
Submitted:
31 October 2024
Posted:
01 November 2024
You are already at the latest version
Abstract
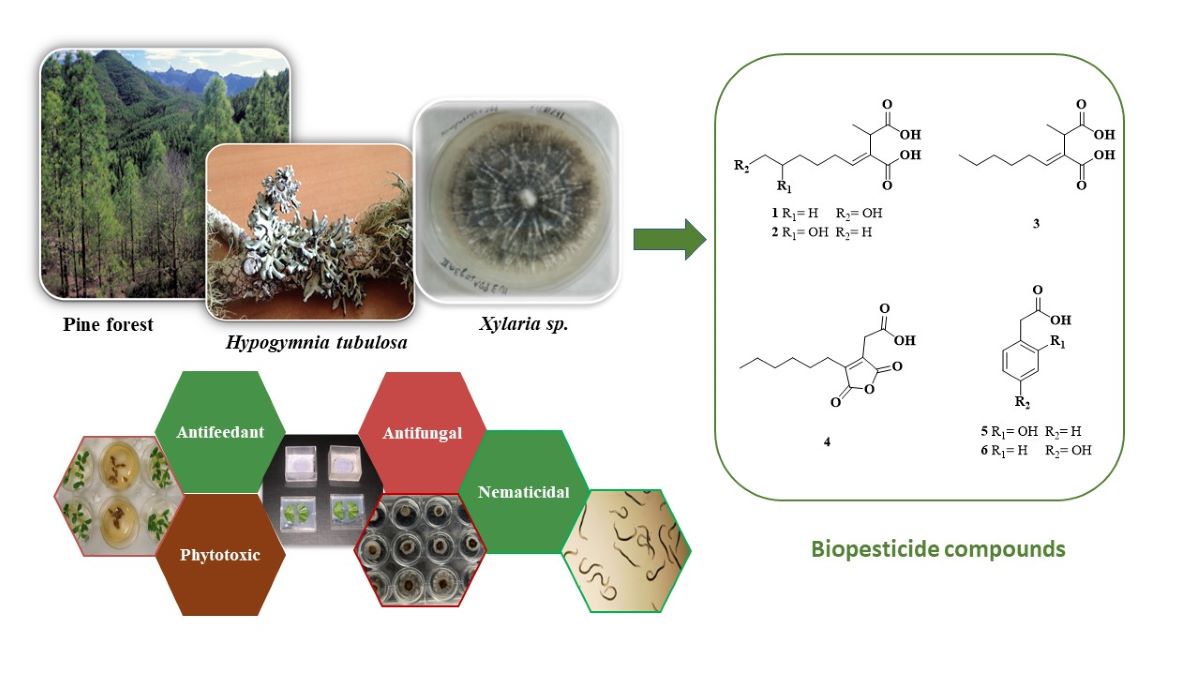
Endolichenic fungi represent an important ecological group of microorganisms that form associ-ations with photobionts in the lichen thallus. These endofungi that live in and coevolve with li-chens are known for synthesizing secondary metabolites with novel structures and diverse chemical skeletons making them an unexplored microbial community of great interest. As part of our search for new phytoprotectans, in this work we studied the endolichenic fungus Xylaria sp. isolated from the lichen Hypogymnia tubulosa, which grow as epiphyte on the bark of the endemic tree Pinus canariensis. From the extract of liquid fermentation, we isolated two new piliformic de-rivatives, (+)-9-hydroxypiliformic acid (1) and (+)-8-hydroxypiliformic acid (2), along four known compounds, (+)-piliformic acid (3), hexylaconitic acid A anhydride (4) and two hydroxy-phenylacetic derivatives (5-6). Their structures were elucidated based on NMR and HRESIMS data. The extract and compounds were tested for their antifeedant (Myzus persicae, Rhopalosiphum padi and Spodoptera littoralis), antifungal (Alternaria alternata, Botrytis cinerea and Fusarium ox-ysporum), nematicidal against Meloidogyne javanica and phytotoxic effects on mono and dicotyle-donous plant models (Lolium perenne and Lactuca sativa). Among these compounds 4, 5 and 6 were effective antifeedants against M. persicae and 4 was also active against R. padi. Moreover, com-pound 4 and 3 showed antifungal activity against B. cinerea and 4 was the only nematicidal. Phytotoxic test showed that the extract has strong effect against L. sativa and L. perenne growth. The phytotoxic compounds were 3, 4 and 5. (+)-Piliformic acid (3) and hexylaconitic A anhydride (4) displayed root growth stimulation on low doses and inhibitory effect at higher doses of di-cotyledon specie L. sativa.
Keywords:
Xylaria sp.
; endolichenic fungi
; (+)-9-hydroxypiliformic acid
; (+)-8-hydroxypiliformic acid
; biopesticide activity
1. Introduction
Endolichenic fungi represent an important ecological group of microorganisms that form associations with photobionts in the lichen thalli [1]. These fungi live symbiotically inside the lichen, similar to how endophytic fungi inhabit plant tissues [2]. In this symbiosis the lichen provides the living environment and nutrients for the survival of the endolichenic fungi and in turn, they produce a variety of secondary metabolites that help the lichen to accelerate their growth and to protect their host during biotic and abiotic stress. The potential of endolichenic fungi to produce novel structures, diverse skeletons, and extensive bioactivities based on their mutualistic symbiosis with their hosts and unique living environment, has gained significant research attention due to their possible applications in medicine and agriculture [3].
Fungi from the Xylariaceae family are frequent saprotrophs, and have been described as endophytes of phylogenetically diverse plant and lichens species from a variety of ecosystems [4,5,6,7,8]. Furthermore, xylarialean endophytes can be symbiotic or saprotrophic [9]. Endolichenic isolates from the Xylaria genus have demonstrated the ability to produce a great diversity of secondary metabolites with varied bioactivities. Several novel antimicrobial eremophilane sesquiterpenes, named eremoxylarins D-J, have been isolated from X. hypoxylon from the lichen Rhizocarpon geographicum [10]. Additionally, the nematicidal polyketide grammicin was isolated from X. grammica associated with the lichen Menegazzia sp. [11]. A variety of compounds, including aplysinopsin, ophiocerin B, and the polyketides piliformic acid and methyl xylariate C, were isolated from X. venustula, associated to the lichen Usnea baileyi [6]. Furthermore, endolichenic Xylaria strains have also been reported to produce novel cyclic peptides, such as the cyclic depsipeptide xylaroamide A from Xylaria sp. associated with Usnea sp. [12] and a new nitrogen containing phenolic compound with cytotoxic activity has been discovered from X. psidii, [13].
Secondary metabolites produced by Xylaria species have also shown phytoprotectant activities, including herbicidal, fungicidal and insecticidal [14]. Various endophytic isolates of Xylaria sp. have been reported to produce griseofulvin [15,16,17], a metabolite that inhibits the growth of several species of fungal plant pathogens [17]. Xylaria sp., isolated from Vitis labrusca, produce diplosporin [18], a compound toxic to the polyphagous insect Spodoptera frugiperda when added to artificial diet [19]. Additionally, cytochalasin E isolated from the endophytic fungus Xylaria sp. showed phytotoxic effects on Lactuca sativa and Raphanus sativus seedlings [20].
The excessive use of synthetic pesticides needed to increase crop production and productivity by controlling pathogens that harm the crops [21] has led to pathogenic resistance, environmental problems affecting soil biodiversity, beneficial insects, aquatic life, and has been linked to detrimental effects on human health [22]. Biopesticides, of natural origin, are an alternative to control plant pathogens with less toxic effects on the ecosystem [23].
In this work, the endolichenic fungal strain HYP6 isolated from the lichen Hypogymnia tubulosa was fermented in malt liquid medium and extracted with ethyl acetate (EtOAc). The fractionation of the extract gave two new compounds and four known metabolites that have been identified based on their spectroscopic data. The extract and pure compounds were tested for their antifeedant (Myzus persicae, Rhopalosiphum padi and Spodoptera littoralis), nematicidal against Meloidogyne javanica, fungicidal (Alternaria alternata, Botrytis cinerea and Fusarium oxysporum) and phytotoxic effects on mono and dicotyledonous plant models (Lolium perenne and Lactuca sativa).
2. Results and Discussion
2.1. Fungal Identification
The endolichenic fungal strain HYP6 was isolated from the lichen Hypogymnia tubulosa from the bark of the endemic tree, Pinus canariensis, collected in Tenerife Island (Spain). The rDNA ITS was sequenced and compared with those deposited in the NCBI-Genbank. To refine the taxonomic assignment a total of 55 related sequences were used to conduct maximum likelihood phylogenetic analysis using Tamure-3 parameter (T92+G) model and a bootstrap test with 5000 runs. The results indicated that strain HYP6 is closely related to Xylaria arbuscula (Figure 1). Based on multigene phylogenetic analyses [24], X. arbuscula is actually considered a species complex, which predominantly including species from the tropical Xylaria group [25]. According to these studies and the results described above, the fungal strain HYP6 was identified as Xylaria sp. within the X. arbuscula complex.
