
Submitted:
20 November 2024
Posted:
20 November 2024
You are already at the latest version
Abstract
The Spanish Merino is the most significant sheep breed globally due to its economic and cultural importance in human history. It has also had a substantial influence on the development of other Merino and Merino-derived breeds. Historical sources indicate that crossbreeding to produce finer, higher-quality wool was already taking place in the south of the Iberian Peninsula during the Roman era. This evidence suggests that individuals with a racial pattern very similar to that of the modern Merino may have already existed on the peninsula. The presence of skeletal remains of these animals at various human settlements dated to the late fourth and third millennia BC could provide insights into the genomics of these ancestral sheep. This study analyzes ancient DNA extracted from nine skeletal remains from three archaeological sites in southern Iberia, dated to the third millennium BC. The samples were sequenced and aligned with the ovine genome. The genetic distances observed among the samples indicate a closer relationship between several animals from the Marinaleda (Seville) and Grañena Baja (Jaén) sites, suggesting possible shared livestock practices among these communities, while this was not evident at the La Minilla site (La Rambla, Córdoba). The varying ages of sacrifice or death identified in the faunal studies of the sites producing the samples suggest a focus on meat exploitation at La Minilla (La Rambla, Córdoba), while in Grañena Baja (Jaén) and Marinaleda (Sevilla), the emphasis appears to have been on the use of secondary products. The genomic results of the nine Ovis aries individuals support this hypothesis.
Keywords:
Late fourth and third millennia BC
; livestock production
; sheep
; genomics
; Ancient DNA
; sequencing
1. Introduction
The study of the economic strategies developed during the late fourth and third millennia BC in southern Iberian Peninsula through the analysis of animal bone remains constitutes an essential historical approach, though it encounters various challenges. On the one hand, the number of archaeological sites has increased, most of which are settlements or open-air occupied areas which have been identified and partially excavated over recent decades due to an unprecedented expansion of urban and infrastructural development. However, research, analytical studies, and the publication of results have not expanded to a comparable extent, particularly in the case of with archeozoological studies. Additionally, due to the chronological challenges faunal assemblages are often grouped into very broad periods of time, further complicating a scenario in which synthetic studies are sparse [1,2].
Moreover, in addition to the evident scarcity of specific published studies on the mammalian fauna from certain sites in southern Iberia, we must also consider the weakness of the assemblages analyzed, which typically do not exceed one hundred remains. In our specific case, small ruminant livestock, commonly referred to as ovicaprine (belonging to the subfamily Caprinae), a combined value is given for the minimum number of individuals (MNI).
Despite these limitations, based on the available data, and applying an ecological approach, this study aims to provide a general overview using chronological filters and focusing on Copper Age archaeological sites where faunal assemblages have been analyzed and both the bone remains of Ovis aries and a minimum number of individuals of this species have been identified (Figure 1). Subsequently, utilizing studies conducted on faunal assemblages excavated at the archaeological sites of Grañena Baja (Jaén), Marinaleda (Seville), and La Minilla (La Rambla, Córdoba) [3-5], we will focus on the genetic analysis of nine Ovis aries individuals.
The current evidence, derived from the existing small ruminant bone remains discovered in the southern Iberian Peninsula, could indicate that both species were managed concurrently in single herds during the late fourth and third millennia BC. It is a challenging task to establish comparisons with contemporary sites in the Guadalquivir Valley area, as, despite evidence of substantial occupation of this territory during the third millennium BC [6], few sites have published faunal studies. For this reason, providing an overview of ovine livestock exploitation across different territorial contexts in the southern Iberian Peninsula is not currently feasible. However, despite the limited records, we believe this can serve as a foundation for more robust interpretive proposals focused on the exploitation of this type of livestock in the Guadalquivir Depression during the late fourth and third millennia BC.
1.1. The State of the Art
On reviewing the published data related to faunal lists in which Ovis aries remains from the late fourth and third millennia BC in southern Iberia have been identified [3,5,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25], a recurrent observation is the predominance of domestic fauna, with the caprine-bovine-porcine triad standing out [26].
The current number of Late Neolithic-Copper Age archaeological sites where faunal remains have been studied and a minimum number of Ovis aries individuals have been identified is shown in Table 1.
Several faunal studies have been conducted on the archaeological site of Valencina de la Concepción (Seville), the most comprehensive of which was carried out by Hain (1982), who found that the small ruminant stock (sheep and goat) was the second most exploited. A subsequent study with a smaller sample determined that sheep and goat exploitation ranked third in terms of NR (number of remains), although it was the second most represented in terms of MNI [15]. In the latest study, the sheep and goat herds were found to be the second most represented [15]. In all the studies, sheep were more frequently represented than goats. In the Sierra Norte of Seville, the Cueva de los Covachos provides intriguing insights into livestock practices during the transition from the 4th to the 3rd millennium BC. The predominance of caprine bones discovered suggests their primary role in the economy of that society, with only sheep identified at the site ([31]
The faunal study results from the Cabezo Juré site (Alosno, Huelva) confirm that sheep and goats represent the second most prevalent livestock group, both in terms NISP and MNI [2]. At Papa Uvas, sheep/goat livestock is the most represented along with bovids, with sheep being the most represented species in the first group [24,27,28]. In the archaeological zone of Marroquíes Bajos (Jaén), at the IA Ciudad de la Justicia site, the faunal study determined that sheep and goat husbandry ranked third [34], with sheep being more represented than goats. However, at the Late Neolithic site of Polideportivo de Martos (Martos, Jaén), sheep/goat livestock predominated over bovine and porcine, with the minimum number of individuals (MNI) for goats twice that of sheep [29,30].
In the Copper Age sequence at Los Castillejos de Montefrío (Montefrío, Granada), small ruminant livestock (sheep and goats) predominated in both the early and late phases, and then was relegated to a secondary role during the intermediate period [12]. Throughout much of the third millennium BC, goats dominated the small ruminant herds; however, by the end of this millennium, sheep became predominant.
In Phase I of Cerro de la Virgen (Galera, Granada), in the domestic huts, sheep husbandry held the primary position, with more sheep remains identified than goat remains [8] (Table 1). The maximum lengths of six complete metatarsals recovered indicate that they belong to six different individuals [8].
At the site of Les Moreres (Crevillent, Alicante), during the construction phase of Moreres III, sheep and goat husbandry ranked second, becoming the predominant livestock in Moreres IV, where sheep were the only species identified in both phases [23].
One of the largest Copper Age settlements in southern Iberia is Zambujal (Torres Vedras, Portugal), where one of the most extensive faunal studies, with a significant number of identified remains, was conducted [7]. Sheep and goats were the primary focus of livestock exploitation, although there is no significant difference in the number of remains compared to pig remains [7], with sheep more numerous than goats [7]. The faunal study of Penedo do Lexim (Mafra) shows that small ruminants occupied an almost equal position to suids, with sheep being much more represented than goats. [20].
The archaeozoological record from Ota (Alenquer, Portugal) indicates that the sheep and goat herds constituted the primary source of exploitation, followed by bovines, with more sheep than goats identified [21].
In the sample recovered from Perdigões (Reguengos de Monsaraz, Portugal), sheep and goat husbandry was the second most represented domestic group after suids, although it did not constitute the second highest dietary contribution, due to deer hunting. Within the sheep and goat herds, sheep were more represented than goats [16,22]. A similar situation was found at the Chalcolithic settlement of Monte da Tumba (Torrão, Portugal), where ovine and caprine husbandry ranked second in terms of livestock exploitation, with deer hunting also making a considerable contribution, with significantly more sheep than goats [32]. The same was true at Mercador (Mourão, Portugal), where ovine and caprine livestock ranked second, far behind swine exploitation, with a significant contribution from deer and rabbit hunting [19].
Several other sites, though with a much smaller number of identified remains, can help complete the overall picture. At the Iglesia Antigua de Alcolea site (Córdoba), the primary livestock was pigs, with caprines (sheep and goats) occupying the second place. Among the caprines, more sheep than goats were identified [26]. In Gilena (Seville), the sheep and goat herds were the most represented, with sheep being the most represented species within this group [11]. At the Torreparedones site (Baena), ovine and caprine husbandry ranked third, with sheep being more represented than goats [18]. In Phase B of IAP El Corte Inglés, ovine and caprine livestock ranked second, with sheep being more represented than goats [13]. However, at the Alcalar site (Portimão, Portugal), there was a parity between sheep and goats, with the sheep and goat herds being the second most exploited, after swine farming [33].
In this panoramic view of the late fourth and third millennia BC in southern Iberia, some settlements that undoubtedly had significant territorial influence are notably absent, such as the case of Los Millares. Around 20,000 bone remains from domestic livestock have been studied there, and we know that ovine and caprine husbandry held a priority position, with more sheep identified than goats. However, we do not yet have an estimate of the minimum number of individuals [35]. In the faunal study of Vila Nova de São Pedro (Azambuja, Portugal), no distinction was made between Ovis and Capra [36]. These are just a few of the most significant late fourth and third millennia BC settlements that will likely provide interesting results soon.
Currently, the general impression is that the prevalence of suids at Copper Age sites in southern Iberia depends more on the type of context than on their actual role. This is likely to be related to feasting and communal consumption [37], with caprines actually being the primary livestock. In any case, the number of suids appears to have increased proportionally at sites in the southwestern Iberia [26].
1.2. Geographical Area
The bone remains used for the genomic study of sheep farming in the Guadalquivir Valley were collected from Copper Age archaeological contexts at the sites of Grañena Baja (Jaén), Marinaleda (Seville), and La Minilla (La Rambla, Córdoba) (Figure 2).
The archaeological site of Grañena Baja is located 9 km north of the city of Jaén, on the left bank of the Guadalbullón River. Phase III is defined by the discovery of 108 structures of various types, including ditches, habitation sites, storage facilities, combustion areas, funerary structures, etc. Among these, a considerable number of faunal remains were found [38]. Samples of Ovis aries were recovered from non-ritual contexts NE 25 and NE 131 [3], dating to a period close to the transition from the 4th to the 3rd millennium cal BC.
As a result of the casual discovery of a series of bell beaker vessels in the surrounding area (1986) and with plans for new construction (1989), an emergency archaeological excavation was conducted at the La Minilla site (La Rambla, Córdoba). The faunal remains come from the pit structures, with both structures aligned parallel to each other, which, given the chronology, likely represent an enclosure that would have encompassed a habitation area roughly coinciding with the present-day village [39]. The faunal study indicates that of the domestic fauna, goats were the second most represented, with a greater number of sheep remains identified than goat remains [5]. The two samples of Ovis aries were recovered from trench 1 (Z1) and can be dated to 2834-2470 BC.
The Marinaleda site (Seville) is situated near the eponymous locality, and the fauna remains recovered originate from an archaeological excavation conducted prior to the construction of a school. In the highest area of the hill occupied by the site, sector C was excavated, which proved to be the most interesting due to its significant concentration of structures and material remains. Given the urgent nature of the intervention, it was necessary to select certain structures for excavation, including a large six-meter-wide ditch (Ditch 2), a wall complex adjacent to this ditch, and finally, a trilobulated underground structure [4].
