
Submitted:
06 November 2024
Posted:
06 November 2024
You are already at the latest version
Abstract
In angiosperms, paradormancy, a phenomenon that the growth of axillary buds be arrested by the biochemical signals from other organs or tissues, has been extensively investigated in deciduous perennial woody species. However, little attention has been paid to paradormancy set and release in perennial evergreen trees. Camellia sinensis cv Huangdan develops axillary floral buds and axillary shoot buds within same leaf node simultaneously, with growing floral buds repressing shoot buds leading to their paradormancy. In this study, we manually removed the axillary floral buds, then harvested the axillary shoot buds after 1- and 3- days of the floral bud removal, untargeted metabolomics were conducted to identify metabolome remodeling patterns during paradormancy-to-growth transition. Our data suggest that the restricted supply of reducing hexose and ammonium to the axillary shoot buds could be the major cause leading to their paradormancy. The axillary shoot buds were released from paradormancy one day after the axillary floral bud removal, this transition was accompanied with mobilization and reconfiguration for multiple primary and secondary metabolites. These metabolome reprogramming could help establish the reactive oxygen species gradients along leaf bud meristem, thus facilitate paradormancy set or release. This study provides novel insights about paradormancy set and release in a perennial evergreen tree species.
Keywords:
axillary floral bud
; axillary shoot bud
; Camellia sinensis
; metabolomics
; paradormancy
1. Introduction
In angiosperms, the axillary buds are formed from the axillary meristems initiated at the base of emerging leaves, and contain preformed elements of adult shoots including shoot meristems and leaf primordia [1]. Under certain environmental conditions, the axillary buds can enter into dormancy rather than growing out immediately to form a branch. Bud dormancy is a state that meristem-containing plant structures temporarily suspend growth when environmental or endogenous conditions are unfavorable for sustained growth. Thus, dormancy has adaptive advantage for reproductive success and survival. Multiple environmental and endogenous factors could induce bud dormancy set and release, including nutrient or water availability, temperature, light quality, day-length, sink and source activities, hormone levels and epigenetic modification of growth related genes [2,3,4,5]. Based on the types of stimulus that promote growth arrest, Lang et al. (1987) classified bud dormancy into paradormancy, endodormancy, and ecodormancy [6]. Paradormancy is a temporary bud growth suspension caused by biochemical signals from other organs or tissues.
The metabolite reconfiguration of sugars, sugar alcohols, amino acids, organic acids, and flavonoids have been demonstrated participating endodormancy release [7,8,9,10,11,12,13]. Paradormancy release of axillary buds in hybrid aspen is induced by decapitation, gibberellin and plasmodesmata communication play important roles during paradormancy-to-growth transition [14]. Similarly, early defoliation promotes paradormancy release of pear floral buds, and auxin efflux is involved in this process [15]. Redox metabolism has a large impact on endodormancy release [16,17,18,19,20,21,22]. ROS exhibit different gradient distributions across meristem zones and regulate meristem niche, cell cycle, cytokinesis and cell proliferation. Superoxide dismutase (SOD), peroxidase, peroxidases (PRXs) and NADPH oxidase are responsible for ROS gradient formation in meristem [23,24]. By now, it remains unclear whether non-enzymatic scavenger systems are involved in dormancy set and release.
In woody plants, the axillary buds undergo transitions between different dormant states throughout the year. So far, deciduous perennial woody or herbaceous species, including peach, Japanese apricot, grape, kiwifruit, poplar, raspberry or leaf spurge, are mostly used for dormancy studies [16,18,20,21,22,25,26,27], floral buds or shoot buds are applied as study materials. In contrast, less studies have been conducted in perennial evergreen woody species. As a result, it remains unclear whether evergreen trees and deciduous ones share similar mechanisms for dormancy regulation. Tea plant (Camellia sinensis) is a thermophilic perennial evergreen woody tree species, which are cultivated between latitudes 45° N and 35° S [2]. Tea plant shows two types of dormancy: ‘Banjhi dormancy’ and winter dormancy [2,5,28]. The former is a phenomenon of bud growth suspension under favorable conditions, usually take several times during a growing season, and be regarded as a form of endogenous rhythmic growth [28]; the latter occurs in all tea plants growing beyond 16° N or S latitudes when day length is below 11 h during winter [2,5]. Transcriptomic and metabolic studies have demonstrated that multiple factors are involved in winter dormancy, including carbohydrates, auxin, gibberellin, cytokines, abscisic acid (ABA), polyamines, callose-related cellular communication and epigenetic regulation [29,30,31,32,33,34,35,36].
Transcriptional analysis in woody and herbaceous species suggest that bud dormancy could be caused by carbon starvation [37,38,39]. Sugars are involved in cold acclimation and winter dormancy in tea plants [40,41,42]. Trehalose-6-phosphate (Tre6P) acts as a proxy for carbon status participating in sugar sensing [43,44]. SnRK1 is a master transcriptional regulator in energy signaling, it promotes catabolism and represses anabolism, cell division and growth [45]. Tea tree is a short-day plant, with the initiation of day length shorting at late June, tea floral buds start to emerge at leaf base. At late August, the axillary floral buds and axillary shoot buds in Camellia sinensis cv Huangdan are both emerged from the base of lateral leaves, with axillary shoot bud siting in the center and contained by two axillary floral buds (Figure 1). The axillary floral buds grow normally, while the axillary shoot buds are arrested. This developmental behavior provides a system model to tackle the relations between resource allocation and paradormancy in a perennial evergreen tree species. In this study, the axillary floral buds were removed first, then the axillary shoot buds were harvested at 1- and 3- days after floral bud removal (DAF), untargeted metabolomics analysis was followed. This study offers novel insights about metabolomics reconfigurations and regulations during paradormancy-to-growth transitions in a perennial evergreen woody tree species.
2. Results
2.1. The Axillary Shoot Bud Metabolome Was Altered One Day After Local Floral Bud Removal
Camellia sinensis cv Huangdan develops the axillary floral buds and the axillary shoot bud from the base of same leaf node simultaneously, the axillary floral buds grow normally while the growth of the axillary shoot bud was suspended (Figure 1). We reasoned that the growing floral buds could act as strong sink and deprive most of the imported nutrient, this could result in carbon starvation and paradormancy to the axillary shoot bud. To test this hypothesis, we manually removed the axillary floral buds and kept the local axillary shoot buds intact, at 1- and 3- DAF there was no obvious growth for the axillary shoot buds (Figure 2). The samples of the harvested axillary shoot buds (HD_0, HD_1, and HD_3) were extensively extracted, metabolites were analyzed by UPLC-QTOF MS. In total, 1882 and 2779 metabolic signatures were detected from negative ion mode and positive ion mode, 467 and 696 of them showed differential abundance (P<0.05), respectively (Supplementary data 1; Supplementary data 2). PLS-DA analysis showed that the X-component explained 20.9-24.7% of the variance and clearly distinguished HD_0 from HD_1 and HD_3, the Y-component explained 15.2-16% of the variance (Figure 3A-3B), indicating that the metabolic difference between HD_1 and HD_3 samples were smaller compared to HD_0 samples. Apparently, these three groups of the axillary shoot buds were in divergent physiological states.
Statistical analyses were performed to compare metabolite changes in the axillary shoot buds before and after the axillary floral bud removal. Under negative ion mode, 300, 93 and 188 metabolite signatures showed differential abundance for the HD_1 vs. HD_0, HD_3 vs. HD_1 and HD_3 vs. HD_0 comparison; under positive ion mode, 431, 152 and 352 metabolite signatures showed differential abundance for the same set comparisons. The distribution of metabolite signatures that are unique and common among these three comparisons is shown in the Venn diagram (Figure 3C-3D). Under negative ion mode, 207 out of 300 metabolites with differential abundance were unique for HD_1 vs. HD_0, 42 out of 93 were unique for HD_3 vs HD_1, and 110 out of 188 were unique for HD_3 vs. HD_0 (Figure 3C). Under positive ion mode, 231 out of 431 metabolites with differential abundance were unique for HD_1 vs. HD_0, 72 out of 152 were unique for HD_3 vs. HD_1 , and 174 out of 352 were unique for HD_3 vs. HD_0 (Figure 3D). These data suggest that the physiological states of the axillary shoot buds experienced most dramatic metabolic remodeling at 1 DAF, an indication of the paradormancy release for the axillary shoot buds.
By query with an in-house authentic standard library and other online sources, 86 and 63 metabolites were chemically identified from the negative ion mode and the positive ion mode, respectively (Supplementary data 3; Supplementary data 4). Secondary metabolites were over-represented, including phenolic acids and their derivatives, flavanol, flavone and flavonol glycosides, and proanthocyanidin. In addition, several alkaloids were identified only from the positive ion mode. For those metabolites that were identified from both ion modes, they showed a general similar changing trends.
2.2. Flavonoids and Alkaloids
Gallic acid (GA) can be methylated into methyl gallate [46] for storage or converted into galloylglucose serving as substrate for ester catechin synthesis [47,48]. Here, we found that the paradormant axillary shoot buds contained lower levels of GA and its derivatives (monogalloylglucose isomer 1, methylgallate, methyl galloylglucose, trigalloylglucose isomer 2), their contents increased steadily after paradormancy release. In contrast, theogallin, a gallate ester with quinic acid, kept fairly consistent (Figure 4A). Interestingly, all catechins and flavonol glycoside contents showed similar changing trends as GA (Figure 4B). This is consistent with previous report that GA availability could be the major bottleneck for EGCG synthesis [49]. One exception is epigallocatechin digallate which showed higher levels in the paradormant shoot buds followed by steady decrease after paradormancy release (Figure 4B), suggesting its role for paradormancy maintenance. Chicoric acid, a tartaric acid ester of two caffeic acid, increased after the axillary floral bud removal (Figure 4A), this changing trend is consistent with its proposed role in wound healing [50]. Chicoric acid is also an effective antioxidant [51], and regulate glucose and lipid homeostasis [52,53].
Flavone glycosides (vitexin 2”-O-rhamnoside, apigenin 6-C-glucoside 8-C-arabinoside), dihydrocaffeic acid 3-O-glucuronide and majority of the proanthocyanidins showed opposite changing trends as GA and catechins, with higher contents present at the paradormant shoot buds, then decreased after paradormancy release (Figure 5A-5B). Proanthocyanidins (PAs) are synthesized from flavanol oligomerization, it’s not surprising that a higher proanthocyanidin contents were accompanied by a concurrent lower flavanol contents (Figure 4B; Figure 5B). Our data suggest that flavanol oligomerization was enhanced in the paradormant axillary shoot buds. PAs polymerization is a non-enzymatic process in planta, requiring sequestration of reactive/toxic pathway intermediates from cellular components. Once formed, PAs possess protective functions against oxidative stress, herbivory, and pathogen attack [54] . Thus, a higher level of PAs may help quench ROS and confer protection against oxidative stresses in the paradormant shoot buds.
Two alkaloids (theobromine and caffeine) were identified only from the positive ion mode (Supplementary data 4). Theobromine can be derived from caffeine degradation or serve as precursor for caffeine synthesis [55,56]. Since theobromine showed steady increase after paradormancy release, whereas caffeine kept constant (Supplementary data 4), our data suggest that caffeine synthase, which convert theobromine into caffeine [56], was inactive during paradormancy-to-growth transition.
As we described above, the metabolites identified from UPLC-MS platform are over-represented by secondary metabolites (Supplementary data 3; Supplementary data 4). Since primary metabolites are prone to be detected and identified by GC-MS platform [57], these samples were also derivatized and analyzed by GC-TOF-MS platform. In total, 394 metabolites were detected (Supplementary data 5), 146 metabolites were chemically identified (Supplementary data 6). Based on chemical structure, the identified metabolites were clustered into four groups, they are: amino acids, organic acids, sugars and sugar alcohols, free fatty acids and sterols. At below, we describe them individually.
2.3. Amino Acids
Twenty-one amino acids were identified, six of them showed higher contents in the paradormant axillary shoot buds, then decreased following paradormancy release, they are: L-glutamic acid (Glu), L-aspartic acid (Asp), asparagine 1 (Asn), asparagine 4, L-leucine (Leu), L-valine (Val) and L-threonine (Thr) (Figure 6A); meanwhile, nine amino acids, including theanine 1, theanine 2, γ-aminobutyric acid (GABA), glutamine (Gln), ornithine, citrulline, glycine (Gly), serine, phenylalanine (Phe) and L-tyrosine (Tyr), showed lower contents in the paradormant axillary shoot buds, increased to higher levels at 1 DAF followed by decrease at 3 DAF (Figure 6A). In tea tree, Glu, Asp and Asn are abundant proteinaceous amino acids, while theanine is the most abundant non-proteinaceous amino acid [58]. Apparently, theanine synthesis was enhanced following paradormancy release, which use Glu as substrate [59]. Glu also is the precursor for the synthesis of Gln, GABA, ornithine, citrulline and proline [60,61,62,63]. These substrate-products relations could explain why a reduction in Glu was accompanied with concurrent increase of theanine, GABA, Gln, ornithine, citrulline and proline at 1 DAF (Figure 6A). The decrease of Asp and Glu was accompanied with concurrent increase of Gln after paradormancy release at 1 DAF (Figure 6A), this raise the possibility that malate-aspartate shuttle is operating, which lead to the net mitochondrial export of α-ketoglutarate at the expense of Asp, then the exported α-ketoglutarate be converted to glutamate for further Gln synthesis. Asp also serves as the precursor for the synthesis of branched chain amino acids (BCAAs) including Leu, Val, Thr and Lys (Figure S1) [64]. At 1 DAF, Leu, Val and Thr showed similar changing trends as Asp (Figure 6A), suggesting that BCAAs synthesis is closely coordianted with Asp supply. The BCAAs contents decreased at 1 DAF (Figure 6A), suggesting that they could be degraded during paradormancy-to-growth transition. L-threonine aldolase degrade Thr into glycine [65], glycine can be further converted into serine, hydroxypyruvic acid and glyceric acid through partial C2 photorespiratory carbon oxidation cycle, glyceric acid can be fueled into Calvin cycle to generate 1,5-ribulose diphosphate (RuBP) , a key intermediate to boost photo carbon assimilation. Consistent with these potenital interconversions, Thr reduction was accompanied by a concurrent increase of glycine, serine, hydroxypyruvic acid and glyceric acid (Figure 6A).
