
Submitted:
06 November 2024
Posted:
08 November 2024
You are already at the latest version
Abstract
Poultry production system is usually exposed to important infections that could be prevented by vaccination programs. Conventional methods of vaccination such as drinking water, spray, eye or nose inoculation, and injection are usually given after hatching and have many disadvantages. Therefore, there is a great need for searching of alternative ways for vaccination process. In ovo vaccination technology is now regarded as an alternative approach to post-hatch vaccination in modern poultry operations. This technique is effective, fast, provides uniform vaccine dose and delivery, suitable for a massive production, and reduces the labor costs. Routine in ovo vaccination is applied during the late stage of embryonic development between days 17.5 and 19.25 of egg incubation. The best route of inoculation of the vaccine is in the amniotic fluid or in the embryo’s muscles without causing any hatchability or chick quality losses. Accordingly, the inoculation site, the age of the embryos and breeders, presence of maternal antibodies, and the sanitation of equipment’s and the environment during the vaccination process affect the efficiency of the in ovo vaccination technique. In ovo vaccination technology is currently applied for vaccination against several economically important viral diseases such as Newcastle, infectious bursal disease, Marek’s disease, infectious laryngotracheitis, infectious bronchitis, avian influenza, and avian metapneumovirus. Moreover, vaccines used for prevention of mycoplasmosis and coccidiosis could be applied in ovo instead of post hatching application. It could be concluded that in ovo vaccination is a rapidly growing trend of vaccine technology and it can replace the post-hatching vaccination conventional methods.
Keywords:
bacterial
; in ovo vaccination
; parasitic
; poultry
; technique
; viral
1. Introduction
Poultry industry is exposed to variety of stressors and disease conditions that drastically affect the production. Therefore, vaccination of birds is considered a way for reducing the diseases severity, enhancing the bird’s health, protecting the poultry industry, and providing the consumers with safe meat and eggs [1]. The conventional post-hatch vaccination techniques including drinking water, spray, injection, or feed may result in vaccine delivery failure and poor vaccine efficacy [2]. Thus, these techniques, where possible, are being replaced by in ovo vaccination technology which is more effective and faster and provides a more uniform virus vaccine dose and delivery [3]. Besides, this technique is suitable for a large-scale or massive poultry production system with a reduction in labor costs and worker’s involvement [4].
The in ovo vaccination technology was introduced over three decades ago, however, it is currently used in more than 90% of the commercial hatcheries world-wide including United States of America, Europe, and Asia [5]. The first description of this technology was in 1982 as an efficient method of vaccination against Marek’s disease (MD) [6]. The efficiency and success of this way of vaccination may be influenced by many factors including the timing of the inoculation relative to the stage of embryonic development and the precise site of vaccine inoculation in the developing embryo. Routine in ovo vaccination is usually applied between days 17.5 and 19.25 of egg incubation with a direct inoculation in the amniotic fluid or in the embryo’s body without causing any hatchability or chick quality losses [7]. In comparison with other post-hatch vaccination techniques, vaccination at day 18 of embryogenesis reduces the time between early exposure to infectious agents and vaccination [4], which helps in stimulation of an early chick’s resistance and a good protection [8]. The in ovo vaccination technique, particularly for living poultry vaccines, has proven successful for preventing of different economically significant viral [9,10,11,12,13,14,15,16,17], bacterial [18,19,20], and parasitic [21,22] diseases.
In this context, this review article was designed to spotlight on in ovo vaccination technology regarding the mechanism, affecting factors, advantages, and the role of this trend in controlling some important viral, bacterial, and parasitic diseases of poultry.
2. Mechanism of In Ovo Inoculation Technology
The technique used for the administration of in ovo vaccines is critical. Delivering the vaccine in an incorrect location may result in ineffective vaccination, thus reducing its benefits with severe losses. The in ovo vaccination is applied when the incubated eggs are transformed from the setter to the hatchery between 17-19 days of incubation. At the top of the blunt end of the egg shell, a hole is pierced into the eggshell, and then the egg membrane is punctured using the inoculated needle [23,24]. The vaccine should be injected slightly beneath the membranes at the bottom of the air cell [25,26]. Sometimes, the embryo’s position in eggs is not equally the same due to the individual congenital differences, hence, the injection location may be different [27]. During vaccination, the embryo should be in hatching position where the head under the right wing and the yolk sac should be completely absorbed. However, during the direct puncturing of egg membranes, embryo may be injured causing reduced hatchability, survival rate, and chick’s health. The electromagnetic needle-free in ovo injection automation system has been successfully used for accurate inoculation of embryos in eggs [24]. Vaccines should be deposited into the extra-embryonic sacs mainly into the amniotic sac, however, they could be also inoculated in the breast, neck, or shoulder of the embryo [28]. The deposition of the inoculated vaccine in the embryonic tissues or in the extra-embryonic compartment depends on the duration of eggs incubation and the needle size.
An automatic multiple-head egg injection vaccination system has been distributed in industries world-wide and replaced the conventional method of post-hatch injections at hatchery [29]. In this technique, the liquid jet pressure can be adjusted to the target embryonic tissue, thus, it replaces the traditional needle-type and in consequence reduces embryo injury and cross-infection, as well as, it prevents replacing needles and needles cleaning steps. The egg injection system permits inoculation of 20.000-30.000 eggs per hour [25] with inoculation dose of 50-200 microliter of the vaccine per egg [29]. Leaving a hole in the eggs for few days before hatching along with poor sanitized hatcheries enhance the poor viability of hatched chicks due to bacterial or fungal contamination [29]. Hence, the needles should be sterilized after the completion of inoculation and before the following inoculation to avoid cross-infection of inoculated embryos [19]. Following vaccination, the embryo swallows the dissolved vaccine in the surrounding fluids and the gut associated lymphoid tissues begin to react with the antigen of the vaccine [30,31]. Moreover, the embryo’s choanal cleft can aspirate the vaccine into the nasal cavity and the respiratory tract [28], where the vaccine antigen can stimulate the nasal and bronchial associated lymphoid tissues. The cloacal sucking of the inoculated substances was also detected [5].
3. Factors Affecting In Ovo Vaccination Technology
3.1. The Inoculation Site
The inoculation site of the egg differs according to the type of vaccine [27,32]. A precise and an accurate inoculation site of eggs are required to obtain an optimal response by the embryo [33].
The air cell, amnion, allantois, yolk sac, and embryo’s body are the sites where the vaccine could be injected. The selection of the inoculation site may affected by the survival of embryos and their response to the vaccine [3]. It has been reported that vaccine inoculation in the amnion could be absorbed by the embryo prior to hatching [5]. For instance, vaccine deposition into the air cell or allantois didn’t provide adequate protection of chicks against the subsequent MD virus (MDV) challenge [33], while the amnion inoculation provided the highest vaccine effectiveness [34]. Moreover, the injection might be made directly into the right breast area of the embryo when embryo around 19 days of incubation. Breast inoculation may be considered usual and acceptable, however, injection of another site of the embryos body can result in either lack of vaccine efficacy or damage to the embryos [7]. The protection rates against MDV vaccines were 0%, 28.3%, 94.4%, and 93.9% following inoculation in the air cell, allantoic fluid, amniotic fluid, and embryo’s body, respectively [11]. Furthermore, Alqhtani et al. [20] demonstrated the ts-11 Mycoplasma gallisepticum (MG) vaccine strain gave better protection rate after injection in the amnion (97.2%) when compared with intramuscular injection (2.8%) of the embryos. Intracranial, intraorbital, or intraabdominal inoculations of embryos are regarded as unsatisfactory and ineffective methods and perhaps cause embryonic death [11,35]. Besides, it has been found that the location of vaccine deposition in eggs can have potential consequences on the immune response of the hatched chick [11,36].
3.2. The Age of the Inoculated Embryos
The age of the embryos at the vaccination time plays an important role in ova vaccination success. In other words, the timing of in ovo vaccine inoculation should be in synchrony with the stage of embryonic development to achieve the required vaccine outcome [2]. Moreover, appropriate embryonic development stage allows selection of the optimum size and position of the embryos for the exact and correct vaccine inoculation. Therefore, the late stage of embryogenesis, at approximately 18 days of age, is appropriate due to maturation of systems including the immune organs [37] and the best immune response [22,38,39]. No detectable lesions could be observed when inoculating the vaccine to 17 to 19-day-old embryos or older due to development of protective immunity and resistance as well as supporting the virus replication [28]. Additionally, vaccination at this time has a minimum impact on the hatching rate with as stable the physiological status of embryos. Rapid replication of infectious bursal disease virus (IBDV) and infectious bronchitis virus (IBV) has been detected in embryos tissues when inoculated on the day 18 of development [40-42].
Early in ovo vaccination may increase the risk of contamination which compromises the livability and quality of the chicks post-hatching [43]. When herpesvirus of turkeys (HVT) vaccine was administered in 10 to 12-day-old embryos, the virus was much replicated and caused severe lesions and deaths of embryos [44]. Similarly, the in ovo vaccination of 13-day-old embryos resulted in higher embryonic mortality compared with vaccination of 18-day-old embryos [8]. Reduced hatchability around an average 1% was observed in hatcheries when in ovo vaccination was applied on day 17 of incubation [29]. In a similar study [7], the results indicated that the in ovo vaccination on day 16 of incubation reduced the hatchability in 4.68% in comparison with the other vaccination days. Furthermore, Williams [45] demonstrated that too early inoculation of eggs (before day 17.5) might be very invasive causing embryonic damage and the vaccination process could be overwhelmed by the high microbial challenge during hatch. However, the ideal time for safely inoculation of eggs could be from day 17 and 12-14 hrs of i[40–42ncubation to day 19 and 2-4 hrs of incubation [45]. On the other hands, inoculation of eggs after day 19 of age may induce embryo’s injury and mortalities.
3.3. The Age of Breeders
Breeder’s age may also influence the determination of in ovo vaccination timing [46]. Day-old chicks of old breeders, which were usually subjected to a vigorous vaccination program, presented a lower immunity when compared with chicks of young breeders. However, other studies showed that the antibody titers against IBDV and fowlpox virus (FPV) in day-old chicks and their performance at slaughtering were not affected by the in ovo vaccination time or by the breeder’s age [7].
3.4. The Maternal Immunity
Maternal immunity may show some adverse effects on the in ovo vaccination technology. In the presence of high titers of maternal antibodies in vaccinated chicks, the vaccine couldn’t be able to override immunity with a low vaccination efficiency. For example, maternal immunity can affect the efficiency of in ovo vaccination against certain diseases such as MD. Therefore, developing insensitive vaccines to maternal antibodies or vaccination of chick embryos on a day 18 of development is important to overcome this problem [7].