The fill sediment of the three entry structures was very homogeneous, clayey, and dark, with some stone blocks, allowing for the differentiation of various stratigraphic units. In all cases, the walls sloped until they ultimately converged, forming an impressive underground space with three interconnected compartments reaching a maximum depth of 2.10 meters. Among the materials recovered from the stratigraphy (UE82) of this trilobulated structure was the sample of Ovis aries presented in this work, which has also been subjected to radiocarbon dating (C14 AMS), yielding a date with two chronological intervals ranging from 2402 to 2138 cal BC (Table 2). This chronology aligns with the archaeological materials recovered throughout the sequence, not only from UE82 but from the entire site, where bell beaker vessels with impressed decoration and highly burnished black ceramics stand out, accompanied by a wider variety of undecorated elements.
2. Materials and Methods
2.1. Archaeological Approach
The archaeozoological methodology used relied on anatomical and taxonomic identifications using both the reference collections from the Faculty of Veterinary Medicine and the Prehistory Laboratory of the University of Córdoba, as well as specific atlases and studies on agrotypes and domestic forms [12,41-43]. The total number of bones remains (NISP) was used to measure the relative quantity of each taxon. The minimum number of individuals (MNI) per taxon was estimated by comparing the best-represented anatomical element, laterality (right or left), and the state of epiphyseal fusion and dental eruption and wear [44,45]. The Ovis aries taxon was determined following the criteria of various authors [46-50], using a comparative reference collection.
2.2. DNA Purification
The 9 bone remains were processed in the Ancient DNA Laboratory of the MERAGEM research group (University of Cordoba, Cordoba, Spain), where the aDNA was purified following the Yang protocol [51]. Samples from 3 autochthonous Spanish breeds from the south of the Peninsula were employed for comparison with the aDNA. Two Spanish Merino (ME) were selected for their wool and/or wool-meat use, two Black Merino (MN) were selected due to their ancestral origin and two individuals from the Segureña breed were analyzed for their high meat use. Blood from the 6 individuals were collected by jugular venipuncture in vacutainers, with EDTA K3 as the anticoagulant. The genomic DNA was purified with the DNeasy Blood and Tissue Kit (QIAGEN, Hilden, Germany), following the manufacturer’s instructions.
2.3. Sequencing, Alignment and Quality Control
The aDNA from the 9 bone remains, and DNA from the 6 modern sheep, were sent to the National Center of Genomic Assays (CNAG) (Barcelona, Spain). Paired-end sequencing libraries were constructed according to the manufacturer’s instructions (Illumina Inc., San Diego, CA, USA), and then analyzed for size distribution using TapeStation, quantified using Qubit, and sequenced on the Illumina NovaSeq 6000 platform (Illumina Inc., San Diego, California, USA) at PE150. A total of 222.7 GB of data was generated. The quality control of the raw data was conducted with fastqc V0.11.9 software (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ accessed on 24th August, 2024). After filtering, the adapters were removed using fastp v0.23.4 software [52]. The remaining high-quality sequences were aligned with the GCA_016772045.1_ARS-UI_Ramb_v2.0 sheep genome (previously indexed), using the Burrows-Wheeler Aligner (BWA) v0.7.18 software with the “-men” command [53]. Next, the bam files generated for each sample were sorted using the “sort” command in Samtools v1.12 [54]. Duplicates were marked and removed using the “MarkDuplicates” and “RemoveDuplicates” commands in Picard v3.1 software (Picard Toolkit. Cambridge, UK, 2018). The samples were then filtered to keep only the reads with paired reads mapped with quality values of 20, and an alignment score above 100, using Samtools v1.12 software [54].
2.4. Variant Calling
The genotype likelihood was calculated by adding the allelic depth, genotype depth, and strand bias, using the Ovis aries reference genome GCA_016772045.1_ARS-UI_Ramb_v2.0 to generate a bcf file using bcftools (https://github.com/samtools/bcftools accessed on 24th March, 2023). Finally, the genotypes were called with bcftools to generate the vcf file, which was converted to a binary file using plink v1.9 software [55].
2.5. Genomic Comparison Between Ancient Samples
In order to compare the ancient samples, Nei’s genetic distance [56] was calculated with the StAMPP-1.6.3 R package [57] and visualized in a tree using the ape 5.5 package [58] using the Unweighted Pair Group Method with the Arithmetic mean (UPGMA). Samples were sexed using the plinkv1.9 software [4], and inbreeding coefficient (F) and multilocus heterozygosity were estimated with the “calcdiversity” function of R package Sambar [59]. Thereafter, Nei’s genetic distance was used for the principal coordinate analyses (PCoAs) performed using the function ‘pcoa’ of the R package ape-5.7.1 [58].
2.6. Genomic Homology with Modern Breeds
Each of the remains were independently compared with the modern breeds to check similarities through a Principal Components Assay (PCA), analyzed using plink v1.9 software [55].
3. Results
3.1. Genomic Comparison Between Samples from the Different Sites
The nine samples were sexed as females. Figure 3 shows the individual genomic diversity as multi-locus heterozygosity (Figure 3a) and inbreeding coefficient (Figure 3b). Samples from Grañena showed the highest mean levels of Multi locus heterocigosity, while Marinaleda showed the lowest. The dispersion of values was highly significant in the Marinaleda remains.
The Principal Coordinates Analyses (PCoA) calculated for the ancient samples showed how the sheep cluster was in the same position in the different sites, although one sample from Marinaleda (FOS082) was located out of the cloud formed by the rest of samples from this site and nearer to the Grañena samples. The first component captured 35.4% of the variability, while the second PC axis accounted for 28.6% of the total variance (Figure 4).
The dendrogram between individuals indicates a high level of similarity between animals from the same location that are in the same branch. However, sample FOS082 belonging to the Marinaleda site is included in the branch of samples belonging to the Grañena site (Figure 5a). Overall, the genetic distances did not show major differences between the different sites (Figure 5b).
Comparison of the genetic distances of Fst from Wright 1943 and Nei (D) are shown in Figure 6. Both genetic distances show the same tendency, with the most similar sites being the Marinaleda and La Minilla sites, while the most genetically different animals are those belonging to the Minilla and Grañena sites.
3.2. Genomic Comparison Between Ancient Remains and Modern Breeds
The comparison between ancient samples and modern breeds was conducted both jointly, to provide an overall view, and individually, to maximize the number of available DNA markers for each bone remain. When the aDNA were compared with the modern DNA samples, it was observed that all the bone remains clustered in a cloud, regardless of their site, distinct from the modern sheep, which, in turn, were also clustered according to their breed (Figure 7).
The individualized analysis indicated that FOS69 and FOS92 samples from the Marinaleda site exhibited significant genetic distance and equidistance from modern breeds, particularly when compared to the Merino and Black Merino. On the other hand, samples FOS66, FOS82, FOS113, and FOS115, from the Marinaleda and Grañena Baja sites, displayed a significant genetic divergence from both the White Merino and the Segureña sheep, yet clustered more closely with the Black Merino. Finally, the samples from La Minilla demonstrated a greater genetic affinity with one White Merino individual and the Segureña sheep (Figure 8).
4. Discussion
So far, based on the faunal records of Late Neolithic-Copper Age settlements in southern Iberia, it is clear that sheep and goat husbandry held a prominent position alongside pig farming. Pig remains are often found in large quantities, which can be interpreted as evidence of collective consumption events [37]. Domestic caprines were the best-adapted animals to the environment and held a predominant economic role in the region from the Neolithic to the Roman periods. On the other hand, pig remains are usually easier to identify due to their distinct anatomical features, whereas many domestic caprine remains are classified as indeterminate mesomammals, leading to their underrepresentation [3].
In general terms, without delving into the environmental and geographical characteristics of each habitat, it seems that the primary meat source for these human communities was provided by pigs, supplemented by ovicaprid and bovine sacrifices.
Regarding ovicaprids, during this period, there is no evidence to refute the hypothesis that goats and sheep were managed in separate flocks. However, recent studies have shown that mixed herds enhance efficient resource utilization in pasture systems. This synergy arises from the complementary feeding habits of the two species: sheep primarily graze on low-lying grasses, while goats exhibit a preference for browsing on shrubs and higher vegetation. This grazing behavior not only reduces competition for food but also promotes a more extensive growth of ground-level grasses, as goats consume shrubs and reduce vegetative overgrowth, which frees space for grass growth [60]. Goats exhibit more exploratory and aggressive behavior then sheep, and due to their vocalizations and high state of alertness, can serve as effective sentinels, potentially warning the herd of the presence of a predator. Additionally, in extensive grazing systems, the role of an experienced, older goat often proves valuable in guiding movements of the herd, thus facilitating group cohesion [61]. All these reasons suggest that sheep and goats could have been managed together in single, mixed herds during the late fourth and third millennia BC. Recent advances in geoarchaeological studies of stabling floors could shed light on this issue; however, for now, the ethnoarchaeological results are promising [62].
Regarding the different sites, based on the number of identified bones remains and the minimum number of individuals, in most cases, sheep had higher prevalence than goats. A study on aDNA and the application of ZooMS analysis suggest a possible overrepresentation of goats when using the osteomorphological criteria of Zeder and Pilaar (2010), which are based on mandibular teeth to distinguish between sheep and goats [63]. In any case, even when accounting for a possible overestimation of goat remains compared with sheep, herds contained a majority of sheep over goats during the late fourth and third millennia BC. This preference may be attributed to the generally docile nature of sheep, which likely facilitated their management and grazing especially in the Guadalquivir Basin, which is rich in pastures and gentle orography (Figure 2).
In other regions of Europe, in Neolithic and Copper Age contexts, sheep farming seems to have focused on meat production [64]. The same occurred at different sites of the Iberian Peninsula such as Mercador (Mouräo, Portugal), Penedo do Lexim (Mafra, Portugal) and La Minilla (La Rambla, Córdoba), where the mortality rate of individuals younger than 24 months might indicate a meat-based use [5,20].
However, in the cases of Cabezo Jure (Alosno, Huelva), Ciudad de la Justicia (Jaén), IA Corte Inglés (Jaén), Les Moreres (Crevillent, Alicante), Monte da Tumba (Torräo, Portugal), and Perdigões (Reguengos de Monsaraz, Portugal), the slaughter or death of the sheep predominantly occurs during juvenile or adult stages [16,22,32,34], which suggests a comprehensive exploitation of the livestock, not only for meat, but also for secondary resources such as wool, milk, hides, manure, etc. Lastly, data from the mid-3rd millennium BC at Los Castillejos de Montefrío (Montefrío, Granada), Marinaleda (Sevilla), and Ota (Alenquer, Portugal) reveal a high number of adult individuals, which could indicate a trend towards the exploitation of these secondary products. Faunal studies of Late Neolithic-Copper Age settlements with the most identified and individualized sheep remains show the difficulty in differentiating between Ovis/Capra in infant and juvenile specimens. At Valencina de la Concepción (Sevilla), the age of death or sacrifice is assessed based on the ovicaprid group, with one-fifth of the animals being slaughtered before the age of nine months, and slightly less than half being older than two years [9]. The same was found at Zambujal, where a joint assessment of the age of ovicaprids estimated 180 individuals under two years of age and 194 individuals older than two years [7].