Chalcone is an upstream intermediate for flavonoid synthesis, which require 3 acetyl-CoAs for 1 chalcone synthesis (Figure S2). In tea leaves, the flavonoid pathway is a high flux rout, it’s anticipated that large supply of cytosolic acetyl-CoAs is required. Currently, it remains elusive in tea tree which pathway(s) are responsible to generate cytosolic acetyl-CoA to fuel flavonoid synthesis. Considering that BCAAs degradation produce acetyl-CoA, a reduction of BCAAs at 1 DAF was accompanied with concurrent increase of phenolic acids, flavanols and flavonols (Figure 4; Figure 5A), we speculate that BCAAs degradation could provide acetyl-CoA to fuel flavonoid synthesis during paradormancy-to-growth transition. Aromatic amino acids (Phe, Tyr, Trp) are derived from shikimic acid, their contents concurrently increased at 1 DAF (Figure 6A). However, this correlation no longer existed at 3 DAF except tyrosine, in which shikimic acid reached peak level whereas Phe and Trp were on their nadir (Figure 6A-6B). Phe is the precursor for flavonoids synthesis, its reduction from 1 DAF to 3 DAF was accompanied with concurrent increase of GCG and EC (Figure 4B; Figure 6B). Trp is the precursor for auxin synthesis, its decline at 3 DAF raises the possibility that auxin synthesis could be elevated at this time point.
2.4. Organic Acids
Citric acid, succinic acid, malic acid and niacin are intermediates of TCA and glyoxalate cycle (Figure S3), they exhibited two different changing trends: citric acid showed higher contents in the paradormant shoot buds followed by steady decrease through 1- and 3- DAF; in contrast, the contents of succinic, malic acid and niacin increased at 1 DAF, then slightly decreased at 3 DAF (Figure 6B). These data suggest that both TCA cycle and glyoxalate cycle are operating at 1 DAF simultaneously, which lead to the net export of α-ketoglutarate from mitochondrial to sustain malate-aspartate shuttle for the synthesis of glutamate and reducing power NADH at the expense of acetyl-CoA (Figure S3). Acetyl-CoA may originate from BCAAs degradation, β-oxidation of free fatty acids or glycolysis. Considering that citric acid inhibits glycolysis through its negative effects on the activities of phosphofructokinase and pyruvate kinase [67], the higher levels of citric acid at paradormant shoot buds are expected to suppress glycolysis leading to lower cell energy charge. After floral bud removal, the decline of citric acid would relieve its inhibition on glycolysis, more trios phosphate can be produced to fuel into TCA. At 3 DAF, the reduction of these TCA intermediates could be related to the growth initiation of the axillary shoot buds, since there is a general negative correlation between the intermediates of central metabolic pathways and plant growth rate [68].
Five organic acids, including quinic acid (QA), salicylic acid (SA), gallic acid (GA), shikimic acid (SHA), and benzoic acid (BA), are the intermediates of shikimate pathway and phenylpropanoid pathway (Figure S2). They enxibited two different changing trends: QA and SA showed higher content in the paradormant shoot buds, then decreased at 1 DAF; in contrast, GA, SHA and BA are steadily increased through 1- to 3- DAF (Figure 6B). Considering that the synthesis pathways of QA, GA and SHA share 3-dehydroquinate as common precursor (Figure S2) [69], these opposite changing patterns suggest that QA may play roles in paradormant release. Quinate and p-coumaroyl-CoA can be convert into p-coumaroyl quinate through quinate shunt which is an intermediate for chlorogenic acid synthesis [70,71,72]. Accordingly, quinate and p-coumaroyl quinate showed opposite changing trends during paradormancy-to-growth transition (Figure 4A; Figure 6B). SA is synthesized from chorismic acid through two routs (Figure S2) [73]: in the phenylalanine-benzoic acid rout, chorismic acid is sequentially converted into phenylalanine, trans-cinnamic acid and BA, then BA serves as direct substrate for SA synthesis; in the isochorismic acid-IC-9-Glu rout, chorismic acid is sequentially converted into isochorismic acid and IC-9-Glu, then IC-9-Glu serves as the direct substrate for SA synthesis (Figure S2). SHA and BA showed similar changing trends with lowest levels in the paradormant shoot buds (Figure 6B), while SA present at higher levels in the paradormant shoot buds, then decreased at 1 DAF (Figure 6B). These data suggest that phenylalanine-benzoic acid rout contribute to SA synthesis during paradormancy-to-growth transition, while the contributions of isochorismic acid-IC-9-Glu rout to SA synthesis can’t be clarified from our current dataset.
Four organic acids, including methyl phosphate, propanoic acid, malonic acid and ethylmalonic acid, increased during paradormancy-to-growth transition (Figure 6B). Methyl phosphate is Met degradation product, while propanoic acid and malonic acid are degradation intermediates of Val and Ile [74]. Consistent with this substrate-product relation, Val content showed opposite change patterns with propanoic acid and malonic acid during paradormancy-to-growth transition (Figure 6A-6B).
Ascorbic acid is one of the major components of cell’s non-enzymatic antioxidants. It uses L-galactono-1,4-lactone as precursor which is generated through an alternative galactonic acid pathway [75], L-galactonic acid is derived from D-galacturonic acid which is a main constituent of pectin. D-galacturonic acid can also be oxidized into mucic acid [76]. During paradormancy-to-growth transition, D-galacturonic acid and galactonic acid contents kept unchanged, while mucic acid content increased (Figure 6B), suggesting that ascorbic acid synthesis may not be affected. 2-keto-L-gulonic acid is an intermediate of ascorbic acid degradation [77], its contents decreased at 1 DAF (Figure 6B), suggesting that ascorbic acid degradation was reduced during paradormancy-to-growth transition .
3 -hydroxybutyric acid (3-HB) synthesis in plants have two possible routs : 1) it is derived from the synthesis of plant endophyte; 2) it is synthesized in planta from acetyl-CoA through thiolase and acetoacetyl-CoA reductase [78,79]. Considering that Lys and Thr degradation produce 3-hydroxybutyryl-CoA and 2-amino-3-oxobutyrate, respectively [74], that raises an open question whether Lys and Thr degradation could also contribute to 3-HB synthesis. 3-HB shows multiple functions in plants: 1) 3-HB can be catabolized to acetyl-CoA which raise intracellular acetyl-CoA levels; 2) 3-HB acts as a direct antioxidant for hydroxyl radical, and inhibit mitochondrial ROS production [80]; 3) 3-HB can modify the lysine residues on histone and non-histone proteins to regulate gene expression epigenetically [81]; 4) 3-HB inhibits histone deacetylasesand activates gene expression. Genes involved in the phenylpropanoid pathway was identified to be one of the recipients of epigenetic changes induced by 3-HB [79]. We found that 3-HB showed a transit increase during paradormancy-to-growth transition followed by a decline at 3 DAF (Figure 6B), this coincide with dramatic metabolic remodeling at 1 DAF (Figure 3). Thus, our data provide independent evidence that 3-HB could regulate gene expression through epigenetic reprogramming during paradormancy-to-growth transition of tea buds.
2.5. Sugars and Sugar Alcohols
Forty-two sugars and sugar alcohols were identified, 8 of them showed the highest contents in the paradormant axillary shoot buds, then decreased at 1 DAF, they are: trehalose, raffinose, galactinol, fructose, methyl galactoside, α-D-xylopyranose, α-L-(-)-fucopyranose, and cellobiose (Figure 7A). Trehalose can be degraded by trehalase (TRE1) into α-D-glucose and β-D-glucose, then converted into D-glucose 6-phosephate by glucokinase [82]. Raffinose can be degraded into galactose-1-phosphate via galactose kinase (GALK), then converted into glucose-1-phosphate via galactose transferase (GALT) [83]; alternatively, raffinose can be degraded by acid or alkaline α-galactosidases into galactose and sucrose or by β-fructosidases into D-fructose and melibiose [84,85]. Galactinol can be degraded into myo-inositol by raffinose synthase [86]. Thus, trehalose, raffinose and galactinol could act as reserve carbohydrates in the paradormant axillary shoot buds; in addition, these storage carbohydrates could play roles for paradormancy maintainence. For example, trehalose (Tre) can act as a cryoprotectant, protein and membrane stabilizer and antioxidant [87]; its phosphate form, trehalose-6-phosphate (Tre6P), plays a central role in the coordination of metabolism with development and stress response [44,88]; raffinose (RFO) and galactinol can scavenge hydroxyl radicals to protect bud cells from oxidative damage [89], also act as membrane stabilizers, stress tolerance mediators, osmoregulator and mobile oligosaccharide signaling molecule [90]; galactionol is involved in drought tolerance in tea tree [91]. Cellulose degradation generate cellobiose , it’s higher content (Figure 7A) suggest that cell wall could be mobilized as alternative carbon source in the paradormant shoot buds.
In sharp contrast to above 8 sugars, 24 sugars or sugar alcohols showed lower contents in the paradormant shoot buds, and followed by an increase at 1- and 3- DAF, they are: glucose 2, glucose-6-phose, glucose-1-phosphate, gluconic lactone, myo-inositol 1, sedoheptulose, D-xylopyranose, 3-(1-ethoxy-ethoxy)-butan-1-ol, 2-butene-1,4-diol, fucose, α-hydroxypyruvic acid, D-(+)-cellobiose, L-rhamnose, ethyl α-D-glucopyranoside, α-L-galactopyranoside, tetrahydrofuran, myo-inositol 2, D-myo-inositol, D-xylose, D-(-)tagatofuranose, scyllo-inositol, 2-deoxyerythritol, methyl-4-O-methyl-α-D-glucopyranoside and D-galactose 2 (Figure 7A). In view of the substrate-product relations as we discussed above, it’s not surprising that the increase of glucose, G-6-P, G-1-P, and myo-inositol were accompanied by a concurrent reduction of trehalose, raffinose and galactinol (Figure 7A). G-6-P and G-1-P are activated substrates for glycolysis, their up-regulation at 1 DAF (Figure 7A) suggest that carbon status was improved in the axillary shoot buds after the axillary floral bud removal. Sedoheptulose is the precursor for shikimate synthesis, it reached the highest levels at 1 DAF (Figure 7A) which coincide with the up-regulation of phenolic acids and flavanols (Figure 4A-4B).
2.6. Free Fatty Acids and Sterols
Five free fatty acids (FFAs), including two saturate fatty acids (SFAs, palmitic acid, stearic acid) and three unsaturated fatty acids (UFAs, pentenoic acid, linoleic acid, and α-linolenic acid), were detected (Figure 7B). SFAs maintained constant during paradormancy-to-growth transition, while UFAs were kept at low level in paradormant shoot bud, then increased after paradormancy release (Figure 7B). Increased concentrations of free palmitic acid or decreased concentrations of free linolenic acid is reported to reduce plant membrane peroxidation [92], the low levels of UFAs may protect membrane lipids from oxidation in the paradormant shoot buds. Pentenoic acid has the activity of inhibiting FFA β-oxidation [93,94], the increased pentenoic acid levels at 1 DAF could reduce FFA β-oxidation leading to their accumulation (Figure 7B). These observation suggest that in the paradormant shoot buds UFAs could be preferably oxidized as an alternative carbon skeleton and energy source in response to carbon restriction.
Ethanolamine, a precursor for the synthesis of major membrane lipid phosphatidylethanolamine (PE), was lower in the paradormant shoot buds, then increased during paradormancy-to-growth transition (Figure 7B). Serine is the precursor for ethanolamine synthesis [95]; accordingly, both serine and ethanolamine shared similar changing trends during paradormancy-to-growth transition (Figure 6A; Figure 7B). At 3 DAF, the concurrent increase of FFAs, ethanolamine and membrane sterols (stigmasterol, α-sitosterol) (Figure 7B) would provide sufficient substrates for enhanced membrane lipid synthesis and assembly for the initiation of bud meristem cell division and expansion.
3. Discussion
3.1. Tre6P-SA Signaling May Participate Carbon Starvation-Induced Paradormancy in Tea Axillary Shoot Buds
Previous transcriptomic studies in herbaceous and deciduous perennial wood species demonstrate that carbon starvation is the initial cue to trigger bud dormancy [39,96,97]. In perennial evergreen wood species such as Camellia sinensis cv Huangdan, the growing auxiliary floral buds could act as a stronger carbon sink, and deprive carbon availability to the axillary shoot bud grown at the same leaf node (Figure 1), this could be the direct trigger leading to axillary shoot bud paradormancy. Unexpected, sucrose, the major transported carbon, were even slightly higher in paradormant shoot buds (Supplementary data 3; Supplementary data 4). At same time, several non-reducing disaccharides and trisaccharides, including trehalose, raffinose, galactinol, α-D-xylopyranose and cellobiose, were also higher in the paradormant leaf buds (Figure 7A). In addition, Glu, Asp, Asn and BCAAs also showed higher contents in the paradormant shoot buds (Figure 6A) whose catabolism can provide respiration substrates in mitochondria [98]. These data indicate that there was significant carbon reserve in the paradormant axillary shoot buds. In contrast, the reducing hexose pools, including glucose, G-1-P, G-6-P, D-xylose, myo-inositol and its stereoisomer scyllo-inositol, D-galactose and it isomer D-(-)-Tagatofuranose, were lower in the paradormant shoot buds (Figure 7A). Based on these observations, for evergreen plant species, the reducing hexose pool size could be a more reliable marker of carbon status compared to sucrose. The lower reducing hexose pool could lead to reduced respiration regardless the abundant presence of carbon reserve in the forms of oligomer or amino acids. This raise a question how subsets of sugar oligomers or amino acids are accumulated to higher levels in paradormant shoot buds when the carbon availability is restricted? One possibility is that the buildup of these subsets of primary metabolites could be resulted from enhanced de novo synthesis or reduced catabolism in the axillary shoot buds; alternatively, these metabolites could be transported from the nearby floral buds. Considering their competitive relationships for nutrients transported from the same source leaves, it seems unreasonable that the axillary floral buds “give” these carbon source to its competitor. However, this does not exclude the possibility that the axillary floral buds could deliver some unknown signal(s) to their nearby axillary shoot buds and manipulate the metabolism there.
