
Submitted:
06 November 2024
Posted:
07 November 2024
You are already at the latest version
Abstract
Telomerase presents over-expression in most of cancer cells and has been a near-universal marker of cancer. Studies revealed that inhibiting telomerase activity by utilizing oligonucleotides to down-regulate the expression of intracellular human telomerase reverse transcriptase (hTERT) mRNA was an effective method to achieve anti-tumor therapy. Considering oncogenic microRNA-21 has been proved to indirectly up-regulate hTERT expression and drive cancer metastasis and aggression through increased telomerase activity, here we constructed a AS1411-functionallized oligonucleotide-conjugated gold nanoprobe (Au-nanoprobe) to simultaneously down-regulate intracellular microRNA-21 and hTERT mRNA by using anti-sense oligonucleotide technology to explore their targeted anti-tumor therapy effect. In vitro cell studies demonstrated that Au-nanoprobes could effectively induce apoptosis and inhibit the proliferation of cancer cells through down-regulating intracellular hTERT activity. In vivo imaging and anti-tumor studies revealed that Au-nanoprobes could accumulate at the tumor site and inhibit the growth of MCF-7 tumor xenografted on balb/c nude mice, thus exert the potential of anti-tumor therapy.
Keywords:
gene regulation
; microRNA-21
; hTERT mRNA
; gold nanoparticles
; anti-tumor
1. Introduction
Considering the high-expression of telomerase activity in most cancer cells, cancer treatment specifically targeting telomerase has attracted people’s interest [1,2]. As one of the main subunits of telomerase, human telomerase reverse transcriptase catalytic subunit (hTERT) has been proved to be the rate limiting component of telomerase activity and thus become an important target for telomerase regulation [3,4]. Some studies have pointed out that the regulation of hTERT mRNA by anti-sense technology can effectively induce apoptosis of cancer cells [5], and the drug–Imetelstat (GRN163L) based on this principle has long been in clinical trials [6].
Many studies have shown that a variety of microRNAs in cancer cells, such as carcinogenic microRNAs including microRNA-19b, microRNA-346 and microRNA-21, could directly or indirectly up-regulate the expression of hTERT so as to promote cancer invasiveness, oxidative stress, genomic instability, and cell proliferation, as well as evasion of apoptosis [7,8,9]. Among them, microRNA-21 was found to be indirectly associated with hTERT up-regulation in colorectal cancer and malignant melanoma cells by down-regulating PTEN (a tumor suppressor gene) and then activating PI3K/Akt pathway [10]. Also, microRNA-21 was proved to enhance carcinogenesis through STAT3, and reduced expression of both hTERT and STAT3 as well as slowed tumor growth was observed when microRNA-21 was knocked down in murine glioblastoma xenografts [11]. Moreover, if the inhibition of carcinogenic microRNAs is combined with telomerase therapy, the resistance effect in telomerase therapy can be reduced [12]. Therefore, down-regulation of carcinogenic microRNAs and hTERT mRNA by anti-sense oligonucleotide technology will play a positive role in combating cancer. However, the nucleic acid fragment is not easy to enter the cell directly, appropriate carriers are needed to improve the intake rate of anti-sense oligonucleotides.
The structure of tumor vessels is abnormal with wide interendothelial junctions and a large number of fenestration. Therefore, after administered intravenously, nanoparticles easily extravasate through leaky vasculature and accumulate in the tumor [13]. In addition, targeting ligands modified on the nanoparticles surface can specifically recognize tumor and bind to over-expressed receptors with a high affinity in the target region, which would induce nanomedicines extravasating into tumors through active trans-endothelial mechanisms [14]. Among the nanoparticle-based drug delivery systems that have been developed, gold nanoparticles (AuNPs) have become the preferred carrier for researchers [15,16] due to their several advantages of good biosecurity, stability, surface functionization and fluorescence quenching property. Also, AuNPs have been proved to penetrate tumor vascular system and engineering tumoral vascular leakiness and increase tumoral accessibility of anti-tumor therapeutics and subsequently enhancing therapeutic efficiency [17]. Altogether, targeting ligands modified AuNP-based drug delivery systems present great application potential in the field of targeted anti-tumor therapy.
To expand the application of AuNPs in the tumor theranostics, previously we designed two AuNP-based nucleic acid probes loaded with anti-sense oligonucleotide sequences to achieve in situ detection, fluorescence imaging and down-regulation of intracellular hTERT mRNA or microRNA-21 [18,19]. While the effectiveness of AuNP-based nucleic acid probes in inducing cancer cell apoptosis by anti-sense technology has been verified in the cell level, in vivo studies are needed to show their anti-tumor therapy potential. As a continuation of this series of research work, here we designed a gold nanoprobe (Au-nanoprobe) containing two types of anti-sense sequences, in which one type of anti-sense sequence is used for down-regulating hTERT mRNA and the other is designed for silencing microRNA-21. In addition, in order to achieve targeted delivery to tumors and improving cancer cell uptake of nanomedicines, the aptamer AS1411 that recognizes cancer cells through high affinity with the surface over-expressed nucleolin [20] was modified on the surface of Au-nanoprobes with disulfide bonds as linkers. The in vitro and in vivo anti-cancer effect of Au-nanoprobes was investigated here.
2. Materials and Methods
2.1. Preparing Au NPs and Au-Nanoprobes
AuNPs were synthesized according to the literature previously reported [21]. Firstly, 50 ml of HAuCl4 (1 mM) solution was heated to 100 °C. Next, 10 ml of trisodium citrate solution (38.8 mM) is quickly added the above boiled HAuCl4 solution and the mixed solution was stirred thoroughly at 100 °C for 15 min to obtain the dark red AuNPs solution. The size and morphology of AuNPs were characterized by the JEM-2100 transmission electron microscope (JEOL, Japan).
Au-nanoprobes are prepared according to the following procedure. All DNA sequences (1 OD) (Table S1) are individually dissolved in 100 μL DEPC water. HS-anti-hTERT-DNA and Cy3-hTERT-DNA were mixed in the molar ratio of 1: 1.2 and the mixture was heated to 75 °C and hold for 10 min. Then the mixture was naturally cooled to room temperature and incubated under dark conditions for 12 h to obtain the hybridized hTERT-related DNA duplexes (HS-anti-hTERT-DNA/Cy3-hTERT-DNA). With the same procedure, the hybridized microRNA-21-related DNA duplexes (HS-miRNA-21-DNA/Cy5-AS1411-anti-miRNA-21-DNA) was also prepared. Next, the hybridized hTERT- and microRNA-21-related DNA duplexes systems were mixed together and further reacted with 4 mL AuNPs solution. The final mixture were incubated with rotation at room temperature for 24 h and further inactivated with PBS solution (400 μL) thrice with the interval time of 10 h. Then the reaction mixture was centrifuged and the precipitate was washed with PBS solution thrice to discard the unbound DNA sequences. Finally, the newly prepared Au-nanoprobes were dispersed in PBS solution (4 mL) and store at 4 °C for future studies. The ultraviolet visible (UV-Vis) absorption spectrum of Au-nanoprobes were determined and the concentration of Au-nanoprobes was calculated by measuring their extinction at 524 nm (ε = 2.7 × 108 L·mol-1·cm-1).
2.2. Evaluating the Amount of DNA Duplexes Bound on Each Au-Nanoprobe
The amount of HS-anti-hTERT-DNA/Cy3-hTERT-DNA or HS-miRNA-21-DNA/Cy5-AS1411-anti-miRNA-21 duplexes bound on each Au-nanoprobe was evaluated according to the previously reported protocol [22]. Briefly, different concentrations of mercaptoethanol (0, 1, 1.5, 2, 3, 5, and 10 mM) were individually added to the probe solutions (1.5 nM). After incubation overnight with shaking at room temperature, DNA duplexes were gradually released by the competitive binding of mercaptoethanol with AuNPs. Then the released DNA duplexes were separated from AuNPs through centrifugation and the fluorescence intensity of suspension was determined by using the F-7000 spectrofluorometer (Hitachi, Japan) with the excitation wavelengths of 530 and 630 nm. The standard linear calibration curve was prepared with known concentrations of two types of DNA duplexes (0, 10, 20, 40, 60, 80, 100, 120, 150, and 200 nM) with identical buffer pH, ionic strength and mercaptoethanol concentrations. The amount of HS-anti-hTERT-DNA/Cy3-hTERT-DNA or HS-miRNA-21-DNA/Cy5-AS1411-anti-miRNA-21 duplexes bound on each Au-nanoprobe was calculated by referring the fluorescence intensity of the supernatant containing DNA duplexes collected after the prepared probes were incubated with mercaptoethanol (10 mM) to the standard curve.
