
Submitted:
05 November 2024
Posted:
06 November 2024
You are already at the latest version
Abstract
Hard yellow maize is a crucial crop in Peruvian agriculture, playing a significant role in food security and livestock production. However, intensive fertilization practices in agronomic management have negatively impacted soil health. To explore more sustainable agricultural technologies, researchers have investigated using microorganisms to enhance plant growth. This study assessed the synergistic effects of microbial inoculants and mineral fertilization on INIA 619 and Dekal B-7088 maize varieties' yield and nutritional quality. A split-plot design was employed, incorporating four inoculation treatments—no inoculant, Bacillus subtilis, Trichoderma viride, and Pseudomonas putida—combined with fertilization levels of 0%, 50%, 75%, and 100%. The findings revealed that Bacillus subtilis boosted yields by 13.1% in INIA 619 and 55.5% in Dekal B-7088. Additionally, combined with 100% fertilization, microbial inoculation increased protein content by 47% and carbohydrates by 6% in INIA 619, while maintaining nutritional quality with 75% fertilization. Similarly, in Dekal B-7088, inoculation with total fertilization enhanced protein content by 54% and fiber by 27%. These results demonstrate that microbial inoculation can reduce mineral fertilization by up to 25% while sustaining high yields and improving the nutritional quality of maize.