3.5. Other Factors
4. Advantages of In Ovo Vaccination Technology
The in ovo vaccination technique should be easily applied, achieves vaccine efficiency, enhances embryos health, and ensures chick quality [35]. Furthermore, this approach allows an even and uniform mass vaccination with a fast vaccine delivery, provides a precise inoculation, reduces the stressful conditions on chicks, enhances bird’s welfare, decreases the contamination and the time between hatching and placement on the farm, reduces the labor’s costs, as well as allows application of other biological materials that may influence broiler performance [29,43].
The in ovo embryonic vaccination has a positive effect of on the chick’s immune response. It has been supposed that this technique contributes to reduce the period between early exposure to infectious agent and vaccination [4] which helps in evasion of maternal immunity interference and stimulation of earlier embryonic immunity [7,33]. By the time of hatch, vaccinated chicks develop a more significant degree of protection in the form of innate and adaptive immunity when compared with vaccinated chicks after hatching [4,35].
5. Application of In Ovo Vaccination Technology Against Some Important Diseases of Poultry (Figure 1).
Figure 1.
Application of in ovo vaccination technology against some important diseases of poultry.
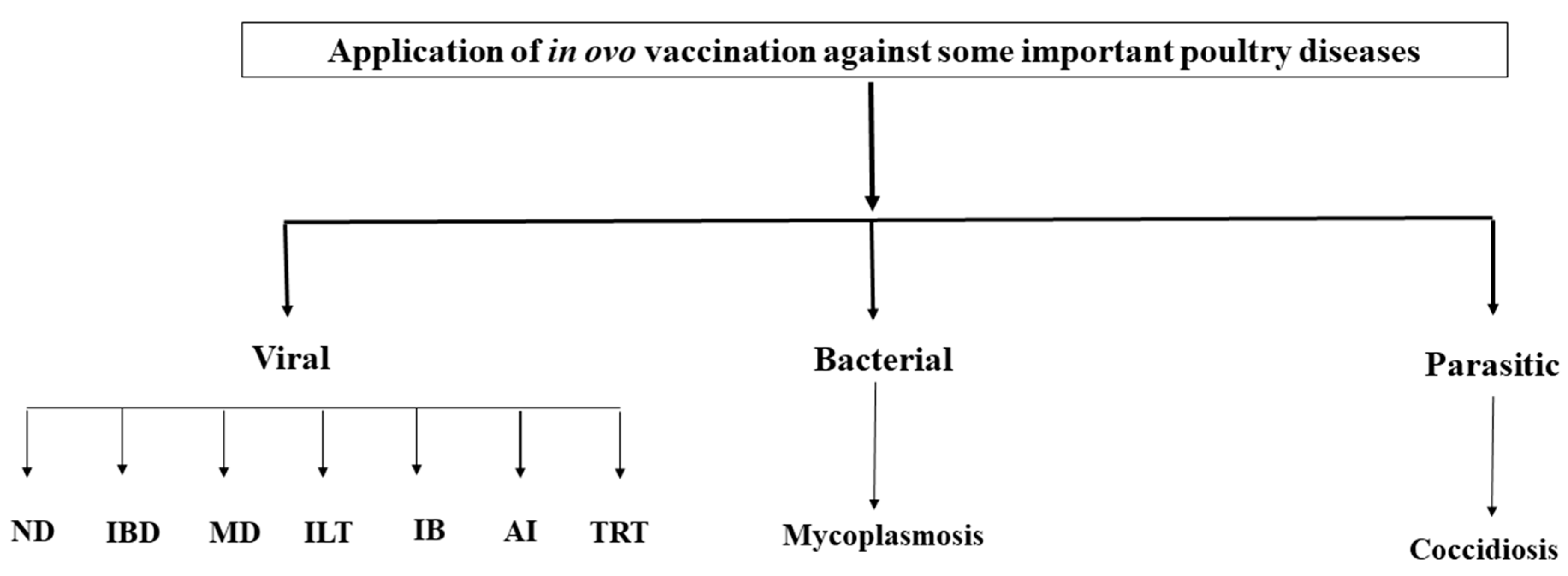
5.1. Viral Diseases
Newcastle Disease
Newcastle disease is a devastating viral infection of most avian species and it caused by avian Orthoavulavirus 1 (previously paramyxovirus 1) [48]. The biosecurity measures and the widespread vaccination programs have been unable to prevent the outbreaks of NDV [49]. Despite application of extensive NDV vaccination protocols with more virulent vaccines particularly in endemic countries, the outbreaks continue to occur in many areas all over the world [50,51,52].
The ideal vaccine against NDV should be able to fill the gap and protects chicks in the first weeks of life. Moreover, it should stimulate both cell-mediated and humoral immunity to reduce the replication of the virulent field viruses, the shedding rate, and the possible horizontal transmission. Living and inactivated vaccines against NDV are commercially available world-wide [53]. Oral or intraocular application of living NDV vaccines provides neutralizing immunity and induces mucosal immunity [54], but, some of them may cause some respiratory manifestations and reduce the productivity [55]. Though inactivated vaccines induce long-lasting immunity [56], they neither replicate in the host nor elicit strong cell-mediated immunity [54].
There are some attends to develop another ways for NDV vaccination. Vaccination against NDV using a living vaccine in ovo faces many limitations and challenges such as high embryos mortality, presence of maternal derived antibodies, and incomplete protection during the early weeks of life [8,57,58,59,60,61]. Low geometric mean titers against NDV vaccine have been observed in specific pathogen free (SPF) chickens vaccinated against IBDV and NDV using in ovo techniques [62]. The best timing of in ova vaccination against NDV is on 18 day of embryogenesis when the last and third wave of thymus colonization occurs [63]. The study of Ohta et al. [64] showed that in ovo administration of aluminum hydroxide-adsorbed attenuated NDV vaccine resulted in improved hatchability and survival rate and better antibody responses of protection-level immunity compared with the administration of NDV alone.
There are about 13 recombinant viral vector vaccines against NDV. The first generation consisted of 2 FPV vectors, followed by 6 second generation, and 5 third generation vaccines incorporating an HVT vector with the fusion (F) protein or the hemagglutinin (HA)-neuraminidase (NA) protein of NDV. Most of these vaccines are administered using in ovo route [65,66]. A recombinant (rFPV) expressing the F and HA-NA glycoproteins of NDV as well as chicken type I or type II interferon (IFN) were applied in the eggs of turkeys [67]. They didn’t affect the hatchability and survival rates, the performance, the weight gain, and the immune response of hatched chicks. A recombinant HVT expressing NDV F protein (rHVT-ND) (Meleagrid alpha herpesvirus 1) has been successfully used for in ovo vaccination against NDV infection in chickens. Despite a single application of rHVT-ND vaccine provided a long-lasting protection against the virulent strain of NDV with no obvious clinical manifestations, is takes up to 4 weeks to mount a protective immune response, can’t be used as a primary vaccine with other rHVT-vectored vaccines, and expensive [68,69,70]. Recently, Marcano et al. [17] reported that in ovo vaccination of 19-day-old embryos with a recombinant live attenuated NDV vaccine containing the antisense sequence of chicken interleukin 4 (IL-4) (rZJ1*L-IL4R) induced the highest degree of protection against NDV challenge without adverse effect on the survival rate of the hatched chicks. However, the antisense RNA may cause transcriptional interference via different mechanisms, so modulating gene expression by reducing or completely silencing a gene is important [71,72]. A recombinant FPV co-expressing type I IFN as well as the NDV HA-NA and F proteins enhanced the protective efficacy of the vaccine against NDV challenge when given in ovo and at hatch [73]. Moreover, a recombinant virus vaccine containing rNDV, rNDV49, and rNDVGu was compared with a living NDV Clone 30 vaccine following in ovo injection [74]. The findings indicated that only one bird from the NDV Clone 30 vaccine group was survived for 3 weeks; whereas, the survival rate of hatched chicks from other groups receiving recombinant NDV vaccine were between 40% and 80%. Moreover, in ovo administration of antigen-antibody complexed live B1-La Sota NDV vaccine provided full immunity against phylogenetically distant virulent viruses at an early age in the presence of maternal immunity [75].
Infectious Bursal Disease
Infectious bursal disease is regarded as an important disease of broiler chickens that caused by Avibirna virus. Due to the resistant nature of the virus, biosecurity measures along with vaccination protocols are routinely used for prevention of IBDV infection.
The classical live attenuated IBDV vaccines are usually applied in the drinking water, while the vector-based and immune complex vaccines are applied either in ovo or subcutaneously. The immune complex vaccines containing live intermediate plus IBDV strains have been mixed with hyper-immune sera to produce virus-antibody complex vaccines. Application of this type of vaccines in ovo provided a good protection against the virulent IBDV challenge even in the presence of high levels of maternal derived antibodies to the virus [76]. In Spain, a single in ovo immunization of broiler chickens with an immune complex IBDV vaccine induced high and uniform protective immunity between 35 and 51 days of age with mean antibody titers values ranging between 6.331 and 7.426 [16]. Combining live IBDV vaccine virus with the immune complex vaccine could be used in ovo or at hatching [77,78], as it can be replicated at the most suitable time and provided active immunity [43,79].
Despite presence of high persistent levels of maternal antibodies can protect chicks during the first weeks of life, they prevented the classical living IBDV vaccines from the active protection following single or repeated applications. Live IBDV strains might be neutralized or break through maternal derived immunity and induced constant damage chick's immune response [80]. In four field trials of Ashash et al. [81], a conventional live intermediate plus IBDV vaccine strain and the IBDV MB-1 were applied in maternally immunized embryos and in one-day-old chicks using subcutaneous route. The findings confirmed that the in ovo application of live IBDV MB-1 vaccine induced a full protection of broilers against a very virulent virus challenge at 22 and 36 days of age as indicated by the bursa lesion score, the q polymerase chain reaction, and the virus genotyping. Furthermore, in ovo vaccination with an intermediate live IBDV vaccine was found to be able to evoke active immune response and protection of 21-day-old SPF chickens as testified by bursal lesions, bursal index after challenge, and vaccine immuno-response [62]. In addition, the study of Lara et al. [82] proved that inoculation of an intermediate live IBDV vaccine strain in 18-day-old embryos or in day-old hatched chicks derived from vaccinated breeders induced no effect on the feed intake, feed conversion ratio, and weight gain till 40 days of age. Hence, it could be concluded that the in ovo vaccination against IBDV didn’t adversely affect hatchability or survival of hatched chicks [83]. The immuno-pathogenesis of IBDV following in ovo vaccination using an intermediate live IBDV vaccine strain was described [84]. The results indicated that inoculation of living IBDV vaccine in ovo induced a faster recovery from the bursal lesions and similar protection against the virus challenge when compared with post-hatch vaccinated chickens. Recently, Zaheer et al. [85] compared immuno-pathological effects of in ovo vaccination with a post-hatch vaccination in white Leghorn chicks and they concluded that the in ovo vaccination with live IBDV vaccines provoked protective immunity even in the presence of IBD-specific maternal immunity.