2.2. Structure Elucidation
The fractionation of the extract from the culture broth of endolichenic fungi Xylaria sp fermented in malt liquid medium led to the isolation and structural identification of two new piliformic derivatives, (+)-9-hydroxypiliformic acid (1) and (+)-8-hydroxypiliformic acid (2) together with the known compounds, (+)-piliformic acid (3) as the main metabolite of extract, hexylaconitic acid A anhydride (4), 2-hydroxyphenylacetic acid (5) and 4-hydroxyphenylacetic acid (6) (Figure 2).
Compound 1 was isolated as colourless oil. Its molecular formula was determined as C11H18O5 by the positive ion mode HRESIMS data at m/z 253.1059 [M+Na]+ (calcd. for C11H18O5Na, 253.1052), indicating three degrees of unsaturation. The IR spectrum showed absorption bands at vmax 3355, 1689 and 1637 cm-1, which indicated the presence of a hydroxyl group, carboxylic acid and double bound, respectively. In the 1H RMN spectrum (Table 1 and Figure S1) was observed resonances of one methyl group at δ 1.31 (d, J=7.1 Hz, H-11) coupled with a methine at δ 3.64 (dq, J=7.1 Hz, H-2) and one olefinic proton at δ 6.85 (t, J=7.6Hz). Other signals observed were multiplets attributed to the protons of three overlapped methylenes of an aliphatic chain at δ 1.42-1.66 and the signal at δ 3.55 (t, J=6.6Hz, H-9) of hydroxymethylene group. Analysis the 13C NMR (Table 1 and Figure S2) data in combination with gHSQC spectrum (Figure S4) showed eleven carbon signals, two carbonyl groups at δ 178.0 (C-1) and 170.3 (C-10), two olefinic carbons at δ 134.4. (C-3) and 144.7 (C-4) of a trisubstituted double bond, one oxygenated methylene at δ 62.8 (C-9), one methine at δ 39.0 (C-2), one methyl group at δ 16.4 (C-11), as well as four methylene carbons at δ 29.4 (C-5), 29.6 (C-6), 26.7 (C-7) and 33.4 (C-8) in aliphatic region. The 1H-1H COSY NMR spectrum of 1 (Figure S3) showed coupling between the H-2 and H-11 and also established a partial structure from the olefinic proton at δ 6.85 (H-4) along the aliphatic chain with a spin system of methylenic protons H-5/ H-6/H-7/H-8/H-9.
The low field resonance of a methine proton at δ 3.64 (H-2) in the CH-CH3 grouping indicated deshielding effects due to carboxyl groups in the molecule. The carboxyl groups are located on C-1 and C-10, as supported by the HMBC correlations between H-2 and C-1/ C-3/ C-4/C-10/C-11 and between H-11 methyl with C-1/C-2/C-3 (Figure 3). The position of the double bond was confirmed by the correlations previously indicated for H-2 and those observed between H-4 and C-2/ C-3/ C-5/ C-10. The HMBC experiment (Figure S5) showed also connectivities of H-5 with C-2, C-3, C-4, C-6 and C-7, and H-9 with C-7 and C-8, which confirmed the structure of compound 1 closely related to the known fungal metabolite (+) piliformic acid (3) [26,27], also isolated from this fungal strain and whose structure has been obtained by chemical synthesis [28]. The configuration of double bound was assigned E based on the NOESY between H-3 and H-5 (Figure S6), however with the available spectroscopic data the configuration at C-2 in compound 1 could not be defined. Thus, this new compound was identified as (+)-(E)-9-hydroxypiliformic acid (1).
Compound 2 exhibited an ion peak in its HRMS (ESI+) at m/z 253.1058 [M+Na]+, corresponding to the molecular formula C11H18O5Na, which was identical to that compound 1. The main difference observed in its 1H-NMR (Table 1 and Figure S8) is the absence of signal of hydroxymethylene group at C-9 in 1 that was replaced by a new signal due to hydroxyl group at δ H 3.74 coupled with a new doublet methyl signal at δ 1.16 (3H, d, J=6.2 Hz). These data suggested that these two compounds differ in the position of hydroxyl group in 2. Furthermore, the 13C NMR spectrum showed an additional methyl carbon at δ 23.5.8 (C-9) and a methine group at 68.2 (C-8) (Figure S9).
The location of hydroxyl group at C-8 was confirmed by the 1H-1H COSY crosspeak of H-9 (δ 1.16)/ H-8 (δ 3.74) together the HMBC correlations of H-9 methyl group with C-7 and C-8 (Figure 3 and Figure S12). A detailed analysis of the 13C NMR (Table 1 and Figure S9) showed double signals for C-5, C-6, C-7 and C-8, which indicated that compound 2 was the mixture of two epimers at C-8 with a ratio 2:1 according to the height of double peaks. These observations, together with a careful analysis of the COSY, HMBC and NOESY experiments, led to the identification compound 2 as a mixture (ratio 2:1) of two epimers of (+)-(E)-8-hydroxypiliformic acid.
The known compound 4 was identified as 2-carboxymethyl-3-n-hexyl-maleic acid anhydride and named hexylaconitic acid A anhydride. Its molecular formula was determined as C12H16O5 in accordance with its HRESIMS (m/z 239.0920 [M-H]-, calculated for C12H15O5 239.0919) (Figure S18). The 1H NMR of compound 4 revealed signals representative of one n-alkyl chain (n-hexyl moiety) and a deshielded singlet signal that integrated for two protons at δ 3.55 (H-11). The 13C NMR spectrum (Figure S17) revealed the presence of three carboxyl carbons δ 173.2 (C-12), 165.3 (C-1), 165.2 (C-4), two olefinic quaternary carbons at δ 148.3 (C-2), 135.6 (C-3) and one methyl group at δ 14.1 (C-10) alongside with the presence of six methylenes δ 31.4 (C-8), 27.6 (C-6), 25.1 (C-5), 22.5 (C-9) and two overlapped carbons at δ 29.3 (C-7, C-11). The 1H and 13C NMR spectral data are consistent with those reported for 2-carboxymethyl-3-n-hexyl-maleic acid anhydride, which was first isolated as a natural product from Aspergillus niger [29] and later obtained by chemical synthesis [30,31]. Now in this work, we have completed the assignments of the proton and carbon resonances in the 1H and 13C spectra using 2D-NMR experiments (COSY, HSQC and HMBC), which had not been previously assigned. The configuration Z of double bound was also confirmed by the crosspeak of H2-5/H2-11 in the NOESY spectrum (Figure S27).
Another known compounds isolated from this fungus were identified as (+)-piliformic acid (3) [26,27,28], a fungal metabolite isolated previously from different fungi belonging to the Xylaria genus [32], and the phenolic compounds, 2-hydroxyphenylacetic acid (5) [33] and 4-hydroxyphenylacetic acid (6) [34]. In the experimental part we provide complementary NMR and MS data of these compounds.
2.3. Biopesticide Activity
The EtOAc extract and compounds obtained from the fermentation of HYP6 were tested for their biopesticide properties against insect pest (Myzus persicae, Rhopalosiphum padi and Spodoptera littoralis), the plant parasitic nematode Meloidogyne javanica and fungal phytopathogens (Alternaria alternata, Botrytis cinerea and Fusarium oxysporum).The extract showed strong antifeedant effects against M. persicae (EC50 value of 10.9 µg/cm2) followed by a moderate activity against R. padi and it was not active on S. littorallis. This extract was also antifungal to B. cinerea and moderately active on F. oxysporum and A. alternata but it did not have any nematicidal effect (Table 2).
When tested the compounds on insect pests (Table 2), 4 showed a strong antifeedant activity against M. persicae (EC50 value of 1.6 µg/cm2) and R. padi (EC50 value of 8.9 µg/cm2), being more effective than the extract. Additionally, compounds 5 and 6 showed a selective antifeedant activity against M. persicae with EC50 value of 4.5 µg/cm2 and EC50 value of 15.5 µg/cm2, respectively. None of the tested compounds showed any antifeedant effects against S. littoralis. Compound 4 also exhibited a strong inhibition of mycelial growth of B. cinerea (EC50 value of 0.12 mg/ml) and moderate against A. alternata (EC50 value of 0.24 mg/ml), whereas compound 3 was moderately active on B. cinerea (EC50 value of 0.36 mg/ml). When tested against M. javanica, compound 4 was very active with an LD50 value of 0.10 mg/ml.
The extract and compounds were also tested for phytotoxic effects on seeds of mono- and dicotyledoneous plant species (L. perenne and L. sativa) (Figure 4). The extract of Xylaria sp. strongly inhibited the germination of L. perenne with 88.2% inhibition after 7 days compared to the control. The extract also exhibited phytotoxic effects on L. perenne, with 89.3% and 100% inhibition of leaf and root growth, respectively and affected the root growth of L. sativa (66.8% inhibition). Among the compounds tested, compound 3 affected the germination of L. sativa (86.7% inhibition), decreased moderately root growth of L. perenne (56.8% inhibition) and affected the root growth of L. sativa (72.6% inhibition). Compound 4 reduced significantly the germination of L. perenne and L. sativa (72.2% and 94.7% inhibition, respectively) with a strong inhibition of root growth (100% and 97.9% inhibition, respectively). Additionally, this compound also decreased the leaf growth of L. perenne (73.1% inhibition). Compound 5, with reported phytotoxic effects [35,36], showed significant effect on root growth on L. perenne and L. sativa root growth (85.4% and 91.3% inhibition, respectively) (Figure 4). The most phytotoxic compounds (3, 4) were further tested in dose-response experiments against the two plant species. At lower concentrations, both compounds stimulated the root growth of L. sativa. Compound 3 increased the root length up to 170% at a concentration of 0.1 mg/ ml and 4 up to 200% at 0.05 mg/ml.