G-1-P is the precursor for UDP-glucose synthesis, UDP-glucose and G-6-P are the substrates for Tre6P synthesis. With the lower levels of G-1-P and G-6-P in the paradormant shoot buds (Figure 7A), a lower Tre6P synthesis is expected. Meanwhile, Tre6P can be converted into trehalose [44]. The paradormant shoot bud showed the highest contents of trehalose (Figure 7A), suggesting an enhanced conversion of Tre6P to trehalose. With reduced synthesis of Tre6P and its enhanced conversion to trehalose, a lower level of Tre6P is expected in the paradormant shoot buds. Tre6P acts as a proxy for carbon status, may be involved in the sensing of low energy status [44]. Lower Tre6P levels reduce cell division and sucrose degradation, but enhance lipid mobilization and gluconeogenesis [99,100]. Accordingly, the paradormant shoot buds showed higher sucrose and lower FFAs contents (Supplementary data 3; Supplementary data 4; Figure 7B). The arrest of cell division would diminish the requirement of pectin and cellulose for cell wall synthesis [99], this may explain why the sugars involved in cell wall synthesis, including methyl galactoside, α-D-xylopyranose, α-L-(-)-fucopyranose, and D-(+)-galacturonic acid, showed higher contents in the paradormant shoot buds (Figure 7A). The floral bud removal essentially eliminates a strong competing sink, and make more carbon available for the axillary shoot buds, this was supported by the elevated G-6-P and G-1-P levels at 1 DAF (Figure 7A). Overall, our data support the notion that Tre6P could be involved in the sensing of the low carbon status in the paradormant shoot buds, and carbon status improvement could be the key event leading to paradormancy release of the axillary shoot buds.
SA exhibits multiple activities, such as inhibiting lipoxygenase (LOX), reducing JA synthesis [101,102], repressing auxin signaling [102], and promoting quiescent center cell division and intracellular redox homeostasis [73,103]. A higher SA levels in HD_0 samples (Figure 6B) could help paradormancy maintenance, a transit SA decline at 1 DAF (Figure 6B) could facilitate paradormancy release. We speculate that this transit SA decline at 1 DAF could be due to: 1) Considering that SA is easily transported across tissues [104,105], this raise the possibility that some SAs in the axillary shoot buds could be transported from the nearby axillary floral buds. Under this scenario, the floral bud removal concurrently eliminates SA supply leading to this transit decline in the axillary shoot buds at 1 DAF (Figure 6B); 2) the wounding caused by floral bud removal could activate the synthesis of jasmonic acid (JA) or ethylene in the local axillary shoot buds, JA and ethylene cross-talk with SA pathway and down-regulate SA levels [106,107]. In support this view, above ground shoot removal in herbaceous perennial leaf spurge lead to JA increase followed by gradual decrease [108]; in popular vegetative buds, SA and JA associated genes were also down-regulated during paradormancy-to-endodormancy transition [109].
Based on our results in this study and other reports discussed here, we propose a model in Figure 8 to explain how carbon status changes affect paradormancy set or release in tea axillary shoot buds. In response to carbon deprivation by the nearby growing axillary floral buds, hexose pool size in the axillary leaf buds was reduced (Figure 7A), this lead to reduced Tre6P levels through reduced synthesis and enhanced conversion into trehalose. Since Tre6P inhibit SnRK1 activity [110,111,112,113], the lower Tre6P level activates SnRK1 activity, which repress gibberellin and brassinosteroid signaling [114,115], and positively regulate SA pathway through bZIP11/63 [116,117], and result in SA accumulation (Figure 6B). Higher SA levels repress auxin signaling [102]. Meanwhile, higher SA inhibit CAT and APX activity [118], which lead to ROS buildup and antioxidant remodeling. These signaling events collectively maintain the axillary shoot bud in paradormancy state. When the axillary floral buds were removed, the carbon supply to the axillary shoot buds is recovered as supported by elevated hexose pool size (Figure 7A), the above processes were reversed. Meanwhile, wound-induced JAs synthesis in the axillary shoot buds could also be activated, which inhibit SA synthesis leading to its transit decline at 1 DAF (Figure 6B), these signaling events help break the bud paradormancy.
Exogenous application of SA has been reported to activate gene expression in quercetin and apigenin synthesis pathway and increased lutin contents [119]. Consistent with this report, a higher SA levels at HD-0 (Figure 6B) was accompanied with higher vitexin 2′-O-rhamonsie and apiegnin 6-C-glucoside 8-C-arabinoside contents followed by concurrently decrease at 1 DAF (Figure 5A). However, this correlation was not observed at 3 DAF, in which SA reached the highest level (Figure 6B) while Vitexin 2′-O-rhamonsie and apiegnin 6-C-glucoside 8-C-arabinoside decreased (Figure 5A). In contrast, shikimic pathway intermediates, including shikimic acid, benzoic acid (Figure 6B), gallic acid, methygallate, catechins and 5-p-coumaroylquinic acid (Figure 4A) reached to their highest levels. Considering that SA shows dosage-dependent effect [120], higher SA levels at 3 DAF could activate shikimate, flavonol and monolignin synthesis, thus alter carbon flux among branch routs for various flavonoid synthesis.
3.2. C and N Coordination in the Axillary Shoot Buds
In plants, sugars and amino acids are the major forms of carbon and nitrogen source, respectively. The paradormant axillary shoot buds showed characteristic carbon and nitrogen distribution patterns: for carbon, the non-reducing di- and tri- polysaccharides were highly accumulated while the reducing hexose levels decreased (Figure 7A); for nitrogen, Glu, Asp, Asn and branched chain amino acids were highly accumulated while other amino acids showed lower contents (Figure 6A). Even though theanine is the most abundant non-proteinaceous amino acid, its contents were low in the paradormant shoot buds, then accumulated to its highest level at 1 DAF followed by decline at 3 DAF (Figure 6A), suggesting that theanine play role to coordinate nitrogen metabolism during paradormancy-to-growth transition. Unlike theanine, Glu, Asp and Asn are the convenient storage forms of nitrogen, and can be easily converted into other amino acids or nitrogen-containing compounds. Similarly, the non-reducing di- or tri- polysaccharide can also be easily remobilized. These data suggest that the paradormant shoot buds are prepared to growth initiation once nutrients become available. That raises a question what are the driving force to shape this carbon and nitrogen distribution patterns in tea paradormant shoot buds? The initial step of nitrogen fixation starts with transamination to α-ketoglutaric acid which are diverted from TCA cycle (Figure S5). In the paradormant shoot buds, the higher citric acid content but lower malic acid and succinic acid contents (Figure 6B) suggest that α-ketoglutaric acid is diverted from TCA cycle, this may explain why Glu, Asp and Asn accumulate in the paradormant shoot buds to higher levels (Figure 6A). Threonine and lysine are derived from Asp (Figure S1), both also showed higher contents in paradormant shoot buds as Asp (Figure 6A). Val and Leu use glycolytic intermediate pyruvate as carbon skeleton for their synthesis (Figure S4). Accordingly, both of them showed similar changing trends during paradormancy-to-growth transition (Figure 6A). Val and Leu allosteric inhibit acetohydroxy acid synthase activity [64], the first enzyme in their synthesis pathway (Figure S4). A higher Val and Leu levels in the paradormant shoot buds would feedback inhibiting their own synthesis, thus divert pyruvate into TCA cycle which provide carbon skeleton for Asp and Glu synthesis. Val allosteric activates Asp kinase [121], the first enzyme for BCAAs synthesis (Figure S1); in addition, Val allosteric inhibits threonine deaminase (OMR1) activity [122], the first enzyme to convert threonine into isoleucine (Figure S1). These regulation loops could lead to the accumulation of Asp, Asn, Glu, Val and Leu simultaneously. Thus, even though the carbon supply is restricted in the paradormant shoot buds, carbon and nitrogen still can be preferably diverted to synthesize a specific set of amino acids.
Gln is synthesized from NH3 transamination to Glu (Figure S5). In the paradormant shoot buds, Glu was at the highest level while Gln was at the lowest level, this is in sharp contrast with Asn which showed the highest level (Figure 6A; Figure S5). These data suggest that nitrogen (NH3) supply in the paradormant shoot buds also was restricted. α-ketoglutarate has been suggested acting as nitrogen starvation signal in deciduous species, our data suggest that glutamine could act as proxies for N status in tea tree, both pyruvate and α-ketoglutarate could act as proxy for C status. Glutamine activates N-acetyl-L-glutamate kinase (NAGK) [60], the key enzyme of ornithine synthesis pathway. This may explain why a lower Gln, ornithine and citrulline were concurrently showed up in the paradormant shoot buds, and their contents were up-regulated simultaneously after paradormancy release (Figure 6A; Figure S5).
3.3. The Potential Roles of 3-HB for Phenylpropanoid Reconfiguration During Paradormancy-to- Growth Transition in the Axillary Shoot Buds
Bud development involves an extensive reconfiguration of metabolism [123]. The association of the phenylpropanoid with bud dormancy has been reported in apricot [8]. In almond flower buds, phenylpropanoid compounds are involved in the endodormancy release [11]. In this study, we found that flavonoids in the axillary shoot buds were remodeled dramatically after the axillary floral bud removal (Figure 4; Figure 5), these data suggest that phenylpropanoids are involved in paradormancy release in tea plants. One intriguing question is that how tea buds reconfigure phenylpropanoid metabolism? Here, we suggest that 3-HB could be involved in this process for the following reasons: 1) 3-HB has the potential to regulate gene expression epigenetically. 3-HB is an endogenous activator of methyltransferase 3 (CMT3), decrease the activities of DNA methylation I (DDM1) and DEMETER DNA glycosylase (DME), and act as an inhibitor of histone deacetylase sirtuin 1 (SRT1) [79,81,124]. 2) Increasing 3-HB levels by transgenic overexpression of bacterial β-ketothiolase in planta or exogenous application of 3-HB activated most genes involved in the lignin and phenolic acid synthesis pathway [79]. 3) The lower 3-HB levels in the paradormant shoot buds (Figure 6B) could reduce monolignol synthesis, thus push caffeate into the synthesis of dihydrocaffeic acid 3-O-glucuronide (Figure S2). Similarly, reduced carbon flux into flavanol and flavonol synthesis is expected to push more carbon into flavone synthesis (Figure S2) leading to higher contents of apigenin glycosides and vitexin glycosides as we observed (Figure 5A). 4) 3-HB was increased transit at 1 DAF (Figure 6B). Accordingly, flavonol, flavanol and flavone accumulation were reversed (Figure 5B), and a dramatic increase in shikimic acid, benzoic acid, salicylic acid was followed at 3 DAF (Figure 6B). Thus, our data provide independent evidence that 3-HB could activate genes in the phenylpropanoid pathway as reported in other plant species [79].
3.4. The Non-Enzymatic Antioxidants Remodeling Could Regulate Meristematic Cell Activity and the Paradormancy-to-Growth Transition in the Axillary Shoot Buds
Reactive oxygen species (ROS) have different forms including free radical (O2•-, superoxide anion radical; OH•, hydroxyl radical; HO2•, perhydroxy radical and RO•, alkoxy radicals) and non-radical forms (H2O2, hydrogen peroxide and 1O2, single oxygen) [125], O2•- and H2O2 are the most stable forms of ROS, while OH• and1O2 have shorter lifetimes [126]. At normal state ROS are kept at low levels, thus, a small increase can result in large changes, this feature makes ROS acting as signaling molecules in plants. However, high levels in ROS can cause oxidative damage. Accordingly, plants possess enzymatic and non-enzymatic antioxidant defense system to scavenge ROS [127]. The non-enzymatic antioxidants include flavonoids, sugar alcohols, amino acids, alkaloids, carotenoids, α-tocopherols, ascorbic acid, and glutathione (GSH) [125,128,129,130,131].
Recently, ROS are recognized as signaling component regulating meristem activities [23]. ROS distribute along meristematic tissues in a gradient manner to maintain stem cell niche, O2•− is markedly enriched in the central zone of SAM to maintain stemness, while H2O2 accumulates in the peripheral zone to promote stem cell differentiation [23,24,132,133]. Since ROS homeostasis is maintained by ROS-producing and scavenging systems, it is expected that the non-enzymatic antioxidants could play important role to shape the ROS gradient along meristematic tissues, thus, affect stem cell division and differentiation and facilitate bud paradormancy set or release.
Flavonoids include flavonols, flavones, flavanols (catechins), flavanones, flavanonols, anthocyanins, proanthocyanidins and isoflavonoids [134]. The flavonoid backbones can be modified by hydroxylation, methylation, glycosylation, and/or acylation, which alter their ROS scavenge ability. As a result, some flavonoid subclasses possess higher antioxidant capacity and have a more significant role in redox-regulated processes. For example, flavanols (catechins) show high scavenging ability for H2O2 but very low scavenge ability to 1O2 [135,136]. Flavonol compounds (kaempferol, quercetin, rutin and myricetin) have distinct antioxidant avtivities [136], myricetin perform better at scavenging 1O2 than quercetin, while quercetin has the highest antioxidant capacity for H2O2 followed by kaempferol, myricetin [135]. In addition, quercetin, rutin and myricetin can suppress lipid peroxidation and reduce ROS generation in peroxisome. Flavonols have the potential to locally regulate ROS levels in cytoplasmic and nuclear [137,138]. In contrast, flavone compounds (apigenin, apigenin glycosides and vitexin glycosides) are the least effective with no antioxidant capacity for H2O2 and very little capacity for 1O2 [72,139]. Proanthocyanidins are derived from flavanol oligomerization [140], it also can bind proteins and metals to reduce metal-mediated ROS production [54]. During paradormancy-to-growth transition, phenolics, flavanol and flavonol contents increased, while proanthocyanidin and flavone contents decreased (Figure 4; Figure 5). These flavonoid reconfigurations could be an important aspect to regulate ROS homeostasis in the axillary shoot buds.
Besides flavonoids, sugar and sugar alcohols (trehalose, raffinose, galactinol and myo-inositol) also have antioxidant activities [83,89,141,142]. In paradormancy state, their higher levels (Figure 7A) could compensate the lower levels of phenolic compounds (Figure 4) to maintain ROS homeostasis. In addition, some amino acid (theanine and proline) also possess anti oxidative activities [142,143,144]. During paradormancy-to-growth transition, theanine contents increased while proline contents kept constant (Figure 6A). Alkaloids (theobromine and caffeine) possesses potent antioxidant activities [145]. During paradormancy-to-growth transition, theobromine content increased while caffeine content kept constant (Supplementary Data 4). These diverse changes from different subclasses of non-enzymatic antioxidants may provide a robust system to fine-tune various types of ROS. In view that different types of ROS accumulate in different meristematic zones and show diverse activities toward cell division or differentiation [146], the reconfigurations of these diverse non-enzymatic antioxidants could regulate bud meristem growth and paradormancy-to-growth transition.