Some previous studies showed presence of IBDV in the bursa of Fabricius with bursal lesions after in ovo vaccination with a modified live IBDV [86,87]. In ovo vaccination with a classic or Delaware E variant strains of IBDV produced a 6 days delay in the onset of IBDV infection [88]. However, McCarty et al. [89] revealed that in ovo inoculation of modified live IBDV vaccine infected the bursa of Fabricius in maternal-antibody-negative and maternal-antibody-positive eggs. Besides, in ovo vaccination of antibody free broiler chickens with modified live IBDV vaccine induced immunity to post-hatching IBDV exposure [62,90].
The efficacy of the in ovo prime-boost vaccination against IBDV using a DNA vaccine followed by a killed-vaccine boost post hatching, as well as the adjuvant effects of plasmid-encoded chicken IL-2 (ChIL-2) and chicken IFN-γ (ChIFN-γ) in conjunction with the vaccine were tested [91]. The findings revealed that the in ovo DNA vaccination followed by a killed vaccine could completely protect chickens after challenge with very virulent IBDV in terms of absence of mortality and reduction of both bursal damage and lesion score. Besides, the addition of ChIL-2 or ChIFN-γ did not enhance the protective immunity. Both IBDV and MDV vaccines could be jointly administered in ovo without interference with each other [29]. The study of Gagic et al. [92] showed that a single in ovo inoculation of a vaccine containing serotypes 1, 2, and 3 of MDV, serotype 1 of IBDV, and a recombinant FPV vaccine with HA-NA and F genes of NDV (rFP-NDV) produced protection against all these viruses. Furthermore, the inoculation of a vaccine containing serotypes 1, 2, and 3 of MDV and IBDV did not affect hatchability, although the addition of rFP-NDV to the mixture reduced hatchability by 23% to 26%.
It could be summarize that vaccination against IBDV using in ovo inoculation technology decreases the stress of birds handling, induces better immune response, and minimizes the risks of contamination and disease spreading [93].
Marek’s Disease
It has been shown that in ovo vaccination could give greater protection against an early MDV challenge than subcutaneous vaccination at hatching [6]. Since the success of this initial study, the in ovo vaccination technology was developed and distributed world-wide to combat MDV infections [25,29]. However, the exact successful mechanism of in ovo vaccination with HVT has not been fully determined till 2014 [94].
Early infection with MDV and consequently early immune response and protection against infection were obtained after in ovo vaccination [40]. It has been reported that vaccination against MDV on days 17 and 18 of incubation did not interfere with the immune response, decreased the susceptibility of chicks to the onset of disease lesions, and supported the virus replication by the embryos [28]. Similarly, the in ovo vaccination against MDV after 18 days of incubation provoked earlier protection than vaccinations on the first day of life because the vaccine can take 8 days to become effective [95]. Chickens given serotype 1 of MDV (CVI988) during embryogenesis showed better protection against virulent MDV challenge than those given MDV CVI988 at hatch as the virus could be expressed in the embryo to initiate pre-hatch immunologic response [96]. On the other site, Schat [70] reported on immune interference following vaccination with an HVT vaccine in the embryo as antibodies can neutralize a second HVT vector vaccine after hatching. The site of MDV vaccine inoculation in ovo may also affect the protection rate of embryos. In the study of Wakenell et al. [11], the findings showed that when HVT/SB-1 vaccine was injected in the air cell, allantoic fluid, amniotic fluid, and embryo’s body, the protection percentages were 0%, 28.3%, 94.4%, and 93.9%, respectively.
Ricks et al. [29] reported that both IBDV and MDV vaccines could be administered together in ovo with no interference with each other. The in ovo vaccination with a vaccine containing both MDV and IBDV vaccine viruses did not enhance the inhibitory effect of individual viral agents on the humoral and cellular immune competence [92]. In addition, an HVT vector vaccine expressing IL-2 enhanced the protection induced by IBDV and IBV live attenuated vaccines when given via in ovo route [97]. Moreover, the protective efficacy of in ovo vaccination with rHVT against NDV and MDV in chickens was evaluated with successful results [98].
Infectious Laryngotracheitis
Infectious laryngotracheitis virus (ILTV) is taxonomically classified as Gallid herpesvirus 1 [99], and it is associated with expectoration of bloody exudate, gasping, and asphyxia in chickens [100]. The disease usually generates severe production losses as a result of increased mortality, decreased egg production, loss of body weight, and predisposition to other respiratory pathogens [101].
Prevention of ILTV infection is based on biosecurity measures and vaccination using living attenuated virus vaccines produced in emberyonated chicken eggs (ECEs) or tissue culture [102,103,104,105]. However, new generations of recombinant viral vector ILTV vaccines containing FPV and HVT as vectors have been developed [106]. Vector vaccines reduce the possibility of viral transmission from bird to another, beside absence of latent infection after vaccination or reversion to virulence. There are different immunization routes for ILTV vaccines, but the most practical route being in ovo vaccination with HVT, MDV, or the GaHV-1 ‘meq’ gene-deleted vector. Vectored ILTV vaccines are now applied in ovo [107]. It has been documented that in ovo inoculation of recombinant HVT-ILTV was ineffective to break the chain of viral transmission [108]. Though the study of Johnson et al. [109] indicated that the in ovo vaccination with FPV-ILT and HVT-ILT vector vaccines slightly reduced the clinical signs of ILTV, but the vaccine did not significantly reduce the replication of the virus in the trachea. In ovo vaccination with either HVT-ILTV or FPV-ILTV vaccine did not provide as much protection as ECEs derived vaccines which given at 2 weeks of age. Besides, Vagnozzi et al. [107] found that inoculation of the recombinant ILTV vectored vaccines in eggs or subcutaneously at hatching provided more protection and reduction of the severity of clinical signs and virus replication in the trachea when compared with eye drop vaccination of broiler chickens with living attenuated ILTV vaccine at 14 days of age. Moreover, inoculation of HVT-ILTV vaccine in ovo results in inhalation or swallowing the vaccine from the amniotic fluid the embryo and the vaccine can reach to the trachea [11], while injection of the vaccine in day-old chick results in deposition and absorption of the vaccine by the tissues and systematic distribution in the blood. The HVT-ILTV vaccine was more efficient than the FPV-ILTV vaccine in alleviating the disease and reducing the challenge virus level when applied either in ovo or subcutaneously in chicks. Moreover, it has been found that the reactions to FPV-ILTV vector vaccine can be obtained when administered at 17.5 to 18 days of embryonic age rather than at 19 days [110].
The safety and efficacy of a novel “glycoprotein J gene-deleted strain of ILTV” were evaluated in broiler chickens following in ovo vaccination at 18 days of embryo age [111], and the results indicated that this way of vaccination was safe and it did not affect the hatchability or survivability of the hatched chicks. Moreover, vaccination of broilers using either in ovo alone or in ovo combined with eye drop routes revealed significant reduction in signs and the ILTV load post-challenge [111].
The study of Thapa et al. [112] showed that in ovo delivered CpG DNA ILTV vaccine reduced the pre-hatch viral infection in correlation with the expression of IL-1β, increased macrophages in lungs, as well as decreased the mortality and morbidity resulting from ILTV infection post-hatch. Gimeno et al. [113] found that inoculation of rHVT-ILTV vaccine via inoculation of the amnion at day 18 and 19 day of egg incubation resulted in a more virus replication in spleen with a detectable glycoprotein I gene transcripts than subcutaneous inoculation of a day-old chicks.
Infectious Bronchitis
Infectious bronchitis is a highly contagious and an acute upper respiratory tract disease in chickens. The disease is caused by a virus which belongs to the genus corona virus and family coronaviridae [114]. Infection of chickens operations with IBV induces severe economic losses in terms of drop in egg production and egg quality of layers as well as poor weight gain and increased mortalities and condemnation rates of broilers [115]. Prevention of the disease depends mainly on application of both live and inactivated IBV vaccines. However, the emerge of new IBV variant strains [116], the labor, and the costs of vaccine production increase the need of other types of vaccines such as the recombinant DNA types [117].
The in ovo vaccination with IBV vaccines are successfully applied [42,118,119,120,121]. The early study of Wakenell et al. [118] revealed that embryos or day-old chicks which vaccinated with tissue culture serially passaged IBV vaccines had similar lesions in trachea, primary and secondary immune response, and absence of the effector cytotoxic cells to IBV. In addition, Chew et al. [122] studied that effect of inoculation of live IBV vaccine and attenuated kidney cell culture IBV in 18-day-old embryos and the results showed that the hatchability rate was significantly lower in group vaccinated in ovo (27%) than those received attenuated virus (45-58%). Moreover, inoculation of the vaccine and the attenuated IBV in day-old chicks with maternal antibodies induced no pathological lesions in the oviduct and the respiratory signs were seen in 61% and 5% of chicks inoculated with the vaccine in the eggs and at hatch, respectively. In other study [123], hatched chicks from inoculated 17-days-old embryos with 8 different strains of IBV which represented 7 different serotypes showed strict epitheliotropic nature and wide tissue tropism of IBV strains in the chicken embryos and the universality of riboprobe.
Broilers received HVT/SB-1 and HVT plus CVI-988 MDV vaccines either in ovo or at hatch showed better protection rate to IBV challenge than those received IBV vaccines (Ark and Mass serotypes) at hatch [124]. Besides, the MDV vaccines that inoculated either in ovo or at hatch did not affect the efficiency of IBV vaccine given by eye drop at hatch. It has been noticed that in ovo vaccination at 18 days of age with IBV M-41 vaccine could affect the hatchability, therefore using of automatic injectovac system that capable of handling 60.000 eggs at a time could improve the hatching rate [125].
Vaccination of chickens with the DNA vaccine containing the S1 glycoprotein gene from IBV induced a protective immunity against infection [126,127]. Inoculation of this vaccine in ovo or intramuscularly after hatch provided chickens with some protection against clinical signs following homologous virus challenge [120]. Babapoor et al. [117] studied the effect of the in ovo inoculation of a plasmid DNA pTracer-CVM2-IBVS (pCMV-S) vaccine that expresses the immunogenic S glycoprotein genes of IBV serotype Massachusetts 41 and a recombinant chicken interferon type I alpha (rChIFN) as an adjuvant. The results showed that chicks received pCMV-S vaccine had mild signs and were protected at the level of 66%, but those received a combination of the pCMV-S with 2000 IU or 500 IU of rChIFN revealed protection at the levels of 83% and 89% respectively. Furthermore, boostering of pCMV-S and 500 IU rChIFN inoculated chickens with live attenuated IBV vaccine indicated a significant protection (more than 92%) against IBV challenge. In ovo delivered resiquimod, toll-like receptor 7 ligand agonist, enhanced host responses and produced secretory immunoglobulin as well as antigen-specific humoral responses against IBV vaccine [128].