The presence of loom weights, crescent-shaped objects, and 'cheese strainers' in archaeological contexts could also be associated with the exploitation of secondary products (wool and milk), whereas their absence might indicate otherwise. One clear example of this is the settlement of Les Moreres (Crevillent, Alicante), where a large number of crescents and loom weights were recovered from two huts dating to the final centuries of the Copper Age [65], which are related to textile and weaving production. The presence of Ovis aries has been confirmed in this context [23]. Conversely, at Mercador, the absence of these artifacts suggests a meat-based exploitation of sheep [20].
However, it is essential to approach these findings with caution, as the earliest known textile discoveries in the Iberian Peninsula were of plant-based fibers. [66]. In some cases, the bias in faunal information that reaches us through the archaeological record should be supplemented with other analytical methods to clarify whether sheep and goat husbandry were being used. A recent study on lipids found inside Neolithic ceramics demonstrates that in the northern and Atlantic regions of the Iberian Peninsula, ovicaprine livestock was already being exploited for both meat and dairy production [67].
The isolated discovery of the “flint dagger” from Wiepenkathen (Germany), found in a peat bog by farmers in 1935 and typologically dated to the Late Neolithic, was encased in a sheath made of sheep leather, decorated with a vegetal pattern. The straps wrapping the artifact were made of cattle hide, the wooden handle was covered with textile fibers from sheep, horse, cow, and goat, and the warp threads were made of plant fibers. This represents the oldest evidence of the use of wool in Europe [68]. However, there has always been an ongoing debate among scholars about the role of sheep as wool producers [69,70], with a more conservative view suggesting the probable absence of woolly sheep breeds in Western Europe before the 3rd millennium BCE [71-73]. The earliest evidence of woolen textiles in southern Iberia is associated with an early Bronze Age burial [74].
The secondary resources of ovicaprids are principally milk and wool. In ovicaprid husbandry, it has historically been shown that goats are more productive in terms of milk yield [75]. Non-improved sheep breeds provide milk to their offspring for approximately 135 days, with a daily milk yield ranging between 0.21 and 0.33 liters per animal [75]. In this context, goats produce more milk than sheep, in some cases doubling the annual milk production in kilograms [76]. Ewes start producing milk between 12 and 18 months after their first lambing, with the age of highest production being between 3 and 5 years [77]. On the other hand, sheep start to produce wool from 12 months, but it is between 2 and 4 years of age that they are most productive. Normally it is from the age of 6 years that ewes start to produce less, and poorer quality, wool [78]. Therefore, the prevalence of sheep in these herds could be due, in addition to their greater docility, to the exploitation of wool, rather than for dairy production, which is higher in goats.
Several studies have attempted to describe the domestication processes of sheep and the origins of current breeds through genomic analyses [79-81]. However, only few works analyze ancient DNA (aDNA) from bone remains, and all of them employed only mitochondrial approaches [63,82,83]. However, recently, paleogenomic techniques have been used to infer domestication process on Caprinae subfamily from Neolithic [84]. In fact, to our knowledge, this is the first study to employ genomic approaches in ancient DNA focused on production uses during the late fourth and third millennia BC.
Currently, the Spanish Merino breed is considered the origin of all contemporary Merino breeds and their derivatives [85]. The oldest written reference to sheep from southern Iberia comes from Columella (42 A.D.), in his work Res Rustica. Here, he describes the crossbreeding practices utilized by his uncle Marcus used to obtain higher-quality wool, noting that white wool is preferable for its ease of dyeing. Additionally, he highlights the value of the dark and blackish fleece of sheep in Italy (Polentia) and in Baetica (Córdoba). In this study, we go back to the late fourth and third millennia BC to investigate the native sheep of the Iberian Peninsula, which later would serve as the basis used by the Romans to establish the genetic stock that would later give rise to the Spanish Merino sheep [85]. Genomic assays showed how individuals from the Marinaleda breed were highly heterogeneous both in terms of multilocus heterozigosity and inbreeding coefficiency, reflecting individuals with different genetic variants (Figure 3a,b). In fact, three of the samples had similar values, which could indicate that they may belong to the same herd. Individuals from La Minilla reflected this trend more clearly and explicitly in its two individuals, which, according to the inbreeding coefficient, could be related in some degree.
The young males kill-off model is one of the strongest arguments used to support domestication events, and has been observed in several Neolithic sites in south-western Asia [86]. The Chalcolithic context of the present study situates it in a time of management events of already domesticated livestock. Thus, one key element that supports the concept of a managed herd structure is the finding that 100% of the samples analyzed were female. This observation aligns with the management practices that have been in place from historical periods to the present, which typically maintain a high female-to-male ratio, estimated at approximately 1 male for every 30 to 50 females [87].
Moreover, it is important to note that the samples used in the genomic analysis comprised adult animals rather than juveniles, where one might expect to observe a female-to-male ratio closer to 1. This discrepancy further emphasizes the implications of management strategies on population dynamics and genetic diversity. However, in the prehistoric communities we have studied, males held a prominently symbolic role, as evidenced by the deposits of butchered crania found at some sites in southern Iberia [3,30].
The aDNA samples from the studied sheep are grouped by their genomic homology based on the site of origin (Figure 4 and Figure 5a). The differences observed may be attributed to a temporal separation of as much as 941 years between the earliest and latest dates at the various sites, according to the radiocarbon dating (Table 2), which may have led to a substantial accumulation of genomic variants. This could have resulted in sheep we assume proceed from a common genetic base becoming highly differentiated due to breeding processes related to the management practices of each time period and geographic locations. However, the archaeological records suggest that, during this period, management practices were conducted in a highly similar way, and that these populations did not accumulate as much genetic change as is seen in modern breeds. Thus, we can assume that the differences found are due to the redistribution of allele frequencies that occurred due to the particularities of management and the specific environmental conditions of each geographical location, as observed in modern herds of the same breed [88].
Although the genetic distances between sites are not very large (approximately between 0.08 and 0.12), it is evident that the most genetically similar animals are those from La Minilla and Marinaleda, while the most genetically distant are those from Grañena Baja and La Minilla (Figure 6). Nevertheless, we have detected a significant level of genetic homology between samples from two different sites. Specifically, an individual from Marinaleda (FOS82) shows genetic similarity with an individual from Grañena Baja (FOS113), as illustrated in the dendrogram of genetic distances (Figure 5a,b).
The fact that all the Copper Age samples have more genetic similarities between them than with some of the modern breeds can be explained by the extensive process of selection that has taken place over the last few centuries in these breeds (Figure 7). In fact, in the last decades, increased professionalization and new diagnostic techniques have meant that selection has developed into genetic improvement. Proof of this is the large number of livestock breeds which have their own breeding program. However, when we analyze the samples individually, we can determine, based on genetic distances, the level of homology they may have with samples of current breeds (Figure 8).
At the Marinaleda site, most of the samples analyzed are equidistant from the current breeds, but with greater similarity to the Black Merino, a trend that is also observed in the Grañena Baja site (Figure 8a,c). However, one of the samples (FOS059) showed similar genetic distances to ME and MN. This could indicate that this individual resembles the endemic animals, but with some features of breeds with greater wool aptitude.
At the La Minilla site, however, the remains are genetically closer to one of the white Merino samples, and very close to the two samples of the Segureña breed (Figure 8b). In other words, these sheep are genetically close to modern breeds with a marked aptitude for meat use. In the case of the Minilla, the animals slaughtered in general were young (males and females), which could indicate a management more focused on meat production where secondary products such as milk and wool would not have such importance for obtaining local resources [5].
This is the first work to analyze the aDNA of sheep from the late fourth and third millennia BC in the Iberian Peninsula, in which we suggest the presence of different types of management, both of primary and secondary products. This may provide an important knowledge base for further research on the sheep that are believed to have derived from the first Merinos.
5. Conclusions
The ovicaprine livestock during the late fourth and third millennia BC, with some exceptions, would have been composed of a greater number of sheep than goats due to their docility and/or wool production. In our work we have sequenced 9 samples of adult individuals from the archaeological sites of Marinaleda, Grañena Baja and La Minilla for genomic study, all of which were sexed as females. This leads us to believe that the ratio of males to females was very similar to that of the current herds. In the case of La Minilla, the slaughtering patterns of young animals indicates that we they were used mainly for meat production, as is also suggested by the genomic analysis by homology with breeds of greater meat or joint meat-wool use. In contrast, at Marinaleda and Grañena Baja, the presence of a greater number of skeletal remains of adult female individuals (over two years of age) is more in line with the exploitation of secondary resources (milk and wool), which is supported by the greater genetic resemblance to the ancestral Merino Negro breed. The hypothesis of wool production at the sites we studied does not contradict the dates accepted for the first appearance of sheep’s wool in Europe [71]. Nevertheless, we consider our work to be an initial approach to the study of ancestral sheep in the south of the Iberian Peninsula. Following the genetic evidence of the present study, we suggest that the Iberian sheep of the late fourth and third millennia BC, which constitute a possible genetic basis for the Merino sheep and its derived breeds, may have been used for wool production since 5000 years ago. The livestock breeding interests of our ancestors in each era clearly has some genomic reflection in the traces of selection in these sheep. We believe that future research is needed to construct a diachronic journey to detect these genomic traces and situate them in their corresponding historical eras.
Author Contributions
“Conceptualization, J.M.G and G.A.; methodology, G.A., A.M., J.A.R. and R.Mª.M.; software, G.A.; formal analysis, G.A. and A.M.; investigation, J.M.G., J.A.R., R.Mª.M., A.P., J.A.C. and A.R. ; resources, J.A.R., R.Mª.M., A.P., J.A.C.; data curation, A.R.; writing—original draft preparation, J.M.G and G.A; writing—review and editing, J.M.G and G.A; supervision, A.M. and J.C.M.; project administration, A.M. and J.C.M.; funding acquisition, A.M. and J.C.M. All authors have read and agreed to the published version of the manuscript.”.
Funding
This study has been funded through the Andalusian Regional Government's call for Excellence Projects 2011 as part of the Project "Patrimonio Arqueozoológico de Andalucía: Análisis genético de las interacciones culturales y de su aplicación a la conservación del patrimonio ganadero actual", led by the research groups INREPA (HUM-262) and MERAGEM (AGR-158).
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Harrison, R.J.; Moreno Lopez, G. El policultivo ganadero o la revolución de los productos secundarios. Trabajos de Prehistoria 1985, 42, 51.
- Riquelme, J.A.; Calvo, F. Aspectos socioeconómicos basados en el estudio de los restos óseos del yacimiento del III Milenio de Cabezo Juré, Alosno (Huelva). In Proceedings of the II-III simposios de prehistoria Cueva de Nerja, Málaga, 2004; pp. 379-385.
- Martínez, R.M.; Conlin, E.; Delgado, A.; Guijo, M.; Granados, A.; Cámara, J.A. Into the circle. Animal and human deposits in a new Upper Guadalquivir site from the beginning of the 3rd millennium Cal BC (Grañena Baja, Jaén). Archaeofauna 2023, 32 (1), 113-128.
- Caro, J.A.; Cruz-Aúñon, R.; García, L. Excavación de urgencia en el asentamiento de la edad del cobre de Marinaleda (Marinaleda, Sevilla). In Proceedings of the Anuario arqueológico de Andalucía 2001, Sevilla, 2004; pp. 920-928.