2.3. Determining the Fluorescence Response of Au-Nanoprobes to Target DNA
In order to verify the responsiveness of Au-nanoprobes to the hTERT mRNA or microRNA-21-related target DNA, a series of Target-hTERT-DNA or Target-miRNA-21-DNA (0, 100, 200, 600, and 1000 nM) were individually mixed with Au-nanoprobes (200 μL, 1.5 nM). All mixtures were incubated at 37 °C for 4 h. Then the fluorescence intensity of different system was determined by using the F-7000 spectrofluorometer (Hitachi, Japan) with the excitation wavelengths of 530 and 630 nm.
2.4. Determining the Fluorescence Response of Au-Nanoprobes to Intracellular MicroRNA-21 and hTERT mRNA
Two telomerase-positive cancer cell lines, including HeLa (human cervical cancer cells) and MCF-7 (human breast cancer cells), were cultured in the DMEM medium (GIBCO) containing fetal bovine serum and Penicillin-Streptomycin (100 µg·ml−1) with the ratio of 9 : 1 : 0.1 at 37 °C in a humidified atmosphere containing 5% CO2. The cell number was determined using the Petroff-Hausser cell counter (USA).
The fluorescence responsiveness of Au-nanoprobes to intracellular microRNA-21 and hTERT mRNA was studied with the cell lysates and living cells, respectively. The experimental process were shown as follows.
(i) Cell lysate analysis: Cell lysates were obtained by breaking down MCF-7 cells (1 × 106) by using ultrasonic disruptor. Au-nanoprobes (1.5 nM) were incubated with the freshly prepared cell extracts at 37 °C for 4 h. The fluorescence intensity of the experimental systems were determined by using the F-7000 spectrofluorometer (Hitachi, Japan) with the excitation wavelengths of 530 and 630 nm.
(ii) In situ fluorescence imaging: MCF-7 or HeLa cells (0.4 mL, 1 × 106 mL-1) were respectively seeded in a 20-mm glass-bottom confocal dish. After 24 h, Au-nanoprobes (1.5 nM) were incubated with cells for 4 h. Then, cells were washed with PBS thrice and observed by LSM880 confocal laser scanning microscopy (CLSM, Zeiss, Germany). The fluorescence signals of Cy3 and Cy5 of Au-nanoprobes responsive to hTERT mRNA and microRNA-21 were excited with the wavelengths of 543 and 633 nm, respectively.
2.5. Analyzing the Intracellular hTERT mRNA and microRNA-21 Level
HeLa and MCF-7 cells (5 × 106) in the logarithmic growth phase were treated with AuNP-probes (1.5 nM) or HS-control-DNA/Control-DNA duplex-functionalized probes (Control-Au-nanoprobes, 1.5 nM) for different times (12, 24, 48, or 72 h). Then, total RNA from the tested cells was extracted using Trizol total RNA isolation reagent (TIANGEN) according to the manufacturer’s instructions. The cDNA was reverse- transcribed using a QuantiNova Reverse Transcription Kit (Qiagen, Duesseldorf, Germany). The reactions were incubated in a thermal cycler for 60 min at 37 °C, 5 min at 95 °C, and then held at 4 °C. Real-time quantitative, reverse-transcription polymerase chain reaction (qRT-PCR) was performed using the QuantStudio™ 5 Real-Time PCR system (Applied Biosystems, USA) with specific microRNA-21 primers from the commercial kit (miScript Primer Assays, Qiagen) and miScript SYBR® Green PCR Kit (Qiagen). Relative level of microRNA-21 was calculated from the quantity of microRNA-21 PCR products and the quantity of RNU6B PCR products and normalized to the expression level in untreated cells using the 2-ΔΔCT method [ΔΔCT = (CTmiRNA-21−CTU6-RNA)experimental group − (CTmiRNA-21−CTU6-RNA)untreated group]. The reaction proceeded as follows: 1 cycle of 95 °C for 15 min, followed by 40 cycles of 94 °C for 15 s, 55 °C for 30 s, and 70 °C for 30 s.
With the similar procedure, the relative level of hTERT mRNA was also determined by using qRT-PCR. The sequences of forward and reverse primers of hTERT and GAPDH are given in Table S1. The reaction proceeded as follows: 1 cycle of 50 °C for 2 min and 1 cycle of 95 °C for 2 min were followed by 40 cycles of 95 °C for 15 s, 55 °C for 15 s, and 72 °C for 1 min.
2.6. Analyzing the Intracellular hTERT Activity
MCF-7 cells (5 × 105 cells/well) were inoculated in 6-well plates and cultured for 24 h. Then all cell samples were divided into three groups and were incubated with PBS, Control-Au-nanoprobes (1.5 nM), or Au-nanoprobes (1.5 nM), respectively for 48 or 72 h. After the treatment, hTERT in different cell samples was extracted according to the following procedure. Firstly, 1 × 106 cells were dispensed in a 1.5 mL EP tube, washed thrice with ice-cold PBS (0.1 M, pH 7.4) through centrifugation, and resuspended in ice-cold CHAPS lysis buffer (200 µL) containing 10 mM Tris-HCl, pH 7.5, 1 mM MgCl2, 1 mM EGTA, 0.1 mM PMSF, 0.5% CHAPS and 10% glycerol. The mixture was incubated for 30 min on ice and centrifuged at 16000 rpm at 4 °C for 20 min. The supernatant was collected as cell extract for analysis. To quantify the hTERT activity in different samples, a standard curve was constructed using a commercial hTERT activity ELISA Kit (Shanghai Kepeirui Biotech. Co. Ltd.). The hTERT activity level in different cell extracts was determined according to the procedure given by the ELISA Kit and referring to the standard curve.
2.7. Determining the Pro-Apoptosis Effect and In Vitro Cytotoxicities of Au-Nanoprobes
Pro-apoptosis effect of Au-nanoprobes to MCF-7 cells was investigated by using the AnnexinV-FITC/PI method. MCF-7 (5 × 105 cells/well) were inoculated in 6-well plates. Cells were incubated with PBS (blank group) or Au-nanoprobes (1.5 nM), respectively, for 24, 48, or 72 h. Then, cells were stained according to the procedure given by the commercial Annexin V-FITC/PI apoptosis kit (Beyotime) and collected after the trypsinization treatment for the cell apoptosis analysis determined by Guava easyCyte 5HT flow cytometer (Millipore, USA). The apoptosis data were analyzed by FlowJo v10 software.
The in vitro cytotoxicities of Au-nanoprobes against HeLa and MCF-7 cells were determined by using the MTT method. Firstly, a certain number of cells were inoculated in 96-well plates (1 × 105 cells/well). 24 h later, cells were treated with Au-nanoprobes (1.5 or 2 nM) for 24, 36, 48, or 72 h. For the blank or control groups, cells were treated with PBS or Control-Au-nanoprobes (1.5 or 2 nM), respectively. Then, the cell medium was removed and replaced with 100 μl fresh medium containing 2.5 mg/ml of MTT. 4 h later, 100 μl DMSO was added to dissolve the formazan crystals after the removal of MTT solution. The absorbance at the wavelength of 490 nm was measured with the microplate reader. Cell survival was calculated from subtracting the optical density (OD) value of each well by that of blank group.
2.8. In Vivo and Ex Vivo Fluorescence Imaging
Xenograft tumor models of MCF-7 were built by subcutaneously injecting MCF-7 cells (1×106) in 200 µL Matrigel into the right flank of female balb/c nude mice (3 ~ 4 weeks old). MCF-7 tumor bearing balb/c nude mice with the tumor volume of ~300 mm3 were randomly divided into three groups (n = 3) and fasted 12 h before the experiment with free access to water. Then, biodistribution of Au-nanoprobes was investigated after a single intravenous injection at a dose of 50 μL (6.5 nM). As controls, the other two groups of mice were individually administered with PBS or Control-Au-nanoprobes (50 μL, 6.5 nM). At timed intervals, the mice were anesthetized and then imaged by using the PerkinElmer IVIS Spectrum In Vivo Imaging System for tracking the Cy5 modified anti-miRNA-21-DNA (excitation: 640 nm; emission: 680 nm; epi-illumination). In addition, at 60 min, two representative mice that were treated individually with PBS or Au-nanoprobes, were sacrificed by cervical dislocation and the tumors as well as main organs (hearts, livers, spleens, lungs, kidneys) were excised and imaged by the IVIS Spectrum system.
2.9. In Vivo Anti-Tumor Study
When the tumors reached approximately 100 mm3 (set as Day 0), MCF-7 tumor bearing balb/c nude mice were randomly divided into three groups (n = 5): PBS blank group, Au-nanoprobes group, and Control-Au-nanoprobes group. Next, at different time intervals (Day 0, 2, 4, 6, 8, 10, and 12), mice were injected via tail vein with 50 µL different systems: PBS, Au-nanoprobes (3 nM), and Control-Au-nanoprobes (3 nM). The tumor volumes and the mice body weights were determined synchronously every two days. On day 28, all mice was sacrificed by cervical dislocation. All experiments were carried out in accordance with the National Guide for Care and Use of Laboratory Animals.