Avian Influenza
Avian influenza virus (AIV) is an enveloped, negative-sense single-stranded segmented RNA virus in the family Orthomyxoviridae [129]. Despite immunization against AIV has been established as one of the most significant control strategy in poultry production system all over the world, AIV has go through antigenic drift due to the presence of immune pressure [130]. Therefore, it is difficult to select a vaccine strain that has the potency to face the widely circulating mutants of AIV.
In ovo inoculation of 18-day-old white rock and white leghorn chicken embryos with oil-emulsion inactivated NDV or AIV H5N9 vaccines induced acceptable hatchability, seroconversion rates, and protective immunity if they are prepared with sufficient antigen and properly administered [131]. Many types of AIV vaccines have been studied for in ovo administration, including; non-replicating adenovirus vectors expressing hemagglutinin proteins [132,133,134], attenuated vaccines [135], and recombinant attenuated vaccines [136]. Chicken embryos received 109 infectious units of an adenovirus vector AIV vaccine showed that less than 1% of hatched chickens displayed serum antibody titers by 10 days post-hatch, while, the percentage rose to 65% and 85% by 20 and 40 days post-hatch, respectively [133]. A previous study showed that in ovo vaccinated chicken embryos with a recombinant attenuated H5N1 AIV vaccine induced humoral immune response to the H5 protein in 30% of the hatched chickens and 80% of vaccinated chickens were survived post-challenge [136]. Moreover, vaccination of 18-days-old embryos with a live AIV H9N2 vaccine produced higher antibody response at young age and showed better protection upon challenge than eggs inoculated with n inactivated oil emulsion AIV vaccine [137].
It has been reported that in ovo administration of CpG ODN decreased the replication of H4N6 AIV and IBV in the inoculated eggs [138,139]. Recently, the immune responses in chickens against a beta-propiolactone inactivated H9N2 AIV vaccine after a primary vaccination on embryonic day 18, and a secondary intramuscular vaccination on day 14 post-hatch were detected [15]. Also, the toll-like receptor 21 ligand, CpG ODN 2007, and an oil emulsion adjuvant were included [15]. The secondary vaccine induced antibody-mediated immune responses and the beta-propiolactone inactivated virus combined with adjuvant provoked cell-mediated immune responses.
Avian Metapneumovirus Infection
Metapneumovirus infection in turkeys is termed as turkey rhinotrachitis in turkeys and swollen head syndrome in chickens.
Vaccination of turkey eggs with avian metapneumovirus subtypes A and B vaccine revealed that the hatched turkey poults showed resistance to the virulent virus challenge [13]. It has been also found that hatched chicks from inoculated eggs with metapneumovirus vaccine showed earlier immunity than day-old vaccinated turkey poults [140]. Inoculation of a living attenuated metapneumovirus vaccine subtype C in the amnion of turkey eggs showed no effect on the hatching rate and the vaccine virus was able to replicate in the upper respiratory tract and the lungs of vaccinated birds, along with low histopathological lesions were seen in vaccinated birds after challenge with virulent strain of avian metapneumovirus at 3-week-old [141]. Moreover, high resistance to the virus replication was observed which was correlated with the low count of the virus in the upper respiratory tract.
5.2. Bacterial diseases
Mycoplasmosis
Mycoplasma gallisepticum (MG) is the main cause chronic respiratory disease of chickens or infectious sinusitis of turkeys with subsequent severe adverse impacts on production worldwide [142]. Affected layers show drastic drop in gg production and broiler show low growth rate [143,144]. Eradication of MG infection could be achieved through vaccination, isolation, and biosecurity measures [145]. Commercially available live MG vaccines include F strain of MG (FMG), 6/85 strain of MG (6/85MG), and ts-11 strain of MG (ts-11MG) have different features and they might be used to effectively control and eradicate field strains of MG. Vaccination of MG is usually applied for pullets before the start of egg production at 9-weeks-old through spray, eye drop, wing web, or intramuscular or subcutaneous ways [146,147].
The effects of the in ovo vaccination with FMG vaccine on layer chicks hatchability [148] and post-hatch survivability, immunity, and horizontal transmission were studied [144]. The results revealed that injection of FMG vaccine in 18-day-old embryonated eggs induced higher embryonic [19,148] and post-hatch [144] mortalities in comparison with control non-injected eggs. The bacterium was detected in the trachea, mouth, esophagus, yolk sac, and duodenum of 22-day-old chick embryos [148]. However, a signification decrease in the growth rate [148] and depression of the humoral immune response [144] were observed in post-hatching chicks till 6 weeks of age. Furthermore, other study showed that the F strain of MG was horizontally transmitted from vaccinated to unvaccinated layer pullets through the direct contact [149]. However, Alqhtani et al. [20] demonstrated that hatched chicks from eggs injected with ts-11 of MG vaccine remained MG free till 21 days post-hatch and the vaccine had no impact either on the hemagglutination inhibition titers or on the post-hatch serology of layers. The timing of the humoral immune response in the layer chicks during the first 4 weeks of age following of in ovo vaccination with FMG was evaluated [150]. The results of the previous study indicated early detection of MG antibodies at 7 days of age. Moreover, male layer chickens were more susceptible to the effects of an in ovo FMG vaccine than females.
5.3. Parasitic diseases
Coccidiosis
Avian coccidiosis is a host, tissue, and immune-specific intestinal protozoon parasitic disease that caused by Eimeria species. The disease is associated with high economic losses including reduced performance parameters, increased mortalities, and high medication costs [151,152]. In addition, coccidiosis is regarded as a major predisposing factor to necrotic enteritis that is caused by the Clostridium perfringens [153]. In the field, prevention strategy of coccidiosis is based on using anticoccidial drugs in feed as well as living vaccines [154,155]. It is important to mention that the vaccine strains of Eimeria are sensitive to anticoccidial drugs.
Live Eimeria oocysts vaccines are applied in the early life of chicks to help in the development of immunity against the wild-type of Eimeria species following oocyst cycling [156]. Live vaccines against coccidiosis could be applied through the in ovo injection of embryos during incubation [157,158]. These vaccines are administered in 18-days-old embryos [159,160]. Living oocysts vaccine containing a mixture of E. acervulina, E. tenella, and two strains of E. maxima was used.
Commercially, the ideal time for in ovo vaccination against coccidiosis is during late embryonic development (between 17.5 and 19.0 days of emberyonation) +14 hrs with a maximum amniotic fluid [3,27]. Moreover, inoculation of embryos with infective Eimeria strains vaccines at 18.0, 18.5, or 19.0 day of emberyonation did not effect on hatchability or chicks quality [2,38,159]. Sokale et al. [22] demonstrated no significant difference in cycling of coccidiosis following vaccination at 18.5 and 19.0 day of incubation.
Following in ovo inoculation of Eimeria oocysts, they absorbed from the amniotic fluid and remained dormant in the embryo’s intestine without occurrence of life cycle till chicks hatching [161]. Weber and Evans [159] reported that the in ovo inoculation of Eimeria species oocysts resulted in a shedding peak at approximately 7 days post-hatch.
6. Conclusion
Nowadays, in ovo vaccination is regarded as an alternative approach to post-hatch conventional vaccination of birds, especially in broiler chickens. It is a rapidly growing trend of vaccine technology. Currently, in ovo vaccinations against some avian viral, bacterial, and parasitic diseases are still under investigations and there is a progress for production of safe and efficient vaccines.
Author Contributions
WAA designed and review of the literature. The author has read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All prepared data are presented in the present article.
Conflicts of Interest
The author provides the publisher a copyright license, granting the publisher the exclusive right to publish and sell the study findings in whole or in part in any language. The author has no relevant financial or non-financial interests to disclose.
References
- García, M. Current and future vaccines and vaccination strategies against infectious laryngotracheitis (ILT) respiratory disease of poultry. Vet. Microbiol. 2017, 206, 157–162. [Google Scholar] [CrossRef]
- Sokale, A.O.; Zhai, W.; Pote, L.M.; Williams, C.J.; Peebles, E.D. Effects of coccidiosis vaccination administered by in ovo injection on the hatchability and hatching chick quality of broilers. Poult. Sci. 2017, 96, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Williams, C. In ovo vaccination for disease prevention. Int. Poult. Prod. 2007, 15, 7–8. Available at: http://www.positiveaction.info/pdfs/articles/pp15.8p7.pdf.
- Negash, T.; Al-garib, S.O.; Gruqays, E. Comparison of in-ovo and post-hatch vaccination with particular reference to infectious bursal disease. A review. Vet. Q. 2004, 26, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Jochemsen, P.; Jeurissen, S.H.M. The localization and uptake of in ovo injected soluble and particulate substances in the chicken. Poult. Sci. 2002, 81, 1811–1817. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.M.; Burmester, B.R. Resistance of Marek’s disease at hatching in chickens vaccinated as embryos with the turkey herpesvirus. Avian Dis. 1982, 26, 134–149. [Google Scholar] [CrossRef]
- Fernandes, J.I.M.; Prokoski, K.I.; Oliveira, B.C.I.; Oro, C.S.I.; Oro, P.J.I.I. Evaluation of incubation yield, vaccine response, and performance of broilers submitted to in-ovo vaccination at different embryonic ages. Rev. Bras. Cienc. Avic. 2016, 18, 55–63. [Google Scholar] [CrossRef]
- Okwor, G.O.; El-Yuguda, A.; Baba, S.S. Profile of maternally derived antibody in broiler chicks and in-ovo vaccination of chick embryo against Newcastle disease. World J. Vaccines 2014, 4, 72–80. [Google Scholar] [CrossRef]
- Ahmad, J.; Sharma, J.M. Protection against hemorrhagic enteritis and Newcastle disease in turkeys by embryo vaccination with monovalent and bivalent vaccines. Avian Dis. 1993, 37, 485–491. [Google Scholar] [CrossRef]
- McMillen, J.K.; Cochran, M.D.; Junker, D.E.; Reddy, D.N.; Valencia, D.M. The safe and effective use of fowlpox virus as a vector for poultry vaccine. Dev. Biol. Stand. 1994, 82, 137–145. [Google Scholar] [PubMed]
- Wakenell, P.S.; Bryan, T.; Schaeffer, J.; Avakian, A.; Williams, C.; Whitfill, C. Effect of in ovo vaccine delivery route on HVT/SB-1 efficacy and viremia. Avian Dis. 2002, 46, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.Y.; Giambrone, J.J.; Dormitorio, T.V.; Wu, H. Influence of a reovirus-antibody complex vaccine on efficacy of Marek's disease vaccine administered in ovo. Avian Dis. 2003, 47, 1362–1367. Available at: http://www.jstor.org/stable/1593177.
