
Submitted:
06 November 2024
Posted:
06 November 2024
You are already at the latest version
Abstract
Background:
We hypothesized that the maximum residual concentrations of trimethoprim allowed in food (Acceptable Daily Intake – ADI) could select for de novo trimethoprim resistance in Escherichia coli in vivo.
Methods:
We developed models of chronic Escherichia coli infection in Galleria mellonella and exposed them to sub-ADI doses of trimethoprim via single-dosing regimens. The emergence of trimethoprim resistance was assessed via isolation of the target bacteria in selective agar plates. Species identity was confirmed using MALDI-TOF mass spectrometry, and the antibiotic susceptibility of E. coli to trimethoprim was evaluated using the E-test to determine the minimum inhibitory concentration (MIC).
Results:
Exposure to as low as one-tenth of the ADI dose of the single-dosing regimen of trimethoprim resulted in the selection of trimethoprim resistance in E. coli.
Conclusion:
We found that trimethoprim doses ten-fold lower than those classified as acceptable for daily intake could induce resistance to trimethoprim in E. coli. These results suggest that it would be prudent to include the induction of antimicrobial resistance as a significant criterion for determining ADIs in food.
Keywords:
Escherichia coli
; antimicrobial consumption
; minimum selection concentration
; AMR
; trimethoprim
; MSC
; resistance
Introduction
In recent years, several studies have established that subinhibitory concentrations of antimicrobials can induce de novo antimicrobial resistance (AMR) [1,2,3]. The minimum selection concentration (MSC) is defined as the minimum concentration of an antimicrobial that can select for resistance [1,4]. MSC can be categorized into two types: MSCselect, which is the lowest concentration of an antimicrobial that selects for a resistant phenotype over a susceptible one [5] and MSCde novo, defined as the minimum concentration of an antimicrobial that can induce de novo resistance. Antimicrobials are frequently detected in the environment at sub-MIC concentrations, and numerous studies have found that these low concentrations can select for AMR [4,6,7,8]. For example, Gullberg et al. found that the ciprofloxacin MSC for Escherichia coli can be up to 260 times lower than the MIC of the susceptible strain (MICsusc) [4]. Another study performed by Gullberg et al. used an E. coli plasmid that conferred resistance to a range of antibiotics, including tetracycline and trimethoprim, and found that the multidrug-resistant plasmid was selected for at concentrations far below the MICsusc [1].
An emerging body of research has suggested that numerous foodstuffs contain low concentrations of antimicrobials that are above the MSCs [8,9]. More recently, in vivo models have been developed to directly test if the concentrations of antimicrobials allowed in food could induce AMR [8]. These studies have assessed if the acceptable daily intake (ADI) of an antimicrobial can induce AMR. The ADI is defined by the Food and Agriculture Organization/World Health Organization (FAO/WHO) as “an estimate of the amount of a food additive in food or beverages expressed on a body weight (bw) basis that can be ingested daily over a lifetime without appreciable health risk to the consumer” [10]. Thus far, these studies have found that ciprofloxacin and erythromycin doses as low as 1/10th of the ADI can induce resistance in Klebsiella pneumoniae and Streptococcus pneumoniae in a Galleria mellonella model [8,11]. In the current study, we use the same model, G. mellonella, to assess if sub-ADI doses of trimethoprim can induce resistance to trimethoprim in E. coli. We chose trimethoprim as it is one of the five most commonly used antibiotics for food animals worldwide [12].
High concentrations of trimethoprim have been reported in untreated municipal wastewater systems in numerous studies, with levels between 0.17-8.8 µg/L in regions of South Africa [13] and Scandinavia [14] and concentrations as high as 28 µg/L in surface waters in Pakistan [15]. Furthermore, a study in the United Kingdom found that trimethoprim was the second most frequently detected antimicrobial in a range of animal-based food and drink products, with concentrations ranging from 55.2 - 461.7 µg/kg [16].
The European Medicines Agency (EMA) determines the acceptable daily ingestion (ADI) of a medicinal compound based on studies assessing microbiological and cellular toxicity thresholds [17,18,19,20,21,22]. For antimicrobials, ADIs are predominately derived from microbiological toxicity data, typically established by evaluating the MICs for common human bacterial commensals or pathobionts, such as E. coli, and estimating dose exposure levels in the human colon [17,18,19,22,23]. Notably, the potential for induction of- or selection for AMR is not directly included [22]. The ADIs serve as the basis for setting maximum residue limits (MRLs) that represent the maximum concentration of the compound allowed in food products based on the average consumption patterns of those food products [22,24]. According to the latest EMA reports, the ADI for trimethoprim is established at 4.2 μg/kg [25].
Trimethoprim is a synthetic antimicrobial mainly used to treat urinary tract infections (UTIs) [26]. It is also commonly used in human and veterinary medicine, frequently in combination with sulphonamide [25], to treat a range of other infections [27]. It is either bactericidal (in combination with sulphonamide) or bacteriostatic via inhibiting the folic acid synthesis pathway [28] in which the reduction of dihydrofolate to tetrahydrofolate is blocked, causing disordered nucleic acid synthesis [26].
We hypothesized that the EMA ADI dose of trimethoprim could induce resistance in vivo. We tested this hypothesis using a G. mellonella model of E. coli infection treated with peri-ADI doses of trimethoprim. We tested three doses of trimethoprim: ADI dose (6,2 μg/kg), a 10-fold higher dose and a 1/10th dose.
