
Submitted:
31 October 2024
Posted:
01 November 2024
You are already at the latest version
Abstract
The white poplar (Populus alba L.) is an economically significant tree species with a natural distribution spanning extensive region of Eurasia. Nevertheless, there is currently no hypothesis regarding the historical shaping of this range. In this study, we collected and sequenced 36 individuals of white poplar from disparate regions of Russia and Kazakhstan. Additionally, we employed available genomic data of white poplars from Italy, Hungary, and China. A genomic approach was employed to collate data on the location of glaciers in different periods, along with information on the natural and artificial distribution of white poplar. This enabled the formulation of the first hypothesis regarding the formation of the modern range of this plant. It is hypothesized that during the period of maximum glaciation, three refugia existed: the South European, Transcaucasian, and Altai-Middle Asian refugia. Post-glacial migration from these refugia led to the formation of modern populations of P. alba in Eastern Europe (including the European part of Russia), the Caucasus, and Siberia, respectively.
Keywords:
Populus alba
; natural range
; historical formation
; Russia
; Europe
; genomics
; Quaternary glaciation
; glaciers
1. Introduction
Poplars are a widespread genus of woody plants with pronounced economic value. They have long been used as a source of pulp and wood, and the International Poplar Commission (IPC) was established in 1947 to promote the sustainable management of fast-growing trees by its 38 member countries [1]. At the same time, due to their fast growth, poplars can be used to combat climate change as they store atmospheric carbon in their wood [2,3]. Poplars have traditionally been used for landscaping rural and urban areas, including in Russia. This is partly due to their ability to withstand heavy soil pollution, which allows poplar to be used for phytoremediation, cleaning both urban and industrial soils [4,5]. Finally, poplars are used by scientists as model objects among trees - for example, Populus trichocarpa became the first tree the genome of which was sequenced in 2006 [6]. Due to all these reasons, the geographical distribution of selected valuable poplar species has become wider, through introduction in selected regions.
White poplar (Populus alba) is a well-known member of the genus with a native range from Central and Southern Europe to Xinjiang and the Western Himalayas (https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:776573-1, accessed 13 October, 2024), and is currently present on five continents due to its biological characteristics that allow it to be grown as a source of cellulose and lignin or for landscaping in urban areas. Poplars are dioecious plants, much less common than monoecious plants. The ARABIDOPSIS RESPONSE REGULATOR 17 (ARR17) gene, located in the sex-determining region (SDR), is the main regulator of sex, which ensures female phenotype development [7], while male development is regulated by suppression of this gene expression [8]. White poplar is an exception in the sense that males do not have genetic mechanisms to suppress ARR17 expression, but instead have a deletion of this gene [9], making it possible to use genome editing tools to change the sex of a tree by inserting or deleting the gene, which could allow the cultivation of trees of the preferred sex to, for example, reduce economic losses by reducing the number of trees of the preferred sex.
The complex history of white poplar range formation, including the reduction of its habitat area during the glacial period and the subsequent development of new territories, has led to the formation of a large intraspecific genetic diversity, which arose both because of range disjunction and the need to adapt to the conditions of very diverse regions in terms of climate and biota. Populations from different regions are characterized by very contrasting indicators of growth rate, winter hardiness, frost resistance, drought tolerance, salt tolerance, as well as resistance to pests and pathogens, primarily those that cause stem rot. In addition, populations of different origins differ in their ability to vegetative reproduction and the set of symbiotic microorganisms, which is very important in connection with the increasing use of biotechnology methods in the cultivation of these tree species. The current intraspecific diversity has both great practical applications in breeding work and can be used to reconstruct the history of white poplar dispersal after the retreat of glaciers, to identify primary and secondary ranges of the species, the history of origin and dispersal of individual forms, and to solve problems of intraspecific systematics.
In this study, we present the first insights into the formation of the modern P. alba region during the postglacial era, based on the genomes of 36 white poplar individuals collected in Russia and Kazakhstan, and using available genomic data of white poplars from Italy, Hungary, and China. Our findings shed light on the evolutionary history of this species in the East European Plain, the Caucasus, and Western Siberia. The data may also be employed to model the range dynamics of other species of the genus in Eurasia and North America, as poplars exhibit analogous strategies in the development of their populations.
2. Results
To understand the process of modern white poplar range formation, we used a genomic approach. To do so, we collected genome data from plants originating from a range of countries and geographic locations and then evaluated clusters of genetically similar samples based on SNP profiling. First, we mapped the reads to the reference genome of white poplar and then performed SNP calling as described in the Materials and Methods section. For the most samples (73 of 87) more than 85% reads were successfully mapped to the reference sequence. About 23 million SNPs (at least in one sample) were identified. Based on the profiles of per-sample SNP occurrence, we performed PCA analysis (Figure 1). The phylogenetic tree corresponding to this graph is shown in Supplementary Figure S1.
Based on this PCA graph, we identified three clusters. First, samples collected in the Caucasus (in the vicinity of Sochi, Pyatigorsk, and in Dagestan) were most strongly separated from all other points and formed a separate cluster. Chinese (growing in Xinjiang and the Irtysh River valley) and Siberian trees were also grouped separately. All other plants formed a large third cluster. We will now note that the Italian specimens are furthest away in it, as well as plants from Beijing (which were imported). And 6 poplars, 4 of which were pyramidal (these are all pyramidal poplars in our study), formed a separate subcluster inside this third cluster.
Having studied the PCA plot, we hypothesized that there were probably 3 ancestral populations according to the 3 identified clusters. Based on these genotypes, the entire modern diversity of white poplars could have been formed. To test this hypothesis, we used the NGSAdmix program as written in the Materials and Methods section, the results of which are presented in Figure 2.
We also used NGSAdmix with the number of genotypes k = 2, 4, 5, and 6, and the results are shown in Supplementary Figures 1, 2, 3, and 4, respectively. However, while at k = 3 the split corresponds to geography (Europe, Caucasus, and Chinese regions), at k = 4 Chinese poplars split into two genotypes, and at k = 5 a cluster appears in the Southern Federal District, where it is unlikely that a refugium existed during glaciation due to the harsh climatic conditions. Thus, we hypothesize that there were three major refugia in terms of the global population of P. alba.
We interpreted the results of this analysis as follows. A1 is the most probable ancestor of poplars in China, largely in Western Siberia, and has also contributed to the gene pool of plants in the vicinity of Uryupinsk. A2 is a key ancestor of trees in the Transcaucasia and Caucasus, and its participation in the artificial settlement of CFD territories and the creation of pyramidal forms is also notable. A3 is an ancestor of poplars from Italy, a key member of the gene pool of the central and lower Volga region, secondary area of central part of Russia, as well as contributing to populations in south of Western Siberia and the Caucasus and to the creation of pyramidal forms, known as the variation pyramidata sovietica. We reveal our theory of white poplar distribution in more detail in the discussion section.
3. Discussion
The family Salicaceae separated about 128 million years ago. It includes 54 genera and about 1,400 species, with most genera represented by a small number of species and distributed in Southeast Asia, the likely center of origin of the family [10]. The most evolutionary successful genera are Salix and Populus. Their common ancestor about 60-65 million years ago underwent Salicoid whole genome duplication, which affected approximately 92% of the genome and resulted in more than 8000 pairs of paralogous genes [6]. This is probably why the genera Salix and Populus were able to spread almost all over the Northern Hemisphere, especially in the boreal regions, and now number around 450 and 50 species, respectively [11,12].