- Worthington, K.J.; Sargent, B.A.; Davelaar, F.G.; Jones, R.C. Immunity to avian pneumovirus infection in turkeys following in ovo vaccination with an attenuated vaccine. Vaccine 2003, 21, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Hess, M.; Huggins, M.B.; Heincz, U. Hatchability, serology and virus excretion following in ovo vaccination of chickens with an avian metapneumovirus vaccine. Avian Pathol. 2004, 33, 576–580. [Google Scholar] [CrossRef] [PubMed]
- Astill, J.; Alkie, T.; Yitbarek, A.; Abdelaziz, K.T.; Bavananthasivam, J.; Nagy, J.J.; Petrik, É.; Sharif, S. Induction of immune response in chickens primed in ovo with an inactivated H9N2 avian influenza virus vaccine. BMC Res. Notes 2018, 11, 428. [Google Scholar] [CrossRef]
- García, C.; Soriano, J.M.; Cortés, V.; Sevilla-Navarro, S; Marin, C.; Balaguer, J.L.; Catalá-Gregori, P. Monitoring serologic response to single in ovo vaccination with an immune complex vaccine against infectious bursal disease in broilers. Poult. Sci. 2021, 100, 100999. [CrossRef]
- Marcano, V.C.; Cardenas-Garcia, S.; Diel, D.G.; Antoniassi da Silva, L.H.; Gogal, R.M. Jr.; Miller, P.J.; Brown, C.C.; Butt, S.L.; Goraichuk, I.V.; Dimitrov, K.M.; Taylor, T.L.; Williams-Coplin, D.; Olivier, T.L.; Stanton, J.B.; Afonso, C.L. A novel recombinant Newcastle disease vaccine improves post- In ovo vaccination survival with sustained protection against virulent challenge. Vaccines (Basel). 2021, 9, 953. [Google Scholar] [CrossRef]
- Noor, S.M.; Husband, A.J.; Widders, P.R. In ovo oral vaccination with Campylobacter jejuni establishes early development of intestinal immunity in chickens. Br. Poult. Sci. 1995, 36, 563–573. [Google Scholar] [CrossRef]
- Elliott, K.E.C.; Branton, S.L.; Evans, J.D.; Peebles, E.D. Evaluation of the potential influence of the disinfection cycle on the efficacy of strain F Mycoplasma gallisepticum vaccine administered by in ovo injection to layer hatching eggs. J. Appl. Poult. Res. 2020, 29, 673–683. [Google Scholar] [CrossRef]
- Alqhtani, A.H.; Fatemi, S.A.; Elliott, K.E.C.; Branton, S.L.; Evans, J.D.; Leigh, S.A.; Gerard, P.D.; Peebles, E.D. Effects of the In ovo vaccination of the ts-11 Strain of Mycoplasma gallisepticum in layer embryos and posthatch chicks. Animals 2022, 12, 1120. [Google Scholar] [CrossRef]
- Hornok, S.; Szell, Z.; Sreter, T.; Kovacs, A.; Varga, I. Influence of in ovo administered Cryptosporidium baileyi oocyst extract on the course of homologous infection. Vet. Parasitol. 2000, 89, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Sokale, A.O.; Williams, C.J.; Hoerr, F.J.; Collins, K.E.C.; Peebles, E.D. Effects of administration of an in ovo coccidiosis vaccine at different embryonic ages on vaccine cycling and performance of broiler chickens, Poult. Sci. 2021, 100, 100914. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cai, K.; Mishra, R.; Jha, R. In ovo supplementation of chitooligosaccharide and chlorella polysaccharide affects cecal microbial community, metabolic pathways, and fermentation metabolites in broiler chickens. Poult. Sci. 2020, 99, 4776–4785. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.J.; Li, C.H.; Tsai, P.K.; Lai, C.C.; Kuo, Y.R.; Hsieh, M.K.; Cheng, C.W. Electromagnetic force-driven needle-free in ovo injection device. Vet Sci. 2022, 9, 147. [Google Scholar] [CrossRef]
- Gildersleeve, R.P.; Hoyle, C.M.; Miles, A.M.; Murray, D.L.; Ricks, C.A.; Secrest, M.N.; Williams, C.J.; Womack, C.L. Developmental performance of an egg injection machine for administration of Marek's disease vaccine. J. Appl. Poult. Res. 1993, 2, 337–346. [Google Scholar] [CrossRef]
- Sarma, G.; Greer, W.; Gildersleeve, R.P.; Murray, D.L.; Miles, A.M. Field safety and efficacy of in ovo administration of HVT + SB-1 bivalent Marek’s disease vaccine in commercial broilers. Avian Dis. 1995, 39, 211–217. [Google Scholar] [CrossRef]
- Sokale, A.O.; Williams, C.J.; Triplett, M.D.; Hoerr, F.J.; Peebles, E.D. Effects of stage of broiler embryo development on coccidiosis vaccine injection accuracy, and subsequent oocyst localization and hatchling quality. Poult. Sci. 2020, 99, 189–195. [Google Scholar] [CrossRef]
- Sharma, J.M.; Lee, L.F.; Wakenell, P.S. Comparative viral, immunologic, and pathologic responses of chickens inoculated with herpesvirus of turkeys as embryos or at hatch. Am. J. Vet. Res. 1984, 45, 1619–1623. [Google Scholar] [PubMed]
- Ricks, C.A.; Avakian, A. , Bryan, T. ; Gildersleeve, R.; Haddad, E.; Llich, R.; King, S.; Murray, L.; Phelps, P.; Poston, R.; Whitfill, C.; Williams, C.. In ovo vaccination technology. Adv. Vet. Med. 1999, 41, 495–515. [Google Scholar] [PubMed]
- Sharma, J.M.; Tizard, I. Avian cellular immune effector mechanisms. Avian Pathol. 1984, 13, 357–376. [Google Scholar] [CrossRef]
- Sharma, J.M. Delayed replication of Marek’s disease virus following in ovo inoculation during the late stage of embryonal development. Avian Dis. 1987, 31, 570–567. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, C.D.; McDaniel, C.D.; Abdelhamed, H.; Karsi, A.; Kiess, A.S. Evaluating bacterial colonization of a developing broiler embryo after in ovo injection with a bioluminescent bacteria. Poult. Sci. 2019, 98, 2997–3006. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.J., Hopkins, B.A. Field evaluation of the accuracy of vaccine deposition by two different commercially available in ovo injection systems. Poult. Sci. 2011, 90, 223–226. [CrossRef]
- Peebles, E.D. In ovo applications in poultry: A review. Poult. Sci. 2018, 97, 2322–2338. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.J.; Zedek, A.S. Comparative field evaluations of in-ovo applied technology. Poult. Sci. 2010, 89, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Avakian, A. Understanding in ovo vaccination. Int. Hatch. Pract, 2006, 20, 15–17. Available at: http://www.positiveaction.info/pdfs/articles/hp20.5p15.pdf.
- Tong, Q.; Romanini, C.E.; Exadaktylos, V.; Bahr, C.; Berckmans, D.; Bergoug, H.; Eterradossi, N.; Roulston, N.; Verhelst, R.; McGonnell, I.M.; Demmers, T. Embryonic development and the physiological factors that coordinate hatching in domestic chickens. Poult. Sci. 2013, 92, 620–662. [Google Scholar] [CrossRef]
- Sokale, A.O.; Williams, C.J.; Cummings, T.S.; Gerard, P.D.; Bello, A.; Peebles, E.D. Effects of in ovo injection of different doses of coccidiosis vaccine and turn-out times on broiler performance. Poult. Sci. 2018, 97, 1891–1898. [Google Scholar] [CrossRef]
- Vandeputte, J.; Martel, A.; Van Rysselberghe, N.; Antonissen, G.; Verlinden, M.; De Zutter, L.; Heyndrickx, M.; Haesebrouck, F.; Pasmans, F.; Garmyn, A. In ovo vaccination of broilers against Campylobacter jejuni using a bacterin and subunit vaccine. Poult. Sci. 2019, 98, 5999–6004. [Google Scholar] [CrossRef]
- Sharma, J.M. Embryo vaccination with infectious bursal disease virus alone or in combination with Marek's disease vaccine. Avian Dis. 1985, 29, 1155–1169. [Google Scholar] [CrossRef]
- Sharma, J.M. Embryo vaccination of specific pathogen free chickens with infectious bursal disease virus: tissue distribution of the vaccine virus and protection of the hatched chicken against disease. Avian Dis. 1986, 30, 776–780. [Google Scholar] [CrossRef]
- Wakenell, P.S.; Sharma, J.M. Chicken embryonal vaccination with avian infectious bronchitis virus. Am. J. Vet. Res. 1986, 47, 933–938. [Google Scholar] [PubMed]
- Berchieri, Jr.A.; Bolis, D.A. Vacinações e aplicações de produtos intra-ovo. In: Macari, M.; Gonzales, E.; editores. Manejo da incubação.Campinas:, FACTA; 2003. p., 267-283.
- Sharma, J.M.; Coulson, B.D.; Young, E. Effect of in vitro adaptation of MDV on pock induction on the chorioallantoic membrane of embryonated chicken eggs. Infect. Immun. 1976, 13, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.J. In-ovo vaccination and chick quality. Int. Hatch. Practice 2005, 19, 7–13. Available at: http://www.positiveaction.info/pdfs/articles/hp19.2p7.pdf.
