumar, R. Evaluating eggplant (Solanum melongena L) genotypes for bioactive properties: A chemometric approach. Food Res Int 2014, 60, 205-211. [CrossRef]
- Ng, R.C.; Kassim, N.K.; Yeap, Y.S.; Ee, G.C.L.; Yazan, S.L.; Musa, K.H. Isolation of carbazole alkaloids and coumarins from Aegle marmelos and Murraya koenigii and their antioxidant properties. Sains Malays 2018, 47, 1749-1756. [CrossRef]
- Benítez-Estrada, A.; Villanueva-Sánchez, J.; González-Rosendo, G.; Alcántar-Rodríguez, V.E.; Puga-Díaz, R.; Quintero-Gutiérrez, A.G. Determinación de la capacidad antioxidante total de alimentos y plasma humano por fotoquimioluminiscencia: Correlación con ensayos fluorométricos (ORAC) y espectrofotométricos (FRAP). Tip rev. espec. cienc. quím. -biol. 2020, 23. [CrossRef]
- Bisby, R.H.; Brooke, R.; Navaratnam, S. Effect of antioxidant oxidation potential in the oxygen radical absorption capacity (ORAC) assay. Food Chem. 2008, 108, 1002-1007. [CrossRef]
- Tian, W.; Zhi, H.; Yang, C.; Wang, L.; Long, J.; Xiao, L.; Liang, J.; Huang, Y.; Zheng, X.; Zhao, S. Chemical composition of alkaloids of Plumula nelumbinis and their antioxidant activity from different habitats in China. IC&P 2018, 125, 537-548. [CrossRef]
- Tiong, S.H.; Looi, C.Y.; Hazni, H.; Arya, A.; Paydar, M.; Wong, W.F.; Cheah, S.C.; Mustafa, M.R.; Awang, K. Antidiabetic and antioxidant properties of alkaloids from Catharanthus roseus (L.) G. Don. Molecules 2013, 18, 9770-9784. [CrossRef]
- Goboza, M.; Meyer, M.; Aboua, Y.G.; Oguntibeju, O.O. In vitro antidiabetic and antioxidant effects of different extracts of Catharanthus roseus and its indole alkaloid, vindoline. Molecules 2020, 25, 5546. [CrossRef]
- Siddique, M.A.B.; Brunton, N. Food glycoalkaloids: distribution, structure, cytotoxicity, extraction, and biological activity. In Alkaloids, Kurek, J., Ed.; IntechOpen: Rijeka, 2019.
- Li, H.; Deng, Z.; Liu, R.; Loewen, S.; Tsao, R. Bioaccessibility, in vitro antioxidant activities and in vivo anti-inflammatory activities of a purple tomato (Solanum lycopersicum L.). Food Chem. 2014, 159, 353-360. [CrossRef]
- Velderrain-Rodríguez, G.R.; Palafox-Carlos, H.; Wall-Medrano, A.; Ayala-Zavala, J.F.; Chen, C.-Y.O.; Robles-Sánchez, M.; Astiazaran-García, H.; Alvarez-Parrilla, E.; González-Aguilar, G.A. Phenolic compounds: their journey after intake. J. Funct. Foods 2014, 5, 189-197. [CrossRef]
- Timilsena, Y.P.; Haque, M.A.; Adhikari, B. Encapsulation in the food industry: A brief historical overview to recent developments. Food sci. nutr. 2020, 11, 481-508. [CrossRef]
- Sarabandi, K.; Jafari, S.M.; Mahoonak, A.S.; Mohammadi, A. Application of gum Arabic and maltodextrin for encapsulation of eggplant peel extract as a natural antioxidant and color source. Int. J. Biol. Macromol. 2019, 140, 59-68. [CrossRef]
- Arrazola, G.; Herazo, I.; Alvis, A. Microencapsulación de Antocianinas de Berenjena (Solanum melongena L.) mediante Secado por Aspersión y Evaluación de la Estabilidad de su Color y Capacidad Antioxidante. Inf. Tecnol. 2014, 25, 31-42. [CrossRef]
- Vidović, S.S.; Vladić, J.Z.; Vaštag, Ž.G.; Zeković, Z.P.; Popović, L.M. Maltodextrin as a carrier of health benefit compounds in Satureja montana dry powder extract obtained by spray drying technique. Powder Technol. 2014, 258, 209-215. [CrossRef]
- Vergara, C.; Pino, M.T.; Zamora, O.; Parada, J.; Pérez, R.; Uribe, M.; Kalazich, J. Microencapsulation of anthocyanin extracted from purple flesh cultivated potatoes by spray drying and its effects on in vitro gastrointestinal digestion. Molecules 2020, 25, 722. [CrossRef]