The genus Populus is divided into four sections: Abaso, Turanga, Populus and ATL, the latter including representatives of the traditional sections Aigeiros, Tacamahaca and Leucoides. Abaso is the most primitive section, including P. mexicana, common in Mexico. Turanga includes P. euphratica, known to grow in deserts in the Middle East and named after the Euphrates River. Populus and ATL are the most evolutionary advanced groups. Populus includes the aspens: P. tremula, P. tremuloides, as well as P. alba, P. tomentosa, P. qiongdaoensis, and some other poplars. The ATL group includes P. trichocarpa, P. balsamifera, P. deltoides, P. nigra, P. lasiocarpa, and others [13]. Interestingly, poplars, like willows, are dicotyledonous plants, and some representatives of the Populus section, including P. alba, have a ZW system of sex determination, while most poplars are characterized by an XY system [14,15]. Moreover, only 2 species are tropical in the genus Populus, namely P. qiongdaoensis and P. ilicifolia, distributed in Hainan and Africa (Kenya and Tanzania), respectively [15]. P. trichocarpa and P. balsamifera are distributed in temperate forests of North America. P. trichocarpa is spread along the west coast of the United States and Canada, while P. balsamifera occurs somewhat northward, from Alaska to Labrador; however, there is a significant degree of hybridization between these closely related species in the habitat overlap zone [16,17,18,19]. P. tomentosa is a Chinese endemic [20]. The range of P. alba extends from Western Europe to Altai and Xinjiang, bounded in the north by mixed forests and taiga, where the climate is too cold for it, and in the south by steppes, semi-deserts and deserts, where it is too dry, as can be seen in Figure 3. P. alba did not spread across the Altai, probably due to the harsh sharply continental climate of Eastern Siberia and competition from other species [21,22,23,24].
We did not find articles describing a possible scenario for the formation of the modern range and population structure of P. alba, there was only a research devoted to Central and Southern Europe. It was shown that the two main refugia in this region were in Italy and Romania. At the same time, P. tremula survived in more northerly and harsh conditions, near the ice shield of the glaciated Alps. When the glaciers retreated, the ranges of both species expanded, and in the contact zone they began to hybridize actively, forming the natural hybrid P. x canescens [25]. White poplars in China, growing in the Irtysh River basin, have also been studied. Interestingly, the heterozygosity of populations there is lower than in Italy and Hungary [26]. In addition, in this region, as in Europe, P. alba also hybridized significantly with P. tremula. This process, in turn, was significantly limited by plastid-nuclear incompatibility [27].
We will also consider analogous research for two other species, namely P. cathayana in China and P. balsamifera in Canada and USA, just for example.
A recent study of P. cathayana revealed 4 genetically distinct populations, named by the authors for growing in the Southwest (NW), Northwest (SW), and North China (NC), and the Taihang Mountains (TH). Splitting of the ancestral population began 1649 thousand years ago (kya), with one branch dividing 1430 kya into TH and NW and the other branch splitting 987 kya into NC and SW. The distribution and adaptation of P. cathayana has been linked to climate changes [28].
Another interesting evolutionary story concerns the postglacial distribution of P. balsamifera in North America after the last glacial maximum, which occurred about 18,000 years ago. The authors identified three demes, namely the Central, Northern, and Eastern ones. The Central deme is the most genetically diverse, occupies the largest area, and is probably directly related to the ancestral population in the refugium. The Northern deme occupies Alaska, and the Eastern deme inhabits Quebec and Labrador [18]. In another paper, it was shown that the boundary between the Eastern and Central demes is clearer than between the Central and Northern demes (there referred as Western), which can probably be explained by the earlier separation of the Eastern deme from the general evolutionary branch [19].
Quaternary glaciation appears to have been a significant force in shaping the present-day range of P. alba. It began 2.58 million years ago and has continued to modern times, with periods of glaciation alternating with much warmer periods of interglaciation [29,30]. For example, the last glaciation ended 11,700 years ago, followed by the Holocene, which is an interglacial period [31,32], and the next glaciation is projected to begin in 50,000 years [33]. During the maximum glaciation glaciers were in the northern part of Europe (most of Great Britain, the territory of modern Germany, Poland, Belarus, northern Ukraine and part of Russia), also the centers of glaciation were mountain systems: the Alps and the Caucasus. But in Eastern Siberia glaciation also occurred but covered quite a little percentage of all surface - probably, the dry continental climate did not allow the formation of a glacier [34,35,36].
Based on our findings and all the above information, as well as on the data on the current distribution of white poplar we formulated a hypothesis of how the formation of its modern range occurred, which is shown in Figure 3, and the numbers in this section of the text coincide with the numbers in the figure for ease of perception.
In the late Pliocene, about 2.5 million years ago, a single population of white poplar probably existed across vast territories of Eurasia. However, after a series of glaciations in the Pleistocene, this tree became extinct in a significant part of its historical range, surviving, among others, in three regions on the territory of Russia and neighboring countries. These regions, those we consider as refugia, were never covered by glaciers and correspond to putative ancestors A1, A2, and A3. The first (5) is the Altai Region and mountains of Central Asia, located in present-day Russia, Kazakhstan, China (Xinjiang), Kyrgyzstan, Uzbekistan, Tajikistan and Afghanistan, which matches A1. The second (4) is Transcaucasia (since the Caucasus Mountains themselves were covered by glaciers at that time and the southern coast of the Caspian Sea on the territory of modern Azerbaijan, Armenia, eastern Turkey and northern Iran, it conforms to ancestor A2. The third region (3) was in the north of Africa and in the south of Europe, on the Iberian, Apennine and Balkan peninsulas, on some large islands, including Sicily, Sardinia and Crete, and on peninsula Asia Minor, settling on the territory of modern Spain, France, Italy, Croatia, Greece, Turkey and some other countries, and corresponded to ancestor A3. We named these regions Altai-Middle Asian refugium, Transcaucasian refugium, and South European refugium, respectively.
When glaciers began to retreat, white poplars started to spread outside these refugia. It was the third, Southern European refugium that became the most important for the formation of poplars in the European part of Russia. Migrations (6) from it in the northeastern direction led to the settlement of a significant part of the East European Plain, including the vicinities of Nizhny Novgorod and Uryupinsk. At the same time, steppes (10) in the south of modern Ukraine and the Pre-Caucasus, as well as semi-deserts and deserts (11) in the territory of modern Russia, Kazakhstan and other countries became a natural obstacle to the spread of poplar in certain directions. It penetrated the respective regions only along river valleys (12), such as the Don (12a), Volga (12b), and Ural (12c). From the European part of Russia, poplar migrated further eastward to Western Siberia, making a certain contribution to the gene pool of this region, as can be seen from the results of genomic analysis of samples collected near Novosibirsk in Figure 3. In parallel with this migration, plants from the Black Sea coastline of Turkey reached the Caucasus, when the glaciers retreated from here. This event had an impact on the local genofond, as some of the Caucasian samples are also carriers of European genes.
The main contribution to the gene pool of modern populations of white poplar in the Caucasus comes from the Transcaucasian refugium. When the glacier in the Caucasus receded, trees from Transcaucasia penetrated northward (7), but did not spread further due to steppe and semi-desert zones north of the Caucasus Mountains.
The Altai-Middle Asian refugium may have expanded somewhat (8) after the end of glaciation; the poplars living there are the ancestors of modern Chinese plants and, to a large extent, the poplars of Western Siberia. There was also their migration westward, at least to the East European Plain, since all plants collected in the vicinity of Uryupinsk are carriers of Altai genes.
We separately note that although the plants we collected in the Central Federal District (Moscow and Moscow Region, Obninsk, Tula, etc.) are genetically close to other European populations and also have a certain percentage of Caucasian genes, in these regions, poplars appeared as a result of introduction in the nineteenth and twentieth centuries (9), so this part of their habitat cannot be called natural.