4. Materials and Methods
4.1. Plant Material
Camellia sinensis cv Huangdan was grown in the tea garden at Fujian Agriculture and Forestry University campus (Fuzhou, China; 26°08′19″ N, 119°24′06″ E) for 25 years. On August 30, 2016, axillary floral buds were carefully removed from each leaf node and kept axillary shoot bud in place, then harvested on August 31(HD_1 sample) and September 2 (HD_3 sample). Some axillary shoot buds were harvested on August 30 right before floral bud removal as control (HD_0 sample).
4.2. Metabolite Extraction
~40 mg axillary shoot buds were weighed into a 1.5 mL centrifuge tube, grounded into powder with a plastic pestle in the presence of liquid nitrogen, a 200 μL of 80% (v/v) methanol (chromatographic grade) was added and ultrasonicated for 30 min under 25 ℃, then centrifuged at 12000 g for 15 min (25℃), the supernatant was collected into a 1.5 mL tube; the pellet was re-extracted 2 more times as above, the supernatant was pooled into the same tube. Then, the pellet was re-extracted 3 more times by 200 μL of 80% methanol containing 0.1% (v/v) formic acid as extraction solvent, the supernatant was pooled and filtered through 0.22 μm membrane, then divided into two parts for UPLC-MS and GC-TOF MS analysis, respectively. For LC-MS analysis, 20 μL filtrate was transferred into a fresh 2-mL GC vial, diluted 50 x by 80% methanol/0.1% (v/v) formic acid solution. For GC-TOF MS analysis, 500 μL filtrate was aliquot into a new GC vial, dried down completely in CentriVap Console (Labconco, KS, USA). 80 μL methoxyl amine (20 mg/mL) in pyridine was added to the pellet, ultrasonicated to dissolve the pellet, kept at room temperature for 30 min, then 80 μL N-methyl-N-(trimethylsiyl) trifluoroacetamide (MSTFA) plus 1% (v/v) trimethylchlorosilane (TMCS) was added, placed in 70℃ oven for 60 min, then left at room temperature under dark for additional 2 h, 160 μL pyridine was added into the derivitized sample before sample injection [147].
4.3. GC-TOF MS Analysis
Derivitized samples were analyzed using an Agilent 7890B gas chromatography (Agilent, Santa Clara, CA, USA) coupled with a Pegasus HT time-of-flight mass spectrometer (LECO, Saint Joseph, MI, USA). The capillary column is Rxi-5Sil (30 m x 0.25 mm x 0.25 μm, Restek, USA), helium is used as the carrier gas with a flow rate of 1 mL min-1. One μL of sample was injected in splitless mode. The temperature settings are as following: injector, 270 ℃; transmission line, 275 ℃; ion source, 250 ℃. The oven was initiated at 80 ℃ with 5 min solvent delay, then ramped at 5 ℃ min-1 to 310 ℃, held for 6 min before next sample injection. The electron collision energy was -70 eV, the detector voltage was 1440 V. The scan range was 45-600 atomic mass unit (AMU) with acquisition rate of 10 spectra s-1. Raw data were processed by Chroma TOF software (ver 4.51.6, LECO, St. Joseph, MI, USA), metabolites were identified by quest NIST library or comparing with authentic standards, the mean area of the selected characteristic ions were used for quantification.
4.4. UPLC-QTOF MS Analysis
Tea metabolites also was analyzed by Waters Acquity ultra-performance liquid chromatography coupled to a SYNAPT G2-Si HDMS QTOF mass spectrometry (Waters, Manchester, UK). One μL of sample was injected into Acquity UPLC HSS T3 column (100 x 2.1 mm, 1.8 μm, Waters, Manchester, UK) for chromatographic separation. The flow rate was 0.3 mL min-1. The mobile phase A and B were water/0.1% (v/v) formic acid and acetonitrile/0.1% (v/v) formic acid, respectively. The linear elution gradient was: 0 min, 1% B; 2 min, 7% B; 13 min, 40% B; 14 min, 99% B; 18 min, 99% B; 18.1 min, 1% B; held for 5 min before next sample injection. Each sample was analyzed in the positive ion mode and the negative ion mode, respectively. The MS was acquired in continuum mode with 10 to 40 eV collision energy. The mass parameter setting was: capillary voltage, 1.5 kV (ESI+) and 2.5 kV (ESI-); ion source temperature, 120 ℃; desolvation temperature, 500 ℃; cone gas flow rate, 50 L/h; desolvation gas flow, 800 L/h; m/z range, 50-1200 Da. The instrument was operated by MassLynx software (version 4.1, Waters, Milford, MA, USA) [148]. UPLC-MS data were processed in Progenesis QI (v2.1, Nonlinear Dynamics, Newcastle upon Tyne, UK) with default settings, normalized peak areas were exported to Microsoft Office Excel, outliers were manually removed. Metabolites were identified by query an in-house authentic standard spectra library and online spectral databases (Metlin; MassBank; HMDB; KNApSAcK; ReSpect) [149,150,151,152,153].
4.5. Data Processing
MS peak areas were extracted, and normalized to sample weight. To compare individual metabolite changes across time, the average peak area of HD_0 samples was used as dividend, such that each individual metabolite contents in HD_0 samples were artificially set as 1.
4.6. PLS-DA and Heat-Map Analysis
Partial least squares discriminate analysis (PLS-DA) was performed by using SIMCA-P+ 14.1 software (Umetrics, Umeå, Sweden). For heat-map analysis, the metabolite contents were log2 transformed, log2 values equal to 0, <0, and >0 were presented as black, green and red color, respectively, such that the green color represents content lower than that of HD_0 control, while red color represents content higher than that of HD_0 control.
5. Conclusions
Camellia sinensis cv Huangdan develops the axillary floral buds and the axillary shoot buds at the same leaf nodes simultaneously, the growing axillary floral buds deprive nutrient from the axillary shoot buds leading to their carbon starvation and paradormancy. We took advantage of this developmental event, and dissect the metabolic regulations of paradormancy set and release in a perennial evergreen tree species. We performed an extensive metabolomics comparison in the axillary shoot buds before and after the axillary floral bud removal, and demonstrated that the axillary shoot buds exhibit characteristic carbon and nitrogen distribution patterns indicating restricted carbon and nitrogen availability, this could be the major cause leading to axillary shoot bud paradormancy. Reducing hexose pool size could be a reliable marker for carbon status, while glutamine could act as proxy of cell’s nitrogen status in tea tree buds. In paradormant shoot buds, cell wall and membrane lipids could be mobilized as alternative carbon source. The paradormancy-to-growth transition involve a complex metabolome reconfiguration, here we mainly discussed metabolic remodeling of phenylpropanoids, amino acids, organic acids, sugars and sugar alcohols, free fatty acids and sterols. We propose that remodeling of a variety of non-enzymatic antioxidants could be an important aspect to shape the ROS gradients along the meristematic tissues which signal cell division and differentiation. This research revealed novel insights about paradormancy set and release in a perennial evergreen tree species.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Schematic presentation of aspartate-derived amino acid synthesis and regulation. Red line, activation; black line, inhibition; Figure S2: Schematic presentation of phenylpropanoid synthesis pathway; Figure S3: Schematic presentation of TCA cycle and glyoxalate cycle; Figure S4: Schematic presentation of valine and leucing synthesis pathway and regulation. Pyr: pyruvate; 2AL: 2-acetolactate; 2,3DH3MB: 2,3-dihydroxy-3-methylbutanoate; 3MOB: 3-methyl-2-oxobutanoate; 4MOP: 4-methyl-2-oxopentanoate; 2-IPM: 3-isopropylmalate; 3-IPM: 2-isopropylmalate; AHASL, acetohydroxy acid synthase large subunit; AHASS1 and AHASS2, acetohydroxy acid synthase small subunit 1 and 2; Figure S5: Schematic presentation of nitrogen assimilation pathway in plants; Supplementary data 1: UPLC-QTOF MS metabolomic data collected in negative ion mode; Supplementary Data 2: UPLC-QTOF MS metabolomic data collected in positive ion mode; Supplementary Data 3: Chemically identified metabolites from UPLC-QTOF MS in negative ion mode; Supplementary Data 4: Chemically identified metabolites from UPLC-QTOF MS in positive ion mode; Supplementary Data 5: GC-TOF MS metabolomic data; Supplementary Data 6: Chemically identified metabolites from GC-TOF MS.
Author Contributions
Conceptualization, M.C. and C.C.; methodology, M.C., Z.D., W.Y., X.K., Q.X., D.F.; software, Z.D. and W.Y.; validation, Z.D. and W.Y.; formal analysis, M.C., W.Y., X.K., Q.X. and D.F.; investigation, Z.D., X.K., Q.X. and D.F.; resources, M.C., C.C., X.K.; data curation, Z.D., M.C., W.Y., Q.X.; writing—original draft preparation, M.C.; writing—review and editing, M.C., Z.D., W.Y., X.K. and C.C.; visualization, M.C., W.Y.; supervision, M.C.; project administration, M.C., Z.D., C.C.; funding acquisition, M.C., C.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was sponsored and supported by the National Natural Science Foundation of China (grant number 31870803) and Fujian Provincial Natural Science Foundation(grant number 2021J01491).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials. Further inquiries can be directed to the corresponding author.
Acknowledgments
The metabolomics analysis was conducted by the Center for Horticultural Biology and Metabolomics, Haixia Institute of Science and Technology, Fujian Agriculture and Forestry University.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Garrison, R. Studies in the development of axillary buds. Am. J. Bot. 1955, 42, 257–266. [Google Scholar] [CrossRef]
- Barua, D.N. Seasonal Dormancy in Tea (Camellia sinensis L.). Nature 1969, 224, 514–514. [Google Scholar] [CrossRef]
- Cooke, J.E.K.; Eriksson, M.E.; Junttila, O. The dynamic nature of bud dormancy in trees: environmental control and molecular mechanisms. Plant, Cell Environ. 2012, 35, 1707–1728. [Google Scholar] [CrossRef]
- Horvath, D.P. , Anderson, J. V., Chao, W.S., Foley, M.E. Knowing when to grow: signals regulating bud dormancy. Trends Plant Sci. 2003, 8, 534–540. [Google Scholar]
- Paul, A.; Kumar, S. Responses to winter dormancy, temperature, and plant hormones share gene networks. Funct. Integr. Genom. 2011, 11, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo-, Para-, and Ecodormancy: Physiological Terminology and Classification for Dormancy Research. HortScience 1987, 22, 371–377. [Google Scholar] [CrossRef]
- Anderson, J.V.; Gesch, R.W.; Jia, Y.; Chao, W.S.; Horvath, D.P. Seasonal shifts in dormancy status, carbohydrate metabolism, and related gene expression in crown buds of leafy spurge. Plant, Cell Environ. 2005, 28, 1567–1578. [Google Scholar] [CrossRef]
- Conrad, A.; Yu, J.; E Staton, M.; Audergon, J.-M.; Roch, G.; Decroocq, V.; Knagge, K.; Chen, H.; Zhebentyayeva, T.; Liu, Z.; et al. Association of the phenylpropanoid pathway with dormancy and adaptive trait variation in apricot (Prunus armeniaca). Tree Physiol. 2019, 39, 1136–1148. [Google Scholar] [CrossRef]
- Del Cueto, J.; Ionescu, I.A.; Pičmanová, M.; Gericke, O.; Motawia, M.S.; Olsen, C.E.; Campoy, J.A.; Dicenta, F.; Møller, B.L.; Sánchez-Pérez, R. Cyanogenic Glucosides and Derivatives in Almond and Sweet Cherry Flower Buds from Dormancy to Flowering. Front. Plant Sci. 2017, 8, 800. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, J.; Chen, Y.; Tang, H.; Wang, Y.; He, Y.; Ou, Y.; Sun, X.; Wang, S.; Yao, Y. Tomato SlAN11 regulates flavonoid biosynthesis and seed dormancy by interaction with bHLH proteins but not with MYB proteins. Hortic. Res. 2018, 5, 1–18. [Google Scholar] [CrossRef]