- Martínez, R.M.; Ruiz, D. Bones in ditches: animal remains at the Copper Age enclosure of La Minilla (La Rambla, Andalusia) in the middle Guadalquivir Valley. Estudos & Memórias 2022, 77. [CrossRef]
- Martínez, R.M. El IV milenio ANE en el Guadalquivir Medio. Intensificación agrícola y fragua de la comunidad doméstica aldeana; Archaeopress: Oxford, 2013.
- Driesch, A.v.d.; Boessneck, J. Die fauna vom Castro do Zambujal. Studien über frühe Tierknochenfunde von der Iberischen Halbinsel 1976, 5, 4-129.
- Driesch, A.v.d. Osteoarchäologische untersuchungen auf der Iberischen Halbinsel; Uni-Druck Muünchen: Muünchen, 1972.
- Hain, F.H. Kupferzeitliche tierknochenfunde aus Valencina de la Concepción; 1982.
- ANTUNES, M.T. O povoado fortificado calcolítico do Monte da Tumba. IV-Mamíferos (nota preliminar). Setúbal Arqueológica 1987, 8, 103-144.
- Cruz-Auñón, R.; Moreno, E.; Rivero, E. Experiencias arqueológicas en Gilena (Sevilla). In Proceedings of the IInd Deya international conference of prehistory. Recent developments in Western Mediterranean prehistory: archaeological techniques, technology and theory, 1991; pp. 313-337.
- Riquelme, J.A. Contribución al estudio arqueofaunístico durante el Neolítico y la Edad del Cobre en las Cordilleras Béticas: el yacimiento arqueológico de Los Castillejos en Las Peñas de los Gitanos, Montefrío (Granada); Universidad de Granada: Granada, 1996.
- Riquelme, J.A. Informe sobre los restos óseos recuperados en la IAP El Corte Inglés" de Jaén. Historia de un arroyo. De Marroquíes Bajos al Centro Comercial El Corte Inglés de Jaén 2011, 310-331.
- Morales, A.; Riquelme, J.A. Faunas de mamíferos del Neolítico Andaluz. In Proceedings of the II-III simposios de prehistoria Cueva de Nerja, 2004; pp. 41-51.
- Abril, D.; Nocete, F.; Riquelme, J.A.; Bayona, M.; Inacio, N. Zooarqueología del III Milenio A.N.E.: El barrio metalúrgico de Valencina de la Concepción (Sevilla). Complutum 2010, 21, 87-100.
- Costa, C. Tafonomia em contexto pré-histórico: a zooarqueologia como recurso para a compreensão das" estruturas em negativo" da pré-história recente. 2013.
- Pajuelo Pando, A.; López Aldana, P.M. Estudio arqueozoológico de estructuras significativas de c/Mariana de Pineda s/n (Valencina de la Concepción, Sevilla). In Proceedings of the El asentamiento prehistórico de Valencina de la Concepción (Sevilla): Investigación y Tutela en el 150 Aniversario del Descubrimiento de La Pastora, 2013; pp. 445-458.
- Martínez, R.M.; Pérez, G.; Peña-Chocarro, L. La campiña de Córdoba entre el IV y el I milenio ANE. Apuntes sobre la ocupación prehistórica del yacimiento de Torreparedones (Baena-Castro del Río, Córdoba). El sondeo 3, al norte del foro. ANTIQVITAS 2014, 26, 135-153.
- Moreno-García, M. Estudo arqueozoológico dos restos faunísticos do povoado calcolítico do Mercador (Mourão). As sociedades agropastoris na margem esquerda do Guadiana (2ª metade do IV e inícios do II milénio AC), Memórias d’Odiana, 2ª série 2013, 321-349.
- Moreno-Garcia, M.; Sousa, A.C. A exploração de recursos faunísticos no Penedo do Lexim (Mafra) durante o Neolítico Final. In Proceedings of the 5. º Congresso do Neolítico Peninsular. Actas., 2015; pp. 67-76.
- Almeida, N.J.; Texugo, A.; Basilio, A.C. ‘Animal farm’: the faunal record from the Chalcolithic Ota site (Alenquer, Portugal) and its regional significance. Documenta Praehistorica 2022, 49, 124-149. [CrossRef]
- Almeida, N.J.; Valera, A.C. Animal consumption and social change: the vertebrates from Ditch 7 in the context of a diachronic approach to the faunal remains at Perdigöes enclosure (3400-2000 BC). Archaeofauna 2021, 30, 75-106. [CrossRef]
- Iborra Eres, M.P. Caza y animales domésticos en el poblado de Les Moreres: análisis arqueozoológico. In Proceedings of the El poblado calcolítico de Les Moreres (Crevillent, Alicante), 2023; pp. 301-310.
- Rojano Simón, M. Principios de estratigrafía arqueológica y arqueometría. Contrastación y validación empírica en Papa Uvas (Aljaraque, Huelva). 2024.
- Morán, M.E. El asentamiento prehistórico de Alcalar (Portimāo, Portugal). La organización del territorio y el proceso de formación de un Estado prístino en la bahía de Lagos en el tercer milenio A.N.E.; UNIARQ WAPS: Lisboa, 2018; Volume 12.
- Martínez, R.M. Cerdos, caprinos y náyades. Aproximación a la explotación ganadera y fluvial en el Guadalquivir entre el Neolítico y la Edad del Cobre (3500-2200 ane). Spal. Revista de Prehistoria y Arqueología (2013, Vol. 22, p. 29-46) 2013.
- Morales Muñiz, A.; Cereijo Pecharromán, M. Consideraciones faunísticas en la transición Neolítico Final-Calcolítico: el yacimiento arqueológico de Papa Uvas (Huelva). Archaeofauna 1992, 1, 87-104.
- Álvarez, M.; Chaves, P. Informe faunístico del yacimiento de Aljaraque (Huelva). Cortes A- 7. 2 y A- 10.4 del Sector A. In Papa Uvas II. Aljaraque. Huelva. Campañas de 1981 a 1983. , Martín de la Cruz, J., Ed.; Ministerio de Cultura: Madrid, 1986.
- Lizcano Prestel, R.; Cámara Serrano, J.A.; Riquelme Cantal, J.A.; Cañabate Guerrero, M.L.; Sánchez Vizcaíno, A.; Afonso Marrero, J.A. El polideportivo de Martos. Producción económica y símbolos de cohesión en un asentamiento del Neolítico Final en las campiñas del Alto Guadalquivir. Cuadernos de prehistoria y arqueología de la Universidad de Granada 1991, 16, 5-101.
- Cámara Serrano, J.A.; Riquelme Cantal, J.A.; Pérez Bareas, C.; Lizcano Prestel, R.; Burgos Juárez, A.; Torres Torres, F. Sacrificio de animales y ritual en el polideportivo de Martos-La Alberquilla (Martos, Jaén). Cuadernos de Prehistoria y Arqueología de la Universidad de Granada 2012, 20, 295-328. [CrossRef]
- Pando, A.P. Usos pecuarios en la transición del IV al III milenio aC en la Sierra Norte de Sevilla. In Proceedings of the Actas del VII Congreso sobre Neolítico en la península ibérica, 2023; pp. 381-393.
- Antunes, M.T. O povoado fortificado calcolítico do Monte da Tumba IV: mamíferos (nota preliminar). Setúbal Arqueológica 1987, 8, 103-144.
- Riquelme, J.A. Anexo 2. Estudio de los restos óseos de vertebrados procedentes de las excavaciones arqueológicas del Poblado Calcolítico de Alcalar, sectores 15L y 16 L. . In El asentamiento prehistórico de Alcalar (Portimāo, Portugal). La organización del territorio y el proceso de formación de un Estado prístino en la bahía de Lagos en el tercer milenio A.N.E., Morán, M.E., Ed.; Estudios & Memorias; UNIARQ WAPS: Lisboa, 2018; Volume 12, pp. 254-302.
- Riquelme, J.A. Una aproximación a la utilización por el hombre de las especies animales documentadas en la Ciudad de la Justicia de Jaén. Ciudad de la Justicia de Jaén. Excavaciones Arqueológicas 2010, 117-133.
- Peters, J.; Driesch, A.v.d. Archäozoologische untersuchung der tierreste aus der kupferzeitlichen siedlung von Los Millares (Prov. Almeria). 1990.
- Detry, C.; Francisco, A.C.; Diniz, M.; Martins, A.; Neves, C.; Arnaud, J.M. Estudo zooarqueológico das faunas do Calcolítico final de Vila Nova de São Pedro (Azambuja, Portugal): Campanhas de 2017 e 2018. Arqueologia em Portugal 2020-Estado da Questão 2020, 925-941.
- Albarella, U.; Davis, S.; Detry, C.; Rowley-Conwy, P. Pigs of the “Far West”: the biometry of Sus from archaeological sites in Portugal. Anthropozoologica 2005, 40, 27-54.
- Conlin, E.; Mercado, L. Actividad Arqueológica Preventiva de excavación arqueológica extensiva y control de movimiento de tierras: “Proyecto de construcción de la Línea de Alta Velocidad Madrid-Alcázar de San Juan, Tramo Grañena-Jaén. Anuario Arqueológico de Andalucía 2013 2013.
- Ruiz, D. Excavación arqueológica de urgencia en La Minilla (La Rambla, Córdoba). Campaña de 1989. In Anuario arqueológico de Andalucía 89.III; Junta de Andalucia, Spain: Sevilla, 1991; pp. 157-163.
- Reimer, P.J.; Austin, W.E.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Ramsey, C.B.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 2020, 62, 725-757.
- Schmid, E. Atlas de huesos de animales Para prehistoriadores, arqueólogos y geólogos del Cuaternario. Knochenatlas. Für prähistoriker, archäologen und quartärgeologen. ; Elsevier: Ámsterdam, Nueva York, 1972.
- Barone, R. Anatomie comparée des mammifères domestiques-ostéologie; Vigot frères éditeurs: París, 1976.
- Rowley-Conwy, P.; Albarella, U.; Dobney, K. Distinguir a los jabalíes de los cerdos domésticos en la prehistoria: una revisión de los enfoques y los resultados recientes. Revista de Prehistoria Mundial 2012, 25, 1-44.
- Reitz, E.; Wing, E. Zooarqueología. Manuales de Arqueología de Cambridge; Cambridge University Press: Cambridge, 2008.
- Grayson, D. Zooarqueología cuantitativa; Elsevier: 1984.
- Boessneck, J.; Müller, H.H.; Teichert, M. Osteologische Unterscheidungsmerkmale zwischen Schaf (Ovis aries Linné) und Ziege (Capra hircus Linné). Kün-Archiv 1964, 78, 1-129.
- Prummel, W.; Frisch, H.-J. A guide for the distinction of species, sex and body side in bones of sheep and goat. Journal of Archaeological Science 1986, 13, 567-577. [CrossRef]
- Halstead, P.; Collins, P.; Isaakidou, V. Sorting the Sheep from the Goats: Morphological Distinctions between the Mandibles and Mandibular Teeth of AdultOvis and Capra. Journal of Archaeological Science 2002, 29, 545-553. [CrossRef]
- Zeder, M.A.; Lapham, H.A. Assessing the reliability of criteria used to identify postcranial bones in sheep, Ovis, and goats, Capra. Journal of Archaeological Science 2010, 37, 2887-2905. [CrossRef]
- Zeder, M.A.; Pilaar, S.E. Assessing the reliability of criteria used to identify mandibles and mandibular teeth in sheep, Ovis, and goats, Capra. Journal of Archaeological Science 2010, 37, 225-242. [CrossRef]
- Yang, D.Y.; Eng, B.; Waye, J.S.; Dudar, J.C.; Saunders, S.R. Improved DNA extraction from ancient bones using silica-based spin columns. Am. J. Phys. Anthropol. 1998, 105, 539-543. [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884-i890.
