Submitted:
14 November 2024
Posted:
15 November 2024
You are already at the latest version
Abstract
Carotenoids, the bright yellow, orange, and red pigments of many fruits and vegetables, are essential components of the human diet as bioactive compounds not synthesized in animals. As a staple crop potato has the potential to deliver substantial amounts of these nutraceuticals despite their lower concentration in tubers compared to edible organs of other plant species. Even small gains in tuber carotenoid levels could have a significant impact on the nutritional value of potatoes. This review will focus on the current status and future perspectives of carotenoid biofortification in potato with conventional breeding and biotechnological approaches. The high biodiversity of tuber carotenoid levels and composition is presented, with an emphasis on the under-exploited native germplasm that represents a wide reservoir of useful genetic variants to breed carotenoid-rich varieties. The following section describes the structural genes involved in carotenoid metabolism and storage known to have a major impact on carotenoid accumulation in potato, together with the strategies that harnessed their expression changes to increase tuber carotenoid content. Finally, the little information available on the regulation of carotenoid metabolism and the desirable future advances in potato carotenoid biofortification are discussed.
Keywords:
potato
; carotenoids
; antioxidants
; biofortification
; conventional breeding
; genetic engineering
1. Introduction
Potato (Solanum sect. Petota, Solanaceae) is the fifth most important food crop worldwide. There has been an increasing trend in the global production of potatoes over the past 25 years, particularly significant in the two most populous countries: India (+125%) and China (+44%), which together represent more than 40% of the total share [1].
The potato germplasm includes more than 100 species, of which only 4 domesticated, and an estimated 4000 cultivars (cvs.) grown for food, feed, and non-food uses. Around 70% of the wild species are diploid. Conversely, commercial varieties are mostly autotetraploid and comprise highly heterozygous S. tuberosum genotypes clonally propagated through seed tubers [2,3].
Besides energy (87 Kcal/100 g FW) and proteins of high biological value, selected potato cultivars provide substantial amounts of essential micronutrients both organic (phenolic acids, flavonoids, carotenoids, vitamins B6, B9, and C) and inorganic (K, Ca, Mg, Fe, Zn) that help meet the relative daily requirements and fight hidden hunger. Low phytate and high vitamin C levels in potato tubers ensure proper bioavailability of key minerals including Fe and Zn [4,5,6,7].
Carotenoids are terpenoid compounds produced by all photosynthetic organisms and some heterotrophic prokaryotes and fungi.
In plants, they perform multiple functions essential for growth, development, and response to environmental stimuli and stress conditions. In photosynthesis, carotenoids act as light-harvesting pigments and have a key role in the assembly of the photosynthetic apparatus and its protection against photodamage. As precursors of a wide array of secondary metabolites known as apocarotenoids, including the two classes of plant hormones ABA and strigolactones, they participate in developmental and physiological processes throughout the plant life cycle. In addition, carotenoids and their derivatives contribute to the attractive colours and scents of flowers and fruits thus promoting pollination and seed dispersal [8,9,10].
Whereas carotenoid composition in photosynthetic tissues is rather constant, comprising lutein, β-carotene, neoxanthin, and violaxanthin as the main constituents [11], in non-photosynthetic organs such as fruits, flowers, and some types of stems and roots, carotenoid content and profile can be quite variable even within the same species [12].
Rising carotenoid levels in edible organs of crop plants, especially in staple crops, is desirable since these antioxidant compounds play specific roles in human health [13].
Alpha- and β-carotene and β-cryptoxanthin are the main precursors of vitamin A, whose deficiency causes impaired vision and increased susceptibility to infection and inflammation. About 500,000 children suffer annually from early blindness due to vitamin A deficiency [14].
Lutein and zeaxanthin are the main pigments of the human eye with a maximum concentration found in the macula lutea of the eye retina. By filtering blue light, they protect the eye against cataracts and age-related macular degeneration (AMD), a major cause of blindness in the elderly [15].
Due to their antioxidant activity, carotenoids have also been implicated in the prevention of oxidative stress-related diseases, including some types of cancer and dementia [16,17].
With rare exceptions [18], carotenoids cannot be produced in animals, therefore they must be assimilated regularly through the diet.
Carotenoids have also a widespread use as natural colorants in the feed, food, pharmaceutical, and cosmetic industries. For instance, carotenoid-rich feed is employed to enhance the pigmentation of dairy products, egg yolk and aquaculture fish [19].
This review highlights the biodiversity of potato carotenoid content and profile and current knowledge on the genetic basis of carotenoid accumulation in tubers as a foundation for carotenoid biofortification of potatoes with conventional breeding and new biotechnological approaches.
2. Carotenoid Biosynthesis in Plants
Carotenoids are a vast group of over 1200 tetraterpenoid compounds comprising carotenes, linear or cyclized hydrocarbons with a variable number of conjugated double bond systems, and xanthophylls, their oxygenated derivatives [20].
Plant carotenoids are produced in all types of differentiated plastids, primarily chloroplasts in green tissues and chromoplasts in flowers, fruits, and roots. In potato tubers, amyloplasts are the main site of carotenoid biosynthesis and storage [21].
The core carotenoid biosynthetic pathway is depicted in Figure 1.
The immediate precursor of carotenoids is the C20 isoprenoid geranylgeranyl diphosphate (GGPP) produced from the condensation of isopentenyl diphosphate (IPP) and dimethylallyl diphosphate (DMPP) units, derived from the 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway.
GGPP acts as a substrate in other competing pathways leading to the synthesis of several secondary metabolites including gibberellins (GAs), tocopherols, plastoquinones, and chlorophyll side chains and consequently its metabolism represents a primary control point of carotenoid production.
In the first committed step of carotenoid biosynthesis, two molecules of GGPP condensate to form phytoene, by means of phytoene synthase (PSY) activity.
The two ensuing desaturations catalysed by phytoene desaturase (PDS) followed by an isomerization step carried out by ζ-carotene isomerase (ZISO) convert phytoene to ζ -carotene.
A similar pattern of two desaturations and one isomerization follows, leading to the synthesis of all-trans lycopene. These steps involve ζ-carotene desaturase (ZDS) and carotenoid isomerase (CRTISO) activity, although in green tissues the isomerization is induced directly by light and requires chlorophyll as a sensitizer.
The extended conjugated double-bond system (chromophore) of carotenoids determines their color, which can vary from yellow to orange and red, except for the colorless phytoene and phytofluene, and their photochemical features, including light harvesting and photoprotection properties
The carotenoid pathway branches downstream all-trans lycopene, which is cyclized at both ends of the acyclic molecule following two different patterns.
In the β,ε−branch, lycopene β-cyclase (LYCB) and lycopene ε-cyclase (LYCE) carry out the synthesis of α-carotene with one β-ionone and one ε-ionone end group whereas in the β,β-branch LYCB introduces two β-ionone rings to generate β-carotene, the main precursor of vitamin A.
Hydroxylation of the terminal rings of α- and β-carotene leads to xanthophyll synthesis.
In the β,ε−branch, cytochrome p450-type enzymes (CIP97A andCYP97C) convert α-carotene into lutein, which accumulates on plastidial membranes.
In the other branch, a two-step reaction by the non-heme di-iron β-carotene hydroxylase (BCH/CHY) turns β-carotene sequentially into β-cryptoxanthin and zeaxanthin.
The ensuing two-step epoxidation of zeaxanthin catalysed by zeaxanthin epoxidase (ZEP) produces antheraxanthin and violaxanthin and can be reverted by violaxanthin de-epoxidase (VDE) in the so-called xanthophyll (or violaxanthin) cycle that represents a pivotal photoprotection system.
Alternatively, neoxanthin synthase (NSY) converts violaxanthin into neoxanthin: both xanthophylls are precursors of the plant hormone ABA.
The oxidative cleavage of several carotenoids, performed by carotenoid cleavage oxygenase (CCOs, also named CCDs and NCEDs in plants according to their substrate and cleavage sites), gives rise to a vast group of apocarotenoid molecules involved in various aspects of plant physiology and development. Among these are the pigments bixin and crocetin and the hormones ABA and strigolactones. In addition, CCO activity controls carotenoid homeostasis and recycling [10].
3. Carotenoid content and composition in potato tubers
In potatoes, carotenoid level and profile closely correlate with the hue and intensity of tuber flesh color in the yellow-orange range [22,23,24,25,26,27]. The total amount of potato tuber carotenoids has been reported to vary from less than 20 to more than 2000 μg /100 g fresh weight (FW) with the highest levels found in diploid South American native cultivars of S. phureja, S. stenotomum, and S. goniocalyx (also indicated as S. tuberosum Group Phureja, Stenotomum and Goniocalyx) [5,22,24,26,28,29,30,31,32].
The xanthophylls antheraxanthin, lutein, neoxanthin, violaxanthin, and zeaxanthin, in varying proportions, constitute the bulk of tuber carotenoids in wild and cultivated potatoes, with β-carotene as a minor component (less than 3% of total carotenoid content) [22,33,34], although in some native Andean cultivars relative high levels of β−carotene were detected [25,26,28,31].
Xanthophyll esters are present in variable amounts, reaching more than 60% of total carotenoids. Their concentration positively correlates with tuber carotenoid levels, in accordance with the proposed role of esterification as a means to stabilize xanthophylls and promote their sequestration within plastids [25,28,33,35].
Carotenoid concentration and profile of diploid and tetraploid potatoes with contrasting tuber flesh color are summarized in Table 1.
White-flashed genotypes show the lowest levels of total carotenoids with a reported maximum of 450 μg /100 g FW (recalculated from [25] considering 25% dry matter) and lutein as the main constituent. In S. phureja accessions an inverse correlation between β−carotene and total carotenoid levels was found. The average concentration of β-carotene in white-flashed tubers exceeded that of their coloured counterparts in both relative and absolute terms [26,31].
Yellow tubers display the widest range of carotenoid concentration, with recorded levels from 14 μg /100 g FW in an old-Spanish cultivar (recalculated from [28]) to 1435 μg /100 g FW [24]. Also, the carotenoid profile is highly variable with either violaxanthin [22,26,28,33,34], lutein [24,25,26] or antheraxanthin [36,38,39] as the main component.
Finally, orange-fleshed genotypes, all diploid, are carotenoid-rich, with maximum levels exceeding 2000 μg /100g FW [23], and show a distinctive profile characterized by remarkably elevated levels of zeaxanthin (up to 90% of total carotenoids) followed by antheraxanthin or lutein [26,32,40,41].