- Guillamón, J.G.; Prudencio. S.; Yuste, J.E.; Dicenta, F.; Sánchez-Pérez, R. Ascorbic acid and prunasin, two candidate biomarkers for endodormancy release in almond flower buds identified by a nontargeted metabolomic study. Hortic. Res. 2020, 7, 1–13. [Google Scholar] [CrossRef]
- Kaufmann, H.; Blanke, M. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. J. Plant Physiol. 2017, 218, 1–5. [Google Scholar] [CrossRef]
- Ruttink, T.; Arend, M.; Morreel, K.; Storme, V.; Rombauts, S.; Fromm, J.; Bhalerao, R.P.; Boerjan, W.; Rohde, A. A Molecular Timetable for Apical Bud Formation and Dormancy Induction in Poplar. Plant Cell 2007, 19, 2370–2390. [Google Scholar] [CrossRef] [PubMed]
- Rinne, P.L.; Paul, L.K.; Vahala, J.; Kangasjärvi, J.; van der Schoot, C. Axillary buds are dwarfed shoots that tightly regulate GA pathway and GA-inducible 1,3-β-glucanase genes during branching in hybrid aspen. J. Exp. Bot. 2016, 67, 5975–5991. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Yang, Q.; Ni, J.; Gao, Y.; Tang, Y.; Bai, S.; Teng, Y. Early defoliation induces auxin redistribution, promoting paradormancy release in pear buds. Plant Physiol. 2022, 190, 2739–2756. [Google Scholar] [CrossRef]
- Ibáñez, C.; Kozarewa, I.; Johansson, M.; Ögren, E.; Rohde, A.; Eriksson, M.E. Circadian Clock Components Regulate Entry and Affect Exit of Seasonal Dormancy as Well as Winter Hardiness in Populus Trees. Plant Physiol. 2010, 153, 1823–1833. [Google Scholar] [CrossRef]
- Kuroda, H.; Sugiura, T.; Ito, D. Changes in Hydrogen Peroxide Content in Flower Buds of Japanese Pear(Pyrus pyrifolia Nakai) in Relation to Breaking of Endodormancy. J. Jpn. Soc. Hortic. Sci. 2002, 71, 610–616. [Google Scholar] [CrossRef]
- Mazzitelli, L.; Hancock, R.D.; Haupt, S.; Walker, P.G.; Pont, S.D.A.; McNicol, J.; Cardle, L.; Morris, J.; Viola, R.; Brennan, R.; et al. Co-ordinated gene expression during phases of dormancy release in raspberry (Rubus idaeus L.) buds. J. Exp. Bot. 2007, 58, 1035–1045. [Google Scholar] [CrossRef]
- Pérez, F.J.; Vergara, R.; Rubio, S. H2O2 is involved in the dormancy-breaking effect of hydrogen cyanamide in grapevine buds. Plant Growth Regul. 2008, 55, 149–155. [Google Scholar] [CrossRef]
- Sreekantan, L.; Mathiason, K.; Grimplet, J.; Schlauch, K.; Dickerson, J.A.; Fennell, A.Y. Differential floral development and gene expression in grapevines during long and short photoperiods suggests a role for floral genes in dormancy transitioning. Plant Mol. Biol. 2010, 73, 191–205. [Google Scholar] [CrossRef]
- Wu, R.-M.; Walton, E.F.; Richardson, A.C.; Wood, M.; Hellens, R.P.; Varkonyi-Gasic, E. Conservation and divergence of four kiwifruit SVP-like MADS-box genes suggest distinct roles in kiwifruit bud dormancy and flowering. J. Exp. Bot. 2011, 63, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Shi, T.; Gao, Z.; Zhang, Z.; Zhang, J. Differential expression of proteins associated with seasonal bud dormancy at four critical stages in Japanese apricot. Plant Biol. 2012, 15, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q. ROS: Important factor in plant stem cell fate regulation. J. Plant Physiol. 2023, 289, 154082. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Dong, Z.; Wu, H.; Tian, Z.; Zhao, Z. Redox regulation of plant stem cell fate. EMBO J. 2017, 36, 2844–2855. [Google Scholar] [CrossRef]
- Horvath, D.P.; Chao, W.S.; Suttle, J.C.; Thimmapuram, J.; Anderson, J.V. Transcriptome analysis identifies novel responses and potential regulatory genes involved in seasonal dormancy transitions of leafy spurge (Euphorbia esula L.). BMC Genom. 2008, 9, 536–536. [Google Scholar] [CrossRef]
- Leida, C.; Terol, J.; Martí, G.; Agustí, M.; Llácer, G.; Badenes, M.L.; Ríos, G. Identification of genes associated with bud dormancy release in Prunus persica by suppression subtractive hybridization. Tree Physiol. 2010, 30, 655–666. [Google Scholar] [CrossRef]
- Zhong, W.; Gao, Z.; Zhuang, W.; Shi, T.; Zhang, Z.; Ni, Z. Genome-wide expression profiles of seasonal bud dormancy at four critical stages in Japanese apricot. Plant Mol. Biol. 2013, 83, 247–264. [Google Scholar] [CrossRef]
- Tanton, T.W. The Banjhi (Dormancy) Cycle in Tea (camellia sinensis). Exp. Agric. 1981, 17, 149–156. [Google Scholar] [CrossRef]
- Arana, M.V.; la Rosa, N.M.-D.; Maloof, J.N.; Blázquez, M.A.; Alabadí, D. Circadian oscillation of gibberellin signaling in Arabidopsis. Proc. Natl. Acad. Sci. 2011, 108, 9292–9297. [Google Scholar] [CrossRef]
- Hao, X.; Yang, Y.; Yue, C.; Wang, L.; Horvath, D.P.; Wang, X. Comprehensive Transcriptome Analyses Reveal Differential Gene Expression Profiles of Camellia sinensis Axillary Buds at Para-, Endo-, Ecodormancy, and Bud Flush Stages. Front. Plant Sci. 2017, 8, 553. [Google Scholar] [CrossRef]
- Nagar, P.K.; Kumar, A. Changes in endogenous gibberellin activity during winter dormancy in tea (Camellia sinensis (L.) O. Kuntze). Acta Physiol. Plant. 2000, 22, 439–443. [Google Scholar] [CrossRef]
- Nagar, P.K. Changes in endogenous abscisic acid and phenols during winter dormancy in tea [Camellia sinensis L. (O) Kuntze]. Acta Physiol. Plant. 1996, 18, 33–36. [Google Scholar]
- Nagar, P.K.; Sood, S. Changes in endogenous auxins during winter dormancy in tea (Camellia sinensis L.) O. Kuntze. Acta Physiol. Plant. 2006, 28, 165–169. [Google Scholar] [CrossRef]
- Nandi, S.K. , Palni, L.M.S. Shoot growth and winter dormancy in tea. In: NAIR, M. K (ed.): PLACROSYM X. 1992. J. Plant Crops 1993, 21 (Suppl.), 328–332.
- Wang, X.-C.; Zhao, Q.-Y.; Ma, C.-L.; Zhang, Z.-H.; Cao, H.-L.; Kong, Y.-M.; Yue, C.; Hao, X.-Y.; Chen, L.; Ma, J.-Q.; et al. Global transcriptome profiles of Camellia sinensis during cold acclimation. BMC Genom. 2013, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hao, X.; Ma, C.; Cao, H.; Yue, C.; Wang, L.; Zeng, J.; Yang, Y. Identification of differential gene expression profiles between winter dormant and sprouting axillary buds in tea plant (Camellia sinensis) by suppression subtractive hybridization. Tree Genet. Genomes 2014, 10, 1149–1159. [Google Scholar] [CrossRef]
- Chao, W.S.; Serpe, M.D. Changes in the expression of carbohydrate metabolism genes during three phases of bud dormancy in leafy spurge. Plant Mol. Biol. 2009, 73, 227–239. [Google Scholar] [CrossRef]
- Horvath, D.P. Role of mature leaves in inhibition of root bud growth inEuphorbia esulaL. Weed Sci. 1999, 47, 544–550. [Google Scholar] [CrossRef]
- Tarancón, C.; González-Grandío, E.; Oliveros, J.C.; Nicolas, M.; Cubas, P. A Conserved Carbon Starvation Response Underlies Bud Dormancy in Woody and Herbaceous Species. Front. Plant Sci. 2017, 8, 788. [Google Scholar] [CrossRef]
- Li, N.-N.; Qian, W.-J.; Wang, L.; Cao, H.-L.; Hao, X.-Y.; Yang, Y.-J.; Wang, X.-C. Isolation and expression features of hexose kinase genes under various abiotic stresses in the tea plant ( Camellia sinensis ). J. Plant Physiol. 2017, 209, 95–104. [Google Scholar] [CrossRef]
- Qian, W.; Yue, C.; Wang, Y.; Cao, H.; Li, N.; Wang, L.; Hao, X.; Wang, X.; Xiao, B.; Yang, Y. Identification of the invertase gene family (INVs) in tea plant and their expression analysis under abiotic stress. Plant Cell Rep. 2016, 35, 2269–2283. [Google Scholar] [CrossRef]
- Yue, C.; Cao, H.-L.; Wang, L.; Zhou, Y.-H.; Huang, Y.-T.; Hao, X.-Y.; Wang, Y.-C.; Wang, B.; Yang, Y.-J.; Wang, X.-C. Effects of cold acclimation on sugar metabolism and sugar-related gene expression in tea plant during the winter season. Plant Mol. Biol. 2015, 88, 591–608. [Google Scholar] [CrossRef] [PubMed]
- Lunn, J.E.; Delorge, I.; Figueroa, C.M.; Van Dijck, P.; Stitt, M. Trehalose metabolism in plants. Plant J. 2014, 79, 544–567. [Google Scholar] [CrossRef]
- Paul, M.J.; Primavesi, L.F.; Jhurreea, D.; Zhang, Y. Trehalose Metabolism and Signaling. Annu. Rev. Plant Biol. 2008, 59, 417–441. [Google Scholar] [CrossRef]
- Baena-González, E.; Lunn, J.E. SnRK1 and trehalose 6-phosphate – two ancient pathways converge to regulate plant metabolism and growth. Curr. Opin. Plant Biol. 2020, 55, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zeng, L.; Chen, Y.; Wang, X.; Liao, Y.; Xiao, Y.; Fu, X.; Yang, Z. Metabolism of Gallic Acid and Its Distributions in Tea (Camellia sinensis) Plants at the Tissue and Subcellular Levels. Int. J. Mol. Sci. 2020, 21, 5684. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, L.; Liu, L.; Yang, Q.; Lu, Z.; Nie, Z.; Wang, Y.; Xia, T. Purification and Characterization of a Novel Galloyltransferase Involved in Catechin Galloylation in the Tea Plant (Camellia sinensis). J. Biol. Chem. 2012, 287, 44406–44417. [Google Scholar] [CrossRef]
- Niemetz, R.; Gross, G.G. Enzymology of gallotannin and ellagitannin biosynthesis. Phytochemistry 2005, 66, 2001–2011. [Google Scholar] [CrossRef]
- Chen, M.; Kong, X.; Zhang, Y.; Wang, S.; Zhou, H.; Fang, D.; Yue, W.; Chen, C. Metabolomic Profiling in Combination with Data Association Analysis Provide Insights about Potential Metabolic Regulation Networks among Non-Volatile and Volatile Metabolites in Camellia sinensis cv Baijiguan. Plants 2022, 11, 2557. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Barberán, F.A.; Gil, M.I.; Castañer, M.; Artés, F.; Saltveit, M.E. Effect of Selected Browning Inhibitors on Phenolic Metabolism in Stem Tissue of Harvested Lettuce. J. Agric. Food Chem. 1997, 45, 583–589. [Google Scholar] [CrossRef]
- Schlernitzauer, A.; Oiry, C.; Hamad, R.; Galas, S.; Cortade, F.; Chabi, B.; Casas, F.; Pessemesse, L.; Fouret, G.; Feillet-Coudray, C.; et al. Chicoric Acid Is an Antioxidant Molecule That Stimulates AMP Kinase Pathway in L6 Myotubes and Extends Lifespan in Caenorhabditis elegans. PLOS ONE 2013, 8, e78788. [Google Scholar] [CrossRef]
- Kim, M.; Yoo, G.; Randy, A.; Kim, H.S.; Nho, C.W. Chicoric acid attenuate a nonalcoholic steatohepatitis by inhibiting key regulators of lipid metabolism, fibrosis, oxidation, and inflammation in mice with methionine and choline deficiency. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Lipchock, J.M.; Hendrickson, H.P.; Douglas, B.B.; Bird, K.E.; Ginther, P.S.; Rivalta, I.; Ten, N.S.; Batista, V.S.; Loria, J.P. Characterization of Protein Tyrosine Phosphatase 1B Inhibition by Chlorogenic Acid and Cichoric Acid. Biochemistry 2016, 56, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Sarnala, S. Proanthocyanidin Biosynthesis—a Matter of Protection. Plant Physiol. 2020, 184, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Mohanpuria, P.; Kumar, V.; Ahuja, P.S.; Yadav, S.K. Agrobacterium-Mediated Silencing of Caffeine Synthesis through Root Transformation in Camellia sinensis L. Mol. Biotechnol. 2010, 48, 235–243. [Google Scholar] [CrossRef]
- Xia, E.-H.; Zhang, H.-B.; Sheng, J.; Li, K.; Zhang, Q.-J.; Kim, C.; Zhang, Y.; Liu, Y.; Zhu, T.; Li, W.; et al. The Tea Tree Genome Provides Insights into Tea Flavor and Independent Evolution of Caffeine Biosynthesis. Mol. Plant 2017, 10, 866–877. [Google Scholar] [CrossRef]
- Chen, M.; Rao, R.S.P.; Zhang, Y.; Zhong, C.; Thelen, J.J. Metabolite variation in hybrid corn grain from a large-scale multisite study. Crop. J. 2016, 4, 177–187. [Google Scholar] [CrossRef]
- Sun, M.; Yang, F.; Hou, W.; Jiang, S.; Yang, R.; Zhang, W.; Chen, M.; Yan, Y.; Tian, Y.; Yuan, H. Dynamic Variation of Amino Acid Contents and Identification of Sterols in Xinyang Mao Jian Green Tea. Molecules 2022, 27, 3562. [Google Scholar] [CrossRef]
- Wei, C. , Yang, H., Wang, S., Zhao, J., Liu, C., Gao, L., Xia, E., Lu, Y., Tai, Y., She, G., et al., 2018. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA, 115, E4145–E4158.