In this work, (+)-piliformic acid (3) showed moderate antifungal effects against B. cinerea. Similarly, previous reports showed moderate activity of 3 against Colletotrichum gloeosporioides, one of the phytopathogenic fungus responsible for the anthracnose disease [37]. Compound 4, with aphid antifeedant, antifungal and nematicidal effects was identified as hexylaconitic A anhydride. This compound (4) isolated from Aspergillus niger[29,38] and Aspergillus tubigensis [39] has been previously reported for its fungicidal activity against Neurospora crassa [39], but it was found inactive against other fungal plant pathogens (Gaeumannomyces graminis var. tritici, Rhizoctonia solani, and Phytophthora cinnamomic) [38]. This may indicate a species-dependent antifungal effects that could be used to target specific fungal pathogens. However, this is the first report on the aphid antifeedant and nematicidal effects of compound 4. Phenolic acids related to 5 and 6, are known for their insecticidal activities [40] and play an important role in the plant resistance against insect pest [41]. For example, previous reports showed that phenylacetic compounds isolated from Streptomyces gramineus have insecticidal activity against Thrips palmi, also known as melon thrips, a sap-sucking phytophagous insect [42].
The phytotoxic effects observed are consistent with those of the analogous compounds of alkylitaconic acid derivatives, which promoted radicle growth at low doses, while inhibited the seedlings growth at higher doses particularly in dicotyledonous species [35,43]. Additionally, Mondal et al. [44] reported that compound 4 stimulates germination and seedling growth in cauliflower at low ppm concentrations. This behaviour could be explained by hormetic effects of toxic agent showing a biphasic response, promoting early seedling development (and potentially later plant growth) at low concentrations, while inhibiting growth at high concentrations [45,46]. This work has shown for the first time the potent phytotoxic effects of (+)-piliformic acid (3) and hexylaconitic A anhydride (4) on mono and dicotyledonous plant (L. perenne and L. sativa) and stimulating effect the growth of L. sativa root at lower dose.
3. Materials and Methods
3.1. General Experimental Procedures
Optical rotations were measured on MCP 150-Anton Paar polarimeter (Anton Paar, Seelze-Letter, Germany) at a temperature of 25°C. IR spectra were recorded with a Cary 630 FTIR spectrophotometer (Agilent, Santa Clara, CA, USA). 1H ,13C and 2D (COSY, HSQC, HMBC, and NOESY) NMR spectra were recorded in CDCl3 or CD3OD solution (Aldrich, St. Louis, MO, USA) on a Bruker Avance II-500 spectrometer equipped with a 5 mm TCI inverse detection cryo-probe (Bruker Biospin, Falländen, Switzerland). Chemical shifts are given in ppm (δ), referenced to solvent signal (CDCl3, δH 7.26 and δC 77.0; CD3OD3, δH 3.31 and δC 49.0. The HRESIMS mass spectra were obtained using a Waters LCT Premier XE mass spectrometer (Manchester, UK). Sephadex LH-20 (Sigma-Aldrich, St. Louis, MO, USA) and silica gel 60 (40±63 μm, Merck, Darmstadt, Germany) was used for column chromatography. TLC was performed with precoated silica gel 60 F254 plates (Merck, Darmstadt, Germany). Flash chromatography separations were carried out on a Biotage Isolera Prime (BIOTAGE, Uppsala, Sweeden) apparatus equipped with a UV detector (200-400 nm) using a prepacked RediSep flash column of 35g of silica gel (90 ml, 13 cm x 3 cm; Teledyne Isco, Lincoln, NEB, USA). Semipreparative HPLC was performed on a Beckman System Gold 125P equipped with a diode-array detector Beckman Coulter 168 (Brea, CA, USA) using a semi-preparative Beckman Ultrasphere ODS (10.0 × 250 mm, 5 μm) column
3.2. Lichen Material and Isolation of the Endolichenic Fungus HYP6
The fungal strain HYP6 was isolated from the lichen Hypogymnia tubulosa collected in Tenerife Island (Las Lagunetas, 28°24'47.2"N, 16°24'11.2"W) from the bark of endemism Pinus canariensis, dominant tree of the Canarian pine forest ecosystem. The samples were placed into sterile polybags and transported under refrigeration in a box container until isolation processing, within 24 hours of collection. Collected lichen thalli were initially washed with distilled water to remove excess dirt. For surface sterilization, thalli were cut into manageable portions followed by successively dipping in a 70% ethanol solution for 1 minute, 1% sodium hypochlorite for 10 minutes and finally washing with 70% ethanol for 1 minute. The surface sterilized thalli were dried on sterilized paper and cut into 1cm pieces. Then, lichen explants were inoculated in Potato Dextrose Agar (PDA) and Malt extract Agar (MEA) media in petri dishes, containing 50 mg/L of the antibiotic chloramphenicol to avoid bacterial contamination. The plates were incubated at 24°C for 2-3 weeks in darkness with daily observation of the emerging fungal colonies. The new fungal mycelia that grew from the lichen explants were transferred to fresh plates containing the respective medium and various subcultures were performed until pure cultures were obtained.
3.3. Molecular Characterization of HYP6 Strain
For the extraction of DNA of the fungal strain the DNeasy Plant mini kit (Qiagen GmbH, Hilden, Germany, Cat. No 69104) was used. The ITS rDNA region was PCR-amplified using the oligonucleotide primers ITS1 (5'-TCCGTAGGTGAACCTGCGG-3') and ITS4 (5'-TCCTCCGCTTA TTGATATGC-3') with the following steps: Genomic DNA (100–200 ng) was amplified on a PTC-200 Thermal Cycler (MJ Research, San Diego, CA, USA) in a 25 µL final volume with the AmpONE Taq DNA polymerase PCR kit (GeneAll, Seoul, Korea) for 35 cycles (95 ◦C, 1 min; 50 ◦C, 20 s; 72 ◦C, 1.5 min) after an initial denaturation (95 ◦C, 2 min) and followed by a final extension (72 ◦C, 7 min). Amplicons were checked by agarose gel (1%) electrophoresis, purified using the EXO-SAP-IT kit (Affimetrix-USB; Thermo Fisher Scientific, Waltham, MS, USA), and sequenced on an AB 3500 Genetic Analyzer (Thermo Fisher Scientific, Waltham, J. Fungi 2021, 7, 109 3 of 11 MS, USA) at the University of La Laguna (La Laguna, Spain) genomic service. The consensus sequence of the rDNA ITS region was generated using aligner software Bioedit (version 5.09; Tom Hall, Department of Microbiology, North Carolina State University, Raleigh, NC, USA) and compared with those published in NCBI (https://www.ncbi.nlm. nih.gov/) database by using the online BLAST program (ncbi.nlm.nih. gov/BLAST, accessed on 29 May 2024 ). The related sequences with high percent identity were aligned using the MEGA11.0 program (Mega Limited, Auckland, New Zealand, Tamura K. 2021) [47] and a phylogenetic tree was constructed using maximum likehood method with a Tamure-3 parameter (T92+G) model and a bootstrap test with 5000 runs. The display and annotation of the phylogenetic tree was performed using the online tool: The Interactive Tree Of Life (https://itol.embl.de, accessed on 31 May, 2024).
3.4. Cultivation of HYP6
The endophytic strain HYP6 was cultivated on PDA solid medium for 10 days at 25 °C. Sterile water (10ml) was added to each petri dishes and the surface of mycelium was gently srcaped with a spatula to obtain a suspension of mycelium. This suspension was cultivated in 250 ml Erlenmeyer flasks containing 50 ml of MEB medium (17 g/l of malt extract, 3 g/l of peptone, pH adjusted at 5.4±0.2) for 5 days at 25 ◦C on a rotary shaker (120 rpm) to prepare the seed culture. Ten Erlenmeyer flasks (500 ml) with 200 ml of fresh MEB medium each were inoculated with 10 ml of seed culture and cultured at 25 °C under continuous agitation (120 rpm) for 14 days.
3.5. Extraction and Isolation of Compounds
The fermentation broth was filtered under reduced pressure through a Buchner funnel using cheesecloth (25 µm pore diameter) to separate the mycelium. The resulting filtrate was subjected to a three-cycle liquid/liquid extraction with ethyl acetate (EtOAc), dried over Na2SO4, and the organic phase concentrated under reduced pressure to yield the crude HYP6 extract (817.5 mg).