High broad-sense heritability for carotenoid content was estimated for both total (0.96) and individual carotenoids (from 0.51 for neoxanthin to 0.93 for antheraxanthin) pointing to the genotype as the main determinant of this trait [36]. Nevertheless, according to other studies, differences in environmental conditions linked to location or year of cultivation can have a significant impact on tuber carotenoid concentration and/or profile [27,29,32,37,42,43,44,45,46]. As to the influence of the growing method on carotenoid accumulation mixed results were reported. In some cases, a positive effect of organic and biodynamic vs. conventional farming on total and individual carotenoid accumulation was observed [47,48], while in others no differences or the opposite effect emerged [27,49]. Carotenoid levels, composition, and retention in tubers are also affected by postharvest storage and thermal treatments [32,39,50,51,52]. For instance, long-term cold storage was found to either increase, decrease, or leave total carotenoid content almost unaffected [29,35,53,54]. Moreover, either pairwise or triple interactions of genotype, environmental conditions, and processing method turned out to contribute to the variability in tuber carotenoid pattern and concentration, suggesting that phenotypic stability in different environments should be considered when selecting for carotenoid-rich potato genotypes [32,36,42].
3. Genetic Analysis of Carotenoid Accumulation in Tubers
For the genetic dissection of carotenoid accumulation in potato tuber, genome-wide scanning and candidate gene approaches were used.
Through conventional genetic analysis the Y (yellow) locus was identified as the main determinant of tuber flesh color and mapped to chromosome 3 (Chr. 3) [55] in the same region where a major QTL for flesh color and carotenoid content was later detected with bi-parental populations analysis and Genome-Wide Association Studies (GWAS). This QTL, disclosed in both diploid (Phureja) [56] and tetraploid (Tuberosum) germplasm [57,58,59], was estimated to explain 26% of the phenotypic variance for flesh colour [59].
Other minor QTLs were found scattered over the potato genome [56,57,59,60,61]. Most of them do not map to genomic regions encompassing carotenogenic genes, as is the case for tomato [62].
Candidate gene analysis for tuber carotenoid levels resulted in the identification of a few structural genes with a key role in carotenoid accumulation in potatoes, as described below. At the same time, manipulation of their expression levels led to the production of transgenic varieties showing high concentration of total and/or specific carotenoids.
3.1. Carotenoid Biosynthesis
3.1.1. Psy
The first committed, and often rate-limiting, step in carotenoid biosynthesis is catalysed by phytoene synthase (PSY) [63,64].
Overexpression of a bacterial phytoene synthase gene in potato tubers of a low carotenogenic variety resulted in a substantial increase of total carotenoids (up to 6.3-fold, 35.50 μg/g DW) and specific components, notably β-carotene (up to 10.30 μg/g DW) and lutein (up to 11 μg/g DW). Comparable results were obtained with a carotenoid-rich variety suggesting that also in potato PSY activity represents a bottleneck in carotenogenesis. [65].
In addition, a positive correlation was found between Psy2 transcript levels and tuber carotenoid content at early stages of tuber development [35] and in Zep down-regulated lines [66].
In mature tubers contrasting results were reported. Psy expression correlated with total carotenoid concentration in three yellow-fleshed cvs. [67] but not in a group of eight tetraploid varieties with contrasting flesh color [68].
No QTL for tuber color maps to the same genomic region as either Psy1 (SOLTU DM 03G002970) on Chr. 3, 40 Mb from the Chy2 locus, or Psy2 (SOLTU DM 02G020840), on Chr. 2, the genes coding for the two isoforms of potato PSY. This questions the existence of superior alleles at these loci that can be exploited in breeding programs for carotenoid biofortification of potato tubers.
3.1.2. Chy2
Several investigations pointed to Chy2 (or Bch2) as the putative gene corresponding to the Y locus [56,69,70].
Chy1 and Chy2 code for the two isoforms of potato β-carotene hydroxylase, the enzyme that converts β-carotene to zeaxanthin in the β,β-branch of the carotenoid pathway. Down-regulation of both genes in tubers led to an increase of β-carotene (up to 38-fold, 0.085 μg/g DW) and total carotenoids (up to 4.5-fold) with a concomitant decrease of zeaxanthin [71].
Conversely, high Chy2 expression, supposedly conditioned by the presence of a specific dominant allele (allele-3 or B), is associated with yellow tuber flesh color and determines a consistent flux through the β,β-branch of the carotenoid pathway leading to the synthesis of zeaxanthin and downstream xanthophylls, the main carotenoids in yellow-fleshed tubers [38,69,72,73]. Usually, the dominant Chy2 allele-3 is not found in white-fleshed cvs., which primarily accumulate lutein and β-carotene [56,70,74].
However, other genetic factors can override the effect of this allele on tuber colour and carotenoid accumulation given that white-fleshed clones harbouring Chy2 allele-3 as well as yellow ones lacking it were described [75,76]. For instance, a recent analysis of potato genome structure unravelled a 5.8 Mb paracentric inversion on the long arm of Chr. 3 spanning 464 genes, among which Chy2. An analysis of 22 diploid accessions found a strong association between tuber flesh color, Chy2 transcript level, and the orientation of the 5.8-Mb inversion, suggesting that this structural variation is a major determinant of tuber color through its effect on Chy2 transcription. The occurrence of Chy2 allele-3 in the genotypes examined was not investigated [77].
3.1.3. Zep
The metabolic step following CHY activity is the stepwise conversion of zeaxanthin to antheraxanthin and violaxanthin catalysed by zeaxanthin epoxidase (ZEP).
Orange-fleshed, high-carotenoid cvs., which are particularly rich in zeaxanthin, show low steady-state levels of Zep transcript compared to yellow and white-fleshed clones [35]. Accordingly, down-regulation of Zep in a low-carotenogenic variety led to a significant increase of zeaxanthin (up to 130-fold, 40 μg/g DW) but also of total tuber carotenoids (up to 5.7-fold) implying that the whole carotenoid biosynthetic pathway was affected [66].
Homozygosity for a recessive allele of this locus (zep allele-1) in the presence of the dominant Chy2 allele-3 determines the naturally occurring orange phenotype. zep allele-1 is usually found in the diploid germplasm whereas among a panel of 230 tetraploid cvs. it only occurred in 5 varieties, always in single copy, thus explaining the absence of the orange phenotype from the known tetraploid potato germplasm [72,78].
3.1.4. Lcye
LCYE catalyses the committed step in the β,ε-branch of the carotenoid pathway leading to lutein synthesis.
In potato, transcript levels of Lyce positively correlate with lutein concentration. Antisense-mediated Lyce silencing in a white-fleshed variety significantly raised levels of β,β-branch carotenoids, in particular β-carotene which showed a 14-fold increase (up to 0.0436 μg/g DW) [35,79].
No correlation between specific Lcye allelic variants disclosed in diploid potato germplasm and tuber flesh colour was found [72]. A thorough investigation of the tetraploid germplasm could clarify whether natural genetic variation at this locus can impact carotenoid content of potato.
3.2. Carotenoid. Degradation
3.2.1. Ccd4a
In Chrysanthemum (Chrysanthemum morifolium) the carotenoid cleavage dioxygenase encoded by the Ccd4a gene negatively regulates carotenoid accumulation in flowers [80].
Similarly, the potato homologous StCcd4a is involved in carotenoid degradation in tubers and flower petals, as assessed in RNAi studies [81]. In stable transformants, down-regulation of Ccd4a led to a 5.6-fold increase of total tuber carotenoids in the most strongly affected line without altering carotenogenic gene expression. Among the xanthophylls analysed in the RNAi lines, zeaxanthin showed the largest relative variation (up to 16x), suggesting that it may represent the primary substrate of CCD4a activity, although in vitro and in vivo studies pointed to β-carotene as the most likely candidate [82]. Suppressed lines often displayed aberrant phenotypes including elongated or chain tubers with a variable degree of premature spouting, ruling out Ccd4a silencing as an effective means to increase carotenoid levels in tubers of commercial varieties.
In agreement with its function in carotenoid degradation, in wt potatoes StCcd4a shows a higher expression in mature tubers from white cultivars compared to yellow, carotenoid-rich ones [81]. A negative correlation was also found between carotenoid concentration in tubers and transcript levels of Ccd1a, another member of the potato CCD family [41].
3.2.2. Lox
Plant lipoxygenases (LOXs) are dioxygenases that catalyse the oxidation of polyunsaturated fatty acids (PUFAs) leading to the synthesis of biologically active compounds, such as the defence-related metabolites jasmonic acid and methyl jasmonate [83]. The hydroperoxyl fatty acid intermediates produced by LOX activity can oxidate carotenoid molecules, targeting their conjugated double-bond systems [84,85]. In cereals, this co-oxidation process is responsible for β-carotene degradation during grain storage and processing [86,87].
LOX gene expression and activity were detected in potato tubers and related to the tuberization process [88,89,90]. In addition, LOX activity was found to increase in stored tubers [91]. Therefore, besides CHY2 and CCD4a, LOX might contribute to the observed turnover of β-carotene-derived xanthophylls during long-term cold storage [29,35].
3.3. Carotenoid. Storage
In cauliflower (Brassica oleracea) the gain-of-function Orange (Or) mutation induces β-carotene accumulation in otherwise unpigmented tissues by triggering the differentiation of non-colored plastids into chromoplasts without affecting the expression of carotenogenic genes [92,93]. The Or gene is highly conserved across the plant kingdom and encodes a plastid-associated DnaJ-like protein with a cysteine-rich zinc finger domain functional in protein-protein interaction.
Overexpression of the cauliflower Or allele in a low-carotenoid potato variety under the control of a tuber-specific promoter resulted in a six-fold increase of total tuber carotenoids and the rise of β-carotene levels up to 5 μg/g DW from negligible amounts in the untransformed control. This implies that also in potato sink capacity constrains carotenoid accumulation. Alternatively, the carotenoid sequestering structures found in the distinctive chromoplasts of Or-transformed tubers may help slow down carotenoid degradation. Or-transformed lines also showed enhanced carotenoid accumulation and stability during long term-cold storage [94,95]. Nevertheless, the genetic background conditions the outcome of the BoOr allele insertion into potato. Transforming two carotenoid-rich Phureja clones with the same construct used with the white-fleshed variety resulted in a limited increase of tuber carotenoids (up to 60%), mainly due to higher xanthophylls levels, while β-carotene concentration was almost unaffected [54].
A naturally occurring point mutation in the Or gene of melon (Cucumis melo), causing a single amino acid substitution, turned out to be functionally equivalent to the Or gain-of-function mutation of cauliflower [96]. Overexpression in Arabidopsis of the potato Or gene (StOr) or its mutagenized variant bearing the melon “golden SNP” mutation led to enhanced β-carotene accumulation and increased abiotic stress tolerance, making StOr a candidate target for the improvement of β-carotene accumulation and environmental stress adaptation in potato [97].
3.4. Additional Transgenic Strategies to Increase Tuber Carotenoid Content
3.4.1.“. Golden” Potato
The coordinated overexpression under a tuber-specific promoter of three bacterial genes coding for phytoene synthase (CrtB), phytoene desaturase (CrtI), and lycopene β-cyclase (CrtY) was reported to induce a 20–fold increase of total tuber carotenoids with outstanding amounts of β-carotene (up to 47 μg/g DW, the highest level ever recorded in potato tubers) [98].