All pyramidal poplars, two of which were collected by us in Kazakhstan and two in CFD, are artificially bred based on European and Caucasian genotypes and are genetically relatively close to each other.
Therefore, we consider Southern Europe, Transcaucasia and the Altai-Middle Asian system as potential refugia where poplar could have survived during glaciations. During glacial retreat, poplar spread to regions suitable for it in terms of climatic conditions. This probably happened repeatedly. Our study is the first hypothesis about the formation of the modern range of white poplar. Further research in this direction involving more data is required.
4. Materials and Methods
4.1. Plant Material
A collection of samples was obtained from 36 trees, representing a range of geographical conditions, for the purposes of the study. The study encompassed a multitude of regions within the European part of the Russian Federation, including the Central Federal District, Volga Federal District, Southern Federal District, and North Caucasus Federal District. Additionally, two specimens were procured from the Republic of Kazakhstan, and three were obtained from the collection of the Central Siberian Botanical Garden of the Siberian Branch of the Russian Academy of Sciences (CSBS SB RAS), situated in the vicinity of the city of Novosibirsk. The geographical distribution of the collected samples is illustrated in Figure 4. To ensure accurate sex determination, samples were collected during the flowering period of poplars.
Branches were cut from the trees and transported for two days before DNA extraction. Leaf material for DNA isolation was collected in laboratory conditions and frozen in 2 ml eppendorf tubes at -70 C.
4.2. DNA Isolation, Library Preparation, and Whole-Genome Sequencing
For DNA isolation, 0.2 g of leaf material was homogenized using a MagNA Lyser automated homogenizer (Roche, Switzerland) in 1 ml of lysis modified CTAB buffer (100 mM Tris-HCl pH 8. 0; 3% CTAB; 3M NaCl; 20 mM EDTA pH 8.0; 1% PVP K30) with 5 μl β-mercaptoethanol and solid beads. The homogenate was incubated at 65°C for 1 h, stirring every 20 min. Two consecutive chloroform purifications were then performed: an equal volume of chloroform was added to the homogenate and centrifuged for 15 min at 10,000 g and 20°C. The aqueous phase was transferred to clean tubes, two volumes of 96% alcohol were added to precipitate DNA and incubated for 60 min at -20°C. Then centrifuged for 20 min at 10,000 g and 4°C. The supernatant was carefully removed without touching the precipitate. The precipitate was washed three times with 70% alcohol and centrifuged for 5 min at 10,000 g and 4°C. DNA was quantified using a Qubit®2.0 fluorometer (Thermo Fisher Scientific, Waltham, MA, USA); quality control was performed on a NanoDrop® ND-1000 spectrophotometer (NanoDrop Technologies Inc., Wilmington, DE, USA).
The A260/A280 ratio in the DNA samples was 1.8-2.0. Approximately 500 ng of genomic DNA was fragmented into 400 bp double-stranded fragments using the Covaris S220 system (Covaris Inc., Woburn, MA, USA). Double-stranded DNA library was prepared using the VAHTS Universal Plus DNA Library Prep Kit for Illumina V2 (Nanjing Vazyme Biotech Co., Red Maple Hi-tech Industry Park, Nanjing, PRC) according to the manufacturer’s recommendations. AMPure XP beads were used to select DNA libraries by size (500-600 bp). Samples were quantified at standard concentration dilution to 4 nM using a Qubit®2.0 fluorometer (Thermo Fisher Scientific, Waltham, MA, USA) and verified on an Agilent 2100 Bioanalyzer using the High Sensitivity DNA Kit (Agilent Technologies, Santa Clara, CA, USA). Genomic DNA of bacteriophage PhiX was used as an internal control. Sequencing was performed using the NextSeq 500/550 High Output Kit v2.5 (300 cycles) on the Illumina NextSeq 500 platform (Illumina, San Diego, CA, USA) according to the manufacturer’s instructions.
4.3. Read Mapping and SNP-Based Analysis
A phylogenetic analysis was conducted based on SNPs profiling, including point SNVs and short insertions/deletions (indels). Reads were mapped to the white poplar reference genome (GCF_005239225) using Bowtie2 [37] with increased sensitivity parameters -D 20 -R 3 -N 0 -L 20 -i S,1,0.50). Secondary (supplementary) alignments were removed with samtools. FreeBayes was used for variant calling [38]. Using per-sample variant allele frequency (VAF) data, principal components analysis (PCA) was performed to identity groups of genetically related samples. Also, the Euclidean pairwise distance matrix between samples was calculated for variant allele frequency (VAF) vectors, and samples were clustered using UPGMA algorithm (with bootstrapping), thus considering the possibility of polyploidy.
The assessment of population mixing was performed using NGSAdmix software, based on the assessment of the probability of genotypes for all SNPs (-K 3) [39]. For the accuracy of the analysis, additional samples of white poplar trees were used, growing in the Xinjiang province of China, as well as in Italy and Hungary. The raw genomic data for these samples was obtained from the NCBI SRA (accession numbers can be found in Supplementary Table 1). These data correspond to several research articles [9,12,40,41,42,43].
5. Conclusions
In this study, using a genomic approach and data from different regions of Russia, as well as Italy, Hungary, Kazakhstan, and China, we first hypothesized the formation of the modern range of white poplar. Probably, before glaciation, the range of P. alba was uniform and then divided into three refugia: South European, Transcaucasian, and Altai-Middle Asian, meaning that poplar was preserved in places where glaciers did not occur. After the glaciers retreated, populations from these refugia gave rise to modern poplar populations in Europe, the Caucasus, and Siberia, respectively, which eventually spread and restored the integrity of the area. To confirm and refine this hypothesis, further studies aimed at increasing the number of samples and expanding the study areas are needed.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Figure S1 depicts the phylogenetic tree corresponding to the PCA plot in Figure 1. Supplementary Figures S2, S3, S4, and S5 show NGSAdmix results with k = 2, 4, 5, and 6, respectively. Supplementary Table S1 contains information about reference P. alba genomes used in our study.
Author Contributions
Conceptualization, M.I.P., N.S.G., M.S.F. and A.V.K.; methodology, M.I.P., N.S.G. and V.V.V.; software, A.V.K.; validation, G.S.K., A.I.S. and A.S.B.; formal analysis, V.V.V.; investigation, M.A.K. and A.V.K.; resources, A.V.K.; data curation, G.S.K. and M.A.K.; writing—original draft preparation, M.I.P., N.S.G. and M.A.K.; writing—review and editing, D.S.K. and A.V.K.; visualization, G.S.K. and M.A.K.; supervision, A.V.K.; project administration, A.V.K. and N.L.B.; funding acquisition, N.L.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported financially by the Russian Science Foundation, grant number 22-14-00404 (collection of P. alba samples and their genomic sequencing (Figure 1 and Figure 4, Table 1)) and by the Ministry of Science and Higher Education of the Russian Federation, grant no. 075-15-2019-1660 (Bioinformatics analysis of P. alba genomes (Figure 2 and Figure 3).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data obtained in our study are in the public domain. Whole genome sequencing results for 36 P. alba individuals is available at NCBI under accession number https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1109753.
Acknowledgments
This work was performed using the equipment of EIMB RAS “Genome” center (http://www.eimb.ru/ru1/ckp/ccu_genome_ce.php).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Overview | International Commission on Poplars and Other Fast-Growing Trees Sustaining People and the Environment | Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/ipc/overview/en/ (accessed on 14 October 2024).