The EtOAc extract was fractionated by column chromatography on silica gel eluting with a gradient of increasing polarity of n-hexane/EtOAc and then Cl2CH2/MeOH. The fractions obtained were analysed by TLC and combined those with high similarity to afford five main fractions. Fraction 2 was separated by media pressure cromatography using a Biotage Isolera Prime equipment through on 35g Si pre-packed flash cartridge column, eluted with n-hexane/EtOAc mixtures of increasing polarity (70:30–25:75) at 10 ml/min to afford compound 3 (33.7 mg) and 4 (13.5 mg). Fraction 3 was further purified by Sephadex LH-20 with a mixture of CH2Cl2:MeOH (1:1) to give compounds 5 (11.3 mg) and 6 (8.4mg). Fraction 4 was separated by Sephadex-LH-20 using an isocratic mixture of n-hexane:CH2Cl2:MeOH (1:1:1) to yield four fractions (Fr. 4.1-Fr.4.4). Fr.4.3 was further separated by reverse semi-preparative HPLC (Ultrasphere ODS, 10.0 × 250 mm, 5 μm) using a gradient solvent system of H2O/MeOH (80:20-30:70) at a flow rate of 3 ml/min to afford compound 2 (18.4 mg, tR=42.6 min). Similarly, the reverse semi-preparative HPLC of subfraction FR.4.4 eluted with H2O:MeOH (60:40) gave compound 1 (4.6 mg, tR=32.0 min).
3.6. Spectroscopic Data of Compounds
3.6.1. (+)-9-. Hydroxypiliformic Acid (1)
3.6.2. (+)-8-. Hydroxypiliformic Acid (2)
3.6.3. (+)-Piliformic Acid (2-hexylidene-3-methylsuccinic Acid) (3)
White solid; +24 (c 0.5, CHCl3); IR νmax: 2927, 2859, 1685, 1637, 1418, 1241 cm-1; 1H NMR (CDCl3, 500 MHz,): δ 0.89 (3H, t, J = 6.9 Hz, H-9), 1.31 (4H, m, H-7, H-8), 1.35 (3H, d, J =7.1 Hz, H-11), 1.48 (2H, br t, J =6.8 Hz, H-6), 2.23 (2H, m, H-5), 3.63 (1H, q, J =7.1 Hz, H-2), 7.01 (1H, t, J =7.5 Hz, H-4); 13C NMR (CDCl3, 125 MHz): δ 180.4 (C-1), 37.7 (C-2), 131.4 (C-3), 147.4 (C-4), 28.9 (C-5), 28.2 (C-6), 31.6 (C-7), 22.6 (C-8), 14.1 (C-9), 172.3 (C-10), 15.1 (C-11); HRMS (ESI-) m/z 213.1129 [M-H]- (calcd. C11H17O4, 213.1127).
3.6.4. Hexylaconitic Acid A Anhydride (2-carboxymethyl-3-n-hexyl-maleic acid anhydride) (4)
Yellowish oil; IR νmax: 2929, 2859, 1855, 1825, 1763, 1718, 1273 cm-1; 1H NMR (CDCl3, 500 MHz): δ 0.88 (3H, t, J=6.7 Hz, H-11), 1.30 (6H, m, H-8, H-9, H-10), 1.59 (2H, quint, J=7.5 Hz, H-7), 2.49 (2H, t, J=7.7 Hz, H-6), 3.56 (2H, s, H-11), 8.45 (1H, br s, -OH); 13C NMR (CDCl3, 125 MHz,): δ 165.3 (C-1), 148.3 (C-2), 135.6 (C-3), 165.2 (C-4), 25.1 (C-5), 27.6 (C-6), 29.3(C-7, C-11), 31.4 (C-8), 22.5 (C-9), 14.1 (C-10), 173.2 (C-12); HRMS (ESI-) m/z 239.0920 [M-H]- (calcd. C12H15O5, 239.0919).
3.6.5. 2-Hydroxyphenylacetic Acid (5)
White solid; 1H NMR (CD3OD, 500 MHz,): δ 3.58 (2H, s, H-7), 6.77 (2H, m, H-3, H-5), 7.08 (1H, dd, J= 7.4, 1.2 Hz, H-4), 7.11 (1H, td, J=7.4, 1.6 Hz, H-6); 13C NMR (CD3OD, 125 MHz): δ 122.8 (C-1), 156.7 (C-2), 115.9 (C-3), 129.3 (C-4), 120.5 (C-5), 132.1 (C-6), 36.6 (C-7), 176.3 (C-8); HRMS (ESI-) m/z 151.0400 [M-H]- (cacld. C8H7O3, 151.0395).
3.6.6. 4-Hydroxyphenylacetic Acid (6)
White solid; 1H NMR (CD3OD, 500 MHz): δ 3.48 (2H, s, H-7), 6.73 (2H, d, J=8.5 Hz, H-3 and H-5), 7.08 (2H, d, J=8.5 Hz, H-2 and H-6); 13C NMR (CD3OD, 125 MHz): δ 126.8 (C-1), 131.3 (C-2, C-6), 116.2 (C-3, C-5), 157.3 (C-4), 41.1 (C-7), 176.4 (C-8). HRMS (ESI-) m/z 151.0395 [M-H]- (calcd. C8H7O3, 151.0395).
3.7. Antifungal Bioassay
To evaluate the antifungal activity three different phytopathogenic fungi were used: Fusarium oxysporum, Alternaria alternata and Botrytis cinerea. These strains come from the fungal collection at Instituto de Productos Naturales y Agrobiologia-CSIC (Tenerife, Spain) and the colonies were maintained on PDA medium plates in darkness at 25 ◦C.
Based on the protocol describe before [48], the in vitro mycelial growth inhibition assay was conducted in 12-well plates (Falcon) using a modified PDA-dilution method supplemented with the addition of 0.05 mg/ml of methyltetrazolium salts (MTT). Extract and pure compounds dissolved in ethanol (EtOH) were tested at different concentrations (extract at 1 mg/ml and compounds at 0.5, 0.25, 0.1, and 0.05 mg/ml) and were incorporated into the culture medium before plates were poured. EtOH was used as a negative control and all treatments were replicated four times. Fungal colonies were digitalized and measured using ImageJ (http://imagej.nih.gov/ij/, accessed on March 18, 2024) [49] after the incubation the plates at 25 ◦C in darkness for 48 h. The mycelial growth inhibition (%MGI) was calculated as: %MGI = (C − T/C) × 100, where C is the diameter of the control colonies and T is the diameter of the test colonies. Data were analysed with STATGRAPHICS statistical analysis software (Centurion XVI, version 16.1.03) and EC50 values (effective dose to obtain 50% of inhibition) were determined by regression analysis (%MGI on Log-dose).
3.8. Antifeedant Bioassay
Spodoptera littoralis colonies were reared on an artificial diet [50], while Myzus persicae and Rhopalosiphum padi colonies were maintained on bell pepper (Capsicum annuum) and barley (Hordeum vulgare) plants, respectively. The plants were grown from seeds in pots with commercial substrate and regularly infected for aphid feeding (bell pepper plants were infected at a 4 leaves stage and barley plants when they reached approximately a length of 10 cm). Both the insect colonies and their host plants were maintained in a growth chamber at 22 ± 1 ºC, > 70% relative humidity with a 16:8 h light photoperiod.
Bioassays were conducted with 1.0 cm2 leaf disks/fragments of C. annuum (M. persicae, S. littoralis) or H. vulgare (R. padi) as described previously [51]. The tests (10 μL of the solution) were applied at initial doses of 10 or 5 mg/ml (extract or compound) to the upper surface of the leaf fragments. Two sixth-instar larvae (>24 h after moulting) of S. littoralis were placed in 6 Petri dishes (9 cm in diameter) with 2 leaf disks (treatment and control) and allowed to feed at room temperature until 75% larval consumption of the paired control or treatment disks. The leaf disk surface consumption was measured using ImageJ (http://imagej.nih.gov/ij/, accesed on March 18, 2024) [49]. In the case of aphids, twenty (2x2 cm) ventilated plastic boxes containing 10 apterous aphid adults (24-48 h old) were used. Aphids were allowed to feed in a growth chamber under the described environmental conditions for 24 hours. Settling was quantified by counting the number of aphids settled on each leaf fragment. All experiments were repeated twice (SE<10%).
The feeding or settling inhibition (%FI or %SI) was calculated as [1 - (T/C) x 100], where T and C represent treated and control leaf fragments, respectively. The effects (%SI / %FI) were analyzed by the non-parametric Wilcoxon Signed-Rank Test. Extracts and compounds with an effect > 70% were tested in dose-response experiments (3-5 serial dilutions) to calculate their EC50 (the effective dose causing a 50% settling / feeding reduction) with linear regression models (%FI/SI on Log-dose).