Materials and Methods
Bacterial Strains and Growth Conditions
The strain used in this study was the E. coli strain ATCC 25922, commonly used in quality control for antimicrobial susceptibility testing [29,30] and susceptible to trimethoprim with a MIC of 1µg/mL [31]. It belongs to serotype O6, biotype 1, and was initially isolated from a clinical sample in Seattle, Washington 1946 [29]. The assembled genome of this strain is 5.20 Mb, comprising two plasmids (48,488 and 24,185-bp, respectively) and a chromosome (5,130,767-bp) [29]. The MIC for trimethoprim was confirmed as 1µg/mL using an E-test (AB bioMerieux, France) prior to performing the main experiments.
Preparation of Live Microbial Inocula for Infection
E. coli strains were cultured from frozen stocks onto BDTM Columbia Agar supplemented with 5% Sheep Blood for ≤16 h at 37°C with 5% (v/v) CO2. Single colonies were selected and spread onto fresh agar plates that were incubated at 37°C with 5% (v/v) CO2 for 6 h. Suspension of E. coli was made with phosphate buffer saline (PBS) and then injected into the hemocoel of the G. mellonella larva. The dose of E. coli was optimized to allow the recovery of the bacteria up to 3 days post-inoculation, minimizing larval mortality (data not shown).
Injection of G. mellonella Larvae
The study included control groups of 10 larvae and experimental groups with 30 larvae per condition. Healthy, non-discoloured larvae in their last larval stage, weighing 250-450 mg, were selected and placed into sterile petri dishes in groups of 10 per petri dish. These larvae were incubated at 37°C with 5% (v/v) CO2 throughout the experiment.
In the experimental groups, each larva was injected with 30 µL of bacterial suspension into the last right pro-leg. After 10-20 minutes, the larvae were injected in the last left pro-leg with various doses of trimethoprim. Injections were administered using 0.3mL U-100 insulin syringes (BD Micro-Fine), with one syringe and needle used per petri dish (10 larvae per petri dish).
One control group followed the same procedure as the experimental groups, receiving E. coli inoculation in the last right pro-leg followed by 10 µL/larva of phosphate-buffered saline (PBS) in the last left pro-leg (positive control). The other control group received only 10 µL/larva of PBS in the last left pro-leg (negative control).
Concentration of Trimethoprim Injected
As previously mentioned, the EMA ADI for trimethoprim is 4.2 µg/kg bw. Based on this, the equivalent dose of trimethoprim for these experiments was calculated to be 1.57 ng, using the average weight of the G. mellonella larvae (380 mg). The doses tested include 15.7 ng (10xADI), 1.57 ng (ADI) and 0.157 ng (0.1xADI) per larva.
Upon completion of each experiment, both surviving and dead G. mellonella larvae were kept at −80°C overnight to sedate and euthanize them. Following this, the larvae were autoclaved at 121°C for 15 min and discarded.
Retrieval of E. coli from G. mellonella
One to 5 larvae from each group of 10 were randomly selected at 24 and 48 hours after the injection for the retrieval of E. coli. Half of the surviving larvae were used at 24 hours, and the remaining live larvae were used at 48 hours. The selected larvae were then placed in -80°C refrigeration for 60 seconds until no movement could be observed. They were then put into a petri dish for an incision to be made between two segments close to the larval tail using a scalpel. The haemolymph was then squeezed into 1.8ml centrifuge tubes containing 50 µl PBS, after which the tubes were vortexed. E. coli from the G. mellonella haemolymph extraction were cultured by plating equal volumes of the extraction onto MacConkey agar with and without trimethoprim. The plates containing trimethoprim were prepared with a concentration of three times their MIC (3µg /mL trimethoprim).
MacConkey plates were incubated at 37°C with 5% (v/v) CO2 for 24 hours, and the number of E. coli colonies that appear dark blue to violet was counted. Growth was assessed for up to 48 hours post-haemolymph extraction. A subset of colonies from the antibiotic plates and a random selection of colonies from the non-antibiotic plates were selected and sub-cultured on MacConkey plates without antibiotic for species identification by MALDI-TOF-MS and for determining the trimethoprim MIC by E-test (AB bioMerieux, France). The E-tests for MIC determination of trimethoprim were performed on Mueller-Hinton agar (MHA) plates and incubated for 18-20h at 37°C with a 5% (v/v) CO2. The EUCAST guidelines were followed in defining the trimethoprim resistance as >2 µg/ml for E. coli (https://mic.eucast.org/search/). The E-test strips were placed on a plate inoculated with 0.5 McFarland concentration of E. coli isolates and read at 100% inhibition of bacterial growth at 18-20h.
For the stability experiment, trimethoprim-resistant isolates were passaged daily for five consecutive days on 5% Sheep Blood BDTM Columbia Agar plate, and MIC was assessed post-passage using E-tests in order to determine if the observed resistance phenotype was stable over time.
MALDI-TOF MS Species Identification
Species identification of the isolates were carried out using Matrix-Assisted Laser Desorption/Ionization-Time-of-Flight mass spectrometry (MALDI-TOF MS) on a MALDI Biotyper® Sirius IVD system using the MBT Compass IVD software and library (Bruker Daltonics, Bremen, Germany). This was done by spreading each bacterial isolate onto a polished steel target plate, covering it with 1 µL of α-cyano-4-hydroxycinnamic acid (CHCA) matrix solution, drying it, and loading the target plate. The results of the identification were classified as either reliable or unreliable based on the recommended cut-off values of 1.7 and 2 for genus and species levels, respectively.