- Liberloo, M.; Calfapietra, C.; Lukac, M.; Godbold, D.; Luo, Z.; Polle, A.; Hoosbeek, M.R.; Kull, O.; Marek, M.; Raines, C.; et al. Woody Biomass Production during the Second Rotation of a Bio-energy Populus Plantation Increases in a Future High CO 2 World. Global Change Biology 2006, 12, 1094–1106. [Google Scholar] [CrossRef]
- Macaya-Sanz, D.; Chen, J.; Kalluri, U.C.; Muchero, W.; Tschaplinski, T.J.; Gunter, L.E.; Simon, S.J.; Biswal, A.K.; Bryan, A.C.; Payyavula, R.; et al. Agronomic Performance of Populus Deltoides Trees Engineered for Biofuel Production. Biotechnol Biofuels 2017, 10, 253. [Google Scholar] [CrossRef] [PubMed]
- Tőzsér, D.; Horváth, R.; Simon, E.; Magura, T. Heavy Metal Uptake by Plant Parts of Populus Species: A Meta-Analysis. Environ Sci Pollut Res 2023, 30, 69416–69430. [Google Scholar] [CrossRef] [PubMed]
- Di Lonardo, S.; Capuana, M.; Arnetoli, M.; Gabbrielli, R.; Gonnelli, C. Exploring the Metal Phytoremediation Potential of Three Populus Alba L. Clones Using an in Vitro Screening. Environ Sci Pollut Res 2011, 18, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The Genome of Black Cottonwood, Populus Trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef]
- Cronk, Q.; Müller, N.A. Default Sex and Single Gene Sex Determination in Dioecious Plants. Front. Plant Sci. 2020, 11, 1162. [Google Scholar] [CrossRef]
- Leite Montalvão, A.P.; Kersten, B.; Fladung, M.; Müller, N.A. The Diversity and Dynamics of Sex Determination in Dioecious Plants. Front. Plant Sci. 2021, 11, 580488. [Google Scholar] [CrossRef]
- Müller, N.A.; Kersten, B.; Leite Montalvão, A.P.; Mähler, N.; Bernhardsson, C.; Bräutigam, K.; Carracedo Lorenzo, Z.; Hoenicka, H.; Kumar, V.; Mader, M.; et al. A Single Gene Underlies the Dynamic Evolution of Poplar Sex Determination. Nat. Plants 2020, 6, 630–637. [Google Scholar] [CrossRef]
- Ogutcen, E.; De Lima Ferreira, P.; Wagner, N.D.; Marinček, P.; Vir Leong, J.; Aubona, G.; Cavender-Bares, J.; Michálek, J.; Schroeder, L.; Sedio, B.E.; et al. Phylogenetic Insights into the Salicaceae: The Evolution of Willows and Beyond. Molecular Phylogenetics and Evolution 2024, 199, 108161. [Google Scholar] [CrossRef]
- Zhang, Z.-S.; Zeng, Q.-Y.; Liu, Y.-J. Frequent Ploidy Changes in Salicaceae Indicates Widespread Sharing of the Salicoid Whole Genome Duplication by the Relatives of Populus L. and Salix L. BMC Plant Biol 2021, 21, 535. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, J.; Li, E.; Xu, S.; Zhan, Z.; Zhang, X.; Yang, Z.; Guo, F.; Liu, K.; Liu, D.; et al. Phylogenomics and Biogeography of Populus Based on Comprehensive Sampling Reveal Deep-Level Relationships and Multiple Intercontinental Dispersals. Front. Plant Sci. 2022, 13, 813177. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, L.; Zhang, Z.; Li, M.; Wang, D.; Zhang, X.; Xi, Z.; Keefover-Ring, K.; Smart, L.B.; DiFazio, S.P.; et al. Phylogenomics of the Genus Populus Reveals Extensive Interspecific Gene Flow and Balancing Selection. New Phytologist 2020, 225, 1370–1382. [Google Scholar] [CrossRef] [PubMed]
- Gladysh, N.S.; Kovalev, M.A.; Lantsova, M.S.; Popchenko, M.I.; Bolsheva, N.L.; Starkova, A.M.; Bulavkina, E.V.; Karpov, D.S.; Kudryavtsev, A.A.; Kudryavtseva, A.V. The Molecular and Genetic Mechanisms of Sex Determination in Poplar. Mol Biol 2024, 58, 178–191. [Google Scholar] [CrossRef]
- Li, Y.; Wang, D.; Wang, W.; Yang, W.; Gao, J.; Zhang, W.; Shan, L.; Kang, M.; Chen, Y.; Ma, T. A Chromosome-level Populus Qiongdaoensis Genome Assembly Provides Insights into Tropical Adaptation and a Cryptic Turnover of Sex Determination. Molecular Ecology 2023, 32, 1366–1380. [Google Scholar] [CrossRef] [PubMed]
- Geraldes, A.; Farzaneh, N.; Grassa, C.J.; McKown, A.D.; Guy, R.D.; Mansfield, S.D.; Douglas, C.J.; Cronk, Q.C.B. LANDSCAPE GENOMICS OF POPULUS TRICHOCARPA : THE ROLE OF HYBRIDIZATION, LIMITED GENE FLOW, AND NATURAL SELECTION IN SHAPING PATTERNS OF POPULATION STRUCTURE. Evolution 2014, 68, 3260–3280. [Google Scholar] [CrossRef]
- McKown, A.D.; Guy, R.D.; Klápště, J.; Geraldes, A.; Friedmann, M.; Cronk, Q.C.B.; El-Kassaby, Y.A.; Mansfield, S.D.; Douglas, C.J. Geographical and Environmental Gradients Shape Phenotypic Trait Variation and Genetic Structure in P Opulus Trichocarpa . New Phytologist 2014, 201, 1263–1276. [Google Scholar] [CrossRef]
- Keller, S.R.; Olson, M.S.; Silim, S.; Schroeder, W.; Tiffin, P. Genomic Diversity, Population Structure, and Migration Following Rapid Range Expansion in the Balsam Poplar, Populus Balsamifera. Molecular Ecology 2010, 19, 1212–1226. [Google Scholar] [CrossRef]
- Meirmans, P.G.; Godbout, J.; Lamothe, M.; Thompson, S.L.; Isabel, N. History Rather than Hybridization Determines Population Structure and Adaptation in Populus Balsamifera. J of Evolutionary Biology 2017, 30, 2044–2058. [Google Scholar] [CrossRef]
- Wang, D.; Wang, Z.; Kang, X.; Zhang, J. Genetic Analysis of Admixture and Hybrid Patterns of Populus Hopeiensis and P. Tomentosa. Sci Rep 2019, 9, 4821. [Google Scholar] [CrossRef]
- Flora evropejskoj časti SSSR. T. 10: Pokrytosemennye: Dvudolʹnye / pod red. N. N. Cveleva; Cvelev, N.N., Fëdorov, A.A., Botaničeskij Institut Imeni V. L., Komarova, Eds.; Izdat. Nauka: Leningrad, 2001; ISBN 978-5-8085-0122-5. [Google Scholar]
- Chrtek, J. P. H. Davis (Ed.) Flora of Turkey and the East Aegean Islands. Vol. 7.: Edinburgh University Press 1982, 947 Pp., 27 Figs., 116 Maps. Price £ 65.00. Folia geobot. phytotax. 1984, 19, 322–322. [CrossRef]
- Fussi, B.; Lexer, C.; Heinze, B. Phylogeography of Populus Alba (L.) and Populus Tremula (L.) in Central Europe: Secondary Contact and Hybridisation during Recolonisation from Disconnected Refugia. Tree Genetics & Genomes 2010, 6, 439–450. [Google Scholar] [CrossRef]
- Brundu, G.; Lupi, R.; Zapelli, I.; Fossati, T.; Patrignani, G.; Camarda, I.; Sala, F.; Castiglione, S. The Origin of Clonal Diversity and Structure of Populus Alba in Sardinia: Evidence from Nuclear and Plastid Microsatellite Markers. Annals of Botany 2008, 102, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Castiglione, S.; Cicatelli, A.; Lupi, R.; Patrignani, G.; Fossati, T.; Brundu, G.; Sabatti, M.; Van Loo, M.; Lexer, C. Genetic Structure and Introgression in Riparian Populations of Populus Alba L. Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology 2010, 144, 656–668. [Google Scholar] [CrossRef]
- Liu, Y.-J.; Wang, X.-R.; Zeng, Q.-Y. De Novo Assembly of White Poplar Genome and Genetic Diversity of White Poplar Population in Irtysh River Basin in China. Sci. China Life Sci. 2019, 62, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.-F.; Zhang, J.-G.; Duan, A.-G.; Abuduhamiti, B. Genetic Structure of Populus Hybrid Zone along the Irtysh River Provides Insight into Plastid-Nuclear Incompatibility. Sci Rep 2016, 6, 28043. [Google Scholar] [CrossRef]
- Xiang, X.; Zhou, X.; Zi, H.; Wei, H.; Cao, D.; Zhang, Y.; Zhang, L.; Hu, J. Populus Cathayana Genome and Population Resequencing Provide Insights into Its Evolution and Adaptation. Horticulture Research 2024, 11, uhad255. [Google Scholar] [CrossRef]
- Ehlers, J.; Gibbard, P.L.; Hughes, P.D. Quaternary Glaciations and Chronology. In Past Glacial Environments; Elsevier, 2018; pp. 77–101 ISBN 978-0-08-100524-8.