Keywords:
1. Introduction
2. Materials and Methods
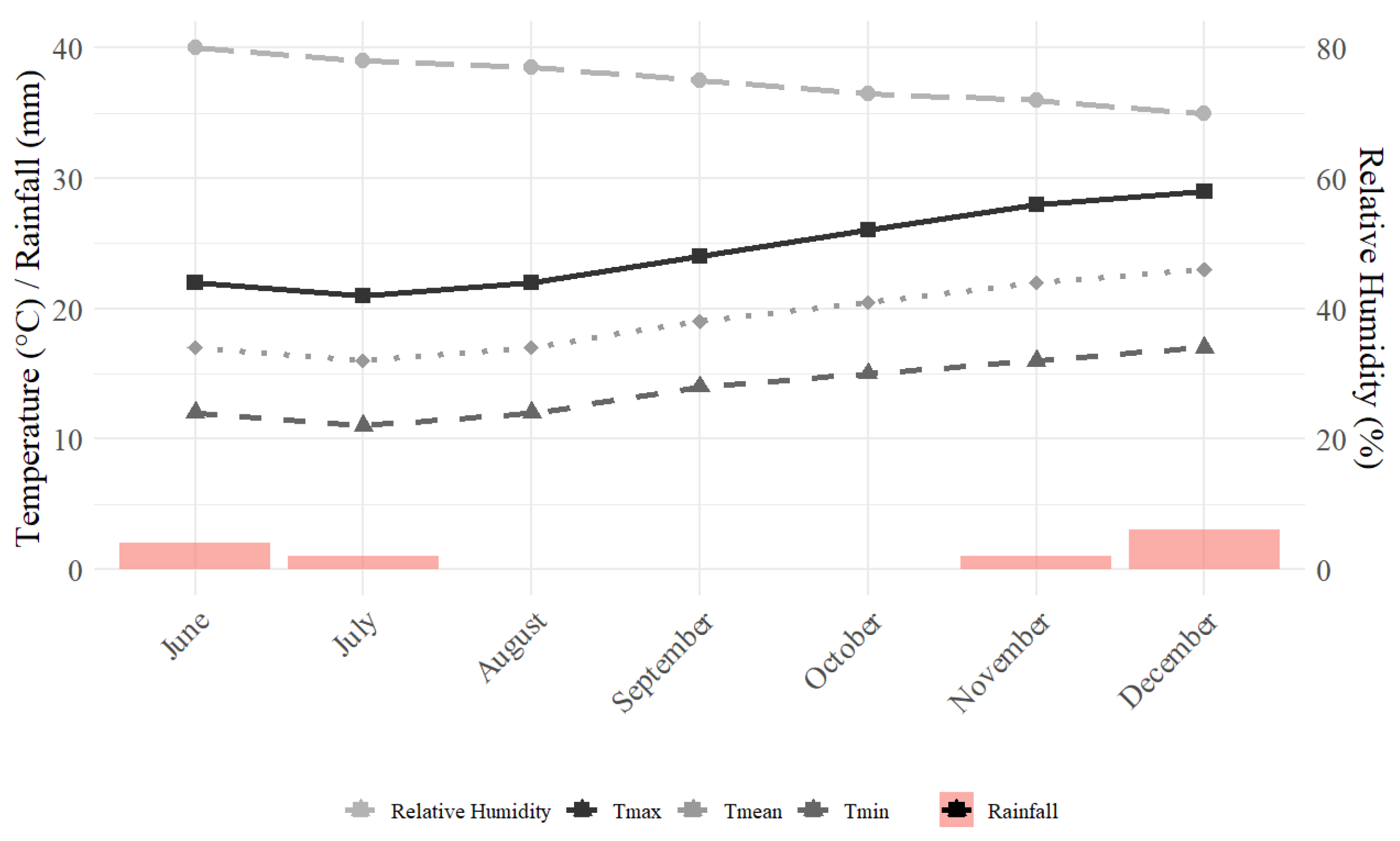
2.1. Trial Set-Up
2.2. Soil Characteristics
2.3. Experimental Design
2.4. Crop Fertilization
2.5. Microorganism Inoculation
2.6. Phytosanitary Management
2.7. Evaluated Parameters
2.8. Statistical Analysis
3. Results
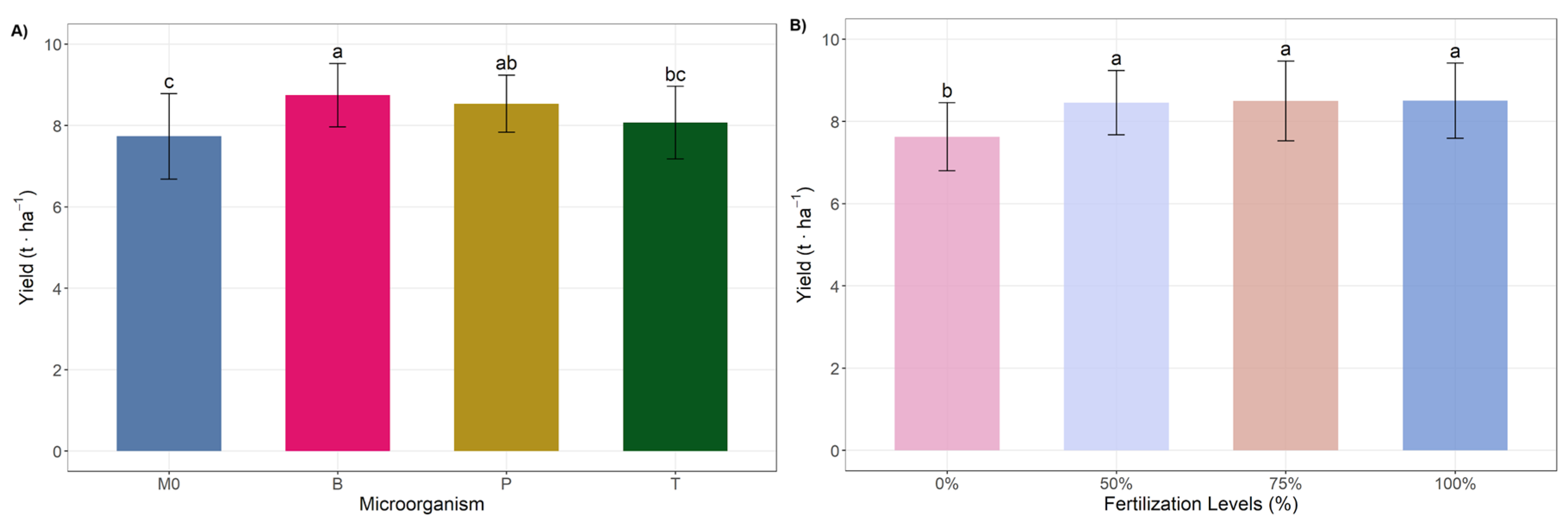
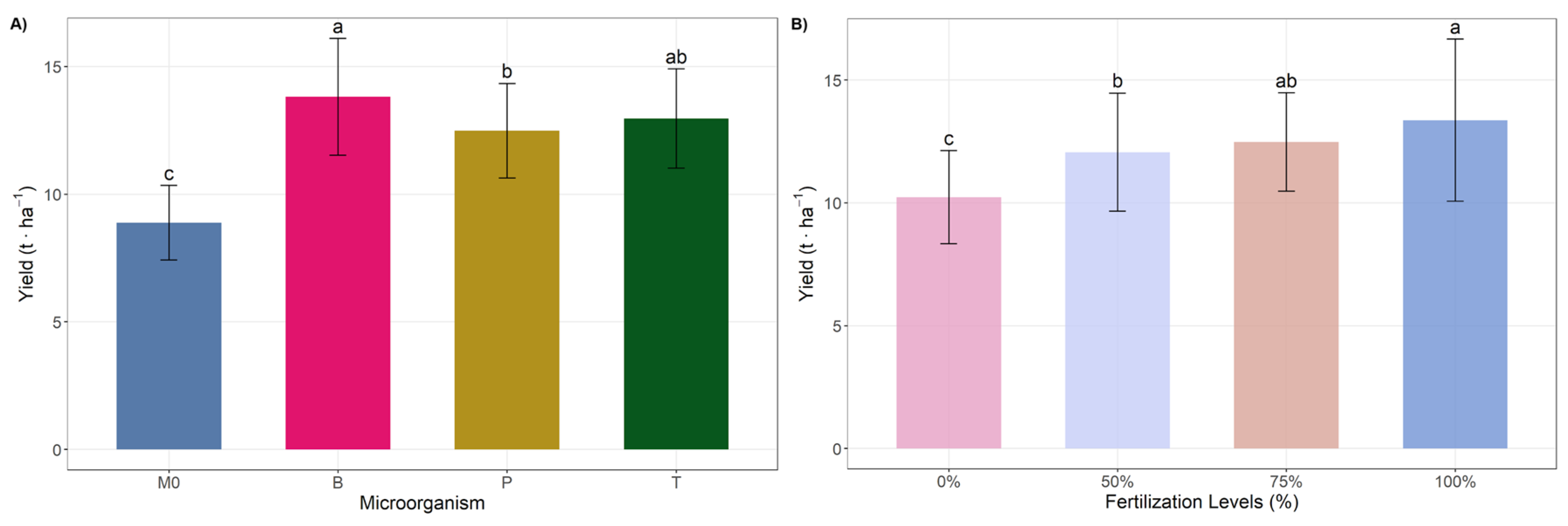
3.1. Ear Characteristics and Yield

| Factor | EL | ED | RE | GR | EW | GW | CW |
|---|---|---|---|---|---|---|---|
| (cm) | (cm) | (g) | (g) | (g) | |||
| Factor 1. Microorganisms | |||||||
| M0 | 15.48 ± 1.31 | 4.01 ± 0.15 | 13.65 ± 0.35 | 29.22 ± 2.55 | 158.72 ± 20.6 b | 135.15 ± 17.14 b | 23.57 ± 4.06 |
| B | 15.5 ± 0.88 | 4.04 ± 0.07 | 13.75 ± 0.4 | 29.33 ± 2.29 | 185.31 ± 18.16 a | 159.79 ± 16.79 a | 25.52 ± 2.15 |
| T | 15.63 ± 1.21 | 4.02 ± 0.1 | 13.77 ± 0.45 | 29.21 ± 2.74 | 178.19 ± 18.44 a | 151.86 ± 16.1 a | 25.66 ± 3.01 |
| P | 15.6 ± 0.89 | 4.08 ± 0.14 | 13.9 ± 0.34 | 29.57 ± 2.73 | 186.48 ± 17.66 a | 158.96 ± 15.03 a | 26.69 ± 4.44 |
| (%) | Factor 2. Fertilization Dose | ||||||
| 0 | 14.99 ± 1.33 | 4.04 ± 0.15 | 13.86 ± 0.33 | 27.34 ± 2.11 b | 160.85 ± 20.42 b | 136.75 ± 17.47 b | 24.1± 3.48 |