Results
An overview of the study methodology is provided in Figure 1.
Colonization
E. coli were successfully recovered on a selective agar plate for up to 2 days following inoculation (Figure 2). Species identity were confirmed through MADLI-TOF MS (Table 1). Each haemolymph extraction was plated in triplicate for each condition across all experiments performed, with 50 µl of haemolymph plated onto two MacConkey agar plates – one with and one without trimethoprim. E. coli was isolated on all non-trimethoprim plates following the 2 and 48-hour extraction.
Colony Emergence and Identification
No growth was observed on any of the trimethoprim plates after the initial 24 h incubation, but after an additional 24 h incubation, bright pink colonies appeared on all the plates except those from the positive and negative control groups. More specifically, no colonies emerged on the trimethoprim plates from the positive or negative control groups at 24 h or 48 h. The colonies from the trimethoprim plates were all sub-cultured onto MacConkey agar for MIC determination and species identification. All these colonies were identified as E. coli via MALDI-TOF (Table 1). The only exception was a single colony from the 10x ADI trimethoprim group, identified as Escherichia marmotae, which is phenotypically close to E. coli (Table 1) [32].
Two of 6 colonies (2/6) from the 1xADI group and both (2/2) colonies from the 0.1xADI group from the 24 h trimethoprim plates along with two of 6 colonies (2/6) from the 0.1x ADI group from the 48 h trimethoprim plate and one from the (1/2) 0.1x ADI positive control samples from both 24 and 48 h were randomly selected for MIC determination via E-test. Elevated trimethoprim MICs were observed for two colonies - 0.1xADI-48hr-TMP1.1 (16 µg/mL) and 0.1xADI-48hr-TMP2.1 (12 µg/mL) samples (Figure 3).
Stability of Trimethoprim-Resistant Isolates
Two trimethoprim-resistant isolates (0.1xADI-48hr-TMP1.1 and 0.1xADI-48hr-TMP2.1) along with one isolate from the 0.1x ADI control group were used to assess if the observed increase in MIC was a stable phenotypic change. Only isolates from the 0.1x ADI group were passaged, as this was the lowest concentration that induced resistance.
At day 5, the isolate (0.1xADI-48hr-TMP1.1) with the pre-passage trimethoprim MIC of 16 µg/mL displayed two ellipses in the post-passage E-test, with one reading at 1.5 µg/mL and the other at 12 µg/mL (Supplementary Figure 1). The second isolate (0.1xADI-48hr-TMP2.1) with a pre-passage MIC of 12 µg/mL retained this MIC post-passage. The MIC of the 0.1x ADI control group also remained unchanged at 1 µg/mL after passaging.
Discussion
In this study, the lowest dose that induces de novo resistance to trimethoprim in E. coli in the G. mellonella model was found to be 0.157 ng/mL. This concentration is 6,369-fold lower than the MIC and 10-fold lower than the EMA ADI for trimethoprim. Forty-eight hours after receiving a single dose of one-tenth of the ADI (0.1X ADI) of trimethoprim, resistant colonies emerged with an increase in MIC of up to 16-fold (MIC-16µg/mL). These findings suggest that single doses of subinhibitory concentrations of trimethoprim, even 10 times below the acceptable daily intake, can select for de novo resistance in vivo. These results are consistent with other MSC studies involving E. coli. For example, a study by Gullberg et al. determined the MSCdenovo for ciprofloxacin and tetracycline to be 0.1 ng/mL and 15 ng/mL [4]. Another study performed by Gullberg et al. investigated the selective effect of sub-MIC concentrations on multidrug-resistant plasmids and found the MSC of trimethoprim to be 1/6th (33 ng/mL) of the MIC of the susceptible strain (MICsusc.) [1]. The MSCdenovo was not assessed in this study. One possible explanation for the higher MSC observed in the plasmid study is the higher fitness cost of a resistance mechanism being carried on a plasmid rather than the chromosome [1]. In 2014, Gullberg et al. noted that moving the resistance genes from the plasmid to the chromosome reduces the fitness cost associated with resistance, which in turn reduced the MSC for the tetracycline, trimethoprim and erythromycin by 2- to 15-fold [1]. Our study’s MSC result for trimethoprim is significantly lower than either of these estimates, which may be due to chromosomal mutations with little to no fitness costs, differences in the E. coli strain used or be related to peculiarities of the G. mellonella infection model.
We did not conduct whole genome sequencing (WGS) to evaluate emergent resistance-associated mutations, but previous studies have found that mutations at residues P21, A26, D27, L28, W30, I94, and F153 of the dihydrofolate reductase (DHFR) enzyme play an important role in trimethoprim resistance in E. coli [33]. Of these, the L28R mutation is the most frequent mutation in the coding region of folA [33]. It not only increases the trimethoprim MIC but also acts as a compensatory mutation for the reduced catalytic activity caused by other DHFR mutations [33]. Brolund et al. investigated the distribution of dfr-genes and integrons in E. coli and found that the prevalence of integrons was 85% and 57% for integron class I and II, respectively [34]. Sequencing analysis revealed dfrA1 - the most common trimethoprim resistance present together with either dfrA5, dfrA7, dfrA14 or dfrA17 genes [34]. The likelihood of integron carriage increased with the number of resistance determinants [34]. mgrB, a gene involved in trimethoprim resistance, was identified by screening the E. coli single-gene knockout library [35], and Shi et al. investigated the mechanism of trimethoprim resistance, showing that its deletion upregulated the PhoP/Q system, leading to folA overexpression and DHFR-related resistance [36,37]. Mutations in DHFR’s promoter and ribosome-binding site also contributed to trimethoprim resistance [38,39]. Finally, deletion of glyA, a gene encoding serine hydroxy methyltransferase from the folate pathway, increased sensitivity to trimethoprim, indicating its implication in trimethoprim resistance [42].