2.10. Statistical Analysis
All experiments were performed in triplicate and all data presented as mean and standard deviation. Data were analyzed using IBM SPSS Statistics 25. Values of P < 0.05 and P < 0.01 were considered statistically significant.
3. Results and Discussion
3.1. Design Mechanism of Au-Nanoprobes
The Au-nanoprobes are constructed by combining two types of DNA duplexes on AuNPs through the Au-S bonds (Scheme 1). Among the two types of DNA duplexes, the HS-anti-hTERT-DNA/Cy3-hTERT-DNA duplex is composed of a thiol group modified DNA (HS-anti-hTERT-DNA) in which the part fragment at the 5’ end is designed as the hTERT mRNA anti-sense sequence and a Cy3 labeled DNA (Cy3-hTERT-DNA) that has the same sequence with part fragment of hTERT mRNA [23]. In the other DNA duplexes, HS-miRNA-21-DNA/Cy5-AS1411-anti-miRNA-21-DNA, the thiol group modified DNA (HS-miRNA-21-DNA) is designed having the same sequence with part fragment of microRNA-21 [24]. In Cy5-AS1411-anti-miRNA-21-DNA, the Cy5 is labeled at the 3’ end of microRNA-21 anti-sense sequence and the AS1411 aptamer is linked with the anti-sense sequence through a disulfide bond. When Au-nanoprobes reach the tumor tissue, the high affinity and specificity of AS1411 with nucleolin over-expressed on the cancer cell membrane [25,26] will promote the endocytosis of the probes by cancer cells and enhance the uptake of Au-nanoprobes in tumor tissue. Above the special optical property of AuNPs, the fluorescence signals of Cy3 and Cy5 can be quenched in the Au-nanoprobes [18,19]. Upon encountering target hTERT mRNA and microRNA-21, Cy3-hTERT-DNA and Cy5-AS1411-anti-miRNA-21-DNA will dissociate from Au-nanoprobes because of the competing hybridization between HS-anti-hTERT-DNA and hTERT mRNA as well as Cy5-AS1411-anti-miRNA-21-DNA and microRNA-21 by forming more stable duplexes with longer hybridizing fragments. This will induce the recovery of the fluorescence signals of Cy3 and Cy5 and achieve the in situ monitoring and silencing of intracellular hTERT mRNA and microRNA-21. At the same time, under the action of intracellular glutathione (GSH), the disulfide bond in the Cy5-AS1411-anti-miRNA-21-DNA sequences will be broken. After separating with AS1411, the antisense sequence anti-miRNA-21-DNA can play a better role of silencing microRNA-21. Finally, under the synergism of tumor-targeting delivery and the down-regulation of the expression of intracellular hTERT mRNA and microRNA-21, Au-nanoprobes can exert an effective anti-tumor effects in vivo.
3.2. Characterization of AuNPs and Au-Nanoprobes
The size (~ 13 nm) and the spherical morphology of AuNPs was characterized by transmission electron microscopy (TEM) (Figure S1A). The UV-Vis absorption spectra of both AuNPs and Au-nanoprobes was determined and the results show the characteristic peak of AuNPs at 520 nm in the two spectral curves (Figure S1B). But for Au-nanoprobes, the characteristic absorption peak of nucleic acid at 260 nm is observed. In addition, dynamic light scattering (DLS) was used to verify the hydrodynamic changes after AuNPs were modified with the nucleic acid sequences, which increased from 13.3 ± 0.4 nm (Figure S1C) to 43.0 ± 0.3 nm (Figure S1D). All these results indicate that Au-nanoprobes have been successfully prepared.
3.3. Evaluation of the Amount of DNA Duplexes Bound on Each Au-Nanoprobe
Due to the competition reaction between mercaptoethanol and thiol group modified DNA [22], HS-anti-hTERT-DNA/Cy3-hTERT-DNA or HS-miRNA-21-DNA/Cy5-AS1411-anti-miRNA-21 duplexes bound on AuNPs can be displaced by mercaptoethanol and dissociate from Au-nanoprobes. This will induce the fluorescence recovery of Cy3 and Cy5 in two types of DNA duplexes. The studies revealed that mercaptoethanol with the concentration of 10 mM is enough to replace all DNA sequences bound on AuNPs. Therefore, based on the fluorescence intensity (Figure 1A and B) of the supernatant containing DNA duplexes collected from the incubation solution of Au-nanoprobes and mercaptoethanol (10 mM) and the standard curve of two types of DNA duplexes (Figure S2), the amount of HS-anti-hTERT-DNA/Cy3-hTERT-DNA and HS-miRNA-21-DNA/ Cy5-AS1411-anti-miRNA-21 duplexes on each probe was estimated to be around 47 and 26, respectively.
3.4. In Vitro Response of Au-Nanoprobes to hTERT mRNA and microrna-21-Related Target DNA
In order to play the role of gene silencing, partial fragments of HS-anti-hTERT-DNA and Cy5-AS1411-anti-miRNA-21-DNA in the Au-nanoprobes are designed as the anti-sense sequences of hTERT mRNA and microRNA-21. Therefore, when Au-nanoprobes were incubated with the two target DNA that has the same sequences as microRNA-21 or the key fragment of hTERT mRNA (5’-GGUCGAUUGUGAACAUGGA-3’) (4015 nucleotides; accession no. AF015950) [23] that can be used as gene silence site, Cy3-hTERT-DNA and Cy5-AS1411-anti-miRNA-21-DNA will dissociate from Au-nanoprobes and induce the recovery of the fluorescence signals of Cy3 and Cy5. As shown in Figure 2C and D, with the increase of concentrations of two types of target DNA, the fluorescence intensity of Cy3 and Cy5 correspondingly increased. This indicates more Cy3-hTERT-DNA and Cy5-AS1411-anti-miRNA-21-DNA were released from the Au-nanoprobes due to the competition hybridization between HS-anti-hTERT-DNA and Target-hTERT-DNA as well as Cy5-AS1411-anti-miRNA-21-DNA and Target-miRNA-21-DNA. In addition, the fluorescence intensity of Cy3 (Figure 1E) and Cy5 (Figure 1F) was also significantly increased after the addition of cell extract (curve b). These results demonstrate the successful response of the Au-nanoprobe to intracellular hTERT mRNA and microRNA-21.
3.5. In situ Fluorescence Imaging of Intracellular hTERT mRNA and microRNA-21 by Using Au-Nanoprobes
After validating the response of Au-nanoprobes to two DNA or RNA targets in PBS solution or cell lysate, next by using CLSM we investigated the in situ fluorescence recovery of Au-nanoprobes in two cancer cell lines, HeLa and MCF-7, which highly-express hTERT mRNA and microRNA-21 [18,19,27]. As shown in Figure 2A, after incubated with Au-nanoprobes for 4 h, fluorescence signals of both Cy3 (green) and Cy5 (red) under the excitation of 543 and 633 nm could be observed in two cell lines. This reveals that the hybridization between target RNA (hTERT mRNA and microRNA-21) and Au-nanoprobes happened in two types of cancer cells.
3.6. Analysis of the Expression Level of hTERT mRNA and microRNA-21 as Well as hTERT Activity in Cancer Cells Treated with Au-Nanoprobes
As mentioned above, anti-hTERT-DNA and anti-miRNA-21-DNA in Au-nanoprobes was designed as the anti-sense sequences to reduce the expression of hTERT mRNA and microRNA-21 in cancer cells by using anti-sense oligonucleotide technology. After verifying the responsiveness of Au-nanoprobes to intracellular hTERT mRNA and microRNA-21, next we further evaluate their gene silence efficiency by qRT-PCR technique. The PCR results (Figure 2B and C) indicated that both hTERT mRNA and microRNA-21 were down-regulated in the Au-nanoprobe-treated HeLa and MCF-7 cells, especially for the samples that were treated with the Au-nanoprobes for 72 h. In contrast, there is no obvious down-regulation for two RNA targets when these two types of cancer cells were treated with the Control-Au-nanoprobes. This demonstrates that Au-nanoprobes designed here can effectively down-regulate the expression of intracellular hTERT mRNA and microRNA-21 simultaneously in cancer cells through gene silencing technology. Consistent with this, our studies also revealed that repression of hTERT mRNA and microRNA-21 could significantly inhibit hTERT expression in MCF-7 cells (Figure 2D), which has been proved to be caused by the regulation of STATS or PTEN expression [11,28].