- Abudabos, A. The Effect of broiler breeder strain and parent flock age on hatchability and fertile hatchability. Int. J. Poult. Sci. 2010, 9, 231–235. [Google Scholar] [CrossRef]
- Avakian, A.; Wakenell, P.S.; Bryan, T.; Schaeffer, J.L.; Williams, C.J.; Whitfill, C.E. In ovo administration of Marek’s disease vaccine: Importance of vaccine deposition site in the fertile egg. Proceedings of 51º Western Poultry Disease Conference. Puerto Vallarta. México: Veterinary Software Publishing O’Fallon, 2002. [Google Scholar]
- Amarasinghe, G.K.; Ceballos, N.G.A.; Banyard, A.C.; Basler, C.F.; Bavari, S.; Bennett, A.J.; Blasdell, K.R.; Briese, T.; Bukreyev, A.; Caì, Y.; et al. Taxonomy of the order Mononegavirales: Update 2018. Arch. Virol. 2018, 163, 2283–2294. [Google Scholar] [CrossRef]
- Perozo, F.; Marcano, R.; Afonso, C. Biological and phylogenetic characterization of a genotype VII Newcastle disease virus from Venezuela: Efficacy of field vaccination. J. Clin. Microbiol. 2012, 50, 1204–1208. [Google Scholar] [CrossRef]
- Jaganathan, S.; Ooi, P.T.; Phang, L.Y.; Allaudin, Z.N.B.; Yip, L.S.; Choo, P.Y.; Lim, B.K.; Lemiere, S.; Audonnet, J.C. Observation of risk factors, clinical manifestations and genetic characterization of recent Newcastle disease virus outbreak in West Malaysia. BMC Vet. Res. 2015, 11, 219. [Google Scholar] [CrossRef]
- Wajid, A.; Dimitrov, K.M.; Wasim, M.; Rehmani, S.F.; Basharat, A.; Bibi, T.; Arif, S.; Yaqub, T.; Tayyab, M.; Ababneh, M.; Sharma, P.; Miller, P.J.; Afonso, C.L. Repeated isolation of virulent Newcastle disease viruses in poultry and captive non-poultry avian species in Pakistan from 2011 to 2016. Prev. Vet. Med. 2017, 142, 1–6. [CrossRef]
- Absalon, A.E.; Cortés-Espinosa, D.V.; Lucio, E.; Miller, P.J.; Afonso, C.L. Epidemiology, control, and prevention of Newcastle disease in endemic regions: Latin America. Trop. Anim. Health Prod. 2019, 51, 1033–1048. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Afonso, C.L.; Yu, Q.; Miller, P.L. Newcastle disease vaccines-A solved problem or a continuous challenge? Vet. Microbiol. 2017, 206, 126–136. [Google Scholar] [CrossRef]
- Schijns, V.E.J.C.; van de Zande, S.; Lupiani, B.; Reffy, S.M. Practical Aspects of Poultry Vaccination. In Avian Immunology; Schat, K.A., Kaspers, B., Kaiser, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 345–362. [Google Scholar]
- Gallili, G.E.; Ben-Nathan, D. Newcastle disease vaccines. Biotechnol. Adv. 1998, 16, 343–366. [Google Scholar] [CrossRef]
- Jansen, T.; Hofmans, M.P.M.; Theelen, M.J.G.; Cshijns, V.E.J.C. Structure-activity relations of water-in-oil vaccine formulations and induced antigen-specific antibody responses. Vaccine 2005, 23, 1053–1060. [Google Scholar] [CrossRef]
- Giambrone, J.J.; Closser, J. Effect of breeder vaccination on immunization of progeny against Newcastle disease. Avian Dis. 1990, 34, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, J.; Sharma, J.M. Evaluation of a modified-live virus vaccine administered in ovo to protect chickens against Newcastle disease. Am. J. Vet. Res. 1992, 53, 1999–2004. [Google Scholar] [CrossRef] [PubMed]
- Mast, J.; Nanbru, C.; Decaesstecker, M.; Lambrecht, B.; Couvreur, B.; Meulemans, G.; van den Berg, T. Vaccination of chicken embryos with escape mutants of La Sota Newcastle disease virus induces a protective immune response. Vaccine 2006, 24, 1756–1765. [Google Scholar] [CrossRef] [PubMed]
- Dilaveris, D.; Chen, C.; Kaiser, P.; Russel, P.H. The safety and immunogenicity of an in ovo vaccine against Newcastle disease virus differ between two lines of chicken. Vaccine 2007, 25, 3792–3799. [Google Scholar] [CrossRef]
- Cardenas-Garcia, S.; Dunwoody, R.P.; Marcano, V.; Diel, D.G.; Williams, R.J.; Gogal, R.M., Jr.; Brown, C.C.; Miller, P.J.; Afonso, C.L. Effects of chicken interferon gamma on Newcastle disease virus vaccine immunogenicity. PLoS One 2016, 11, e0159153. [Google Scholar] [CrossRef]
- Coletti, M.; Del Rossi, E.; Franciosini, M.P.; Passamonti, F.; Tacconi, G.; Marini, C. Efficacy and safety of an infectious bursal disease virus intermediate vaccine in ovo. Avian Dis. 2001, 45, 1036–1043. [Google Scholar] [CrossRef]
- Dunon, D.; Courtois, D.; Vainio, O.; Six, A.; Chen, C.H.; Cooper, M.D.; Dangy, J.P.; Imhof, B.A. Ontogeny of the immune system: Gamma/delta and alpha/beta T cells migrate from thymus to the periphery in alternating waves. J. Exp. Med. 1997, 186, 977–988. [Google Scholar] [CrossRef]
- Ohta, H.; Ezoe, S.; Yamazaki, K.; Kawai, T.; Honda, T. Application of aluminum hydroxide for an in ovo live Newcastle disease vaccine. Avian Dis. 2009, 53, 392–395. [Google Scholar] [CrossRef]
- Romanutti, C.; Keller, L.; Zanetti, F.A. Current status of virus-vectored vaccines against pathogens that affect poultry. Vaccine 2020, 38, 6990–7001. [Google Scholar] [CrossRef]
- Hein, R.; Koopman, R.; Garcia, M.; Armour, N.; Dunn, J.R.; Barbosa, T.; Martinez, A. Review of poultry recombinant vector vaccines. Avian Dis. 2021, 65, 438–452. [Google Scholar] [CrossRef] [PubMed]
- Rautenschlein, S.; Sharma, J.M.; Winslow, B.J.; McMillen, J.; Junker, D.; Cochran, M. Embryo vaccination of turkeys against Newcastle disease infection with recombinant fowlpox virus constructs containing interferon's as adjuvant. Vaccine 2000, 18, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Palya, V.; Kiss, I.; Tatár-Kis, T.; Mató, t.; Felföldi, B.; Gardin, Y. Advancement in vaccination against Newcastle disease: Recombinant HVT NDV provides high clinical protection and reduces challenge virus shedding with the absence of vaccine reactions. Avian Dis. 2012, 56, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Esaki, M.; Godoy, A.; Rosenberger, J.K.; Rosenberger, S.C.; Gardin, Y.; Yasuda, A.; Dorsey, K.M. Protection and antibody response caused by turkey herpesvirus vector Newcastle disease vaccine. Avian Dis. 2013, 57, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Schat, K.A. Back to the Past: Do Vector Vaccines Represent the Future? Department of Microbiology and Immunology College of Veterinary Medicine, Cornell University: Ithaca, NY, USA, 2015; pp. 1–12.
- Xu, Z.; Wei, W.; Gagneur, J.; Clauder-Münster, S.; Smolik, M.; Huber, W.; Steinmetz, L.M. Antisense expression increases gene expression variability and locus interdependency. Mol. Syst. Biol. 2011, 7, 468. [Google Scholar] [CrossRef]
- Pelechano, V.; Steinmetz, L.M. Gene regulation by antisense transcription. Nat. Rev. Genet. 2013, 14, 880–893. [Google Scholar] [CrossRef]
- Karaca, K.; Sharma, J.M.; Winslow, B.J.; Junker, D.E.; Reddy, S.; Cochran, M.; McMillen, J. Recombinant fowlpox viruses co-expressing chicken type I IFN and Newcastle disease virus HN and F genes: influence of IFN on protective efficacy and humoral responses of chickens following in ovo or post-hatch administration of recombinant viruses. Vaccine, 1998, 16, 1496–1503. [Google Scholar] [CrossRef]
- Ramp, K.; Topfstedt, E.; Wackerlin, R.; Hoper, D.; Ziller, M.; Mettenleiter, T.C.; Grund, C.; Romer-Oberdorfer, A. Pathogenicity and immunogenicity of different recombinant Newcastle disease virus clone 30 variants after in ovo vaccination. Avian Dis. 2012, 56, 208–217. [Google Scholar] [CrossRef]
- Kapczynski, D.R.; Martin, A.; Haddad, E.E.; King, D.J. Protection from clinical disease against three highly virulent strains of Newcastle disease virus after in ovo application of an antibody-antigen complex vaccine in maternal antibody-positive chickens. Avian Dis. 2012, 56, 555–560. [Google Scholar] [CrossRef]
- Jeurissen, S.H.; Janse, E.M.; Lehrbach, P.R.; Haddad, E.E.; Avakian, A.; Whitfill, C.E. The working mechanism of an immune complex vaccine that protects chickens against infectious bursal disease. Immunology 1998, 95, 494–500. [Google Scholar] [CrossRef]
- Shirai, J.; Seki, R.; Kamimura, R.; Mitsubayashi, S. Effects of invert soap with 0. 05% sodium hydroxide on infectious bursal disease virus. Avian Dis. 1994, 38, 240–243. [Google Scholar] [PubMed]
- Corley, M.M.; Giambrone, J.J.; Dormitorio, T.V. Detection of infectious bursal disease vaccine viruses in lymphoid tissues after in ovo vaccination of specific-pathogen-free embryos. Avian Dis. 2001, 45, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Kelemen, M.; Forgách, K.; Iván, J.; Palya, V.; Süveges, T.; Tóth, B.; Mészáros, J. Pathological and immunological study of an in ovo complex vaccine against infectious bursal disease. Acta Vet. Hung. 2000, 48, 443–454. [Google Scholar] [CrossRef] [PubMed]
- de Wit, J.J.; Jorna, I.; Finger, A.; Loeb, V.; Dijkman, R.; Ashash, U.; Ifrah, M.; Raviv, Z. In ovo application of a live infectious bursal disease vaccine to commercial broilers confers proper immunity. Avian Pathol. 2021, 50, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Ashash, U.; Noach, C.; Perelman, B.; Costello, C.; Sansalone, P.; Brazil, T.; Raviv, Z. In ovo and day of hatch application of a live infectious bursal disease virus vaccine to commercial broilers. Avian Dis. 2019, 63, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Lara, L.J.C.; Michell, B.C.; Baião, N.C.; Resende, M,; Gomes, A.D.; Martins, N.R.S,. Effect of maternally-derived antibodies on the performance and immunity of broilers induced by in ovo or post-hatching immunizations with a live vaccine against infectious bursal disease. Braz. J. Poult. Sci. 2009, 11, 57–63. [CrossRef]
- Riaz, M.N.; Hussain, I.; Akhtar, M.; Rasool, M.H.; Mansoor, M.K. Haq SEU. Evaluation of in ovo vaccination against infectious bursal disease virus in commercial broilers in Pakistan. Int. J. Agric. Biol. 2004, 6, 984–986. Available at: https://www.fspublishers.org/published_papers/97468_..pdf.