Earlier, it was assumed that the mutant selective window (MSW) lies between the MICsusc and the MICres and that sub-MIC concentrations of antibiotics did not confer a selection pressure [43]. However, results from this study, along with previous MSC studies, have established that the sub-MIC selective window is significantly wider and extends to the MSC. Sub-MIC concentrations can select for resistant strains or induce de novo resistance [4]. Long-term persistence of resistance due to low concentrations of antibiotics in the environment may be sufficient to maintain resistance in a population of bacteria for bacterial pathogens whose life cycle involves periodic growth in the environment, such as E. coli [4]. The initial fitness costs associated with resistance mutations are often compensated for by secondary mutations, which allows these resistance mutations to spread in various settings [44].
In our trimethoprim experiments, an emergent trimethoprim-resistant isolate (MIC of 16 µg/mL) exhibited a double ellipse after 5 days of in vitro passaging. One ellipse was at 1.5 µg/mL and the other at 12 µg/mL, which is suggestive of a heterogenous subpopulation, whereby the population of E. coli with the MIC of 16 µg/mL was more dominant pre-passage, potentially due to low or cost-free mutations as a result of the antibiotic selective pressure. However, post-passage, the population with a MIC of 12 µg/mL was more fit without the selective pressure and, therefore, became more dominant. Such heterogeneous subpopulations are sometimes observed in gradient tests as a presence of bacterial colonies within the growth inhibition zone (ellipse) [45], as observed in our experiment.
In addition to the lack of WGS data, another limitation of this study was the fact that we evaluated the effect of trimethoprim on one strain of one bacterial species. Furthermore, this ATCC 25922 strain is used mainly for quality control purposes. It would, therefore, be useful to include other bacterial species and other strains of E. coli that are more pathogenic or clinically relevant in future experiments. It would be particularly important to include other commensal bacterial species, such as commensal Neisseria spp., who by virtue of their high prevalence, would be most exposed to antimicrobials in food [46]. Our model relied on establishing a chronic haemolymph infection in G. mellonella to determine whether low doses of trimethoprim could induce AMR. A more applicable approach would be to examine if these low doses ingested orally could induce AMR in bacteria in humans or other mammals [47]. Moreover, our study only explored the effect of low antibiotic doses on the emergence of de novo resistance without assessing the potential enrichment of pre-existing resistant strains or the spread of AMR through mobile genetic elements [1,5]. Future studies should investigate whether these low doses could interact with other substances in food, such as heavy metals, to induce and select for AMR as has been established in vitro [48]. Long-term, daily dosing regimens should also be implemented to evaluate if this would have a more pronounced effect on the emergence of AMR.
Despite these limitations, the study is the first of its kind to assess if low doses of trimethoprim can select for AMR in vivo. The positive findings from this study together with those of previous similar studies, suggest the need for equivalent studies in mammals [8,11,49]. Finally, our results should encourage the relevant authorities to include MSCs of antimicrobials in their determination of ADIs and MRLs. While the G. mellonella model is useful for testing numerous bug-drug combinations, further validation in mouse and human models will be required to validate the findings.
Authors’ contributions:
JKM, CK, SB, and ZG conceptualized the study. JKM conducted the MSC experiments. All authors read and approved the final draft.
Competing interests
The authors declare that they have no competing interests.
References
- Gullberg, E., et al., Selection of a multidrug resistance plasmid by sublethal levels of antibiotics and heavy metals. mBio, 2014. 5(5): p. e01918-14. [CrossRef]
- Hjort, K., et al., Antibiotic Minimal Selective Concentrations and Fitness Costs during Biofilm and Planktonic Growth. mBio, 2022. 13(3): p. e0144722. [CrossRef]
- Gestels, Z., et al., Ciprofloxacin Concentrations 100-Fold Lower than the MIC Can Select for Ciprofloxacin Resistance in Neisseria subflava: An In Vitro Study. Antibiotics, 2024. 13(6): p. 560.
- Gullberg, E., et al., Selection of Resistant Bacteria at Very Low Antibiotic Concentrations. PLOS Pathogens, 2011. 7(7): p. e1002158. [CrossRef]
- González, N., et al., Ciprofloxacin Concentrations 1/1000th the MIC Can Select for Antimicrobial Resistance in N. gonorrhoeae-Important Implications for Maximum Residue Limits in Food. Antibiotics (Basel), 2022. 11(10). [CrossRef]
- Kraupner, N., et al., Selective concentrations for trimethoprim resistance in aquatic environments. Environment International, 2020. 144: p. 106083. [CrossRef]
- Stanton, I.C., et al., Evolution of antibiotic resistance at low antibiotic concentrations including selection below the minimal selective concentration. Communications Biology, 2020. 3(1): p. 467. [CrossRef]
- Baranchyk, Y., et al., Effect of erythromycin residuals in food on the development of resistance in Streptococcus pneumoniae: an in vivo study in Galleria mellonella. PeerJ, 2024. 12: p. e17463. [CrossRef]
- Wu-Wu, J.W.F., et al., Antibiotic resistance and food safety: perspectives on new technologies and molecules for microbial control in the food industry. Antibiotics, 2023. 12(3): p. 550.