This “push” strategy was aimed at relieving the main bottlenecks of carotenoid biosynthesis, enabling a steady metabolic flux through the pathway. Conversely, overexpression of lycopene β-cyclase (StLCYb) alone only doubled β-carotene levels [99].
3.4.2. Dxs
In another attempt to push potato carotenoid biosynthesis a bacterial gene coding for 1-deoxy-D-xylulose 5-phosphate synthase (DXS), the first enzyme of the MEP pathway, was overexpressed in tubers of a low-carotenogenic variety. Despite the relatively high Dxs transcript levels in some transformed lines, carotenoid content only doubled mainly due to a 7-fold increase in phytoene concentration whereas levels of downstream carotenoids were not significantly affected [100].
3.4.3. Astaxanthin
Genetic engineering of potato tubers was also employed to produce the valuable ketocarotenoid astaxanthin, which is not naturally synthetized in potatoes.
In one study, the CrtO ketolase gene from the cyanobacterium Synechocystis was overexpressed in a transgenic potato line high in zeaxanthin. Ketocarotenoids accumulated up to 10-12% of total carotenoids in both leaves and tubers. However, astaxanthin only accounted for 1.8% of the total carotenoid content [101].
Better results were obtained by transforming a high carotenogenic S. phureja clone with an algal β-carotene ketolase gene (bkt1): astaxanthin levels in tubers reached 14 μg/g DW (47% of total carotenoids) [102].
A further increase in astaxanthin levels was achieved by stacking in a selected S. phureja background the cauliflower Or allele and two bacterial genes coding for β-carotene hydroxylase (crtZ) and β-carotene ketolase (crtW). Compared to previous attempts with the bkt1 gene alone, a six-fold higher tuber astaxanthin content was obtained (up to 77 μg/g DW, a level considered nutritionally significant [54]).
4. Genome Editing
Until now, genome editing technologies have not been harnessed to improve tuber carotenoid concentration and composition because of the possible systemic side-effects of knocking out carotenogenesis genes, primarily on photosynthesis. Indeed, CRISPR-Cas9 silencing of the potato Pds gene led to albino mutants devoid of both carotenoids and chlorophylls, whose synthesis is coordinated [103].
In contrast, CRISPR/CAS9-based editing of the banana (Musa spp.) Lcye gene caused a drastic reduction of lutein, the most abundant carotenoid in chloroplasts, but with no or little effects on photosynthesis. Only a slight decrease in chlorophyll a/b ratio and reduced growth of fruit bunches were observed in the most affected knockout line grown under controlled conditions [104].
It should be noted, however, that in higher plants lutein is required for effective photoprotection only under strong light and the Arabidopsis lutein-deficient mutant lut2, impaired in LCYE activity, is viable under non-stress conditions. Likely, the observed increase in β,β-carotenoids, particularly those involved in the xanthophyll cycle (zeaxanthin, antheraxanthin, and violaxanthin) functionally compensates for lutein deficiency [105].
To avoid systemic side effects, genome editing could be used to target single components of multi-gene families displaying different expression patterns. For instance, the potato paralogue genes Chy1 and Chy2 encode the main β-carotene hydroxylase isoform of leaves and tubers respectively [71].
Alternatively, especially with polyploid genotypes, the production of heterozygous mutants with the proper balance of wt and edited alleles may lead to a modulation of the knockout effect on plant phenotype.
Genome editing could also be harnessed to replicate natural mutations with a proven effect on carotenoid accumulation, such as the Or mutation of melon, which boosts carotenoid storage without affecting photosynthesis. Another example is the zep-1 leaky mutation of potato, almost completely restricted to the diploid Phureja germplasm, which could be functionally reproduced in tetraploid commercial varieties to enhance tuber zeaxanthin content.
5. Regulation of Carotenoid Metabolism
Genetic manipulation of carotenoid accumulation in tubers has relied so far on a basic knowledge of the structural genes involved in carotenoid synthesis, storage, and turnover. Understanding the regulation of carotenoid metabolism is required to further extend the qualitative and quantitative improvement of tuber carotenoid content through biotechnological approaches.
The dynamic process of carotenoid accumulation in plants is the net result of biosynthesis, turnover, and sequestration capacity of a particular tissue or organ and is regulated by both environmental and developmental cues in coordination with related metabolic pathways, such as those sharing the same isoprenoid precursors [12].
Several transcription factors that transactivate specific metabolic genes of the carotenoid pathway have been described in both photosynthetic and non-photosynthetic organs of many plant species. However, most of these transcriptional regulators are not integral to the carotenoid pathway but have a broader role in signal transduction of both external and internal stimuli, including light, temperature, redox status, circadian clock, and phytohormones [107].
Additional mechanisms of control of carotenoid metabolism and sequestration act at the posttranscriptional and posttranslational levels. These include alternative mRNA splicing events, formation of multi-enzyme complexes and other protein-protein interactions, topological distribution of key enzymes within the plastids, and positive or negative feedback loops involving metabolite intermediates or end products [108,109].
Compared to other species like Arabidopsis and tomato, limited information is available on the regulation of potato carotenoid accumulation, a process which is likely to present distinctive features. Potato tubers are peculiar organs that have few equivalents in other cultivated or model plants [110] and the carotenoids they store, mostly xanthophylls, differ from those found in carotenoid-rich organs of closely related species such as tomato (Solanum lycopersicum, lycopene and β-carotene [111]) and pepper (Capsicum annuum, capsanthin, lutein and β-carotene [112]). Moreover, the underground growth habit of potato tubers rules out a direct role of light in the regulation of carotenoid metabolism, whereas in fruits, flowers, and photosynthetic organs light is a major cue [113,114].
As described above, metabolic engineering has shown that some structural genes are key in determining the amount and profile of tuber carotenoids and represent possible flux-controlling steps of natural carotenoid homeostasis. On the other hand, since most of the QTL identified so far underlying carotenoid accumulation in potato tubers are not linked to carotenogenic genes, a complex regulatory network is anticipated to control this metabolic process [73].
5.1. Transcriptional Regulation
Analysis of carotenoid gene expression has given limited information on the transcriptional regulation of carotenoid levels in tubers.
In many species PSY activity, as the committed step of carotenoid biosynthesis, is tightly regulated transcriptionally and post-transcriptionally. For example, during photomorphogenesis, a light-dependent pathway involving phytochrome and cryptochrome photoreceptors modulates Psy expression through transcriptional activators/repressors that bind G-box motifs in the Psy promoter. [64]. In potato, higher Psy transcript levels were found to correlate with higher carotenoid content in the early stages of tuberization [35] but no transcriptional regulator was shown to directly control Psy expression in response to developmental or environmental factors.
Chy2 is believed to play a more significant role in tuber carotenoid accumulation, and its mRNA steady-state levels correlate with carotenoid concentration in this storage organ [38,41]. A sugar-responsive cis-element in the promoter of the dominant Chy2 allele-3 together with relative high concentration of tuber glucose and sucrose was suggested to explain the increased transcription of this allele in a carotenoid-rich cultivar [41]. However, a subsequent study did not find any significant correlation between the levels of total or individual soluble sugars and the amount of tuber carotenoids [56]. The putative effect of the Chr. 3 genomic inversion on Chy2 expression has not been analysed in detail.
A transcriptional regulation was suggested also for the main carotenoid degradative gene of potato tubers, CCcd4a, given the negative relationship between its mRNA level and carotenoid concentration in tubers with contrasting flesh colours. [81]. Functional analysis of the Ccd4a promoter has not been reported.
5.2. Post-Transcriptional Regulation
Additional structural genes with a putative role in carotenoid regulation in tubers are Zep and Or.
Besides the expected increase in zeaxanthin levels, downregulation of Zep stimulates the whole carotenoid biosynthesis suggesting a broader role for this gene in controlling carotenoid accumulation in potato tubers [66]. Accordingly, an inverse correlation between Zep transcript level and total carotenoid accumulation was observed in a range of potato germplasm [35]. Similarly, the tomato high-pigment 3 (hp3) mutant impaired in ZEP activity shows higher carotenoid accumulation in fruit and leaves associated with enlargement of the plastid compartment size [115] and the zep mutation in the aba1-1 mutant of Arabidopsis conditions a 60% higher carotenoid content in leaves [116]. It is not known whether Zep silencing in potato determines an increased sink capacity for carotenoid accumulation, as is the case for the tomato hp3 mutant. The observed enhancement of Psy transcript levels in potato Zep-suppressed lines could be the main determinant of carotenoid overproduction. The natural recessive zep-1 mutation found in native orange fleshed high-carotenoid germplasm is due to the insertion of a non LTR-retrotransposon-like sequence in the first intron which may affect mRNA processing [38,72].
In addition to regulate plastid differentiation into chromoplasts, the OR protein was shown to control post-transcriptionally PSY level and activity in Arabidopsis by directly interacting with PSY in plastids [117]. In transgenic potato overexpressing the Or gene higher stability of PSY was observed pointing to a similar OR-dependent mechanism of PSY regulation promoting carotenoid biosynthesis [95].
6. Conclusions
Although French fries' consumption may be associated with an increased risk of obesity and type 2 diabetes [118], potatoes can help meet daily caloric requirements and food security in countries where undernutrition is a concern [119]. Moreover, they provide a variety of health-promoting micronutrients including various antioxidant metabolites like the tuber colouring pigments anthocyanins and carotenoids [7,120,121]. A negative correlation between tuber carotenoid and anthocyanin content was reported in native South American potato germplasm [30]. Nevertheless, pigmented varieties high in both types of antioxidants were described, suggesting that there is no reciprocal interference in the accumulation of these two classes of nutraceuticals [37,39].
Even though metabolic engineering of potato has been successfully employed to increase both total and specific tuber carotenoids, regulatory constraints and consumers’ diffidence towards GM food crops may limit the actual marketability of the engineered varieties [122,123,124].
Exploiting new plant breeding techniques (NPBTs), in particular genome editing, represents the next step in carotenoid biofortification of potatoes. Conventional genetic manipulation has already disclosed some major structural genes controlling the metabolic flux through the carotenoid pathway, making them the obvious targets of editing approaches. NPBTs have been effectively used for site-specific genome editing of agronomic and quality traits in tetraploid potato cultivars. Transient CRISPR-Cas9 expression or CRISPR-Cas9 ribonucleoprotein delivery into protoplasts has proven useful to regenerate edited clones devoid of foreign DNA [125]. Genome editing represents a potential tool to boost tuber β-carotene levels in a transgene-free background through the introduction of the gain-of-function Or mutation into the potato genome, provided the proper genotype is targeted.