3.7. Studies of the Pro-Apoptosis Effect and In Vitro Cytotoxicity of Au-Nanoprobes
As hTERT expression is tightly associated with telomerase activity which is critical to the growth of cancer cells, down-regulation of hTERT activity would induce apoptosis and inhibit the proliferation of cancer cells. Previously, we have investigated the pro-apoptosis effect of AuNP-based nucleic acid probes through down-regulation of microRNA-21 or hTERT mRNA individually [18,19] by using anti-sense oligonucleotide technology. However, when relying solely on single regulation of microRNA-21 or hTERT mRNA, its ability to inhibit cell proliferation is weak. The experimental results showed that after down-regulating hTERT mRNA in HeLa cells with AuNP-based nucleic acid probes for 72 h, the cell proliferation inhibition rate was only 28% [18]. Similarly, when only down-regulating microRNA-21 in MCF-7 cells, the inhibition rate of cell proliferation was just 20% [19]. To further deepen the research system, here we investigated the cellular apoptosis under the circumstances of simultaneously reducing the expression of microRNA-21 and hTERT mRNA. The flow cytometric analysis revealed that the apoptosis rate of cells treated by Au-nanoprobes gradually increases with the prolong of action time (34.54% at 24 h, 44.12% at 48 h, and 45.68% at 72 h) (Figure 3A and B). Also, this pro-apoptosis efficacy surpass that caused by solely down-regulating microRNA-21 or hTERT mRNA with inhibitors (10 ~ 20%) [11,28].
Next, we further investigated the in vitro anticancer activity of Au-nanoprobes with MTT method. The studies indicate that Au-nanoprobes significantly inhibited the proliferation of HeLa and MCF-7 cells in a concentration- and time-dependent manner. After 72 h of interaction with 2 nM Au-nanoprobes, the proliferation inhibition rates of HeLa and MCF-7 cells reached 43% and 57% (Figure 3C and D), respectively, which were significantly higher than the cell proliferation inhibition rates achieved by solely down-regulating microRNA-21 or hTERT mRNA [18,19]. Comparatively, Control-Au-nanoprobes did not present obvious in vitro cytotoxicities against HeLa and MCF-7 cells in the absence of effective gene regulation. Combining the above gene level and hTERT activity analysis, we can conclude that simultaneously decreased microRNA-21 and hTERT mRNA level caused by Au-nanoprobes through anti-sense oligonucleotide technology could lead to a more significant decrease in hTERT activity, thus more effectively inducing cancer cell apoptosis and exerting anti-proliferative effects.
3.8. In Vivo and Ex Vivo Imaging of Au-Nanoprobes
Numerous studies have shown that the enhanced permeability and retention effect (EPR) related to nanostructures has a very limited impact on the targeted delivery of drugs to tumors. Therefore, researchers have turned to active targeted delivery by binding targeting ligands on the surface of nanomedicines. Among them, DNA aptamer-modified nanostructures as a tumor targeted drug delivery platform [29,30] have been widely reported. Among these researches, high binding affinity and specificity of the aptamer AS1411 with necleolin that is highly expressed on the surface of cancer cells have been widely studied to achieve targeted anti-tumor therapy [31,32]. In this work, to endow Au-nanoprobes with tumor targeting, AS1411 with the sequence of 5′-d(GGT GGT GGT GGT TGT GGT GGT GGT GGA AAA AAA AA)-3′ has been designed to link with anti-miRNA-21-DNA through a disulfide bond. This will realize the tumor targeted delivery and improve the cancer cell uptake rate of Au-nanoprobes. Additionally, besides the passive and active tumor targeting delivery effect of AS1411 modified nanomedicines [33,34], AuNPs themselves have been proved to break through the tumor vascular basement membrane and cross the endothelial barrier through endothelial leakiness to improve nanotherapeutic delivery to tumors [35]. Therefore, Au-nanoprobes designed here should access the tumor to exert the therapeutic outcome. To verify this speculation, next we assessed the biodistribution of the Au-nanoprobes after a single intravenous injection into MCF-7 tumor-bearing mice by utilizing in vivo fluorescence imaging. Considering the limit of weak penetration of Cy3 fluorescence, the fluorescence signal of Cy5 was used to perform the in vivo imaging for tracking the response of Au-nanoprobes to microRNA-21 in vivo and the biodistribution of Cy5-labeled anti-miRNA-21-DNA sequences. The results showed that after intraveneous administration of Au-nanoprobes in MCF-7 tumor bearing mice, the Cy5 fluorescence signal was quickly observed at the abdomen and peritumoral sites within 10 min and the intensity increased over a period of 40 min (Figure 4A and B). Also, the obvious Cy5 fluorescence appeared at the tumor sites in a period of 15 ~ 30 min. Furthermore, ex vivo fluorescence imaging of the main organs and tumors dissected from MCF-7 tumor-bearing mice after 60 min of administration showed bright fluorescence signal in the liver, kidney and tumor (Figure 4C). All these results demonstrate the effective accumulation of Au-nanoprobes at the tumor site and then the residues can be excreted through the liver and kidney.
3.9. In Vivo Targeted Anti-Tumor Therapy
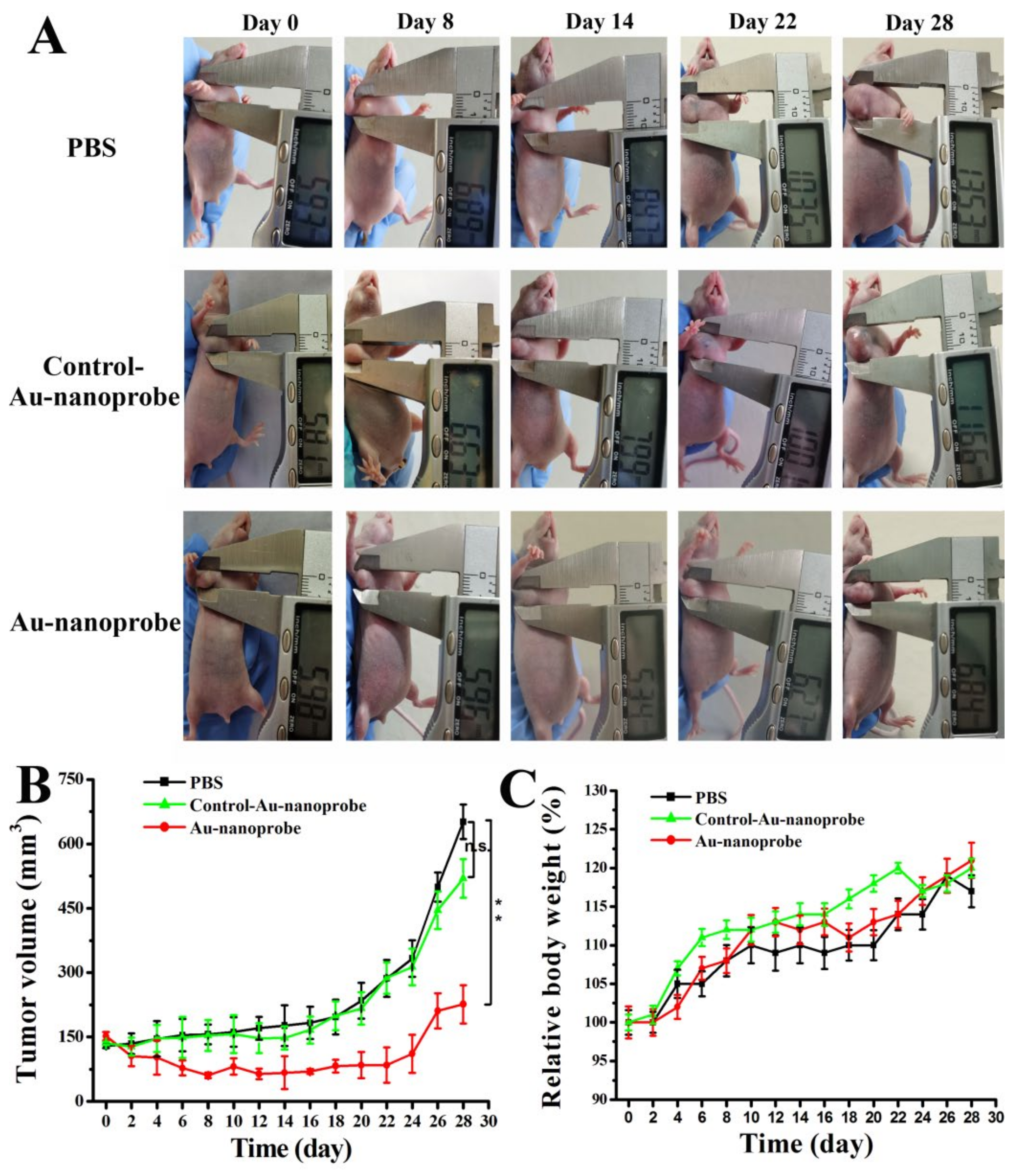
Encouraged by the above impressive in vitro pro-apoptosis and tumor targeting results of Au-nanoprobes, next we further investigated its in vivo anti-tumor therapeutic efficacy with MCF-7 tumor model built by subcutaneously injecting MCF-7 cells into the right flank of balb/c nude mice. Au-nanoprobes at a dosage of 3 nM were intraveneously administered to mice every other day for seven times. The tumor growth rate in terms of average tumor volume was recorded in a period of 29 days. The results showed that MCF-7 tumors in the PBS group grew gradually during the experiments. Similar phenomenon was also observed in the Control-Au-nanoprobe treated groups (Figure 5A and B). Although AuNPs have been reported to be able to induce innate immune responses through the process of immunogenic cell death [36], but obviously, relying solely on this effect has not played an effective anti-tumor role here. Comparatively, the tumor volumes regressed gradually during the Au-nanoprobes treatment and this inhibition state of tumor proliferation could also keep in the following 10 days even after stopping administration. Afterwards, in the absence of the effect of Au-nanoprobes, the tumor volume gradually increased. Thus, we can conclude that Au-nanoprobes could efficiently inhibited the MCF-7 tumor growth and the simultaneous down-regulation of intratumoral microRNA-21 and hTERT mRNA caused by Au-nanoprobes play a leading role in the anti-tumor therapy.