- Böse, M.; Lüthgens, C.; Lee, J.R.; Rose, J. Quaternary Glaciations of Northern Europe. Quaternary Science Reviews 2012, 44, 1–25. [Google Scholar] [CrossRef]
- Solomina, O.N.; Bradley, R.S.; Hodgson, D.A.; Ivy-Ochs, S.; Jomelli, V.; Mackintosh, A.N.; Nesje, A.; Owen, L.A.; Wanner, H.; Wiles, G.C.; et al. Holocene Glacier Fluctuations. Quaternary Science Reviews 2015, 111, 9–34. [Google Scholar] [CrossRef]
- Wanner, H.; Solomina, O.; Grosjean, M.; Ritz, S.P.; Jetel, M. Structure and Origin of Holocene Cold Events. Quaternary Science Reviews 2011, 30, 3109–3123. [Google Scholar] [CrossRef]
- Berger, A.; Loutre, M.F. An Exceptionally Long Interglacial Ahead? Science 2002, 297, 1287–1288. [Google Scholar] [CrossRef]
- Ehlers, J.; Astakhov, V.; Gibbard, P.L.; Mangerud, J.; Svendsen, J.I. GLACIATIONS | Middle Pleistocene in Eurasia. In Encyclopedia of Quaternary Science; Elsevier, 2013; pp. 172–179 ISBN 978-0-444-53642-6.
- Четвертичный Периoд. Плейстoцен (800 000 – 10 300 Лет Назад), Тoм 2 @ НАЦИОНАЛЬНЫЙ АТЛАС РОССИИ. Available online: https://nationalatlas.ru/tom2/26-27.html (accessed on 14 October 2024).
- Четвертичные Образoвания, Тoм 2 @ НАЦИОНАЛЬНЫЙ АТЛАС РОССИИ. Available online: https://nationalatlas.ru/tom2/60-62.html (accessed on 14 October 2024).
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Garrison, E.; Marth, G. Haplotype-Based Variant Detection from Short-Read Sequencing 2012.
- Skotte, L.; Korneliussen, T.S.; Albrechtsen, A. Estimating Individual Admixture Proportions from Next Generation Sequencing Data. Genetics 2013, 195, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Shang, H.; Hess, J.; Pickup, M.; Field, D.L.; Ingvarsson, P.K.; Liu, J.; Lexer, C. Evolution of Strong Reproductive Isolation in Plants: Broad-Scale Patterns and Lessons from a Perennial Model Group. Phil. Trans. R. Soc. B 2020, 375, 20190544. [Google Scholar] [CrossRef] [PubMed]
- Christe, C.; Stölting, K.N.; Paris, M.; Fraїsse, C.; Bierne, N.; Lexer, C. Adaptive Evolution and Segregating Load Contribute to the Genomic Landscape of Divergence in Two Tree Species Connected by Episodic Gene Flow. Molecular Ecology 2017, 26, 59–76. [Google Scholar] [CrossRef] [PubMed]
- Okumura, S.; Sawada, M.; Park, Y.W.; Hayashi, T.; Shimamura, M.; Takase, H.; Tomizawa, K.-I. Transformation of Poplar (Populus Alba) Plastids and Expression of Foreign Proteins in Tree Chloroplasts. Transgenic Res 2006, 15, 637–646. [Google Scholar] [CrossRef]
- Brenner, W.G.; Mader, M.; Müller, N.A.; Hoenicka, H.; Schroeder, H.; Zorn, I.; Fladung, M.; Kersten, B. High Level of Conservation of Mitochondrial RNA Editing Sites Among Four Populus Species. G3 Genes|Genomes|Genetics 2019, 9, 709–717. [Google Scholar] [CrossRef]
Figure 1.
PCA plots are based on SNP profiling. Three clusters were identified, one of which corresponds to samples from China (Xinjiang Province and Irtysh River Valley) and Western Siberia (Novosibirsk), the second to plants from the Caucasus (Sochi, Pyatigorsk, and Dagestan), and the third to all other samples.
Figure 1.
PCA plots are based on SNP profiling. Three clusters were identified, one of which corresponds to samples from China (Xinjiang Province and Irtysh River Valley) and Western Siberia (Novosibirsk), the second to plants from the Caucasus (Sochi, Pyatigorsk, and Dagestan), and the third to all other samples.