| 50 | 15.97 ± 1.07 | 4.05 ± 0.1 | 13.91 ± 0.22 | 30.31 ± 2.53 a | 182.11 ± 19.57 a | 155.78 ± 17.32 a | 26.33± 2.62 |
| 75 | 15.77 ± 0.69 | 4.04 ± 0.09 | 13.65 ± 0.38 | 29.88 ± 1.93 a | 179.53 ± 17.87 a | 154.39 ± 15.27 a | 24.31± 3.33 |
| 100 | 15.47 ± 0.9 | 4.01 ± 0.14 | 13.65 ± 0.52 | 29.8 ± 2.51 a | 186.2 ± 20.35 a | 158.82 ± 18.02 a | 26.7± 4.42 |
| Interaction | 0.95 | 0.61 | 0.51 | 0.86 | 0.88 | 0.92 | 0.66 |
| Factor | EL | ED | RE | GR | EW | GW | CW |
|---|---|---|---|---|---|---|---|
| (cm) | (cm) | (g) | (g) | (g) | |||
| Factor 1. Microorganisms | |||||||
| M0 | 11.1 ± 4.01 | 4.39 ± 0.1 | 16.64 ± 0.47 | 31.83 ± 2.79 | 180.53 ± 14.52 | 164.3 ± 13.25 | 15.33 ± 2.03 |
| B | 11.47 ± 4.31 | 4.44 ± 0.1 | 16.81 ± 0.7 | 33.11 ± 1.76 | 191.14 ± 18.02 | 175.09 ± 16.4 | 16.42 ± 3.76 |
| T | 11.29 ± 4.22 | 4.42 ± 0.13 | 16.65 ± 0.37 | 32.55 ± 1.19 | 187.55 ± 18.13 | 172.22 ± 16.29 | 16.22 ± 3.64 |
| P | 11.13 ± 4.06 | 4.39 ± 0.12 | 16.73 ± 0.71 | 32.46 ± 1.27 | 185.66 ± 11.65 | 169.24 ± 10.3 | 16.05 ± 1.89 |
| (%) | Factor 2. Fertilization dose | ||||||
| 0 | 13.16 ± 0.42 b | 4.35 ± 0.11 b | 16.36 ± 0.35 | 31.71 ± 1.31 | 176.38 ± 12.29 b | 161.65 ± 10.97 c | 14.73 ± 1.61 |
| 50 | 13.4 ± 0.69 b | 4.41 ± 0.11 ab | 16.64 ± 0.64 | 32.84 ± 2.37 | 184.32 ± 13.81 ab | 168.09 ± 12.6 bc | 16.86 ± 3.6 |