During the experiments, the body weight of all mice were recorded and the results showed that there is no significant difference for the average body weight of all three groups of mice (Figure 5C), indicating that the Au-nanoprobes has no significant toxicity.
4. Conclusions
In conclusion, we constructed a AS1411 aptamer modified tumor-targeted Au-nanoprobe by binding DNA duplexes on the AuNPs with Au-S bonds. Under the synergism of passive and active targeting effect as well as the endothelial leakiness effect attributed to AuNPs, Au-nanoprobes were able to accumulate at the tumor site. The response of Au-nanoprobes to intracellular hTERT mRNA and microRNA-21 visually monitored by confocal fluorescence imaging has caused the simultaneous down-regulation of the expression of hTERT mRNA and microRNA-21 by anti-sense oligonucleotide technology and induced the cell apoptosis. The in vivo experiments demonstrate that Au-nanoprobes could effectively inhibit the tumor growth, showing great potential for the development of AuNP-based nanomedicines associated with telomerase therapy by using gene regulation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: DNA sequences used in this work.; Figure S1: (A) TEM characterization of AuNPs. (B) UV-Vis absorption spectra of AuNPs and Au-nanoprobes. Hydrodynamic size distribution of AuNPs (C) and Au-nanoprobes (D) determined by DLS; Figure S2: The fluorescence spectra of HS-anti-hTERT-DNA/Cy3-hTERT-DNA (A) or HS-miRNA-21-DNA/Cy5-AS1411-anti- miRNA-21 duplexes (C) with the concentrations of 0, 10, 20, 40, 60, 80, 100, 120, 150, and 200 nM. The relationship between the fluorescence intensity and different concentrations of HS-anti-hTERT-DNA/Cy3-hTERT-DNA (B) or HS-miRNA-21-DNA/Cy5-AS1411-anti- miRNA-21 duplexes (D).
Author Contributions
Conceptualization, M.H.; methodology, Q.J. and M.H.; software, Q.Y.; formal analysis, M.O.; investigation, Q.J. and Q.Y.; resources, Q.J.; data curation, M.O.; writing—original draft preparation, Q.J. and M.H.; writing—review and editing, M.H.; visualization, Q.Y.; supervision, M.H.; project administration, M.H.; funding acquisition, M.H. All authors have read and agreed to the published version of the manuscript.
Funding
The work was supported by the Natural Science Foundation of Shandong Province (No. ZR2015BM024), the Tai-Shan Scholar Research Fund of Shandong Province and Research Foundation of Liaocheng University (No. 318012026).
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Berei, J.; Eckburg, A.; Miliavski, E.; Anderson, A.D.; Miller, R.J.; Dein, J.; Giuffre, A.M.; Tang, D.; Deb, S.; Racherla, K.S.; et al. Potential telomere-related pharmacological targets. Curr. Top. Med. Chem. 2020, 20, 458–484. [Google Scholar] [CrossRef]
- Hrdličková, R.; Nehyba, J.; Bargmann, W.; Bose, H.R. Multiple tumor suppressor microRNAs regulate telomerase and TCF7, an important transcriptional regulator of the Wnt pathway. PLoS ONE 2014, 9, e86990–87001. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.D.; Tam, J.; Wu, R.A.; Greber, B.J.; Toso, D.; Nogales, E.; Collins, K. Cryo-EM structure of substrate-bound human telomerase holoenzyme. Nature 2018, 557, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Long, W.; Zeng, Y.X.; Zheng, B.X.; Li, Y.B.; Wang, Y.K.; Chan, K.H.; She, M.T.; Lu, Y.J.; Cao, C.; Wong, W.L. Targeting hTERT Promoter G-Quadruplex DNA Structures with Small-Molecule Ligand to Downregulate hTERT Expression for Triple-Negative Breast Cancer Therapy. J. Med. Chem. 2024, 67, 13363–13382. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, A.; Dein, J.; Berei, J.; Schrank, Z.; Puri, N. Oligonucleotides and microRNAs targeting telomerase subunits in cancer therapy. Cancers (Basel) 2020, 12, 2337–2343. [Google Scholar] [CrossRef]
- Salloum, R.; Hummel, T.R.; Kumar, S.S.; Dorris, K.; Li, S.; Lin, T.; Daryani, V.M.; Stewart, C.F.; Miles, L.; Poussaint, T.Y.; et al. A molecμLar biology and phase II study of imetelstat (GRN163L) in children with recurrent or refractory central nervous system malignancies: a pediatric brain tumor consortium study. J. Neuro-Oncol. 2016, 129, 443–451. [Google Scholar] [CrossRef]
- Ohira, T.; Naohiro, S.; Nakayama, Y.; Osaki, M.; Okada, F.; Oshimura, M.; Kugoh, H. MiR-19b regulates hTERT mRNA expression through targeting PITX1 mRNA in melanoma cells. Sci. Rep. 2015, 5, 8201–8210. [Google Scholar] [CrossRef]
- Song, G.; Wang, R.; Guo, J.; Liu, X.; Wang, F.; Qi, Y.; Wan, H.; Liu, M.; Li, X.; Tang, H. miR-346 and miR-138 competitively regulate hTERT in GRSF1- and AGO2-dependent manners, respectively. Sci. Rep. 2015, 5, 15793–15808. [Google Scholar] [CrossRef]
- Melnik, B.C. MiR-21: an environmental driver of malignant melanoma? J. Transl. Med. 2015, 13, 202–218. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, J.J.; Tao, H.; Jin, W.S. MicroRNA-21 controls hTERT via PTEN in human colorectal cancer cell proliferation. J. Physiol. Biochem. 2015, 71, 59–68. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Sun, G.; Luo, H.; Wang, X.F.; Lan, F.M.; Yue, X.; Fu, L.S.; Pu, P.Y.; Kang, C.S.; Liu, N.; et al. MiR-21 Modulates hTERT Through a STAT3-Dependent Manner on Glioblastoma Cell Growth. CNS Neurosci. Ther. 2012, 18, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Schrank, Z.; Khan, N.; Osude, C.; Singh, S.; Miller, R.J.; Merrick, C.; Mabel, A.; Kuckovic, A.; Puri, N. Oligonucleotides Targeting Telomeres and Telomerase in Cancer. Molecules 2018, 23, 2267. [Google Scholar] [CrossRef] [PubMed]
- Nel, A.; Ruoslahti, E.; Meng, H. New insights into “Permeability” as in the enhanced permeability and retention effect of cancer nanotherapeutics. ACS Nano 2017, 11, 9567–9569. [Google Scholar] [CrossRef] [PubMed]
- Byrne, J.D.; Betancourt, T.; Brannon-Peppas, L. Active targeting schemes for nanoparticle systems in cancer therapeutics. Adv. Drug Deliv. Rev. 2008, 60, 1615–1626. [Google Scholar] [CrossRef]
- Llevot, A.; Astruc, D. Applications of vectorized gold nanoparticles to the diagnosis and therapy of cancer. Chem. Soc. Rev. 2012, 41, 242–57. [Google Scholar] [CrossRef]
- Kyriazi, M.E.; Giust, D.; El-Sagheer, A.H.; Lackie, P.M.; Muskens, O.L.; Brown, T.; Kanaras, A.G. Multiplexed mRNA Sensing and Combinatorial-Targeted Drug Delivery Using DNA-Gold Nanoparticle Dimers. ACS Nano 2018, 12, 3333–3340. [Google Scholar] [CrossRef]