The improvement of commercial potato varieties with conventional breeding approaches is hampered by their tetraploid nature, self-incompatibility, and high heterozygosity typical of an inbreeding-sensitive species. In addition, the widespread occurrence of toxic steroidal glycoalkaloids (SGA), primarily α-solanine and α-chaconine, among wild potato genotypes is a detrimental trait that must be counter-selected in breeding programs. Nevertheless, the genetic potential of wild potato germplasm has been successfully exploited to introgress resistance and quality genes into economically important cultivars, overcoming hybridization barriers based on differences in ploidy [3,126,127,128]. Potato breeding to enhance tuber carotenoid content can harness the high heritability of the trait and the wide genetic variation in carotenoid level and composition of the native germplasm [4,5,30,77]. For instance, the tetraploid variety Nagasaki Kogane with high levels of tuber carotenoids (845 μg/100g FW, mainly zeaxanthin and lutein) has a diploid, carotenoid-rich S. phureja genotype in its ancestry [129].
A recent breakthrough in the context of genomics-assisted breeding of potatoes is the sequencing of a few modern cultivars which has shed light on the complex structural and functional organization of their highly heterozygous autotetraploid genome [130,131,132]. The knowledge drawn from these sequencing efforts will ease the construction of homozygous potato genotypes devoid of the deleterious alleles linked to inbreeding depression and of the large genomic rearrangements underlying regions of extensive linkage disequilibrium. Indeed, ongoing efforts to redesign potato as a diploid, seed-propagated crop have the potential to overcome major drawbacks in potato breeding, easing the introgression of favourable genes into elite germplasm and opening the way to hybrid production [133,134].
Such advances, together with a deeper understanding of the regulatory mechanisms underlying carotenoid accumulation in tubers, may open a new era in the field of potato carotenoid biofortification, and further improve the nutritional value of this staple crop.
Funding
This research received no external funding
Conflicts of Interest
The author declares no conflicts of interest
References
- Available online: http://www.fao.org/faostat/en/#data (accessed on 22 June 2024).
- Spooner, D.M., Ghislain, M.; Simon, R.; Jansky, S.H.; Gavrilenko, T. Systematics, Diversity, Genetics, and Evolution of Wild and Cultivated Potatoes. Bot. Rev. 2014, 80, 283–383. doi: 10.1007/s12229-014-9146-y.
- Machida-Hirano, R. Diversity of potato genetic resources. Breed. Sci. 2015, 65(1), 26–40. doi: 10.1270/jsbbs.65.26.
- Brown C.R. Breeding for Phytonutrient Enhancement of Potato. Am. J. Pot. Res. 2008, 85, 298–307. [CrossRef]
- Andre, C.M.; Ghislain, M.; Bertin, P.; Oufir, M.; Herrera Mdel, R.; Hoffmann, L.; Hausman, J.F.; Larondelle, Y.; Evers, D. Andean potato cultivars (Solanum tuberosum L.) as a source of antioxidant and mineral micronutrients. J. Agric. Food Chem. 2007, 55(2), 366-378. [CrossRef]
- Camire, M.E.; Kubow, S.; Donnelly, D.J. Potatoes and human health. Crit. Rev. Food Sci. Nutr. 2009, 49(10), 823-840. [CrossRef]
- Singh, B.; Goutam, U.; Kukreja, S.; Sharma, J.; Sood, S.; Bhardwaj, V. Potato biofortification: an effective way to fight global hidden hunger. Physiol. Mol. Biol. Plants. 2021, 27(10), 2297-2313. [CrossRef]
- Zhu, C.; Bai, C., Sanahuja, G.; Yuan, D.; Farré, G.; Naqvi, S.; Shi, L.; Capell, T.; Christou, P. The regulation of carotenoid pigmentation in flowers. Arch. Biochem. Biophys. 2010, 504(1), 132-141. [CrossRef]
- Domonkos, I.; Kis, M.; Gombos, Z.; Ughy, B. Carotenoids, versatile components of oxygenic photosynthesis. Prog. Lipid Res. 2013, 52(4), 539-561. [CrossRef]
- Ahrazem, O.; Gómez-Gómez, L.; Rodrigo, M.J.; Avalos, J.; Limón, M.C. Carotenoid Cleavage Oxygenases from Microbes and Photosynthetic Organisms: Features and Functions. Int. J. Mol. Sci. 2016, 17(11), 1781. [CrossRef]
- Pogson, B.J.; Niyogi, K.K.; Björkman, O.; DellaPenna, D. Altered xanthophyll compositions adversely affect chlorophyll accumulation and nonphotochemical quenching in Arabidopsis mutants. Proc. Natl. Acad. Sci. U.S.A. 1998, 95(22), 13324-13329. [CrossRef]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid metabolism and regulation in horticultural crops. Hortic. Res. 2015, 2, 15036. [CrossRef]
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55(3), 207-216. [CrossRef]
- Carazo, A.; Macáková, K.; Matoušová, K.; Krčmová, L.K.; Protti, M.; Mladěnka, P. Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity. Nutrients. 2021, 13(5), 1703. [CrossRef]
- Mares, J. Lutein and Zeaxanthin Isomers in Eye Health and Disease. Annu. Rev. Nutr. 2016, 36, 571-602. [CrossRef]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017,174(11), 1290-1324. [CrossRef]
- Polidori, M.C.; Stahl, W.; Griffiths, H.R. Nutritional cognitive neuroscience of aging: Focus on carotenoids and cognitive frailty. Redox Biol. 2021, 44, 101996. [CrossRef]
- Cobbs, C.; Heath, J.; Stireman, J.O. 3rd; Abbot, P. Carotenoids in unexpected places: gall midges, lateral gene transfer, and carotenoid biosynthesis in animals. Mol. Phylogenet. Evol. 2013, 68(2), 221-228. [CrossRef]
- Barreiro, C.; Barredo, J.L. Carotenoids Production: A Healthy and Profitable Industry. Methods Mol. Biol. 2018,1852, 45-55. [CrossRef]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; Ribot, J.; Rodrigo, M.J.; Zacarias, L.; Zhu, C. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62-93. [CrossRef]
- Pasare, S.; Wright, K.; Campbell, R.; Morris, W.; Ducreux, L.; Chapman, S.; Bramley, P.; Fraser, P.; Roberts, A.; Taylor, M. The sub-cellular localisation of the potato (Solanum tuberosum L.) carotenoid biosynthetic enzymes, CrtRb2 and PSY2. Protoplasma. 2013, 250(6), 1381-1392. [CrossRef]
- Iwanzik, W.; Tevini, M.; Stute, R.; Hilbert, R. Carotinoidgehalt und-zusammensetzung verschiedener deutscher Kartoffelsorten und deren Bedeutung für die Fleischfarbe der Knolle. Potato Res. 1983, 26, 149–162. [CrossRef]
- Brown, C.R.; Edwards, C.G.; Yang, C.; Dean, B.B. Orange Flesh Trait in Potato: Inheritance and Carotenoid Content. J. Amer. Soc. Hort. Sci. 1993, 118(1), 145-150. doi: 10.21273/JASHS.118.1.145.
- Lu, W.; Haynes, K.; Wiley, E., Clevidence, B. Carotenoid Content and Color in Diploid Potatoes. J. Amer. Soc. Hort. Sci. 2001, 126(6), 722-726. doi: 10.21273/JASHS.126.6.722.
- Andre, C.M.; Oufir, M.; Guignard, C.; Hoffmann, L.; Hausman, J.F.; Evers, D.; Larondelle, Y. Antioxidant profiling of native Andean potato tubers (Solanum tuberosum L.) reveals cultivars with high levels of beta-carotene, alpha-tocopherol, chlorogenic acid, and petanin. J. Agric. Food Chem. 2007, 55(26), 10839-10849. [CrossRef]
- Burgos, G.; Salas, E.; Amoros, W.; Auqui, M.; Munoa, L.; Kimura, M.; Bonierbale, M. Total and individual carotenoid profiles in Solanum phureja of cultivated potatoes: I. Concentrations and relationships as determined by spectrophotometry and HPLC. J. Food Comp. Anal. 2009, 22, 503–508. doi: 10.1016/j.jfca.2008.08.008.
- Tatarowska, B., Milczarek, D.; Wszelaczyńska, E.; Pobereżny, J.; Keutgen, N.; Keutgen, A.J.; Flis, B. Carotenoids Variability of Potato Tubers in Relation to Genotype, Growing Location and Year. Am. J. Potato Res. 2019, 96, 493–504. [CrossRef]
- Fernandez-Orozco, R.; Gallardo-Guerrero, L.; Hornero-Méndez, D. Carotenoid profiling in tubers of different potato (Solanum sp) cultivars: accumulation of carotenoids mediated by xanthophyll esterification. Food Chem. 2013, 141(3), 2864-72. [CrossRef]
- Griffiths, D.W.; Dale, M.F.; Morris, W.L.; Ramsay, G. Effects of season and postharvest storage on the carotenoid content of Solanum phureja potato tubers. J. Agric. Food Chem. 2007, 55(2), 379-85. [CrossRef]
- Brown, C.R.; Culley, D.; Bonierbale, M.; Amoro´s, W. Anthocyanin, carotenoid content, and antioxidant values in native South American potato cultivars. HortScience. 2007, 42, 1733–1736. doi: 10.21273/HORTSCI.42.7.1733.
- Bonierbale, M.; Grüneberg, W.; Amoros, W.; Burgos, G., Salas, E.; Porras, E.; Zum Felde, T. Total and individual carotenoid profiles in Solanum phureja cultivated potatoes: II. Development and application of near-infrared reflectance spectroscopy (NIRS) calibrations for germplasm characterization. J. Food Compos. Anal. 2009, 22, 509–516. [CrossRef]
- Cuéllar-Cepeda, F.A.; Parra-Galindo, M.A.; Urquijo, J.; Restrepo-Sánchez, L.P.; Mosquera-Vásquez, T.; Narváez-Cuenca, C.E. Influence of genotype, agro-climatic conditions, cooking method, and their interactions on individual carotenoids and hydroxycinnamic acids contents in tubers of diploid potatoes. Food Chem. 2019, 288, 127-138. [CrossRef]
- Breithaupt, D.E.; Bamedi, A. Carotenoids and carotenoid esters in potatoes (Solanum tuberosum L.): new insights into an ancient vegetable. J. Agric. Food Chem. 2002, 50(24), 7175-7181. [CrossRef]
- Nesterenko, S.; Sink, K.C. Carotenoid Profiles of Potato Breeding Lines and Selected Cultivars. HortScience. 2003, 38(6), 1173-1177. doi: 10.21273/HORTSCI.38.6.1173.
- Morris, W.L.; Ducreux, L.; Griffiths, D.W.; Stewart, D.; Davies, H.V.; Taylor, M.A. Carotenogenesis during tuber development and storage in potato. J. Exp. Bot. 2004, 55(399), 975-82. [CrossRef]
- Haynes, K.G.; Clevidence, B.A.; Rao, D.; Vinyard, B.T.; White, J.M. Genotype × Environment Interactions for Potato Tuber Carotenoid Content. J. Amer. Soc. Hort. Sci. 2010, 135(3), 250-258. doi: 10.21273/JASHS.135.3.250.