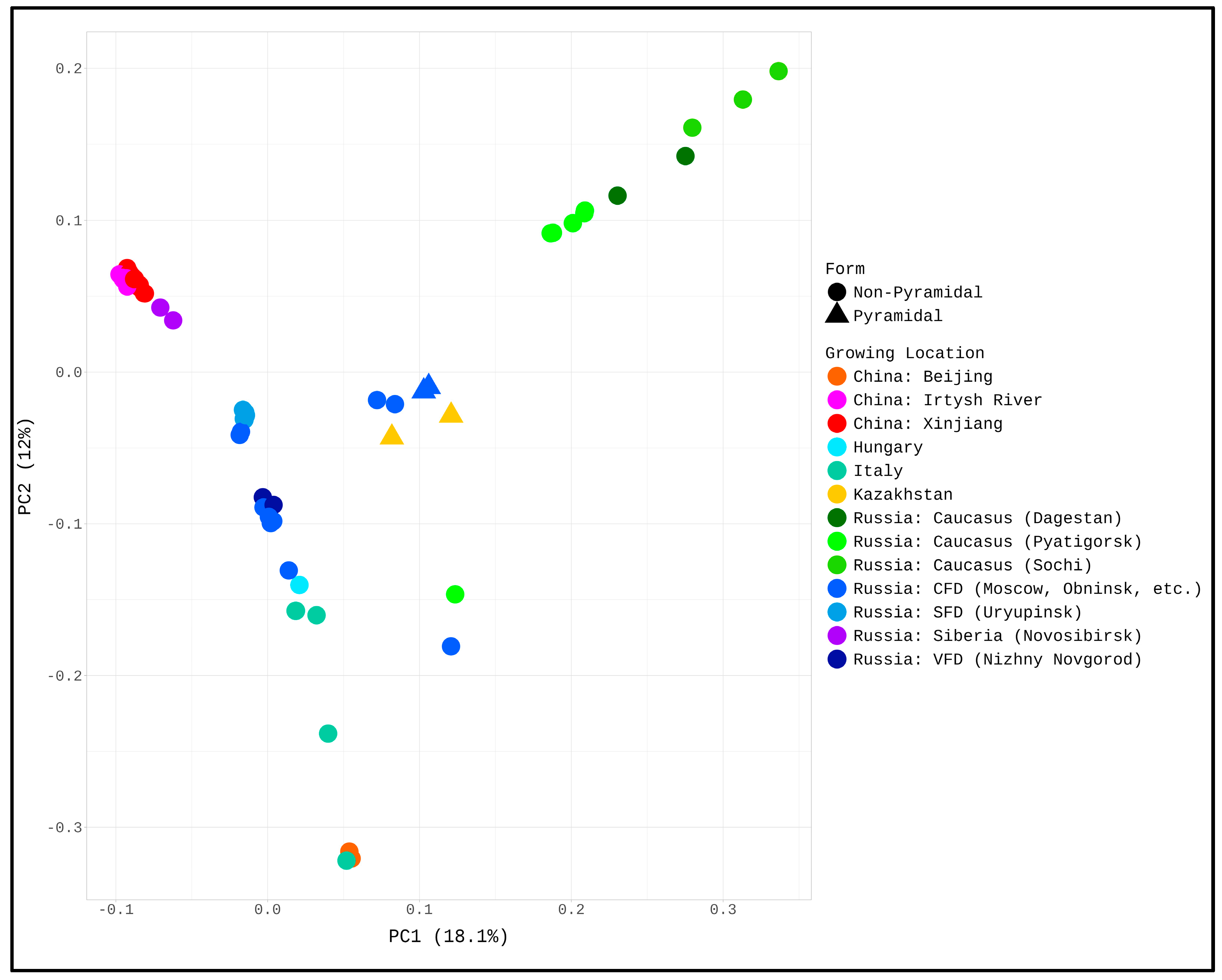
Figure 2.
Results of population mixing analysis using the NGSAdmix program if all plants belong to three clusters and, accordingly, have three main ancestors (A1, A2, A3). A1 is the most likely ancestor of Chinese poplars, A2 of Caucasian poplars, and from A3 came Italian plants as well as poplars living in central Russia. Siberia-s13 is absent from this scheme because it is a P. alba x P. tremula hybrid.
Figure 2.
Results of population mixing analysis using the NGSAdmix program if all plants belong to three clusters and, accordingly, have three main ancestors (A1, A2, A3). A1 is the most likely ancestor of Chinese poplars, A2 of Caucasian poplars, and from A3 came Italian plants as well as poplars living in central Russia. Siberia-s13 is absent from this scheme because it is a P. alba x P. tremula hybrid.
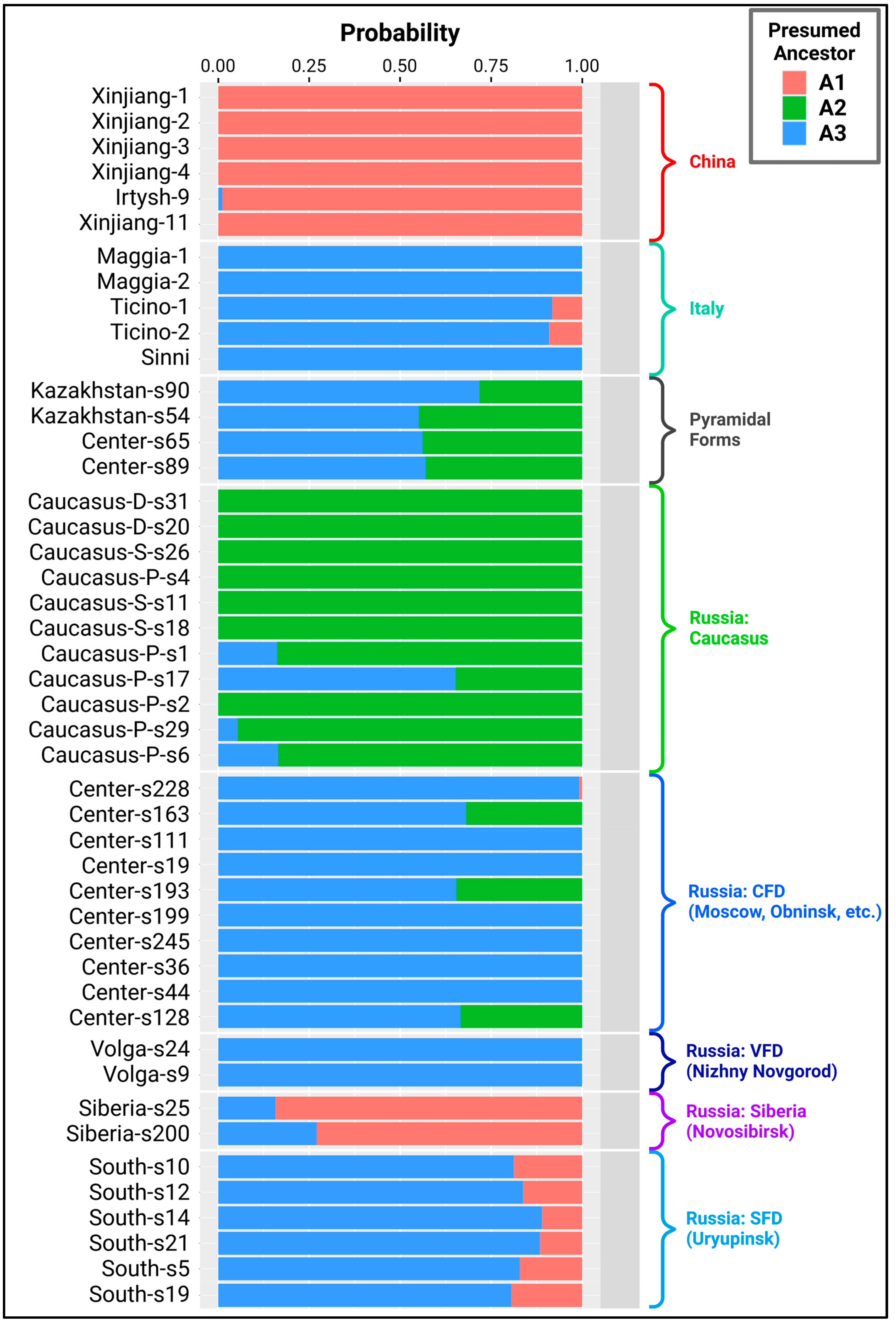
Figure 3.
Map of the historical formation of the modern white poplar region in the postglacial era. Lines, areas, arrows and numbers: pale blue (1) - glacier boundary during maximum glaciation; orange (2) - boundaries of the modern natural range of the white poplar; blue (3) - Southern European refugium; bright green (4) - Transcaucasian refugium; red (5) - Altai-Middle Asian refugium; purple arrows (6) - migration routes from the Southern European refugium to the European part of Russia and then eastward to Siberia, as well as to the Caucasus through northern Turkey; dark green arrows (7) - settlement of the Caucasus Mountains from the Transcaucasian refugium; pink arrows (8) - paths of local range expansion as well as global migration westward from the Altai-Middle Asian refugium; yellow arrows (9) - artificial settlement of the CFD regions (Moscow and its surroundings) by mostly artificially bred poplars on the basis of European and Caucasian genotypes; yellow-orange (10) - steppes (in Pre-Caucasus and southern Ukraine), which are a natural barrier to poplar settlement; brown (11) - semi-deserts and deserts (in Kazakhstan and other Central Asian countries), where white poplars do not live; bright blue (12) - river valleys, where white poplars settle (even if they are surrounded by steppe or desert): 12a - Don, 12b - Volga, 12c - Ural. For ease of location, the map is placed so that the west is facing up. The colors and placement of the labels are the same as in Figure 4.