- Setyawati, M.I.; Wang, Q.; Ni, N.; Tee, J.K.; Ariga, K.; Ke, P.C.; Ho, H.K.; Wang, Y.; Leong, D.T. Engineering tumoral vascular leakiness with gold nanoparticles. Nature Comm. 2023, 14, 4269. [Google Scholar] [CrossRef]
- Sun, H.; Hong, M.; Yang, Q.; Li, C.; Zhang, G.; Yue, Q.; Ma, Y.; Li, X.; Li, C.Z. Visualizing the down-regulation of hTERT mRNA expression using gold-nanoflare probes and verifying the correlation with cancer cell apoptosis. Analyst 2019, 144, 2994–3004. [Google Scholar] [CrossRef]
- Hong, M.; Sun, H.; Yang, Q.; Cheng, S.; Yu, S.; Fan, S.; Li, C.; Cui, C.; Tan, W. A microRNA-21-responsive doxorubicin-releasing sticky-flare for synergistic anticancer with silencing of microRNA and chemotherapy. Sci. China Chem. 2021, 64, 1009–1019. [Google Scholar] [CrossRef]
- Mehrnia, S.S.; Hashemi, B.; Mowla, S.J.; Nikkhah, M.; Arbabi, A. Radiosensitization of breast cancer cells using AS1411 aptamer-conjugated gold nanoparticles. Radiat. Oncol. 2021, 6, 33–45. [Google Scholar] [CrossRef]
- Prigodich, A.E.; Seferos, D.S.; Massich, M.D.; Giljohann, D.A.; Lane, B.C.; Mirkin, C.A. Nano-flares for mRNA Regulation and Detection. ACS Nano 2009, 3, 2147–2152. [Google Scholar] [CrossRef] [PubMed]
- Demers, L.M.; Mirkin, C.A.; Mucic, R.C.; Reynolds, R.A.; Leitsinger, R.L.; Viswanadham, G. A fluorescence-based method for determining the surface coverage and hybridization efficiency of thiol-capped oligonucleotides bound to gold thin films and nanoparticles. Anal. Chem. 2000, 72, 5535–5541. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, K.; Fuessel, S.; Schmidt, U.; Kotzsch, M.; Schwenzer, B.; Wirth, M.P.; Meye, A. Antisense-mediated hTERT Inhibition Specifically Reduces the Growth of Human Bladder Cancer Cells. Clin. Cancer Res. 2003, 9, 3794–3800. [Google Scholar] [PubMed]
- Ryoo, S.R.; Lee, J.; Yeo, J.; Na, H.K.; Kim, Y.K.; Jang, H.; Lee, J.H.; Han, S.W.; Lee, Y.; Ki, V.N.; et al. Quantitative and Multiplexed MicroRNA Sensing in Living Cells Based on Peptide Nucleic Acid and Nano Graphene Oxide (PANGO). ACS Nano 2013, 7, 5882–5891. [Google Scholar] [CrossRef]
- Carvalho, J.; Mergny, J.L.; Salgado, G.F.; Queiroz, J.A.; Cruz, C. G-quadruplex, friend or foe: the role of the g-quartet in anticancer strategies. Trends Mol. Med. 2020, 26, 848–861. [Google Scholar] [CrossRef]
- Hosseini, N.F.; Amini, R.; Ramezani, M.; Saidijam, M.; Hashemi, S.M.; Najafi, R. AS1411 aptamer-functionalized exosomes in the targeted delivery of doxorubicin in fighting colorectal cancer. Biomed. Pharmacother. 2022, 155, 113690. [Google Scholar] [CrossRef]
- Hong, M.; Sun, H.; Xu, L.; Yue, Q.; Shen, G.; Li, M.; Tang, B.; Li, C.Z. In situ monitoring of cytoplasmic precursor and mature microRNA using gold nanoparticle and graphene oxide composite probes. Anal. Chim. Acta 2018, 1021, 129–139. [Google Scholar] [CrossRef]
- Zhu, H.Y.; Li, C.; Bai, W.D.; Su, L.L.; Liu, J.Q.; Li, Y.; Shi, J.H.; Cai, W.X.; Bai, X.Z.; Jia, Y.H.; et al. MicroRNA-21 Regulates hTERT via PTEN in Hypertrophic Scar Fibroblasts. PLoS ONE 2014, 9, e97114. [Google Scholar] [CrossRef]
- Ravichandran, G.; Rengan, A.K. Aptamer-Mediated Nanotheranostics for Cancer Treatment: A Review. ACS Appl. Nano Mat. 2020, 3, 9542–9559. [Google Scholar] [CrossRef]
- Berger, C.M.; Gaume, X.; Bouvet, P. The roles of nucleolin subcellular localization in cancer. Biochimie 2015, 113, 78–85. [Google Scholar] [CrossRef]
- Carvalho, L.S.; Gonçalves, N.; Fonseca, N.A.; Moreira, J.N. Cancer stem cells and nucleolin as drivers of carcinogenesis. Pharmaceuticals 2021, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, N.F.; Amini, R.; Ramezani, M.; Saidijam, M.; Hashemi, S.M.; Najaf, R. AS1411 aptamer-functionalized exosomes in the targeted delivery of doxorubicin in fighting colorectal cancer. Biomed. Pharmacother. 2022, 155, 113690. [Google Scholar] [CrossRef] [PubMed]
- Latorre, A.; Posch, C.; Garcimartín, Y.; Celli, A.; Sanlorenzo, M.; Vujic, I.; Ma, J.; Zekhtser, M.; Rappersberger, K.; Ortiz-Urda, S.; et al. DNA and aptamer stabilized gold nanoparticles for targeted delivery of anticancer therapeutics. Nanoscale 2014, 6, 7436–7442. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Zhang, J.; Gao, J.; Zhang, Z.; Zhu, H.; Wang, D. Gold Nanoparticles in Cancer Theranostics. Front. Bioeng. Biotech. 2021, 9, 647905. [Google Scholar] [CrossRef]
- Wang, Q.; Liang, Q.; Dou, J.; Zhou, H.; Zeng, C.; Pan, H.; Shen, Y.; Li, Q.; Liu, Y.; Leong, D.T.; et al. Breaking through the basement membrane barrier to improve nanotherapeutic delivery to tumours. Nature Nanotech. 2024, 19, 95–105. [Google Scholar] [CrossRef]
- Chen, X.Y.; Yung, L.Y.L.; Tan, P.H.; Bay, B.H. Harnessing the Immunogenic Potential of Gold Nanoparticle-Based Platforms as a Therapeutic Strategy in Breast Cancer Immunotherapy: A Mini Review. Front. Immun. 2022, 13, 865554. [Google Scholar] [CrossRef]
Scheme 1.
Illustration of the construction of Au-nanoprobes, the pro-apoptosis and targeted anti-tumor mechanism that is involved with the simultaneous down-regulation of intracellular microRNA-21 and hTERT mRNA through anti-sense technology.
Scheme 1.
Illustration of the construction of Au-nanoprobes, the pro-apoptosis and targeted anti-tumor mechanism that is involved with the simultaneous down-regulation of intracellular microRNA-21 and hTERT mRNA through anti-sense technology.

Figure 1.
The fluorescence recovery phenomena of Cy3 (A) and Cy5 (B) after Au-nanoprobes (1.5 nM) being treated by different concentrations of mercaptoethanol (a to g: 0, 1, 1.5, 2, 3, 5, and 10 mM). Fluorescence spectra of Au-nanoprobes after incubating with different concentrations of (C) Target-hTERT-DNA (0, 100, 200, 600, and 1000 nM), (D) Target-miRNA-21-DNA (0, 100, 200, 600, and 1000 nM), or (E and F) cell lysate.
Figure 1.
The fluorescence recovery phenomena of Cy3 (A) and Cy5 (B) after Au-nanoprobes (1.5 nM) being treated by different concentrations of mercaptoethanol (a to g: 0, 1, 1.5, 2, 3, 5, and 10 mM). Fluorescence spectra of Au-nanoprobes after incubating with different concentrations of (C) Target-hTERT-DNA (0, 100, 200, 600, and 1000 nM), (D) Target-miRNA-21-DNA (0, 100, 200, 600, and 1000 nM), or (E and F) cell lysate.