- Hejtmánková, K.; Kotikova, Z.; Hamouz, K.; Pivec ,V.; Vacek, J.; Lachman, J. Influence of flesh colour, year and growing area on carotenoid and anthocyanin content in potato tubers. J. Food Compos. Anal. 2013, 32, 20–27. doi: 10.1016/j.jfca.2013.07.001.
- Sulli, M.; Mandolino, G.; Sturaro, M.; Onofri, C.; Diretto, G.; Parisi, B.; Giuliano, G. Molecular and biochemical characterization of a potato collection with contrasting tuber carotenoid content. PLoS One. 2017, 12(9), e0184143. [CrossRef]
- Kotíková, Z.; Šulc, M.; Lachman, J.; Pivec, V.; Orsák, M.; Hamouz, K. Carotenoid profile and retention in yellow-, purple- and red-fleshed potatoes after thermal processing. Food Chem. 2016, 197(Pt A), 992-1001. [CrossRef]
- Kobayashi, A.; Ohara-Takada, A.; Tsuda, S.; Matsuura-Endo, C.; Takada, N.; Umemura, Y.; Nakao, T.; Yoshida, T.; Hayashi, K.; Mori, M. Breeding of potato variety “Inca-no-hitomi” with a very high carotenoid content. Breed. Science. 2008, 58(1), 77-82. [CrossRef]
- Zhou, X.; McQuinn, R.; Fei, Z.; Wolters, A.A.; Van Eck, J.; Brown, C.; Giovannoni, J.J.; Li, L.I. Regulatory control of high levels of carotenoid accumulation in potato tubers. Plant Cell Environ. 2011, 34(6), 1020-1030. [CrossRef]
- Kotíková, Z.; Hejtmánková, A.; Lachman, J.; Hamouz, K.; Trnková, E.; Dvořák, P. Effect of selected factors on total carotenoid content in potato tubers (Solanum tuberosum L.). Plant Soil Environ. 2007, 53(8), 355-360. doi: 10.17221/2214-PSE.
- Reddivari, L.; Hale, A.L.; Miller, J.C. Jr. Genotype, location, and year influence antioxidant activity, carotenoid content, phenolic content, and composition in specialty potatoes. J. Agric. Food Chem. 2007, 55(20), 8073-8079. [CrossRef]
- Haynes, K.G.; Clevidence, B.A.; Rao, D.; Vinyard, B.T. Inheritance of Carotenoid Content in Tetraploid × Diploid Potato Crosses. J. Amer. Soc. Hort. Sci. 2011, 136(4), 265-272. doi: 10.21273/JASHS.136.4.265.
- Payyavula, R.S.; Navarre, D.A.; Kuhl, J.C.; Pantoja, A.; Pillai, S.S. Differential effects of environment on potato phenylpropanoid and carotenoid expression. BMC Plant Biol. 2012, 12, 39. [CrossRef]
- Hamouz, K.; Pazderů, K.; Lachman, J.; Čepl, J.; Kotíková, Z. Effect of cultivar, flesh colour, locality and year on carotenoid content in potato tubers. Plant Soil Environ. 2016, 62(2), 86-91. [CrossRef]
- Tatarowska, B.; Milczarek, D.; Jakuczun, H.; Stochmal, A.; Pecio, Ł.; Flis, B. The potential for the improvement of carotenoid level in potato-effect of the genotype and environment. J. Food Agric. Environ. 2014, 12(2), 536-540. doi: 10.1234/4.2014.5193.
- Vaitkevičienė, N.; Kulaitienė, J.; Jarienė, E.; Levickienė, D.; Danillčenko, H.; Średnicka-Tober, D.; Rembiałkowska, E.; Hallmann, E. Characterization of Bioactive Compounds in Colored Potato (Solanum Tuberosum L.) Cultivars Grown with Conventional, Organic, and Biodynamic Methods. Sustainability. 2020, 12(7), 2701. [CrossRef]
- Kazimierczak, R.; Średnicka-Tober, D.; Hallmann, E.; Kopczyńska, K.; Zarzyńska, K. The impact of organic vs. conventional agricultural practices on selected quality features of eight potato cultivars. Agronomy. 2019, 9(12), 799. doi: 10.3390/agronomy9120799.
- Blessington, T.; Nzaramba, M.N.; Scheuring, D.C.; Hale, A.L.; Reddivari, L.; Miller, J.C. Cooking Methods and Storage Treatments of Potato: Effects on Carotenoids, Antioxidant Activity, and Phenolics. Am. J. Pot. Res. 2010, 87, 479–491. [CrossRef]
- Burgos, G.; Amoros, W.; Salas, E.; Muñoa, L.; Sosa, P.; Díaz, C.; Bonierbale, M. Carotenoid concentrations of native Andean potatoes as affected by cooking. Food Chem. 2012, 133(4), 1131-1137. doi: 10.1016/j.foodchem.2011.09.002.
- Tian, J.; Chen, J.; Lv, F.; Chen, S.; Chen, J.; Liu, D.; Ye, X. Domestic cooking methods affect the phytochemical composition and antioxidant activity of purple-fleshed potatoes. Food Chem. 2016, 197 Pt B, 1264-1270. [CrossRef]
- Andre, C.M.; Schafleitner, R.; Guignard, C.; Oufir, M.; Aliaga, C.A.; Nomberto, G.; Hoffmann, L.; Hausman, J.F.; Evers D.; Larondelle Y. Modification of the health-promoting value of potato tubers field grown under drought stress: emphasis on dietary antioxidant and glycoalkaloid contents in five native andean cultivars (Solanum tuberosum L.). J. Agric. Food Chem. 2009, 57(2), 599-609. [CrossRef]
- Campbell, R.; Morris, W.L.; Mortimer, C.L.; Misawa, N.; Ducreux, L.J.; Morris, J.A.; Hedley, P.E.; Fraser, P.D.; Taylor, M.A. Optimising ketocarotenoid production in potato tubers: effect of genetic background, transgene combinations and environment. Plant Sci. 2015, 234, 27-37. [CrossRef]
- Bonierbale, M.W.; Plaisted, R.L.; Tanksley, S.D. RFLP Maps Based on a Common Set of Clones Reveal Modes of Chromosomal Evolution in Potato and Tomato. Genetics. 1988, 120(4), 1095-1103. [CrossRef]
- Campbell, R.; Pont, S.D.; Morris, J.A.; McKenzie, G.; Sharma, S.K.; Hedley, P.E.; Ramsay, G.; Bryan, G.J.; Taylor, M.A. Genome-wide QTL and bulked transcriptomic analysis reveals new candidate genes for the control of tuber carotenoid content in potato (Solanum tuberosum L.). Theor. Appl. Genet. 2014, 127(9), 1917-1933. [CrossRef]
- D'hoop, B.B.; Keizer, P.L.; Paulo, M.J.; Visser, R.G.; van Eeuwijk, F.A.; van Eck, H.J. Identification of agronomically important QTL in tetraploid potato cultivars using a marker-trait association analysis. Theor. Appl. Genet. 2014, 127(3), 731-748. [CrossRef]
- Sharma, S.K.; MacKenzie, K.; McLean, K.; Dale, F.; Daniels, S.; Bryan, G.J. Linkage Disequilibrium and Evaluation of Genome-Wide Association Mapping Models in Tetraploid Potato. G3 (Bethesda). 2018, 8(10), 3185-3202. [CrossRef]
- Pandey, J.; Scheuring, D.C.; Koym, J.W.; Vales, M.I. Genomic regions associated with tuber traits in tetraploid potatoes and identification of superior clones for breeding purposes [published correction appears in Front. Plant. Sci. 2024, 22, 15:1396479. doi: 10.3389/fpls.2024.1396479]. Front. Plant Sci. 2022, 13, 952263. [CrossRef]
- D’hoop, B.B.; Paulo, M.J.; Mank, R.; van Eck, H.J.; van Eeuwijk, F.A. Association mapping of quality traits in potato (Solanum tuberosum L.). Euphytica. 2008, 161, 47–60. [CrossRef]
- Śliwka, J.; Wasilewicz-Flis, I.; Jakuczun, H.; Gebhardt, C. Tagging quantitative trait loci for dormancy, tuber shape, regularity of tuber shape, eye depth and flesh colour in diploid potato originated from six Solanum species. Plant Breed. 2008, 127, 49-55. doi: 10.1111/j.1439-0523.2008.01420.x.
- Liu, Y.S.; Gur, A.; Ronen, G.; Causse, M.; Damidaux, R.; Buret, M.; Hirschberg, J.; Zamir, D. There is more to tomato fruit colour than candidate carotenoid genes. Plant Biotechnol. J. 2003, 1(3), 195-207. [CrossRef]
- Thorup, T.A.; Tanyolac, B.; Livingstone, K.D.; Popovsky, S.; Paran, I.; Jahn, M. Candidate gene analysis of organ pigmentation loci in the Solanaceae. Proc. Natl. Acad. Sci. U.S.A. 2000, 97(21), 11192-11197. [CrossRef]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15(5), 266-274. [CrossRef]
- Ducreux, L.J.; Morris, W.L.; Hedley, P.E.; Shepherd, T.; Davies, H.V.; Millam, S.; Taylor, M.A. Metabolic engineering of high carotenoid potato tubers containing enhanced levels of beta-carotene and lutein. J. Exp. Bot. 2005, 56(409), 81-89. [CrossRef]
- Römer, S.; Lübeck, J.; Kauder, F.; Steiger, S.; Adomat, C.; Sandmann, G. Genetic engineering of a zeaxanthin-rich potato by antisense inactivation and co-suppression of carotenoid epoxidation. Metab. Eng. 2002, 4(4), 263-272. doi:.10.1006/mben.2002.0234.
- Goo, Y.M.; Kim, T.W.; Ha, S.H.; Back, K.W.; Bae, J.M.; Shin, Y.W.; Lee, C.H.; Ahn, M.J.; Lee, S.W. Expression Profiles of Genes Involved in the Carotenoid Biosynthetic Pathway in Yellow-Fleshed Potato Cultivars (Solanum tuberosum L.) from South Korea. J. Plant Biol. 2009, 52, 49–55. doi: 10.1007/s12374-008-9003-9.