- Rezende, Y.R.R.S.; Nogueira, J.P.; Narain, N. Microencapsulation of extracts of bioactive compounds obtained from acerola (Malpighia emarginata DC) pulp and residue by spray and freeze drying: Chemical, morphological and chemometric characterization. Food Chem 2018, 254, 281-291. [CrossRef]
- Đorđević, V.; Balanč, B.; Belščak-Cvitanović, A.; Lević, S.; Trifković, K.; Kalušević, A.; Kostić, I.; Komes, D.; Bugarski, B.; Nedović, V. Trends in encapsulation technologies for delivery of food bioactive compounds. Food Eng. Rev. 2015, 7, 452-490. [CrossRef]
- Santos-Silva, G.; Gouveia-Gomes, M.H.; Moreira de Carvalho, L.; Leite-Abreu, T.; dos Santos Lima, M.; Suely-Madruga, M.; Kurozawa, L.E.; Alencar-Bezerra, T.K. Microencapsulation of organic coffee husk polyphenols: Effects on release, bioaccessibility, and antioxidant capacity of phenolics in a simulated gastrointestinal tract. Food Chem 2023, 137435. [CrossRef]
- Ortega-Hernández, E.; Camero-Maldonado, A.V.; Acevedo-Pacheco, L.; Jacobo-Velázquez, D.A.; Antunes-Ricardo, M. Immunomodulatory and Antioxidant Effects of Spray-Dried Encapsulated Kale Sprouts after In Vitro Gastrointestinal Digestion. Foods 2023, 12, 2149. [CrossRef]
- Cardona-Tangarife, D.P.; Patiño-Arias, L.P.; Ormaza-Zapata, A.M. Aspectos tecnológicos de la microencapsulación de compuestos bioactivos en alimentos mediante secado por aspersión. Cienc. Tecnol. Agropecuaria. 2021, 22, 1-21. [CrossRef]
- Iturri, M.S.; Barros-Calado, C.M.; Prentice, C. Microparticles of Eugenia stipitata pulp obtained by spray-drying guided by DSC: An analysis of bioactivity and in vitro gastrointestinal digestion. Food Chem. 2021, 334, 127557. [CrossRef]
- Tomé-Sánchez, I.; Martín-Diana, A.B.; Peñas, E.; Frias, J.; Rico, D.; Jiménez-Pulido, I.; Martínez-Villaluenga, C. Bioprocessed wheat ingredients: Characterization, bioaccessibility of phenolic compounds, and bioactivity during in vitro digestion. Front. Plant Sci. 2021, 12, 790898. [CrossRef]
- Hameed, A.; Ijaz, S.; Mohammad, I.S.; Muhammad, K.S.; Akhtar, N.; Khan, H.M.S. Aglycone solanidine and solasodine derivatives: A natural approach towards cancer. Biomed. Pharmacother. 2017, 94, 446-457. [CrossRef]
- Merkel, S.; Dib, B.; Maul, R.; Köppen, R.; Koch, M.; Nehls, I. Degradation and epimerization of ergot alkaloids after baking and in vitro digestion. Anal. Bioanal. Chem. 2012, 404, 2489-2497. [CrossRef]
- Alminger, M.; Aura, A.M.; Bohn, T.; Dufour, C.; El, S.N.; Gomes, A.; Karakaya, S.; Martínez-Cuesta, M.C.; McDougall, G.J.; Requena, T. In vitro models for studying secondary plant metabolite digestion and bioaccessibility. Compr. Rev. Food Sci. Food Saf. 2014, 13, 413-436. [CrossRef]
- Vronen, P.J.E. Potato glycoalkaloids as starting material for the synthesis of steroid hormones; Wageningen University and Research: 2003.
Figure 1.
Scanning electron microscopy of the microcapsules of the alkaloid-rich extract of eggplant fruit. Specifications: (A), magnification: 2.5 K X, beam intensity (HV): 10.00 kV, sample to leans distance (WD): 6.0 mm. (B), magnification: 5.0 K X, beam intensity (HV): 10.00 kV, sample to lens distance (WD): 6.0 mm.
Figure 1.
Scanning electron microscopy of the microcapsules of the alkaloid-rich extract of eggplant fruit. Specifications: (A), magnification: 2.5 K X, beam intensity (HV): 10.00 kV, sample to leans distance (WD): 6.0 mm. (B), magnification: 5.0 K X, beam intensity (HV): 10.00 kV, sample to lens distance (WD): 6.0 mm.