3.9. Nematicidal Bioassay
The Meloidogyne javanica population was maintained on tomato plants (Solanum lycopersicum var. Marmande) cultivated in pot cultures and kept in environmentally controlled growth chambers (at 25 ± 1 ◦C, >70% relative humidity). Egg masses of M. javanica were handpicked from infected tomato roots two months after seedling inoculation. Second-stage juveniles (J2) were obtained by incubating egg masses in a water suspension at 25 ◦C for 24 h. The tests were carried out in 96-well plates (BD Falcon, San Jose, CA, USA) and the extract and compounds were dissolved in distilled water containing 5% of a DMSO-Tween solution (0.2% Tween 20 in DMSO) according to Andres et al. [52]. The initial concentrations tested were of 1 and 0.5 mg mL for extract and pure compound, respectively, and four replicates were used for each test. Water containing 5% of a DMSO-Tween solution (0.2% Tween 20 in DMSO) was used for the negative control. The mortality rates after 72 h of incubation are presented as percentage of dead J2 corrected according to Scheider-Orelli’s formula. Serial dilutions were used to calculate the effective lethal doses (LD50) of the active compound by Probit Analysis (STATGRAPHICS Centurion XVI, version 16.1.03)
3.10. Phytotoxic Bioassay
The phytotoxic test was conducted with Lolium perenne and Lactuca sativa seeds placed in 12-well microplates (40 seeds for test), as described [53]. The extract or compounds dissolved in EtOH (negative control) were tested at concentrations of 0.4 or 0.2 mg/ml (final concentration in the well) and diluted serially if needed. Juglone (Sigma) was used as positive control (0.1 mg/ml), resulting in 100% germination inhibition. Briefly, the test solution (20µl) and 300 µl of H2O were added to a 2.5 cm diameter filter paper into each well plates. The seeds (10/5 of L. sativa/L. perenne soaked in distilled water for 8 h) were placed in every well and the parafilm-sealed plates were incubated in a plant growth chamber (25◦C, 70% RH, 16:8 L:D). Germination was monitored for 7 days and leaf length (for L. perenne) and root length (for both species) were measured at the end of the experiment on 25 randomly selected digitalized seedlings with the ImageJ application (http://rsb.info.nih.gov/ij/, acceded on May 20, 2024).
4. Conclusions
In this study the endophytic fungus HYP6, isolated from the epiphytic lichen Hypogymnia tubulosa, was identified as Xylaria sp. belonging to the X. arbuscula complex. The EtOAc extract from the liquid culture was chromatographed to afford (+)-piliformic acid (3) as the main metabolite and two new hydroxylated derivatives, (+)-9-hydroxypiliformic acid (1) and (+)-8-hydroxypiliformic acid (2), along the known compounds, hexylaconitic acid A anhydride (4) and the hydroxyphenylacetic derivatives (5-6). The extract showed significant aphid antifeedant, antifungal and phytotoxic effects. The aphid antifeedant activity of the extract against M. persicae can be explained by compounds 4-6, whereas their moderately active against R. padi probably due to content of antifeedant compound 4. Compound 4 showed potent fungicide effect against B. cinerea and the nematode M. javanica. The phytotoxicity observed can be attributed to the compounds 3-5 affecting negatively the growth of L. perenne and L. sativa. In addition, compound 3 and 4 showed stimulating effect the growth of L. sativa root at low dose. Endolichenic fungus Xylara sp. represents an important source for the biotechnological production of biopesticide compounds for plant disease control. Further optimization of the fermentation process is needed to increase the production of these active metabolites.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Data sequence of Xylaria sp.; Figures S1-S7: 1H-NMR, 13C-NMR, COSY, gHSQC, HMBC, NOESY and HREIMS spectra for compound 1; Figures S8-S14: 1H-NMR, 13C-NMR, COSY, gHSQC, HMBC, NOESY and HREIMS spectra for compound 2; Figures S15-S21: 1H-NMR, 13C-NMR, COSY, gHSQC, HMBC, NOESY and HREIMS spectra for compound 3; Figures S22-S28: 1H-NMR, 13C-NMR, COSY, gHSQC, HMBC, NOESY and HREIMS spectra for compound 4; Figures S29-S31: 1H-NMR, 13C-NMR and HREIMS spectra for compound 5; Figures S32-S34: 1H-NMR, 13C-NMR and HREIMS spectra for compound 6.
Author Contributions
Conceptualization, C.E.D. and A.G-C.; Methodology, M.F.A., F.A.L., C.E.D., A.G-C.; Validation, M.F.A., C.E.D. and A.G.-C; Formal analysis, M.F.A., F.A.L., C.E.D., A.G-C.; Investigation, F.A.L., M.F.A, A.G.-C. and C.E.D.; Supervision, C.E.D. and A.G.-C.; Writing—Original draft, F.A.L., C.E.D., A.G-C.; Writing—Review & editing, C.E.D. and A.G.-C. Funding acquisition, A.G.-C. and C.E.D. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been supported by grant PID2019-106222RB-C31, MINECO/FEDER, MINECO/FEDER. Work is co-financed by the Canary Islands Agency for Research, Innovation and Information Society of the Regional Ministry of Economy, Knowledge and Employment and by the European Social Fund (ESF) Canary Islands Integrated Operational Program 2014–2020, Axis 3 Priority Theme 74 (85%).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the article and Supplementary Materials
Acknowledgments
F.A.L. acknowledge to pre-doctoral contract from the “Agencia Canaria de Investigación, Innovación y Sociedad de la Información (ACIISI) del Gobierno de Canarias”. We gratefully acknowledge to E. Moreno (ICA-CSIC) for his technical support in insect bioassays.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kellogg, J.; Raja, H. Endolichenic fungi: a new source of rich bioactive secondary metabolites on the horizon. Phytochem. Rev. 2017, 16, 271–293. [Google Scholar] [CrossRef]
- Agrawal, S.; Deshmukh, S.K.; Reddy, M.S.; Prasad, R.; Goel, M. Endolichenic fungi: A hidden source of bioactive metabolites, S. Afr. J. Bot. 2020, 134, 163–186. [Google Scholar] [CrossRef]
- Zhang, W.; Ran, Q.; Li, H.; Lou, H. Endolichenic Fungi: A promising medicinal microbial resource to discover bioactive natural molecules-An update. J. Fungi 2024, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Pushpavathi, D.; Krishnamurthy, Y.L. Study on endolichenic fungal assemblage in Parmotrema and Heterodermia lichens of Shivamoga, Karnataka. Mol. Biol. Rep. 2024, 51, 549. [Google Scholar] [CrossRef]
- Miral, A.; Jargeat, P.; Mambu, L.; Rouaud, I.; Tranchimand, S.; Tomasi, S. Microbial community associated with the crustose lichen Rhizocarpon geographicum L. (DC.) living on oceanic seashore: A large source of diversity revealed by using multiple isolation methods. Environ. Microbiol. Rep. 2022, 14, 856–872. [Google Scholar] [CrossRef]
- Santiago, K.A.A; dela Cruz, T.E.E.; Ting, A.S.Y. Endolichenic fungi from common Usnea lichens found in a montane forest in Malaysia: a study on diversity and bioactivity profiling. Asian J. Mycol. 2022, 5, 18–37. [Google Scholar] [CrossRef]
- Vandegrift, R. Xylariales (sordariomycetes, Ascomycota) of the Boston Harbor Islands. Northeastern Nat. 2021, 25, 150–199. [Google Scholar] [CrossRef]
- Thomas, D.C.; Vandegrift, R.; Ludden, A.; Carroll, G.C.; Roy, B.A. Spatial ecology of the fungal genus Xylaria in a tropical cloud forest. Biotropica 2016, 48, 381–393. [Google Scholar] [CrossRef]
- Franco, M.E.E.; Wisecaver, J.H.; Arnold, A.E.; Ju, Y.M.; Slot, J.C.; Ahrendt, S.; Moore, L.P.; Eastman, K.E.; Scott, K.; Konkel, Z.; Mondo, S.J.; Kuo, A.; Hayes, R.D.; Haridas, S.; Andreopoulos, B.; Riley, R.; LaButti, K.; Pangilinan, J.; Lipzen, A.; Amirebrahimi, M.; U'Ren, J.M. Ecological generalism drives hyperdiversity of secondary metabolite gene clusters in xylarialean endophytes. The New phytol. 2022, 233, 1317–1330. [Google Scholar] [CrossRef]
- Miral, A.; Ferron, S.; Rouaud, I.; Slyambayev, D.; Bousarghin, L.; Camuzet, C.; Belouzard, S.; Séron, K.; Le Pogam, P.; Tranchimand, S.; Tomasi, S. Eremoxylarins D-J, antibacterial eremophilane sesquiterpenes discovered from an endolichenic strain of Xylaria hypoxylon. J. Nat. Prod. 2023, 86, 730–738. [Google Scholar] [CrossRef]
- Kim, T.Y.; Jang, J.Y.; Yu, N.H.; Chi, W.J.; Bae, C.H.; Yeo, J.H.; Park, A.R.; Hur, J.S.; Park, H.W.; Park, J.Y.; Park, J.H.; Lee, S.K.; Kim, J.C. Nematicidal activity of grammicin produced by Xylaria grammica KCTC 13121BP against Meloidogyne incognita. Pest Manag. Sci. 2018, 74, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Chang, S.; Li, Y.; Xi, X.; Chen, M.; He, N.; Wang, M.; Zhao, W.; Xie, Y. Molecular networking-based screening led to the discovery of a cyclic heptadepsipeptide from an endolichenic Xylaria sp. J. Nat. Prod., 2022, 85, 972–979. [Google Scholar] [CrossRef]
- Santhirasegaram, S.; Wickramarachchi, S.R.; Attanayake, R.N.; Weerakoon, G.; Samarakoon, S.; Wijeratne, K.; Paranagama, P.A. A novel cytotoxic compound from the endolichenic fungus, Xylaria psidii inhabiting the lichen, Amandinea medusulina. Nat. Prod. Commun. 2020, 15, 1934578X20933017. [Google Scholar] [CrossRef]
- Macías-Rubalcava, M.L.; Sánchez-Fernández, R.E. Secondary metabolites of endophytic Xylaria species with potential applications in medicine and agriculture. World J. Microbiol. Biotechnol. 2017, 33, 15. [Google Scholar] [CrossRef]