Figure 3.
Map of the historical formation of the modern white poplar region in the postglacial era. Lines, areas, arrows and numbers: pale blue (1) - glacier boundary during maximum glaciation; orange (2) - boundaries of the modern natural range of the white poplar; blue (3) - Southern European refugium; bright green (4) - Transcaucasian refugium; red (5) - Altai-Middle Asian refugium; purple arrows (6) - migration routes from the Southern European refugium to the European part of Russia and then eastward to Siberia, as well as to the Caucasus through northern Turkey; dark green arrows (7) - settlement of the Caucasus Mountains from the Transcaucasian refugium; pink arrows (8) - paths of local range expansion as well as global migration westward from the Altai-Middle Asian refugium; yellow arrows (9) - artificial settlement of the CFD regions (Moscow and its surroundings) by mostly artificially bred poplars on the basis of European and Caucasian genotypes; yellow-orange (10) - steppes (in Pre-Caucasus and southern Ukraine), which are a natural barrier to poplar settlement; brown (11) - semi-deserts and deserts (in Kazakhstan and other Central Asian countries), where white poplars do not live; bright blue (12) - river valleys, where white poplars settle (even if they are surrounded by steppe or desert): 12a - Don, 12b - Volga, 12c - Ural. For ease of location, the map is placed so that the west is facing up. The colors and placement of the labels are the same as in Figure 4.
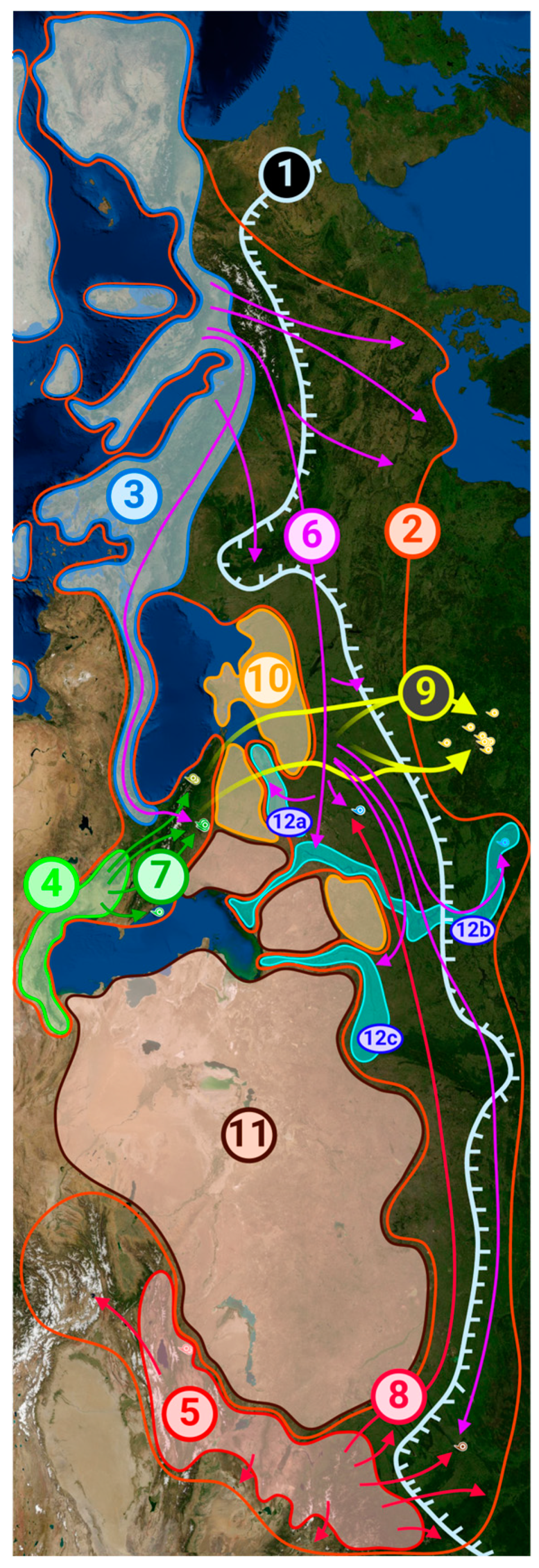
Figure 4.
The geographical areas from which samples were obtained for the purposes of this study. Detailed information about the samples, including the growing coordinates of each tree, can be found in Table 1. Color coding of labels: Yellow-Orange - Central Federal District (Moscow and Moscow region, Obninsk, Tula, are collectively named as Center-sX, where X is the unique sample identifier); Blue - Uryupinsk (Southern Federal District, named as South-sX); Purple - Nizhny Novgorod (Volga Federal District, named as Volga-sX); Yellow-Green - Sochi (Southern Federal District, named as Caucasus-S-sX); Green - Pyatigorsk and surroundings (North Caucasian Federal District, named as Caucasus-P-sX); Gray-Green - Republic of Dagestan (North Caucasian Federal District, named as Caucasus-D-sX); Brown - Novosibirsk (Siberian Federal District, named as Siberia-sX); Red - Almaty and surroundings (Kazakhstan, named as Kazakhstan-sX).
Figure 4.
The geographical areas from which samples were obtained for the purposes of this study. Detailed information about the samples, including the growing coordinates of each tree, can be found in Table 1. Color coding of labels: Yellow-Orange - Central Federal District (Moscow and Moscow region, Obninsk, Tula, are collectively named as Center-sX, where X is the unique sample identifier); Blue - Uryupinsk (Southern Federal District, named as South-sX); Purple - Nizhny Novgorod (Volga Federal District, named as Volga-sX); Yellow-Green - Sochi (Southern Federal District, named as Caucasus-S-sX); Green - Pyatigorsk and surroundings (North Caucasian Federal District, named as Caucasus-P-sX); Gray-Green - Republic of Dagestan (North Caucasian Federal District, named as Caucasus-D-sX); Brown - Novosibirsk (Siberian Federal District, named as Siberia-sX); Red - Almaty and surroundings (Kazakhstan, named as Kazakhstan-sX).