- FAO/WHO, Codex Alimentarius: Guidelines for the simple evaluation of dietary exposure to food additives. CAC/GL 3-1989 Adopted 1989. Revision 2014. 2014.
- Gestels, Z., et al., Could traces of fluoroquinolones in food induce ciprofloxacin resistance in Escherichia coli and Klebsiella pneumoniae? An in vivo study in Galleria mellonella with important implications for maximum residue limits in food. Microbiology Spectrum, 2024: p. e03595-23.
- Van Boeckel, T.P., et al., Global antibiotic consumption 2000 to 2010: an analysis of national pharmaceutical sales data. The Lancet Infectious Diseases, 2014. 14(8): p. 742-750. [CrossRef] [PubMed]
- Faleye, A.C., et al., Concentration and reduction of antibiotic residues in selected wastewater treatment plants and receiving waterbodies in Durban, South Africa. Science of The Total Environment, 2019. 678: p. 10-20. [CrossRef] [PubMed]
- Flach, C.-F., et al., A Comprehensive Screening of Escherichia coli Isolates from Scandinavia’s Largest Sewage Treatment Plant Indicates No Selection for Antibiotic Resistance. Environmental Science & Technology, 2018. 52(19): p. 11419-11428. [CrossRef]
- Khan, G.A., et al., Occurrence and Abundance of Antibiotics and Resistance Genes in Rivers, Canal and near Drug Formulation Facilities – A Study in Pakistan. PLOS ONE, 2013. 8(6): p. e62712. [CrossRef]
- Seo, J., et al. Antibiotic Residues in UK Foods: Exploring the Exposure Pathways and Associated Health Risks. Toxics, 2024. 12, DOI: 10.3390/toxics12030174.
- The European Agency for the Evaluation of Medicinal Products, COMMITTEE FOR VETERINARY MEDICINAL PRODUCTS: ENROFLOXACIN SUMMARY REPORT (5). 2002.
- Murray, A.K., et al., Dawning of a new ERA: Environmental Risk Assessment of antibiotics and their potential to select for antimicrobial resistance. Water Res, 2021. 200: p. 117233. [CrossRef]
- FAO/WHO. Codex Alimentarius Commission (CAC): Maximum Residue Limits. Available from: http://www.fao.org/fao-who-codexalimentarius/codex-texts/maximum-residue-limits/en/.
- Elder, H., et al., Human studies to measure the effect of antibiotic residues. Veterinary and human toxicology, 1993. 35: p. 31-36.
- European Food Safety Authority, The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2018/2019. EFSA Journal, 2021. 19(4).
- Mitchell, J., et al., Antimicrobial drug residues in milk and meat: causes, concerns, prevalence, regulations, tests, and test performance. Journal of food protection, 1998. 61(6): p. 742-756. [CrossRef]
- ATCC. Escherichia coli (Migula) Castellani and Chalmers 25922 ™. FDA strain Seattle 1946 [DSM 1103, NCIB 12210]]. Available from: https://www.atcc.org/products/25922#detailed-product-information.
- Gu, B., et al., Comparison of the prevalence and changing resistance to nalidixic acid and ciprofloxacin of Shigella between Europe–America and Asia–Africa from 1998 to 2009. International journal of antimicrobial agents, 2012. 40(1): p. 9-17. [CrossRef]
- Agency, T.E.M., The European Agency for the Evaluation of Medicinal Products. COMMITTEE FOR VETERINARY MEDICINAL PRODUCTS: TRIMETHOPRIM SUMMARY REPORT (2).
- Gleckman, R., N. Blagg, and D.W. Joubert, Trimethoprim: Mechanisms of Action, Antimicrobial Activity, Bacterial Resistance, Pharmacokinetics, Adverse Reactions, and Therapeutic Indications. Pharmacotherapy: The Journal of Human Pharmacology and Drug Therapy, 1981. 1(1): p. 14-19. [CrossRef]
- Papich, M.G., Saunders Handbook of Veterinary Drugs. Fourth Edition ed. 2016.