- Rautenschlein, S.; Haase, C. Differences in the immunopathogenesis of infectious bursal disease virus (IBDV) following in ovo and post-hatch vaccination of chickens. Vet. Immunol Immunopathol. 2005, 106, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Zaheer, I.; Chen, W.; Khan, A.; Elokil, A.; Saleemi, M.K.; Zaheer, T.; Khan, M.Z. Immunopathological comparison of in ovo and post-hatch vaccination techniques for infectious bursal disease vaccine in layer chicks. Front. Vet. Sci. 2022, 9, 947522. [Google Scholar] [CrossRef]
- Fahey, K.J.; Crooks, J.K.; Fraser, R.A. Assessment by ELISA of passively acquired protection against infectious bursal disease virus in chickens. Aust. Vet. J. 1987, 64, 203–207. [Google Scholar] [CrossRef]
- Giambrone, J.J.; Dormitorio, T.; Brown, T. Safety and efficacy of in ovo administration of infectious bursal disease viral vaccines. Avian Dis. 2001, 45, 144–148. [Google Scholar] [CrossRef]
- McCarty, J.E.; Brown, T.P.; Giambrone, J.J. Delay of infectious bursal disease virus infection by in ovo vaccination of antibody-positive chicken eggs. J. Appl. Poult. Res. 2005, 14, 136–140. [Google Scholar] [CrossRef]
- McCarty, J.; Newman, L.; Brown, T.P.; Giambrone, J. The effect of in ovo IBDV vaccination when administered in antibody positive and negative chickens. Page 31 in Program and Abstr. XIII Congr. World Poult. Assoc., Denver, CO, 2003.
- Corley, M.M.; Giambrone, J.J.; Dormitorio, T.V. Evaluation of the immune response and detection of infectious bursal disease viruses by reverse transcriptase-polymerase chain reaction and enzyme-linked immunosorbent assay after in ovo vaccination of commercial broilers. Avian Dis. 2002, 46, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H; Sung, H.W.; Yoon, B.I.; Kwon, H.M. Protection of chicken against very virulent IBDV provided by in ovo priming with DNA vaccine and boosting with killed vaccine and the adjuvant effects of plasmid-encoded chicken interleukin-2 and interferon-gamma. J. Vet. Sci. 2009, 10, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Gagic, M.; St Hill, C.A.; Sharma, J.M. In ovo vaccination of specific-pathogen-free chickens with vaccines containing multiple agents. Avian Dis. 1999, 43, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Muller, H.; Mundt, E.; Eterradossi, N.; Rafiqul Islam, M. Current status of vaccines against infectious bursal disease. Avian Pathol. 2012, 41, 133–139. [Google Scholar] [CrossRef]
- Davison, F. The Importance of the Avian Immune System and its Unique Features: Embryonic (In ovo) Vaccination. Karel A. Schat, Bernd Kaspers, Pete Kaiser (Eds.), Page 7 in Avian Immunology (2nd ed), Elsevier Ltd, San Diego, CA, 2014.
- Comte, S.; Borne P. Vacinas e vacinação na produção avícola. São Paulo: Ceva Santa Animale, 2003, p. 140.
- Zhang, Y.; Sharma, J.M. Early post-hatch protection against Marek's disease in chickens vaccinated in ovo with a CVI988 serotype 1 vaccine. Avian Dis. 2001, 45, 639–645. [Google Scholar] [CrossRef]
- Tarpey, I.; van Loon, A.A.; de Haas, N.; Davis, P.J.; Orbell, S.; Cavanagh, D.; Britton, P.; Casais, R.; Sondermeijer, P.; Dundick, R. A recombinant turkey herpesvirus expressing chicken interleukin-2 increases the protection provided by in ovo vaccination with infectious bursal disease and infectious bronchitis virus. Vaccine 2007, 25, 8529–8535. [Google Scholar] [CrossRef]
- Reddy SK, Sharma JM, Ahmed J, Reddy DN, McMillan JK, Cook SM, Wild MA and Schwartz RD. Protective efficacy of a recombinant herpesvirus of turkeys as an in ovo vaccine against Newcastle and Marek's diseases in specific pathogen free chickens. Vaccine 1996, 14, 469–477. [CrossRef]
- Davison, A.J.; Eberle, R.; Hayward, G.S.; Mcgeoch, D.J.; Minson, A.C.; Pellet, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2009, 154, 171–177. [Google Scholar] [CrossRef]
- García, M.; Spatz, S.; Guy, J. Infectious Laryngotracheitis. In Swayne, D.E.; Glisson, J.R.; McDougald, L.R.; Nolan, L.K.; Suarez, D.L.; Nair, V.L. (Eds). Diseases of Poultry 13th edn (pp.). Ames, Iowa: Wiley- Blackwell, 2013, pp. 161–179.
- Guy, J.; Garcıa, M. Infectious Laryngotracheitis Virus. In Saif, Y.M.; Glisson, J.R.; Fadly, A.M.; McDougald, L.R.; Nolan, L.K.; Swayne, D.E. (Eds). Diseases of Poultry, 12th ed., Ames, AI: Blackwell Publishing, 2008, pp. 137–152.
- Gelenczei, E.F.; Marty, E.W. Studies on a tissue-culture modified infectious larygotracheitis virus. Avian Dis. 1964, 8, 105–122. [Google Scholar] [CrossRef]
- Fulton, R.M.; Schrader, D.L.; Will, M. Effect of route of vaccination on the prevention of infectious laryngotracheitis in commercial egg-laying chickens. Avian Dis. 2000, 44, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Han, M.G.; Kim, S.J. Efficacy of live virus vaccines against infectious laryngotracheitis assessed by polymerase chain reaction restriction fragment length polymorphism. Avian Dis. 2003, 47, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Rodr´ıguez-Avila, A.; Oldoni, I., Riblet, I.; Riblet, S.M.; Garc´ıa, M. Evaluation of the protection elicited by direct and indirect exposure to live attenuated infectious laryngotracheitis virus (ILTV) vaccines against a recent challenge strain from the United States. Avian Pathol. 2008, 37, 287–292. [CrossRef]
- Davison, S.; Gingerich, E.; Casavant, N.S.; Eckroade, R.J. Evaluation of the efficacy of a live fowlpox-vectored infectious laryngotracheitis/avian encephalomyelitis vaccine against ILT viral challenge. Avian Dis. 2006, 50, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Vagnozzi, A.; Zavala, G.; Riblet, S.M.; Mundt, A.; García, M. Protection induced by commercially available live-attenuated and recombinant viral vector vaccines against infectious laryngotracheitis virus in broiler chickens. Avian Pathol. 2012, 41, 21–31. [Google Scholar] [CrossRef]
- Maekawa, D.; Beltrán, G.; Riblet, S.M.; García, M. Protection efficacy of a recombinant herpesvirus of turkey vaccine against infectious laryngotracheitis virus administered in ovo to broilers at three standardized doses. Avian Dis. 2019, 63, 351–358. [Google Scholar] [CrossRef]
- Johnson, D.I.; Vagnozzi, A.; Dorea, F.; Riblet, S.M.; Mundt, A.; Zavala, G.; Garc´ıa, M. Protection against infectious laryngotracheitis virus (ILTV) by in ovo vaccination by commercially available viral vector recombinant vaccines. Avian Dis. 2010, 54, 1251–1259. [Google Scholar] [CrossRef]
- Williams, S.M.; Smith, J.A.; Garc´ıa, M.; Brinson, D.; Kiupel, M.; Hofacre, C. Severe histiolymphocytic and heterophilic bronchopneumonia as a reaction to in ovo fowlpox vaccination in broiler chicks. Vet. Pathol. 2010, 47, 177–180. [Google Scholar] [CrossRef]
- Mashchenko, A.; Riblet, S.M.; Zavala, G.; García, M. In ovo vaccination of commercial broilers with a glycoprotein J gene-deleted strain of infectious laryngotracheitis virus. Avian Dis. 2013, 57, 523–531. [Google Scholar] [CrossRef]
- Thapa, S.; Cader, M.S.A.; Murugananthan, K.; Nagy, E.; Sharif, S.; Czub, M.; Abdul-Careem, M.F. In ovo delivery of CpG DNA reduces avian infectious laryngotracheitis virus induced mortality and morbidity. Viruses. 2015, 7, 1832–1852. [Google Scholar] [CrossRef]
- Gimeno, I.M.; Cortes, A.L.; Guy, J.S.; Turpin, E.; Williams, C. Replication of recombinant herpesvirus of turkey expressing genes of infectious laryngotracheitis virus in specific pathogen free and broiler chickens following in ovo and subcutaneous vaccination. Avian Pathol. 2011, 40, 395–403. [Google Scholar] [CrossRef]
- Cavanaugh, D.; Madwort, K.; Welchman, D.; Britton, P.; Gough, R.E. Coronaviruses from pheasants (Phasianus colchicus) are genetically closely related to coronaviruses of domestic fowl (infectious bronchitis virus) and turkeys. Avian Pathol. 2002, 31, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Naqi, S. Infectious bronchitis. In: Calnek, B.W.; Barnes, H.J., McDougald, L.R.; Saif, Y.M. (Eds.), Diseases of Poultry, 10th Edn., London, 1997, pp. 511–526.
- Ignjatovic, J.; Gould, G.; Sapats, S. Isolation of a variant infectious bronchitis virus in Australia that further illustrates diversity among emerging strains. Arch. Virol. 2003, 151, 1567–1585. [Google Scholar] [CrossRef] [PubMed]
- Babapoor, S.; Almeida, D.O.; Fabis, J.J.; Helal, Z.H.; Wang, X.; Girshick, T.; Khan, M.I. Protective effect of in ovo vaccination with IBV-spike-recombinant DNA and chicken interferon as an adjuvant. Int. J. Poult. Sci. 2009, 8, 1034–1041. [Google Scholar] [CrossRef]
- Wakenell, P.S.; Sharma, J.M.; Slocombe, R.F. Embryo vaccination of chickens with infectious bronchitis virus: Histologic and ultrastructural lesion response and immunologic response to vaccination. Avian Dis. 1995, 39, 752–765. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Fabis, J.J. In ovo vaccination for IBV, using DNA vaccine. A preliminary study. 139th annual American Veterinary Medical Association Convention and Meeting, Nashville, KY., 2002, pp. 73.
- Kapczynski, D.R.; Hilt, D.A.; Shapiro, D.; Sellers, H.S.; Jackwood, M.W. Protection of chickens from infectious bronchitis by in ovo and intramuscular vaccination with a DNA vaccine expressing the S1 glycoprotein. Avian Dis. 2003, 47, 272–285. [Google Scholar] [CrossRef]
- Fabis, J.J.; Khan, M.I. In vivo expression of IBV-S gene in chicks inoculated with recombinant DNA vaccine in ovo. IV. Symposium on Avian Corona and Pneumovirus infections, Rauischholzhausen, Germany, 2004, pp: 232-236.