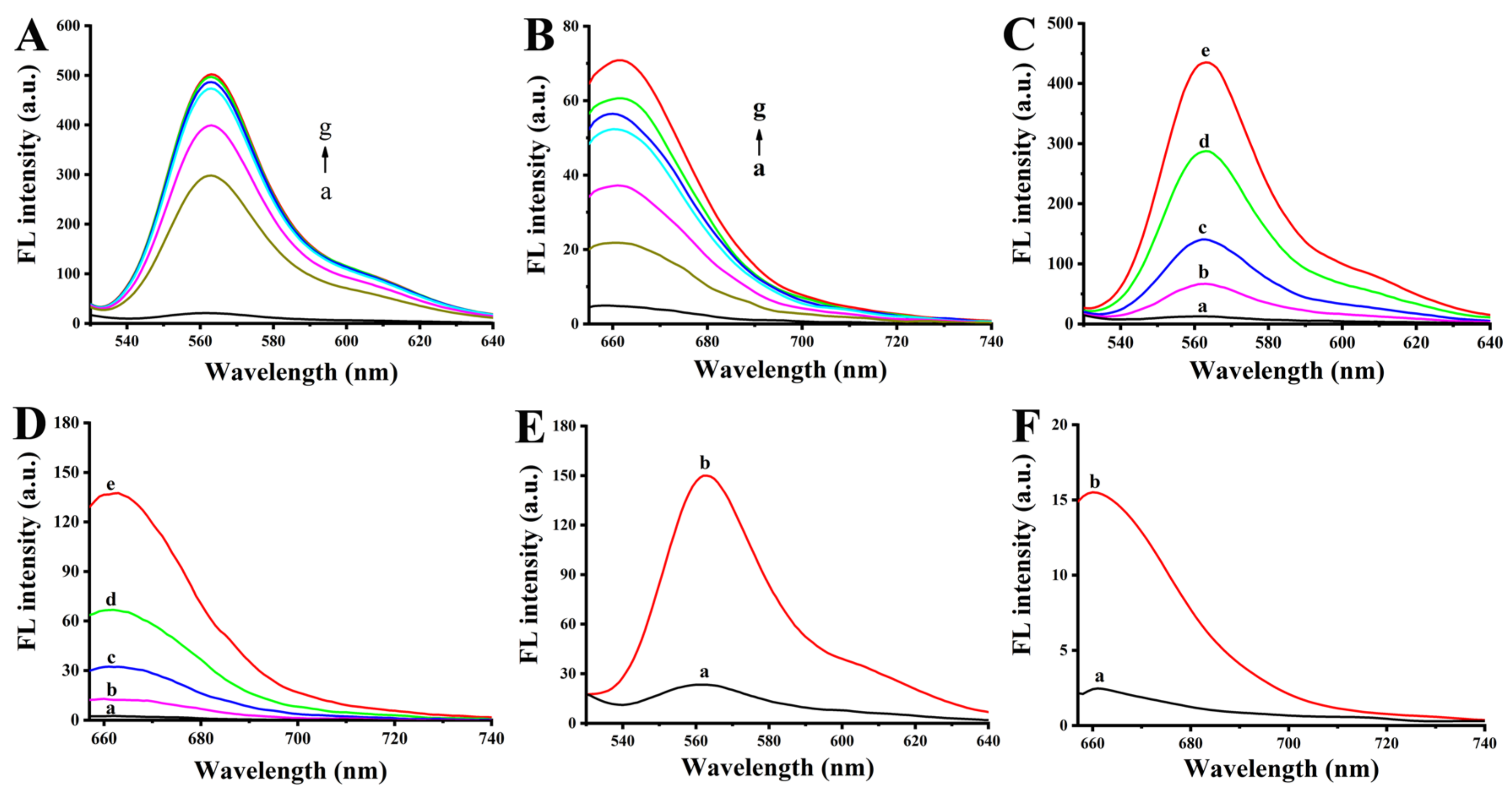
Figure 2.
(A) CLSM imaging of HeLa and MCF-7 cells after incubating with Au-nanoprobes (1.5 nM) for 4 h. Relative expression level of microRNA-21 (B) and hTERT mRNA (C) in HeLa and MCF-7 cells after treated with Au-nanoprobes or Control-Au-nanoprobes for different times. (D) hTERT activity in MCF-7 cells after treated by PBS, Au-nanoprobes or Control-Au-nanoprobes for 48 or 72 h.
Figure 2.
(A) CLSM imaging of HeLa and MCF-7 cells after incubating with Au-nanoprobes (1.5 nM) for 4 h. Relative expression level of microRNA-21 (B) and hTERT mRNA (C) in HeLa and MCF-7 cells after treated with Au-nanoprobes or Control-Au-nanoprobes for different times. (D) hTERT activity in MCF-7 cells after treated by PBS, Au-nanoprobes or Control-Au-nanoprobes for 48 or 72 h.
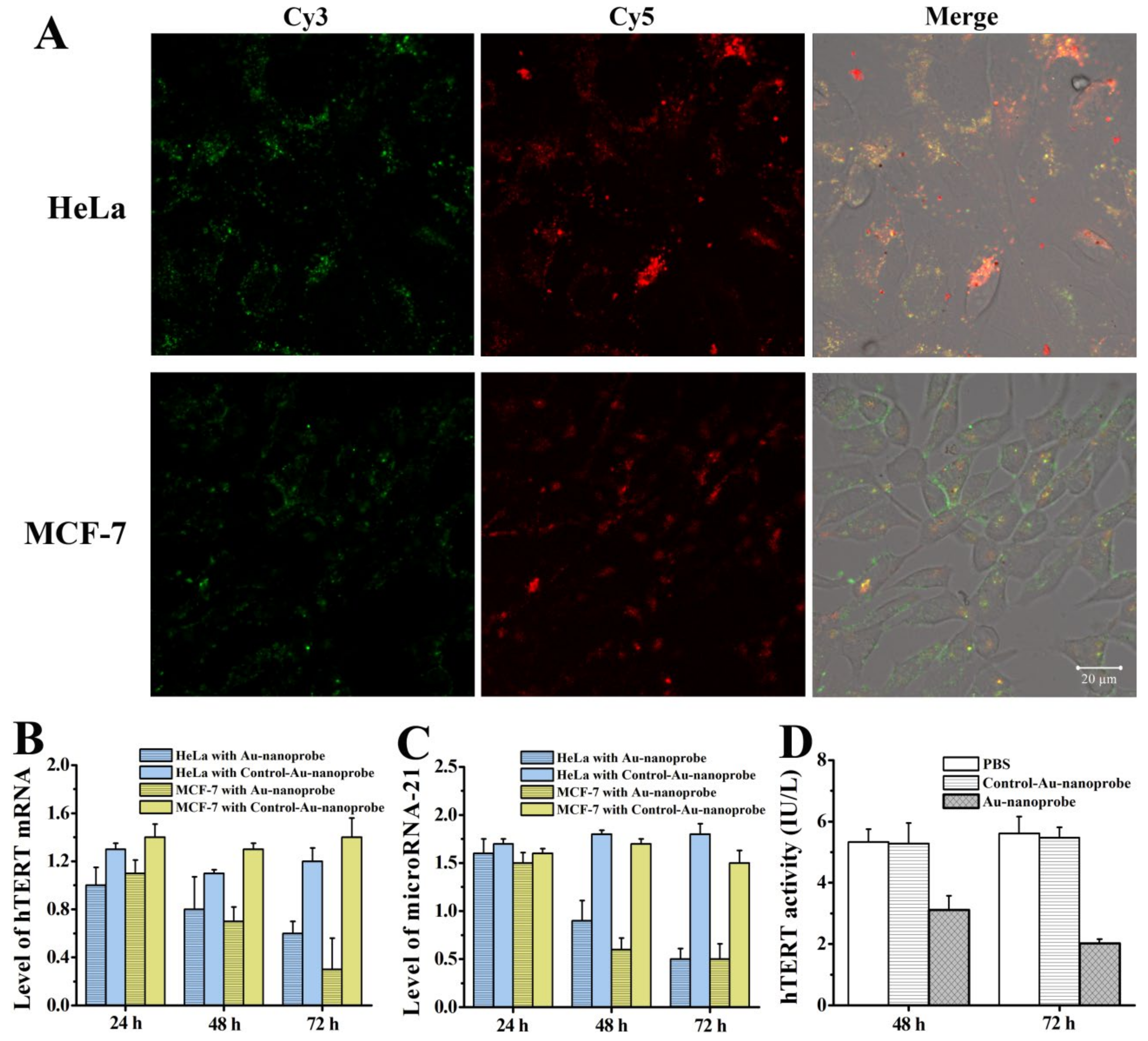
Figure 3.
(A) Apoptosis analysis of MCF-7 cells treated with Au-nanoprobes (1.5 nM) for different times by flow cytometry. (B) Early and late apoptosis rate of MCF-7 shown in (A). Cell viabilities of (C) HeLa and (D) MCF-7 cells after treated with Au-nanoprobes (1.5 or 2.0 nM) or Control-Au-nanoprobes (1.5 or 2.0 nM) for different times (24, 36, 48, or 72 h).
Figure 3.
(A) Apoptosis analysis of MCF-7 cells treated with Au-nanoprobes (1.5 nM) for different times by flow cytometry. (B) Early and late apoptosis rate of MCF-7 shown in (A). Cell viabilities of (C) HeLa and (D) MCF-7 cells after treated with Au-nanoprobes (1.5 or 2.0 nM) or Control-Au-nanoprobes (1.5 or 2.0 nM) for different times (24, 36, 48, or 72 h).