| 75 | 13.48 ± 0.71 ab | 4.43 ± 0.1 a | 16.86 ± 0.61 | 32.05 ± 1.65 | 189.8 ± 14.99 a | 172.94 ± 13.9 ab | 16.22 ± 3.79 |
| 100 | 13.92 ± 0.64 a | 4.44 ± 0.11 a | 16.98 ± 0.49 | 33.36 ± 1.7 | 194.38 ± 17.36 a | 178.17 ± 15.85 a | 16.21 ± 1.8 |
| Interaction | 0.55 | 0.39 | 0.22 | 0.95 | 0.83 | 0.66 | 0.85 |
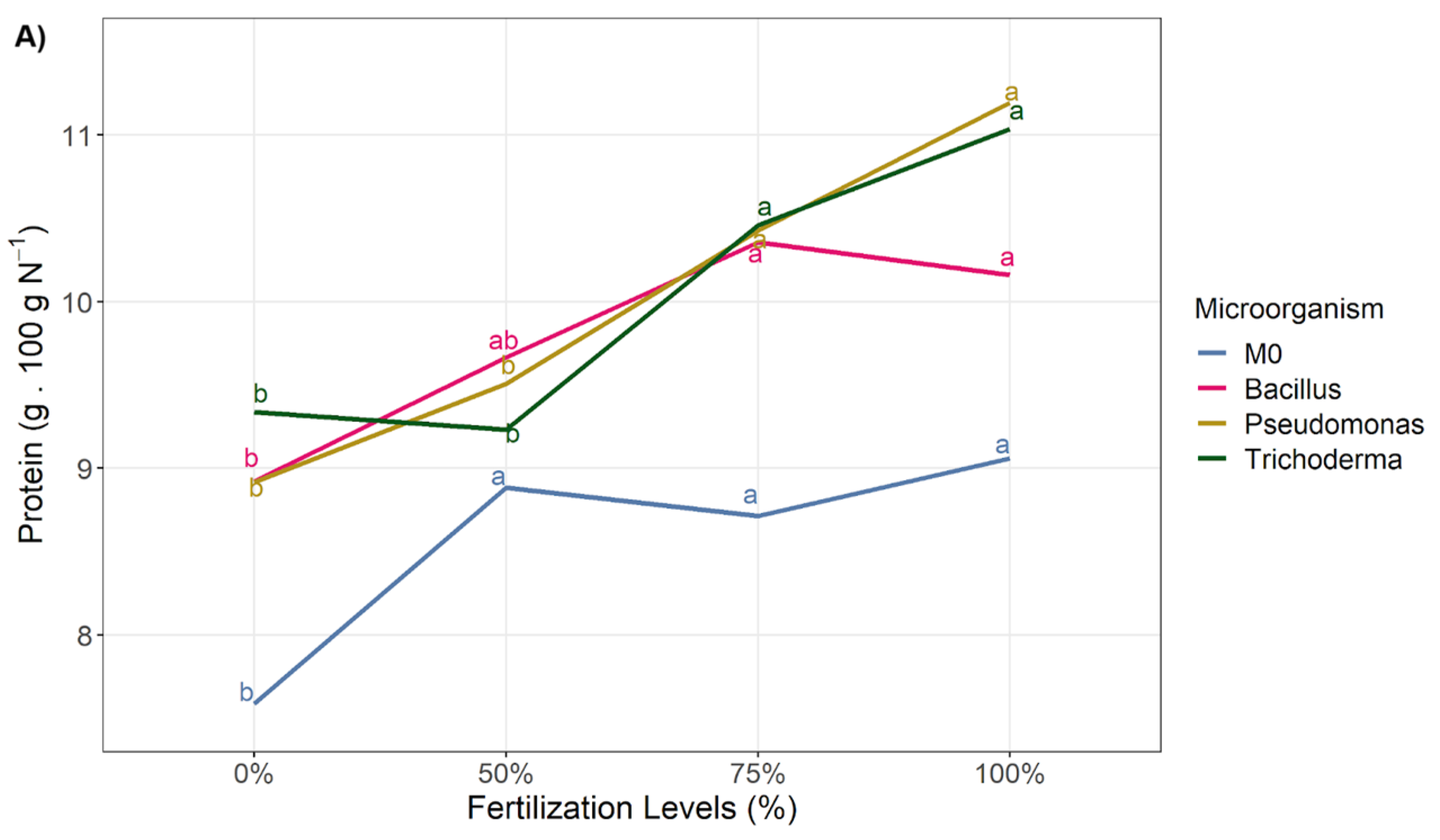
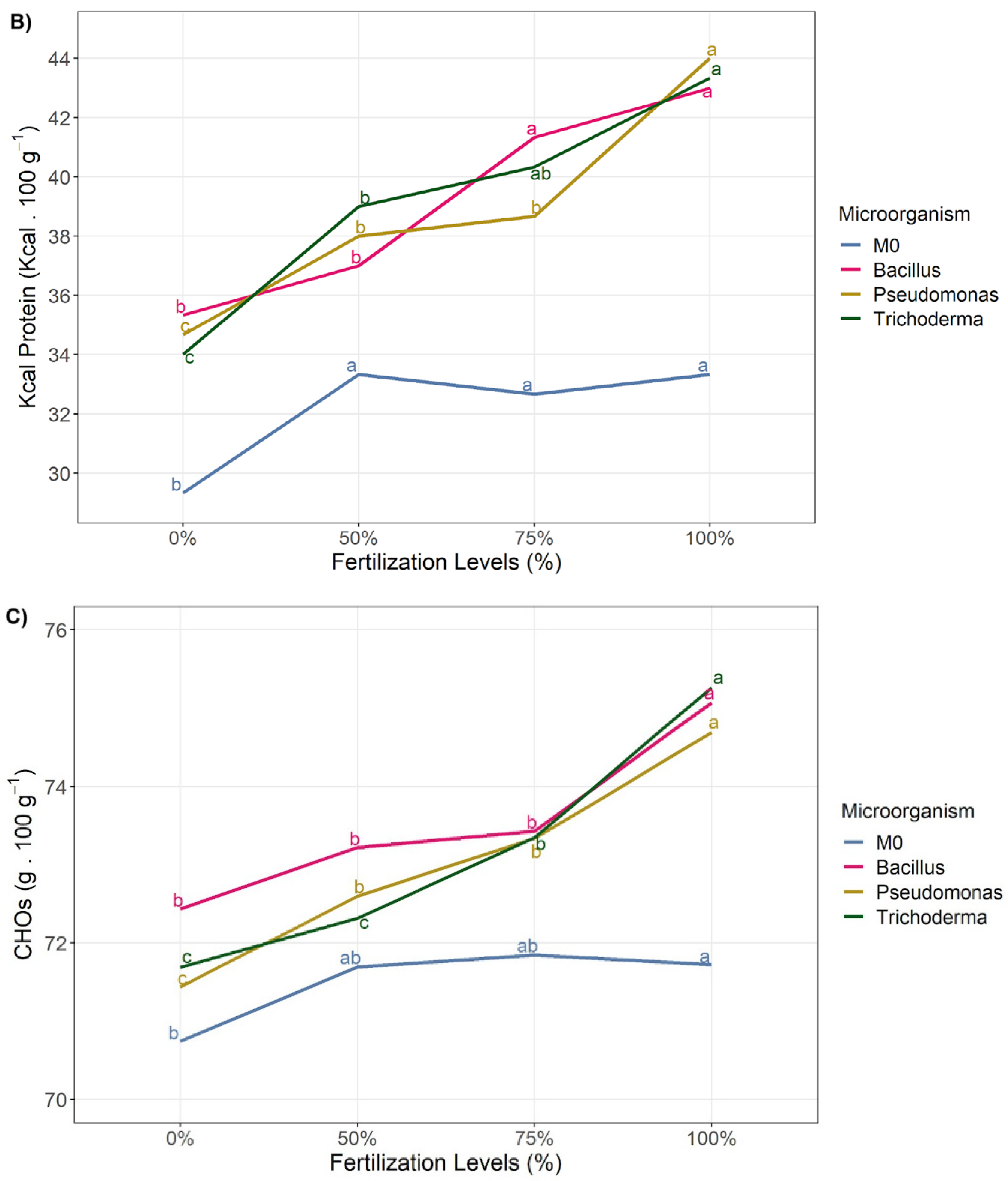
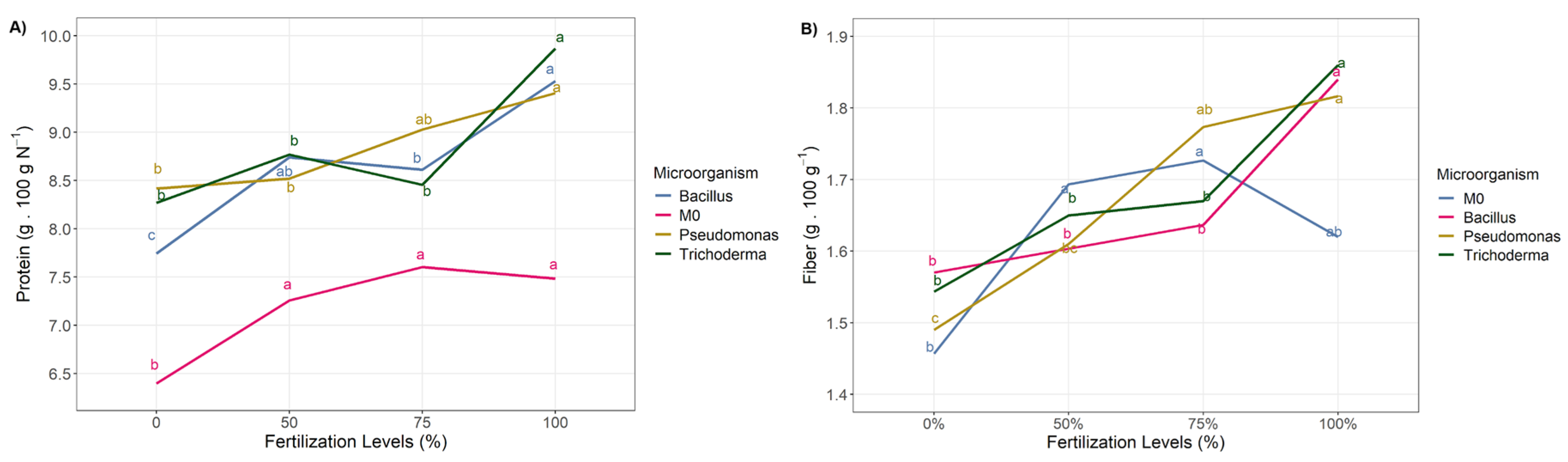
3.2. Nutritional Quality
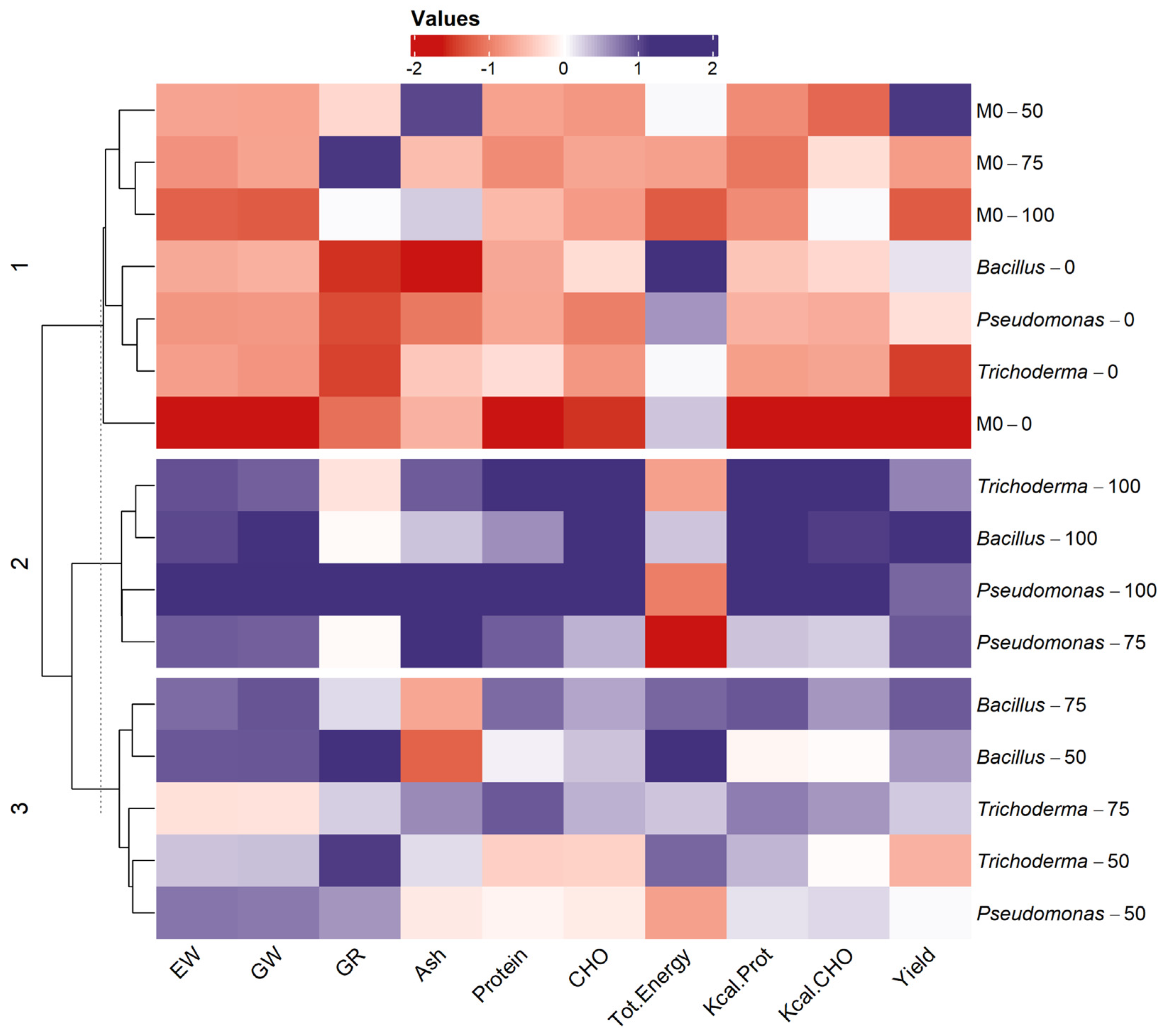
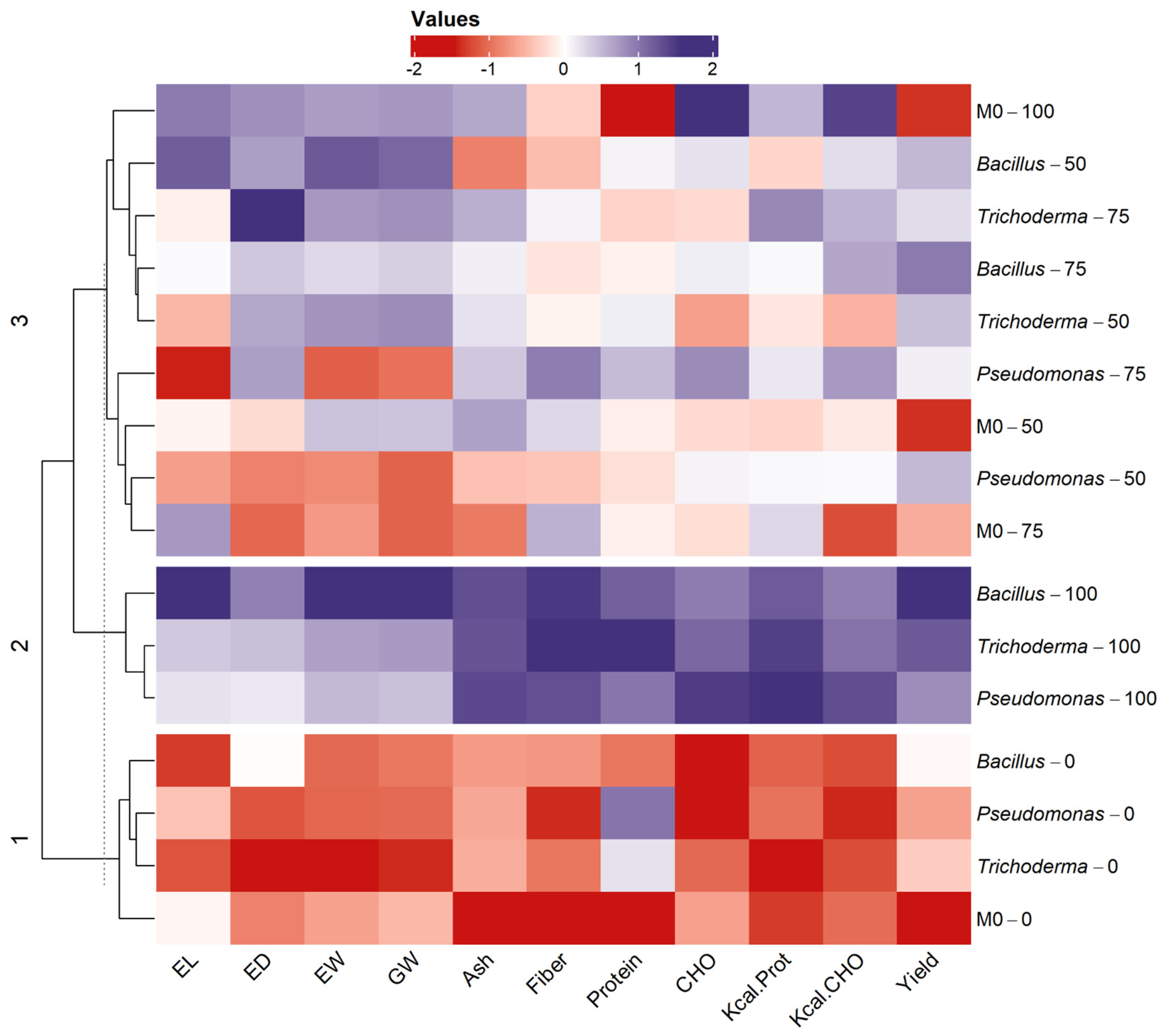
3.3. Heatmps
3.3.1. INIA 619
3.3.2. DEKAL B-7088
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shah, T.R., T.; Prasad, K.; Kumar, P. Maize — A Potential Source of Human Nutrition and Health: A Review. Cogent Food Agric. 2016, 2, 1166995. [CrossRef]
- Acosta, L.; Barreda, C.; Becerra, J.; Galarreta, L.; Huaman, O.; Moreyra, J.; Romero, C.; Rospigliosi, J. Marco Orientador de Cultivos, Campaña 2024/2025 2024.
- Perfil Productivo y Competitivo de los Principales Cultivos del Sectir Available online: https://app.powerbi.com/view?r=eyJrIjoiYjYwYTk5MDgtM2M0MS00NDMyLTgzNDEtMjNhNjEzYWQyOTNlIiwidCI6IjdmMDg0NjI3LTdmNDAtNDg3OS04OTE3LTk0Yjg2ZmQzNWYzZiJ9 (accessed on 4 October 2024).
- Barandiarán, M. Manual Técnico Del Cultivo de Maíz Amarillo Duro; 1; First.; Instituto Nacional de Innovación Agraria: Perú, 2020; ISBN 978-9972-44-051-9.