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754-1760. [CrossRef]
- Danecek, P.; Bonfield, J.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.; Whitwham, A.; Keane, T.; McCarthy, S.; Davies, R. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, giab008.
- Chang, C.; Chow, C.; Tellier, L.; Vattikuti, S.; Purcell, S.; Lee, J. Second-generation PLINK: rising to the challenge of larger and richer datasets. GigaScience 2015, 4. [CrossRef]
- Nei, M. Genetic Distance between Populations. The American Naturalist 1972, 106, 283-292. [CrossRef]
- Pembleton, L.W.; Cogan, N.O.I.; Forster, J.W. StAMPP: an R package for calculation of genetic differentiation and structure of mixed-ploidy level populations. Mol. Ecol. Resour. 2013, 13, 946-952. [CrossRef]
- Paradis, E.; Schliep, K. ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2018, 35, 526-528. [CrossRef]
- de Jong, M.J.; de Jong, J.F.; Hoelzel, A.R.; Janke, A. SambaR: An R package for fast, easy and reproducible population-genetic analyses of biallelic SNP data sets. Mol. Ecol. Resour. 2021, 21, 1369-1379.
- Animut, G.; Goetsch, A.L. Co-grazing of sheep and goats: Benefits and constraints. Small Rumin. Res. 2008, 77, 127-145. [CrossRef]
- Miranda-de la Lama, G.C.; Mattiello, S. The importance of social behaviour for goat welfare in livestock farming. Small Rumin. Res. 2010, 90, 1-10. [CrossRef]
- Égüez, N.; Zerboni, A.; Biagetti, S. Microstratigraphic analysis on a modern central Saharan pastoral campsite. Ovicaprine pellets and stabling floors as ethnographic and archaeological referential data. Quaternary International 2018, 483, 180-193. [CrossRef]
- Pilaar Birch, S.E.; Scheu, A.; Buckley, M.; Çakırlar, C. Combined osteomorphological, isotopic, aDNA, and ZooMS analyses of sheep and goat remains from Neolithic Ulucak, Turkey. Archaeological and Anthropological Sciences 2019, 11, 1669-1681. [CrossRef]
- Serjeantson, D. Review of animal remains from the Neolithic and Early Bronze Age of Southern Britain. 2011.
- Basso, R.E. Actividad textil en el asentamiento calcolítico de Les Moreres: Las pesas de telar y los crecientes de las campañas de 1988-1993. In El poblado calcolítico de Les Moreres (Crevillent, Alicante), González Prats, A., Lorrio Alvarado, A.J., Eds.; Universidad de Alicante: Alicante, 2024; pp. 277-300.
- Gleba, M.; Bretones-García, M.D.; Cimarelli, C.; Vera-Rodríguez, J.C.; Martínez-Sánchez, R.M. Multidisciplinary investigation reveals the earliest textiles and cinnabar-coloured cloth in Iberian Peninsula. Sci. Rep. 2021, 11, 21918.
- Cubas, M.; Lucquin, A.; Robson, H.K.; Colonese, A.C.; Arias, P.; Aubry, B.; Billard, C.; Jan, D.; Diniz, M.; Fernandes, R. Latitudinal gradient in dairy production with the introduction of farming in Atlantic Europe. Nat. Commun. 2020, 11, 1-9.
- Möllers, S. Jungsteinzeit hautnah – Der Flintdolch von Wiepenkathen. In Fundsache: Archäologie zwischen Oste und Elbe, Nösler, D.a.S., A. , Ed.; MCE Verlag in Zusammenarbeit mit der Archäologischen Denkmalpflege des Landkreises Stade und der Stadtarchäologie der Hansestadt Stade.: Drochtersen, 2013; pp. 42-43.
- Prestel, R.L.; Serrano, J.A.C.; Cantal, J.A.R.; Guerrero, M.L.C.; Vizcaíno, A.S.; Marrero, J.A.A. El polideportivo de Martos. Producción económica y símbolos de cohesión en un asentamiento del Neolítico Final en las campiñas del Alto Guadalquivir. Cuadernos de prehistoria y arqueología de la Universidad de Granada 1991, 16, 5-101.
- Cámara, J.A.; Lizcano, R.; Pérez, C.; Gómez del Toro, E. Apropiación, sacrificio, consumo y exhibición ritual de los animales en el Polideportivo de Martos. Sus implicaciones en los orígenes de la desigualdad social. 2008.
- Sherratt, A. Plough and pastoralism: aspects of the secondary products revolution. 1981.
- Sherratt, A. La traction animale et la transformation de l’Europe néolithique. P. Pétrequin 2006, 329-360.
- Davis, S. La arqueología de los animales; Bellaterra: 1989.
- Molina González, F.; Rodríguez-Ariza, M.O.; Jiménez, S.; Botella López, M.C. La sepultura 121 del yacimiento argárico de el Castellón Alto (Galera, Granada). 2003.
- Alling, S. Foragers and Farmers. Prehistoric Archaeology Series 1988.
- Haenlein, G.F.W. About the evolution of goat and sheep milk production. Small Rumin. Res. 2007, 68, 3-6. [CrossRef]
- Gootwine, E.; Pollott, G.E. Factors affecting milk production in Improved Awassi dairy ewes. Animal Science 2000, 71, 607-615. [CrossRef]
- Medhi, D.; Das, P. Nutritional Influences on Wool Growth and Development. 2018.
- Kijas, J.W.; Lenstra, J.A.; Hayes, B.; Boitard, S.; Porto Neto, L.R.; San Cristobal, M.; Servin, B.; McCulloch, R.; Whan, V.; Gietzen, K.; et al. Genome-wide analysis of the world's sheep breeds reveals high levels of historic mixture and strong recent selection. PLoS Biol. 2012, 10, e1001258. [CrossRef]
- Ciani, E.; Lasagna, E.; D’Andrea, M.; Alloggio, I.; Marroni, F.; Ceccobelli, S.; Delgado, J.V.; Sarti, F.; Kijas, J.; Lenstra, J.; et al. Merino and Merino-derived sheep breeds: a genome-wide intercontinental study. Genet. Sel. Evol. 2015, 47, 64. [CrossRef]
- Ceccobelli, S.; Landi, V.; Senczuk, G.; Mastrangelo, S.; Sardina, M.T.; Ben-Jemaa, S.; Persichilli, C.; Karsli, T.; Bâlteanu, V.; Raschia, M.; et al. A comprehensive analysis of the genetic diversity and environmental adaptability in worldwide Merino and Merino-derived sheep breeds. Genet. Sel. Evol. 2023, 55, 24. [CrossRef]
- Scheu, A.; Hartz, S.; Schmölcke, U.; Tresset, A.; Burger, J.; Bollongino, R. Ancient DNA provides no evidence for independent domestication of cattle in Mesolithic Rosenhof, Northern Germany. Journal of Archaeological Science 2008, 35, 1257-1264. [CrossRef]
- Kahila Bar-Gal, G.; Ducos, P.; Kolska Horwitz, L. The application of ancient DNA analysis to identify neolithic caprinae: a case study from the site of Hatoula, Israel. International Journal of Osteoarchaeology 2003, 13, 120-131. [CrossRef]
- Kaptan, D.; Atağ, G.; Vural, K.B.; Morell Miranda, P.; Akbaba, A.; Yüncü, E.; Buluktaev, A.; Abazari, M.F.; Yorulmaz, S.; Kazancı, D.D.; et al. The Population History of Domestic Sheep Revealed by Paleogenomes. Mol. Biol. Evol. 2024, 41. [CrossRef]
- Granero, A.; Anaya, G.; Demyda-Peyrás, S.; Alcalde, M.J.; Arrebola, F.; Molina, A. Genomic Population Structure of the Main Historical Genetic Lines of Spanish Merino Sheep. Animals 2022, 12, 1327.
- Arbuckle, B.; Atici, L. Initial diversity in sheep and goat management in Neolithic south-western Asia. Levant 2013, 45, 219-235. [CrossRef]
- Lianou, D.T.; Vasileiou, N.G.C.; Michael, C.K.; Valasi, I.; Mavrogianni, V.S.; Caroprese, M.; Fthenakis, G.C. Patterns of Reproductive Management in Sheep and Goat Farms in Greece. Animals 2022, 12, 3455.
- Anaya, G.; Granero, A.; Alcalde, M.J. Situación genética de las principales líneas puras del merino español. ITEA-Informacion Tecnica Economica Agraria 2024, 120 (2), 133-143. [CrossRef]
Figure 1.
Sites from the late fourth and third millennia BC in southern Iberia with a minimum number of Ovis aries individuals identified. Spain: 1. Valencina de la Concepción (Sevilla), 2. Cabezo Juré (Alosno, Huelva), 3. Papa Uvas (Aljaraque, Huelva), 4. Gilena (Sevilla), 5. Marinaleda (Sevilla), 6. La Minilla (La Rambla, Córdoba), 7. Torreparedones (Baena, Córdoba), 8. Antigua Iglesia de Alcolea (Córdoba), 9. Grañena Baja (Jaén), 10. IA Corte Inglés (Jaén), 11. Ciudad de la Justicia (Jaén), 12. Polideportivo de Martos (Martos, Jaén), 13. Los Castillejos (Montefrío, Granada), 14. Cerro de la Virgen (Galera, Granada), 15. Les Moreres (Crevillent, Alicante). 16. Cueva de los Covachos (Almadén de la Plata, Sevilla). Portugal: 17. Zambujal (Torres Vedra), 18. Penedo do Lexim (Mafra), 19. Ota (Alenquer), 20. Perdigões (Reguengos de Monsaraz), 21. Monte da Tumba (Torrão), 22. Mercador (Mourão), 23. Alcalar (Portimão). Reference map: https://www.ign.es/iberpix/visor/. Iberpix is a cartographic viewer published by the National Center for Geographic Information (CNIG) and the National Geographic Institute of Spain (IGN) used for the consultation and visualization of maps and layers of geographic information.
Figure 1.
Sites from the late fourth and third millennia BC in southern Iberia with a minimum number of Ovis aries individuals identified. Spain: 1. Valencina de la Concepción (Sevilla), 2. Cabezo Juré (Alosno, Huelva), 3. Papa Uvas (Aljaraque, Huelva), 4. Gilena (Sevilla), 5. Marinaleda (Sevilla), 6. La Minilla (La Rambla, Córdoba), 7. Torreparedones (Baena, Córdoba), 8. Antigua Iglesia de Alcolea (Córdoba), 9. Grañena Baja (Jaén), 10. IA Corte Inglés (Jaén), 11. Ciudad de la Justicia (Jaén), 12. Polideportivo de Martos (Martos, Jaén), 13. Los Castillejos (Montefrío, Granada), 14. Cerro de la Virgen (Galera, Granada), 15. Les Moreres (Crevillent, Alicante). 16. Cueva de los Covachos (Almadén de la Plata, Sevilla). Portugal: 17. Zambujal (Torres Vedra), 18. Penedo do Lexim (Mafra), 19. Ota (Alenquer), 20. Perdigões (Reguengos de Monsaraz), 21. Monte da Tumba (Torrão), 22. Mercador (Mourão), 23. Alcalar (Portimão). Reference map: https://www.ign.es/iberpix/visor/. Iberpix is a cartographic viewer published by the National Center for Geographic Information (CNIG) and the National Geographic Institute of Spain (IGN) used for the consultation and visualization of maps and layers of geographic information.