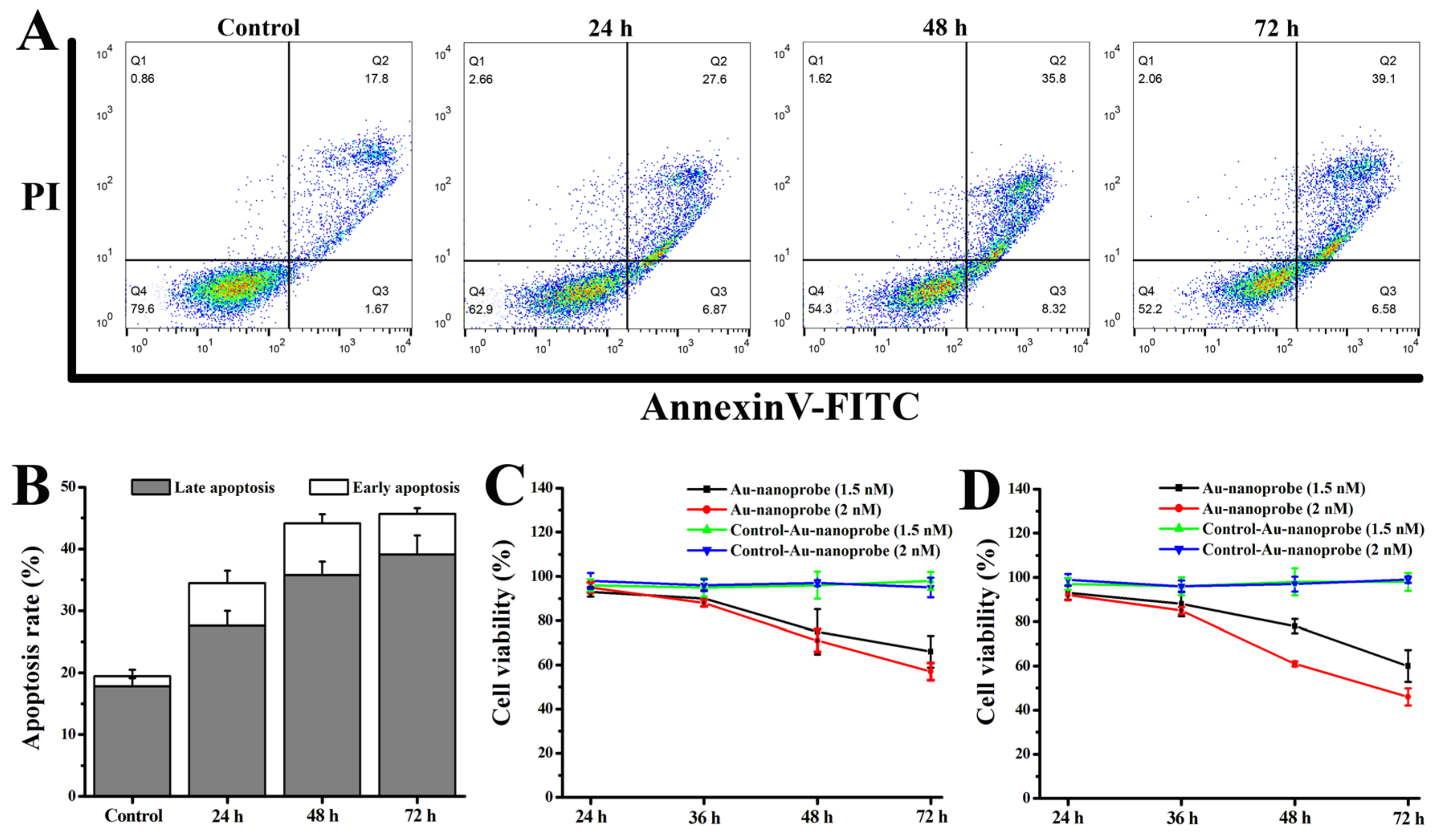
Figure 4.
(A and B) In vivo real-time fluorescence imaging of balb/c nude mice after intravenous administration of PBS, Control-Au-nanoprobes (6.5 nM) or Au-nanoprobes (6.5 nM). (C) Ex vivo fluorescence imaging of the main organs and tumors dissected from the representative balb/c nude mice after oral administration of PBS or Au-nanoprobes for 60 min shown in panel (B).
Figure 4.
(A and B) In vivo real-time fluorescence imaging of balb/c nude mice after intravenous administration of PBS, Control-Au-nanoprobes (6.5 nM) or Au-nanoprobes (6.5 nM). (C) Ex vivo fluorescence imaging of the main organs and tumors dissected from the representative balb/c nude mice after oral administration of PBS or Au-nanoprobes for 60 min shown in panel (B).
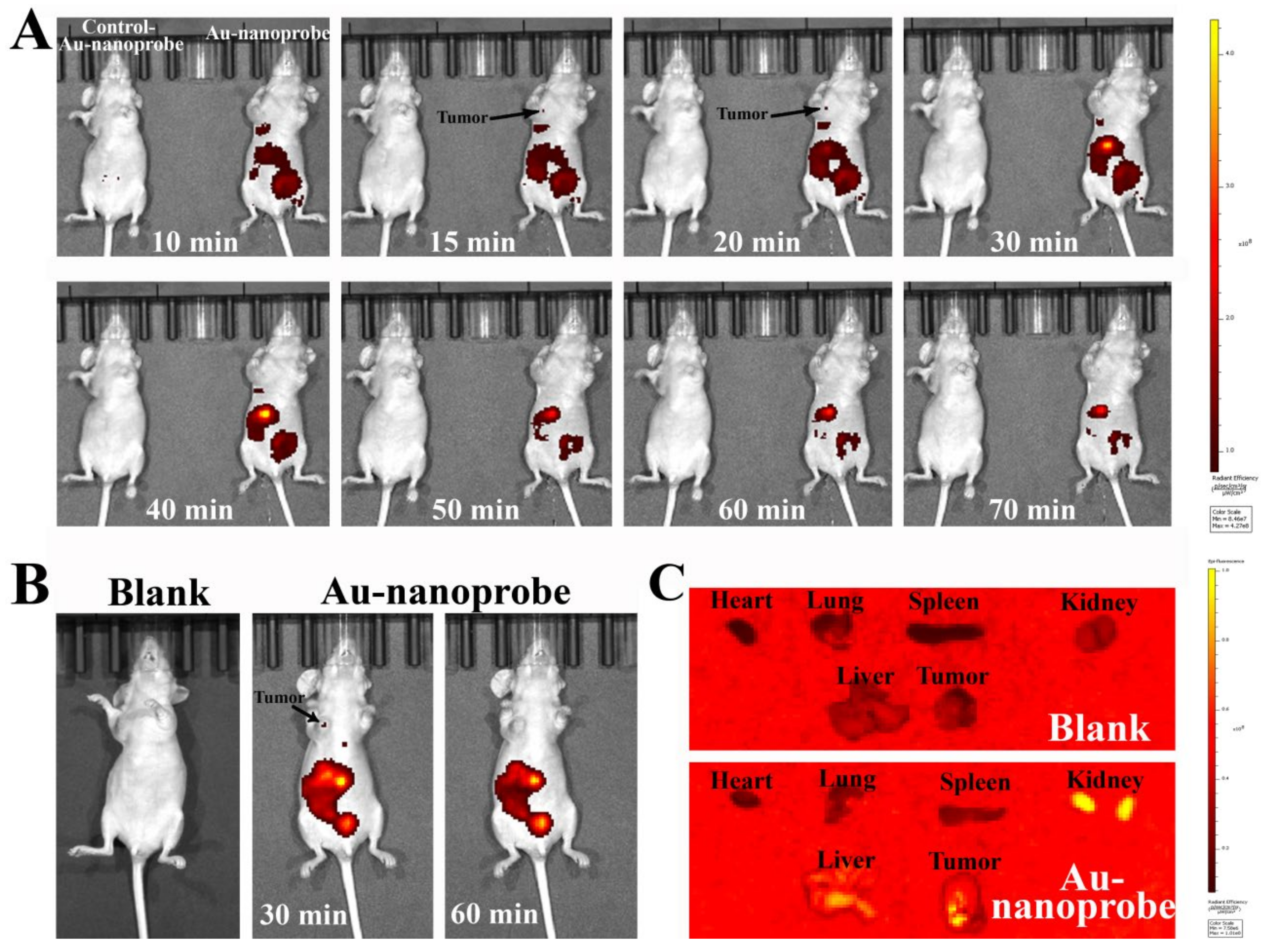
Figure 5.
Anti-tumor study of Au-nanoprobes against subcutaneous MCF-7 xenograft mouse tumors: (A) representative tumor-bearing mouse images, (B) tumor growth curve, and (C) averaged tumor weight of each group during the experiments. During the first 12 days of the experiment, Au-nanoprobes at a dosage of 3 nM (50 μL) were intraveneously administered to mice every other day for seven times. (n.s. P > 0.05, *P < 0.05, **P < 0.01.).
Figure 5.
Anti-tumor study of Au-nanoprobes against subcutaneous MCF-7 xenograft mouse tumors: (A) representative tumor-bearing mouse images, (B) tumor growth curve, and (C) averaged tumor weight of each group during the experiments. During the first 12 days of the experiment, Au-nanoprobes at a dosage of 3 nM (50 μL) were intraveneously administered to mice every other day for seven times. (n.s. P > 0.05, *P < 0.05, **P < 0.01.).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