- Poirel, L., et al., Antimicrobial Resistance in Escherichia coli. Microbiology Spectrum, 2018. 6(4): p. 10.1128/microbiolspec.arba-0026-2017. [CrossRef]
- Minogue, T.D., et al., Complete Genome Assembly of Escherichia coli ATCC 25922, a Serotype O6 Reference Strain. Genome Announc, 2014. 2(5). [CrossRef]
- ATCC. Escherichia coli (Migula) Castellani and Chalmers 25922 ™. FDA strain Seattle 1946 [DSM 1103, NCIB 12210]]. Available from: https://www.atcc.org/products/25922#detailed-product-information. [CrossRef]
- Calarga, A.P., et al., Antimicrobial resistance and genetic background of non-typhoidal Salmonella enterica strains isolated from human infections in São Paulo, Brazil (2000–2019). Brazilian Journal of Microbiology, 2022. 53(3): p. 1249-1262. [CrossRef]
- Sivertsen, A., et al., Escherichia marmotae-a Human Pathogen Easily Misidentified as Escherichia coli. Microbiol Spectr, 2022. 10(2): p. e0203521. [CrossRef]
- Manna, M.S., et al., A trimethoprim derivative impedes antibiotic resistance evolution. Nature Communications, 2021. 12(1): p. 2949. [CrossRef]
- Brolund, A., et al., Molecular Characterisation of Trimethoprim Resistance in Escherichia coli and Klebsiella pneumoniae during a Two Year Intervention on Trimethoprim Use. PLOS ONE, 2010. 5(2): p. e9233. [CrossRef]
- Baba, T., et al., Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol Syst Biol, 2006. 2: p. 2006.0008. [CrossRef]
- Shi, H., et al., MgrB Inactivation Confers Trimethoprim Resistance in Escherichia coli. Frontiers in Microbiology, 2021. 12. [CrossRef]
- Smith, D.R. and J.M. Calvo, Nucleotide sequence of dihydrofolate reductase genes from trimethoprim-resistant mutants of Escherichia coli. Evidence that dihydrofolate reductase interacts with another essential gene product. Mol Gen Genet, 1982. 187(1): p. 72-8. [CrossRef]
- Flensburg, J. and O. Sköld, Regulatory changes in the formation of chromosomal dihydrofolate reductase causing resistance to trimethoprim. J Bacteriol, 1984. 159(1): p. 184-90. [CrossRef] [PubMed]
- Flensburg, J. and O. Sköld, Massive overproduction of dihydrofolate reductase in bacteria as a response to the use of trimethoprim. Eur J Biochem, 1987. 162(3): p. 473-6. [CrossRef]
- Sangurdekar, D.P., Z. Zhang, and A.B. Khodursky, The association of DNA damage response and nucleotide level modulation with the antibacterial mechanism of the anti-folate drug trimethoprim. BMC Genomics, 2011. 12: p. 583. [CrossRef] [PubMed]
- Mitosch, K., G. Rieckh, and T. Bollenbach, Noisy Response to Antibiotic Stress Predicts Subsequent Single-Cell Survival in an Acidic Environment. Cell Syst, 2017. 4(4): p. 393-403.e5. [CrossRef]
- Bhosle, A., et al., A Strategic Target Rescues Trimethoprim Sensitivity in Escherichia coli. iScience, 2020. 23(4): p. 100986. [CrossRef]
- Drlica, K., The mutant selection window and antimicrobial resistance. J Antimicrob Chemother, 2003. 52(1): p. 11-7. [CrossRef]
- Andersson, D.I. and D. Hughes, Antibiotic resistance and its cost: is it possible to reverse resistance? Nat Rev Microbiol, 2010. 8(4): p. 260-71. [CrossRef]
- Kowalska-Krochmal, B. and R. Dudek-Wicher, The Minimum Inhibitory Concentration of Antibiotics: Methods, Interpretation, Clinical Relevance. Pathogens, 2021. 10(2). [CrossRef]
- Laumen, J.G.E., et al., Antimicrobial susceptibility of commensal Neisseria in a general population and men who have sex with men in Belgium. Sci Rep, 2022. 12(1): p. 9.
- Gaulke, C.A., et al., Ecophylogenetics Clarifies the Evolutionary Association between Mammals and Their Gut Microbiota. mBio, 2018. 9(5).
- Seiler, C. and T.U. Berendonk, Heavy metal driven co-selection of antibiotic resistance in soil and water bodies impacted by agriculture and aquaculture. Front Microbiol, 2012. 3: p. 399.
- Kenyon, C., et al., Doxycycline PEP can induce doxycycline resistance in Klebsiella pneumoniae in a Galleria mellonella model of PEP. Front Microbiol, 2023. 14: p. 1208014.
Figure 1.
Schematic overview of study methodology (Figure produced with Biorender).
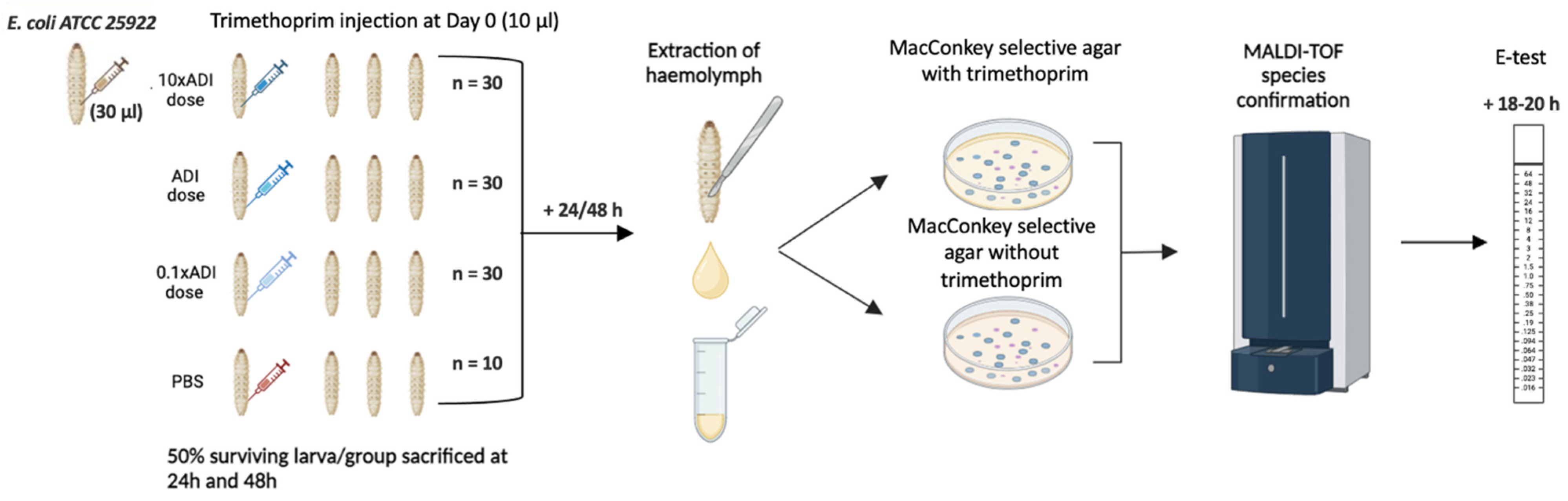
Figure 2.