- Sica, V.P; Rees, E.R.; Tchegnon, E.; Bardsley, R.H.; Raja, H.A.; Oberlies, N.H. Spatial and temporal profiling of griseofulvin production in Xylaria cubensis using mass spectrometry mapping. Front. Microbiol. 2016, 7, 544. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.N.; Walker, A.K.; Nsiama, T.K.; McFarlane, J.; Sumarah, M.W.; Ibrahim, A.; Miller J., D. Griseofulvin-producing Xylaria endophytes of Pinus strobus and Vaccinium angustifolium: evidence for a conifer-understory species endophyte ecology. Fungal Ecology 2014, 11, 107–113. [Google Scholar] [CrossRef]
- Park, J.H.; Choi, G.J.; Lee, H.B.; Kim, K.M.; Jung, H.S.; Lee, S.W.; Jang, K.S; Cho, K.Y.; Kim, J.C. Griseofulvin from Xylaria sp. Strain F0010, an endophytic fungus of Abies holophylla and its antifungal activity against plant pathogenic fungi. J. Microbiol. Biotechnol. 2005, 15, 112–117. [Google Scholar]
- Ibrahim, A.; Sørensen, D.; Jenkins, H.A.; McCarry, B.E.; Sumarah, M.W. New diplosporin and agistatine derivatives produced by the fungal endophyte Xylaria sp. isolated from Vitis labrusca, Phytochem. Lett. 2014, 9, 179-183. [CrossRef]
- Wicklow, D.T.; Rogers, K.D.; Dowd, P.F.; Gloer, J.B. Bioactive metabolites from Stenocarpella maydis, a stalk and ear rot pathogen of maize. Fungal Biol. 2011, 115, 133–142. [Google Scholar] [CrossRef]
- Zhang, Q.; Xiao, J.; Sun, Q.Q.; Qin, J.C.; Pescitelli, G.; Gao, J.M. Characterization of cytochalasins from the endophytic Xylaria sp. and their biological functions. J. Agric. Food Chem. 2014, 62, 10962–10969. [Google Scholar] [CrossRef]
- Fenta, L.; Mekonnen, H. Microbial biofungicides as a substitute for chemical fungicides in the control of phytopathogens: current perspectives and research directions. Scientifica, 2024, 2024, 12. [Google Scholar] [CrossRef]
- Kaur, R.; Choudhary, D.; Bali, S.; Bandral, S.S.; Singh, V.; Ahmad, M.A.; Rani, N.; Singh, T.G.; Chandrasekaran, B. Pesticides: An alarming detrimental to health and environment. Sci. Total Environ 2024, 915, 170113. [Google Scholar] [CrossRef] [PubMed]
- Daraban, G.M.; Hlihor, R.M.; Suteu, D. Pesticides vs. Biopesticides: From pest management to toxicity and impacts on the environment and human health. Toxics 2023, 11, 983. [CrossRef]
- Hsieh, H.M.; Lin, C.R.; Fang, M.J.; Rogers, J.D.; Fournier, J.; Lechat, C.; Ju, Y.M. Phylogenetic status of Xylaria subgenus Pseudoxylaria among taxa of the subfamily Xylarioideae (Xylariaceae) and phylogeny of the taxa involved in the subfamily. Mol. Phylogenet. Evol. 2010, 54, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Fournier, J.; Flessa, F.; Peršoh, D.; Stadler, M. Three new Xylaria species from Southwestern Europe. Mycol. Prog. 2010, 10, 33–52. [Google Scholar] [CrossRef]
- Sonyot, W.; Lamlertthon, S.; Luangsa-Ard, J.J.; Mongkolsamrit, S.; Usuwanthim, K.; Ingkaninan, K.; Waranuch, N.; Suphrom, N. In vitro antibacterial and anti-inflammatory effects of novel insect fungus Polycephalomyces phaothaiensis extract and its constituents against Propionibacterium acnes. Antibiotics 2020, 9, 274. [Google Scholar] [CrossRef] [PubMed]
- Chesters, N.; O´Hagan, D. Biosynthesis of the fungal metabolite, piliformic acid (2-hexylidene-3-methylsuccinic acid). J. Chem. Soc., Perkin Trans. 1997, 6, 827–834. [Google Scholar] [CrossRef]
- Mangaleswaran, S.; Argade, N. An efficient synthesis of (±)-piliformic acid †. J. Chem. Soc., Perkin Trans. 1, 2000, 3290-3291. [CrossRef]
- Weidenmüller, H.L.; Cavagna, F.; Fehlhaber, H.W.; Präve, P. 2-carboxymethyl-3-n-hexyl-maleic acid anhydride, a novel metabolite from an Aspergillus. Tetrahedron Lett. 1972, 13, 3519–3522. [Google Scholar] [CrossRef]
- Viveki, A.B.; Pol, M.D.; Halder, P.; Sonavane, S.R.; Mhaske, S.B. Annulation of enals with carbamoylpropiolates via NHC-catalyzed enolate pathway: Access to functionalized maleimides/iso-maleimides and synthesis of Aspergillus FH-X-213. J. Org. Chem 2021, 86, 9466–9477. [Google Scholar] [CrossRef]
- Boukouvalas, J.; Thibault, C.; Loach, R. Expedient assembly of bioactive maleic anhydrides using click Diels–Alder chemistry. Sylett. 2014, 25, 2139–2142. [Google Scholar] [CrossRef]
- Becker, K.; Stadler, M. Recent progress in biodiversity research on the Xylariales and their secondary metabolism. J. Antibiot. 2021, 74, 1–23. [Google Scholar] [CrossRef]
- Li, Z.; Park, H.S.; Qiao, J.X.; Yeung, K.S.; Yu, J.Q. Ligand-enabled C–H hydroxylation with aqueous H2O2 at room temperature. ACS. 2022, 144, 18109–18116. [Google Scholar] [CrossRef]
- Chen, S.; Wang, F.F.; Chen, Y.; Li, M.S.; Zhang, B.; Luo, J.Z.; Song, X.X.; Li, J.J.; Qin, F. Chemical constituents of Corydalis saxicola. Chem. Nat. Compd. 2022, 58, 119–121. [Google Scholar] [CrossRef]
- Li, Z.H.; Wang, Q.; Ruan, X.; Pan, C.D.; Jiang, D.A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef] [PubMed]
- Hoang, A.L.; Van, Q.N.; Tuan, N.L.; Dang, X.T. Phenolic allelochemicals: Achievements, limitations, and prospective approaches in weed management. Weed Biol. Manag. 2021, 21, 37–67. [Google Scholar] [CrossRef]
- Elias, L.M.; Fortkamp, D.; Sartori, S.B.; Ferreira, M.C.; Gomes, L.H.; Azevedo, J.L.; Montoya, Q.V.; Rodrigues, A.; Ferreira, A.G.; Lira, S.P. The potential of compounds isolated from Xylaria spp. as antifungal agents against anthracnose. Braz. J. Microbiol. 2018, 49, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Almassi, F.; Ghisalberti, E.L.; Rowland, C.Y. Alkylcitrate-derived metabolites from Aspergillus niger. J. Nat. Prod., 1994, 57, 833–836. [Google Scholar] [CrossRef]
- Koch, L.; Lodin, A.; Herold, I.; Ilan, M.; Carmeli, S.; Yarden, O. Sensitivity of Neurospora crassa to a marine-derived Aspergillus tubingensis anhydride exhibiting antifungal activity that is mediated by the MAS1 protein. Mar. Drugs 2014, 12, 4713–4731. [Google Scholar] [CrossRef]
- Czerniewicz, P.; Chrzanowski, G.; Sytykiewicz, H.; Sprawka, I.; Leszczynski, B. Aphidicidal and deterrent activity of phenolic acid extracts from some herbal plants towards Myzus persicae Sulz. and Rhopalosiphum padi L. Fresenius Environ. Bull. 2016, 25, 5714–5721. [Google Scholar]
- Singh, S.; Kaur, S.; Kaur, R.; Kaur, A. Impact of plant symbiotic endophytic fungus, Aspergillus terreus on insect herbivore Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae). Neotrop. Entomol. 2023, 52, 932–944. [Google Scholar] [CrossRef]
- Kim, S.H.; Park, D.H.; Choi, J.Y.; Wang, M.; Liu, S.; Je, Y.H. Characterization of insecticidal compound from Streptomyces gramineus against Thrips palmi. , J. Asia. Pac. Entomol. 2023, 26, 102166. [Google Scholar] [CrossRef]
- Cao, L.; Yan, W.; Gu, C.; Wang, Z.; Zhao, S.; Kang, S.; Khan, B.; Zhu, H.; Li, J.; Ye, Y. New alkylitaconic acid derivatives from Nodulisporium sp. A21 and their auxin herbicidal activities on weed seeds. J. Agric. Food Chem. 2019, 67, 2811–2817. [Google Scholar] [CrossRef]
- Mondal, G.; Dureja, P.; Sen, B. Fungal metabolites from Aspergillus niger AN27 related to plant growth promotion. Indian J. Exp. Biol. 2000, 38, 84–87. [Google Scholar] [PubMed]
- Vargas-Hernandez, M.; Macias-Bobadilla, I.; Guevara-Gonzalez, R.G.; Romero-Gomez, S.J.; Rico-Garcia, E.; Ocampo-Velazquez, R.V.; Alvarez-Arquieta, L.L.; Torres-Pacheco, I. Plant hormesis management with biostimulants of biotic origin in agriculture. Front. Plant Sci. 2017, 8, 1762. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Blain, R.B. Hormesis and plant biology. Environ. Pollut. 2009, 157, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol Biol Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Morales-Sánchez, V.; Díaz, C. E.; Trujillo, E.; Olmeda, S.A.; Valcarcel, F.; Muñoz, R.; Andrés, M.F.; González-Coloma, A. Bioactive metabolites from the endophytic fungus Aspergillus sp. SPH2. J. Fungi. 2021, 7, 109. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC bioinformatics 2017, 18, 529. [Google Scholar] [CrossRef]
- Truzi, C.C.; Vieira, N.F.; De Souza, J.M.; De Bortoli, S.A. Artificial diets with different protein levels for rearing Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Insect Sci. 2021, 21, 1–7. [Google Scholar] [CrossRef]
- González-Coloma, A.; Andres, M.F.; Contreras, R.; Zúñiga, G.E.; Díaz, C.E. Sustainable production of insecticidal com-pounds from Persea indica. Plants 2022, 11, 418. [Google Scholar] [CrossRef]
- Andrés, M.F.; González-Coloma, A.; Sanz, J. ; Burillo, J; & Sainz, P. Nematicidal activity of essential oils: A review. Phytochem. Rev. 2012, 11, 371–390. [Google Scholar] [CrossRef]
- Julio, L.F.; Burillo, J.; Giménez, C.; Cabrera, R.; Díaz, C.E.; Sanz, J.; González-Coloma, A. Chemical and biocidal characterization of two cultivated Artemisia absinthium populations with different domestication levels. Ind. Crops Prod. 2015, 76, 787–792. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic maximum likelihood tree (T92+G) of Xylaria sp. (HYP6) based on selected ITS rDNA sequences obtained from GenBank. Confidence values from a 5000-replicate bootstrap analysis are shown at each branch nodes.