- Valcarcel, J.; Reilly, K.; Gaffney, M.; O'Brien, N.M. Levels of potential bioactive compounds including carotenoids, vitamin C and phenolic compounds, and expression of their cognate biosynthetic genes vary significantly in different varieties of potato (Solanum tuberosum L.) grown under uniform cultural conditions. J. Sci. Food Agric. 2016, 96(3), 1018-1026. [CrossRef]
- Brown, C.R.; Kim, T.S.; Ganga, Z.; Haynes, K.; De Jong, D.; Jahn, M.; Paran, I.; De Jong, W. Segregation of total carotenoid in high level potato germplasm and its relationship to beta-carotene hydroxylase polymorphism. Am. J. Pot. Res. 2006, 83, 365–372. [CrossRef]
- Kloosterman, B.; Oortwijn, M.; uitdeWilligen, J.; America, T.; de Vos, R.; Visser, R.G.; Bachem, C.W. From QTL to candidate gene: genetical genomics of simple and complex traits in potato using a pooling strategy. BMC Genomics. 2010, 11, 158. [CrossRef]
- Diretto, G.; Welsch, R.; Tavazza, R.; Mourgues, F.; Pizzichini, D.; Beyer, P.; Giuliano, G. Silencing of beta-carotene hydroxylase increases total carotenoid and beta-carotene levels in potato tubers. BMC Plant Biol. 2007, 7, 11. [CrossRef]
- Wolters, A.M.; Uitdewilligen, J.G.; Kloosterman, B.A.; Hutten, R.C.; Visser, R.G.; van Eck, H.J. Identification of alleles of carotenoid pathway genes important for zeaxanthin accumulation in potato tubers. Plant Mol. Biol. 2010, 73(6), 659-671. [CrossRef]
- Zhang, Z.; Zhou, D.; Li, S.; Pan, J.; Liang, J.; Wu, X.; Wu, X.N.; Krall, L.; Zhu, G. Multiomics Analysis Reveals the Chemical and Genetic Bases of Pigmented Potato Tuber. J. Agric. Food Chem. 2003, 71(43), 16402–16416. [CrossRef]
- Van Eck, J.; Conlin, B.; Garvin, D.F.; Mason, H.; Navarre, D.A.; Brown, C.R. Enhancing beta-carotene content in potato by rnai-mediated silencing of the beta-carotene hydroxylase gene. Amer. J. Potato Res. 2007, 84, 331–342. [CrossRef]
- Brown, C.R.; Durst, R.W.; Wrolstad, R.; De Jong, W. Variability of Phytonutrient Content of Potato in Relation to Growing Location and Cooking Method. Potato Res. 2008, 51, 259–270. doi: 10.1007/s11540-008-9115-0.
- Tatarowska, B.; Milczarek, D.; Plich, J. The Content of Total Carotenoids, Vitamin C and Antioxidant Properties of 65 Potato Cultivars Characterised under the European Project ECOBREED. Int. J. Mol. Sci. 2023, 24(14), 11716. [CrossRef]
- Tang, D.; Jia, Y.; Zhang, J.; Li, H.; Cheng, L.; Wang, P.; Bao, Z.; Liu, Z.; Feng, S.; Zhu, X.; Li, D.; Zhu, G.; Wang, H.; Zhou, Y.; Zhou, Y.; Bryan, G.J.; Buell, C.R.; Zhang, C.; Huang, S. Genome evolution and diversity of wild and cultivated potatoes. Nature. 2022, 606(7914), 535-541. [CrossRef]
- McCord, P.; Zhang, L.; Brown, C. The Incidence and Effect on Total Tuber Carotenoids of a Recessive Zeaxanthin Epoxidase Allele (Zep1) in Yellow-fleshed Potatoes. Am. J. Pot. Res. 2012, 89, 262–268. doi: 10.1007/s12230-012-9250-7.
- Diretto, G.; Tavazza, R.; Welsch, R.; Pizzichini, D.; Mourgues, F.; Papacchioli, V.; Beyer, P.; Giuliano, G. Metabolic engineering of potato tuber carotenoids through tuber-specific silencing of lycopene epsilon cyclase. BMC Plant Biol. 2006, 6, 13. [CrossRef]
- Ohmiya, A.; Kishimoto, S.; Aida, R.; Yoshioka, S.; Sumitomo, K. Carotenoid cleavage dioxygenase (CmCCD4a) contributes to white color formation in chrysanthemum petals [published correction appears in Plant Physiol. 2016 Oct;172(2):1353doi: 10.1104/pp.16.01388]. Plant Physiol. 2006, 142(3), 1193-1201. [CrossRef]
- Campbell, R.; Ducreux, L.J.; Morris, W.L.; Morris, J.A.; Suttle, J.C.; Ramsay, G.; Bryan, G.J.; Hedley, P.E.; Taylor, M.A. The metabolic and developmental roles of carotenoid cleavage dioxygenase4 from potato. Plant Physiol. 2010, 154(2), 656-664. [CrossRef]
- Bruno, M.; Beyer, P.; Al-Babili, S. The potato carotenoid cleavage dioxygenase 4 catalyzes a single cleavage of β-ionone ring-containing carotenes and non-epoxidated xanthophylls. Arch. Biochem. Biophys. 2015, 572, 126-133. [CrossRef]
- Babenko, L.M.; Shcherbatiuk, M.M.; Skaterna, T.D.; Kosakivska, I.V. Lipoxygenases and their metabolites in formation of plant stress tolerance. Ukr. Biochem. J. 2017, 89(1), 5–21. [CrossRef]
- Wu, Z.; Robinson, D.S.; Hughes, R.K.; Casey, R.; Hardy, D.; West, S.I. Co-oxidation of beta-carotene catalyzed by soybean and recombinant pea lipoxygenases. J. Agric. Food Chem. 1999, 47(12), 4899–4906. [CrossRef]
- Chedea, V. S.; Jisaka, M. Lipoxygenase and carotenoids: A co-oxidation story. Afr. J. Biotechnol. 2013, 12(20), 2786-2791. [CrossRef]
- Gayen, D.; Ali, N.; Sarkar, S.N.; Datta, S.K.; Datta, K. Down-regulation of lipoxygenase gene reduces degradation of carotenoids of golden rice during storage. Planta. 2015, 242(1), 353-363. [CrossRef]
- Leenhardt, F.; Lyan, B.; Rock, E.; Boussard, A.; Potus, J.; Chanliaud, E.; Remesy, C. Wheat lipoxygenase activity induces greater loss of carotenoids than vitamin E during breadmaking. J. Agri. Food Chem. 2006, 54,1710–1715. [CrossRef]
- Galliard, T.; Phillips, D.R. Lipoxygenase from potato tubers. Partial purification and properties of an enzyme that specifically oxygenates the 9-position of linoleic acid. Biochem. J. 1971, 124(2), 431-438. [CrossRef]
- Royo, J.; Vancanneyt, G.; Pérez, A.G.; Sanz, C.; Störmann, K.; Rosahl, S.; Sánchez-Serrano, J.J. Characterization of three potato lipoxygenases with distinct enzymatic activities and different organ-specific and wound-regulated expression patterns. J. Biol. Chem. 1996, 271(35), 21012-21019. [CrossRef]
- Kolomiets, M.V.; Hannapel, D.J.; Chen, H.; Tymeson, M.; Gladon, R.J. Lipoxygenase is involved in the control of potato tuber development. Plant Cell. 2001, 13(3), 613-626. [CrossRef]
- Petersen, M.A.; Poll, L.; Larsen, L.M. Changes in flavor-affecting aroma compounds during potato storage are not associated with lipoxygenase activity. Am. J. Pot. Res. 2003, 80, 397–402. [CrossRef]
- Li, L.; Paolillo, D.J.; Parthasarathy, M.V.; Dimuzio, E.M.; Garvin, D.F. A novel gene mutation that confers abnormal patterns of beta-carotene accumulation in cauliflower (Brassica oleracea var. botrytis). Plant J. 2001, 26(1), 59-67. [CrossRef]
- Lu, S.; Van Eck, J.; Zhou, X.; Lopez, A.B.; O'Halloran, D.M.; Cosman, K.M.; Conlin, B.J.; Paolillo, D.J.; Garvin, D.F.; Vrebalov, J.; Kochian, L.V.; Küpper, H.; Earle, E.D.; Cao, J.; Li, L. The cauliflower Or gene encodes a DnaJ cysteine-rich domain-containing protein that mediates high levels of beta-carotene accumulation. Plant Cell. 2006, 18(12), 3594-3605. [CrossRef]
- Lopez, A.B.; Van Eck, J.; Conlin, B.J.; Paolillo, D.J.; O'Neill, J.; Li, L. Effect of the cauliflower Or transgene on carotenoid accumulation and chromoplast formation in transgenic potato tubers. J. Exp. Bot. 2008, 59(2), 213-223. [CrossRef]
- Li, L.; Yang, Y.; Xu, Q.; Owsiany, K.; Welsch, R.; Chitchumroonchokchai, C.; Lu, S.; Van Eck, J.; Deng, X.X.; Failla, M.; Thannhauser, T.W. The Or gene enhances carotenoid accumulation and stability during post-harvest storage of potato tubers. Mol. Plant. 2012, 5(2), 339-352. [CrossRef]
- Tzuri, G.; Zhou, X.; Chayut, N.; Yuan, H.; Portnoy, V.; Meir, A.; Sa'ar, U.; Baumkoler, F.; Mazourek, M.; Lewinsohn, E.; Fei, Z.; Schaffer, A.A.; Li, L.; Burger, J.; Katzir, N.; Tadmor, Y. A 'golden' SNP in CmOr governs the fruit flesh color of melon (Cucumis melo) [published correction appears in Plant J. 2015 Sep;83(5):940doi: 10.1111/tpj.12950]. Plant J. 2015, 82(2), 267-279. [CrossRef]
- Kang, L.; Zhang, C.; Liu, J.; Ye, M.; Zhang, L.; Chen, F.; Lin, X.; Yang, D.; Ren, L.; Li, Y.; Kim, H.S.; Kwak, S.S.; Li, H.; Deng, X.; Zhang, P.; Ke, Q. Overexpression of potato ORANGE (StOR) and StOR mutant in Arabidopsis confers increased carotenoid accumulation and tolerance to abiotic stress. Plant Physiol. Biochem. 2023, 201, 107809. [CrossRef]
- Diretto, G.; Al-Babili, S.; Tavazza, R.; Papacchioli, V.; Beyer, P.; Giuliano G. Metabolic engineering of potato carotenoid content through tuber-specific overexpression of a bacterial mini-pathway. PLoS One. 2007, 2(4), e350. [CrossRef]
- Song, X.Y.; Zhu, W.J.; Tang, R.M.; Cai, J.H.; Chen, M.; Yang, Q. Over-expression of StLCYb increases β-carotene accumulation in potato tubers. Plant Biotechnol. Rep. 2016, 10, 95-104. [CrossRef]
- Morris, W.L.; Ducreux, L.J.; Hedden, P.; Millam, S.; Taylor, M.A. Overexpression of a bacterial 1-deoxy-D-xylulose 5-phosphate synthase gene in potato tubers perturbs the isoprenoid metabolic network: implications for the control of the tuber life cycle. J. Exp. Bot. 2006, 57(12), 3007-3018. [CrossRef]
- Gerjets, T.; Sandmann, G. Ketocarotenoid formation in transgenic potato. J. Exp. Bot. 2006, 57(14), 3639-3645. [CrossRef]