Colonization of G. mellonella larvae with E. coli up to 2 days post inoculation of E. coli and various concentrations of trimethoprim, with the number of colonies (N) observed on plates without trimethoprim.
Figure 2.
Colonization of G. mellonella larvae with E. coli up to 2 days post inoculation of E. coli and various concentrations of trimethoprim, with the number of colonies (N) observed on plates without trimethoprim.
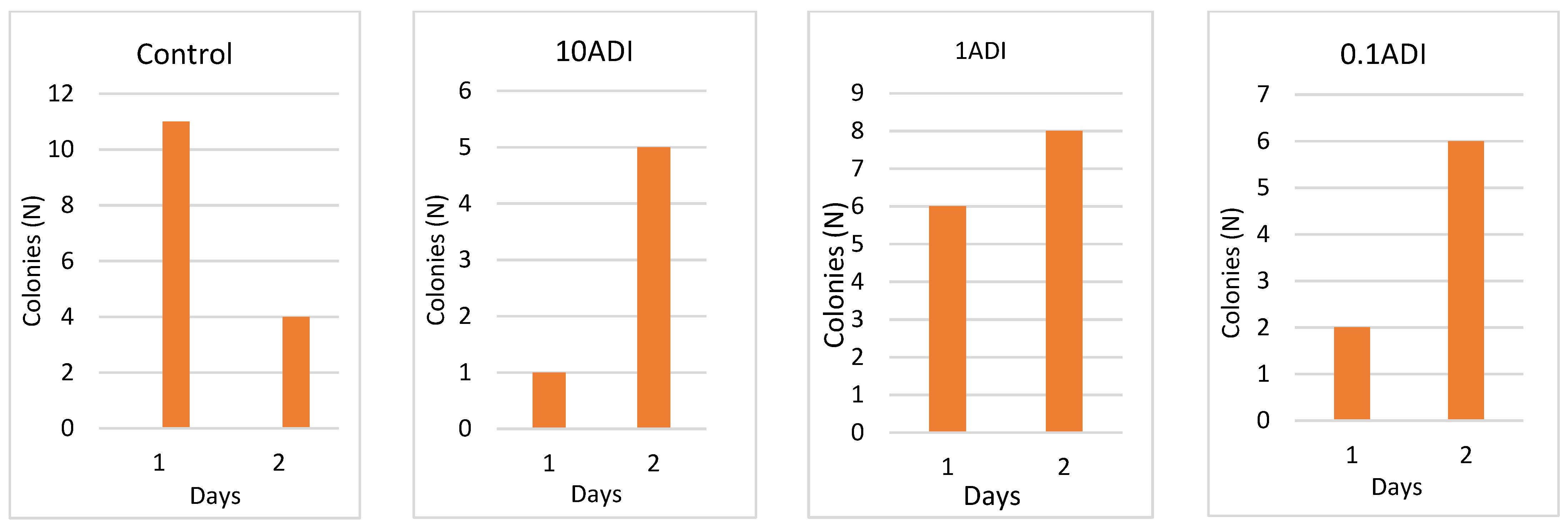
Figure 3.
A graph depicting the difference in trimethoprim MICs of E. coli between the wildtype (WT), 0.1x ADI control, and the resistant isolates.
Figure 3.
A graph depicting the difference in trimethoprim MICs of E. coli between the wildtype (WT), 0.1x ADI control, and the resistant isolates.