- Yan, X.; Chen, X.; Ma, C.; Cai, Y.; Cui, Z.; Chen, X.; Wu, L.; Zhang, F. What Are the Key Factors Affecting Maize Yield Response to and Agronomic Efficiency of Phosphorus Fertilizer in China? Field Crops Res. 2021, 270, 108221. [CrossRef]
- Samaniego, T.; Pérez, W.E.; Lastra-Paúcar, S.; Verme-Mustiga, E.; Solórzano-Acosta, R. The Fermented Liquid Biofertilizer Use Derived from Slaughterhouse Waste Improves Maize Crop Yield. Trop. Subtrop. Agroecosystems 2024, 27. [CrossRef]
- Mulyati; Baharuddin, A.B.; Tejowulan, R.S. Improving Maize (Zea Mays L.) Growth and Yield by the Application of Inorganic and Organic Fertilizers Plus. IOP Conf. Ser. Earth Environ. Sci. 2021, 712, 012027. [CrossRef]
- FAO. El estado de los recursos de tierras y aguas del mundo para la alimentación y la agricultura. La gestión de los sistemas en situación de riesgo; 2011.
- Bhattacharyya, P.N.; Goswami, M.P.; Bhattacharyya, L.H. Perspective of Beneficial Microbes in Agriculture under Changing Climatic Scenario: A Review. J. Phytol. 2016, 8, 26. [CrossRef]
- Parra-Cota, F.I.; Coronel-Acosta, C.-B.; Amézquita-Avilés, C.F.; Santos-Villalobos, S. de los; Escalante-Martínez, D.I. Diversidad metabólica de microorganismos edáficos asociados al cultivo de maíz en el Valle del Yaqui, Sonora. Rev. Mex. Cienc. Agríc. 9, 431–442.