Figure 2.
Geographical location of the archaeological sites.
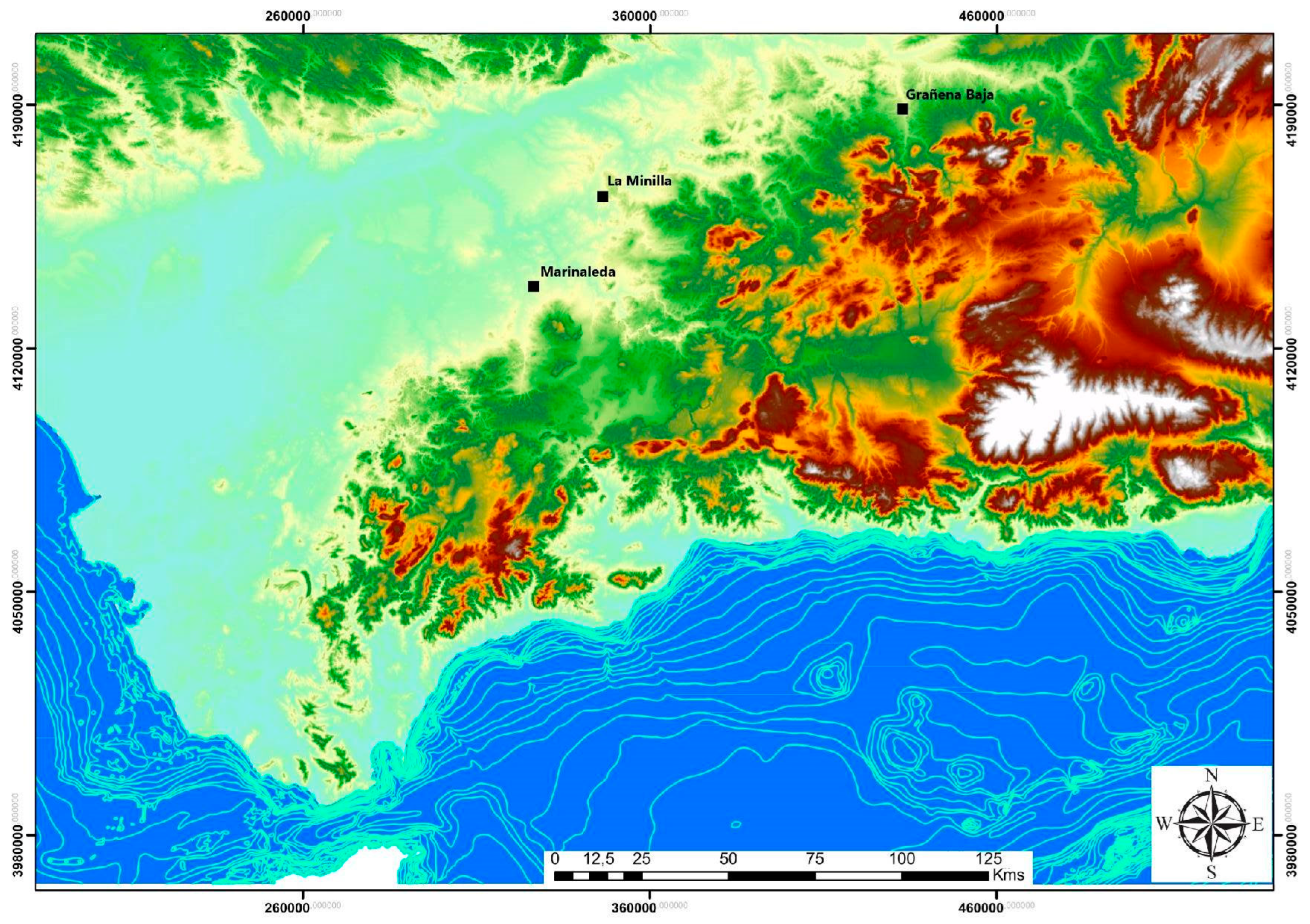
Figure 3.
Genomic diversity of the bone remains by site. (a) Multi-locus heterozygosity at individual level. (b) Inbreeding coefficient at individual level.
Figure 3.
Genomic diversity of the bone remains by site. (a) Multi-locus heterozygosity at individual level. (b) Inbreeding coefficient at individual level.
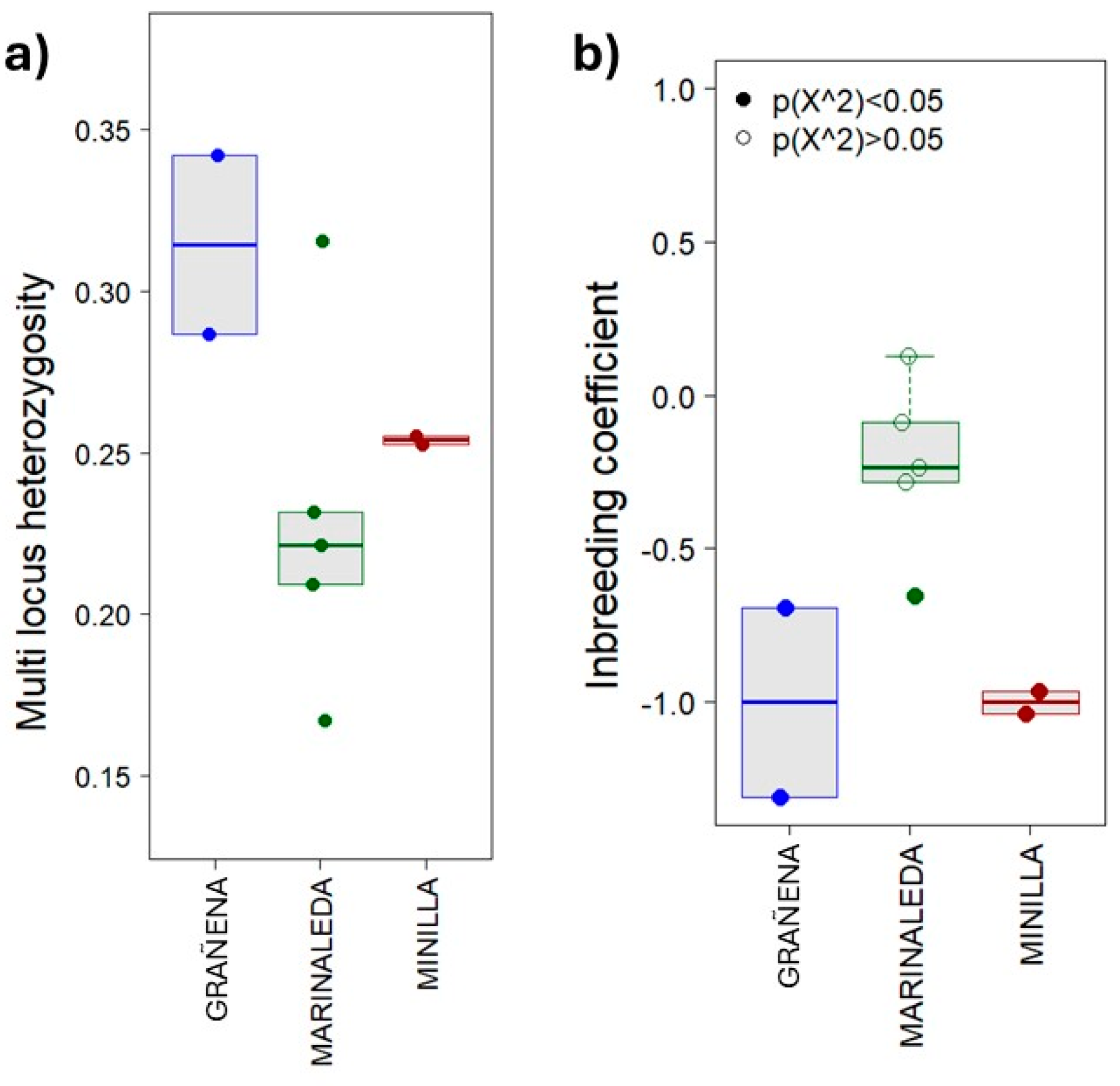
Figure 4.
Principal Coordinates Analysis of the bone remains from the 3 sites of the study. Red ellipse corresponds to the Minilla remains, green ellipse to the Marinaleda bones and blue ellipse to samples from Grañena.
Figure 4.
Principal Coordinates Analysis of the bone remains from the 3 sites of the study. Red ellipse corresponds to the Minilla remains, green ellipse to the Marinaleda bones and blue ellipse to samples from Grañena.
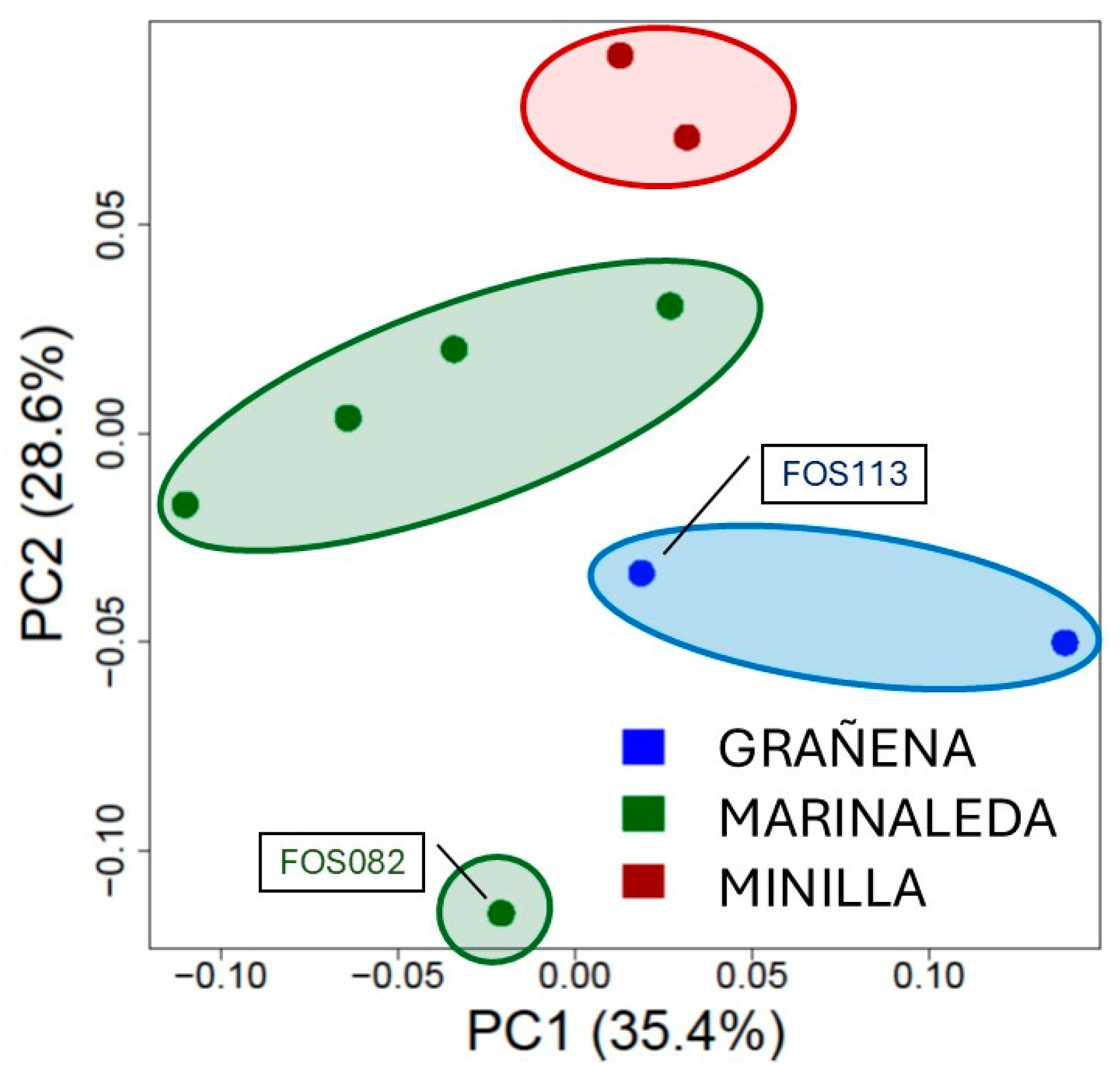
Figure 5.