- Chellamuthu, V.-R.; Ermilova, E.; Lapina, T.; Lüddecke, J.; Minaeva, E.; Herrmann, C.; Hartmann, M.D.; Forchhammer, K. A Widespread Glutamine-Sensing Mechanism in the Plant Kingdom. Cell 2014, 159, 1188–1199. [Google Scholar] [CrossRef]
- Fait, A.; Nesi, A.N.; Angelovici, R.; Lehmann, M.; Pham, P.A.; Song, L.; Haslam, R.P.; Napier, J.A.; Galili, G.; Fernie, A.R. Targeted Enhancement of Glutamate-to-γ-Aminobutyrate Conversion in Arabidopsis Seeds Affects Carbon-Nitrogen Balance and Storage Reserves in a Development-Dependent Manner. Plant Physiol. 2011, 157, 1026–1042. [Google Scholar] [CrossRef]
- Kanamori, T. , Matsumoto, H. Glutamine synthase from rice plant roots. Archives of Biochemistry and Biophysics. 1972, 152, 404–412. [Google Scholar] [CrossRef]
- Lee, Y. , Lee, C. B., Kim, S.G., Kwon, Y.M. Purification carbamoyltransferase and characterization of ornithine from the chloroplasts of Canavalia lineata leaves. Plant Science 1997, 122, 217–224. [Google Scholar]
- Xing, A.; Last, R.L. A Regulatory Hierarchy of the Arabidopsis Branched-Chain Amino Acid Metabolic Network. Plant Cell 2017, 29, 1480–1499. [Google Scholar] [CrossRef] [PubMed]
- Joshi, V.; Laubengayer, K.M.; Schauer, N.; Fernie, A.R.; Jander, G. Two Arabidopsis Threonine Aldolases Are Nonredundant and Compete with Threonine Deaminase for a Common Substrate Pool. Plant Cell 2006, 18, 3564–3575. [Google Scholar] [CrossRef] [PubMed]
- Ben Mohamed, H.; Vadel, A.M.; Geuns, J.M.; Khemira, H. Biochemical changes in dormant grapevine shoot tissues in response to chilling: Possible role in dormancy release. Sci. Hortic. 2010, 124, 440–447. [Google Scholar] [CrossRef]
- Auslender, E.L.; Dorion, S.; Dumont, S.; Rivoal, J. Expression, purification and characterization of Solanum tuberosum recombinant cytosolic pyruvate kinase. Protein Expr. Purif. 2015, 110, 7–13. [Google Scholar] [CrossRef]
- Meyer, R.C.; Steinfath, M.; Lisec, J.; Becher, M.; Witucka-Wall, H.; Törjék, O.; Fiehn, O.; Eckardt. ; Willmitzer, L.; Selbig, J.; et al. The metabolic signature related to high plant growth rate in Arabidopsis thaliana. Proc. Natl. Acad. Sci. 2007, 104, 4759–4764. [Google Scholar] [CrossRef]
- Marsh, K.B.; Boldingh, H.L.; Shilton, R.S.; Laing, W.A. Changes in quinic acid metabolism during fruit development in three kiwifruit species. Funct. Plant Biol. 2009, 36, 463–470. [Google Scholar] [CrossRef]
- Ha, C.M.; Escamilla-Trevino, L.; Yarce, J.C.S.; Kim, H.; Ralph, J.; Chen, F.; Dixon, R.A. An essential role of caffeoyl shikimate esterase in monolignol biosynthesis in Medicago truncatula. Plant J. 2016, 86, 363–375. [Google Scholar] [CrossRef]
- Hoffmann, L.; Maury, S.; Martz, F.; Geoffroy, P.; Legrand, M. Purification, Cloning, and Properties of an Acyltransferase Controlling Shikimate and Quinate Ester Intermediates in Phenylpropanoid Metabolism. J. Biol. Chem. 2003, 278, 95–103. [Google Scholar] [CrossRef]
- Magaña, A.A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic acids: chemistry, biosynthesis, occurrence, analytical challenges, and bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef]
- Vlot, A.C.; Dempsey, D.M.A.; Klessig, D.F. Salicylic acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, T.M.; Nesi, A.N.; Araújo, W.L.; Braun, H.-P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [PubMed]
- Isherwood, F.A.; Chen, Y.T.; Mapson, L.W. Synthesis of l-ascorbic acid in plants and animals. Biochem. J. 1954, 56, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Wang, J.; Liu, Q.; Zheng, Z.; Ouyang, J. [Production of mucic acid from pectin-derived D-galacturonic acid: a review]. . 2022, 38, 666–677. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Z.; Xia, Y.; Kai, G.; Chen, W.; Tang, K. Metabolic Engineering of Plant L-Ascorbic Acid Biosynthesis: Recent Trends and Applications. Crit. Rev. Biotechnol. 2007, 27, 173–182. [Google Scholar] [CrossRef]
- Mahatma, M.K.; Thawait, L.K.; Jadon, K.S.; Thirumalaisamy, P.P.; Bishi, S.K.; Rathod, K.J.; Verma, A.; Kumar, N.; Golakiya, B.A. Metabolic profiling for dissection of late leaf spot disease resistance mechanism in groundnut. Physiol. Mol. Biol. Plants 2021, 27, 1027–1041. [Google Scholar] [CrossRef]
- Mierziak, J.; Wojtasik, W.; Kulma, A.; Dziadas, M.; Kostyn, K.; Dymińska, L.; Hanuza, J.; Żuk, M.; Szopa, J. 3-Hydroxybutyrate Is Active Compound in Flax that Upregulates Genes Involved in DNA Methylation. Int. J. Mol. Sci. 2020, 21, 2887. [Google Scholar] [CrossRef]
- Rojas-Morales, P.; Pedraza-Chaverri, J.; Tapia, E. Ketone bodies, stress response, and redox homeostasis. Redox Biol. 2019, 29, 101395. [Google Scholar] [CrossRef]
- Xie, Z.; Zhang, D.; Chung, D.; Tang, Z.; Huang, H.; Dai, L.; Qi, S.; Li, J.; Colak, G.; Chen, Y.; et al. Metabolic Regulation of Gene Expression by Histone Lysine β-Hydroxybutyrylation. Mol. Cell 2016, 62, 194–206. [Google Scholar] [CrossRef]
- Ji, X. , Van den Ende, W. , Van Laere, A., Cheng, S., Bennett, J. Structure, evolution and expression of the two invertase families of rice. J. Mol. Evol. 2005, 60, 615–634. [Google Scholar]
- Salvi, P.; Varshney, V.; Majee, M. Raffinose family oligosaccharides (RFOs): role in seed vigor and longevity. Biosci. Rep. 2022, 42. [Google Scholar] [CrossRef] [PubMed]
- Keller, F. , Pharr, D.M., 1996. “Metabolism of carbohydrates in sinks and sources: galactosyl-sucrose oligosaccharides,” in Photoassimilate Distribution in Plants and Crops: Source–Sink Relationships, eds E. Zamski and A. A. Schaffer (New York: Marcel Dekker), 157–183.
- dos Santos, R.; Vergauwen, R.; Pacolet, P.; Lescrinier, E.; Ende, W.V.D. Manninotriose is a major carbohydrate in red deadnettle (Lamium purpureum, Lamiaceae). Ann. Bot. 2012, 111, 385–393. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Y.; Liu, Y.; Li, X.; Hao, G.; Han, Q.; Dirk, L.M.A.; Downie, A.B.; Ruan, Y.-L.; Wang, J.; et al. Raffinose synthase enhances drought tolerance through raffinose synthesis or galactinol hydrolysis in maize and Arabidopsis plants. J. Biol. Chem. 2020, 295, 8064–8077. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.J. Trehalose 6-phosphate. Current Opinion in Plant Biology 2007, 10, 303–309. [Google Scholar] [CrossRef]
- Almeida, A.M.; Villalobos, E.; Araújo, S.S.; Leyman, B.; Van Dijck, P.; Alfaro-Cardoso, L.; Fevereiro, P.S.; Torné, J.M.; Santos, D.M. Transformation of tobacco with an Arabidopsis thaliana gene involved in trehalose biosynthesis increases tolerance to several abiotic stresses. Euphytica 2005, 146, 165–176. [Google Scholar] [CrossRef]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and Raffinose Constitute a Novel Function to Protect Plants from Oxidative Damage. Plant Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef]
- Van den Ende, W. Multifunctional fructans and raffinose family oligosaccharides. Front. Plant Sci. 2013, 4, 247. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, Y.; Zhang, Y.; Dong, Y.; Liu, Y.; Liu, L.; Wan, S.; He, J.; Yu, Y. Accumulation of Galactinol and ABA Is Involved in Exogenous EBR-Induced Drought Tolerance in Tea Plants. J. Agric. Food Chem. 2022, 70, 13391–13403. [Google Scholar] [CrossRef]
- Barclay, K.D.; McKersie, B.D. Peroxidation reactions in plant membranes: Effects of free fatty acids. Lipids 1994, 29, 877–882. [Google Scholar] [CrossRef]
- Brendel, K.; Corredor, C.F.; Bressler, R. The effect of 4-pentenoic acid on fatty acid oxidation. Biochem. Biophys. Res. Commun. 1969, 34, 340–347. [Google Scholar] [CrossRef]
- Schulz, H. Inhibitors of fatty acid oxidation. Life Sci. 1987, 40, 1443–1449. [Google Scholar] [CrossRef] [PubMed]
- Mudd, S.H.; Datko, A.H. Synthesis of Ethanolamine and Its Regulation in Lemna paucicostata. Plant Physiol. 1989, 91, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Barbier, F.; Péron, T.; Lecerf, M.; Perez-Garcia, M.-D.; Barrière, Q.; Rolčík, J.; Boutet-Mercey, S.; Citerne, S.; Lemoine, R.; Porcheron, B.; et al. Sucrose is an early modulator of the key hormonal mechanisms controlling bud outgrowth in Rosa hybrida. J. Exp. Bot. 2015, 66, 2569–2582. [Google Scholar] [CrossRef] [PubMed]
- Mason, M.G.; Ross, J.J.; Babst, B.A.; Wienclaw, B.N.; Beveridge, C.A. Sugar demand, not auxin, is the initial regulator of apical dominance. Proc. Natl. Acad. Sci. USA 2014, 111, 6092–6097. [Google Scholar] [CrossRef]
- Kirma, M.; Araújo, W.L.; Fernie, A.R.; Galili, G. The multifaceted role of aspartate-family amino acids in plant metabolism. J. Exp. Bot. 2012, 63, 4995–5001. [Google Scholar] [CrossRef]
- Gómez, L.D. , Baud, S. , Gilday, A., Li, Y., Graham, I. Delayed embryo development in the Arabidopsis trehalose 6-phosphate synthase 1 mutant is associated with altered cell wall structure, decreased cell division and starch accumulation. Plant J. 2006, 46, 69–84. [Google Scholar]
- Schluepmann, H.; Pellny, T.; van Dijken, A.; Smeekens, S.; Paul, M. Trehalose 6-phosphate is indispensable for carbohydrate utilization and growth in Arabidopsis thaliana. Proc. Natl. Acad. Sci. 2003, 100, 6849–6854. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Asghari, M.; Khorsandi, O.; Mohayeji, M. Alleviation of postharvest chilling injury of tomato fruit by salicylic acid treatment. J. Food Sci. Technol. 2012, 51, 2815–2820. [Google Scholar] [CrossRef]
- Lapenna, D.; Ciofani, G.; Pierdomenico, S.D.; Neri, M.; Cuccurullo, C.; Giamberardino, M.A.; Cuccurullo, F. Inhibitory activity of salicylic acid on lipoxygenase-dependent lipid peroxidation. Biochim. et Biophys. Acta (BBA) - Gen. Subj. 2009, 1790, 25–30. [Google Scholar] [CrossRef]
- Wang, Z.; Rong, D.; Chen, D.; Xiao, Y.; Liu, R.; Wu, S.; Yamamuro, C. Salicylic acid promotes quiescent center cell division through ROS accumulation and down-regulation of PLT1, PLT2, and WOX5. J. Integr. Plant Biol. 2020, 63, 583–596. [Google Scholar] [CrossRef]
- Heil, M.; Ton, J. Long-distance signalling in plant defence. Trends Plant Sci. 2008, 13, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Shah, J. Plants under attack: Systemic signals in defense. Curr. Opin. Plant Biol. 2009, 12, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Caarls, L.; Pieterse, C.M.J.; Van Wees, S.C.M. How salicylic acid takes transcriptional control over jasmonic acid signaling. Front. Plant Sci. 2015, 6, 170–170. [Google Scholar] [CrossRef] [PubMed]
- Thaler, J.S.; Humphrey, P.T.; Whiteman, N.K. Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Sci. 2012, 17, 260–270. [Google Scholar] [CrossRef]
- Chao, W.S.; Doğramaci, M.; Horvath, D.P.; Anderson, J.V.; Foley, M.E. Phytohormone balance and stress-related cellular responses are involved in the transition from bud to shoot growth in leafy spurge. BMC Plant Biol. 2016, 16, 1–21. [Google Scholar] [CrossRef]
- Howe, G.T.; Horvath, D.P.; Dharmawardhana, P.; Priest, H.D.; Mockler, T.C.; Strauss, S.H. Extensive Transcriptome Changes During Natural Onset and Release of Vegetative Bud Dormancy in Populus. Front. Plant Sci. 2015, 6, 989–989. [Google Scholar] [CrossRef]
- Nunes, C. , Schluepmann, H. , Delatte, T.L., Wingler, A., Silva, A.B., Fevereio, P.S., Jansen, M., Fiorani, F., Wiese-Klinkenberg, A., Paul, M. Regulation of growth by the trehalose pathway: Relationship to temperature and sucrose. Plant Signal. Behav. 2013, 8, 12. [Google Scholar]
- O’hara, L.E.; Paul, M.J.; Wingler, A. How Do Sugars Regulate Plant Growth and Development? New Insight into the Role of Trehalose-6-Phosphate. Mol. Plant 2013, 6, 261–274. [Google Scholar] [CrossRef]
- Smeekens, S.; Ma, J.; Hanson, J.; Rolland, F. Sugar signals and molecular networks controlling plant growth. Curr. Opin. Plant Biol. 2010, 13, 273–278. [Google Scholar] [CrossRef]
- Zhang, Y.; Primavesi, L.F.; Jhurreea, D.; Andralojc, P.J.; Mitchell, R.A.; Powers, S.J.; Schluepmann, H.; Delatte, T.; Wingler, A.; Paul, M.J. Inhibition of SNF1-Related Protein Kinase1 Activity and Regulation of Metabolic Pathways by Trehalose-6-Phosphate. Plant Physiol. 2009, 149, 1860–1871. [Google Scholar] [CrossRef]
- Baena-González, E.; Rolland, F.; Thevelein, J.M.; Sheen, J. A central integrator of transcription networks in plant stress and energy signalling. Nature 2007, 448, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Radchuk, R.; Radchuk, V.; Weschke, W.; Borisjuk, L.; Weber, H. Repressing the Expression of the SUCROSE NONFERMENTING-1-RELATED PROTEIN KINASE Gene in Pea Embryo Causes Pleiotropic Defects of Maturation Similar to an Abscisic Acid-Insensitive Phenotype. Plant Physiol. 2005, 140, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Filipe, O.; De Vleesschauwer, D.; Haeck, A.; Demeestere, K.; Höfte, M. The energy sensor OsSnRK1a confers broad-spectrum disease resistance in rice. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; De Vleesschauwer, D.; Sharma, M.K.; Ronald, P.C. Recent Advances in Dissecting Stress-Regulatory Crosstalk in Rice. Mol. Plant 2013, 6, 250–260. [Google Scholar] [CrossRef]
- Janda, T.; Szalai, G.; Pál, M. Salicylic Acid Signalling in Plants. Int. J. Mol. Sci. 2020, 21, 2655. [Google Scholar] [CrossRef]
- Kianersi, F.; Abdollahi, M.R.; Mirzaie-Asl, A.; Dastan, D.; Rasheed, F. Identification and tissue-specific expression of rutin biosynthetic pathway genes in Capparis spinosa elicited with salicylic acid and methyl jasmonate. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Tao, S.; Sun, L.; Ma, C.; Li, L.; Li, G.; Hao, L. Reducing basal salicylic acid enhances Arabidopsis tolerance to lead or cadmium. Plant Soil 2013, 372, 309–318. [Google Scholar] [CrossRef]
- Jander, G.; Joshi, V. Aspartate-Derived Amino Acid Biosynthesis inArabidopsis thaliana. Arab. Book 2009, 7, e0121. [Google Scholar] [CrossRef]
- Joshi, V. , Jander, G. Arabidopsis methionine g-lyase is regulated according to isoleucine biosynthesis needs but plays a subordinate role to threonine deaminase. Plant Physiology 2009, 151, 367–378. [Google Scholar]
- Lange, B.M. , Ghassemian, M. Comprehensive post-genomic data analysis approaches integrating biochemical pathwaymaps. Phytochemistry 2005, 66, 413–451. [Google Scholar]
- Arcadi, A.; Morlacci, V.; Palombi, L. Synthesis of Nitrogen-Containing Heterocyclic Scaffolds through Sequential Reactions of Aminoalkynes with Carbonyls. Molecules 2023, 28, 4725. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive Oxygen Species in Plant Signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef]
- Apel, K. , Hirt, H., 2004. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 55, 373–399.