- Gilbert, J.A.; Neufeld, J.D. Life in a World without Microbes. PLOS Biol. 2014, 12, e1002020. [CrossRef]
- Wu, Q.; Chen, Y.; Dou, X.; Liao, D.; Li, K.; An, C.; Li, G.; Dong, Z. Microbial Fertilizers Improve Soil Quality and Crop Yield in Coastal Saline Soils by Regulating Soil Bacterial and Fungal Community Structure. Sci. Total Environ. 2024, 949, 175127. [CrossRef]
- Todeschini, V.; AitLahmidi, N.; Mazzucco, E.; Marsano, F.; Gosetti, F.; Robotti, E.; Bona, E.; Massa, N.; Bonneau, L.; Marengo, E.; et al. Impact of Beneficial Microorganisms on Strawberry Growth, Fruit Production, Nutritional Quality, and Volatilome. Front. Plant Sci. 2018, 9, 1611. [CrossRef]
- Vassilev, N.; Malusà, E. Special Issue: Microorganisms and Plant Nutrition. Microorganisms 2021, 9, 2571. [CrossRef]
- Widnyana, I.K.; Javandira, C. Activities Pseudomonas Spp. and Bacillus Sp. to Stimulate Germination and Seedling Growth of Tomato Plants. Agric. Agric. Sci. Procedia 2016, 9, 419–423. [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability—A Review. Molecules 2016, 21, 573. [CrossRef]
- Tsukanova, K.A.; Сhеbotаr, V.К.; Meyer, J.; Bibikova, T. Effect of Plant Growth-Promoting Rhizobacteria on Plant Hormone Homeostasis. South Afr. J. Bot. 2017, 113, 91–102. [CrossRef]
- Etesami, H.; Glick, B.R. Halotolerant Plant Growth–Promoting Bacteria: Prospects for Alleviating Salinity Stress in Plants. Environ. Exp. Bot. 2020, 178, 104124. [CrossRef]
- Sah, S.; Krishnani, S.; Singh, R. Pseudomonas Mediated Nutritional and Growth Promotional Activities for Sustainable Food Security. Curr. Res. Microb. Sci. 2021, 2, 100084. [CrossRef]
- Efthimiadou, A.; Katsenios, N.; Chanioti, S.; Giannoglou, M.; Djordjevic, N.; Katsaros, G. Effect of Foliar and Soil Application of Plant Growth Promoting Bacteria on Growth, Physiology, Yield and Seed Quality of Maize under Mediterranean Conditions. Sci. Rep. 2020, 10, 21060. [CrossRef]
- Konieczna, W.; Turkan, S.; Warchoł, M.; Skrzypek, E.; Dąbrowska, G.B.; Mierek-Adamska, A. The Contribution of Trichoderma Viride and Metallothioneins in Enhancing the Seed Quality of Avena Sativa L. in Cd-Contaminated Soil. Foods 2024, 13, 2469 . [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Woo, S.L.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Ruocco, M.; Lanzuise, S.; et al. Trichoderma Secondary Metabolites Active on Plants and Fungal Pathogens.The Open Mycology Journal 2014, 8,127-139. [CrossRef]
- Rodríguez-García, D.; Vargas-Rojas, J. Efecto de la inoculación con Trichoderma sobre el crecimiento vegetativo del tomate (Solanum lycopersicum). Agron. Costarric. 2022.
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The Current Status of Its Application in Agriculture for the Biocontrol of Fungal Phytopathogens and Stimulation of Plant Growth. Int. J. Mol. Sci. 2022, 23, 2329. [CrossRef]
- NOM-021-RECNAT-2000; Norma Oficial Mexicana Que Establece Las Especificaciones de Fertilidad, Salinidad y Clasificación de Suelos. Estudios, Muestreo y Análisis. 2002. Available online: https://faolex.fao.org/docs/pdf/mex50674.pdf (accessed on 08 August 2024).
- USEPA. METHOD 9045D. SOIL AND WASTE pH 2004; USEPA: Washington, DC, USA, 2004. Available online:https://www.epa.gov/sites/default/files/2015-12/documents/9045d.pdf (accessed on 14 August 2024).
- ISO 11265:1994; Soil Quality—Determination of the Specific Electrical Conductivity. ISO (International Organization for Standardization): Geneva, Switzerland, 1994. Available online: https://www.iso.org/standard/19243.html (accessed on 08 August 2024).
- Verhulst, Nele; Sayre, Ken; Govaerts, Bram Manual de determinación de rendimiento; 1er ed.; Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT): México, 2012; ISBN 978-607-95844-7-4.
- AACC-Method 46-11 American Association of Cereal Chemists. Crude Protein – Improved Kjeldahl Method, Copper Catalyst Modification; St. Paul, USA, 2009.
- INACAL. Instituto Nacional de Calidad. Norma Técnica Peruana. NTP 205.004:2022. Cereales y Leguminosas. Determinación de Cenizas; 2022.
- INACAL. Instituto Nacional de Calidad. Norma Técnica Peruana. NTP 209.019:1976 . ALIMENTOS BALANCEADOS PARA ANIMALES. (Revisada El 2014); Perú, 2014.
- AOCS (American Oil Chemists’ Society) American Oil Chemists’ Society. Ba 6-84 Crude Fiber in Oilseed By-Products; USA, 2017.
- Collazos, C; Phlip, W.; Viñas, E; Alvistur, J; Urquieta, A; Vásquez, J Ministerio de Salud, Instituto Nacional de Nutrición. Metodología Para Carbohidratos, Por Diferencia de Materia Seca (MS-INN); Lima, Perú, 1993.
- Mendiburu, F. de Agricolae: Statistical Procedures for Agricultural Research 2023. Available online: https://cran.r-project.org/web/packages/agricolae/index.html (accessed on 10 August 2024).