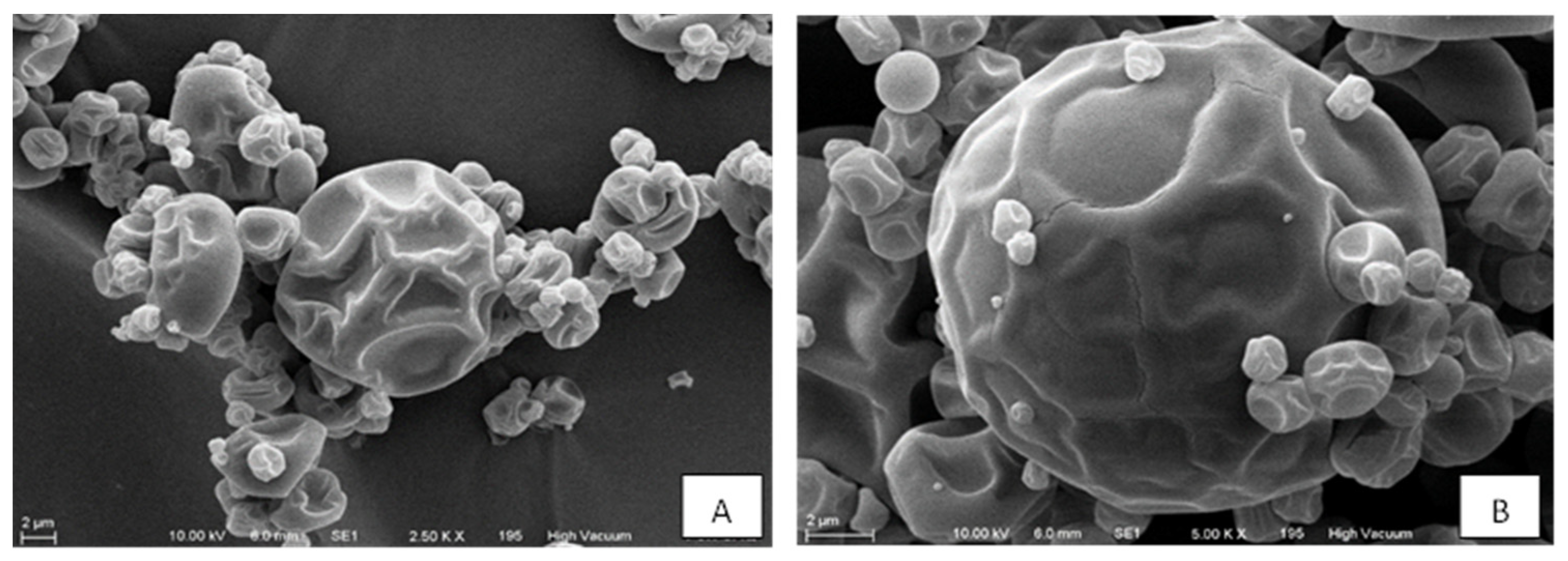

Table 1.
Total alkaloids and antioxidant capacity of alkaloid-rich extract of eggplant biomass fruit.
Table 1.
Total alkaloids and antioxidant capacity of alkaloid-rich extract of eggplant biomass fruit.
| Method | Undigested | Digested | % BA |
|---|---|---|---|
| Total alkaloid content* | 173 ± 8.6a | 7 ±0.8 b | 4 |
| TEAC** | 1195 ± 10a | 141 ± 16b | 12 |
| FRAP** | 800 ± 21a | 50 ± 2b | 6 |
| ORAC** | 1778 ± 43a | 545 ± 5b | 31 |
*mg ESS/g DE; ** µmol TE/g DE. Data is shown as mean ± standard deviation (n=3). Equal letters indicate no statistically significant difference by the Tukey test (p < 0.05). Comparisons were made by row and treatment.
Table 2.
Alkaloids and glycoalkaloids identified and quantified by UPLC-MS in the alkaloid-rich extract of eggplant fruit biomass.
Table 2.
Alkaloids and glycoalkaloids identified and quantified by UPLC-MS in the alkaloid-rich extract of eggplant fruit biomass.
| Compound | Compound type | Molecular mass [M+H]+ | Retention time (min) | Undigested (ng/g) |
Digested (ng/g) |
% BA |
|---|---|---|---|---|---|---|
| Solamargine | Glycoalkaloid | 867.49 | 3.86 | 2485 ± 6a | 431 ± 11b | 17.34 |
| Solasonine | Glycoalkaloid | 883.49 | 3.81 | 1724 ± 35a | 263 ± 9b | 15.26 |
Data is shown as mean ± standard deviation (n=3). Equal letters indicate no statistically significant difference by the Tukey test (p < 0.05). Comparisons were made by row and treatment.
Table 3.
Total alkaloids and antioxidant capacity of encapsulated eggplant alkaloids fruit.
| Method | Undigested | Digested | %BA |
|---|---|---|---|
| Total alkaloid content* | 1.59 ± 0.06a | 0.185 ± 0.04b | 12 |
| TEAC** | 3.90 ± 0.05b | 8 ± 0a | >100 |
| FRAP** | 3.46 ± 0.22a | 2.75 ± 0b | 79 |
| ORAC** | 16 ± 0b | 30 ± 1a | >100 |
*mg ESS/g powder; **µmol TE/g powder. Data is shown as mean ± standard deviation (n=3). Equal letters indicate no statistically significant difference by the Tukey test (p < 0.05).
Table 4.
Alkaloids identified and quantified by UPLC-MS of encapsulated eggplant alkaloids fruit.
| Compound | Alkaloid type | Molecular mass [M+H]+ | Retention time (min) | Undigested (ng/g) |
Digested (ng/g) |
|---|---|---|---|---|---|
| Solamargine | Glycoalkaloid | 867.49 | 3.86 | 6.111 ± 1b | 12.74 ± 0a |
| Solasonine | Glycoalkaloid | 883.49 | 3.81 | 5.169 ± 1b | 9.743 ± 0a |
Data is shown as mean ± standard deviation (n=3). Equal letters indicate no statistically significant difference by the Tukey test (p < 0.05). Comparisons were made by row and treatment. ND: No detected.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