- Morris, W.L.; Ducreux, L.J.; Fraser, P.D.; Millam, S.; Taylor, M.A. Engineering ketocarotenoid biosynthesis in potato tubers. Metab. Eng. 2006, 8(3), 253-263. [CrossRef]
- Butler, N.M.; Jansky, S.H.; Jiang, J. First-generation genome editing in potato using hairy root transformation. Plant Biotechnol. J. 2020, 18(11), 2201-2209. [CrossRef]
- Kaur, N.; Alok, A.; Shivani; Kumar, P.; Kaur, N.; Awasthi, P.; Chaturvedi, S.; Pandey, P.; Pandey, A.; Pandey, A.K.; Tiwari, S. CRISPR/Cas9 directed editing of lycopene epsilon-cyclase modulates metabolic flux for β-carotene biosynthesis in banana fruit. Metab. Eng. 2020, 59, 76-86. [CrossRef]
- Pogson, B.; McDonald, K.A.; Truong, M.; Britton, G.; DellaPenna, D. Arabidopsis carotenoid mutants demonstrate that lutein is not essential for photosynthesis in higher plants. Plant Cell. 1996, 8(9), 1627-1639. [CrossRef]
- Dall'Osto, L.; Lico, C.; Alric, J.; Giuliano, G.; Havaux, M.; Bassi, R. Lutein is needed for efficient chlorophyll triplet quenching in the major LHCII antenna complex of higher plants and effective photoprotection in vivo under strong light. BMC Plant Biol. 2006, 6, 32. [CrossRef]
- Stanley, L.; Yuan, Y.W. Transcriptional Regulation of Carotenoid Biosynthesis in Plants: So Many Regulators, So Little Consensus. Front. Plant Sci. 2019, 10, 1017. [CrossRef]
- Sun, T.; Li, L. Toward the 'golden' era: The status in uncovering the regulatory control of carotenoid accumulation in plants. Plant Sci. 2020, 290, 110331. [CrossRef]
- Kachanovsky, D.E.; Filler, S.; Isaacson, T.; Hirschberg, J. Epistasis in tomato color mutations involves regulation of phytoene synthase 1 expression by cis-carotenoids. Proc. Natl. Acad. Sci. U.S. A. 2012, 109(46), 19021-19026. [CrossRef]
- Zierer, W.; Rüscher, D.; Sonnewald, U., Sonnewald, S. Tuber and Tuberous Root Development. Annu. Rev. Plant Biol. 2021, 72, 551-580. [CrossRef]
- Beecher, GR. Nutrient content of tomatoes and tomato products. Proc. Soc. Exp. Biol. Med. 1998, 218(2), 98-100. [CrossRef]
- Ha, S.H.; Kim, J.B.; Park, J.S.; Lee, S.W.; Cho, K.J. A comparison of the carotenoid accumulation in Capsicum varieties that show different ripening colours: deletion of the capsanthin-capsorubin synthase gene is not a prerequisite for the formation of a yellow pepper. J. Exp. Bot. 2007, 58(12), 3135-3144. [CrossRef]
- Liu, Y.; Roof, S.; Ye, Z.; Barry, C.; van Tuinen, A.; Vrebalov, J.; Bowler, C., Giovannoni, J. Manipulation of light signal transduction as a means of modifying fruit nutritional quality in tomato. Proc. Natl. Acad. Sci. U.S.A. 2004, 101(26), 9897-9902. [CrossRef]
- Llorente, B.; Martinez-Garcia, J.F.; Stange, C.; Rodriguez-Concepcion, M.. Illuminating colors: regulation of carotenoid biosynthesis and accumulation by light. Curr. Opin. Plant Biol. 2017, 37, 49-55. [CrossRef]
- Galpaz, N.; Wan,g Q.; Menda, N.; Zamir, D.; Hirschberg, J. Abscisic acid deficiency in the tomato mutant high-pigment 3 leading to increased plastid number and higher fruit lycopene content. Plant J. 2008, 53(5), 717-730. [CrossRef]
- Duckham, S.C.; Linforth, R.S.T.; Taylor, I.B. (1991), Abscisic-acid-deficient mutants at the aba gene locus of Arabidopsis thaliana are impaired in the epoxidation of zeaxanthin. Plant Cell Env. 1991, 14, 601-606. doi: 10.1111/j.1365-3040.1991.tb01531.x.
- Zhou, X., Welsch; R., Yang; Y., Álvarez, D.; Riediger, M.; Yuan, H.; Fish, T.; Liu, J.; Thannhauser, T.W.; Li, L. Arabidopsis OR proteins are the major posttranscriptional regulators of phytoene synthase in controlling carotenoid biosynthesis. Proc. Natl. Acad. Sci. U.S.A. 2015, 112(11), 3558-63. [CrossRef]
- Borch, D.; Juul-Hindsgaul, N.; Veller, M.; Astrup, A.; Jaskolowski, J.; Raben, A. Potatoes and risk of obesity, type 2 diabetes, and cardiovascular disease in apparently healthy adults: a systematic review of clinical intervention and observational studies. Am. J. Clin. Nutr. 2016, 104(2), 489-498. [CrossRef]
- Zaheer, K.; Akhtar, M.H. Potato Production, Usage, and Nutrition--A Review. Crit. Rev. Food Sci. Nutr. 2016, 56(5), 711-21. [CrossRef]
- Brown, C.R.; Culley, D.; Yang, C.; Durst, R.; Wrolstad, R. Variation of Anthocyanin and Carotenoid Contents and Associated Antioxidant Values in Potato Breeding Lines. J. Amer. Soc. Hort. Sci. 2005, 130(2), 174-180. [CrossRef]
- Bvenura, C.; Witbooi, H.; Kambizi, L. Pigmented Potatoes: A Potential Panacea for Food and Nutrition Security and Health?. Foods. 2022, 11(2), 175. [CrossRef]
- Podevin, N.; Devos, Y.; Davies, H.V.; Nielsen, K.M. Transgenic or not? No simple answer! New biotechnology-based plant breeding techniques and the regulatory landscape. EMBO Rep. 2012, 13(12), 1057-1061. [CrossRef]
- Ahmad, A.; Munawar, N.; Khan, Z.; Qusmani, A.T.; Khan, S.H.; Jamil, A.; Ashraf, S.; Ghouri, M.Z.; Aslam, S.; Mubarik, M.S.; Munir, A.; Sultan, Q.; Abd-Elsalam, K.A.; Qari, S.H. An Outlook on Global Regulatory Landscape for Genome-Edited Crops. Int. J. Mol. Sci. 2021, 22(21), 11753. [CrossRef]
- Entine, J.; Felipe, M.S.S.; Groenewald, J.H.; Kershen, D.L.; Lema, M.; McHughen, A.; Nepomuceno, A.L.; Ohsawa, R.; Ordonio, R.L.; Parrott, W.A.; Quemada, H.; Ramage, C.; Slamet-Loedin, I.; Smyth, S.J.; Wray-Cahen, D. Regulatory approaches for genome edited agricultural plants in select countries and jurisdictions around the world. Transgenic Res. 2021, 30(4), 551-584. [CrossRef]
- Qu, L.; Huang, X.; Su, X.; Zhu, G.; Zheng, L.; Lin, J.; Wang, J.; Xue H. Potato: from functional genomics to genetic improvement. Mol. Hortic. 2024, 4(1), 34. [CrossRef]
- Jansky, S. Overcoming hybridization barriers in potato. Plant breed. 2006, 125(1), 1-12. doi: 10.1111/j.1439-0523.2006.01178.x.
- Krunic, S.L.; Skryhan, K.; Mikkelsen, L.; Ruzanski, C.; Shaik, S.S.; Kirk, H.G.; Palcic, M.; Blennow, A. Non-GMO potato lines with an altered starch biosynthesis pathway confer increased-amylose and resistant starch properties. Starch-Stärke. 2018, 70(1-2), 1600310. doi: 10.1002/star.201600310.
- Rakosy-Tican, E.; Thieme, R.; König, J.; Nachtigall, M.; Hammann, T.; Denes, T.E.; Kruppa, K.; Molnár-Láng, M. Introgression of Two Broad-Spectrum Late Blight Resistance Genes, Rpi-Blb1 and Rpi-Blb3, From Solanum bulbocastanum Dun Plus Race-Specific R Genes Into Potato Pre-breeding Lines. Front. Plant Sci. 2020, 11, 699. [CrossRef]
- Sakamoto, Y.; Mori, K.; Matsuo, Y.; Mukojima, N.; Watanabe, W.; Sobaru, N.; Tamiya, S.; Nakao, T.; Hayashi, K.; Watanuki, H.; Nara, K.; Yamazaki, K.; Chaya, M. Breeding of a new potato variety 'Nagasaki Kogane' with high eating quality, high carotenoid content, and resistance to diseases and pests. Breed. Sci. 2017, 67(3), 320-326. [CrossRef]
- Sun, H.; Jiao, W.B.; Krause, K.; Campoy, J.A.; Goel, M.; Folz-Donahue, K.; Kukat, C.; Huettel, B.; Schneeberger, K. Chromosome-scale and haplotype-resolved genome assembly of a tetraploid potato cultivar. Nat. Genet. 2022, 54(3), 342-348. [CrossRef]
- Hoopes, G.; Meng, X.; Hamilton, J.P.; Achakkagari, S.R.; de Alves Freitas Guesdes, F.; Bolger, M.E.; Coombs, J.J.; Esselink, D.; Kaiser, N.R.; Kodde, L.; Kyriakidou, M.; Lavrijssen, B.; van Lieshout, N.; Shereda, R.; Tuttle, H.K.; Vaillancourt, B.; Wood, J.C.; de Boer, J.M.; Bornowski, N.; Bourke, P.; Douches, D.; van Eck, H.J.; Ellis, D.; Feldman, M.J.; Gardner, K.M.; Hopman, J.C.P.; Jiang, J.; De Jong, W.S.; Kuhl, J.C.; Novy, R.G.; Oome, S.; Sathuvalli, V.; Tan, E.H.; Ursum, R.A.; Vales, M.I.; Vining, K.; Visser, R.G.F.; Vossen, J.; Yencho, G.C.; Anglin, N.L.; Bachem, C.W.B.; Endelman, J.B.; Shannon, L.M.; Strömvik, M.V.; Tai, H.H.; Usadel, B.; Buell, C.R.; Finkers, R. Phased, chromosome-scale genome assemblies of tetraploid potato reveal a complex genome, transcriptome, and predicted proteome landscape underpinning genetic diversity. Mol. Plant. 2022, 15(3), 520-536. [CrossRef]
- Wang, F.; Xia, Z.; Zou, M.; Zhao, L.; Jiang, S.; Zhou, Y.; Zhang, C.; Ma, Y.; Bao, Y.; Sun, H.; Wang, W.; Wang, J. The autotetraploid potato genome provides insights into highly heterozygous species. Plant Biotechnol. J. 2022, 20(10), 1996-2005. [CrossRef]
- Bao, Z.; Li, C.; Li, G.; Wang, P.; Peng, Z.; Cheng, L.; Li, H.; Zhang, Z.; Li, Y.; Huang, W.; Ye, M.; Dong, D.; Cheng, Z.; VanderZaag, P.; Jacobsen, E.; Bachem, C.W.B.; Dong, S.; Zhang, C.; Huang, S.; Zhou, Q. Genome architecture and tetrasomic inheritance of autotetraploid potato [published correction appears in Mol Plant. 2023 Nov 6;16(11):18661866. doi: 10.1016/j.molp.2023.10.005]. Mol. Plant. 2022, 15(7), 1211-1226. [CrossRef]
- Lindhout, P.; Meijer, D.; Schotte, T.; Hutten, R.C.B.; Visser, R.G.F.; van Eck, H.J. Towards F1 Hybrid Seed Potato Breeding. Potato Res. 2011, 54, 301–312 (2011). doi: 10.1007/s11540-011-9196-z.