(a) Dendrogram depicting Euclidean genetic distances between individuals. (b) Nei’s Genetics distances between individuals. Red ellipse corresponds to the Minilla remains, green ellipse to the Marinaleda bones and blue ellipse to samples from Grañena.
Figure 5.
(a) Dendrogram depicting Euclidean genetic distances between individuals. (b) Nei’s Genetics distances between individuals. Red ellipse corresponds to the Minilla remains, green ellipse to the Marinaleda bones and blue ellipse to samples from Grañena.
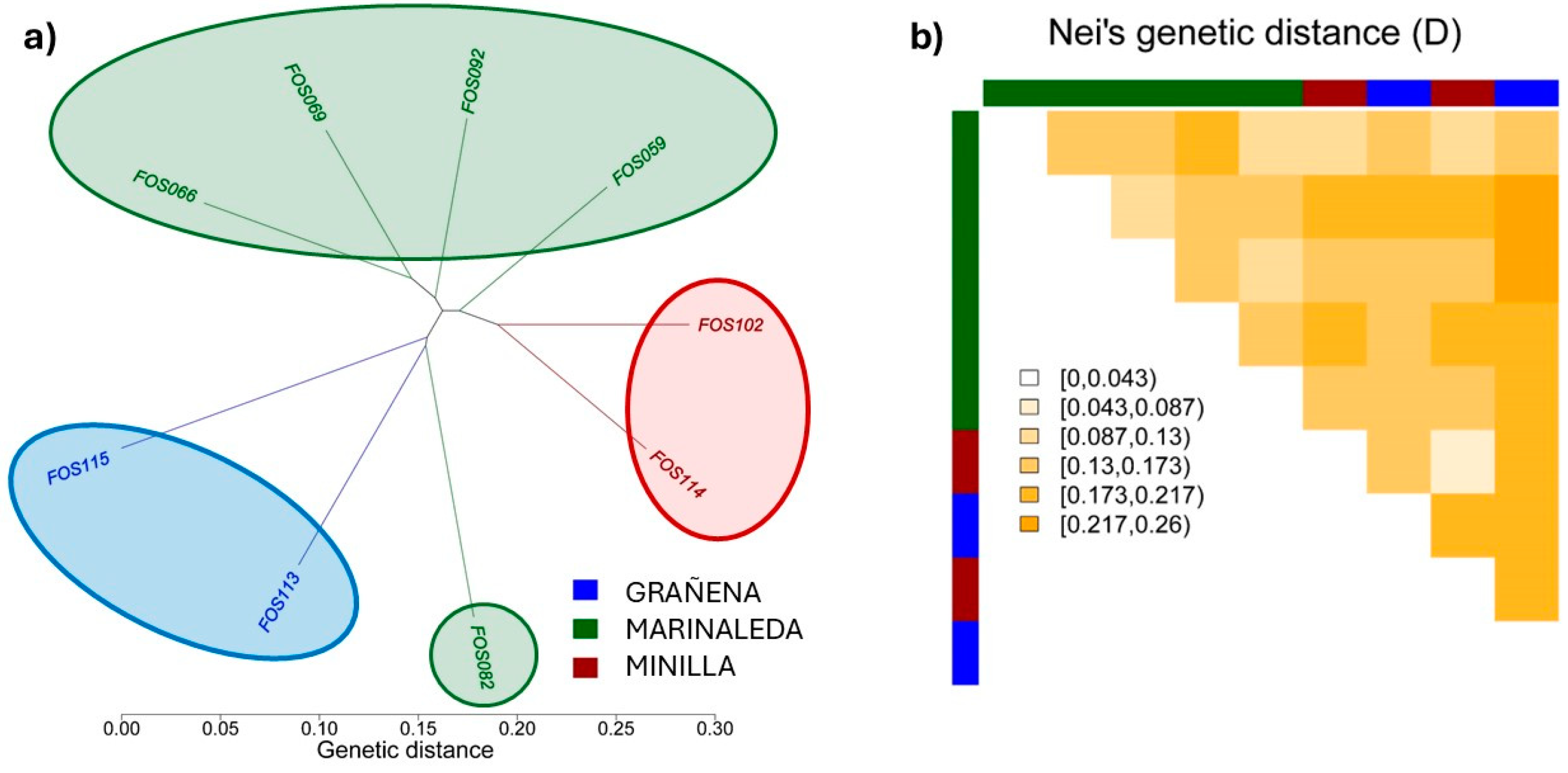
Figure 6.
Representation of the genetic distances of Wright 1943 Fst and Nei (D), between the sites of Grañena, Marinaleda and Minilla.
Figure 6.
Representation of the genetic distances of Wright 1943 Fst and Nei (D), between the sites of Grañena, Marinaleda and Minilla.
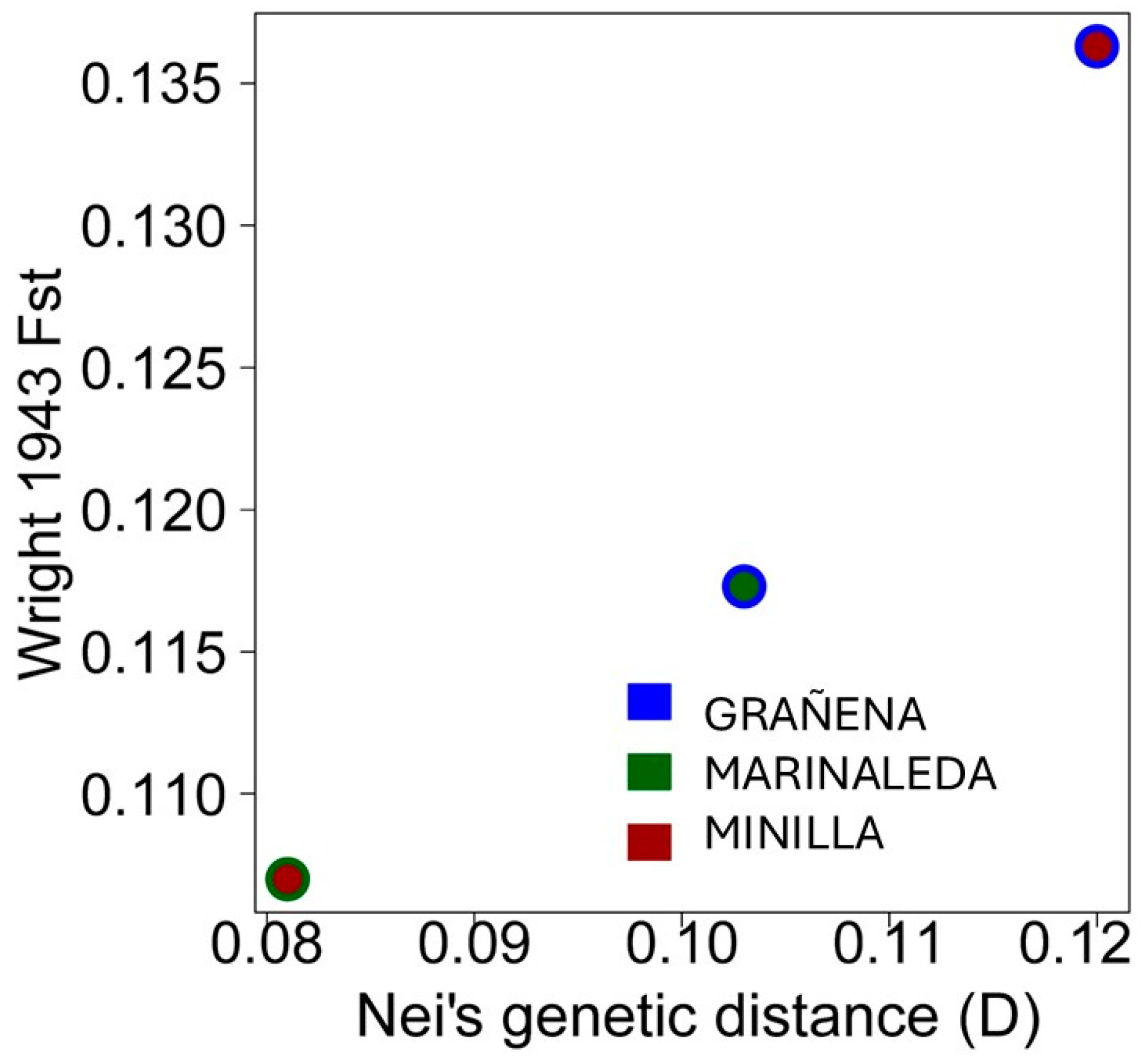
Figure 7.
Principal Component Analysis of the bone remains from the 3 sites of the study compared with the modern Segureña, Merino and Black Merino breeds.
Figure 7.
Principal Component Analysis of the bone remains from the 3 sites of the study compared with the modern Segureña, Merino and Black Merino breeds.