- Paliwoda, D.; Mikiciuk, G. Use of Rhizosphere Microorganisms in Plant Production – A Review Study. Journal of Ecological Engineering 2020, 21, 292-310. [CrossRef]
- dos Santos, R.M.; Diaz, P.A.E.; Lobo, L.L.B.; Rigobelo, E.C. Use of Plant Growth-Promoting Rhizobacteria in Maize and Sugarcane: Characteristics and Applications. Front. Sustain. Food Syst. 2020, 4. [CrossRef]
- Pereira, S.I.A.; Abreu, D.; Moreira, H.; Vega, A.; Castro, P.M.L. Plant Growth-Promoting Rhizobacteria (PGPR) Improve the Growth and Nutrient Use Efficiency in Maize (Zea Mays L.) under Water Deficit Conditions. Heliyon 2020, 6. [CrossRef]
- Huasasquiche, L.; Alejandro, L.; Ccori, T.; Cántaro-Segura, H.; Samaniego, T.; Quispe, K.; Solórzano, R. Bacillus Subtilis and Rhizophagus Intraradices Improve Vegetative Growth, Yield, and Fruit Quality of Fragaria × Ananassa Var. San Andreas. Microorganisms 2024, 12, 1816. [CrossRef]
- Solórzano-Acosta, R.A.; Quispe, K.R. Assessing the Role of Field Isolated Pseudomonas and Bacillus as Growth-promoting Rizobacteria on Avocado ( Persea Americana ) Seedlings. J. Sustain. Agric. Environ. 2024, 3, e12114. [CrossRef]
- Araujo, J.L.; de Mesquita Alves, J.; Rocha, R.H.C.; Santos, J.Z.L.; dos Santos Barbosa, R.; da Costa, F.M.N.; de Lima, G.S.; de Freitas, L.N.; Lima, A.S.; Nogueira, A.E.P.; et al. Beneficial Microorganisms Affect Soil Microbiological Activity and Corn Yield under Deficit Irrigation. Agriculture 2023, 13, 1169. [CrossRef]
- Ning, P.; Li, S.; Yu, P.; Zhang, Y.; Li, C. Post-Silking Accumulation and Partitioning of Dry Matter, Nitrogen, Phosphorus and Potassium in Maize Varieties Differing in Leaf Longevity. Field Crops Res. 2013, 144, 19–27. [CrossRef]
- Wang, J.; Zhao, S.; Zhang, Y.; Lu, X.; Du, J.; Wang, C.; Wen, W.; Guo, X.; Zhao, C. Investigating the Genetic Basis of Maize Ear Characteristics: A Comprehensive Genome-Wide Study Utilizing High-Throughput Phenotypic Measurement Method and System. Front. Plant Sci. 2023, 14, 1248446. [CrossRef]
- Bayer. DEKALB - 7088 Available online: https://www.agro.bayer.pe/es-pe/dekalb/details.html/dekalb_-_7088.html (accessed on 14 October 2024).
- Instituto Nacional de Innovación Agraria Híbrido Simple de Maíz Amarillo Duro INIA 619 - Megahíbrido 2012.
- Ribeiro, V.P.; Gomes, E.A.; De Sousa, S.M.; De Paula Lana, U.G.; Coelho, A.M.; Marriel, I.E.; De Oliveira-Paiva, C.A. Co-Inoculation with Tropical Strains of Azospirillum and Bacillus Is More Efficient than Single Inoculation for Improving Plant Growth and Nutrient Uptake in Maize. Arch. Microbiol. 2022, 204, 143. [CrossRef]
- de Oliveira-Paiva, C.A.; Bini, D.; de Sousa, S.M.; Ribeiro, V.P.; dos Santos, F.C.; de Paula Lana, U.G.; de Souza, F.F.; Gomes, E.A.; Marriel, I.E. Inoculation with Bacillus Megaterium CNPMS B119 and Bacillus Subtilis CNPMS B2084 Improve P-Acquisition and Maize Yield in Brazil. Front. Microbiol. 2024, 15, 1426166. [CrossRef]
- Gholami, A.; Shahsavani, S.; Nezarat, S. The Effect of Plant Growth Promoting Rhizobacteria (PGPR) on Germination, Seedling Growth and Yield of Maize. Engineering and Technology, 2008, 49, 19-24.
- Elhaissoufi, W.; Ghoulam, C.; Barakat, A.; Zeroual, Y.; Bargaz, A. Phosphate Bacterial Solubilization: A Key Rhizosphere Driving Force Enabling Higher P Use Efficiency and Crop Productivity. J. Adv. Res. 2022, 38, 13–28. [CrossRef]
- Sosa-Rodrigues, B.A.; García-Vivas, Y.S. Eficiencia de uso del nitrógeno en maíz fertilizado de forma orgánica y mineral. Agron. Mesoam. 2018, 29, 207. [CrossRef]
- Mpanga, I.K.; Nkebiwe, P.M.; Kuhlmann, M.; Cozzolino, V.; Piccolo, A.; Geistlinger, J.; Berger, N.; Ludewig, U.; Neumann, G. The Form of N Supply Determines Plant Growth Promotion by P-Solubilizing Microorganisms in Maize. Microorganisms 2019, 7, 38. [CrossRef]
- Nepali, B.; Subedi, S.; Bhattarai, S.; Marahatta, S.; Bhandari, D.; Shrestha, J. Bio-Fertilizer Activity of Trichoderma Viride and Pseudomonas Fluorescens as Growth and Yield Promoter for Maize. 2020,191-195. [CrossRef]
- Syamsiyah, J.; Herdiansyah, G.; Hartati, S. Use of Trichoderma as an Effort to Increase Growth and Productivity of Maize Plants. IOP Conf. Ser. Earth Environ. Sci. 2023, 1165, 012020. [CrossRef]
- Khalid, A.; Arshad, M.; Zahir, Z.A. Screening Plant Growth-Promoting Rhizobacteria for Improving Growth and Yield of Wheat. J. Appl. Microbiol. 2004, 96, 473–480. [CrossRef]
- Ochieng’, I.O.; Gitari, H.I.; Mochoge, B.; Rezaei-Chiyaneh, E.; Gweyi-Onyango, J.P. Optimizing Maize Yield, Nitrogen Efficacy and Grain Protein Content under Different N Forms and Rates. J. Soil Sci. Plant Nutr. 2021, 21, 1867–1880. [CrossRef]
- Feng, W.; Xue, W.; Zhao, Z.; Shi, Z.; Wang, W.; Bai, Y.; Wang, H.; Qiu, P.; Xue, J.; Chen, B. Nitrogen Fertilizer Application Rate Affects the Dynamic Metabolism of Nitrogen and Carbohydrates in Kernels of Waxy Maize. Front. Plant Sci. 2024, 15. [CrossRef]
- Monostori, I.; Szira, F.; Tondelli, A.; Árendás, T.; Gierczik, K.; Cattivelli, L.; Galiba, G.; Vágújfalvi, A. Genome-Wide Association Study and Genetic Diversity Analysis on Nitrogen Use Efficiency in a Central European Winter Wheat (Triticum Aestivum L.) Collection. PLoS ONE 2017, 12, e0189265. [CrossRef]
- Aquino, J.P.A. de; Macedo Junior, F.B. de; Antunes, J.E.L.; Figueiredo, M. do V.B.; Alcântara Neto, F. de; Araujo, A.S.F. de Plant Growth-Promoting Endophytic Bacteria on Maize and Sorghum. Pesqui. Agropecuária Trop. 2019, 49, e56241. [CrossRef]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; AL-Harrasi, A. Plant Growth Promoting Bacteria as an Alternative Strategy for Salt Tolerance in Plants: A Review. Microbiol. Res. 2018, 209, 21–32. [CrossRef]
- Lastochkina, O.; Pusenkova, L.; Yuldashev, R.; Babaev, M.; Garipova, S.; Blagova, D.; Khairullin, R.; Aliniaeifard, S. Effects of Bacillus Subtilis on Some Physiological and Biochemical Parameters of Triticum Aestivum L. (Wheat) under Salinity. Plant Physiol. Biochem. 2017, 121, 80–88. [CrossRef]
- Akladious, S.A.; Abbas, S.M. Application of Trichoderma Harzianum T22 as a Biofertilizer Potential in Maize Growth. J. Plant Nutr. 2014, 37, 30–49. [CrossRef]
- Saeed, F.; Hussain, M.; Arshad, M.S.; Afzaal, M.; Munir, H.; Imran, M.; Tufail, T.; Anjum, F. Functional and Nutraceutical Properties of Maize Bran Cell Wall Non-Starch Polysaccharides. Int. J. Food Prop. 2021, 24, 233–248. [CrossRef]
- Mutungi, C.; Tungu, J.; Amri, J.; Gaspar, A.; Abass, A. Nutritional Benefits of Improved Post-Harvest Handling Practices for Maize and Common Beans in Northern Tanzania: A Quantitative Farm-Level Assessment. J. Stored Prod. Res. 2022, 95, 101918. [CrossRef]
- Hawkesford, M.J.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Skrumsager Moller, I.; White, P. Functions of Macronutrients. In; Marschner, P., Ed.; Academic Press Amsterdam, Netherlands, 2012; pp. 135–189 ISBN 978-0-12-384905-2.