- Chew, P.H.; Wakenell, P.S.; Farver, T.B. Pathogenicity of attenuated infectious bronchitis viruses for oviducts of chickens exposed in ovo. Avian Dis. 1997, 41, 598–603. [Google Scholar] [CrossRef]
- Lee, C.W.; Brown, C.; Jackwood, M.W. Tissue distribution of avian infectious bronchitis virus following in ovo inoculation of chicken embryos examined by in situ hyberdization with antisense digoxienin-labeled universal riboprob. J. Vet. Diagn. Invest. 2002, 14, 377–381. [Google Scholar] [CrossRef]
- Avakian, A.P.; Wakenell, P.S,.; Grosse, D.; Whitfill, C.E.; Link, D. Protective immunity to infectious bronchitis in broilers vaccinated against Marek's disease either in ovo or at hatch and against infectious bronchitis at hatch. Avian Dis. 2000, 44, 536-544. [CrossRef]
- Tunio, M.T.; Abro, S.H.; Rind, R.; Wagan, R.; Hashmi, H.A. In ovo-vaccination of chicken embryos with infectious bronchitis virus vaccine. Agric. Trop. Subtrop. 2010, 43, 222–226. [Google Scholar]
- Wang, X.; Schnitzlein, W.M.; Tripathy, D.N., Girshick T.; Khan, M.I. Construction and immunogenicity studies of recombinant fowl poxvirus containing the S1 gene of Massachusetts 41 strain of infectious bronchitis virus. Avian Dis. 2002, 46, 831–838. [CrossRef]
- Johnson, M.A.; Pooley, C.; Ignjatovic, J.; Tyack, S.G. A recombinant fowl adenovirus expressing the S1 gene of infectious bronchitis virus protects against challenge with infectious bronchitis virus. Vaccine. 2003, 21, 2730–2736. [Google Scholar] [CrossRef]
- De Silva Senapathi, U.; Aboelkhair, M.; Puro, K.; Ali, M.; Amarasinghe, A.; Abdul-Cader, M.S.; Van Marle, G.; Czub, M.; Abdul-Careem, M.F. In ovo delivered toll-like receptor 7 ligand, resiquimod enhances host responses against infectious bronchitis corona virus (IBV) infection. Vaccines (Basel). 2020, 8, 186. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Mettenleiter, T.C.; Abdelwhab, E.M. A brief summary of the epidemiology and genetic relatedness of avian infuenza H9N2 virus in birds and mammals in the Middle East and North Africa. Epidemiol. Infect. 2017, 145, 3320–3333. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Pu, J.; Fan, L.; Sun, H.; Wang, J.; Zhang, Y.; Liu, L.; Liu, J. Evaluation of the protective efficacy of a commercial vaccine against different antigenic groups of H9N2 influenza viruses in chickens. Vet. Microbiol. 2012, 156, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Stone, H.; Mitchell, B.; Brugh, M. In ovo vaccination of chicken embryos with experimental Newcastle disease and avian influenza oil emulsion vaccines. Avian Dis. 1997, 41, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Toro, H.; Tang, D.C.; Suarez, D.L.; Sylte, M.J.; Pfeifer, J.; Van Kampen, K.R. Protective avian influenza in ovo vaccination with non-replicating human adeno- virus vector. Vaccine. 2007, 25, 2886–2891. [Google Scholar] [CrossRef]
- Toro, H.; Tang, D.C. Protection of chickens against avian influenza with nonreplicating adenovirus-vectored vaccine. Poult. Sci. 2009, 88, 867–871. [Google Scholar] [CrossRef]
- Mesonero, A.; Suarez, D.L.; van Santen, E.; Tang, D.C.; Toro, H. Avian influenza in ovo vaccination with replication defective recombinant adenovirus in chickens: vaccine potency, antibody persistence, and maternal antibody transfer. Avian Dis. 2011, 55, 285–292. [Google Scholar] [CrossRef]
- Cai, Y.; Song, H.; Ye, J.; Shao, H.; Padmanabhan, R.; Sutton, T.C.; Perez, DR. Improved hatchability and efficient protection after in ovo vaccination with live attenuated H7N2 and H9N2 avian influenza viruses. Virol. J. 2011, 8, 31. [Google Scholar] [CrossRef]
- Steel, J.; Burmakina, S.V.; Thomas, C.; Spackman, E.; García-Sastre, A.; Swayne, D.E.; Palese, P.A. A combination in-ovo vaccine for avian influenza virus and Newcastle disease virus. Vaccine. 2008, 26, 522–531. [Google Scholar] [CrossRef]
- Rafique, S.; Siddique, N.; Qayyum, M.; Abbas, M.A.; Ali, A.; Yasmeen, S.; Naeem, K. In ovo vaccination against avian influenza virus subtype H9N2. Pak. Vet. J. 2015, 35, 299–302. [Google Scholar]
- Dar, A.; Tikoo, S.; Potter, A.; Babiuk, L.A.; Townsend, H.; Gerdts, V.; Mutwiri, G. CpG-ODNs induced changes in cytokine/chemokines genes expression associated with suppression of infectious bronchitis virus replication in chicken lungs. Vet. Immunol. Immunopathol. 2014, 160, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Barjesteh, N.; Brisbin, J.T.; Behboudi, S.; Nagy, É.; Sharif, S. Induction of antiviral responses against avian influenza virus in embryonated chicken eggs with Toll-like receptor ligands. Viral Immunol. 2015, 28, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Tarpey, I.; Huggins, M.B. Onset of immunity following in ovo delivery of avian metapneumovirus vaccines. Vet. Microbiol. 2007, 124, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Cha, R.M.; Khatri, M.; Mutnal, M.; Sharma, J.M. Pathogenic and immunogenic responses in turkeys following in ovo exposure to avian metapneumovirus subtype C. Vet. Immunol. Immunopathol. 2011, 140, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Levisohn, S.; Kleven, S.H. Avian mycoplasmosis (Mycoplasma gallisepticum). Rev. Sci. Tech. 2000, 19, 425–442. [Google Scholar] [CrossRef] [PubMed]
- Kleven, S.H. Mycoplasmas in the etiology of multifactorial respiratory disease. Poult Sci. 1998, 77, 1146–1149. [Google Scholar] [CrossRef]
- Elliott, K.E.C.; Branton, S.L.; Evans, J.D.; Peebles, E.D. Early post-hatch survival and humoral immune response of layer chickens when in ovo vaccinated with strain F Mycoplasma gallisepticum. Poult. Sci. 2018, 97, 3860–3869. [Google Scholar] [CrossRef]
- Kleven, S.H. Changing expectations in the control of Mycoplasma gallisepticum. Acta. Vet. Hung. 1997, 45, 299–305. [Google Scholar] [PubMed]
- Evans, J.D.; Leigh, S.A.; Branton, S.L.; Collier, S.D.; Pharr, G.T.; Bearson, S.M.D. Mycoplasma gallisepticum: current and developing means to control the avian pathogen. J. Appl. Poult. Res. 2005, 14, 757–763. [Google Scholar] [CrossRef]
- Ferguson-Noel, N.; Cookson, K.; Laibinis, V.A.; Kleven, S.H. The efficacy of three commercial Mycoplasma gallisepticum vaccines in laying hens. Avian Dis. 2012, 56, 272–275. [Google Scholar] [CrossRef]
- Elliott, K.E.C.; Branton, S.L.; Evans, J.D.; Gerard, P.D.; Peebles, E.D. Layer chicken embryo survival to hatch when administered an in ovo vaccination of strain F Mycoplasma gallisepticum and locations of bacteria prevalence in the newly hatched chick. Poult. Sci. 2017, 96, 3879–3884. [Google Scholar] [CrossRef] [PubMed]
- Elliott, K.E.C.; Branton, S.L.; Evans, J.D.; Peebles, E.D. Occurrence of horizontal transmission in layer chickens after administration of in ovo strain F Mycoplasma gallisepticum vaccine. Poult. Sci. 2019, 98, 4492–4497. [Google Scholar] [CrossRef] [PubMed]
- Elliott, K.E.C.; Branton, S.L.; Evans, J.D.; Magee, C.L.; Peebles, E.D. Onset of the humoral immune response of layer chicks vaccinated in ovo with strain F Mycoplasma gallisepticum vaccine and evidence of male-biased mortality. Poult. Sci. 2022, 101, 101761. [Google Scholar] [CrossRef] [PubMed]
- McDougald, L.R.; Fitz-coy, S.H. Coccidiosis. In Diseases of Poultry. Saif, Y.M.; Fadley, A.M.; Glisson, J.R.; McDougald, L.R.; Nolan, L.K.; Swayne, D.E. 12th ed. Wiley-Blackwell Publishing, Ames, IA, 2008, pp. 1067–1080.
- Price, K.R. Use of live vaccines for coccidiosis control in replacement layer pullets. J. Appl. Poult. Res. 2012, 21, 679–692. [Google Scholar] [CrossRef]
- Moore, R.J. Necrotic enteritis predisposing factors in broiler chickens. Avian Pathol. 2016, 45, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, M.C.; Parker, C.; Ritter, D. Eimeria oocyst concentrations and species composition in litter from commercial broiler farms during anticoccidial drug or live Eimeria oocyst vaccine control programs. Avian Dis. 2017, 61, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Parent, E.; Fernandez, D.; Boulianne, M. The use of a live non-attenuated coccidiosis vaccine modifies Eimeria spp. excretion in commercial antibiotic-free broiler chicken flocks compared to conventional shuttle anticoccidial programs. Poult. Sci. 2018, 97, 2740–2744. [Google Scholar] [CrossRef]
- Price, K.R.; Hafeez, M.A.; Bulfon, J.; Barta, J.R. Live Eimeria vaccination success in the face of artificial non-uniform vaccine administration in conventionally reared pullets. Avian Pathol. 2016, 45, 82–93. [Google Scholar] [CrossRef]
- Watkins, K.L.; Brooks, M.A.; Jeffers, T.K.; Phelps, P.V.; Ricks, C.A. The effect of in ovo oocyst or sporocyst inoculation on response to subsequent coccidial challenge. Poult. Sci. 1995, 74, 1597–1602. [Google Scholar] [CrossRef]
- Mathis, G.; Schaeffer, J.; Cookson, K.; Dickson, J., LaVorgna, M.; Waldrip, D. Effect of lasalocid or salinomycin administration on performance and immunity following coccidia vaccination of commercial broilers. J. Appl. Poult. Res. 2014, 23, 577–585. [CrossRef]
- Weber, F.H.; Evans, N.A. Immunization of broiler chicks by in ovo injection of Eimeria tenella sporozoites, sporocysts, or oocysts. Poult. Sci. 2003, 82, 1701–1707. [Google Scholar] [CrossRef]
- Weber, F.H.; Genteman, K.C.; LeMay, M.A.; Lewis, D.O.; Evans, N.A. Immunization of broiler chicks by in ovo injection of infective stages of Eimeria. Poult. Sci. 2004, 83, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Weber, F.H.; Farrand, M.; LeMay, M.A.; Lewis, D.O.; Genteman, K.C.; Evans, N.A. Movement of oocysts within chicken embryos after in ovo vaccination with Eimeria maxima. VIIIth International Coccidiosis Conference, Palm Cove, Australia. Australian Society for Parasitology, Queensland, Australia, 2002, p. 184.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