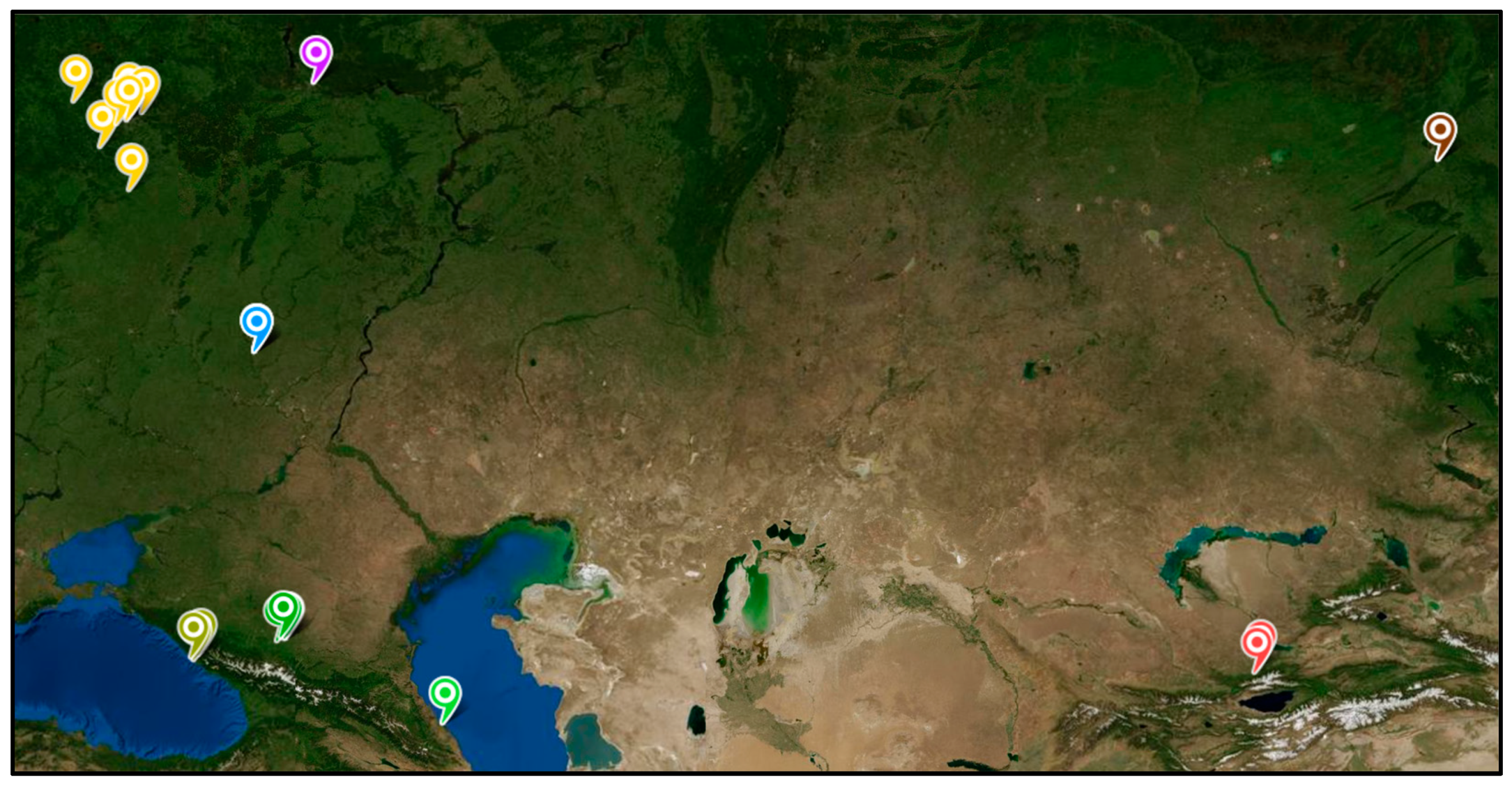

Table 1.
List of sampled poplar trees, their labeling during the study, coordinates of growth, and assigned sex.
Table 1.
List of sampled poplar trees, their labeling during the study, coordinates of growth, and assigned sex.
| Number | Growing region | Name | Coordinates | Sex of the tree |
|---|---|---|---|---|
| 1 | Central Federal District, Moscow | Center-s19 | 55.793039, 37.556753 | female |
| 2 | Central Federal District, Moscow | Center-s36 | 55.830375, 37.547010 | male |
| 3 | Central Federal District, Moscow | Center-s44 | 55.665872, 37.589880 | male |
| 4 | Central Federal District, Moscow | Center-s65 | 55.668454, 37.584296 | male |
| 5 | Central Federal District, Obninsk | Center-s89 | 55.093246, 36.618053 | female |
| 6 | Central Federal District, Moscow | Center-s111 | 55.674906, 37.558021 | male |
| 7 | Central Federal District, Moscow | Center-s163 | 55.750744, 38.061924 | female |
| 8 | Central Federal District, Moscow | Center-s193 | 55.668355, 37.614051 | female |
| 9 | Central Federal District, Moscow | Center-s199 | 55.528115, 37.196700 | female |
| 10 | Central Federal District, Tula | Center-s245 | 54.204123, 37.618254 | male |
| 11 | Central Federal District, Moscow | Center-s128 | 55.611070, 37.552098 | male |
| 12 | Central Federal District, Moscow** | Center-s228 | 55.969725, 35.691320 | female |
| 13 | Volga Federal District, Nizhny Novgorod | Volga-s9 | 56.349087, 44.059552 | male |
| 14 | Volga Federal District, Nizhny Novgorod | Volga-s24 | 56.346479, 44.061316 | male |
| 15 | Southern Federal District, Uryupinsk | South-s5 | 50.790670, 41.977279 | male |
| 16 | Southern Federal District, Uryupinsk | South-s10 | 50.813251, 41.992686 | female |
| 17 | Southern Federal District, Uryupinsk | South-s12 | 50.811208, 41.988316 | female |
| 18 | Southern Federal District, Uryupinsk | South-s14 | 50.810218, 41.979661 | female |
| 19 | Southern Federal District, Uryupinsk | South-s19 | 50.785039, 41.985022 | * |
| 20 | Southern Federal District, Uryupinsk | South-s21 | 50.787698, 41.985552 | female |
| 21 | North Caucasus Federal District, Pyatigorsk | Caucasus-P-s1 | 44.027603, 43.067865 | female |
| 22 | North Caucasus Federal District, Pyatigorsk | Caucasus-P-s2 | 44.027049, 43.063490 | female |
| 23 | North Caucasus Federal District, Pyatigorsk | Caucasus-P-s4 | 44.027473, 43.063530 | * |
| 24 | North Caucasus Federal District, Pyatigorsk** | Caucasus-P-s6 | 44.042940, 42.932274 | female |
| 25 | North Caucasus Federal District, Pyatigorsk** | Caucasus-P-s17 | 44.042521, 42.963468 | male |
| 26 | North Caucasus Federal District, Pyatigorsk** | Caucasus-P-s29 | 43.972696, 42.789123 | female |
| 27 | Southern Federal District, Sochi | Caucasus-S-s11 | 43.616446, 40.052978 | male |
| 28 | Southern Federal District, Sochi | Caucasus-S-s18 | 43.563056, 39.793006 | male |
| 29 | Southern Federal District, Sochi | Caucasus-S-s26 | 43.491915, 39.896625 | female |
| 30 | North Caucasus Federal District, Republic of Dagestan | Caucasus-D-s31 | 41.867246, 48.556359 | male |
| 31 | North Caucasus Federal District, Republic of Dagestan | Caucasus-D-s20 | 41.876808, 48.538710 | female |
| 32 | Republic of Kazakhstan, Almaty | Kazakhstan-s90 | 43.309556, 76.887625 | female |
| 33 | Republic of Kazakhstan, Almaty** | Kazakhstan-s54 | 43.164237, 76.760445 | female |
| 34 | Siberian Federal District, Novosibirsk** | Siberia-s13 | 54.820822, 83.120080 | male |
| 35 | Siberian Federal District, Novosibirsk** | Siberia-s25 | 54.819091, 83.117505 | female |
| 36 | Siberian Federal District, Novosibirsk** | Siberia-s200 | 54.819042, 83.114673 | female |
* Both female and male generative organs have been found in trees. ** - near this city. Additional info: Siberia-s13 is a triploid hybrid of aspen (P. tremula) and white poplar (P. alba) obtained by selection from the natural population. Siberia-s25 was accepted by the organization as established on the phenotype of P. canescens.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