- Havlin, J.L. Soil: Fertility and Nutrient Management. In Landscape and Land Capacity; CRC Press, 2020 ISBN 978-0-429-44555-2.
- Ndukwe, O.K.; Edeoga, H.O.; Omosun, G. Varietal Differences in Some Nutritional Composition of Ten Maize (Zea Mays L.) Varieties Grown in Nigeria. Int. J. Acad. Res. Reflect. 2015, 3, 1–11.
- P, Nirmala Prasadi V.; Joye, I.J. Dietary Fibre from Whole Grains and Their Benefits on Metabolic Health. Nutrients 2020, 12, 3045. [CrossRef]
- Khosravi, A.; Zarei, M.; Ronaghi, A. Effect of PGPR, Phosphate Sources and Vermicompost on Growth and Nutrients Uptake by Lettuce in a Calcareous Soil. J. Plant Nutr. 2018, 41, 80–89. [CrossRef]
- Li, Q.; Ren, Y.; Fu, H.; Li, Z.; Kong, F.; Yuan, J. Cultivar Differences in Carbon and Nitrogen Accumulation, Balance, and Grain Yield in Maize. Front. Plant Sci. 2022, 13, 992041. [CrossRef]
- Wei, Z.; Ying, H.; Guo, X.; Zhuang, M.; Cui, Z.; Zhang, F. Substitution of Mineral Fertilizer with Organic Fertilizer in Maize Systems: A Meta-Analysis of Reduced Nitrogen and Carbon Emissions. Agronomy 2020, 10, 1149. [CrossRef]
- Wu, X.; Tong, L.; Kang, S.; Du, T.; Ding, R.; Li, S.; Chen, Y. Combination of Suitable Planting Density and Nitrogen Rate for High Yield Maize and Their Source–Sink Relationship in Northwest China. J. Sci. Food Agric. 2023, 103, 5300–5311. [CrossRef]
- Ma, R.; Jiang, C.; Shou, N.; Gao, W.; Yang, X. An Optimized Nitrogen Application Rate and Basal Topdressing Ratio Improves Yield, Quality, and Water- and N-Use Efficiencies for Forage Maize (Zea Mays L.). Agronomy 2023, 13, 181. [CrossRef]
- Cvijanović, V.; Cvijanović, G.; Rajičić, V.; Marinković, J.; Đukić, V.; Bajagić, M.; Đurić, N. Influence of Different Methods of Application of Effective Microorganisms in Nutrition of Wheat on Weight by 1000 Grains, Yield, and Content of Crude Wheat Proteins (TRITICUM SP). Cereal Res. Commun. 2022, 50, 1259–1268. [CrossRef]
- Fathollahi, Z.; Gharavi, K.B.; Gasemi, A. Effects of Combined Use of Bio-Fertilizers and Chemical Fertilizers at Different Stages on Physiological and Morphological Characters of Navy Bean. Indian J. Fundam. Appl. Life Sci. 2014, 4, 423–427.
| Treatment | Main Plot Microorganisms |
Subplot Fertilization (%) |
|---|---|---|
| 1 | Without inoculation (M0) | 0 (0-0-0) * |
| 2 | 50 (120-60-70) * | |
| 3 | 75 (180-90-105) * | |
| 4 | 100 (240-120-140) * | |
| 5 | Bacillus subtilis (B) | 0 |
| 6 | 50 | |
| 7 | 75 | |
| 8 | 100 | |
| 9 | Trichoderma viride (T) | 0 |
| 10 | 50 | |
| 11 | 75 | |
| 12 | 100 | |
| 13 | Pseudomonas putida (P) | 0 |
| 14 | 50 | |
| 15 | 75 | |
| 16 | 100 |
| Fertilization Dose | Ash | Fat | Fiber | Total Energy | kcal Fat | kcal Carbohydrate |
|---|---|---|---|---|---|---|
| (%) | (g∙100 g-1) | (g∙100 g-1) | (g∙100 g-1) | (kcal∙100 g-1) | (kcal∙100 g-1) | (kcal∙100 g-1) |
| 0 | 1.17 ± 0.1 c | 3.46 ± 0.27 | 1.88 ± 0.22 | 359.42 ± 1.68 c | 31.08 ± 2.43 | 284.5±5.5 |
| 50 | 1.24 ± 0.13 b | 3.39 ± 0.37 | 1.85 ± 0.26 | 359.83 ± 1.7 bc | 30.5 ± 3.18 | 289±5.19 |
| 75 | 1.25 ± 0.09 ab | 3.36 ± 0.26 | 1.89 ± 0.19 | 360.75 ± 2.09 ab | 30.25 ± 2.38 | 292.08±2.75 |
| 100 | 1.3 ± 0.11 a | 3.33 ± 0.23 | 1.81 ± 0.2 | 361.17 ± 1.85 a | 29.92 ± 2.19 | 296.42±4.1 |
| Interaction | 0.21 | 0.28 | 0.09 | 0.85 | 0.36 | 0.47 |
| Fertilization Dose | Ash | Fat | Carbohydrates | Total Energy | kcal Protein | kcal Fat | Kcal Carbohydrate |
|---|---|---|---|---|---|---|---|
| (%) | (g∙100 g-1) | (g∙100 g-1) | (g∙100 g-1) | (kcal∙100 g-1) | (kcal∙100 g-1) | (kcal∙100 g-1) | (kcal∙100 g-1) |
| 0 | 1.09 ± 0.09 c | 3.87 ± 0.22 | 72.77 ± 0.59 d | 363.92 ± 1.88 | 30.33 ± 3.23 c | 33.5 ± 2.65 | 290.33 ± 2.23 d |
| 50 | 1.15 ± 0.1 b | 3.78 ± 0.31 | 73.55 ± 0.58 c | 362.67 ± 1.61 | 33.67 ± 1.83 b | 34 ± 2.95 | 294.17 ± 2.33 c |
| 75 | 1.16 ± 0.09 b | 3.9 ± 0.31 | 73.75 ± 0.63 b | 363.42 ± 1.56 | 35.17 ± 2.44 b | 35 ± 2.76 | 295.08 ± 8.36 b |
| 100 | 1.24 ± 0.08 a | 3.87 ± 0.28 | 74.6 ± 0.57 a | 363.08 ± 1.08 | 37.58 ± 2.27 a | 34.83 ± 2.33 | 298.5 ± 2.35 a |
| Interacction | 0.18 | 0.92 | 0.19 | 0.86 | 0.85 | 0.90 | 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