- Chapman, J.M.; Muhlemann, J.K.; Gayomba, S.R.; Muday, G.K. RBOH-Dependent ROS Synthesis and ROS Scavenging by Plant Specialized Metabolites To Modulate Plant Development and Stress Responses. Chem. Res. Toxicol. 2019, 32, 370–396. [Google Scholar] [CrossRef] [PubMed]
- Hernández, I.; Chacón, O.; Rodriguez, R.; Portieles, R.; López, Y.; Pujol, M.; Borrás-Hidalgo, O. Black shank resistant tobacco by silencing of glutathione S-transferase. Biochem. Biophys. Res. Commun. 2009, 387, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Kandaswami, C. , Middleton, E., 1994. Free radical scavenging and antiocidant activity of plant flavonoids. Free Radicals in Diagnostic Medicine, Edited by D. Annstrong, Plenum Press, New York.
- Løvdal, T.; Olsen, K.M.; Slimestad, R.; Verheul, M.; Lillo, C. Synergetic effects of nitrogen depletion, temperature, and light on the content of phenolic compounds and gene expression in leaves of tomato. Phytochemistry 2010, 71, 605–613. [Google Scholar] [CrossRef]
- Tsukagoshi, H. Control of root growth and development by reactive oxygen species. Curr. Opin. Plant Biol. 2016, 29, 57–63. [Google Scholar] [CrossRef]
- Yang, S.; Yu, Q.; Zhang, Y.; Jia, Y.; Wan, S.; Kong, X.; Ding, Z. ROS: The Fine-Tuner of Plant Stem Cell Fate. Trends Plant Sci. 2018, 23, 850–853. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Csepregi, K.; Hideg, É. Phenolic Compound Diversity Explored in the Context of Photo-Oxidative Stress Protection. Phytochem. Anal. 2017, 29, 129–136. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Saslowsky, D.E.; Warek, U.; Winkel, B.S. Nuclear Localization of Flavonoid Enzymes in Arabidopsis. J. Biol. Chem. 2005, 280, 23735–23740. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.M.; Hechler, P.J.; Muday, G.K. Ethylene-Induced Flavonol Accumulation in Guard Cells Suppresses Reactive Oxygen Species and Moderates Stomatal Aperture. Plant Physiol. 2014, 164, 1707–1717. [Google Scholar] [CrossRef]
- Liang, N.; Dupuis, J.H.; Yada, R.Y.; Kitts, D.D. Chlorogenic acid isomers directly interact with Keap 1-Nrf2 signaling in Caco-2 cells. Mol. Cell. Biochem. 2019, 457, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A. , Xie, D. -Y., Sharma, S.B. Proanthocyanidins—a final frontier in flavonoid research? New Phytologist 2005, 165, 9–28. [Google Scholar]
- Iqbal, S.; Andrabi, S.M.H.; Riaz, A.; Durrani, A.Z.; Ahmad, N. Trehalose improves semen antioxidant enzymes activity, post-thaw quality, and fertility in Nili Ravi buffaloes ( Bubalus bubalis ). Theriogenology 2016, 85, 954–959. [Google Scholar] [CrossRef]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Motta, M.; Marcone, M.; Baptista, J. Variability of antioxidant properties, catechins, caffeine, L-theanine and other amino acids in different plant parts of Azorean Camellia sinensis. Curr. Res. Food Sci. 2020, 3, 227–234. [Google Scholar] [CrossRef]
- Chen, C.; Dickman, M.B. From The Cover: Proline suppresses apoptosis in the fungal pathogen Colletotrichum trifolii. Proc. Natl. Acad. Sci. USA 2005, 102, 3459–3464. [Google Scholar] [CrossRef]
- Azam, S.; Hadi, N.; Khan, N.U.; Hadi, S.M. Antioxidant and prooxidant properties of caffeine, theobromine and xanthine. Med. Sci. Monit. 2003, 9, 325–330. [Google Scholar]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional Regulation of ROS Controls Transition from Proliferation to Differentiation in the Root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Thelen, J.J. Plastid Uridine Salvage Activity Is Required for Photoassimilate Allocation and Partitioning in Arabidopsis. Plant Cell 2011, 23, 2991–3006. [Google Scholar] [CrossRef]
- Yue, W.; Sun, W.; Rao, R.S.P.; Ye, N.; Yang, Z.; Chen, M. Non-targeted metabolomics reveals distinct chemical compositions among different grades of Bai Mudan white tea. Food Chem. 2019, 277, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Tautenhahn, R.; Cho, K.; Uritboonthai, W.; Zhu, Z.; Patti, G.J.; Siuzdak, G. An accelerated workflow for untargeted metabolomics using the METLIN database. Nat. Biotechnol. 2012, 30, 826–828. [Google Scholar] [CrossRef] [PubMed]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: a public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef]
- Wishart, D.S. , Jewison, T. , Guo, A.C., Wilson, M., Knox, C., Liu, Y., Djoumbou, Y., Mandal, R., Aziat, F., Dong, E., Bouatra, S., Sinelnikov, I., Arndt, D., Xia, J., Liu, P., Yallou, F., Bjorndahl, T., Perez-Pineiro, R., Eisner, R., Allen, F., Neveu, V., Greiner, R., Scalbert, A. HMDB 3.0–the human metabolome database in 2013. Nucleic Acids Res. 2013, 41, D801–D807. [Google Scholar]
- Afendi, F.M.; Okada, T.; Yamazaki, M.; Hirai-Morita, A.; Nakamura, Y.; Nakamura, K.; Ikeda, S.; Takahashi, H.; Amin, A.U.; Darusman, L.K.; et al. KNApSAcK Family Databases: Integrated Metabolite–Plant Species Databases for Multifaceted Plant Research. Plant Cell Physiol. 2011, 53, e1–e1. [Google Scholar] [CrossRef]
- Sawada, Y.; Nakabayashi, R.; Yamada, Y.; Suzuki, M.; Sato, M.; Sakata, A.; Akiyama, K.; Sakurai, T.; Matsuda, F.; Aoki, T.; et al. RIKEN tandem mass spectral database (ReSpect) for phytochemicals: A plant-specific MS/MS-based data resource and database. Phytochemistry 2012, 82, 38–45. [Google Scholar] [CrossRef]
Figure 1.
Photography of Camellia sinensis cv Huangdan. (A) Camellia sinensis cv Huangdan grown in the tea garden, bar = 3 mm; (B) a close-up view showing the arrangement of the auxiliary shoot bud, the auxiliary floral buds and the lateral leaf located at the same node, the stem above the node was removed, bar = 2 mm.
Figure 1.
Photography of Camellia sinensis cv Huangdan. (A) Camellia sinensis cv Huangdan grown in the tea garden, bar = 3 mm; (B) a close-up view showing the arrangement of the auxiliary shoot bud, the auxiliary floral buds and the lateral leaf located at the same node, the stem above the node was removed, bar = 2 mm.

Figure 2.
Photograph of the axillary shoot bud. HD_0, before axillary floral bud removal; HD-1, one day after remove the axillary floral buds; HD_3, three days after remove the axillary floral buds.
Figure 2.
Photograph of the axillary shoot bud. HD_0, before axillary floral bud removal; HD-1, one day after remove the axillary floral buds; HD_3, three days after remove the axillary floral buds.

Figure 3.
Partial least square-discriminant analysis and Veno diagram analysis of metabolome dataset obtained on LC-MS platform. PLS-DA results from the negative ion mode (A) and the positive ion mode (B) , respectively; Veno diagram results from the negative ion mode (C) and the positive ion mode (D), respectively. HD_0, axillary shoot bud samples before remove the axillary floral buds; HD_1 and HD-3, axillary shoot bud samples after remove the axillary floral buds for 1 and 3 days, respectively.
Figure 3.
Partial least square-discriminant analysis and Veno diagram analysis of metabolome dataset obtained on LC-MS platform. PLS-DA results from the negative ion mode (A) and the positive ion mode (B) , respectively; Veno diagram results from the negative ion mode (C) and the positive ion mode (D), respectively. HD_0, axillary shoot bud samples before remove the axillary floral buds; HD_1 and HD-3, axillary shoot bud samples after remove the axillary floral buds for 1 and 3 days, respectively.
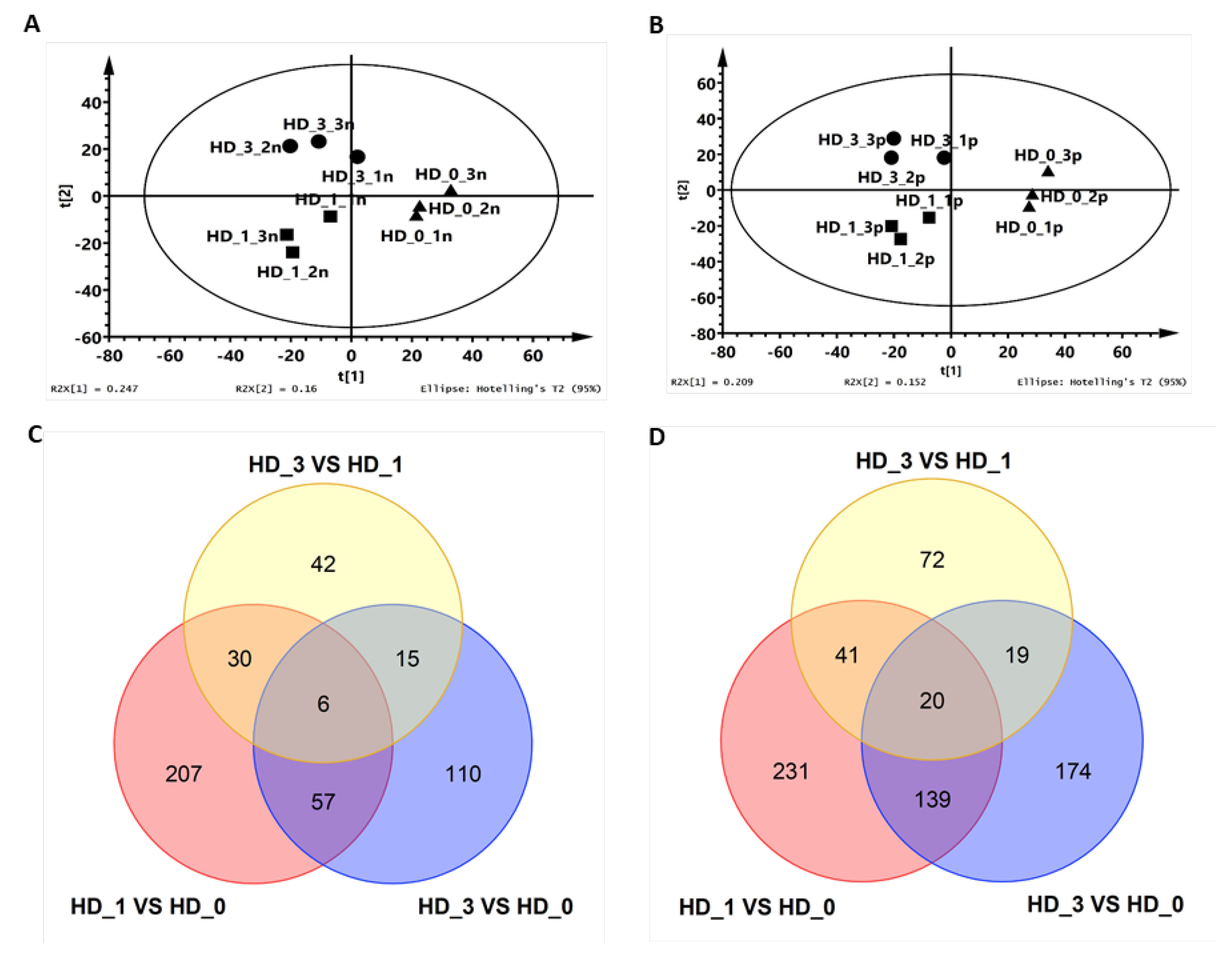
Figure 4.
Heat-map of phenolic acids (A) and flavanols (B) before and after 1- and 3- days of the axillary floral bud removal. HD_0: the samples of the axillary shoot buds harvested before remove the axillary floral buds; HD_1 and HD_3: the samples of the axillary shoot buds harvested after 1- and 3- days of the axillary floral bud removal. .
Figure 4.
Heat-map of phenolic acids (A) and flavanols (B) before and after 1- and 3- days of the axillary floral bud removal. HD_0: the samples of the axillary shoot buds harvested before remove the axillary floral buds; HD_1 and HD_3: the samples of the axillary shoot buds harvested after 1- and 3- days of the axillary floral bud removal. .

Figure 5.