Figure 1.
Phylogenetic maximum likelihood tree (T92+G) of Xylaria sp. (HYP6) based on selected ITS rDNA sequences obtained from GenBank. Confidence values from a 5000-replicate bootstrap analysis are shown at each branch nodes.

Figure 2.
Compounds of endolichenic Xylaria sp. (HYP6).
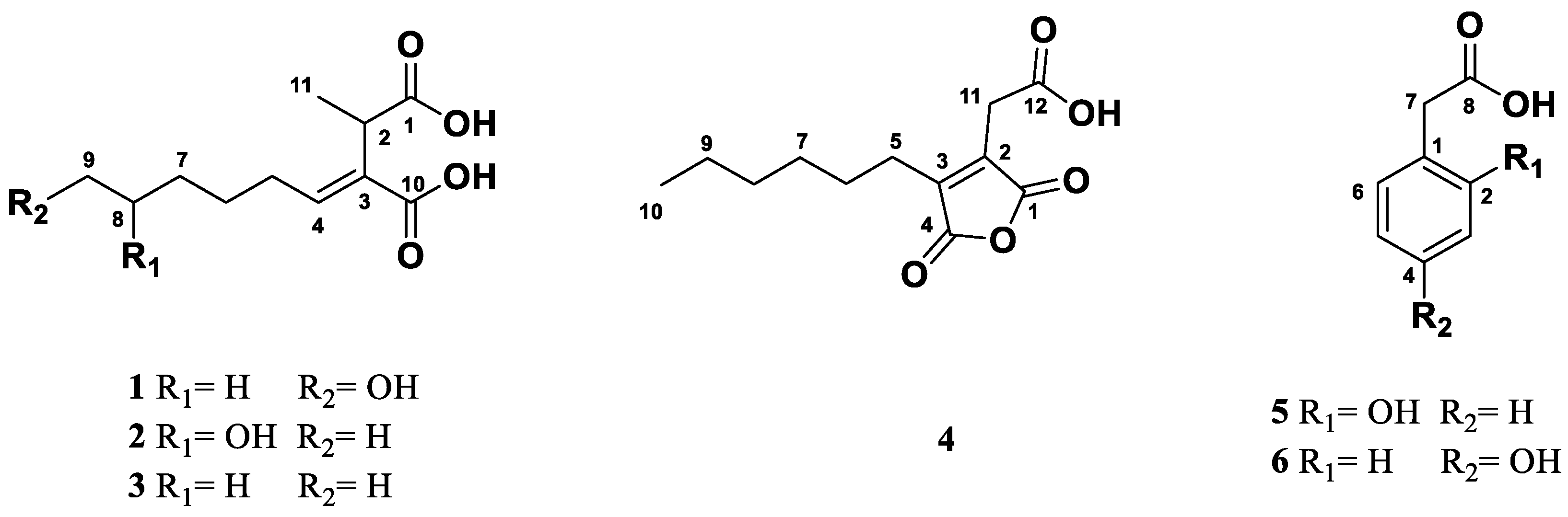
Figure 3.
Key 1H-1H COSY and selected HMBC correlations of compounds 1 and 2.
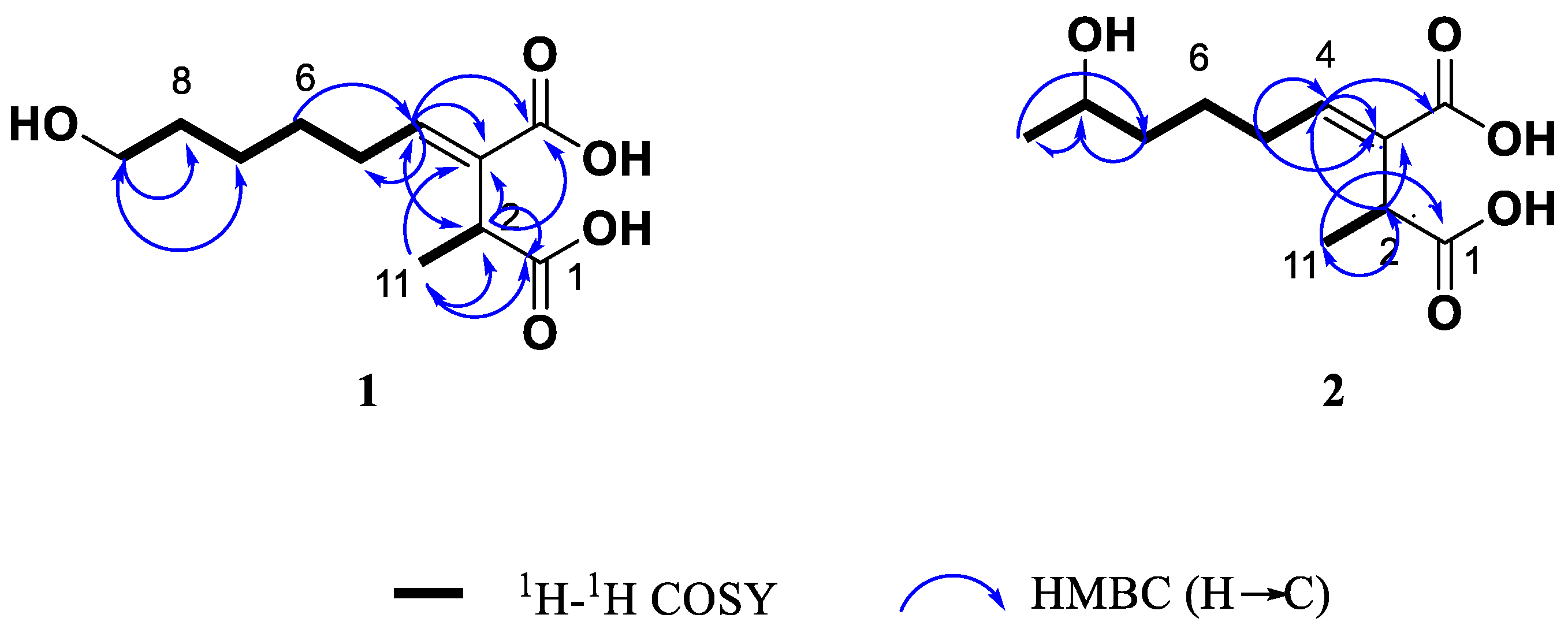
Figure 4.
Phytotoxic effects of extract (EtOAc) and compounds (2-6) on Lolium perenne and Lactuca sativa (doses tested at 0.4 and 0.2 mg/ml for the extract and 2-6 respectively).