- Zhang, C.; Yang, Z.; Tang, D.; Zhu, Y.; Wang, P.; Li, D.; Zhu, G.; Xiong, X.; Shang, Y.; Li, C.; Huang, S. Genome design of hybrid potato. Cell. 2021, 184(15), 3873-3883.e12. [CrossRef]
Figure 1.
Plastidial carotenoid pathway in higher plants. Carotenes are boxed in orange, xanthophylls in yellow. Enzymes of major flux-controlling steps in potato are in dark red, the others in orange. G3P, glyceraldehyde 3-phosphate; MEP, metylerythritol 4-phosphate; IPP, isopentenyl diphosphate; DMAPP, dimethylallyl diphosphate; GGPP, geranylgeranyl diphosphate; GAs, gibberellins; ABA, abscisic acid; PSY, phytoene synthase; PDS, phytoene desaturase; ZISO, ζ-carotene isomerase; ZDS, ζ-carotene desaturase; CRTISO carotenoid isomerase; LYCE, lycopene ε-cyclase; LYCB, lycopene β-cyclase; CYP97A and CYP97C, cytochrome P450 carotene β- and ε-ring hydroxylases; CHY (also known as BCH), β-carotene hydroxylase; ZEP, zeaxanthin epoxidase; VDE, violaxanthin de-epoxidase; NSY, neoxanthin synthase. .
Figure 1.
Plastidial carotenoid pathway in higher plants. Carotenes are boxed in orange, xanthophylls in yellow. Enzymes of major flux-controlling steps in potato are in dark red, the others in orange. G3P, glyceraldehyde 3-phosphate; MEP, metylerythritol 4-phosphate; IPP, isopentenyl diphosphate; DMAPP, dimethylallyl diphosphate; GGPP, geranylgeranyl diphosphate; GAs, gibberellins; ABA, abscisic acid; PSY, phytoene synthase; PDS, phytoene desaturase; ZISO, ζ-carotene isomerase; ZDS, ζ-carotene desaturase; CRTISO carotenoid isomerase; LYCE, lycopene ε-cyclase; LYCB, lycopene β-cyclase; CYP97A and CYP97C, cytochrome P450 carotene β- and ε-ring hydroxylases; CHY (also known as BCH), β-carotene hydroxylase; ZEP, zeaxanthin epoxidase; VDE, violaxanthin de-epoxidase; NSY, neoxanthin synthase. .
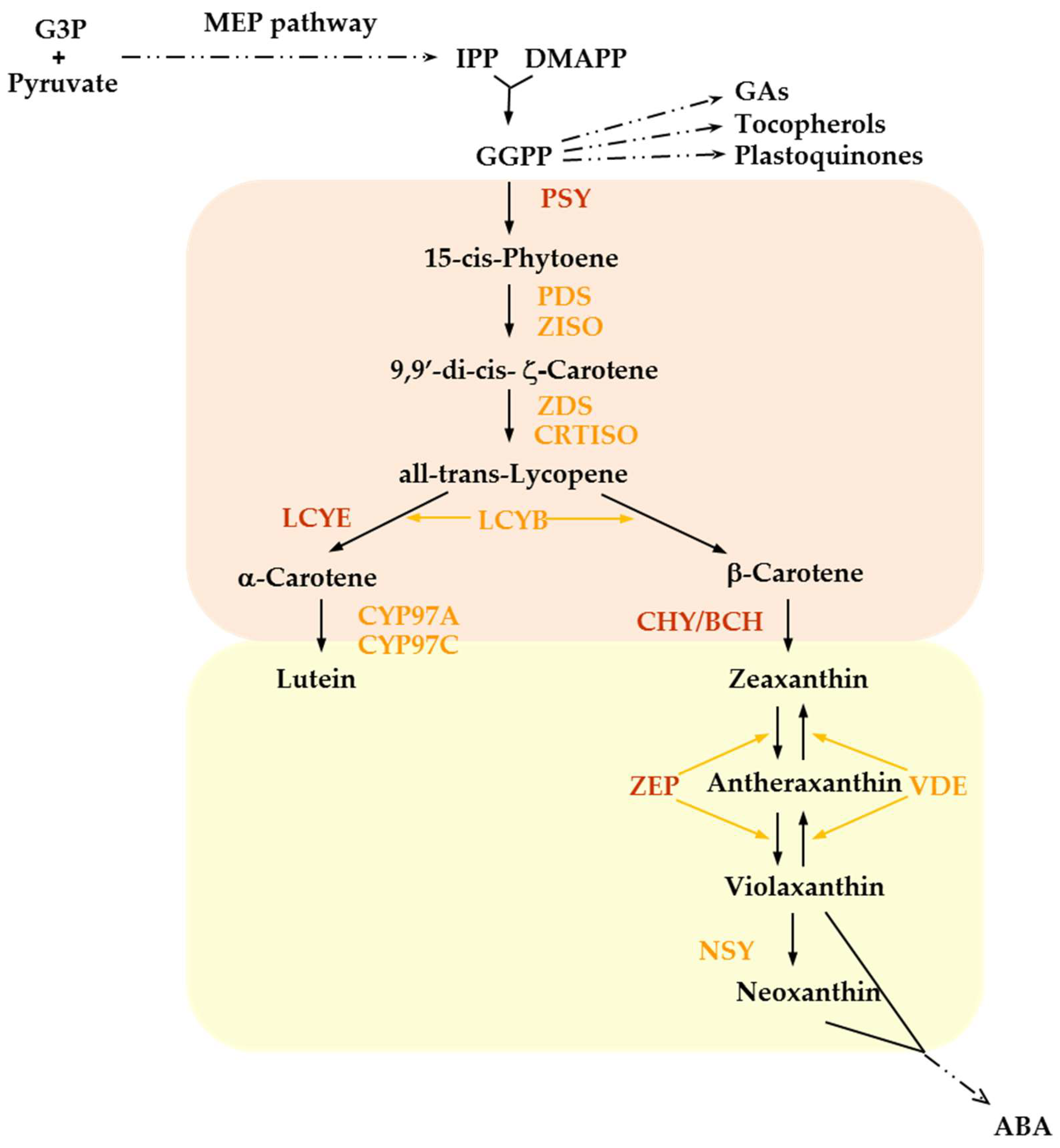

Table 1.
Carotenoid content and profile in potato tubers with different flesh color.
| Taxonomic group (ploidy) | Flesh color (n.) |
TCC | Major components1 | Minor components1 | Refs. |
| S. tuberosum (4n) | W (3) Y (6) DY (4) |
27-74 61 -157 171-343 µg/100g FW |
Lute, Viola Lute-ep. Viola, Lute, Lute-ep., Neo Viola, Lute, Lute-ep., Neo |
Neo, β-Car |
[22] |
|
S. tuberosum (4n) S. phureja x S. stenotomum (2n) |
W (2) Y (11) |
64 and 136 111-1435 µg/100g FW |
Lute, Viola, Lute-ep., Neo Lute-ep., Viola, Lute |
Zea Neo, Zea |
[24] |
| S. tuberosum (4n) | W (4) Y (4) |
38-62 58-175 µg/100g FW |
All: Viola, Anthe, Lute, Zea in different ratios |
All: Neo, β-Cripto, β-Car | [33] |
|
S. tuberosum (4n) S. phureja x S. stenotomum (2n) |
W (7) Y (11) O (1) |
38-265 107-260 878 µg/100g FW |
Lute, Viola Viola, Lute Zea, Anthe, Viola, Lute |
Anthe Neo, Anthe β-Cripto, β-Car |
[34] |
|
S. tuberosum ssp andigena (4n) S. ajanhuiri (2n) S. juzpeczukii (3n) S. phureja (2n) S. stenotomum (2n) |
W (6) C (9) Y (7) P (1) |
1.78-17.90 5.35-18.28 18.85-54.78 16.35 µg/g DW |
All: Lute, Neo, Viola, Zea in different ratios |
All: β-Car, Anthe, β-Cripto |
[25] |
| S. phureja (2n) | C/LY (9) LY/Y (8) Y/DY (6) |
97-262 682-1270 1258-1840 µg/100g FW |
Lute, Viola, Anthe, β-Car Viola, Anthe, Lute, Zea Zea, Anthe |
Zea β-Car Lute, Viola, β-Car |
[26] |
|
S. tuberosum (4n) |
W (3) Y (6) |
101-145 218-511 µg /100g FW |
Lute, Anthe, Viola Anthe, Viola, Zea, Neo |
Zea, Neo Lute |
[36] |
|
S. tuberosum (4n) S. tuberosum ssp andigena (4n) S. phureja (2n) S. stenotomum (2n) S. goniocalix (2n) |
W-P (1) LY (8) Y (46) P-W (5) |
1.66 0.77-6.33 0.54-15.51 0.84-3.27 µg/g DW |
Lute, Neo Lute, Neo or Viola, Neo Viola, Neo Lute, Neo, Viola |
All: Anthe, β-Car, β-Cripto | [28] |
|
S. tuberosum (4n) S. phureja (2n) |
W (2) Y (3) R/P (12) |
All: from 0.779 (W) to 13.3 (Y) µg /g DW |
All: Lute |
All: Viola, Neo, Zea, β-Car | [37] |
|
S. tuberosum (4n) S. chacoense (2n) S. phureja (2n) |
W (2) Y (5) O (3) |
1.37 and 4.1 13.83-32.12 17.42-26.89 µg /g DW |
Lute, Anthe, Neo and Viola, Lute Anthe, Viola, Lute, Zea Zea, Anthe, Lute, |
β-Car, Zea Neo, β-Car Viola, Neo, β-Car |
[38] |
1 = in decreasing order TCC = Total Carotenoid Content, W = White, Y = Yellow, DY = Dark Yellow, O = Orange, C = Cream, P = Purple, R = Red, FW = fresh weight, DW = dry weight, Lute = Lutein, Viola = Violaxanthin, Lute-ep = Lutein-5,6-epoxide, Neo = Neoxanthin, Anthe = Antheraxanthin, Zea = Zeaxanthin, β-Car = β-Carotene, β-Cripto = β-Criptoxanthin. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