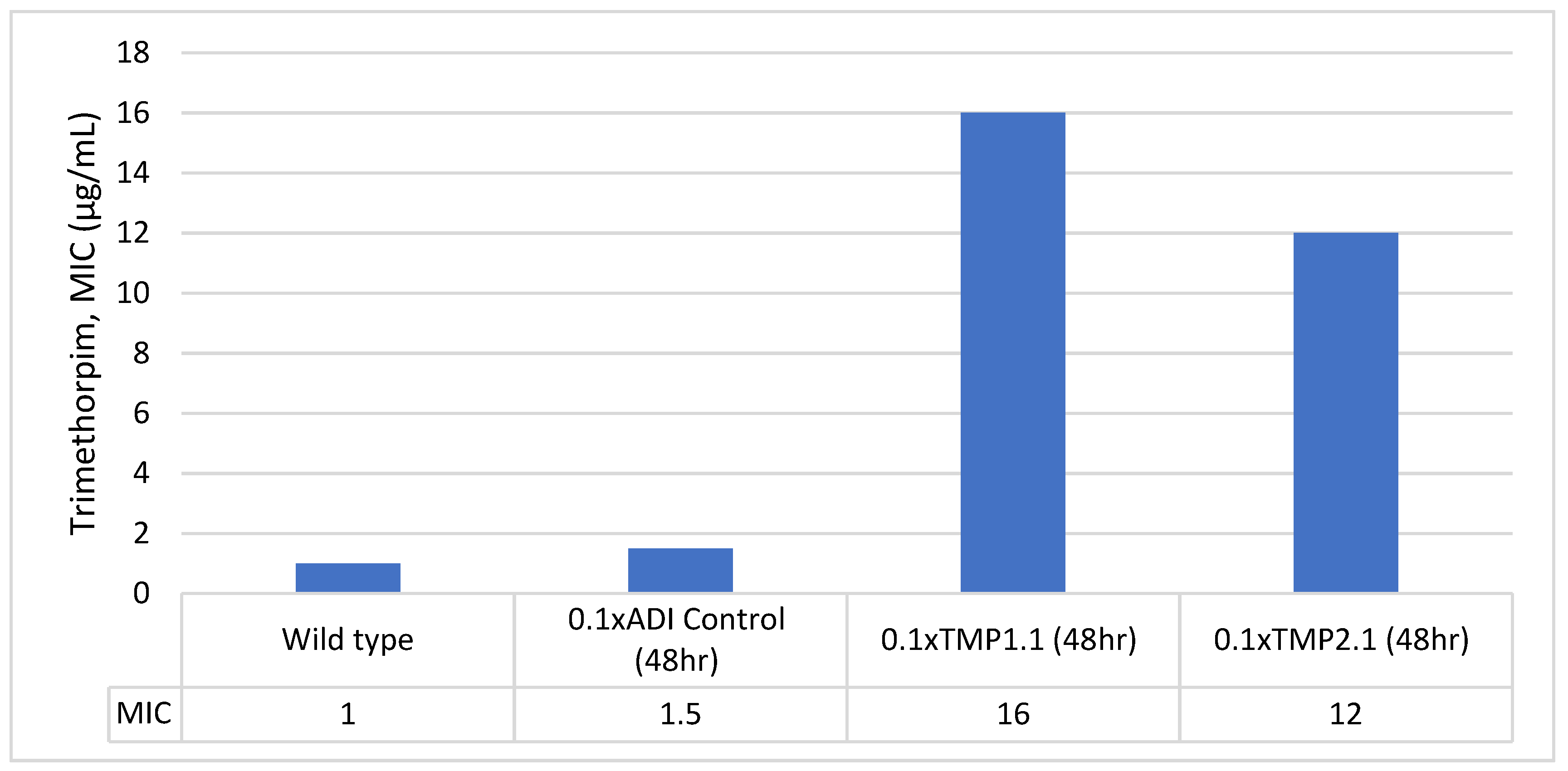

Table 1.
Phenotype and MALDI-TOF species identification with the respective confidence scores for each sample from MacConkey plates. The samples are named according to plate of colony, for example, 0.1x-24hr-TMP1.1 denotes the trimethoprim plate and colony ID (plate 1, colony 1 = 1.1) for the 0.1x ADI condition from the 24 h extraction.
Table 1.
Phenotype and MALDI-TOF species identification with the respective confidence scores for each sample from MacConkey plates. The samples are named according to plate of colony, for example, 0.1x-24hr-TMP1.1 denotes the trimethoprim plate and colony ID (plate 1, colony 1 = 1.1) for the 0.1x ADI condition from the 24 h extraction.
| Sample ID (MacConkey) | Phenotype | Detected species | MALDI-TOF Confidence Score | MIC (µg/mL) |
|---|---|---|---|---|
| 10x-24hr-TMP4.1 | Pink | Escherichia coli | 2.46 | 8 |
| 1x-24hr-TMP1.1 | Pink | Escherichia coli | 2.07 | 1.5 |
| 1x-24hr-TMP2.1 | Pink | Escherichia coli | 2.01 | 1 |
| 1x-24hr-TMP3.1 | Pink | Escherichia coli | 2.2 | 1.5 |
| 0.1x-24hr-TMP1.1 | Pink | Escherichia coli | 2.01 | 1.5 |
| 10x-48hr-TMP1.1 | Pink | Escherichia marmotae | 1.71 | 1.5 |
| 10x-48hr-TMP1.2 | Pink | Escherichia coli | 1.8 | 1.5 |
| 10x-48hr-TMP2.1 | Pink | Escherichia coli | 2.21 | 1.5 |
| 1x-48hr-TMP1.1 | Pink | Escherichia coli | 1.8 | 1.5 |
| 1x-48hr-TMP2.1 | Pink | Escherichia coli | 2.02 | 1.5 |
| 0.1x-48hr-TMP1.1 | Pink | Escherichia coli | 1.92 | 16 |
| 0.1x-48hr-TMP1.2 | Pink | Escherichia coli | 1.91 | 1 |
| 0.1x-48hr-TMP1.3 | Pink | Escherichia coli | 2.09 | 1.5 |
| 0.1x-48hr-TMP2.1 | Pink | Escherichia coli | 1.92 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