Figure 4.
Phytotoxic effects of extract (EtOAc) and compounds (2-6) on Lolium perenne and Lactuca sativa (doses tested at 0.4 and 0.2 mg/ml for the extract and 2-6 respectively).
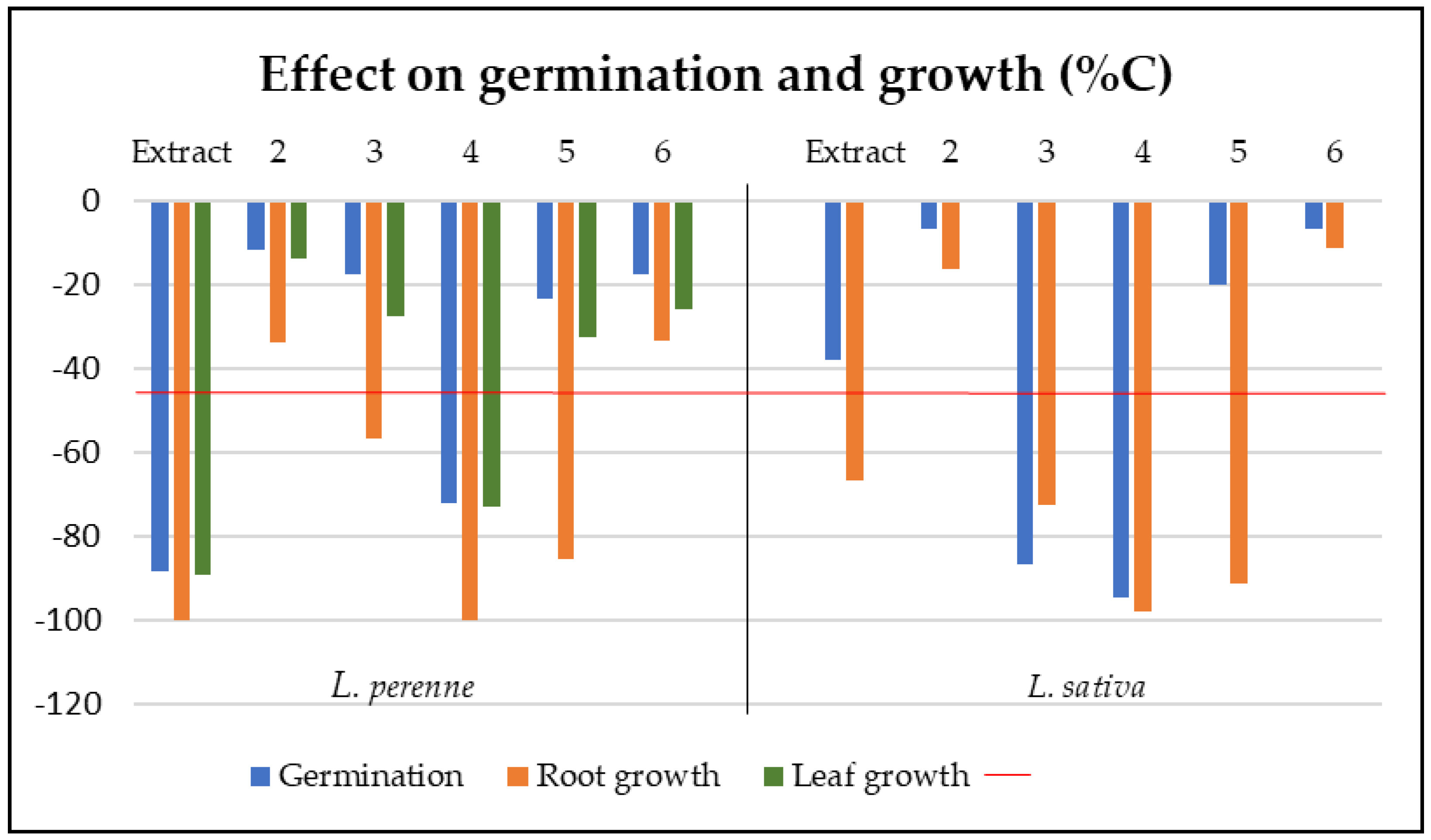

Table 1.
H NMR and 13C NMR spectra of compounds 1 and 2.
| Position | 1 | 2 | ||
|---|---|---|---|---|
| δH, mult (J in Hz) |
δC | δH, mult (J in Hz) |
δC | |
| 1 | - | 178.0 | - | 177.8 |
| 2 | 3.63, q (7.1) | 39.0 | 3.64, qd (7.1, 1.9) | 38.7 |
| 3 | - | 134.5 | 134.3 | |
| 4 | 6.82, t (7.6) | 144.7 | 6.85, t (7.6) | 145.0 |
| 5 | 2.26, m | 29.4 | 2.26, m | 29.4, (29.4)a |
| 6 | 1.51, m | 29.6 | 1.61, m 1.52, m |
26.0, (26.0) a |
| 7 | 1.43, m | 26.7 | 1.50, m | 39.6, (39.7) a |
| 8 | 1.56, m | 33.4 | 3.74, sext (6.1) | 68.2,(68.3) a |
| 9 | 3.55, t (6.6) | 62.8 | 1.16, d (6.1) | 23.5 |
| 10 | - | 170.3 | - | 169.9 |
| 11 | 1.31, d (7.2) | 16.4 | 1.31, d (7.1) | 16.3 |
a data in () for stereoisomer of hydroxyl group in n-alkyl chain.
Table 2.
Biocidal effects of extract (EtOAc) and compounds (2–6) against insect pests (Spodoptera littoralis, Myzus persicae, Rhoplalosiphum padi), the nematode Meloidogyne javanica and phytopathogenic fungi (Alternaria alternata, Botrytis cinerea and Fusarium oxysporum).
Table 2.
Biocidal effects of extract (EtOAc) and compounds (2–6) against insect pests (Spodoptera littoralis, Myzus persicae, Rhoplalosiphum padi), the nematode Meloidogyne javanica and phytopathogenic fungi (Alternaria alternata, Botrytis cinerea and Fusarium oxysporum).
| Target | Effect | Treatment | Activity (%) | EC50c/LD50d |
|---|---|---|---|---|
| S. littoralis | %FIa | Extract | 42.9±11.4 | |
| 2 | 33.0±14.9 | |||
| 3 | 18.0±8.4 | |||
| 4 | 37.8±18.7 | |||
| 5 | 9.1±6.3 | |||
| 6 | 17.7±11.7 | |||
| M. persicae | %SIa | Extract | 81.5±6.8 | 10.9 (7.2-16.4) |
| 2 | 25.5±6.3 | |||
| 3 | 50.4±8.9 | |||
| 4 | 100.0±0.0 | 1.6 (0.5-4.9) | ||
| 5 | 98.9±0.6 | 4.5 (2.0-10.0) | ||
| 6 | 91.9±2.3 | 15.5 (12.0-20.0) | ||
| R. padi | %SIa | Extract | 61.5±6.0 | |
| 2 | 20.6±6.4 | |||
| 3 | 30.3±8.8 | |||
| 4 | 91.0±3.8 | 8.9 (4.6-7.5) | ||
| 5 | 61.2±9.2 | |||
| 6 | 43.0±7.9 | |||
| M. javanica | %Mortalityb | Extract | 2.0±0.7 | |
| 2 | 2.2±0.7 | |||
| 3 | 28.4±6.6 | |||
| 4 | 100.0±0.0 | 0.10 (0.10-0.11) | ||
| 5 | 26.4±5.8 | |||
| 6 | 29.5±5.6 | |||
| A. alternata | % MGIb | Extract | 58.5±2.4 | |
| 2 | 27.3±3.7 | |||
| 3 | 47.3±5.1 | |||
| 4 | 63.7±1.4 | 0.24 (0.20-0.28) | ||
| 5 | 32.2±5.6 | |||
| 6 | 42.5±6.0 | |||
| B. cinerea | % MGIb | Extract | 75.9±9.7 | |
| 2 | 19.7±6.2 | |||
| 3 | 62.1±7.6 | 0.36 (0.25-0.51) | ||
| 4 | 84.3±4.5 | 0.12 (0.09-0.15) | ||
| 5 | 0.5±8.8 | |||
| 6 | 5.8±6.2 | |||
| F. oxysporum | % MGIb | Extract | 61.9±6.5 | |
| 2 | 18.7±3.1 | |||
| 3 | 42.3±3.6 | |||
| 4 | 52.3±2.6 | |||
| 5 | 32.6±9.1 | |||
| 6 | 35.7±3.3 |
a%FI and %SI, doses tested of 100μg/cm2 for the extract and 50.0 μg/cm2 for the compounds on insects. b% Mortality and %MGI doses tested of 1 mg/ml for the extract and 0.5 mg/ml for the compounds. cEfficient dose (μg/cm2) to give 50% feeding inhibition or efficient dose (mg/ml) to give 50% mycelial growth inhibition and 95% confidence limits (lower, upper). dLethal Dose (mg/ml) to give 50% mortality of M. javanica and 95% confidence limits (lower, upper).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