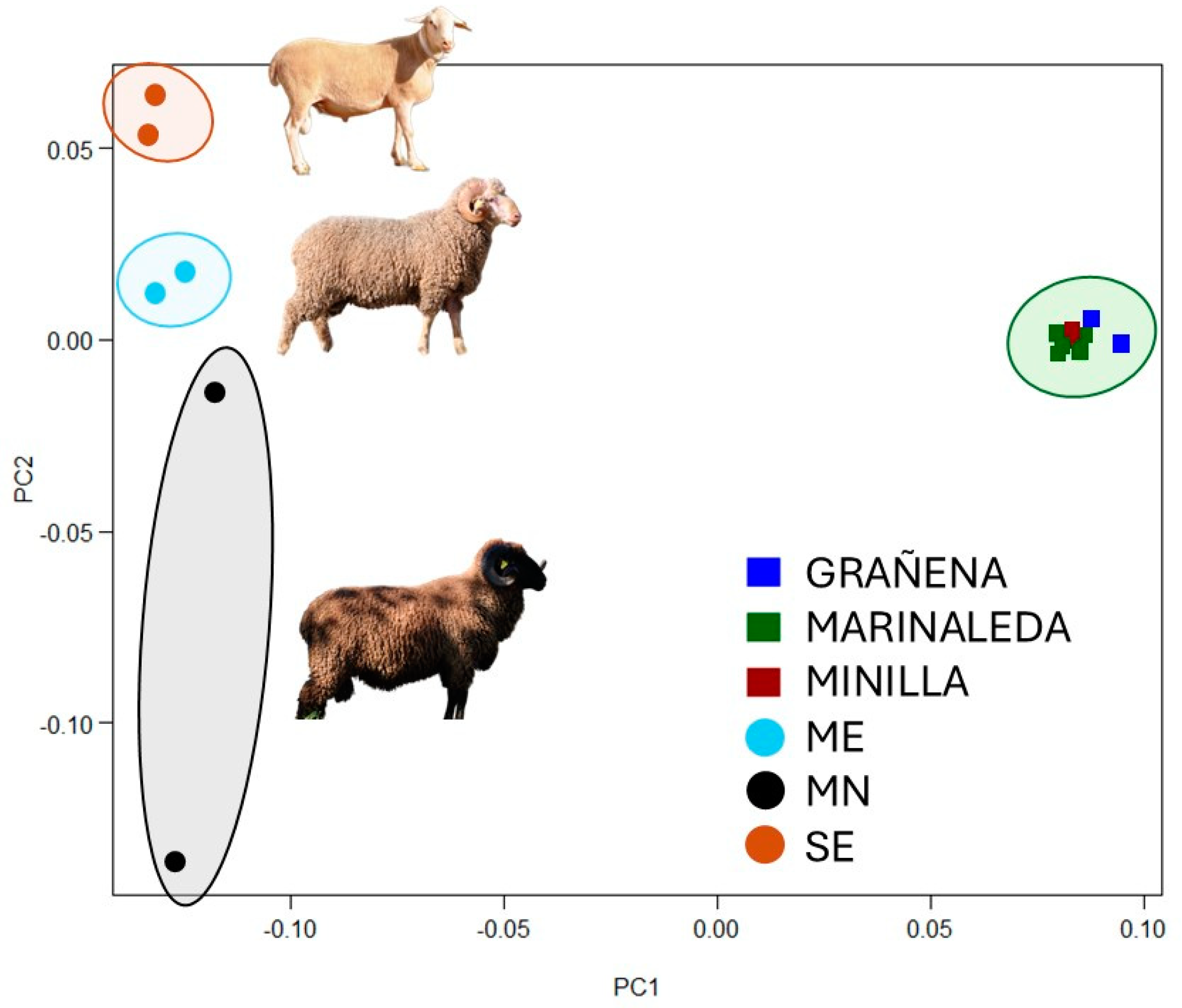
Figure 8.
Figure 8. Individualized, Principal Component Analysis of the bone remains from the Marinaleda (a), La Minilla (b) and Grañena (c) sites of the study compared with the modern Segureña, Merino and Black Merino breeds.
Figure 8.
Figure 8. Individualized, Principal Component Analysis of the bone remains from the Marinaleda (a), La Minilla (b) and Grañena (c) sites of the study compared with the modern Segureña, Merino and Black Merino breeds.
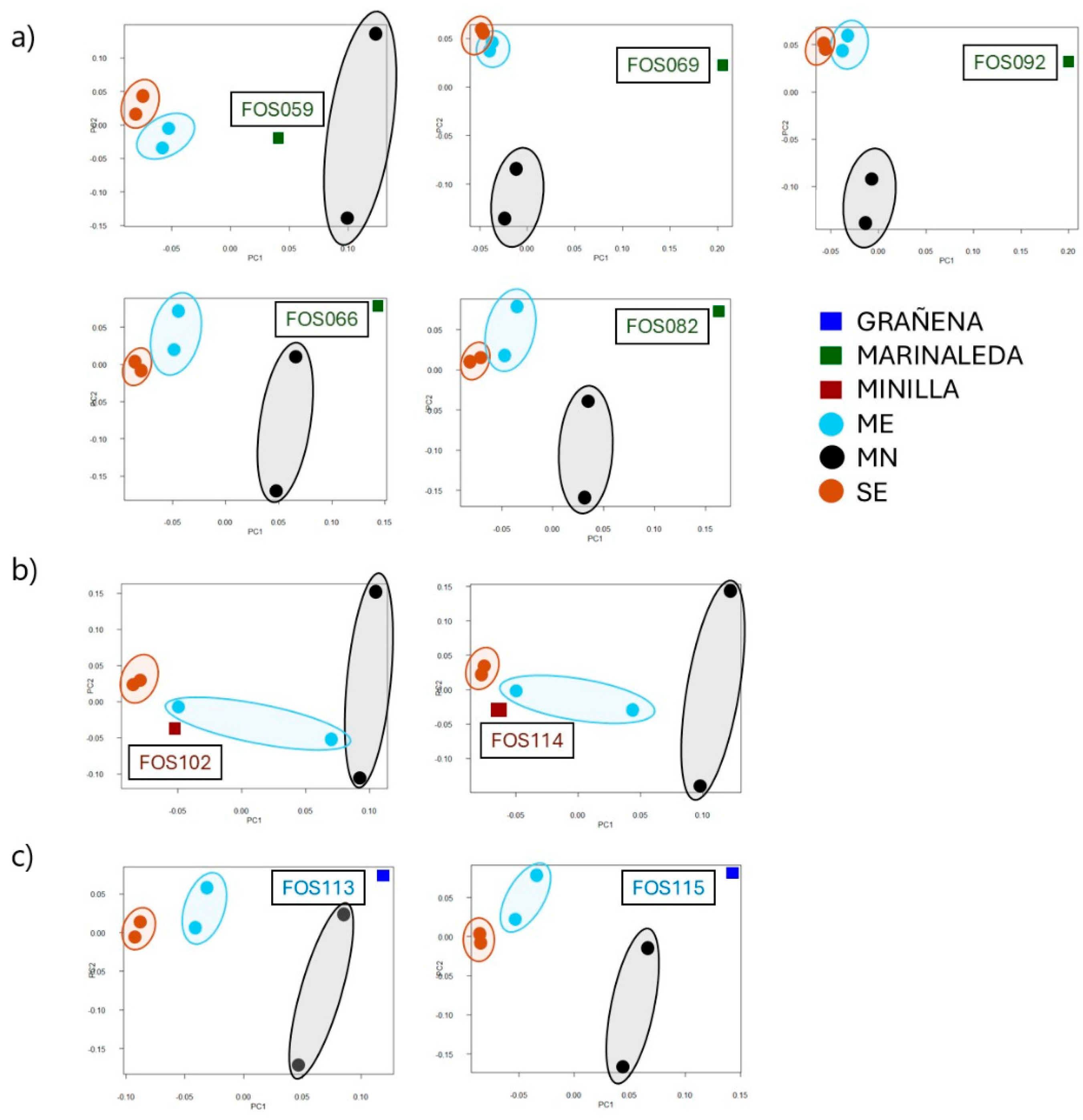

Table 1.
Minimum Number of Ovis aries Individuals found at sites in southern Iberia.
| Site | *MNI | **Dates | |
|---|---|---|---|
| Valencina de la Concepción (Sevilla) | 114 [9], 4 [15], 8 [17] | III millennium BC | |
| Cabezo Juré (Alosno, Huelva) | 34 [2] | 2500 Cal BC. | |
| Papa Uvas (Aljaraque, Huelva) | 6 [27], 5 [28], 1 [24] | 3092-3052 Cal BC | |
| Gilena (Sevilla) | 3 [11] | End IV millennium/ Beginning III millennium BC | |
| Marinaleda (Sevilla) | 4 (this paper) | 2346-2138 Cal BC | |
| La Minilla (La Rambla, Córdoba) | 9 [5] | 2834-2470 Cal BC | |
| Torreparedones (Baena, Córdoba) | 2 [18] | 3020 Cal BC | |
| Iglesia Antigua de Alcolea (Córdoba) | 2 [6] | 3200 Cal BC | |
| Grañena Baja (Jaén) | 2 [3] | Beginning III millennium BC | |
| IA Corte Inglés (Jaén) | 1 [13] | III millennium BC | |
| Ciudad de la Justicia (Jaén) | 10 (Inédito) | III millennium BC | |
| Polideportivo de Martos (Martos, Jaén) | 4 [29,30] | Late IV millennium Cal BC | |
| Los Castillejos de Montefrío (Montefrío, Granada) | 24 [12] | 2325 Cal BC | |
| Cerro de la Virgen (Galera, Granada) | 8 [8] | Mid III millennium BC | |
| Les Moreres (Crevillent, Alicante) | 4 [23] | Mid/late III millennium BC | |
| Cueva de los Covachos (Almadén de la Plata, Sevilla) | 3 [31] | End IV millennium/ Beginning III millennium BC | |
| Zambujal (Torres Vedra) | 108 [7] | 2500 Cal BC | |
| Penedo do Lexim (Mafra) | 5 [20] | 2890-2620 Cal BC | |
| Ota (Alenquer, Portugal) | 6 [21] | First half of III millennium BC | |
| Perdigões (Reguengos de Monsaraz, Portugal). | 3 [16], 2 [22] | III millennium BC | |
| Monte da Tumba (Torrão, Portugal) | 33 [32] | First half of III millennium BC | |
| Mercador (Mourão, Portugal) | 14 [19] | Mid/late III millennium BC | |
| Alcalar (Portimão, Portugal). | 5 [33] | 2577-2335 Cal BC | |
* MNI: Minimum number of individuals. Corresponding reference in square brackets [ ] and numbered. ** Dates of archaeological sites. Cal BC: Date Before Christ calibrated by Radiocarbon; BC: Uncalibrated date Before Christ.
Table 2.
Radiocarbon dates of archaeological samples.
| Sites | Code | BP | SD | Cal BC 95.4 % | m | Bone Sample | References |
|---|---|---|---|---|---|---|---|
| La Minilla | CNA-3151 | 3996 | 35 | 2619-2460 | 2525 | Cervus elaphus | [5] |
| La Minilla | CNA-3153 | 4034 | 36 | 2834-2470 | 2543 | Sus scrofa | [5] |
| La Minilla | CNA-3152 | 4040 | 35 | 2835-2472 | 2552 | Bos taurus | [5] |
| Grañena Baja | Beta-573496 | 4230 | 30 | 2910-2697 | 2858 | Human femur | [3] |
| Grañena Baja | Beta-573497 | 4330 | 30 | 3021-2891 | 2943 | Human tibia | [3] |
| Grañena Baja | CNA-3197 | 4347 | 35 | 3082-2895 | 2967 | Human bone | [3] |
| Grañena Baja | CNA-3194 | 4351 | 33 | 3083-2898 | 2968 | Human femur | [3] |
| Marinaleda | CIRAM-11738 | 3805 | 32 | 2346-2138 | 2244 | Ovis aries | This paper |
Code: Identification of the samples with radiocarbon dating. BP: Before Present; SD: Standard deviation; Cal BC 95.4%: Atmospheric curve employed: IntCal20 [40].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