Heat-map of flavone and flavonol glycosides (A) and proanthocyanidins (B) before and after 1- and 3- days of the axillary floral bud removal. HD_0: the samples of the axillary shoot buds harvested before remove the axillary floral buds; HD_1 and HD_3: the samples of the axillary shoot buds harvested after 1- and 3- days of the axillary floral bud removal, respectively. .
Figure 5.
Heat-map of flavone and flavonol glycosides (A) and proanthocyanidins (B) before and after 1- and 3- days of the axillary floral bud removal. HD_0: the samples of the axillary shoot buds harvested before remove the axillary floral buds; HD_1 and HD_3: the samples of the axillary shoot buds harvested after 1- and 3- days of the axillary floral bud removal, respectively. .
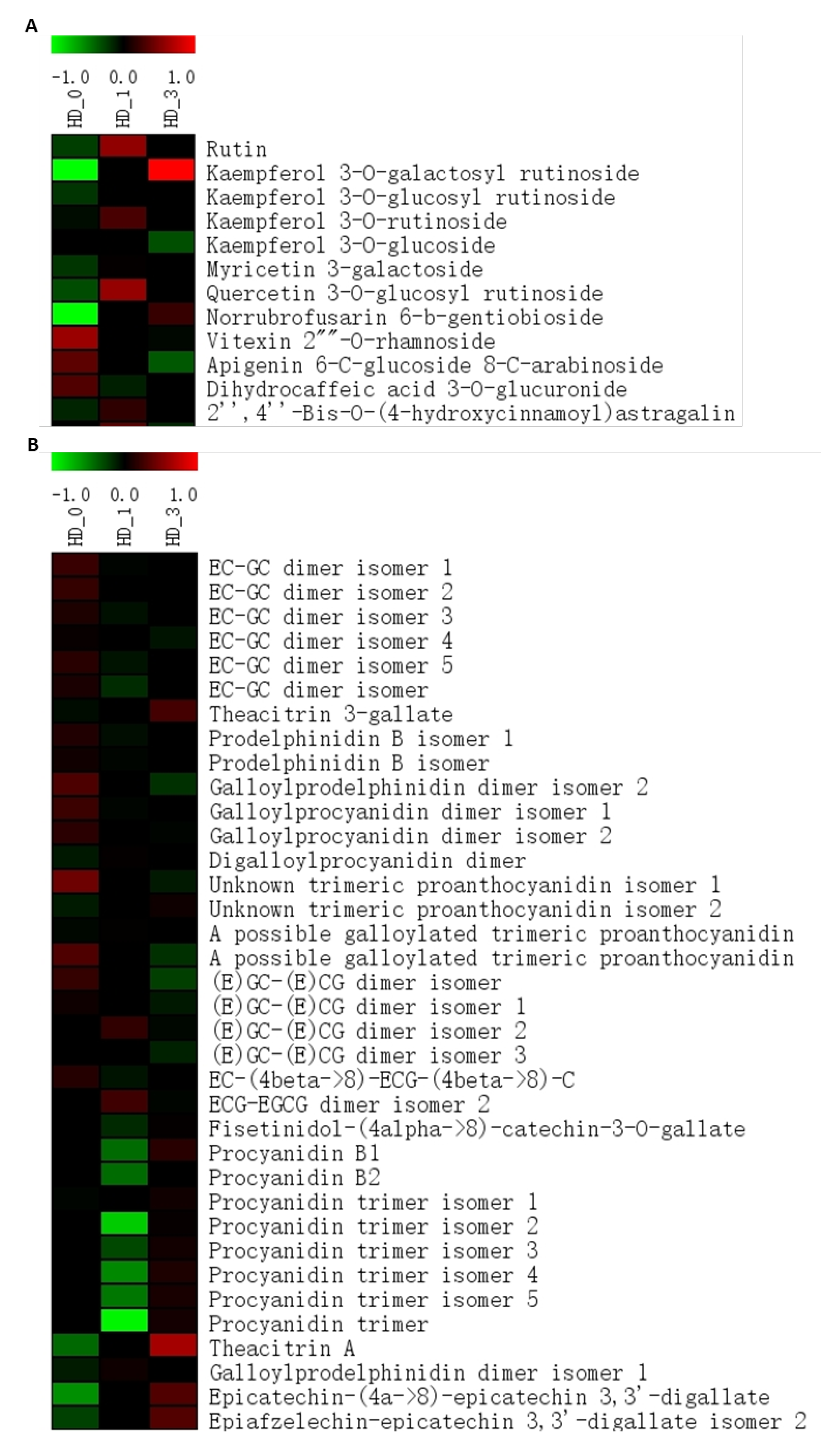
Figure 6.
Heat-map of amino acids (A) and organic acids (B) before and after 1- and 3- days of the axillary floral bud removal. HD_0: the samples of the axillary shoot buds harvested before remove the axillary floral buds; HD_1 and HD_3: the samples of the axillary shoot buds harvested after 1- and 3- days of the axillary floral bud removal, respectively. .
Figure 6.
Heat-map of amino acids (A) and organic acids (B) before and after 1- and 3- days of the axillary floral bud removal. HD_0: the samples of the axillary shoot buds harvested before remove the axillary floral buds; HD_1 and HD_3: the samples of the axillary shoot buds harvested after 1- and 3- days of the axillary floral bud removal, respectively. .

Figure 7.
Heat-map of sugars and sugar alcohols (A) and fatty acids and sterols (B) before and after 1- and 3- days of the axillary floral bud removal. HD_0: the samples of the axillary shoot buds harvested before remove the axillary floral buds; HD_1 and HD_3: the samples of the axillary shoot buds harvested after 1- and 3- days of the axillary floral bud removal, respectively.
Figure 7.
Heat-map of sugars and sugar alcohols (A) and fatty acids and sterols (B) before and after 1- and 3- days of the axillary floral bud removal. HD_0: the samples of the axillary shoot buds harvested before remove the axillary floral buds; HD_1 and HD_3: the samples of the axillary shoot buds harvested after 1- and 3- days of the axillary floral bud removal, respectively.
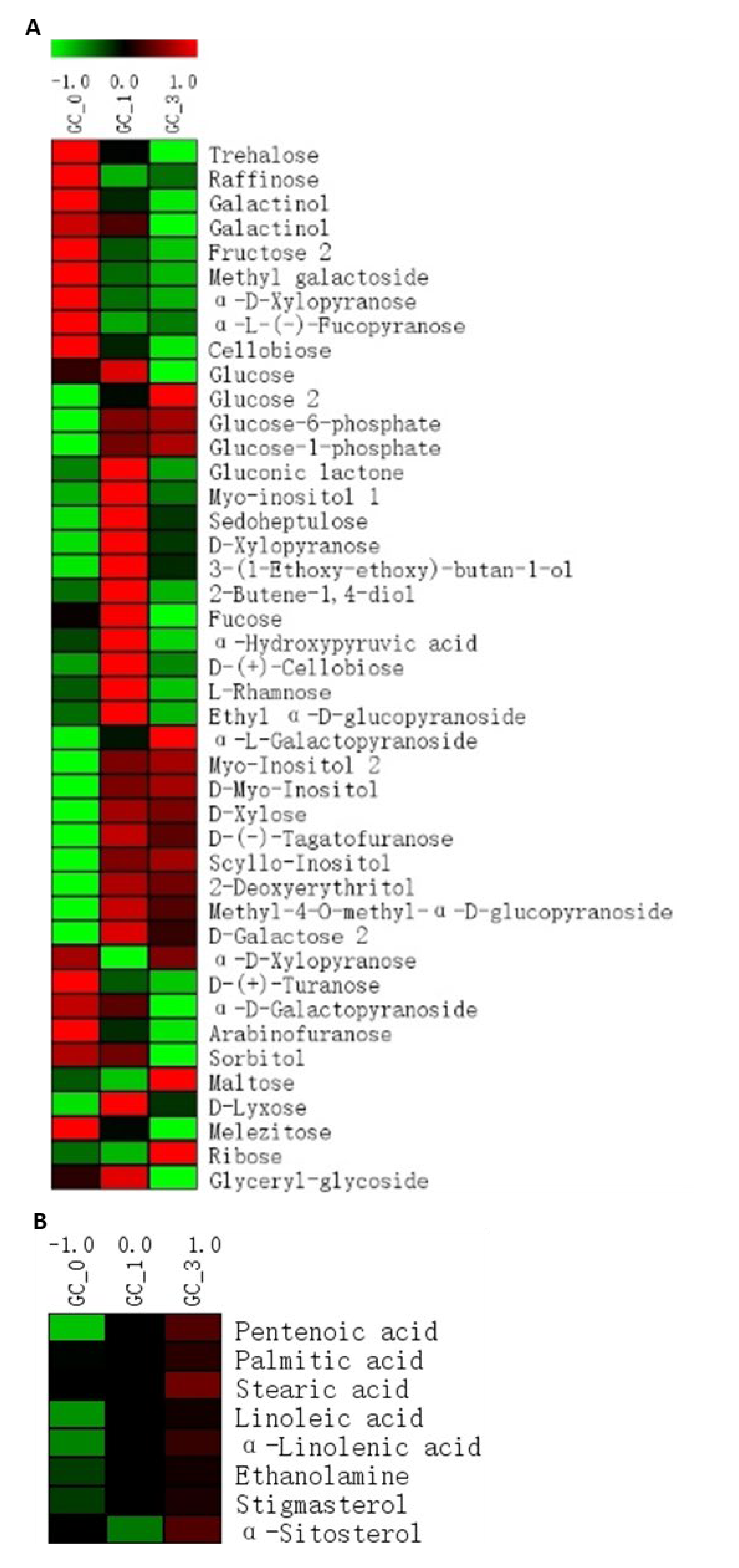
Figure 8.
A proposed model to explain carbon starvation signaling transduction pathway leading to paradormancy set and release in the axillary shoot buds of Camellia sinensis cv Huangdan. G6P: glucose-6-phosphate; G1P: glucose-1-phosphate; Tre6P: trehalose-6-phosphate; Tre: trehalose; GA: gibberellin; BR: brassinosteroid; SA: salicylic acid; JA: jasmonate; APX: ascorbate peroxidase; CAT, catalase; ROS: reactive oxygen species.
Figure 8.
A proposed model to explain carbon starvation signaling transduction pathway leading to paradormancy set and release in the axillary shoot buds of Camellia sinensis cv Huangdan. G6P: glucose-6-phosphate; G1P: glucose-1-phosphate; Tre6P: trehalose-6-phosphate; Tre: trehalose; GA: gibberellin; BR: brassinosteroid; SA: salicylic acid; JA: jasmonate; APX: ascorbate peroxidase; CAT, catalase; ROS: reactive oxygen species.
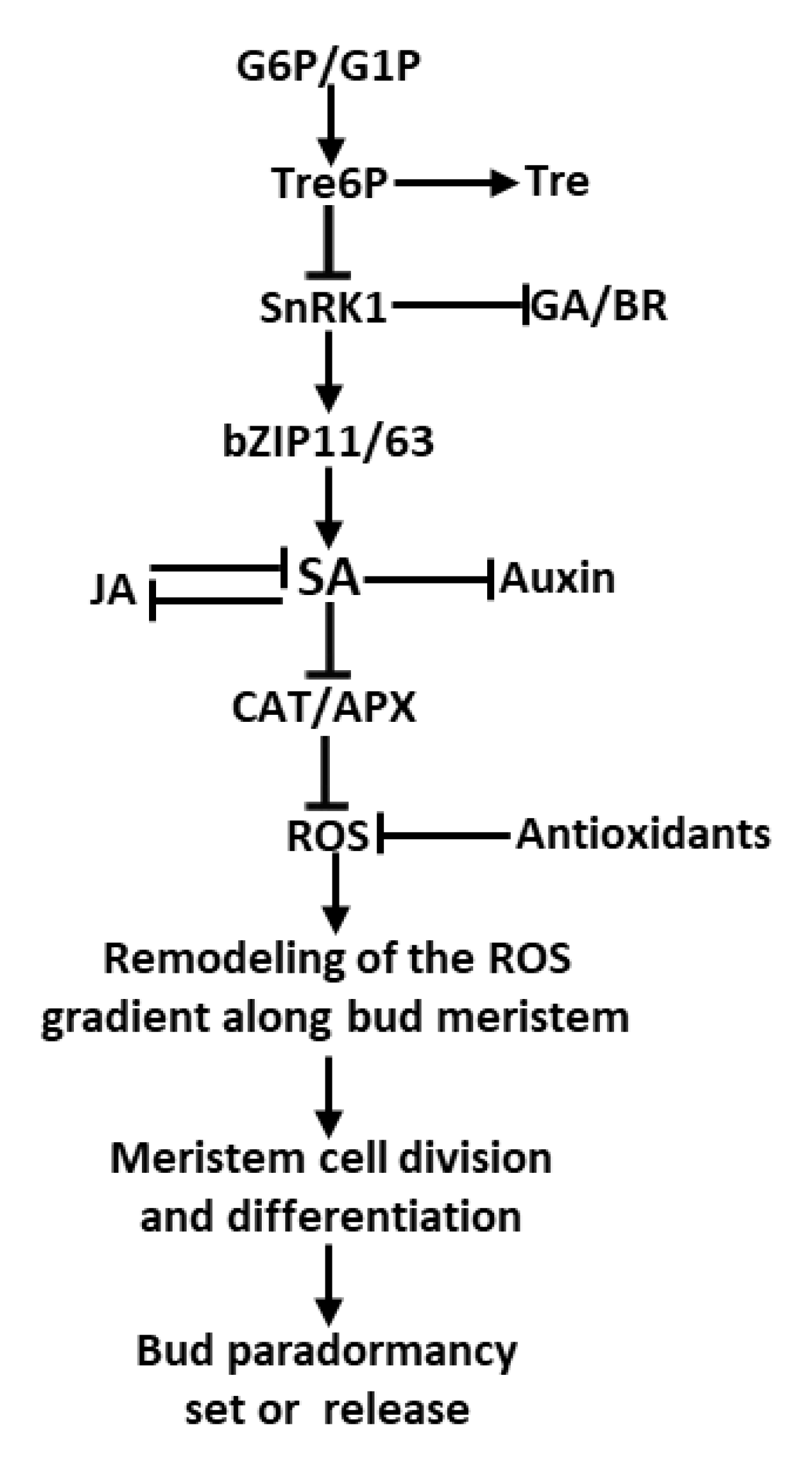
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



